Explore article hub
 Masanori Aikawa1,2,3*†
Masanori Aikawa1,2,3*† Abhijeet R. Sonawane1,2†
Abhijeet R. Sonawane1,2† Sarvesh Chelvanambi1†
Sarvesh Chelvanambi1† Takaharu Asano1
Takaharu Asano1 Arda Halu3
Arda Halu3 Joan T. Matamalas1
Joan T. Matamalas1 Sasha A. Singh1
Sasha A. Singh1 Shizuka Uchida4
Shizuka Uchida4 Elena Aikawa1,2
Elena Aikawa1,2 Alex Arenas5
Alex Arenas5 Jean-Luc Balligand6
Jean-Luc Balligand6 Chiara Giannarelli7
Chiara Giannarelli7 Calum A. MacRae8
Calum A. MacRae8 Neil V. Morgan9
Neil V. Morgan9 Cécile Oury10
Cécile Oury10 Hendrik Tevaearai Stahel11
Hendrik Tevaearai Stahel11 Joseph Loscalzo12
Joseph Loscalzo12- 1Center for Interdisciplinary Cardiovascular Sciences, Division of Cardiovascular Medicine, Department of Medicine, Brigham and Women’s Hospital, Harvard Medical School, Boston, MA, United States
- 2Center for Excellence in Vascular Biology, Division of Cardiovascular Medicine, Department of Medicine, Brigham and Women’s Hospital, Harvard Medical School, Boston, MA, United States
- 3Channing Division of Network Medicine, Department of Medicine, Brigham and Women’s Hospital, Harvard Medical School, Boston, MA, United States
- 4Center for RNA Medicine, Department of Clinical Medicine, Aalborg University, Copenhagen, Denmark
- 5Department of Computer Engineering and Mathematics (DEIM), Rovira I Virgili University (URV), Tarragona, Spain
- 6Institute of Experimental and Clinical Research, Pole of Pharmacology and Therapeutic, Cliniques Universitaires St-Luc, Université Catholique de Louvain, Brussels, Belgium
- 7NYU Cardiovascular Research Center, Division of Cardiology, Department of Medicine, Department of Pathology, New York University Grossman School of Medicine, New York, NY, United States
- 8Division of Cardiovascular Medicine, Department of Medicine, Brigham and Women’s Hospital, Harvard Medical School, Boston, MA, United States
- 9Department of Cardiovascular Sciences, School of Medical Sciences, College of Medicine and Health, University of Birmingham, Birmingham, United Kingdom
- 10Laboratory of Cardiology, GIGA-Cardiovascular Sciences, University of Liège, Liège, Belgium
- 11Bern University Hospital, University of Bern, Bern, Switzerland
- 12Department of Medicine, Brigham and Women’s Hospital, Harvard Medical School, Boston, MA, United States
Abstract
Despite the development of potent drugs for modifiable risk factors and advances in mechanistic biomedical research, cardiovascular diseases (CVDs) collectively remain the leading cause of death globally, indicating a need for new, more effective therapies. A foundational challenge is the multilevel heterogeneity that characterizes CVDs—from their complex pathobiological mechanisms at the molecular and cellular levels, to their clinical presentations and therapeutic responses at the individual and population levels. This variability arises from individuals’ unique genomic and exposomic characteristics, underscoring the need for precision approaches. Other key challenges include the long navigation times, high costs, and low success rates for drug development, often compounded by the poor “druggability” of new targets. In this article, we explore how these challenges have inspired novel technologies that offer promise in improving health outcomes globally through an integrative precision medicine approach. Key to this transformation is the use of systems biology and network medicine, whereby the application of artificial intelligence to “big data”, ranging from clinical information to unbiased multiomics (e.g., genomics, transcriptomics, proteomics, and metabolomics) can elucidate disease mechanisms, yield novel biomarkers for disease progression, and identify potential drug targets. In parallel, new computational approaches are helping translate these discoveries into novel therapies and overcome druggability barriers. The transition to a precision-based research and innovation paradigm in cardiovascular medicine will require greater interdisciplinary collaboration, data science implementation at every stage, and new partnerships between academia and industry. Global policy leadership is also essential to implement suitable models of research funding and organization, data infrastructures and policies, medicines regulations, and patient access policies promoting equity.
Key points
- Heterogeneity in the complex pathobiology and presentation of cardiovascular diseases (CVDs)—the leading cause of death globally—limits the effectiveness of conventional “one-size-fits-all” therapies.
- Integrative systems approaches, which involve unbiased omics analyses (especially proteomics and single-cell analyses), bioinformatics, and network science, now offer the potential for a precision medicine innovation paradigm to tackle CVDs.
- Artificial intelligence (AI) is driving new opportunities in patient profiling and computational precision drug design and development.
- Various RNA therapeutics in development offer promise for effective precision therapy and could help address conventional drug development obstacles, such as cost and time.
- Interdisciplinary and intersectoral collaboration throughout the research and innovation pathway, underpinned by global health policy leadership, is necessary to implement a precision cardiovascular medicine paradigm.
Introduction: challenges are opportunities
“The greater the obstacle, the more glory in overcoming it.” Molière
Challenges are opportunities. Clinical problems and unanswered questions in cardiovascular medicine have driven enormous global efforts aiming to understand the underlying causes of cardiovascular disease (CVD) (1–10). Such needs have also triggered a series of highly valuable technological innovations. Investigation into cardiovascular research in its current form began over a century ago (11–14). In the last several decades, our community has focused on educating the public about lifestyle modifications and dietary interventions to prevent or manage CVD (15). The successful development of effective medicines, such as statins and the newer proprotein convertase subtilisin/kexin type 9 (PCSK9) inhibitors, has significantly contributed to reducing the incidence of certain cardiovascular conditions (16–19). Additionally, advances in basic science have identified various molecules and pathways, including interleukin (IL)-1β and IL-6, which highlight the role of inflammation beyond traditional modifiable cardiovascular risk factors (20).
Despite these efforts and remarkable advancements, a substantial risk persists for many patients (17, 21–23). While some therapies, such as glucagon-like peptide-1 receptor (GLP-1R) agonists, can reduce the burden of chronic diseases (24, 25), there is still a dire need to identify additional strategies to address the complex interplay across these diseases. While there is a benefit to optimal treatment of type 2 diabetes in terms of being able to reduce the excess mortality risk associated with CVD, this effect was only seen in patients with no previous diagnoses of CVD. This suggests a strong rationale for fine-tuning therapeutic strategies to combat complex diseases (26). From 2010 to 2019, the number of deaths caused by hypertension increased among adults aged 35–64 in 86.2% of counties in the United States (27). The burden of CVD is projected to grow over the next few decades. The number of elderly patients with calcific aortic valve stenosis, a major complication of chronic kidney disease, is projected to more than double by 2050 in the United States and Europe (28). Overall, CVD continues to be the leading cause of death globally (17), resulting in 19 million deaths worldwide in 2020. This toll increased by 18.7% in the past decade and will likely rise to 26 million by 2030 (28–30).
Difficult problems require innovative solutions. One of the foundational challenges that complicates CVD treatment is the heterogeneity of the complex pathobiological mechanisms and clinical presentations of CVDs. High-throughput, multimodality, multiomics data have ushered in a new era of big data in CVD research, with each type of data introducing a new layer of complexity to the existing challenge of identifying novel mechanisms and biomarkers (31). Addressing these numerous challenges is necessary for advancing precision CVD medicine.
In this article, we explore how key challenges in cardiovascular medicine have inspired novel technologies that offer promise in improving health outcomes globally. We illustrate how systems biology, network medicine, and artificial intelligence (AI), supported by technological innovations in omics, are providing meaningful biological insights into the complexity of disease heterogeneity—fostering new discoveries and targets for precision medicine interventions. We then discuss computational drug discovery and new classes of medicines that may overcome the limitation of conventional small-molecule drugs. Finally, we outline the interdisciplinary and intersectoral actions, underpinned by global policy leadership, necessary to implement a precision-based paradigm for innovation in cardiovascular medicine globally.
Current challenges in CVD research
Biology of heterogeneity in CVD
CVD heterogeneity is evident in the diversity observed in clinical presentations, underlying causes, and responses to treatments. This variability arises from individuals’ unique characteristics at the genomic (e.g., single-nucleotide polymorphisms) and exposomic (e.g., air pollution or pesticides) levels (32–37) (Figure 1A), underscoring the need to discover new mechanisms and drug targets to advance precision cardiovascular medicine (38). Multiomics approaches have provided more detailed explanations of how such genomic or exposomic signatures contribute to the development of complex diseases (32–36, 39). Recent evidence suggests that human pathobiology is also heterogeneous at multiple levels, from cellular responses to disease mechanisms and clinical features (Figure 1A). Such heterogeneity presents major challenges that have hindered our ability to gain a comprehensive understanding of disease mechanisms and develop more effective therapies for complex human diseases, which have proven to be even more intricately mechanistic than traditionally thought (35, 38, 40–44). Between 1954 and 1977, epidemiological findings of the renowned Framingham Heart Study established risk factors for coronary heart disease and stroke, with a particular emphasis on dyslipidemia and hypertension (45–47). Notably, elevated low-density lipoprotein cholesterol (LDL-C) became a viable target for lowering the incidence of CVD events primarily via statin therapy. Despite the efficacy of statins in reducing LDL-C levels and the incidence of major CVD events, a significant residual risk remains (48).
Figure 1

Figure 1. Precision cardiovascular medicine as a solution to heterogeneity in cardiovascular diseases (CVDs). (A) The heterogeneity in CVDs arises due to many different factors and manifests at multiple levels. (B) A multipronged systems approach leads to precision medicine. On one front, the integration of multiomics data using network medicine techniques unveils molecular pathways and disease biomarkers. Simultaneously, an artificial intelligence (AI)-powered approach utilizes clinical data to offer translational endpoints, such as patient stratification and the development of precision drugs and therapies. Together, these approaches hold the potential to significantly enhance patient outcomes.
Clinical, epidemiological, and genetic evidence also points to the presence of phenotypic heterogeneity within CVDs, encompassing conditions such as myocardial infarction, angina pectoris, hypertrophic cardiomyopathy, and heart failure (49–54). For example, genomic mutations in cytochrome P450 2C (CYP2C9), solute carrier anion transporter family 1B1 (SLCO1B1), and adenosine triphosphate (ATP)-binding cassette super-family G member 2 (ABCG2) impact the response to drugs such as statins (55, 56). Additionally, interacting pathologies can lead to CVD, including patients with a predisposition to venous or arterial thrombosis—there is even evidence of familial clustering in some cases, leading to inherited hypercoagulable states. Genetic, environmental, and other phenotypic modifiers can also contribute to this group of conditions, making the diagnosis and management of these disorders particularly challenging (57). Many pathways that contribute to human diseases influence disease severity by interacting within a large and complex biological network of genes, proteins, and signaling pathways.
In parallel with clinical evidence, findings from single-cell RNA sequencing datasets have suggested that each cell type associated with CVD, such as macrophages, represents a heterogeneous population (58–66). The balance between subsets of immune cells (e.g., pro- vs. anti-inflammatory macrophage subpopulations) in a local microenvironment, such as the atherosclerotic plaque, may determine the risk of disease progression or the onset of clinical complications (e.g., myocardial infarction) (67, 68). As various new technologies capable of unbiased screening for target discovery continue to emerge (e.g., plasma proteomics of clinical samples and single-cell RNA sequencing of cells within cardiovascular tissues or blood), subsequently generated datasets become larger and more complex (69, 70). To identify promising new targets from large datasets analyzing complex factors, we must involve analytic platforms capable of processing the vast amounts of data generated (71).
Complex pathobiological mechanisms for CVD
CVDs are complex entities shaped by multiple factors. It has become increasingly clear that simply focusing on a single molecule or pathway does not allow a comprehensive understanding of the complex interactions and interdependencies among molecules and pathways acting within a larger biological system. Despite their phenotypic diversity, CVDs exhibit common underlying pathophysiologies. This is seen in conditions such as atherosclerosis, which is shared among vascular diseases such as coronary and peripheral artery diseases (CAD and PAD, respectively). The progression of atherosclerosis, however, involves different components of the immune system. CVDs—including heart failure, arrhythmias, hypertension, cardiomyopathies, and thrombosis/embolism—may exhibit varying degrees of complex pathobiological mechanisms that involve factors such as immune response, lipid metabolism, neurohormonal activation, structural variations, sympathetic nervous system modulation, and endothelial dysfunction (72).
Along with lifestyle factors and environmental factors, genetic heterogeneity also plays an important role in complex disease phenotype. Modern technologies such as whole-genome, whole-exome, and targeted sequencing provide sequence information of DNA bases, giving insights into genetic variation. Both allelic and locus heterogeneity can contribute to the development of CVD (43). Large biobanks and databases of multi-ancestry genetic studies now play a crucial role in advancing our understanding of CVDs and thus aid the precision medicine approach (73, 74). While the traditional linear biology approach has been valuable for dissecting individual mechanisms and driving critical discoveries, it does not allow a comprehensive understanding of complex disease pathobiology (75). Preclinical research is powered by many studies built on cell lines and inbred mouse strains (76, 77). The availability of these tools has enabled many mechanistic advances but has failed to capture the heterogeneity observed in the patient population. The use of large animal models and outbred mice could be an important consideration for a systems biology approach. Furthermore, the use of human primary cells to capture the heterogeneity of cells (68) could bring new insights into factors governing inflammation and disease progression. Multi-organ-on-a-chip technologies could be used to mimic the complex cross-organ interactions that regulate cardiovascular diseases (78, 79). These approaches could be bolstered by the utilization of patient-derived induced pluripotent stem cells (iPSCs) reprogrammed into diseased cell types (e.g., endothelial cells, cardiomyocytes, and macrophages) which in turn could be used to perform clinical-trials-on-a-chip for screening patient specific drugs (80, 81). Developing new treatments informed by a holistic view of a complex biological system requires a transition toward a more integrated approach, involving unbiased omics data at multiple levels (e.g., epigenetic, transcriptomic, and proteomic) from the same samples. The identification and understanding of the underlying processes that govern the clinical outcomes are fundamental to the success of precision medicine.
Long navigation time, high costs, and low success rates for drug development
Other major challenges in cardiovascular medicine include the timelines and costs for target discovery and drug development. Estimated development times for new drugs, from target discovery to launch, range from 5 to 20 years—averaging at 9.1 years for innovative medicines (e.g., first-in-class drugs) (82, 83). However, development times for RNA-targeted therapeutics are generally shorter (82, 84, 85). During the COVID-19 pandemic, open science approaches that mobilized the sharing of data and ideas between academic institutions, the pharmaceutical industry, and governmental institutions, helped accelerate the development and implementation of RNA therapeutics (86, 87).
Low success rates of new drugs in clinical development present another major hurdle. Approximately 90% of drugs fail between their entry into phase I trials and regulatory approval (88, 89). If preclinical drug candidates are included, success rates fall below 10%. The most common cause of failure is the lack of anticipated effects, with the exception of genetic disorders, followed by toxicity/side effects, poor pharmacokinetic parameters, and poor overall clinical development strategies (88–91). For example, hidden drug cardiotoxicity can lead to the discontinuation of clinical trials as well as the withdrawal of drugs post-approval (92). The conventional drug discovery approach that targets a single causal factor also shows limited effectiveness in finding new drugs for complex human diseases, as they involve multiple and overlapping molecular pathways and pathologies. This is, in part, due to this approach relying on a simplified hypothesis for a drug target. Such simplistic hypotheses are often tested using monoclonal cell lines and inbred mice, models that do not fully recapitulate the heterogeneity of complex chronic diseases in humans. Phenotype-driven drug discovery, as opposed to target-based strategies, can address diseases for which mechanisms remain incompletely understood and has been widely used in the pharmaceutical industry (93). Network analysis-powered prediction of the potential impact of each candidate target in human disease may also limit the failure of new drugs at the clinical development stage to a certain extent, owing, in part, to its ability to predict off-target adverse effects (94–96). The benefits of such a comprehensive systems approach to drug discovery ultimately lower the overall costs of development and yield a more efficacious and safe therapeutic agent. These approaches can predict the toxicity profiles of drugs, which in turn avoids the selection of compounds that may fail in later drug development stages due to harmful effects (97, 98).
Following the development of numerous drugs targeting previously identified causal proteins, the need to assess the potential ability of a novel target to be modulated positively or negatively by treatments, colloquially known as “druggability,” remains a key challenge in the development of new drugs (99, 100). Fortunately, technological advances have provided solutions to this challenge. The accumulation of biological and chemical data and the rapid evolution of high-performance computing have enabled the development of various computational strategies, including AI, and helped to design new, effective compounds or predict the potential effects of existing drugs via phenotypic screening. The use of such new technologies and novel computational approaches in drug discovery has opened possibilities for making traditionally undruggable targets druggable (101–103). New platforms, particularly those focused on RNA-targeted therapeutics, also offer precise methods for modulating previously undruggable targets while also reducing development timelines and costs (84, 104–106).
Precision cardiovascular medicine: through systems biology
The essence of the solution to these challenges is the concept of precision medicine (Figure 1). The National Institutes of Health and the Food and Drug Administration of the United States define precision medicine as an innovative approach that considers individual differences among patients. The goal is to use the right treatments in the right patients at the right time. Each individual possesses unique genomic characteristics, experiences distinct exposures (i.e., environmental factors), and exhibits various combinations of traditional risk factors (e.g., dyslipidemia, hypertension, diabetes, lifestyle, diet, and sex). Moreover, the same stimuli and therapies may elicit heterogeneous responses in different individuals. Considering the specific characteristics of each patient to tailor medical care and interventions with the goal of optimizing treatment effectiveness and minimizing adverse effects is key to precision medicine (107). This necessitates an approach that enables comprehensive molecular profiling, fosters integration to comprehend complex interactions among diverse components, and facilitates the development of predictive models for diseases and biological processes. Systems biology serves as a valuable framework for fulfilling these objectives: it is an interdisciplinary field where the central tenet is that the behavior of a biological system as a whole arises from the complex interplay between its constituent parts (leading to emergent system properties), which cannot be fully understood by studying them individually (108, 109). Below we discuss different aspects of systems biology and potential solutions to challenges in CVD research.
A systems approach to unraveling the biology of heterogeneity
As discussed, CVD arises from diverse factors affecting complex molecular networks. Such complex mechanisms imply that a traditional reductionist approach to exploring a single cause of disease in the average population, testing a linear hypothesis focused on a single target, and developing a “one-size-fit-all” medicine is overly simplistic (75) and may account for reduced efficacy in a substantial group of patients for which the approved drug has been developed (110). This major challenge has driven our efforts to develop a more holistic, integrative systems approach involving unbiased omics analyses, bioinformatics, and network science to establish precision medicine (Figure 1B) (38, 40, 111, 112). The generation of large amounts of biomedical data, or “big data”, ranging from clinical information in electronic health records (EHRs) to the molecular measurement of analytes using different omics platforms (e.g., genomics, transcriptomics, proteomics, and metabolomics) has also prompted the need for analytical frameworks that can holistically interrogate disease pathobiology. Studying CVDs using multiple omics modalities also requires a systems approach because of the involvement of not only complex tissues, including the heart, vasculature, and valves, but also various common underlying mechanisms—termed endophenotypes—including inflammation, immunity, thrombosis, fibrosis, and calcification.
A systems approach enables the construction of integrated models through the concurrent analysis of entities from different omics layers (e.g., genes, proteins, and metabolites) involved in each disease (Figure 1B). Combining different interacting units (e.g., genes and metabolites) into networks enables the identification of key molecular components and the nature of their interrelationships (e.g., regulatory or co-expression) (112). Similarly, AI and machine learning can pinpoint the key combinations of genomic features that are predictive of disease progression (Figure 1B). A systems approach thus enables simultaneous identification of biomarkers for disease progression and potential drug targets. This includes assessing off-target effects and mechanisms of action, enhancing the translational value of these investigations. Over the past two decades, systems-based research has been effectively applied to cellular systems to reveal a wide variety of emergent biological functions. This approach has complemented and addressed the limitations of the reductionist paradigm prevalent in biomedicine, especially following the influx of high-throughput data during the post-genomic era (109, 113–116). The incorporation of clinical data and disease etiology using AI and an advanced systems approach also allows for better patient stratification into groups based on drug responsiveness, a central tenet of the precision medicine approach (Figure 1B).
Network medicine for multiomics data integration
Networks form the cornerstone of the systems approach in biomedicine (109, 117) and have been used to analyze rich omics data generated in the past few decades. Being multifactorial, complex chronic diseases, CVDs have benefited greatly from the application of systems and network medicine (38, 118, 119). Currently, a myriad of network-based approaches are readily applicable to a vast array of high-throughput molecular, interaction, and ontological data that are publicly available. These approaches are instrumental in inferring key molecules, subnetworks, and pathways related to CVD, providing invaluable information that may help to better identify novel drug targets for preclinical and clinical testing (120). Network methods have been built and implemented for gene regulation (121–125), protein–protein interactions (126–131), and metabolic interplay (132) to shed new light on the drivers of CVDs. Genetic risk loci identified by genome-wide association studies (GWAS) on features such as the PR interval (133), QRS duration (134), and atrial fibrillation (135, 136) have been studied using gene networks. The structural properties of omics-derived networks help in identifying CVD phenotypes that correlate with network features such as functional modules in CVD (137, 138), congenital heart disease (139), cardiac development, hypertrophy, and heart failure (140). Network medicine has helped identify candidate disease genes for CAD (121, 141) or CVD in general (142). Analyzing proteomic data with the assistance of network-based approaches has revealed the global impact of macrophage activation in vascular disease (94) and helped identify the mechanisms behind indoxyl sulfate-triggered pro-inflammatory macrophage activation (95). Similarly, pathway network analysis allows us to study macrophage activation through PCSK9 (143) and in vein graft disease (144), vascular calcification (145), and rheumatic heart valve disease (146). Simultaneous analysis of global transcriptomics and proteomics of calcific aortic valve disease has revealed important associations with various inflammatory diseases (129). Moreover, horizontal integration of the same omics datatypes measured under different conditions or sources reveals correlated features in various layers. For example, integration of proteomics from valvular interstitial cells—either in two-dimensional (2D) cultures on flat surface or three-dimensional (3D) models on hydrogels—with their extracellular vesicles showed correlated proteins relevant to calcification (147). The unbiased proteomics and systems biology of abdominal aortic aneurysms of mouse models and patients revealed potential novel mechanisms (148). Integrative omics analyses have also been used to study blood pressure regulation and hypertension (149, 150). Parallel to the advances on the omics front, the use of systems pharmacology in CVD has gained prominence (151–155).
The systems approach to drug discovery
Systems biology—involving unbiased omics screening, bioinformatics, and network analysis—has facilitated the discovery of both diagnostic and prognostic biomarkers for CVD. A key strength of this approach is its ability to discover promising targets with improved accuracy in predicting drug efficacy in patients (Figure 2) (38, 94, 95, 144, 156–158). This approach has also enabled us to identify potential targets with a higher likelihood of clinical significance through the integration of different omics datasets and holistic analysis of the disease. Using a systems biology approach, we can find connections between drug target genes and a potentially beneficial clinical outcome (94, 112, 159, 160) (Figure 3). Analyzing which proteins have showcased a shared tendency to change within the human interactome predicted potential regulators of macrophage activation (161). Protein network databases also help to identify proteins closely associated with a node or disease target by “proximity” that can be targeted by new or repurposed drugs to interfere with the disease network. Investigating pathways, key driver genes (162), and network modules associated with a potential drug target can provide key information to researchers by allowing them to choose appropriate cell culture methods, animal models, or even patient stratification in clinical trials.
Figure 2

Figure 2. A multilayer systems approach to target discovery and drug development. The hypothesis-driven, reductionist approach may have contributed to low success rates of new therapies based on conventional, basic science-driven clinical development. A systems approach involving unbiased omics, followed by bioinformatics for target prioritization and network medicine-assisted prediction of clinical impact, may facilitate the process and increase the success rate of identifying new targets. Comprehensive in vitro and in vivo experiments substantiate new concepts. Such models enable earlier implementation of drug design than conventional models.
Figure 3
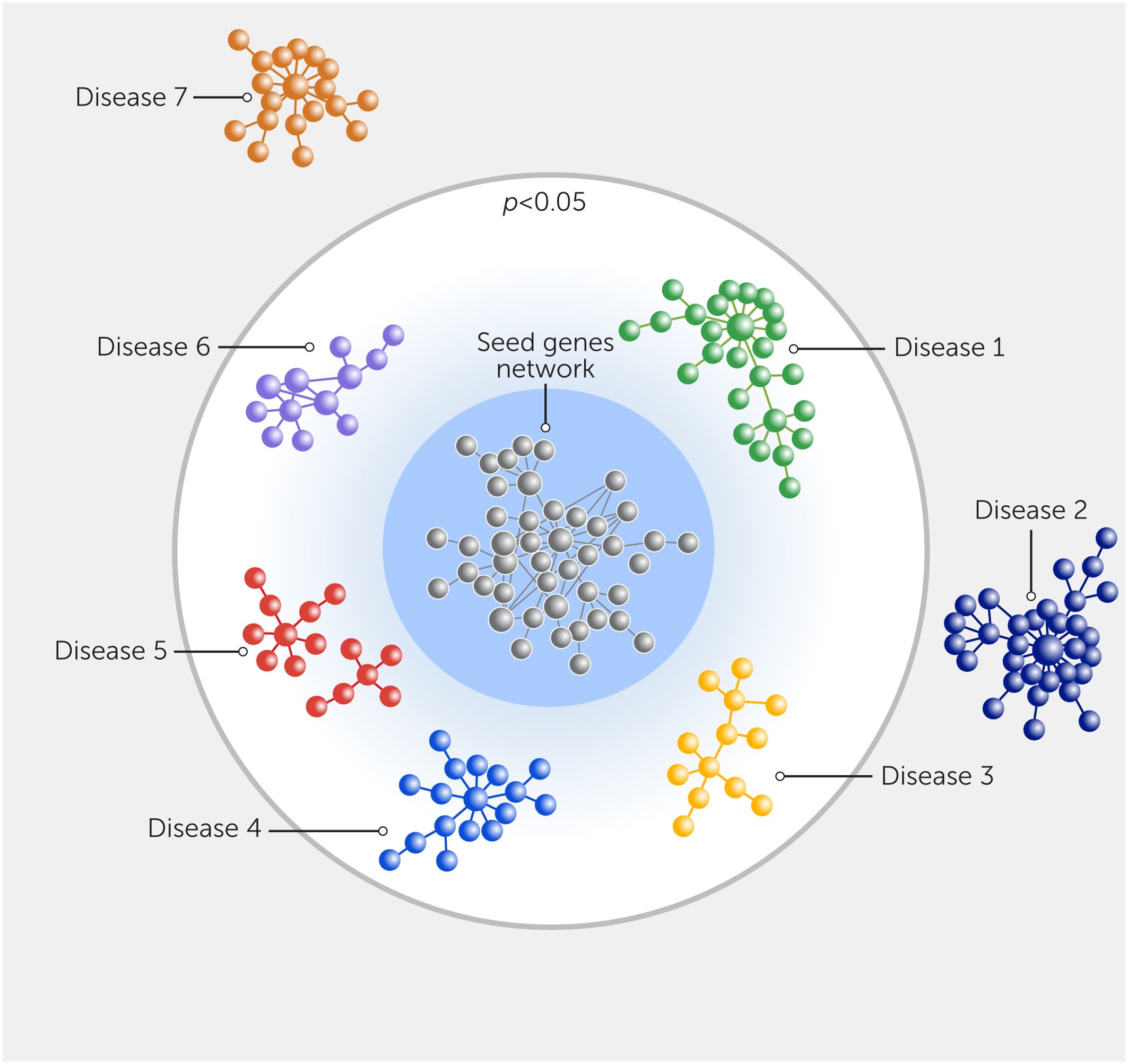
Figure 3. Network prediction of the clinical impact of seed genes on various diseases. The network proximity of the seed genes from analyses (gray), which could be obtained from differentially expressed genes or proteins and various disease gene modules obtained using databases. The p value indicates the significance of proximity to the given disease module and other disorders, as compared with random expectation.
Future perspectives in the systems approach to cardiovascular medicine
One of the important promises of systems medicine is its focus on the patient rather than the disease. Specific genes and individual disease-causing mutations can contribute to an individual’s apparent monogenic CVD phenotype, such as cardiomyopathies. However, in some cases, “additive” modifier genes may also play a role, paving the way for precision genetic medicine (163). For example, a previous study aimed to identify genetic drivers of dilated cardiomyopathy, a diagnosis of exclusion among cardiomyopathies (164). After examining 51 curated genes, they identified 19 that showed high evidence but could only explain a minority of cases, suggesting the need for further studies to unearth the mechanism of disease development. We predict, however, the realization of precision medicine goals at scale in the next decade. The implementation of network-based approaches has accelerated research on integrative single-cell omics (165) and spatially resolved omics (166), which will further unlock the heterogeneity in complex diseases (Figure 1) (38, 111, 167). In the context of CVD, identifying the degree of heterogeneity of cell populations in complex tissues such as atherosclerotic plaques, calcified aortic valves, or cardiac muscle can lead to important insights into pathobiological mechanisms. For example, single-cell RNA sequencing has helped identify the diversity of the cardiac cellulome (168). Studies that combine network medicine and machine learning methods can be used to fine-tune our understanding and help identify a more accurate representation of the regulatory underpinnings of cellular heterogeneity (68, 103, 129–131, 147, 169). Moreover, the transfer of fundamental concepts across fields, such as a newly proposed statistical mechanics framework for single-cell biology (170), can further accelerate advances in systems medicine. As discussed, the integration of multiscale omics data may also facilitate the development of precision therapies. Finally, recent advances in harmonizing network biology methodologies with the power of machine learning (171–173) will soon come to full fruition, as high-resolution molecular data are increasingly converging with corresponding clinical and EHR data from individuals.
Innovative technologies for driving precision cardiovascular medicine
Proteomics headlines the multiomics universe of CVD research
Omics technologies, which offer an unbiased survey of multiple genes and proteins, have significantly increased the likelihood of identifying potential therapeutic targets. This process can be effectively complemented with targeted proteomics and the more recently defined proteoforms resulting from post-translational modifications. Such an approach can facilitate successful target discovery and clinical translation (Figure 4), as discussed later in this section.
Figure 4
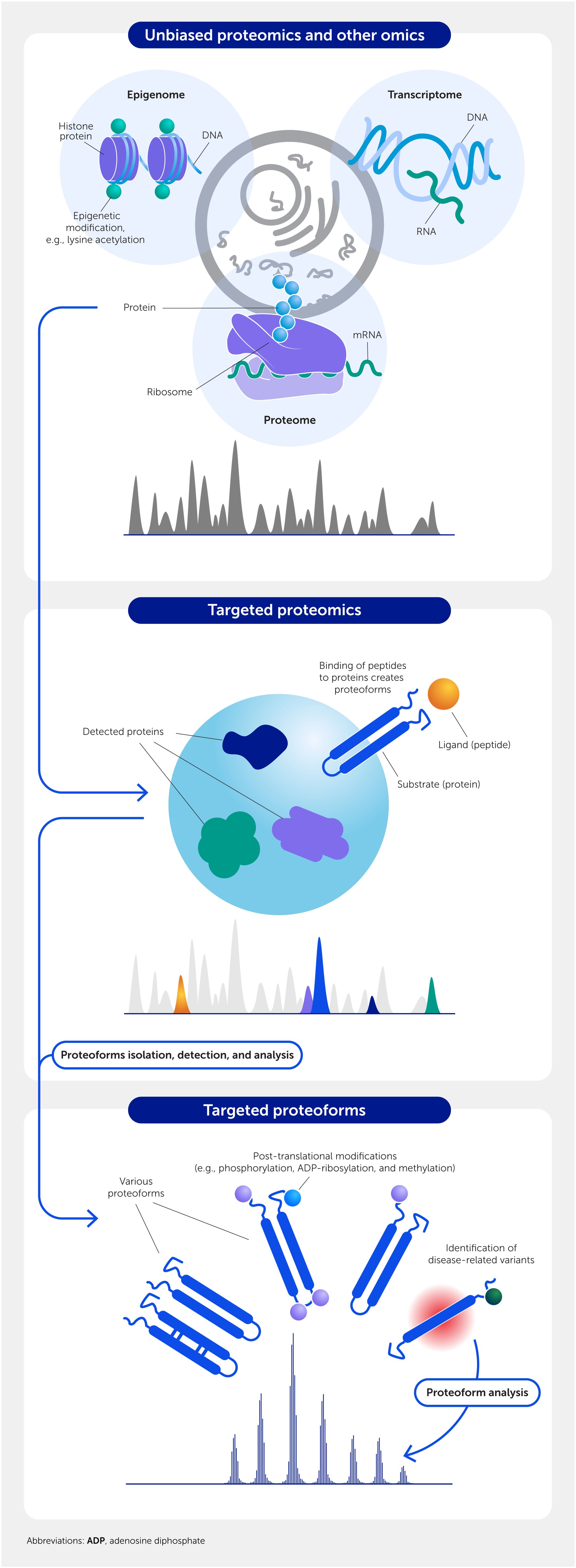
Figure 4. Drug-target discovery depends on multiple omics strategies. Unbiased screening of the epigenome, transcriptome, and/or proteome identifies candidate targets that can be validated using targeted mass spectrometry/proteomics. Ultimately, a specific proteoform (post-translationally modified form) of the protein may be the best target in some cases.
The high demand for continued discovery of additional therapeutic targets has driven the development of global, unbiased platforms such as epigenomics, transcriptomics, proteomics, and metabolomics. In particular, mass spectrometry-enabled protein research has long been recognized as a promising means to identify novel biomarkers and therapeutic targets for CVD (174–176). Today, mass spectrometry is a mainstay not only for proteome profiling (177) but also metabolome profiling (178, 179). When combined with other omics approaches, such as epigenomics (179) or transcriptomics (180), it provides solid foundations for systems biology and multiomics data integration strategies (Figures 1-4) (38, 167, 181).
Proteins are one of the major determinants of the cellular phenotype, driving initiatives such as the Human Proteome Project to facilitate translational research to improve overall human health (182). Unbiased proteomic approaches are consistently used to identify molecular drivers of CVDs, such as coronary heart disease (183), abdominal aortic aneurysms (148), and calcific aortic valve disease (129). In the last example, transcriptomics and proteomics were used to distinguish fibrotic and calcific regions from non-diseased regions of aortic valve leaflets but reported only a weak correlation between the quantified transcripts and proteins (129). These findings emphasize that protein abundances may not necessarily occur in proportion to their transcript abundances (184, 185). Moreover, tissues comprise various cell types such that “bulk RNA and proteome” data provide average signals, thereby eliminating the opportunity to glean potential “disease–driver subpopulations.” While single-cell transcriptomic technologies have been successfully implemented to reveal the extent of cellular subpopulations and heterogeneity in tissues (144), single-cell proteomics (186, 187) is still too recent a technology, requiring extensive expertise to yield a similar widespread implementation.
Targeted proteomics is already valued in a clinical setting, namely, to monitor steady-state kinetics of candidate LDL-C lowering targets such as apolipoprotein B (APOB), cholesteryl-ester transfer protein (CETP), and PCSK9 in cardiovascular outcome trials (188–190). Innovations in targeted mass spectrometry technologies enabled tracer kinetics studies in humans that captured the complex metabolic profiles of several high-density lipoprotein (HDL)-associated proteins, supporting the notion that HDL is a heterogeneous lipoprotein class consistent with its multiple functions (191, 192). These findings underscore that effective CVD drugs may require targeting a subpopulation with distinct functions rather than the entirety of a given molecule or cell class.
Proteins themselves comprise various isoforms—known as proteoforms—that may result from genetic variants, messenger RNA (mRNA) splice variants, and post-translational modifications, of which only one form may be causal to the disease of interest. Therefore, the proteomics community has initiated the Human Proteoform Project, an ambitious endeavor to generate a reference set of proteoforms for the human genome (193). Although mass spectrometry is a central technology supporting this initiative, it is, in essence, a multiomics endeavor (Figure 4) (193).
As first predicted over 20 years ago, mass spectrometry-enabled proteomics is providing CVD researchers multiple avenues through which to identify therapeutic targets. In all likelihood, the next CVD breakthrough targets may be identified using omics. Before its arrival on the market, however, the methodology will require extensive validation studies that, in part, may very well entail one or more additional proteomic technologies.
Single-cell technologies to tackle the complex biology of cellular heterogeneity
As discussed, disease heterogeneity is a major challenge in cardiovascular medicine. For example, statins reduce the risk of acute complications such as myocardial infarction in many, but not all, patients. This can be attributed to factors such as the potency and pharmacodynamics of different statins, as well as patient heterogeneity. This may also result from different patterns of heterogeneity of atherosclerosis-associated cells (e.g., macrophages) among patients. Evidence has linked sustained pro-inflammatory activation of macrophages with vascular disorders (67, 194–196). An earlier paradigm of macrophage heterogeneity proposed a pro-inflammatory M1 phenotype and an anti-inflammatory/pro-resolving M2 phenotype (197, 198). More recent evidence (our own included) however, suggests that macrophage heterogeneity is more complex than the M1/M2 dichotomy, and involves more subpopulations (67, 199–202). While the overall balance of macrophage subpopulations may regulate disease mechanisms or severity, traditional assays only examine average levels of gene or protein expression in the entire population of cells (e.g., Western blot analysis and bulk RNA-sequencing) and cannot assess how individual cells behave. This challenge has driven the development of various platforms for single-cell analysis and their integration (Figure 5).
Figure 5
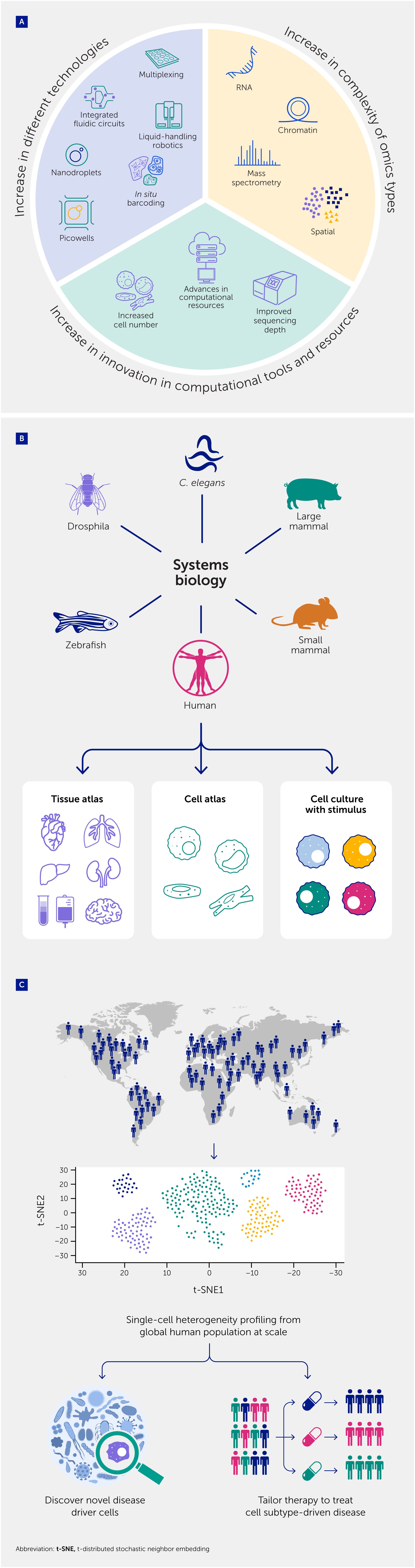
Figure 5. Integration of single-cell analyses for precision medicine. (A) An array of technological advances have occurred in platforms used for single-cell analysis, together with advances that have increased the complexity of omics types analyzed, and innovations in computational tools and resources. (B) Systems biology studies in animals and humans are now translating these advances into multi-tissue, single-cell atlases to provide in vivo landscapes of cellular heterogeneity. (C) Further extension of single-cell technologies to profile populations at scale could shape the future of biomedical research, establishing innovative diagnostics/ biomarkers and tailored therapies for diseases driven by specific cell subtypes.
This challenge has been the catalyst for rapid and expansive development of single-cell technologies that enable deeper sequencing in more cells, platforms for simultaneous surveying of various omics layers in cells, and computational and bioinformatics infrastructure for innovative data analyses (Figure 5A). Technologies such as droplet-based, well-based, and sequential barcoding platforms can be chosen based on the underlying application. Single-cell analysis has extended beyond profiling RNA expression levels at the single-cell level with the ability to map surface expression of receptors (203) and chromatin accessibility (204). While these omics datasets can either be individually mined, they can also be paired with mRNA expression (205) by these cells to provide truly integrated multiomics characterization (206). Furthermore, recent advances in single-cell proteomics (207, 208) powered through mass spectrometry allow unbiased characterization of the proteome at the single-cell level. Spatial transcriptomics and spatial proteomics have also become widely available platforms that provide critical information relating to the spatial disposition of cellular heterogeneity. Leveraging these technologies allows for the construction of multilevel spatial multiomics maps through disease progression within the heart to identify signaling pathways specific to different cell types (59, 130, 209, 210). Assays that characterize different types of omics at single-cell resolution allow for the simultaneous measurement of epigenetics, transcriptomics, and proteomics. These include single-cell assays for transposase-accessible chromatin (ATAC)-sequencing for chromatin accessibility, single-cell profiling of histone modifications (211), spatial transcriptomic profiling (e.g., Slide-seq) (212), and surface receptor profiling (e.g., cellular indexing of transcriptomes and epitopes sequencing; CITE-seq) (213). Recent developments have expanded even into the realm of single-cell metabolomics, which provides opportunities to evaluate substrates and metabolites within the same cell (214–217). Such approaches will significantly advance integrative single-cell omics research.
The wide range of available, free software packages that can be used to analyze these datasets has also removed barriers to entry for many researchers, enabling them to embrace these approaches to address their specific research needs. This transition has coincided with the decreasing cost of cloud computing and the secure computing capabilities provided by research institutions and private companies that allow rapid, cost-effective processing of these large datasets (Figure 5A).
The rapid utilization of single-cell RNA-sequencing technologies (218) has enabled the construction of a wide range of single-cell atlases (219). Multi-tissue cell atlases of various model organisms have provided an important understanding of the in vivo landscape of cellular heterogeneity (220–222). Human-centric atlases have mapped specific organs that have been instrumental in identifying the various cell types and subtypes that make up an organ (219, 223, 224) (Figure 5B). Specific cell atlases discern how the same cell identified in multiple tissues has different underlying transcriptional signaling (58, 62–64, 225–227). Furthermore, recent studies have deployed single-cell RNA-sequencing technology to identify novel cell subtypes that could be disease drivers within specific disease settings (130, 228). These studies help shed light on specific cellular signaling aspects that regulate cellular heterogeneity.
While unbiased single-cell omics datasets have become increasingly cost-effective, generating, annotating, and sharing them remain expensive and resource-intensive. Reproducibility and data access are important aspects of large omics datasets that have made major strides recently. Easy and accessible computational pipelines are now widely available (229, 230). Journal requirements that make detailed single-cell datasets publicly available also help extend their utility once generated, which will also facilitate comparisons of datasets to enhance reproducibility.
Importantly, using publicly available unbiased single-cell omics data, researchers can fine-tune their approaches and generate follow-up experiments using targeted approaches to either validate these findings in a larger dataset or perform hypothesis-testing experiments. In this regard, the development of methods to perform targeted single-cell mRNA sequencing significantly reduces the costs of sequencing and facilitates the sequencing of a large number of cells (231). Similarly, the utilization of high-parameter flow cytometry (232) as well as sequential staining of tissue sections are becoming increasingly attractive (232, 233). Large-scale panels of validated probes and antibodies allow for the rapid adoption of these platforms across a wide range of tissues.
The characterization of cellular heterogeneity through these approaches helps to identify key cell types within in vivo settings. However, future studies can also use both unbiased and targeted approaches within monoculture systems to evaluate cellular heterogeneity in response to classical stimuli (234–236). Past efforts have typically utilized bulk omics studies to identify heterogeneity in responses (94) but are not limited in their capacity to highlight how different cells within a monoculture system can respond differently to the same stimulus (Figure 5B). Single-cell RNA sequencing and single-cell ATAC sequencing (237, 238) will allow the identification of novel subpopulations within a single cell type.
While single-cell technologies have developed rapidly, a few key questions remain (239). It is of critical importance to leverage cellular heterogeneity information to identify new mechanisms that translate into the clinic. We also need to consider the contribution of a small subset of disease driver cells to disease progression. Another point of consideration is the temporal dynamics of measured proteins and genes and their relative contribution to chronic disease. Most importantly, we also need to ask how understanding macrophage heterogeneity can provide molecular bases for the development of new diagnostics and therapies (240, 241). Recent studies offer examples of how cell heterogeneity data can be translated into drug development (169). How can we associate the information of subsets of cells associated with CVD with high-risk patients? Further extension of single-cell technologies, such as high-content live cell tracking for longitudinal monitoring and histologic localization of high-dimensional single-cell data in disease tissues, may help to facilitate clinical translation (31, 242–245). The combined use of single-cell data and computational drug screening methods (68, 169), discussed below, may also lead to potential new therapies. These methods may help us develop new approaches in precision cardiovascular medicine (246), establish innovative diagnostics/biomarkers (247), and enable intelligent enrollment criteria trial design, focused data interpretation, and improved patient safety in clinical trials (248, 249) (Figure 5C).
AI supporting translational discoveries for complex CVD
Medical scientists have faced challenges in analyzing the massive biological and clinical datasets necessary to address the complexity and heterogeneity of human diseases. Exponential technological advances and their integration into basic science and clinical activities, such as omics and EHRs, have accelerated this trend. Over the last several decades, the field of AI has led to a major technological revolution that has already significantly impacted practically every aspect of the human experience, including medical research and practice (250, 251). However, the use of AI-powered technologies in medical sciences is not necessarily new. In the 1980s and 1990s, decision support systems, such as Health Evaluation through Logical Processing (HELP) or DXplain, assisted physicians through the diagnostic process (252, 253). However, with the increase in computational power and the availability of large volumes of data, AI has unveiled its extensive capabilities in the last decade.
The application of AI in cardiovascular sciences has focused on two main tasks: prediction and clustering (Figure 6) (254–256). Prediction tools are used to estimate future prognosis and survivability of CVDs, including heart failure (257–259) and cardiomyopathies (260–263). AI prediction has also been implemented successfully to assist diagnosis (264–266), especially by using medical imaging data (e.g., echocardiogram, computed tomography, or magnetic resonance imaging) (267–274). Although not as common as contemporary AI-based prediction technologies, unsupervised learning has been used to cluster patient populations into different phenotypes (275–278), aiding the design of more precise therapeutic paths. Further, a new AI tool called AlphaMissense, which builds on the protein structure prediction tool AlphaFold2 (279), can be used to evaluate specific genetic variations (e.g., rare missense variations)—addressing the previous “bottleneck” in the bioinformatic analysis and assignment of causality to link a particular candidate genetic variant to the phenotype.
Figure 6
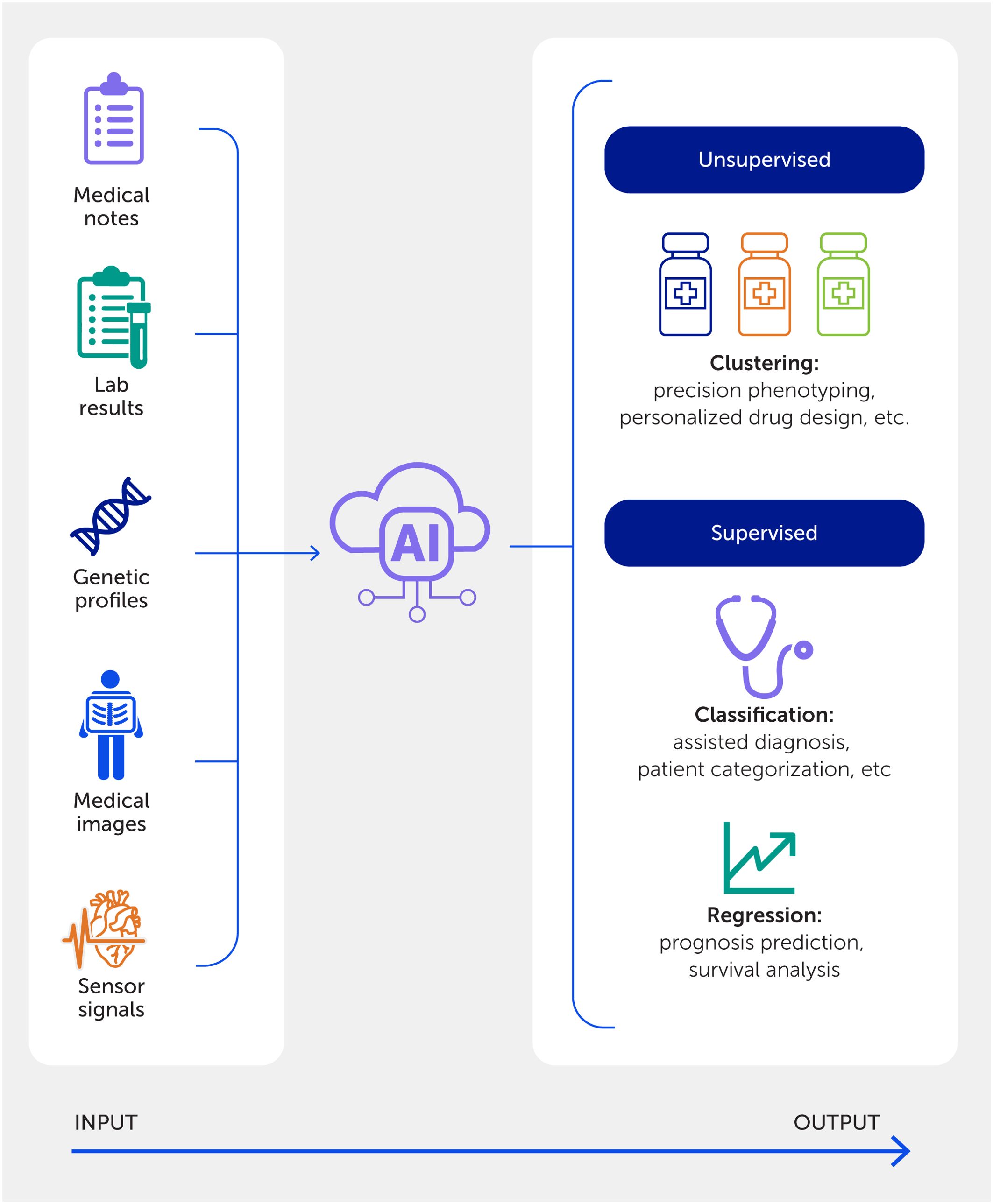
Figure 6. An artificial intelligence (AI)-powered precision medicine approach. An in-depth exploration of the AI workflow in medicine, highlighting diverse data inputs and delineating between supervised and unsupervised learning applications in clinical and research settings.
Considering differences among individuals, recent target discovery efforts have used large clinical data sources such as gene expression datasets associated with specific diseases and EHRs in addition to, or in place of, preclinical samples from cultured cells or animal models to address the biology of disease heterogeneity at the population level (71). The 21st Century Cures Act, initiated by the United States Government in 2016, promoted the use of clinical data sources such as the EHR (“real-world data”) in drug development and regulatory decision-making (280). Generative AI programs can autonomously create new content by learning patterns from existing data. AI’s role in changing the regulatory paradigms will involve improvements at all stages, from creating regulatory documents and designing the protocols to patient and site matching. The ability of AI to navigate huge datasets and construct detailed patient profiles based on demographics, medical history, and genetics to create “digital twins” can be used to simulate outcomes using virtual trials. These are the new AI frontiers: promises of refined therapeutics, improved patient care, and enhanced regulatory processes.
In the near future, integrating health records, clinical medical knowledge, and data provided by “smart” devices, such as phones and watches with AI technologies, will lead to unprecedented changes in our understanding of cardiovascular medicine (281). Technology companies such as IBM (282–284), Microsoft (285), Google (286), and Apple (287) have recognized its potential and have announced significant investments accordingly. The use of AI in continuous real-time monitoring, precision drug design, precision phenotyping, and the precise prediction of the development of CVDs represents a major breakthrough driven by new technology in the history of medicine.
Data science-powered drug development
The cost and time required for drug discovery have increased annually, presenting the pharmaceutical industry with major challenges in developing and marketing new drugs (82–85, 288). Another challenge is the low success rate of new targets progressing to clinical stages and achieving favorable outcomes in clinical trials (88, 89, 91). As discussed earlier, generating compounds for undruggable targets is a major obstacle in the development of innovative drugs (99, 100). On the other hand, valuable medical, biological, and chemical data have accumulated, and the performance of computers in handling “big data” has evolved. To save cost and time, computational approaches have increasingly contributed to various aspects of drug discovery. In particular, the computational exploration and design of effective therapeutic compounds are major emerging fields. Such innovative technologies may also help to make traditionally undruggable targets druggable (101).
One such technique uses the quantitative structure–property relationship (QSPR) and quantitative structure–activity relationship (QSAR)—a prevalent statistical approach that correlates molecular structure with properties or biological activity using quantifiable descriptors. These descriptors are often generated through density functional theory (DFT) (289), a widely applied quantum theory to calculate the electronic structures of atoms and molecules. Selecting the most relevant descriptors among them poses a significant challenge, as they encapsulate molecular characteristics responsible for the observed biological activity or chemical properties (290). Recent advancements have introduced novel QSAR methodologies that enrich the analysis of bioactivity. Nevertheless, QSAR models require rigorous testing and validation to assess their predictive accuracy and practical applicability (291).
One resource for drug identification is the Connectivity Map (CMap) (292). This database includes changes in many gene expression profiles (“signatures”) that occur when various compounds are exposed to various cell types. The CMap has been expanded to include over 1 million signatures using over 20,000 small molecules through the introduction of the L1000 assay, a low-cost, high-throughput, and highly reproducible gene expression profiling method (293, 294). The L1000-based CMap quickly identifies small molecules that modify gene expression signatures by either reversing or mimicking the changes caused by certain diseases. Therefore, such approaches have been widely used for rapid drug repurposing (102, 103, 295–297). This phenotypic screening as a counterstrategy to traditional target-based drug discovery has been successful in the development of “first-in-class” drugs (298, 299).
In target-based drug discovery, which generally favors the development of “best-in-class” drugs, the identification of compounds that interact with target proteins is a key task (300, 301). Drug–target interactions (DTIs) have been experimentally surveyed using high-throughput screening. However, the number of compounds that can be tested this way is limited compared with the theoretical number of drug-like compounds—estimated to range from 1023 to 1060 (302). It is therefore desirable to narrow-down candidate compounds using computational approaches. Computational DTI prediction can be divided into ligand-based, docking-based, and chemogenomic approaches.
Ligand-based approaches exploit the principle that compounds structurally similar to a known binder of a target protein are likely to interact with that protein in a similar manner. While such methods are rational and easy to follow, nothing can be predicted when there is no compound known to bind the target protein.
Docking-based approaches calculate the binding affinities between compounds and target proteins by simulating their 3D structures. Although this approach can evaluate interactions with any compounds, it requires knowledge of the in vivo structure of the target protein. The prediction thus becomes more difficult for compounds that interact with membrane or receptor proteins owing to their complex, flexible structures.
Chemogenomic approaches utilize the compound’s physicochemical features, such as molecular fingerprints, and the protein’s genomic features, such as amino acid sequences. Machine learning models learn the pattern of these features required for the interactions by using known DTI datasets, and then the model predicts whether an unknown compound indeed interacts with the target protein. This approach has attracted attention recently because it overcomes the inherent disadvantages of the ligand-based and docking-based approaches (303). Various frameworks employing classical machine learning methods, such as support vector machine or random forest approaches, and advanced techniques, such as deep learning (DL), have been proposed as relevant computational models and have already improved DTI prediction accuracy significantly (304–306).
Scientists have also attempted to computationally design novel compounds with desired molecular profiles (e.g., bioactivity, drug metabolism, pharmacokinetics, or synthetic accessibility). In this field, denoted de novo molecular design, various generative models based on DL architecture, such as the recurrent neural network, variational autoencoder, and generative adversarial network models, have emerged (307–309). Benefiting from the remarkable development of AI, these AI-powered generative models create feasible, plausible, yet entirely new compounds that have never been synthesized in the real world. These generated compounds can be used seamlessly as a new compound library for DTI prediction. Moreover, by providing the L1000-based CMap signatures to the aforementioned generative models, the design of novel compounds that induce desired gene expression signatures has been used in an attempt to expand the applicable range of L1000-based CMap (310, 311).
Notably, computational approaches are beginning to be used to predict the tertiary structure of proteins. A recently developed AI-based algorithm, AlphaFold2, predicts 3D protein structures from the amino acid sequences with high accuracy (312). AlphaFold2 may accelerate DTI prediction, especially via docking-based and chemogenomic approaches, because it can provide accurate protein structures whenever an experimental protein is unavailable and extract more structural features than the amino acid sequences can when used alone (313). Through such improvements, computational approaches will likely have an increasingly important role in compound exploration for drug discovery.
New therapeutic platforms: from proteins to RNAs
Most CVDs are currently treated with small-molecule drugs that are orally administered to bind to proteins contributing to disease mechanisms. As we discussed, however, some new targets are undruggable with conventional strategies. Therapeutic options other than small molecules include monoclonal antibodies against proteins, e.g., evolocumab and alirocumab targeting PCSK9 for familial hypercholesterolemia (314). While these types of drugs are effective and can overcome some druggability issues, targets are limited to cell membrane proteins or circulating proteins, and their production costs are high. Innovative technologies that enable targeting of undruggable targets include targeted protein degradation, such as proteolysis-targeting chimera (PROTAC) molecules that can degrade a target protein by controlling the ubiquitin–proteasome system. Compared with small molecules that block protein function but leave protein levels unchanged, small interfering RNA (siRNA) and PROTAC-based approaches can help modulate protein levels directly. While PROTAC technology has mainly been used for cancer targets, recent advances have extended its application to non-cancer diseases, particularly immune, inflammatory, and neurological disorders (315).
RNA-targeted interventions, a new class of innovative therapeutics, may overcome some of the aforementioned challenges (84, 104–106). Their advantages include (i) each gene of interest is potentially targetable by RNA therapeutics, whereas protein-targeted small molecules or antibodies can target only 0.05% of the human genome (316); (ii) manufacturing costs are lower than those of protein-targeted therapeutics; and (iii) development times are substantially shorter than those for conventional medicines. RNA interventions include antisense oligonucleotide (ASO), siRNA, clustered regularly interspaced short palindromic repeats (CRISPR)-based genome editing, aptamer, and mRNA vaccines (317–321).
The first ASO drug was fomivirsen, approved by the United States FDA in the late 1990s for the treatment of cytomegalovirus (CMV) retinitis (322, 323). Mipomersen, an ASO targeting apolipoprotein-B-100 mRNA, was the first RNA-targeted therapy approved by the FDA for a CVD—familial hypercholesterolemia (324). Inclisiran, an siRNA targeting PCSK9, has proved safe and effective for lowering LDL (by approximately 50%) and cardiovascular outcome trials are ongoing (325). Another developmental ASO, pelacarsen, is directed against lipoprotein(a) [Lp(a)], which is linked clinically with CVD, including aortic stenosis. Specifically, pelacarsen targets the production of apolipoprotein(a) [Apo(a)], a key component of Lp(a) disulfide-linked to apolipoprotein B100. Pelacarsen proved safe and lowered Lp(a) levels by up to 80% in phase 2 trials (326). Clinical trials of siRNAs that reduce both normal and mutated transthyretin (TTR), causing TTR amyloidosis, reported attenuated progression not only of the associated peripheral neuropathy but also cardiomyopathy (327, 328). In addition to these developments, the COVID-19 mRNA vaccines showed that delivering native or chemically modified (e.g., pseudo-uridine) mRNA by encapsulation in lipid nanoparticles is another potential option to treat various diseases, including CVD. Yet, all such methods are directed toward interacting with proteins.
We know from increased usage of next-generation sequencers that most of our coding genome is transcribed as RNA (329). Only a small percentage codes for proteins, leaving a majority of transcribed RNAs as non-protein-coding RNAs (ncRNAs). In addition to the well-known ncRNAs, ribosomal RNAs (rRNAs), and transfer RNAs (tRNAs), other regulatory ncRNAs have been identified and characterized in recent years, including microRNAs (miRNAs), circular RNAs, and long ncRNAs (lncRNAs) (330). Not surprisingly, the dysregulation of ncRNAs is linked to various CVD etiologies, and hence, these ncRNAs are being investigated as potential CVD diagnostic biomarkers or therapeutic targets (331).
Preclinical and clinical trials of miRNA-based therapeutics for CVDs are ongoing (332) while most projects on lncRNA-targeted therapeutics are still in the preclinical stage. LncRNAs are associated with many human diseases and many efforts are underway to develop technologies to target them therapeutically (333, 334). LncRNAs involve diverse modes of action, providing different opportunities to modify their functions (e.g., via siRNAs, ASOs, CRISPR/Cas9, small molecules). Some mitochondrial lncRNAs have reached clinical trials as cancer therapies (334). Accumulating preclinical evidence has implicated lncRNAs in the pathogenesis of various CVDs, including atherosclerosis, myocardial infarction, heart failure, and arrhythmias, providing molecular bases for their clinical applications as therapeutic targets or biomarkers (331, 335, 336). While lncRNA-targeted therapeutics have high potential, their clinical development is lagging. This may be due to our incomplete understanding of their mechanism of action, necessitating more mechanistic studies of each lncRNA. In addition, innovative computational methods should help elucidate their interactions with miRNAs, coding RNAs, and proteins. The combined use of such targeted systems approaches will help translate advances in lncRNA biology into clinical CVD medicines.
These novel modalities can also be partnered with a wide array of drug delivery strategies to maximize their effectiveness (337) and reduce off-target effects (338). These have been key partnerships for emerging modalities, such as RNA-targeted therapeutics (339). While these drug delivery methods remain unproven in the clinic, they have accelerated preclinical research by serving as powerful tools for in vivo intervention (95, 144).
Transforming cardiovascular medicine: innovative approaches and collaborative initiatives
Interdisciplinary and multistakeholder drug discovery drives innovation
Facilitating drug discovery and development for innovative precision medicine requires new paradigms. New technological developments can help solve specific technical limitations and promote scientific discoveries. These discoveries can be accelerated by models that integrate multiple innovative technologies to holistically address the biology of heterogeneity, identify promising drug targets, predict their clinical impacts, and design, generate, and test new drugs. Dynamic and close collaboration between biologists and data scientists is essential to establish fully integrated drug discovery research, as discussed above. Such seamless approaches also require innovative cross-sector collaboration.
One of the major obstacles is the large gap between target discovery research in academia and drug development in industry (Figure 7). Many ideas or targets identified in academic research do not bridge this gap for various reasons, including the lack of expertise and funding in academia and industry’s unwillingness to invest in early, high-risk projects. To solve these major challenges, several models of academia–industry collaboration have been established to merge the strengths of both sectors (89, 340–344). Establishing novel concepts by exploring uncharted territories and pursuing high-risk projects is a typical strength of academic investigators, while industrial scientists have specific expertise in drug design and development and are more strongly supported by infrastructure and financial resources (341, 342). Indeed, one study indicated that academia–industry collaboration showed higher clinical development success rates than those commonly seen in either academia or industry with no collaboration (89). Other types of collaborative arrangements include precompetitive research between pharmaceutical companies for sharing resources and expertise, and public–private partnerships (345–348).
Figure 7
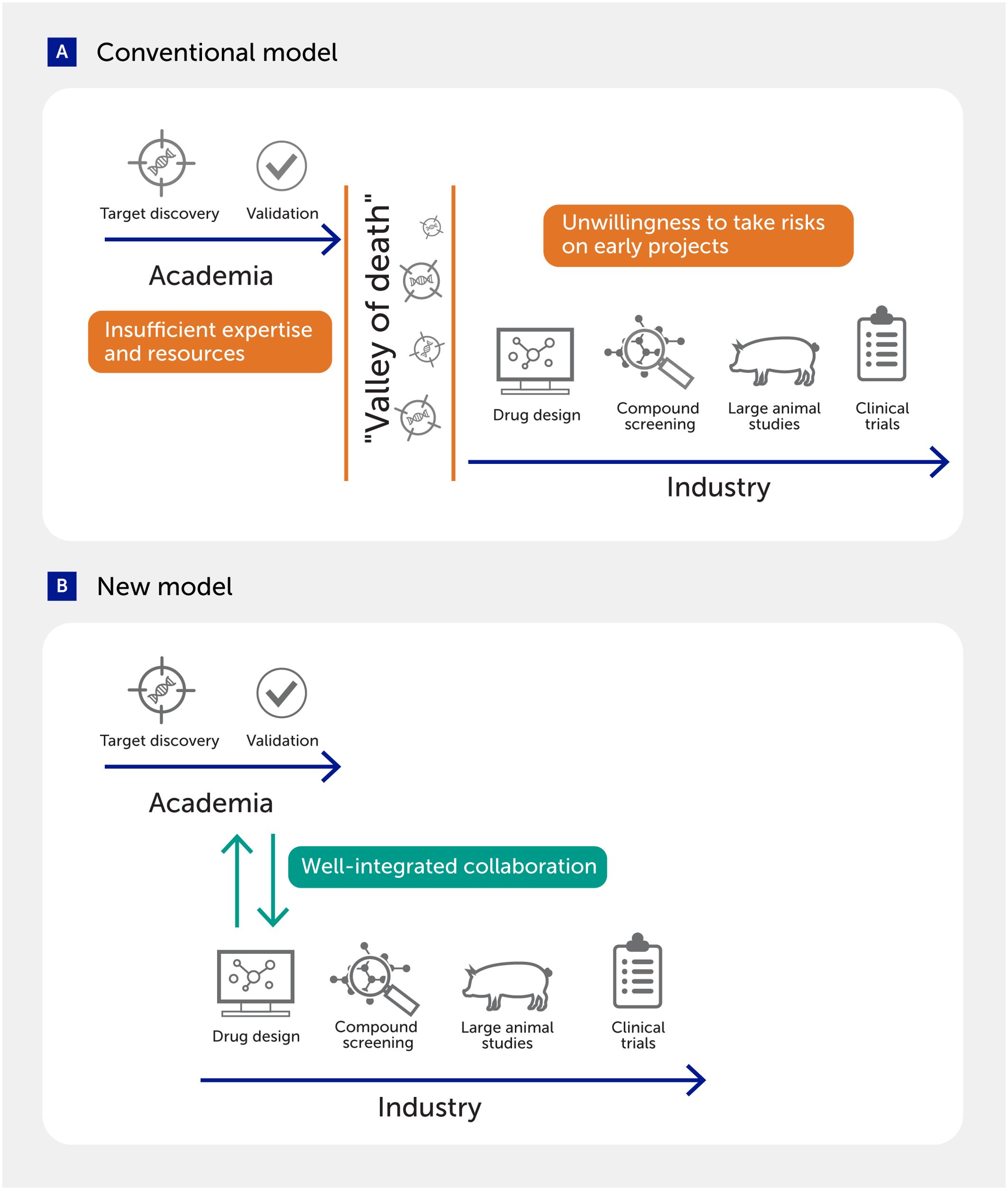
Figure 7. Academia–industry collaboration to fill the gap in drug discovery. (A) Several challenges often impair the transition of targets discovered in academia to drug development in industry. (B) New models that integrate actions in the two sectors may facilitate the translation of discoveries into the clinic.
Collaborative data science: key to precision medicine
As discussed, the major challenges in CVD have prompted precision medicine approaches that in turn necessitate new technologies; these disruptive innovations not only solve challenges, but also generate new concepts. The key, essential component in this synergistic relationship is data science (349). Among new technologies across various disciplines, the evolution of data science has been particularly rapid. It is now critical for us to recognize the importance of implementing this discipline and involving data scientists in every stage of cardiovascular medicine innovation, from basic science, discovery, and translational research to the clinical development of new therapeutics and ultimately their use in clinical practice. We must also recognize the diversity of data science as characterized by various subspecialties (e.g., biostatistics, bioinformatics, biophysics, network science, computational biology, and machine learning-based approaches), which enables the construction of a multidisciplinary data science team to cover a wide range of needs. More resources need to be allocated to support the development of data scientists at the institutional and government levels to bolster future biomedical innovation in both academia and industry. Finally, infrastructures that support interoperability between the multiple data sources involved are also vital to enhance the synergistic relationships between data science and cardiovascular medicine (350).
Lessons learned from COVID-19: are we ready for the next pandemic?
During the COVID-19 pandemic, over 770 million people were infected with the SARS-CoV-2 virus globally, leading to 7 million deaths (351). The scientific community came together to respond to the rapidly evolving demands that arose as a result. Public, political, and scientific awareness enabled resources to be redirected toward combating this global threat. This also triggered research interests in investigating the extensive, long-term consequences of viral infection on a large scale (352).
The United States FDA demonstrated flexibility and innovation during this time by modifying existing regulations to accelerate the approval process of life-changing medications for COVID-19 (353, 354). Similarly, the World Health Organization (WHO) played a central role in generating and distributing guidelines and tools to the global medical community (https://covid19.who.int). As a result, novel technologies, including mRNA vaccines and neutralizing antibodies (355, 356), were deployed with unprecedented rapidity to help reduce disease severity. Although these groundbreaking tools had already been in development (357, 358), the COVID-19 pandemic created the impetus to embrace these platforms to supplement conventional treatments. This new attention toward acute viral infection required the scientific, medical, and regulatory communities to restructure and shorten the drug development and implementation timeline (359). Governmental programs, such as the National Institutes of Health RECOVER program in the United States (360) (https://recovercovid.org/) to characterize post-acute sequelae of SARS-CoV-2 infection (PASC) syndrome or “long COVID” promoted the formation of multi-institutional and multidisciplinary nationwide collaborations by eliminating barriers to interactive science (361). Technologies developed through this global effort will be applied to counter other diseases, such as cardiovascular, pulmonary, and neurological disorders.
Knowledge gained in social, political, and clinical realms has shaped how our scientific community responds to worldwide challenges. The advent of new technologies has heightened the level of responsibility, as they allow the evaluation of interventions more quickly, more precisely, and at greater scales than was previously possible (362, 363). It will be essential to capitalize on the collaborations established during this crisis to address future pandemics successfully.
Revamping global healthcare policies to tackle the leading global cause of mortality
We have discussed that investing in innovative approaches and cutting-edge technologies will help develop treatments and interventions that improve CVD outcomes. However, we cannot afford to miss the forest for the trees. The “domino effect” of anthropogenic causes of mortality is the most pressing human health problem in modern times (364). Climate change leads to increased natural disasters, which, in turn, cause changes in food and water security. It also results in supply chain disruptions and population displacements. All of these consequences collectively strain the healthcare system. This strain exacerbates the impact of both lifestyle and environmental components on the development and progression of CVD. There is also a heightened risk of respiratory and vector-borne diseases due to these interconnected factors (365).
Concerted efforts applied to the population scale of the Millennium Development Goals and the Sustainable Development Goals (SDGs) are needed to increase public awareness of CVD and its risk factors (366, 367). Precision medicine approaches can be a huge asset to meet SDG 3.4, which aims to reduce premature mortality from non-communicable diseases through prevention and treatment. Prevention of these diseases should be emphasized by encouraging healthy lifestyles, tobacco cessation, and more nutritious dietary alternatives. Strengthening healthcare infrastructure, increasing equitable access to quality healthcare, reducing economic disparities (368), and promoting unanimous support to the WHO’s Global Action Plan for the Prevention and Control of NCDs (369, 370) are necessary to, in some measure, mitigate this so-called “disease of civilization” (371).
Political imperative to ensure global equity
The COVID-19 pandemic exposed the vulnerabilities of global healthcare systems: the lack of cooperation between governments and the private sector, unequal distribution of medical resources (including vaccines), and the neglect of needs in the global south. In the context of CVD, neglect of the disease burden in low-income and lower-middle-income countries (LICs/LMICs) and excluding such populations from research (including clinical trials) has led to major healthcare crises (372). Even though the disease burden is high in LICs and LMICs, these countries’ contributions to the global research output are minimal in part due to limited research capacity (373). Moreover, at present, global responses to the major global health threats from both noncommunicable and infectious diseases are hampered by geopolitical division, conflict, and power imbalances. (374) Achieving global health equity requires multisectoral, multifaceted, and multistakeholder engagements (375) through the emergence of global frameworks such as the WHO pandemic agreement adopted in May 2025 (376). While pandemics are temporary, continued cooperation and dedication of financial resources toward research and outreach are critical to mitigate future calamities. Effective global leadership is contingent upon the constructive global healthcare discourse among all member states, alongside proactive contributions from intergovernmental institutions. For this to happen, major policy stakeholders must recognize their lack of attention to the CVD burden and the insufficient healthcare infrastructure of the Global South, consequently amplifying their struggles. Concerted efforts addressing the issues plaguing individuals living in LICs/LMICs will be globally beneficial by reducing the burden of communicable and non-communicable diseases alike. Ultimately, addressing the disparities in global healthcare is not just a moral imperative but also a strategic necessity in safeguarding the well-being of all humans. By fostering collaboration, empathy, and a shared commitment to collective welfare, we can build a world where access to healthcare is a fundamental right for every individual, regardless of geographic location or socioeconomic status.
Therefore, transforming to a precision innovation paradigm in cardiovascular medicine will require more than the scientific and technological advances described above. Interdisciplinary, intersectoral and global collaborations throughout the research and innovation pathway (352, 377, 378), underpinned by global health policy leadership, are necessary to implement suitable models of research funding and organization, data infrastructures and policies, novel clinical trial methods, medicines regulation, and patient access policies (Figure 8).
Figure 8
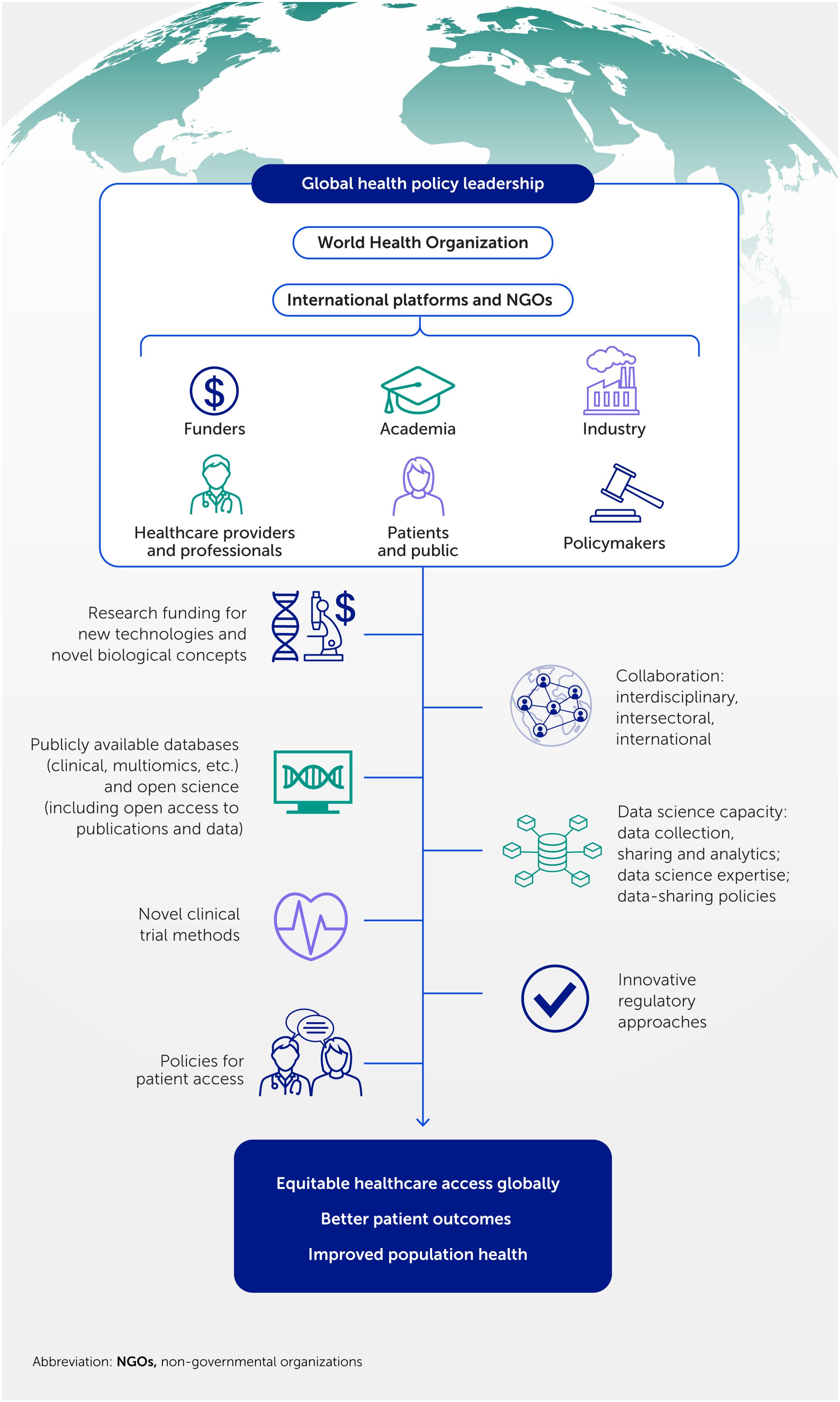
Figure 8. Global policy leadership fuels transformative cardiovascular innovations. Revolutionizing healthcare demands an unprecedented commitment from global policy leaders to foster interdisciplinary collaborations, funding, and support. This will drive innovation, enhance data science capacity, establish patient-centric public health policies, and concert efforts to reduce global inequality, addressing the silent pandemic of cardiovascular disease.
Conclusions
We have discussed how major challenges and needs in clinical cardiovascular medicine have provided opportunities for the scientific and medical communities to implement innovations and develop the systems approach needed to facilitate the search for disease mechanisms and establish unconventional strategies in drug development. The rapid evolution of cutting-edge technologies has recently increased our understanding of the biology of heterogeneity at cellular and patient levels, which would enable the establishment of new paradigms of precision medicine for CVDs. Advanced computational methods help to make traditionally undruggable targets druggable. New platforms, such as RNA therapeutics, facilitate modulation of undruggable targets (379).
Disruptive innovation not only solves challenges, but also leads us to new paradigms. Establishing new concepts in turn requires the development of new technologies. The key to successful cardiovascular innovation is a synergistic relationship between new technology and new paradigms, supported by a dynamic and intimate interplay between biomedical research and data science. In addition, cross-sector (e.g., academia–industry) or international partnerships can help to defend against global residual cardiovascular risks and address unmet medical needs. We saw this in action during the COVID-19 pandemic, where our community was forced to develop “borderless” solutions swiftly. This unprecedented challenge brought about a worldwide effort among scientists in academia and industry, leading to progress in comprehending virus transmission, effectively accelerating the development of novel technologies (e.g., mRNA vaccines), and reorganizing the medical community to improve responses to future crises. Such seamless collaboration across various disciplines, sectors, and nations will shift innovation paradigms to revolutionize borderless cardiovascular medicine and speed up the translation of discoveries into the clinic.
Acknowledgments
We thank Ms. Yennie Lee (United States) for editing the manuscript.
Statements
Author contributions
MA: Conceptualization, Funding acquisition, Project administration, Supervision, Writing – original draft, Writing – review & editing.
ARS: Conceptualization, Funding acquisition, Writing – original draft, Writing – review & editing.
SC: Writing – original draft, Writing – review & editing.
TA: Writing – original draft, Writing – review & editing.
AH: Funding acquisition, Writing – original draft, Writing – review & editing.
JTM: Writing – original draft, Writing – review & editing.
SAS: Writing – original draft, Writing – review & editing.
SU: Writing – original draft, Writing – review & editing.
EA: Funding acquisition, Writing – original draft, Writing – review & editing.
AA: Writing – original draft, Writing – review & editing.
J-LB: Funding acquisition, Writing – original draft, Writing – review & editing.
CG: Writing – original draft, Writing – review & editing.
CAM: Writing – original draft, Writing – review & editing.
NVM: Writing – original draft, Writing – review & editing.
CO: Writing – original draft, Writing – review & editing.
HTS: Writing – original draft, Writing – review & editing.
JL: Conceptualization, Funding acquisition, Writing – original draft, Writing – review & editing.
Data availability statement
The original contributions presented in this work are included in the article. Further inquiries can be directed to the corresponding author.
Funding
The authors declared that financial support was received for this work and/or its publication, from the National Heart Lung and Blood Institute of the United States (nos. R01HL126901, R01HL149302, OT2HL161847, and R01HL 174066 to MA; nos. R01HL136431, R01HL147095, and R01HL141917 to EA; nos. R01HL174066 to MA and EA; nos. R01HL155107, R01HL166137, and U01HG007691 to JL; no. K25HL150336 to AH), from Kowa Company, Ltd. (Nagoya, Japan to MA), from the American Heart Association (no. AHA957729 to JL), and the Sofina Solidarity Fund (Brussels, Belgium to J-LB). The funders were not involved in the writing of this article or the decision to submit it for publication.
Conflict of interest
TA is employed by the Kowa Company Ltd., Nagoya, Japan, and was a visiting scientist at the Brigham and Women’s Hospital. SU is currently an employee of Labcorp Laboratories, Saitama, Japan. These companies were not involved in the writing of this article or the decision to submit it for publication.
J-LB is a principal investigator of the WELRI/WELBIO Institute, Wallonia, Belgium.
The remaining authors declared that this work was conducted in the absence of financial relationships that could be construed as a potential conflict of interest.
The authors MA, ARS, SC, AH, JTM, SAS, SU, EA, AA, CAM, NVM, CO and HTS declared that they were an editorial board member of Frontiers at the time of submission. This had no impact on the peer review process and the final decision.
Generative AI statement
The authors declared that no generative AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
1. Martin SS, Aday AW, Allen NB, Almarzooq ZI, Anderson CAM, Arora P, et al. 2025 heart disease and stroke statistics: a report of US and global data from the American Heart Association. Circulation (2025) 151(8):e41–660. doi: 10.1161/CIR.0000000000001303
2. Mensah GA, Fuster V, Murray CJL, Roth GA, and Global Burden of Cardiovascular Diseases and Risks Collaborators. Global burden of cardiovascular diseases and risks, 1990-2022. J Am Coll Cardiol (2023) 82(25):2350–473. doi: 10.1016/j.jacc.2023.11.007
3. Fuster V, Badimon L, Badimon JJ, and Chesebro JH. The pathogenesis of coronary artery disease and the acute coronary syndromes. N Engl J Med (1992) 326(4):242–50. doi: 10.1056/NEJM199201233260406
4. Libby P. Molecular bases of the acute coronary syndromes. Circulation (1995) 91(11):2844–50. doi: 10.1161/01.cir.91.11.2844
5. Fayad ZA, Robson PM, and Fuster V. Rethinking heart attack prevention: the myth of the “vulnerable plaque” and reality of patient risk. J Am Coll Cardiol (2024) 83(22):2145–7. doi: 10.1016/j.jacc.2024.04.007
6. Kraler S, Mueller C, Libby P, and Bhatt DL. Acute coronary syndromes: mechanisms, challenges, and new opportunities. Eur Heart J (2025) 46(29):2866–89. doi: 10.1093/eurheartj/ehaf289
7. Groenewegen A, Rutten FH, Mosterd A, and Hoes AW. Epidemiology of heart failure. Eur J Heart Fail (2020) 22(8):1342–56. doi: 10.1002/ejhf.1858
8. Rasalam R, Sindone A, Deed G, Audehm RG, and Atherton JJ. State of precision medicine for heart failure with preserved ejection fraction in a new therapeutic age. ESC Heart Fail (2025) 12(3):1544–57. doi: 10.1002/ehf2.15205
9. Gupta P, Bast JA, Razavi AC, Canonico ME, Shahzad A, Naeem M, et al. Hypertension in atherosclerotic cardiovascular disease: insights into epidemiology, management strategies, and outcomes. Curr Opin Cardiol (2025) 40(4):206–12. doi: 10.1097/HCO.0000000000001226
10. Blaser MC, Bäck M, Lüscher TF, and Aikawa E. Calcific aortic stenosis: omics-based target discovery and therapy development. Eur Heart J (2025) 46(7):620–34. doi: 10.1093/eurheartj/ehae829
11. Buja LM and Nikolai N. Anitschkow and the lipid hypothesis of atherosclerosis. Cardiovasc Pathol (2014) 23(3):183–4. doi: 10.1016/j.carpath.2013.12.004
12. Steinberg D. In celebration of the 100th anniversary of the lipid hypothesis of atherosclerosis. J Lipid Res (2013) 54(11):2946–9. doi: 10.1194/jlr.R043414
13. Osler W. Angina pectoris and arteriosclerosis. JAMA (2015) 314(18):1981. doi: 10.1001/jama.2014.12084
14. Classics in arteriosclerosis research: on experimental cholesterin steatosis and its significance in the origin of some pathological processes by N. Anitschkow and S. Chalatow, translated by Mary Z. Pelias, 1913. Arteriosclerosis (1983) 3(2):178–82 doi: 10.1161/01.ATV.3.2.178
15. Mahmood SS, Levy D, Vasan RS, and Wang TJ. The Framingham heart study and the epidemiology of cardiovascular disease: a historical perspective. Lancet (2014) 383(9921):999–1008. doi: 10.1016/S0140-6736(13)61752-3
16. Packard C, Chapman MJ, Sibartie M, Laufs U, and Masana L. Intensive low-density lipoprotein cholesterol lowering in cardiovascular disease prevention: opportunities and challenges. Heart (2021) 107(17):1369–75. doi: 10.1136/heartjnl-2020-318760
17. Aday AW and Ridker PM. Targeting residual inflammatory risk: a shifting paradigm for atherosclerotic disease. Front Cardiovasc Med (2019) 6:16. doi: 10.3389/fcvm.2019.00016
18. Libby P and Aikawa M. Stabilization of atherosclerotic plaques: new mechanisms and clinical targets. Nat Med (2002) 8(11):1257–62. doi: 10.1038/nm1102-1257
19. Corsini A, Ginsberg HN, and Chapman MJ. Therapeutic PCSK9 targeting: inside versus outside the hepatocyte? Pharmacol Ther (2025) 268:108812. doi: 10.1016/j.pharmthera.2025.108812
20. Tokgözoğlu L and Libby P. The dawn of a new era of targeted lipid-lowering therapies. Eur Heart J (2022) 43(34):3198–208. doi: 10.1093/eurheartj/ehab841
21. Dhindsa DS, Sandesara PB, Shapiro MD, and Wong ND. The evolving understanding and approach to residual cardiovascular risk management. Front Cardiovasc Med (2020) 7:88. doi: 10.3389/fcvm.2020.00088
22. Patel KV, Pandey A, and de Lemos JA. Conceptual framework for addressing residual atherosclerotic cardiovascular disease risk in the era of precision medicine. Circulation (2018) 137(24):2551–3. doi: 10.1161/CIRCULATIONAHA.118.035289
23. Cesaro A, De Michele G, Fimiani F, Acerbo V, Scherillo G, Signore G, et al. Visceral adipose tissue and residual cardiovascular risk: a pathological link and new therapeutic options. Front Cardiovasc Med (2023) 10:1187735. doi: 10.3389/fcvm.2023.1187735
24. Skrobucha A, Pindlowski P, Krajewska N, Grabowski M, and Jonik S. Anti-inflammatory effects of glucagon-like peptide-1 (GLP-1) in coronary artery disease: a comprehensive review. Front Cardiovasc Med (2024) 11:1446468. doi: 10.3389/fcvm.2024.1446468
25. Pratley RE, Tuttle KR, Rossing P, Rasmussen S, Perkovic V, Nielsen OW, et al. Effects of semaglutide on heart failure outcomes in diabetes and chronic kidney disease in the FLOW trial. J Am Coll Cardiol (2024) 84(17):1615–28. doi: 10.1016/j.jacc.2024.08.004
26. Garofolo M, Penno G, Solini A, Orsi E, Vitale M, Resi V, et al. Relationship between degree of risk factor control and all-cause mortality in individuals with type 2 diabetes: a prospective cohort study. Eur J Intern Med (2024) 128:53–62. doi: 10.1016/j.ejim.2024.05.034
27. Vaughan AS, Coronado F, Casper M, Loustalot F, and Wright JS. County-level trends in hypertension-related cardiovascular disease mortality-United States, 2000 to 2019. J Am Heart Assoc (2022) 11(7):e024785. doi: 10.1161/JAHA.121.024785
28. Tsao CW, Aday AW, Almarzooq ZI, Alonso A, Beaton AZ, Bittencourt MS, et al. Heart disease and stroke Statistics-2022 update: a report from the American Heart Association. Circulation (2022) 145(8):e153–639. doi: 10.1161/CIR.0000000000001052
29. Benjamin EJ, Blaha MJ, Chiuve SE, Cushman M, Das SR, Deo R, et al. Heart disease and stroke Statistics—2017 update: a report from the American Heart Association. Circulation (2017) 135(10):e146–603. doi: 10.1161/CIR.0000000000000485
30. Roth GA, Mensah GA, Johnson CO, Addolorato G, Ammirati E, Baddour LM, et al. Global burden of cardiovascular diseases and risk factors, 1990–2019: update from the GBD 2019 study. J Am Coll Cardiol (2020) 76(25):2982–3021. doi: 10.1016/j.jacc.2020.11.010
31. Chelvanambi S, Decano JL, Winkels H, Giannarelli C, and Aikawa M. Decoding macrophage heterogeneity to unravel vascular inflammation as a path to precision medicine. Arterioscler Thromb Vasc Biol (2024) 44(11):2253–7. doi: 10.1161/ATVBAHA.124.319571
32. Smith MT, de la Rosa R, and Daniels SI. Using exposomics to assess cumulative risks and promote health. Environ Mol Mutagen (2015) 56(9):715–23. doi: 10.1002/em.21985
33. Vineis P, Robinson O, Chadeau-Hyam M, Dehghan A, Mudway I, and Dagnino S. What is new in the exposome? Environ Int (2020) 143:105887. doi: 10.1016/j.envint.2020.105887
34. Bucher ML, Anderson FL, Lai Y, Dicent J, Miller GW, and Zota AR. Exposomics as a tool to investigate differences in health and disease by sex and gender. Exposome (2023) 3(1):osad003. doi: 10.1093/exposome/osad003
35. Baptiste DL, Turkson-Ocran RA, Ogungbe O, Koirala B, Francis L, Spaulding EM, et al. Heterogeneity in cardiovascular disease risk factor prevalence among white, African American, African immigrant, and Afro-Caribbean adults: insights from the 2010–2018 national health interview survey. J Am Heart Assoc (2022) 11(18):e025235. doi: 10.1161/JAHA.122.025235
36. Rajagopalan S and Landrigan PJ. Pollution and the heart. N Engl J Med (2021) 385(20):1881–92. doi: 10.1056/NEJMra2030281
37. Zhang A,, Wu Z, Wu E, Wu M, Snyder MP, Zou J, et al. Leveraging physiology and artificial intelligence to deliver advancements in health care. Physiol Rev (2023) 103(4):2423–50. doi: 10.1152/physrev.00033.2022
38. Sonawane AR, Aikawa E, and Aikawa M. Connections for matters of the heart: network medicine in cardiovascular diseases. Front Cardiovasc Med (2022) 9:873582. doi: 10.3389/fcvm.2022.873582
39. Chandy M, Obal D, and Wu JC. Elucidating effects of environmental exposure using human-induced pluripotent stem cell disease modeling. EMBO Mol Med (2022) 14(11):e13260. doi: 10.15252/emmm.202013260
40. Kathiresan S and Srivastava D. Genetics of human cardiovascular disease. Cell (2012) 148(6):1242–57. doi: 10.1016/j.cell.2012.03.001
41. Soremekun O, Dib MJ, Rajasundaram S, Fatumo S, and Gill D. Genetic heterogeneity in cardiovascular disease across ancestries: insights for mechanisms and therapeutic intervention. Camb Prism Precis Med (2023) 1:e8. doi: 10.1017/pcm.2022.13
42. Gurdasani D, Barroso I, Zeggini E, and Sandhu MS. Genomics of disease risk in globally diverse populations. Nat Rev Genet (2019) 20(9):520–35. doi: 10.1038/s41576-019-0144-0
43. Woodward AA, Urbanowicz RJ, Naj AC, and Moore JH. Genetic heterogeneity: challenges, impacts, and methods through an associative lens. Genet Epidemiol (2022) 46(8):555–71. doi: 10.1002/gepi.22497
44. Simonetto C, Rospleszcz S, Kaiser JC, and Furukawa K. Heterogeneity in coronary heart disease risk. Sci Rep (2022) 12(1):10131. doi: 10.1038/s41598-022-14013-3
45. Watkin DM, Lawry EY, Mann GV, and Halperin M. A study of serum beta lipoprotein and total cholesterol variability and its relation to age and serum level in adult human subjects. J Clin Invest (1954) 33(6):874–83. doi: 10.1172/JCI102960
46. Castelli WP, Doyle JT, Gordon T, Hames CG, Hjortland MC, Hulley SB, et al. HDL cholesterol and other lipids in coronary heart disease. The cooperative lipoprotein phenotyping study. Circulation (1977) 55(5):767–72. doi: 10.1161/01.cir.55.5.767
47. Castelli WP, Cooper GR, Doyle JT, Garcia-Palmieri M, Gordon T, Hames C, et al. Distribution of triglyceride and total, LDL and HDL cholesterol in several populations: a cooperative lipoprotein phenotyping study. J Chronic Dis (1977) 30(3):147–69. doi: 10.1016/0021-9681(77)90082-0
48. Ridker PM. Evaluating novel cardiovascular risk factors: can we better predict heart attacks? Ann Intern Med (1999) 130(11):933–7. doi: 10.7326/0003-4819-130-11-199906010-00018
49. Crea F and Libby P. Acute coronary syndromes: the way forward from mechanisms to precision treatment. Circulation (2017) 136(12):1155–66. doi: 10.1161/CIRCULATIONAHA.117.029870
50. Dai X, Wiernek S, Evans JP, and Runge MS. Genetics of coronary artery disease and myocardial infarction. World J Cardiol (2016) 8(1):1–23. doi: 10.4330/wjc.v8.i1.1
51. Kaski JC, Crea F, Gersh BJ, and Camici PG. Reappraisal of ischemic heart disease. Circulation (2018) 138(14):1463–80. doi: 10.1161/CIRCULATIONAHA.118.031373
52. Maron BJ, Maron MS, Maron BA, and Loscalzo J. Moving beyond the sarcomere to explain heterogeneity in hypertrophic cardiomyopathy: JACC review topic of the week. J Am Coll Cardiol (2019) 73(15):1978–86. doi: 10.1016/j.jacc.2019.01.061
53. Peters AE, Tromp J, Shah SJ, Lam CSP, Lewis GD, Borlaug BA, et al. Phenomapping in heart failure with preserved ejection fraction: insights, limitations, and future directions. Cardiovasc Res (2023) 118(18):3403–15. doi: 10.1093/cvr/cvac179
54. Simmonds SJ, Cuijpers I, Heymans S, and Jones EAV. Cellular and molecular differences between HFpEF and HFrEF: a step ahead in an improved pathological understanding. Cells (2020) 9(1):242. doi: 10.3390/cells9010242
55. Liu Y, Chen Y, Wei B, Li H, Peng Y, and Luo Z. Impacts of ABCG2 loss of function variant (p. Gln141Lys, c.421 C > A, rs2231142) on lipid levels and statin efficiency: a systematic review and meta-analysis. BMC Cardiovasc Disord (2024) 24(1):202. doi: 10.1186/s12872-024-03821-2
56. Ingelman-Sundberg M and Pirmohamed M. Precision medicine in cardiovascular therapeutics: evaluating the role of pharmacogenetic analysis prior to drug treatment. J Intern Med (2024) 295(5):583–98. doi: 10.1111/joim.13772
57. Chan MY, Andreotti F, and Becker RC. Hypercoagulable states in cardiovascular disease. Circulation (2008) 118(22):2286–97. doi: 10.1161/CIRCULATIONAHA.108.778837
58. Fernandez DM, Rahman AH, Fernandez NF, Chudnovskiy A, Amir ED, Amadori L, et al. Single-cell immune landscape of human atherosclerotic plaques. Nat Med (2019) 25(10):1576–88. doi: 10.1038/s41591-019-0590-4
59. Depuydt MAC, Prange KHM, Slenders L, Örd T, Elbersen D, Boltjes A, et al. Microanatomy of the human atherosclerotic plaque by single-cell transcriptomics. Circ Res (2020) 127(11):1437–55. doi: 10.1161/CIRCRESAHA.120.316770
60. Lin JD, Nishi H, Poles J, Niu X, McCauley C, Rahman K, et al. Single-cell analysis of fate-mapped macrophages reveals heterogeneity, including stem-like properties, during atherosclerosis progression and regression. JCI Insight (2019) 4(4):e124574. doi: 10.1172/jci.insight.124574
61. Vallejo J, Cochain C, Zernecke A, and Ley K. Heterogeneity of immune cells in human atherosclerosis revealed by scRNA-Seq. Cardiovasc Res (2021) 117(13):2537–43. doi: 10.1093/cvr/cvab260
62. Pan H, Xue C, Auerbach BJ, Fan J, Bashore AC, Cui J, et al. Single-cell genomics reveals a novel cell state during smooth muscle cell phenotypic switching and potential therapeutic targets for atherosclerosis in mouse and human. Circulation (2020) 142(21):2060–75. doi: 10.1161/CIRCULATIONAHA.120.048378
63. Kalluri AS, Vellarikkal SK, Edelman ER, Nguyen L, Subramanian A, Ellinor PT, et al. Single-cell analysis of the normal mouse aorta reveals functionally distinct endothelial cell populations. Circulation (2019) 140(2):147–63. doi: 10.1161/CIRCULATIONAHA.118.038362
64. Winkels H, Ehinger E, Vassallo M, Buscher K, Dinh HQ, Kobiyama K, et al. Atlas of the immune cell repertoire in mouse atherosclerosis defined by single-cell RNA-sequencing and mass cytometry. Circ Res (2018) 122(12):1675–88. doi: 10.1161/CIRCRESAHA.117.312513
65. McQueen LW, Ladak SS, Abbasciano R, George SJ, Suleiman MS, Angelini GD, et al. Next-generation and single-cell sequencing approaches to study atherosclerosis and vascular inflammation pathophysiology: a systematic review. Front Cardiovasc Med (2022) 9:849675. doi: 10.3389/fcvm.2022.849675
66. Sonawane AR, Pucéat M, and Jo H. Single-cell OMICs analyses in cardiovascular diseases. Front Cardiovasc Med (2024) 11:1413184. doi: 10.3389/fcvm.2024.1413184
67. Decano JL and Aikawa M. Dynamic macrophages: understanding mechanisms of activation as guide to therapy for atherosclerotic vascular disease. Front Cardiovasc Med (2018) 5:97. doi: 10.3389/fcvm.2018.00097
68. Decano JL, Maiorino E, Matamalas JT, Chelvanambi S, Tiemeijer BM, Yanagihara Y, et al. Cellular heterogeneity of activated primary human macrophages and associated drug-gene networks: from biology to precision therapeutics. Circulation (2023) 148(19):1459–78. doi: 10.1161/CIRCULATIONAHA.123.064794
69. Geyer PE, Holdt LM, Teupser D, and Mann M. Revisiting biomarker discovery by plasma proteomics. Mol Syst Biol (2017) 13(9):942. doi: 10.15252/msb.20156297
70. Eldjarn GH, Ferkingstad E, Lund SH, Helgason H, Magnusson OT, Gunnarsdottir K, et al. Large-scale plasma proteomics comparisons through genetics and disease associations. Nature (2023) 622(7982):348–58. doi: 10.1038/s41586-023-06563-x
71. Leopold JA, Maron BA, and Loscalzo J. The application of big data to cardiovascular disease: paths to precision medicine. J Clin Invest (2020) 130(1):29–38. doi: 10.1172/JCI129203
72. Mohanta SK, Peng L, Li Y, Lu S, Sun T, Carnevale L, et al. Neuroimmune cardiovascular interfaces control atherosclerosis. Nature (2022) 605(7908):152–9. doi: 10.1038/s41586-022-04673-6
73. Caudal A, Snyder MP, and Wu JC. Harnessing human genetics and stem cells for precision cardiovascular medicine. Cell Genom (2024) 4(2):100445. doi: 10.1016/j.xgen.2023.100445
74. Atutornu J, Milne R, Costa A, Patch C, and Middleton A. Towards equitable and trustworthy genomics research. EBiomedicine (2022) 76:103879. doi: 10.1016/j.ebiom.2022.103879
75. Loscalzo J, Barabási AL, and Silverman EK. Network medicine: complex systems in human disease and therapeutics. Cambridge, MA: Harvard University Press (2017).
76. Lusis AJ, Seldin MM, Allayee H, Bennett BJ, Civelek M, Davis RC, et al. The hybrid mouse diversity panel: a resource for systems genetics analyses of metabolic and cardiovascular traits. J Lipid Res (2016) 57(6):925–42. doi: 10.1194/jlr.R066944
77. Daugherty A, Tall AR, Daemen MJAP, Falk E, Fisher EA, García-Cardeña G, et al. Recommendation on design, execution, and reporting of animal atherosclerosis studies: a scientific statement from the American Heart Association. Arterioscler Thromb Vasc Biol (2017) 37(9):e131–57. doi: 10.1161/ATV.0000000000000062
78. Thomas D and Wu JC. Integrative approaches in cardiac tissue engineering: bridging cellular complexity to create accurate physiological models. iScience (2025) 28(8):113003. doi: 10.1016/j.isci.2025.113003
79. Juguilon C, Khosravi R, Radisic M, and Wu JC. In vitro modeling of interorgan crosstalk: multi-organ-on-a-chip for studying cardiovascular-kidney-metabolic syndrome. Circ Res (2025) 136(11):1476–93. doi: 10.1161/CIRCRESAHA.125.325497
80. Yildirim Z, Swanson K, Wu X, Zou J, and Wu J. Next-gen therapeutics: pioneering drug discovery with iPSCs, genomics, AI, and clinical trials in a dish. Annu Rev Pharmacol Toxicol (2025) 65(1):71–90. doi: 10.1146/annurev-pharmtox-022724-095035
81. Wu X, Swanson K, Yildirim Z, Liu W, Liao R, and Wu JC. Clinical trials in-a-dish for cardiovascular medicine. Eur Heart J (2024) 45(40):4275–90. doi: 10.1093/eurheartj/ehae519
82. Brown DG, Wobst HJ, Kapoor A, Kenna LA, and Southall N. Clinical development times for innovative drugs. Nat Rev Drug Discov (2022) 21(11):793–4. doi: 10.1038/d41573-021-00190-9
83. Mohs RC and Greig NH. Drug discovery and development: role of basic biological research. Alzheimers Dement (2017) 3(4):651–7. doi: 10.1016/j.trci.2017.10.005
84. Sasso JM, Ambrose BJB, Tenchov R, Datta RS, Basel MT, DeLong RK, et al. The progress and promise of RNA medicine─an arsenal of targeted treatments. J Med Chem (2022) 65(10):6975–7015. doi: 10.1021/acs.jmedchem.2c00024
85. Hinkson IV, Madej B, and Stahlberg EA. Accelerating therapeutics for opportunities in medicine: a paradigm shift in drug discovery. Front Pharmacol (2020) 11:770. doi: 10.3389/fphar.2020.00770
86. Pilkington EH, Suys EJA, Trevaskis NL, Wheatley AK, Zukancic D, Algarni A, et al. From influenza to COVID-19: lipid nanoparticle mRNA vaccines at the frontiers of infectious diseases. Acta Biomater (2021) 131:16–40. doi: 10.1016/j.actbio.2021.06.023
87. Kalinke U, Barouch DH, Rizzi R, Lagkadinou E, Türeci Ö, Pather S, et al. Clinical development and approval of COVID-19 vaccines. Expert Rev Vaccines (2022) 21(5):609–19. doi: 10.1080/14760584.2022.2042257
88. Dowden H and Munro J. Trends in clinical success rates and therapeutic focus. Nat Rev Drug Discov (2019) 18(7):495–6. doi: 10.1038/d41573-019-00074-z
89. Takebe T, Imai R, and Ono S. The current status of drug discovery and development as originated in United States academia: the influence of industrial and academic collaboration on drug discovery and development. Clin Transl Sci (2018) 11(6):597–606. doi: 10.1111/cts.12577
90. Firoozbakht F, Elkjaer ML, Handy DE, Wang RS, Chervontseva Z, Rarey M, et al. Exploring common mechanisms of adverse drug reactions and disease phenotypes through network-based analysis. Cell Rep Methods (2025) 5(2):100990. doi: 10.1016/j.crmeth.2025.100990
91. Harrison RK. Phase II and phase III failures: 2013–2015. Nat Rev Drug Discov (2016) 15(12):817–8. doi: 10.1038/nrd.2016.184
92. Ferdinandy P, Baczkó I, Bencsik P, Giricz Z, Görbe A, Pacher P, et al. Definition of hidden drug cardiotoxicity: paradigm change in cardiac safety testing and its clinical implications. Eur Heart J (2019) 40(22):1771–7. doi: 10.1093/eurheartj/ehy365
93. Moffat JG, Vincent F, Lee JA, Eder J, and Prunotto M. Opportunities and challenges in phenotypic drug discovery: an industry perspective. Nat Rev Drug Discov (2017) 16(8):531–43. doi: 10.1038/nrd.2017.111
94. Iwata H, Goettsch C, Sharma A, Ricchiuto P, Goh WWB, Halu A, et al. PARP9 and PARP14 cross-regulate macrophage activation via STAT1 ADP-ribosylation. Nat Commun (2016) 7:12849. doi: 10.1038/ncomms12849
95. Nakano T, Katsuki S, Chen M, Decano JL, Halu A, Lee LH, et al. Uremic toxin indoxyl sulfate promotes proinflammatory macrophage activation via the interplay of OATP2B1 and Dll4-notch signaling. Circulation (2019) 139(1):78–96. doi: 10.1161/CIRCULATIONAHA.118.034588
96. Paci P, Fiscon G, Conte F, Wang RS, Handy DE, Farina L, et al. Comprehensive network medicine-based drug repositioning via integration of therapeutic efficacy and side effects. NPJ Syst Biol Appl (2022) 8(1):12. doi: 10.1038/s41540-022-00221-0
97. Gardiner LJ, Carrieri AP, Wilshaw J, Checkley S, Pyzer-Knapp EO, and Krishna R. Using human in vitro transcriptome analysis to build trustworthy machine learning models for prediction of animal drug toxicity. Sci Rep (2020) 10(1):9522. doi: 10.1038/s41598-020-66481-0
98. Fiscon G, Conte F, Farina L, and Paci P. A comparison of network-based methods for drug repurposing along with an application to human complex diseases. Int J Mol Sci (2022) 23(7):3703. doi: 10.3390/ijms23073703
99. Finan C, Gaulton A, Kruger FA, Lumbers RT, Shah T, Engmann J, et al. The druggable genome and support for target identification and validation in drug development. Sci Transl Med (2017) 9(383):eaag1166. doi: 10.1126/scitranslmed.aag1166
100. Hopkins AL and Groom CR. The druggable genome. Nat Rev Drug Discov (2002) 1(9):727–30. doi: 10.1038/nrd892
101. Hopkins AL, Keserü GM, Leeson PD, Rees DC, and Reynolds CH. The role of ligand efficiency metrics in drug discovery. Nat Rev Drug Discov (2014) 13(2):105–21. doi: 10.1038/nrd4163
102. Tanaka T, Asano T, Okui T, Kuraoka S, Singh SA, Aikawa M, et al. Computational screening strategy for drug repurposing identified niclosamide as inhibitor of vascular calcification. Front Cardiovasc Med (2021) 8:826529. doi: 10.3389/fcvm.2021.826529
103. Asano T, Chelvanambi S, Decano JL, Whelan MC, Aikawa E, and Aikawa M. In silico drug screening approach using L1000-based connectivity map and its application to COVID-19. Front Cardiovasc Med (2022) 9:842641. doi: 10.3389/fcvm.2022.842641
104. Crooke ST, Witztum JL, Bennett CF, and Baker BF. RNA-targeted therapeutics. Cell Metab (2018) 27(4):714–39. doi: 10.1016/j.cmet.2018.03.004
105. Kim YK. RNA therapy: rich history, various applications and unlimited future prospects. Exp Mol Med (2022) 54(4):455–65. doi: 10.1038/s12276-022-00757-5
106. Krychtiuk KA, Rader DJ, and Granger CB. RNA-targeted therapeutics in cardiovascular disease: the time is now. Eur Heart J Cardiovasc Pharmacother (2022) 9(1):94–9. doi: 10.1093/ehjcvp/pvac052
107. Agache I and Akdis CA. Precision medicine and phenotypes, endotypes, genotypes, regiotypes, and theratypes of allergic diseases. J Clin Invest (2019) 129(4):1493–503. doi: 10.1172/JCI124611
108. Kitano H. Systems biology: a brief overview. Science (2002) 295(5560):1662–4. doi: 10.1126/science.1069492
109. Loscalzo J and Barabasi AL. Systems biology and the future of medicine. Wiley Interdiscip Rev Syst Biol Med (2011) 3(6):619–27. doi: 10.1002/wsbm.144
110. Schork NJ. Personalized medicine: time for one-person trials. Nature (2015) 520(7549):609–11. doi: 10.1038/520609a
111. Lee LY, Pandey AK, Maron BA, and Loscalzo J. Network medicine in cardiovascular Research. Cardiovasc Res (2021) 117(10):2186–202. doi: 10.1093/cvr/cvaa321
112. Sonawane AR, Weiss ST, Glass K, and Sharma A. Network medicine in the age of biomedical big data. Front Genet (2019) 10:294. doi: 10.3389/fgene.2019.00294
113. Ahn AC, Tewari M, Poon CS, and Phillips RS. The limits of reductionism in medicine: could systems biology offer an alternative? PloS Med (2006) 3(6):e208. doi: 10.1371/journal.pmed.0030208
114. Ahn AC, Tewari M, Poon CS, and Phillips RS. The clinical applications of a systems approach. PloS Med (2006) 3(7):e209. doi: 10.1371/journal.pmed.0030209
115. Wolkenhauer O, Auffray C, Jaster R, Steinhoff G, and Dammann O. The road from systems biology to systems medicine. Pediatr Res (2013) 73(4 Pt 2):502–7. doi: 10.1038/pr.2013.4
116. Clermont G, Auffray C, Moreau Y, Rocke DM, Dalevi D, Dubhashi D, et al. Bridging the gap between systems biology and medicine. Genome Med (2009) 1(9):88. doi: 10.1186/gm88
117. Barabási AL, Gulbahce N, and Loscalzo J. Network medicine: a network-based approach to human disease. Nat Rev Genet (2011) 12(1):56–68. doi: 10.1038/nrg2918
118. Lusis AJ and Weiss JN. Cardiovascular networks: systems-based approaches to cardiovascular disease. Circulation (2010) 121(1):157–70. doi: 10.1161/CIRCULATIONAHA.108.847699
119. MacLellan WR, Wang Y, and Lusis AJ. Systems-based approaches to cardiovascular disease. Nat Rev Cardiol (2012) 9(3):172–84. doi: 10.1038/nrcardio.2011.208
120. Sarajlić A and Pržulj N. Survey of network-based approaches to research of cardiovascular diseases. BioMed Res Int (2014) 2014:527029. doi: 10.1155/2014/527029
121. Zhao Y, Chen J, Freudenberg JM, Meng Q, Rajpal DK, and Yang X. Network-based identification and prioritization of key regulators of coronary artery disease loci. Arterioscler Thromb Vasc Biol (2016) 36(5):928–41. doi: 10.1161/ATVBAHA.115.306725
122. King JY, Ferrara R, Tabibiazar R, Spin JM, Chen MM, Kuchinsky A, et al. Pathway analysis of coronary atherosclerosis. Physiol Genomics (2005) 23(1):103–18. doi: 10.1152/physiolgenomics.00101.2005
123. Ashley EA, Ferrara R, King JY, Vailaya A, Kuchinsky A, He X, et al. Network analysis of human in-stent restenosis. Circulation (2006) 114(24):2644–54. doi: 10.1161/CIRCULATIONAHA.106.637025
124. Schlesinger J, Schueler M, Grunert M, Fischer JJ, Zhang Q, Krueger T, et al. The cardiac transcription network modulated by Gata4, Mef2a, Nkx2.5, Srf, histone modifications, and microRNAs. PloS Genet (2011) 7(2):e1001313. doi: 10.1371/journal.pgen.1001313
125. Skogsberg J, Lundström J, Kovacs A, Nilsson R, Noori P, Maleki S, et al. Transcriptional profiling uncovers a network of cholesterol-responsive atherosclerosis target genes. PloS Genet (2008) 4(3):e1000036. doi: 10.1371/journal.pgen.1000036
126. Jin G, Zhou X, Wang H, Zhao H, Cui K, Zhang XS, et al. The knowledge-integrated network biomarkers discovery for major adverse cardiac events. J Proteome Res (2008) 7(9):4013–21. doi: 10.1021/pr8002886
127. Camargo A and Azuaje F. Linking gene expression and functional network data in human heart failure. PloS One (2007) 2(12):e1347. doi: 10.1371/journal.pone.0001347
128. Camargo A and Azuaje F. Identification of dilated cardiomyopathy signature genes through gene expression and network data integration. Genomics (2008) 92(6):404–13. doi: 10.1016/j.ygeno.2008.05.007
129. Schlotter F, Halu A, Goto S, Blaser MC, Body SC, Lee LH, et al. Spatiotemporal multi-omics mapping generates a molecular atlas of the aortic valve and reveals networks driving disease. Circulation (2018) 138(4):377–93. doi: 10.1161/CIRCULATIONAHA.117.032291
130. Decano JL, Iwamoto Y, Goto S, Lee JY, Matamalas JT, Halu A, et al. A disease-driver population within interstitial cells of human calcific aortic valves identified via single-cell and proteomic profiling. Cell Rep (2022) 39(2):110685. doi: 10.1016/j.celrep.2022.110685
131. Blaser MC, Buffolo F, Halu A, Turner ME, Schlotter F, Higashi H, et al. Multiomics of tissue extracellular vesicles identifies unique modulators of atherosclerosis and calcific aortic valve stenosis. Circulation (2023) 148(8):661–78. doi: 10.1161/CIRCULATIONAHA.122.063402
132. Karlstädt A, Fliegner D, Kararigas G, Ruderisch HS, Regitz-Zagrosek V, and Holzhütter HG. CardioNet: a human metabolic network suited for the study of cardiomyocyte metabolism. BMC Syst Biol (2012) 6:114. doi: 10.1186/1752-0509-6-114
133. Ntalla I, Weng LC, Cartwright JH, Hall AW, Sveinbjornsson G, Tucker NR, et al. Multi-ancestry GWAS of the electrocardiographic PR interval identifies 202 loci underlying cardiac conduction. Nat Commun (2020) 11(1):2542. doi: 10.1038/s41467-020-15706-x
134. van Setten J, Verweij N, Mbarek H, Niemeijer MN, Trompet S, Arking DE, et al. Genome-wide association meta-analysis of 30,000 samples identifies seven novel loci for quantitative ECG traits. Eur J Hum Genet (2019) 27(6):952–62. doi: 10.1038/s41431-018-0295-z
135. Roselli C, Rienstra M, and Ellinor PT. Genetics of atrial fibrillation in 2020: GWAS, genome sequencing, polygenic risk, and beyond. Circ Res (2020) 127(1):21–33. doi: 10.1161/CIRCRESAHA.120.316575
136. Assum I, Krause J, Scheinhardt MO, Müller C, Hammer E, Börschel CS, et al. Tissue-specific multi-omics analysis of atrial fibrillation. Nat Commun (2022) 13(1):441. doi: 10.1038/s41467-022-27953-1
137. Diez D, Wheelock AM, Goto S, Haeggström JZ, Paulsson-Berne G, Hansson GK, et al. The use of network analyses for elucidating mechanisms in cardiovascular disease. Mol Biosyst (2010) 6(2):289–304. doi: 10.1039/b912078e
138. Wheelock CE, Wheelock AM, Kawashima S, Diez D, Kanehisa M, van Erk M, et al. Systems biology approaches and pathway tools for investigating cardiovascular disease. Mol Biosyst (2009) 5(6):588–602. doi: 10.1039/b902356a
139. He D, Liu ZP, and Chen L. Identification of dysfunctional modules and disease genes in congenital heart disease by a network-based approach. BMC Genomics (2011) 12:592. doi: 10.1186/1471-2164-12-592
140. Dewey FE, Perez MV, Wheeler MT, Watt C, Spin J, Langfelder P, et al. Gene coexpression network topology of cardiac development, hypertrophy, and failure. Circ Cardiovasc Genet (2011) 4(1):26–35. doi: 10.1161/CIRCGENETICS.110.941757
141. Zhang L, Li X, Tai J, Li W, and Chen L. Predicting candidate genes based on combined network topological features: a case study in coronary artery disease. PloS One (2012) 7(6):e39542. doi: 10.1371/journal.pone.0039542
142. Sarajlić A, Janjić V, Stojković N, Radak D, and Pržulj N. Network topology reveals key cardiovascular disease genes. PloS One (2013) 8(8):e71537. doi: 10.1371/journal.pone.0071537
143. Katsuki S, K Jha P, Lupieri A, Nakano T, Passos LSA, Rogers MA, et al. Proprotein convertase subtilisin/kexin 9 (PCSK9) promotes macrophage activation via LDL receptor-independent mechanisms. Circ Res (2022) 131(11):873–89. doi: 10.1161/CIRCRESAHA.121.320056
144. Decano JL, Singh SA, Gasparotto Bueno C, Ho Lee L, Halu A, Chelvanambi S, et al. Systems approach to discovery of therapeutic targets for vein graft disease: PPARα pivotally regulates metabolism, activation, and heterogeneity of macrophages and lesion development. Circulation (2021) 143(25):2454–70. doi: 10.1161/CIRCULATIONAHA.119.043724
145. Okui T, Iwashita M, Rogers MA, Halu A, Atkins SK, Kuraoka S, et al. CROT (carnitine O-octanoyltransferase) is a novel contributing factor in vascular calcification via promoting fatty acid metabolism and mitochondrial dysfunction. Arterioscler Thromb Vasc Biol (2021) 41(2):755–68. doi: 10.1161/ATVBAHA.120.315007
146. Passos LSA, Becker-Greene D, Braulio R, Le TD, Gelape CL, de Almeida LFR, et al. Proinflammatory matrix metalloproteinase-1 associates with mitral valve leaflet disruption following percutaneous mitral valvuloplasty. Front Cardiovasc Med (2021) 8:804111. doi: 10.3389/fcvm.2021.804111
147. Clift CL, Blaser MC, Gerrits W, Turner ME, Sonawane A, Pham T, et al. Intracellular proteomics and extracellular vesiculomics as a metric of disease recapitulation in 3D-bioprinted aortic valve arrays. Sci Adv (2024) 10(9):eadj9793. doi: 10.1126/sciadv.adj9793
148. Morgan S, Lee LH, Halu A, Nicolau JS, Higashi H, Ha AH, et al. Identifying novel mechanisms of abdominal aortic aneurysm via unbiased proteomics and systems biology. Front Cardiovasc Med (2022) 9:889994. doi: 10.3389/fcvm.2022.889994
149. Huan T, Esko T, Peters MJ, Pilling LC, Schramm K, Schurmann C, et al. A meta-analysis of gene expression signatures of blood pressure and hypertension. PloS Genet (2015) 11(3):e1005035. doi: 10.1371/journal.pgen.1005035
150. Huan T, Meng Q, Saleh MA, Norlander AE, Joehanes R, Zhu J, et al. Integrative network analysis reveals molecular mechanisms of blood pressure regulation. Mol Syst Biol (2015) 11(1):799. doi: 10.15252/msb.20145399
151. Frades I, Readhead B, Amadori L, Koplev S, Talukdar HA, Crane HM, et al. Systems pharmacology identifies an arterial wall regulatory gene network mediating coronary artery disease side effects of antiretroviral therapy. Circ Genom Precis Med (2019) 12(6):e002390. doi: 10.1161/CIRCGEN.118.002390
152. Halu A, De Domenico M, Arenas A, and Sharma A. The multiplex network of human diseases. NPJ Syst Biol Appl (2019) 5:15. doi: 10.1038/s41540-019-0092-5
153. Liu X, Maiorino E, Halu A, Glass K, Prasad RB, Loscalzo J, et al. Robustness and lethality in multilayer biological molecular networks. Nat Commun (2020) 11(1):6043. doi: 10.1038/s41467-020-19841-3
154. Maiorino E and Loscalzo J. Phenomics and robust multiomics data for cardiovascular disease subtyping. Arterioscler Thromb Vasc Biol (2023) 43(7):1111–23. doi: 10.1161/ATVBAHA.122.318892
155. Wang RS, Maron BA, and Loscalzo J. Multiomics network medicine approaches to precision medicine and therapeutics in cardiovascular diseases. Arterioscler Thromb Vasc Biol (2023) 43(4):493–503. doi: 10.1161/ATVBAHA.122.318731
156. Loscalzo J. Molecular interaction networks and drug development: novel approach to drug target identification and drug repositioning. FASEB J (2023) 37(1):e22660. doi: 10.1096/fj.202201683R
157. Blaser MC, Kraler S, Lüscher TF, and Aikawa E. Multi-omics approaches to define calcific aortic valve disease pathogenesis. Circ Res (2021) 128(9):1371–97. doi: 10.1161/CIRCRESAHA.120.317979
158. Wang Z, Clark NR, and Ma’ayan A. Drug-induced adverse events prediction with the Lincs L1000 data. Bioinformatics (2016) 32(15):2338–45. doi: 10.1093/bioinformatics/btw168
159. Ghiassian SD, Menche J, Chasman DI, Giulianini F, Wang R, Ricchiuto P, et al. Endophenotype Network models: common core of complex diseases. Sci Rep (2016) 6:27414. doi: 10.1038/srep27414
160. Menche J, Sharma A, Kitsak M, Ghiassian SD, Vidal M, Loscalzo J, et al. Disease networks. Uncovering disease-disease relationships through the incomplete interactome. Science (2015) 347(6224):1257601. doi: 10.1126/science.1257601
161. Halu A, Wang JG, Iwata H, Mojcher A, Abib AL, Singh SA, et al. Context-enriched interactome powered by proteomics helps the identification of novel regulators of macrophage activation. eLife (2018) 7:e37059. doi: 10.7554/eLife.37059
162. Yang X. Multitissue multiomics systems biology to dissect complex diseases. Trends Mol Med (2020) 26(8):718–28. doi: 10.1016/j.molmed.2020.04.006
163. Morgan NV. Editorial: case reports in cardiovascular genetics and systems medicine: 2022. Front Cardiovasc Med (2023) 10:1282147. doi: 10.3389/fcvm.2023.1282147
164. Jordan E, Peterson L, Ai T, Asatryan B, Bronicki L, Brown E, et al. Evidence-based assessment of genes in dilated cardiomyopathy. Circulation (2021) 144(1):7–19. doi: 10.1161/CIRCULATIONAHA.120.053033
165. Stuart T and Satija R. Integrative single-cell analysis. Nat Rev Genet (2019) 20(5):257–72. doi: 10.1038/s41576-019-0093-7
166. Crosetto N, Bienko M, and van Oudenaarden A. Spatially resolved transcriptomics and beyond. Nat Rev Genet (2015) 16(1):57–66. doi: 10.1038/nrg3832
167. Joshi A, Rienks M, Theofilatos K, and Mayr M. Systems biology in cardiovascular disease: a multiomics approach. Nat Rev Cardiol (2021) 18(5):313–30. doi: 10.1038/s41569-020-00477-1
168. Skelly DA, Squiers GT, McLellan MA, Bolisetty MT, Robson P, Rosenthal NA, et al. Single-cell transcriptional profiling reveals cellular diversity and intercommunication in the mouse heart. Cell Rep (2018) 22(3):600–10. doi: 10.1016/j.celrep.2017.12.072
169. Amadori L, Calcagno C, Fernandez DM, Koplev S, Fernandez N, Kaur R, et al. Systems immunology-based drug repurposing framework to target inflammation in atherosclerosis. Nat Cardiovasc Res (2023) 2(6):550–71. doi: 10.1038/s44161-023-00278-y
170. Teschendorff AE and Feinberg AP. Statistical mechanics meets single-cell biology. Nat Rev Genet (2021) 22(7):459–76. doi: 10.1038/s41576-021-00341-z
171. Hamilton WL, Ying R, and Leskovec J. Representation learning on graphs: methods and applications. IEEE Data Eng Bull (2017) 40(3):52–74. Available at: http://sites.computer.org/debull/A17sept/p52.pdf
172. Zitnik M, Nguyen F, Wang B, Leskovec J, Goldenberg A, and Hoffman MM. Machine learning for integrating data in biology and medicine: principles, practice, and opportunities. Inf Fusion (2019) 50:71–91. doi: 10.1016/j.inffus.2018.09.012
173. Huang K, Fu T, Gao W, Zhao Y, Roohani Y, Leskovec J, et al. Artificial intelligence foundation for therapeutic science. Nat Chem Biol (2022) 18(10):1033–6. doi: 10.1038/s41589-022-01131-2
174. Ballantyne CM. Newer risk markers and surrogate endpoints in atherosclerosis management. Clin Cardiol (2001) 24(S3):13–7. doi: 10.1002/clc.4960241504
175. Stanley BA, Gundry RL, Cotter RJ, and Van Eyk JE. Heart disease, clinical proteomics and mass spectrometry. Dis Markers (2004) 20(3):167–78. doi: 10.1155/2004/965261
176. Pape ME and Bisgaier CL. Discovering HDL-elevating drugs: are there simplistic approaches to a polygenic disorder? IDrugs (1998) 1(4):442–51
177. Singh SA, Aikawa E, and Aikawa M. Current trends and future perspectives of state-of-the-art proteomics technologies applied to cardiovascular disease research. Circ J (2016) 80(8):1674–83. doi: 10.1253/circj.CJ-16-0499
178. McGarrah RW, Crown SB, Zhang GF, Shah SH, and Newgard CB. Cardiovascular metabolomics. Circ Res (2018) 122(9):1238–58. doi: 10.1161/CIRCRESAHA.117.311002
179. Barallobre-Barreiro J, Chung YL, and Mayr M. Proteomics and metabolomics for mechanistic insights and biomarker discovery in cardiovascular disease. Rev Esp Cardiol (Engl Ed) (2013) 66(8):657–61. doi: 10.1016/j.rec.2013.04.009
180. Wirka RC, Pjanic M, and Quertermous T. Advances in transcriptomics: investigating cardiovascular disease at unprecedented resolution. Circ Res (2018) 122(9):1200–20. doi: 10.1161/CIRCRESAHA.117.310910
181. Leopold JA and Loscalzo J. Emerging role of precision medicine in cardiovascular disease. Circ Res (2018) 122(9):1302–15. doi: 10.1161/CIRCRESAHA.117.310782
182. Vidal M, Chan DW, Gerstein M, Mann M, Omenn GS, Tagle D, et al. The human proteome - a scientific opportunity for transforming diagnostics, therapeutics, and healthcare. Clin Proteomics (2012) 9(1):6. doi: 10.1186/1559-0275-9-6
183. Herrington DM, Mao C, Parker SJ, Fu Z, Yu G, Chen L, et al. Proteomic architecture of human coronary and aortic atherosclerosis. Circulation (2018) 137(25):2741–56. doi: 10.1161/CIRCULATIONAHA.118.034365
184. Vogel C and Marcotte EM. Insights into the regulation of protein abundance from proteomic and transcriptomic analyses. Nat Rev Genet (2012) 13(4):227–32. doi: 10.1038/nrg3185
185. Kustatscher G, Grabowski P, and Rappsilber J. Pervasive coexpression of spatially proximal genes is buffered at the protein level. Mol Syst Biol (2017) 13(8):937. doi: 10.15252/msb.20177548
186. Slavov N. Unlocking the potential of single-cell omics. J Proteome Res (2025) 24(4):1481. doi: 10.1021/acs.jproteome.5c00197
187. Kelly RT. Single-cell proteomics: progress and prospects. Mol Cell Proteomics (2020) 19(11):1739–48. doi: 10.1074/mcp.R120.002234
188. Reyes-Soffer G, Millar JS, Ngai C, Jumes P, Coromilas E, Asztalos B, et al. Cholesteryl ester transfer protein inhibition with anacetrapib decreases fractional clearance rates of high-density lipoprotein apolipoprotein A-I and plasma cholesteryl ester transfer protein. Arterioscler Thromb Vasc Biol (2016) 36(5):994–1002. doi: 10.1161/ATVBAHA.115.306680
189. Lassman ME, McAvoy T, Lee AYH, Chappell D, Wong O, Zhou H, et al. Practical immunoaffinity-enrichment LC-MS for measuring protein kinetics of low-abundance proteins. Clin Chem (2014) 60(9):1217–24. doi: 10.1373/clinchem.2014.222455
190. Millar JS, Reyes-Soffer G, Jumes P, Dunbar RL, deGoma EM, Baer AL, et al. Anacetrapib lowers LDL by increasing ApoB clearance in mildly hypercholesterolemic subjects. J Clin Invest (2015) 125(6):2510–22. doi: 10.1172/JCI80025
191. Singh SA, Andraski AB, Pieper B, Goh W, Mendivil CO, Sacks FM, et al. Multiple apolipoprotein kinetics measured in human HDL by high-resolution/accurate mass parallel reaction monitoring. J Lipid Res (2016) 57(4):714–28. doi: 10.1194/jlr.D061432
192. Singh SA, Andraski AB, Higashi H, Lee LH, Ramsaroop A, Sacks FM, et al. Metabolism of PLTP, CETP, and LCAT on multiple HDL sizes using the Orbitrap Fusion Lumos. JCI Insight (2021) 6(3):e143526. doi: 10.1172/jci.insight.143526
193. Smith LM, Agar JN, Chamot-Rooke J, Danis PO, Ge Y, Loo JA, et al. The Human Proteoform Project: defining the human proteome. Sci Adv (2021) 7(46):eabk0734. doi: 10.1126/sciadv.abk0734
194. Moore KJ, Koplev S, Fisher EA, Tabas I, Björkegren JLM, Doran AC, et al. Macrophage trafficking, inflammatory resolution, and genomics in atherosclerosis: JACC macrophage in CVD series (Part 2). J Am Coll Cardiol (2018) 72(18):2181–97. doi: 10.1016/j.jacc.2018.08.2147
195. Kasikara C, Doran AC, Cai B, and Tabas I. The role of non-resolving inflammation in atherosclerosis. J Clin Invest (2018) 128(7):2713–23. doi: 10.1172/JCI97950
196. Galindo CL, Khan S, Zhang X, Yeh YS, Liu Z, and Razani B. Lipid-laden foam cells in the pathology of atherosclerosis: shedding light on new therapeutic targets. Expert Opin Ther Targets (2023) 27(12):1231–45. doi: 10.1080/14728222.2023.2288272
197. Mantovani A, Sozzani S, Locati M, Allavena P, and Sica A. Macrophage polarization: tumor-associated macrophages as a paradigm for polarized M2 mononuclear phagocytes. Trends Immunol (2002) 23(11):549–55. doi: 10.1016/s1471-4906(02)02302-5
198. Gordon S and Martinez FO. Alternative activation of macrophages: mechanism and functions. Immunity (2010) 32(5):593–604. doi: 10.1016/j.immuni.2010.05.007
199. Nahrendorf M and Swirski FK. Abandoning M1/M2 for a network model of macrophage function. Circ Res (2016) 119(3):414–17. doi: 10.1161/CIRCRESAHA.116.309194
200. Martinez FO and Gordon S. The M1 and M2 paradigm of macrophage activation: time for reassessment. F1000Prime Rep (2014) 6:13. doi: 10.12703/P6-13
201. Bronte V, Brandau S, Chen SH, Colombo MP, Frey AB, Greten TF, et al. Recommendations for myeloid-derived suppressor cell nomenclature and characterization standards. Nat Commun (2016) 7:12150. doi: 10.1038/ncomms12150
202. Kumar Jha P, Aikawa M, and Aikawa E. Macrophage heterogeneity and efferocytosis: beyond the M1/M2 dichotomy. Circ Res (2024) 134(2):186–8. doi: 10.1161/CIRCRESAHA.123.324011
203. Revelo XS, Parthiban P, Chen C, Barrow F, Fredrickson G, Wang H, et al. Cardiac resident macrophages prevent fibrosis and stimulate angiogenesis. Circ Res (2021) 129(12):1086–101. doi: 10.1161/CIRCRESAHA.121.319737
204. Jia G, Preussner J, Chen X, Guenther S, Yuan X, Yekelchyk M, et al. Single cell RNA-seq and ATAC-seq analysis of cardiac progenitor cell transition states and lineage settlement. Nat Commun (2018) 9(1):4877. doi: 10.1038/s41467-018-07307-6
205. Wang L, Yang Y, Ma H, Xie Y, Xu J, Near D, et al. Single-cell dual-omics reveals the transcriptomic and epigenomic diversity of cardiac non-myocytes. Cardiovasc Res (2022) 118(6):1548–63. doi: 10.1093/cvr/cvab134
206. Lilja S, Li X, Smelik M, Lee EJ, Loscalzo J, Marthanda PB, et al. Multi-organ single-cell analysis reveals an on/off switch system with potential for personalized treatment of immunological diseases. Cell Rep Med (2023) 4(3):100956. doi: 10.1016/j.xcrm.2023.100956
207. Leduc A, Khoury L, Cantlon J, Khan S, and Slavov N. Massively parallel sample preparation for multiplexed single-cell proteomics using nPOP. Nat Protoc (2024) 19(12):3750–76. doi: 10.1038/s41596-024-01033-8
208. Huffman RG, Leduc A, Wichmann C, Di Gioia M, Borriello F, Specht H, et al. Prioritized mass spectrometry increases the depth, sensitivity and data completeness of single-cell proteomics. Nat Methods (2023) 20(5):714–22. doi: 10.1038/s41592-023-01830-1
209. Hill MC, Kadow ZA, Long H, Morikawa Y, Martin TJ, Birks EJ, et al. Integrated multi-omic characterization of congenital heart disease. Nature (2022) 608(7921):181–91. doi: 10.1038/s41586-022-04989-3
210. Iqbal F, Schlotter F, Becker-Greene D, Lupieri A, Goettsch C, Hutcheson JD, et al. Sortilin enhances fibrosis and calcification in aortic valve disease by inducing interstitial cell heterogeneity. Eur Heart J (2023) 44(10):885–98. doi: 10.1093/eurheartj/ehac818
211. Bartosovic M, Kabbe M, and Castelo-Branco G. Single-cell CUT&Tag profiles histone modifications and transcription factors in complex tissues. Nat Biotechnol (2021) 39(7):825–35. doi: 10.1038/s41587-021-00869-9
212. Rodriques SG, Stickels RR, Goeva A, Martin CA, Murray E, Vanderburg CR, et al. Slide-seq: a scalable technology for measuring genome-wide expression at high spatial resolution. Science (2019) 363(6434):1463–7. doi: 10.1126/science.aaw1219
213. Stoeckius M, Hafemeister C, Stephenson W, Houck-Loomis B, Chattopadhyay PK, Swerdlow H, et al. Simultaneous epitope and transcriptome measurement in single cells. Nat Methods (2017) 14(9):865–8. doi: 10.1038/nmeth.4380
214. Tang SS, Guillermier C, Wang M, Poczatek JC, Suzuki N, Loscalzo J, et al. Quantitative imaging of selenoprotein with multi-isotope imaging mass spectrometry (MIMS). Surf Interface Anal (2014) 46(Suppl 1):154–7. doi: 10.1002/sia.5625
215. Cairns JL, Huber J, Lewen A, Jung J, Maurer SJ, Bausbacher T, et al. Mass-guided single-cell MALDI imaging of low-mass metabolites reveals cellular activation markers. Adv Sci (Weinh) (2025) 12(5):e2410506. doi: 10.1002/advs.202410506
216. Forte D, Pellegrino RM, Falvo P, Garcia-Gonzalez P, Alabed HBR, Maltoni F, et al. Parallel single-cell metabolic analysis and extracellular vesicle profiling reveal vulnerabilities with prognostic significance in acute myeloid leukemia. Nat Commun (2024) 15(1):10878. doi: 10.1038/s41467-024-55231-9
217. Rappez L, Stadler M, Triana S, Gathungu RM, Ovchinnikova K, Phapale P, et al. SpaceM reveals metabolic states of single cells. Nat Methods (2021) 18(7):799–805. doi: 10.1038/s41592-021-01198-0
218. Tang F, Barbacioru C, Wang Y, Nordman E, Lee C, Xu N, et al. mRNA-Seq whole-transcriptome analysis of a single cell. Nat Methods (2009) 6(5):377–82. doi: 10.1038/nmeth.1315
219. Regev A, Teichmann SA, Lander ES, Amit I, Benoist C, Birney E, et al. The human cell atlas. eLife (2017) 6:e27041. doi: 10.7554/eLife.27041
220. Jovic D, Liang X, Zeng H, Lin L, Xu F, and Luo Y. Single-cell RNA sequencing technologies and applications: a brief overview. Clin Transl Med (2022) 12(3):e694. doi: 10.1002/ctm2.694
221. Cusanovich DA, Hill AJ, Aghamirzaie D, Daza RM, Pliner HA, Berletch JB, et al. A single-cell atlas of in vivo mammalian chromatin accessibility. Cell (2018) 174(5):1309–24.e18. doi: 10.1016/j.cell.2018.06.052
222. Cao J, Packer JS, Ramani V, Cusanovich DA, Huynh C, Daza R, et al. Comprehensive single-cell transcriptional profiling of a multicellular organism. Science (2017) 357(6352):661–7. doi: 10.1126/science.aam8940
223. Jones RC, Karkanias J, Krasnow MA, Pisco AO, Quake SR, Salzman J, et al. The Tabula Sapiens: a multiple-organ, single-cell transcriptomic atlas of humans. Science (2022) 376(6594):eabl4896. doi: 10.1126/science.abl4896
224. Han X, Zhou Z, Fei L, Sun H, Wang R, Chen Y, et al. Construction of a human cell landscape at single-cell level. Nature (2020) 581(7808):303–9. doi: 10.1038/s41586-020-2157-4
225. Kalucka J, de Rooij LPMH, Goveia J, Rohlenova K, Dumas SJ, Meta E, et al. Single-cell transcriptome atlas of murine endothelial cells. Cell (2020) 180(4):764–79.e20. doi: 10.1016/j.cell.2020.01.015
226. Schupp JC, Adams TS, Cosme C Jr, Raredon MSB, Yuan Y, Omote N, et al. Integrated single-cell atlas of endothelial cells of the human lung. Circulation (2021) 144(4):286–302. doi: 10.1161/CIRCULATIONAHA.120.052318
227. Travaglini KJ, Nabhan AN, Penland L, Sinha R, Gillich A, Sit RV, et al. A molecular cell atlas of the human lung from single-cell RNA sequencing. Nature (2020) 587(7835):619–25. doi: 10.1038/s41586-020-2922-4
228. Gawel DR, Serra-Musach J, Lilja S, Aagesen J, Arenas A, Asking B, et al. A validated single-cell-based strategy to identify diagnostic and therapeutic targets in complex diseases. Genome Med (2019) 11(1):47. doi: 10.1186/s13073-019-0657-3
229. Lun ATL, McCarthy DJ, and Marioni JC. A step-by-step workflow for low-level analysis of single-cell RNA-seq data with bioconductor. F1000Res (2016) 5:2122. doi: 10.12688/f1000research.9501.2
230. Amezquita RA, Lun ATL, Becht E, Carey VJ, Carpp LN, Geistlinger L, et al. Orchestrating single-cell analysis with Bioconductor. Nat Methods (2020) 17(2):137–45. doi: 10.1038/s41592-019-0654-x
231. Xu Y, Tan Y, Zhang X, Cheng M, Hu J, Liu J, et al. Comprehensive identification of immuno-related transcriptional signature for active pulmonary tuberculosis by integrated analysis of array and single cell RNA-seq. J Infect (2022) 85(5):534–44. doi: 10.1016/j.jinf.2022.08.017
232. Fox A, Dutt TS, Karger B, Obregón-Henao A, Anderson GB, and Henao-Tamayo M. Acquisition of high-quality spectral flow cytometry data. Curr Protoc Cytom (2020) 93(1):e74. doi: 10.1002/cpcy.74
233. Radtke AJ, Chu CJ, Yaniv Z, Yao L, Marr J, Beuschel RT, et al. IBEX: an iterative immunolabeling and chemical bleaching method for high-content imaging of diverse tissues. Nat Protoc (2022) 17(2):378–401. doi: 10.1038/s41596-021-00644-9
234. Gordon KS, Kyung T, Perez CR, Holec PV, Ramos A, Zhang AQ, et al. Screening for CD19-specific chimaeric antigen receptors with enhanced signalling via a barcoded library of intracellular domains. Nat BioMed Eng (2022) 6(7):855–66. doi: 10.1038/s41551-022-00896-0
235. Xu C, He J, Wang H, Zhang Y, Wu J, Zhao L, et al. Single-cell transcriptomic analysis identifies an immune-prone population in erythroid precursors during human ontogenesis. Nat Immunol (2022) 23(7):1109–20. doi: 10.1038/s41590-022-01245-8
236. Lin JR, Fallahi-Sichani M, Chen JY, and Sorger PK. Cyclic immunofluorescence (CycIF), a highly multiplexed method for single-cell imaging. Curr Protoc Chem Biol (2016) 8(4):251–64. doi: 10.1002/cpch.14
237. Ranzoni AM, Tangherloni A, Berest I, Riva SG, Myers B, Strzelecka PM, et al. Integrative single-cell RNA-seq and ATAC-seq analysis of human developmental hematopoiesis. Cell Stem Cell (2021) 28(3):472–87.e7. doi: 10.1016/j.stem.2020.11.015
238. Ma S, Zhang B, LaFave LM, Earl AS, Chiang Z, Hu Y, et al. Chromatin potential identified by shared single-cell profiling of RNA and chromatin. Cell (2020) 183(4):1103–16.e20. doi: 10.1016/j.cell.2020.09.056
239. Tan WLW, Seow WQ, Zhang A, Rhee S, Wong WH, Greenleaf WJ, et al. Current and future perspectives of single-cell multi-omics technologies in cardiovascular research. Nat Cardiovasc Res (2023) 2(1):20–34. doi: 10.1038/s44161-022-00205-7
240. Eberhardt N and Giannarelli C. How single-cell technologies have provided new insights into atherosclerosis. Arterioscler Thromb Vasc Biol (2022) 42(3):243–52. doi: 10.1161/ATVBAHA.121.315849
241. de Winther MPJ, Bäck M, Evans P, Gomez D, Goncalves I, Jørgensen HF, et al. Translational opportunities of single-cell biology in atherosclerosis. Eur Heart J (2023) 44(14):1216–30. doi: 10.1093/eurheartj/ehac686
242. Skylaki S, Hilsenbeck O, and Schroeder T. Challenges in long-term imaging and quantification of single-cell dynamics. Nat Biotechnol (2016) 34(11):1137–44. doi: 10.1038/nbt.3713
243. Chang Q, Ornatsky OI, Siddiqui I, Loboda A, Baranov VI, and Hedley DW. Imaging mass cytometry. Cytomet A (2017) 91(2):160–9. doi: 10.1002/cyto.a.23053
244. Lin JR, Izar B, Wang S, Yapp C, Mei S, Shah PM, et al. Highly multiplexed immunofluorescence imaging of human tissues and tumors using t-CyCIF and conventional optical microscopes. eLife (2018) 7:e31657. doi: 10.7554/eLife.31657
245. Codeluppi S, Borm LE, Zeisel A, La Manno G, van Lunteren JA, Svensson CI, et al. Spatial organization of the somatosensory cortex revealed by osmFISH. Nat Methods (2018) 15(11):932–5. doi: 10.1038/s41592-018-0175-z
246. Heath JR, Ribas A, and Mischel PS. Single-cell analysis tools for drug discovery and development. Nat Rev Drug Discov (2016) 15(3):204–16. doi: 10.1038/nrd.2015.16
247. Keating SM, Taylor DL, Plant AL, Litwack ED, Kuhn P, Greenspan EJ, et al. Opportunities and challenges in implementation of multiparameter single cell analysis platforms for clinical translation. Clin Transl Sci (2018) 11(3):267–76. doi: 10.1111/cts.12536
248. Gaudillière B, Fragiadakis GK, Bruggner RV, Nicolau M, Finck R, Tingle M, et al. Clinical recovery from surgery correlates with single-cell immune signatures. Sci Transl Med (2014) 6(255):255ra131. doi: 10.1126/scitranslmed.3009701
249. Levitin HM, Yuan J, and Sims PA. Single-cell transcriptomic analysis of tumor heterogeneity. Trends Cancer (2018) 4(4):264–8. doi: 10.1016/j.trecan.2018.02.003
250. Rajkomar A, Dean J, and Kohane I. Machine learning in medicine. N Engl J Med (2019) 380(14):1347–58. doi: 10.1056/NEJMra1814259
251. Nwanosike EM, Conway BR, Merchant HA, and Hasan SS. Potential applications and performance of machine learning techniques and algorithms in clinical practice: a systematic review. Int J Med Inform (2022) 159:104679. doi: 10.1016/j.ijmedinf.2021.104679
252. Barnett GO, Cimino JJ, Hupp JA, and Hoffer EP. DXplain: an evolving diagnostic decision-support system. JAMA (1987) 258(1):67–74. doi: 10.1001/jama.258.1.67
253. Kuperman GJ, Gardner RM, and Pryor TA. HELP: a dynamic hospital information system. New York, NY: Springer-Verlag (1991).
254. Johnson KW, Glicksberg BS, Hodos RA, Shameer K, and Dudley JT. Causal inference on electronic health records to assess blood pressure treatment targets: an application of the parametric g formula. Pac Symp Biocomput (2018) 23:180–91. doi: 10.1142/9789813235533_0017
255. Shameer K, Johnson KW, Glicksberg BS, Dudley JT, and Sengupta PP. Machine learning in cardiovascular medicine: are we there yet? Heart (2018) 104(14):1156–64. doi: 10.1136/heartjnl-2017-311198
256. Lopez-Jimenez F, Attia Z, Arruda-Olson AM, Carter R, Chareonthaitawee P, Jouni H, et al. Artificial intelligence in cardiology: present and future. Mayo Clin Proc (2020) 95(5):1015–39. doi: 10.1016/j.mayocp.2020.01.038
257. Olsen CR, Mentz RJ, Anstrom KJ, Page D, and Patel PA. Clinical applications of machine learning in the diagnosis, classification, and prediction of heart failure. Am Heart J (2020) 229:1–17. doi: 10.1016/j.ahj.2020.07.009
258. Rasmy L, Wu Y, Wang N, Geng X, Zheng WJ, Wang F, et al. A study of generalizability of recurrent neural network-based predictive models for heart failure onset risk using a large and heterogeneous EHR data set. J BioMed Inform (2018) 84:11–6. doi: 10.1016/j.jbi.2018.06.011
259. Choi E, Schuetz A, Stewart WF, and Sun J. Using recurrent neural network models for early detection of heart failure onset. J Am Med Inform Assoc (2017) 24(2):361–70. doi: 10.1093/jamia/ocw112
260. Raghunath S, Ulloa Cerna AE, Jing L, vanMaanen DP, Stough J, Hartzel DN, et al. Prediction of mortality from 12-lead electrocardiogram voltage data using a deep neural network. Nat Med (2020) 26(6):886–91. doi: 10.1038/s41591-020-0870-z
261. Zhao J, Feng Q, Wu P, Lupu RA, Wilke RA, Wells QS, et al. Learning from longitudinal data in electronic health record and genetic data to improve cardiovascular event prediction. Sci Rep (2019) 9(1):717. doi: 10.1038/s41598-018-36745-x
262. Weng SF, Reps J, Kai J, Garibaldi JM, and Qureshi N. Can machine-learning improve cardiovascular risk prediction using routine clinical data? PloS One (2017) 12(4):e0174944. doi: 10.1371/journal.pone.0174944
263. Haro Alonso D, Wernick MN, Yang Y, Germano G, Berman DS, and Slomka P. Prediction of cardiac death after adenosine myocardial perfusion SPECT based on machine learning. J Nucl Cardiol (2019) 26(5):1746–54. doi: 10.1007/s12350-018-1250-7
264. Hannun AY, Rajpurkar P, Haghpanahi M, Tison GH, Bourn C, Turakhia MP, et al. Cardiologist-level arrhythmia detection and classification in ambulatory electrocardiograms using a deep neural network. Nat Med (2019) 25(1):65–9. doi: 10.1038/s41591-018-0268-3
265. Siontis KC, Noseworthy PA, Attia ZI, and Friedman PA. Artificial intelligence-enhanced electrocardiography in cardiovascular disease management. Nat Rev Cardiol (2021) 18(7):465–78. doi: 10.1038/s41569-020-00503-2
266. Arsanjani R, Xu Y, Dey D, Vahistha V, Shalev A, Nakanishi R, et al. Improved accuracy of myocardial perfusion SPECT for detection of coronary artery disease by machine learning in a large population. J Nucl Cardiol (2013) 20(4):553–62. doi: 10.1007/s12350-013-9706-2
267. Siegersma KR, Leiner T, Chew DP, Appelman Y, Hofstra L, and Verjans JW. Artificial intelligence in cardiovascular imaging: state of the art and implications for the imaging cardiologist. Neth Heart J (2019) 27(9):403–13. doi: 10.1007/s12471-019-01311-1
268. de Siqueira VS, Borges MM, Furtado RG, Dourado CN, and da Costa RM. Artificial intelligence applied to support medical decisions for the automatic analysis of echocardiogram images: a systematic review. Artif Intell Med (2021) 120:102165. doi: 10.1016/j.artmed.2021.102165
269. Leclerc S, Smistad E, Pedrosa J, Ostvik A, Cervenansky F, Espinosa F, et al. Deep learning for segmentation using an open large-scale dataset in 2D echocardiography. IEEE Trans Med Imaging (2019) 38(9):2198–210. doi: 10.1109/TMI.2019.2900516
270. Madani A, Ong JR, Tibrewal A, and Mofrad MRK. Deep echocardiography: data-efficient supervised and semi-supervised deep learning towards automated diagnosis of cardiac disease. NPJ Digit Med (2018) 1:59. doi: 10.1038/s41746-018-0065-x
271. van Velzen SGM, Lessmann N, Velthuis BK, Bank IEM, van den Bongard DHJG, Leiner T, et al. Deep learning for automatic calcium scoring in CT: validation using multiple cardiac CT and chest CT protocols. Radiology (2020) 295(1):66–79. doi: 10.1148/radiol.2020191621
272. Zreik M, van Hamersvelt RW, Khalili N, Wolterink JM, Voskuil M, Viergever MA, et al. Deep learning analysis of coronary arteries in cardiac CT angiography for detection of patients requiring invasive coronary angiography. IEEE Trans Med Imaging (2020) 39(5):1545–57. doi: 10.1109/TMI.2019.2953054
273. Zhang N, Yang G, Gao Z, Xu C, Zhang Y, Shi R, et al. Deep learning for diagnosis of chronic myocardial infarction on nonenhanced cardiac cine MRI. Radiology (2019) 291(3):606–17. doi: 10.1148/radiol.2019182304
274. Bernard O, Lalande A, Zotti C, Cervenansky F, Yang X, Heng PA, et al. Deep learning techniques for automatic MRI cardiac multi-structures segmentation and diagnosis: is the problem solved? IEEE Trans Med Imaging (2018) 37(11):2514–25. doi: 10.1109/TMI.2018.2837502
275. Cikes M, Sanchez-Martinez S, Claggett B, Duchateau N, Piella G, Butakoff C, et al. Machine learning-based phenogrouping in heart failure to identify responders to cardiac resynchronization therapy. Eur J Heart Fail (2019) 21(1):74–85. doi: 10.1002/ejhf.1333
276. Segar MW, Patel KV, Ayers C, Basit M, Tang WHW, Willett D, et al. Phenomapping of patients with heart failure with preserved ejection fraction using machine learning-based unsupervised cluster analysis. Eur J Heart Fail (2020) 22(1):148–58. doi: 10.1002/ejhf.1621
277. Verdonschot JAJ, Merlo M, Dominguez F, Wang P, Henkens MTHM, Adriaens ME, et al. Phenotypic clustering of dilated cardiomyopathy patients highlights important pathophysiological differences. Eur Heart J (2021) 42(2):162–74. doi: 10.1093/eurheartj/ehaa841
278. Galli E, Bourg C, Kosmala W, Oger E, and Donal E. Phenomapping heart failure with preserved ejection fraction using machine learning cluster analysis: prognostic and therapeutic implications. Heart Fail Clin (2021) 17(3):499–518. doi: 10.1016/j.hfc.2021.02.010
279. Cheng J, Novati G, Pan J, Bycroft C, Žemgulytė A, Applebaum T, et al. Accurate proteome-wide missense variant effect prediction with AlphaMissense. Science (2023) 381(6664):eadg7492. doi: 10.1126/science.adg7492
280. Burris JF and Puglisi JT. Impact of federal regulatory changes on clinical pharmacology and drug development: the common rule and the 21st century cures act. J Clin Pharmacol (2018) 58(3):281–5. doi: 10.1002/jcph.1026
281. Spatz ES, Ginsburg GS, Rumsfeld JS, and Turakhia MP. Wearable digital health technologies for monitoring in cardiovascular medicine. N Engl J Med (2024) 390(4):346–56. doi: 10.1056/NEJMra2301903
282. Ng K, Steinhubl SR, deFilippi C, Dey S, and Stewart WF. Early detection of heart failure using electronic health records: practical implications for time before diagnosis, data diversity, data quantity, and data density. Circ Cardiovasc Qual Outcomes (2016) 9(6):649–58. doi: 10.1161/CIRCOUTCOMES.116.002797
283. Deo RC and Nallamothu BK. Learning about machine learning: the promise and pitfalls of big data and the electronic health record. Circ Cardiovasc Qual Outcomes (2016) 9(6):618–20. doi: 10.1161/CIRCOUTCOMES.116.003308
284. Erin Lehr Angelini. IBM Watson health and the broad institute launch initiative to help clinicians predict the risk of cardiovascular disease with genomics and AI. PR Newswire (2019). Available at: https://www.prnewswire.com/news-releases/ibm-watson-health-and-the-broad-institute-launch-initiative-to-help-clinicians-predict-the-risk-of-cardiovascular-disease-with-genomics-and-ai-300794505.html
285. Agrawal R. Microsoft AI network for cardiology with Apollo Hospitals to bring new insights in predicting population-based heart diseases. Microsoft News Center India (2018). Available at: https://news.microsoft.com/en-in/features/microsoft-ai-network-healthcare-apollo-hospitals-cardiac-disease-prediction/
286. Poplin R, Varadarajan AV, Blumer K, Liu Y, McConnell MV, Corrado GS, et al. Prediction of cardiovascular risk factors from retinal fundus photographs via deep learning. Nat BioMed Eng (2018) 2(3):158–64. doi: 10.1038/s41551-018-0195-0
287. Turakhia MP, Desai M, Hedlin H, Rajmane A, Talati N, Ferris T, et al. Rationale and design of a large-scale, app-based study to identify cardiac arrhythmias using a smartwatch: the Apple Heart Study. Am Heart J (2019) 207:66–75. doi: 10.1016/j.ahj.2018.09.002
288. Paul SM, Mytelka DS, Dunwiddie CT, Persinger CC, Munos BH, Lindborg SR, et al. How to improve R&D productivity: the pharmaceutical industry’s grand challenge. Nat Rev Drug Discov (2010) 9(3):203–14. doi: 10.1038/nrd3078
289. Sholl DS and Steckel JA. Density functional theory: a practical introduction. Hoboken, NJ: John Wiley & Sons (2009). doi: 10.1002/9780470447710
290. Yaffe D, Cohen Y, Espinosa G, Arenas A, and Giralt F. A fuzzy ARTMAP based on quantitative structure-property relationships (QSPRs) for predicting aqueous solubility of organic compounds. J Chem Inf Comput Sci (2001) 41(5):1177–207. doi: 10.1021/ci010323u
291. Verma J, Khedkar VM, and Coutinho EC. 3D-QSAR in drug design–a review. Curr Top Med Chem (2010) 10(1):95–115. doi: 10.2174/156802610790232260
292. Lamb J, Crawford ED, Peck D, Modell JW, Blat IC, Wrobel MJ, et al. The Connectivity Map: using gene-expression signatures to connect small molecules, genes, and disease. Science (2006) 313(5795):1929–35. doi: 10.1126/science.1132939
293. Subramanian A, Narayan R, Corsello SM, Peck DD, Natoli TE, Lu X, et al. A next generation connectivity map: L1000 platform and the first 1,000,000 profiles. Cell (2017) 171(6):1437–52.e17. doi: 10.1016/j.cell.2017.10.049
294. Halu A, Chelvanambi S, Decano JL, Matamalas JT, Whelan M, Asano T, et al. Integrating pharmacogenomics and cheminformatics with diverse disease phenotypes for cell type-guided drug discovery. Genome Med (2025) 17(1):7. doi: 10.1186/s13073-025-01431-x
295. Duan Q, Reid SP, Clark NR, Wang Z, Fernandez NF, Rouillard AD, et al. L1000CDS2: Lincs L1000 characteristic direction signatures search engine. NPJ Syst Biol Appl (2016) 2:16015. doi: 10.1038/npjsba.2016.15
296. Musa A, Ghoraie LS, Zhang SD, Glazko G, Yli-Harja O, Dehmer M, et al. A review of connectivity map and computational approaches in pharmacogenomics. Brief Bioinform (2017) 18(5):903. doi: 10.1093/bib/bbx023
297. Iwata A, Chelvanambi S, Asano T, Whelan M, Nakamura Y, Aikawa E, et al. Gene expression profiles of precursor cells identify compounds that reduce NRP1 surface expression in macrophages: implication for drug repositioning for COVID-19. Front Cardiovasc Med (2024) 11:1438396. doi: 10.3389/fcvm.2024.1438396
298. Swinney DC. Phenotypic vs. target-based drug discovery for first-in-class medicines. Clin Pharmacol Ther (2013) 93(4):299–301. doi: 10.1038/clpt.2012.236
299. Swinney DC and Anthony J. How were new medicines discovered? Nat Rev Drug Discov (2011) 10(7):507–19. doi: 10.1038/nrd3480
300. Chen R, Liu X, Jin S, Lin J, and Liu J. Machine learning for drug-target interaction prediction. Molecules (2018) 23(9):2208. doi: 10.3390/molecules23092208
301. Sachdev K and Gupta MK. A comprehensive review of feature based methods for drug target interaction prediction. J BioMed Inform (2019) 93:103159. doi: 10.1016/j.jbi.2019.103159
302. Polishchuk PG, Madzhidov TI, and Varnek A. Estimation of the size of drug-like chemical space based on GDB-17 data. J Comput Aid Mol Des (2013) 27(8):675–9. doi: 10.1007/s10822-013-9672-4
303. Ezzat A, Wu M, Li XL, and Kwoh CK. Computational prediction of drug-target interactions using chemogenomic approaches: an empirical survey. Brief Bioinform (2019) 20(4):1337–57. doi: 10.1093/bib/bby002
304. Lim S, Lu Y, Cho CY, Sung I, Kim J, Kim Y, et al. A review on compound-protein interaction prediction methods: data, format, representation and model. Comput Struct Biotechnol J (2021) 19:1541–56. doi: 10.1016/j.csbj.2021.03.004
305. Bagherian M, Sabeti E, Wang K, Sartor MA, Nikolovska-Coleska Z, and Najarian K. Machine learning approaches and databases for prediction of drug-target interaction: a survey paper. Brief Bioinform (2021) 22(1):247–69. doi: 10.1093/bib/bbz157
306. Huang K, Fu T, Glass LM, Zitnik M, Xiao C, and Sun J. DeepPurpose: a deep learning library for drug-target interaction prediction. Bioinformatics (2021) 36(22–23):5545–7. doi: 10.1093/bioinformatics/btaa1005
307. Cheng Y, Gong Y, Liu Y, Song B, and Zou Q. Molecular design in drug discovery: a comprehensive review of deep generative models. Brief Bioinform (2021) 22(6):bbab344. doi: 10.1093/bib/bbab344
308. Tong X, Liu X, Tan X, Li X, Jiang J, Xiong Z, et al. Generative models for de novo drug design. J Med Chem (2021) 64(19):14011–27. doi: 10.1021/acs.jmedchem.1c00927
309. Bian Y and Xie XQ. Generative chemistry: drug discovery with deep learning generative models. J Mol Model (2021) 27(3):71. doi: 10.1007/s00894-021-04674-8
310. Shayakhmetov R, Kuznetsov M, Zhebrak A, Kadurin A, Nikolenko S, Aliper A, et al. Molecular generation for desired transcriptome changes with adversarial autoencoders. Front Pharmacol (2020) 11:269. doi: 10.3389/fphar.2020.00269
311. Méndez-Lucio O, Baillif B, Clevert DA, Rouquié D, and Wichard J. De novo generation of hit-like molecules from gene expression signatures using artificial intelligence. Nat Commun (2020) 11(1):10. doi: 10.1038/s41467-019-13807-w
312. Jumper J, Evans R, Pritzel A, Green T, Figurnov M, Ronneberger O, et al. Highly accurate protein structure prediction with AlphaFold. Nature (2021) 596(7873):583–9. doi: 10.1038/s41586-021-03819-2
313. Thornton JM, Laskowski RA, and Borkakoti N. AlphaFold heralds a data-driven revolution in biology and medicine. Nat Med (2021) 27(10):1666–9. doi: 10.1038/s41591-021-01533-0
314. Pirillo A, Catapano AL, and Norata GD. Monoclonal antibodies in the management of familial hypercholesterolemia: focus on PCSK9 and ANGPTL3 inhibitors. Curr Atheroscler Rep (2021) 23(12):79. doi: 10.1007/s11883-021-00972-x
315. Békés M, Langley DR, and Crews CM. Protac targeted protein degraders: the past is prologue. Nat Rev Drug Discov (2022) 21(3):181–200. doi: 10.1038/s41573-021-00371-6
316. Falese JP, Donlic A, and Hargrove AE. Targeting RNA with small molecules: from fundamental principles towards the clinic. Chem Soc Rev (2021) 50(4):2224–43. doi: 10.1039/d0cs01261k
317. Zhu Y, Zhu L, Wang X, and Jin H. RNA-based therapeutics: an overview and prospectus. Cell Death Dis (2022) 13(7):644. doi: 10.1038/s41419-022-05075-2
318. Oostveen RF, Khera AV, Kathiresan S, Stroes ESG, Fitzgerald K, Harms MJ, et al. New approaches for targeting PCSK9: small-interfering ribonucleic acid and genome editing. Arterioscler Thromb Vasc Biol (2023) 43(7):1081–92. doi: 10.1161/ATVBAHA.122.317963
319. Crooke ST, Baker BF, Crooke RM, and Liang XH. Antisense technology: an overview and prospectus. Nat Rev Drug Discov (2021) 20(6):427–53. doi: 10.1038/s41573-021-00162-z
320. Pickar-Oliver A and Gersbach CA. The next generation of CRISPR-Cas technologies and applications. Nat Rev Mol Cell Biol (2019) 20(8):490–507. doi: 10.1038/s41580-019-0131-5
321. Zhou J and Rossi J. Aptamers as targeted therapeutics: current potential and challenges. Nat Rev Drug Discov (2017) 16(3):181–202. doi: 10.1038/nrd.2016.199
322. Highleyman L. FDA approves fomivirsen, famciclovir, and thalidomide. Food and Drug Administration. Beta (1998) 5
323. Yu AM and Tu MJ. Deliver the promise: RNAs as a new class of molecular entities for therapy and vaccination. Pharmacol Ther (2022) 230:107967. doi: 10.1016/j.pharmthera.2021.107967
324. Fogacci F, Ferri N, Toth PP, Ruscica M, Corsini A, and Cicero AFG. Efficacy and safety of mipomersen: a systematic review and meta-analysis of randomized clinical trials. Drugs (2019) 79(7):751–66. doi: 10.1007/s40265-019-01114-z
325. Ray KK, Wright RS, Kallend D, Koenig W, Leiter LA, Raal FJ, et al. Two phase 3 trials of inclisiran in patients with elevated LDL cholesterol. N Engl J Med (2020) 382(16):1507–19. doi: 10.1056/NEJMoa1912387
326. Tsimikas S, Karwatowska-Prokopczuk E, Gouni-Berthold I, Tardif JC, Baum SJ, Steinhagen-Thiessen E, et al. Lipoprotein(a) reduction in persons with cardiovascular disease. N Engl J Med (2020) 382(3):244–55. doi: 10.1056/NEJMoa1905239
327. Kristen AV, Ajroud-Driss S, Conceição I, Gorevic P, Kyriakides T, and Obici L. Patisiran, an RNAi therapeutic for the treatment of hereditary transthyretin-mediated amyloidosis. Neurodegener Dis Manag (2019) 9(1):5–23. doi: 10.2217/nmt-2018-0033
328. Solomon SD, Adams D, Kristen A, Grogan M, González-Duarte A, Maurer MS, et al. Effects of patisiran, an RNA interference therapeutic, on cardiac parameters in patients with hereditary transthyretin-mediated amyloidosis. Circulation (2019) 139(4):431–43. doi: 10.1161/CIRCULATIONAHA.118.035831
329. Seal RL, Chen LL, Griffiths-Jones S, Lowe TM, Mathews MB, O’Reilly D, et al. A guide to naming human non-coding RNA genes. EMBO J (2020) 39(6):e103777. doi: 10.15252/embj.2019103777
330. Uchida S and Adams JC. Physiological roles of non-coding RNAs. Am J Physiol Cell Physiol (2019) 317(1):C1–2. doi: 10.1152/ajpcell.00114.2019
331. Das S, Shah R, Dimmeler S, Freedman JE, Holley C, Lee JM, et al. Noncoding RNAs in cardiovascular disease: current knowledge, tools and technologies for investigation, and future directions: a scientific statement from the American Heart Association. Circ Genom Precis Med (2020) 13(4):e000062. doi: 10.1161/HCG.0000000000000062
332. Laggerbauer B and Engelhardt S. MicroRNAs as therapeutic targets in cardiovascular disease. J Clin Invest (2022) 132(11):e159179. doi: 10.1172/JCI159179
333. Winkle M, El-Daly SM, Fabbri M, and Calin GA. Noncoding RNA therapeutics - challenges and potential solutions. Nat Rev Drug Discov (2021) 20(8):629–51. doi: 10.1038/s41573-021-00219-z
334. Chen Y, Li Z, Chen X, and Zhang S. Long non-coding RNAs: from disease code to drug role. Acta Pharm Sin B (2021) 11(2):340–54. doi: 10.1016/j.apsb.2020.10.001
335. Uchida S and Dimmeler S. Long noncoding RNAs in cardiovascular diseases. Circ Res (2015) 116(4):737–50. doi: 10.1161/CIRCRESAHA.116.302521
336. Yeh CF, Chang YE, Lu CY, Hsuan CF, Chang WT, and Yang KC. Expedition to the missing link: long noncoding RNAs in cardiovascular diseases. J BioMed Sci (2020) 27(1):48. doi: 10.1186/s12929-020-00647-w
337. Ezike TC, Okpala US, Onoja UL, Nwike CP, Ezeako EC, Okpara OJ, et al. Advances in drug delivery systems, challenges and future directions. Heliyon (2023) 9(6):e17488. doi: 10.1016/j.heliyon.2023.e17488
338. Vargason AM, Anselmo AC, and Mitragotri S. The evolution of commercial drug delivery technologies. Nat BioMed Eng (2021) 5(9):951–67. doi: 10.1038/s41551-021-00698-w
339. Paunovska K, Loughrey D, and Dahlman JE. Drug delivery systems for RNA therapeutics. Nat Rev Genet (2022) 23(5):265–80. doi: 10.1038/s41576-021-00439-4
340. Spicer AJ, Colcomb PA, and Kraft A. Mind the gap: closing the growing chasm between academia and industry. Nat Biotechnol (2022) 40(11):1693–6. doi: 10.1038/s41587-022-01543-4
341. Frye S, Crosby M, Edwards T, and Juliano R. US academic drug discovery. Nat Rev Drug Discov (2011) 10(6):409–10. doi: 10.1038/nrd3462
342. Huryn DM, Resnick LO, and Wipf P. Contributions of academic laboratories to the discovery and development of chemical biology tools. J Med Chem (2013) 56(18):7161–76. doi: 10.1021/jm400132d
343. Brigham and Women’s Hospital. New lab brings BWH, Japanese researchers together [online] (2010). Available at: https://www.bwhpublicationsarchives.org/DisplayCRN.aspx?articleid=1837
344. Brigham and Women’s Hospital. A $75 million ‘brave idea’ to end coronary heart disease [online] (2010). Available at: https://give.brighamandwomens.org/brave-idea-end-coronary-heart-disease/
345. Munos BH and Chin WW. How to revive breakthrough innovation in the pharmaceutical industry. Sci Transl Med (2011) 3(89):89cm16. doi: 10.1126/scitranslmed.3002273
346. Yildirim O, Gottwald M, Schüler P, and Michel MC. Opportunities and challenges for drug development: public-private partnerships, adaptive designs and big data. Front Pharmacol (2016) 7:461. doi: 10.3389/fphar.2016.00461
347. Nwaka S and Ridley RG. Virtual drug discovery and development for neglected diseases through public-private partnerships. Nat Rev Drug Discov (2003) 2(11):919–28. doi: 10.1038/nrd1230
348. Tralau-Stewart CJ, Wyatt CA, Kleyn DE, and Ayad A. Drug discovery: new models for industry-academic partnerships. Drug Discov Today (2009) 14(1–2):95–101. doi: 10.1016/j.drudis.2008.10.003
349. Ferrero E, Brachat S, Jenkins JL, Marc P, Skewes-Cox P, Altshuler RC, et al. Ten simple rules to power drug discovery with data science. PloS Comput Biol (2020) 16(8):e1008126. doi: 10.1371/journal.pcbi.1008126
350. Jodogne S. Client-side application of deep learning models through teleradiology. Stud Health Technol Inform (2023) 302:997–1001. doi: 10.3233/SHTI230325
351. Dong E, Du H, and Gardner L. An interactive web-based dashboard to track COVID-19 in real time. Lancet Infect Dis (2020) 20(5):533–4. doi: 10.1016/S1473-3099(20)30120-1
352. Cauchemez S, Cossu G, Delzenne N, Elinav E, Fassin D, Fischer A, et al. Standing the test of COVID-19: charting the new frontiers of medicine. Front Sci (2024) 2:1236919. doi: 10.3389/fsci.2024.1236919
353. Moreira ED Jr, Kitchin N, Xu X, Dychter SS, Lockhart S, Gurtman A, et al. Safety and efficacy of a third dose of BNT162b2 Covid-19 vaccine. N Engl J Med (2022) 386(20):1910–21. doi: 10.1056/NEJMoa2200674
354. Arbel R, Hammerman A, Sergienko R, Friger M, Peretz A, Netzer D, et al. BNT162b2 vaccine booster and mortality due to Covid-19. N Engl J Med (2021) 385(26):2413–20. doi: 10.1056/NEJMoa2115624
355. Weinreich DM, Sivapalasingam S, Norton T, Ali S, Gao H, Bhore R, et al. REGEN-COV antibody combination and outcomes in outpatients with Covid-19. N Engl J Med (2021) 385(23):e81. doi: 10.1056/NEJMoa2108163
356. O’Brien MP, Forleo-Neto E, Sarkar N, Isa F, Hou P, Chan KC, et al. Effect of subcutaneous casirivimab and imdevimab antibody combination vs placebo on development of symptomatic COVID-19 in early asymptomatic SARS-CoV-2 infection: a randomized clinical trial. JAMA (2022) 327(5):432–41. doi: 10.1001/jama.2021.24939
357. Alameh MG, Tombácz I, Bettini E, Lederer K, Sittplangkoon C, Wilmore JR, et al. Lipid nanoparticles enhance the efficacy of mRNA and protein subunit vaccines by inducing robust T follicular helper cell and humoral responses. Immunity (2022) 55(6):1136–8. doi: 10.1016/j.immuni.2022.05.007
358. Karikó K, Whitehead K, and van der Meel R. What does the success of mRNA vaccines tell us about the future of biological therapeutics? Cell Syst (2021) 12(8):757–8. doi: 10.1016/j.cels.2021.07.005
359. Ward B, Yombi JC, Balligand JL, Cani PD, Collet JF, de Greef J, et al. HYGIEIA: hypothesizing the genesis of infectious diseases and epidemics through an integrated systems biology approach. Viruses (2022) 14(7):1373. doi: 10.3390/v14071373
360. Sun J, Aikawa M, Ashktorab H, Beckmann ND, Enger ML, Espinosa JM, et al. A multi-omics strategy to understand PASC through the RECOVER cohorts: a paradigm for a systems biology approach to the study of chronic conditions. Front Syst Biol (2025) 4:1422384. doi: 10.3389/fsysb.2024.1422384
361. Reese JT, Blau H, Casiraghi E, Bergquist T, Loomba JJ, Callahan TJ, et al. Generalisable long COVID subtypes: findings from the NIH N3C and RECOVER programmes. EBiomedicine (2023) 87:104413. doi: 10.1016/j.ebiom.2022.104413
362. Alfano V and Ercolano S. The efficacy of lockdown against COVID-19: a cross-country panel analysis. Appl Health Econ Health Policy (2020) 18(4):509–17. doi: 10.1007/s40258-020-00596-3
363. García-Basteiro AL and Legido-Quigley H. Evaluation of the COVID-19 response in Spain: principles and requirements. Lancet Public Health (2020) 5(11):e575. doi: 10.1016/S2468-2667(20)30208-5
364. Thawonmas R, Hashizume M, and Kim Y. Projections of temperature-related suicide under climate change scenarios in Japan. Environ Health Perspect (2023) 131(11):117012. doi: 10.1289/EHP11246
365. Agache I, Akdis C, Akdis M, Al-Hemoud A, Annesi-Maesano I, Balmes J, et al. Immune-mediated disease caused by climate change-associated environmental hazards: mitigation and adaptation. Front Sci (2024) 2:1279192. doi: 10.3389/fsci.2024.1279192
366. Anonymous. The best medicine for improving global health? Reduce inequality. Nature (2023) 619:221. doi: 10.1038/d41586-023-02251-y
367. Fuster V and Vedanthan R. Cardiovascular disease and the UN Millennium Development Goals: time to move forward. Nat Clin Pract Cardiovasc Med (2008) 5(10):593. doi: 10.1038/ncpcardio1353
368. MalariaWorld. The best medicine for improving global health? Reduce inequality [online] (2023). Available at: https://malariaworld.org/blogs/the-best-medicine-for-improving-global-health-reduce-inequality
369. World Health Organization. Global action plan for the prevention and control of noncommunicable diseases 2013-2020. Geneva: WHO (2013). Available at: https://www.who.int/publications/i/item/9789241506236
370. World Health Organization. Invisible numbers: the true extent of noncommunicable diseases and what to do about them. Geneva: WHO (2022). Available at: https://www.who.int/publications/i/item/9789240057661
371. Kones R and Rumana U. Cardiometabolic diseases of civilization: history and maturation of an evolving global threat. An update and call to action. Ann Med (2017) 49(3):260–74. doi: 10.1080/07853890.2016.1271957
372. Mocumbi AO. Cardiovascular health care in low- and middle-income countries. Circulation (2024) 149(8):557–9. doi: 10.1161/CIRCULATIONAHA.123.065717
373. Qureshi NQ, Mufarrih SH, Bloomfield GS, Tariq W, Almas A, Mokdad AH, et al. Disparities in cardiovascular research output and disease outcomes among high-, middle- and low-income countries - an analysis of global cardiovascular publications over the last decade (2008-2017). Glob Heart (2021) 16(1):4. doi: 10.5334/gh.815
374. Adeyi O, Yadav P, and Kazatchkine M. Frontiers of medicine unveiled: equitable access is an imperative. Front Sci (2024) 2:1422583. doi: 10.3389/fsci.2024.1422583
375. Global Preparedness Monitoring Board. A fragile state of preparedness: 2023 report on the state of the world’s preparedness. Geneva: World Health Organization (2023). Available at: https://www.gpmb.org/reports/m/item/a-fragile-state-of-preparedness-2023-report-on-the-state-of-the-worlds-preparedness
376. World Health Organization. Pandemic prevention, preparedness and response agreement [online] (2025). Available at: https://www.who.int/news-room/questions-and-answers/item/pandemic-prevention--preparedness-and-response-accord
377. Dzau V, Fuster V, Frazer J, and Snair M. Investing in global health for our future. N Engl J Med (2017) 377(13):1292–6. doi: 10.1056/NEJMsr1707974
378. Davis A, Asma S, Blecher M, Benn C, Ezoe S, Fogstad H, et al. Investing in the future of global health. Lancet (2024) 404(10462):1500–3. doi: 10.1016/S0140-6736(24)02191-3
Keywords: cardiovascular disease, cardiology, precision medicine, systems biology, network medicine, heterogeneity, artificial intelligence
Citation: Aikawa M, Sonawane AR, Chelvanambi S, Asano T, Halu A, Matamalas JT, Singh SA, Uchida S, Aikawa E, Arenas A, Balligand J-L, Giannarelli C, MacRae CA, Morgan NV, Oury C, Tevaearai Stahel H and Loscalzo J. Precision cardiovascular medicine: shifting the innovation paradigm. Front Sci (2025) 3:1474469. doi: 10.3389/fsci.2025.1474469
Received: 01 August 2024; Accepted: 23 July 2025;
Published: 07 October 2025.
Edited by:
Steffen Erhard Petersen, Queen Mary University of London, United KingdomReviewed by:
Rainer Schulz, University of Giessen, GermanyHugo Ten Cate, Maastricht University Medical Centre, Netherlands
Felix B. Engel, University Hospital Erlangen, Germany
Copyright © 2025 Aikawa, Sonawane, Chelvanambi, Asano, Halu, Matamalas, Singh, Uchida, Aikawa, Arenas, Balligand, Giannarelli, MacRae, Morgan, Oury, Tevaearai Stahel and Loscalzo. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Masanori Aikawa, bWFpa2F3YUBid2guaGFydmFyZC5lZHU=
†These authors have contributed equally to this work