 Lydie-Stella Koutika1*
Lydie-Stella Koutika1* Maurício Rumenos Guidetti Zagatto2
Maurício Rumenos Guidetti Zagatto2 Arthur Prudêncio de Araujo Pereira3
Arthur Prudêncio de Araujo Pereira3 Michael Miyittah4
Michael Miyittah4 Silvia Tabacchioni5
Silvia Tabacchioni5 Annamaria Bevivino5
Annamaria Bevivino5 Cornelia Rumpel6
Cornelia Rumpel6- 1CRDPI, Pointe-Noire, Congo
- 2DungTech Biofertilizers LTDA, Santa Bárbara D'Oeste, Piracicaba, Brazil
- 3Department of Soil Science, Federal University of Ceará, Fortaleza, Brazil
- 4Department of Environmental Science, University of Cape Coast, Cape Coast, Ghana
- 5Department for Sustainability, Italian National Agency for New Technologies, Energy and Sustainable Economic Development, ENEA, Casaccia Research Center, Rome, Italy
- 6Centre National de la Recherche Scientifique (CNRS), Paris, France
Plant and/or crop growth rely on nutrient dynamics driven by specific soil biota in different environments. This mini-review aims to provide an overview of interactions between soil organisms, nutrient dynamics, and C sequestration. To this end, we investigated published results from three forest plantations (eucalyptus monocultures and mixed plantations with N2-fixing acacia) on tropical nutrient-poor soils. One case study is located in Central Africa (Congolese coastal plains) and two others in South America (Southeastern Brazil). Overall, the studies showed that soil biota activity exerted positive effects on (i) C accretion, as both soil carbon and belowground and aboveground biomass are driven and enhanced by soil biota; and (ii) on nutrient dynamics and biogeochemical cycles in nutrient-poor soil of tropical ecosystems, which are boosted following C accumulation. On the other hand, the pedoclimatic environment may potentially impact soil functioning of mixed-species plantations through its influence on the composition and activity of bacterial communities. Regardless of the potential risk of acacia invasiveness, benefits such as pulp, fuelwood, electric pole and non-timber products supply, have been reported in Central Africa. We, therefore, conclude that including N2 fixing trees in forestry plantations as reported in this mini-review helps strengthen the links between soil biota, nutrient and SOC dynamics in mixed-species plantations on tropical nutrient-poor soils.
Introduction
Ecosystem services of forest plantations especially when intercropping with N2-fixing trees have been evaluated in Central Africa (1). They may benefit both rural and urban populations (i.e., pulp, fuelwood, non-timber products supply and poles for electricity network), and indirectly be linked to land restoration, environmental services, and conservation of natural forests (1–3). Preference of native species to exotic ones must be considered to avoid the risk of invasiveness, and loss of biodiversity (2, 4).
The introduction of N2-fixing trees in forest plantations was also found to impact soil biota and nutrient dynamic and, therefore, ecosystem functioning and soil carbon sequestration. Interactions among soil biota, nutrient cycling and carbon sequestration have been investigated for decades in different geo-climatic zones (5–7). Soil biota, microbial biomass, community structure and function play a critical role in sustaining the fitness, development, and productivity of plants (8, 9). They regulate ecosystem processes involved in plant litter decomposition, soil organic matter (SOM) turnover, associated nutrient mineralization (10) and soil biodiversity (11). Numerous studies have already been conducted on soil biota and nutrient dynamics in the rhizosphere of acacia and eucalyptus forest plantations, (12–20).
Investigations on the relation of SOC to SOM dynamic on N and P cycles and their connections to soil organic carbon (SOC) sequestration have been performed (21, 22). Stoichiometric limitations (23, 24) for SOM stabilization processes in different ecosystems (3, 25–27) were observed. In highly weathered tropical soils, P availability is especially limited due to strong adsorption of this nutrient to the mineral phase (Al and Fe oxides). However, P availability may be enhanced by intercropping N2-fixing Acacia mangium with Eucalyptus grandis, due to the stimulation of the root colonization by arbuscular mycorrhizal fungi (AMF) and phosphatase activity in the soil (28). Nevertheless, there are still some gaps that need to be filled to better understand the strong link between biota and nutrient dynamics, boosting C sequestration in the mixed-species forest plantations on nutrient-poor tropical soils.
In particular, it is unclear how soil biota drives nutrient dynamics and SOC sequestration in mixed-species plantations on tropical nutrient-poor soils. Recently, it was suggested that soil biota may boost soil C sequestration in forest plantations intercropped with N2-fixing trees by enhancing above and belowground C allocations (29–31) and also reducing old C loss within the ecosystem (32). In general, in mixed plantations with N2 fixing trees, stimulated changes in microbial community activities have been observed (16, 20, 33, 34). This could lead to C and N accumulation (35) in litter and soil by creating distinct microbial communities for respective monocultures and benefitting soil P and nitrate content (33), while P cycling and P nutrition is enhanced by arbuscular mycorrhizal fungi (AMF) colonization and phosphatase activities (28). It also involves a positive balance of nutrients since N2-fixation from acacia benefits eucalyptus (30, 36, 37).
In this mini-review, three case-studies have been selected to give a detailed overview of the role of soil biota for nutrient dynamics and carbon sequestration on nutrient-poor tropical soils of acacia and eucalyptus forest plantations. To this end, we reviewed results from one study in Central Africa (Congolese coastal plains, Republic of the Congo), and two studies in South America (Itatinga, Southeastern Brazil). The first case-study reports the link between bacterial communities, nutrient dynamics and environment in the Congolese coastal plains (Central Africa). The two other case studies from Southeastern Brazil (South America) deal with bacterial, archaeal, and fungal communities related to nutrient cycling on the one hand, and soil faunal communities related to soil quality on the other hand. The three case-studies are characterized by the same experimental design of acacia and eucalyptus [E. urophilla x grandis (Congo) and E. grandis (Brazil)] plantations and different pedoclimatic conditions and forest management. Three main questions arise from the revision of the literature regarding the results of the three selected case-studies:
• How do N2-fixing trees influence the interaction between microbial communities and their impact on SOC and nutrient dynamics in mixed-species forest plantations?
• How are these interactions impacted by soil properties and pedoclimatic conditions?
How can N2-fixing trees be used to benefit other tropical forest plantations?
We hypothesize that based on similarities, differences and benefits (Table 1) of the practice introducing acacia in eucalyptus plantations in the three case- studies will strongly depend on soil pedoclimatic conditions but also on forest management.
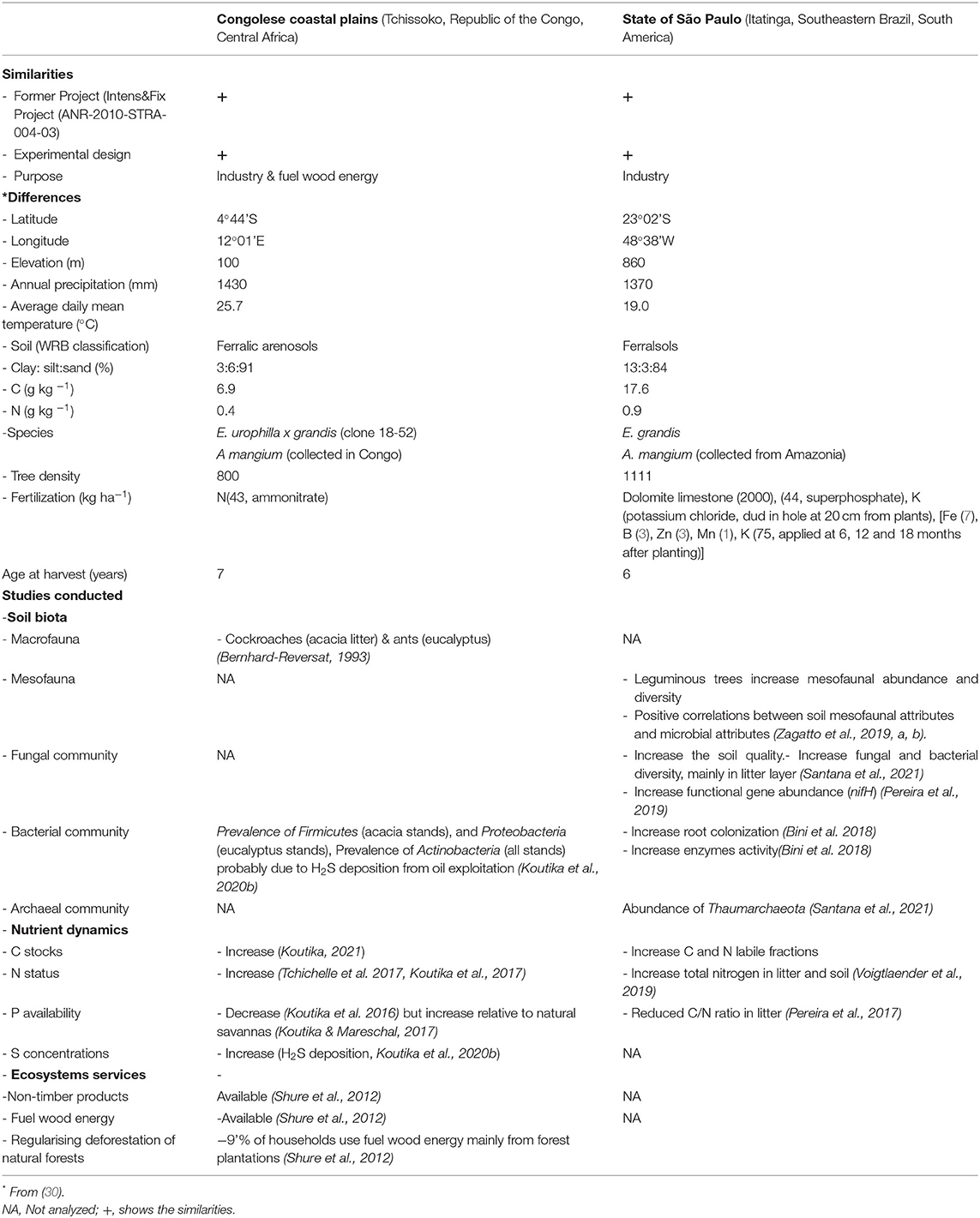
Table 1. Main characteristics of the two locations of the selected case studies.
Presentation of Case Studies
In Brazil, the largest eucalyptus producer in the world, eucalyptus plantations are often implemented on very nutrient-poor soils as monocultures, needing the continuous application of mineral fertilizers. A possible solution is the co-cultivation of eucalyptus and N2-fixing trees in mixed systems to provide an additional supply of N and P for eucalyptus (38, 39) and to create more heterogeneous systems i.e., higher microbial diversity, and supporting an efficient selection of beneficial microorganisms in the rhizosphere (19, 28).
Without neglecting the potential risk of threatening savanna ecosystems (40–42), afforestation of natural savannas on inherently nutrient-poor soils using eucalyptus started in the 1950s in the Congolese coastal plains, [Makany, 1964 cited in (43)]. This has been made to use unsuitable soils for agriculture, provide pulpwood for the industry, and fulfill the important production and consumption of fuelwood energy by the local populations i.e., preserving natural forests and halting the deforestation (1, 3). Since productivity of forest plantations declines after successive rotations and harvests (44), N2-fixing trees have been introduced in the 1990s to restore soil fertility, and improve and sustain forest productivity (12, 30, 45).
To the best of our knowledge, very few studies addressed soil biodiversity (bacterial, fungal and faunal) in the mixed-species plantations in the Congolese coastal plains. Bernhard-Reversat, (12) reported an enhanced activity of edaphic macroarthropod communities i.e., the dominance of cockroaches in acacia litter as opposed to ants in eucalyptus. In general, few investigations have been carried out in tropical environments and especially in sub-Saharan Africa. On the contrary, within the two case-studies in Southeastern Brazil, studies have been conducted on archaeal (20), fungal (35) and faunal communities (46), in addition to investigations on microbial activity, microbial communities, rhizosphere and or root AMF colonization and soil phosphatase activity (19, 28, 34, 35).
How Do N2-Fixing Trees Influence the Interaction Between Microbial Communities and Their Impact on Soc and Nutrient Dynamics in Mixed-Species Forest Plantations?
Bacterial communities structure in soil and litter layers was found to be related to the soil nutrient pools in the first case study at Itatinga (Southeastern Brazil). Indeed, in 27- and 39-months old acacia forest plantations, increased C and N contents in the organic soil fractions (including microbial biomass), bacterial diversity and richness (soil and litter), and nif H gene (related to nitrogen fixation) abundance was observed (34). Furthermore, Rhizobium, Bradyrhizobium and Sphingomonas showed a positive correlation with nif H and total soil N. Additional evidence shows that change in the soil microbial community composition in mixed acacia and eucalyptus plantations is correlated with increased C and N cycling (19).
Archaeal communities are also important contributors to nutrient cycling in mixed-species forest ecosystems. Santana et al. (20) reported that pure stands type influenced the archaeal community structure of the litter layer, as archaeal richness, diversity, and the relative abundance of Thaumarchaeota increased in 27- and 39-months old acacia plantations in Southeastern Brazil. It has also been found that archaeal structure responded to stand types in the litter layer, in which , total-N and the C/N ratio were the most important attributes for community groups differentiation. Young mixed plantations exert a low effect on soil microbial community structure, but archaeal communities may have an important role in nutrient cycling in the litter interface, especially related to the N cycle in the initial stages of tree development (20).
Acacia stimulates the soil phosphatase activity, thereby increasing root AMF colonization and, consequently, P nutrition to the surrounding trees in the same system (28). Pereira et al. (19) showed that the soil organic fraction, phosphorus inorganic fraction, total-P and acid phosphatase activity, were significantly higher in pure stands containing acacia at Itatinga (Southeastern Brazil). Total P, richness, and Shannon diversity of the fungi in the litter was significantly higher in these stands with an intermediate structure between the two pure stands. Also, mixed systems strongly correlated with P dynamics, particularly in the litter layer. Co-occurrence networks of fungal taxa became simpler in pure eucalyptus stands, whereas mixed counterparts showed a more connected and complex network. This result evidenced that mixed forest plantations promote positive responses in the fungal community connections, which are closely related to P availability in the system, prominently in the litter layer.
The second case-study in Southeastern Brazil reported an improvement in soil quality in stands containing acacia compared to pure eucalyptus stands due to their soil mesofaunal and microbial attributes (46). In these planted forests, chemical attributes (especially N, low C/N ratio and macronutrients) are strongly related to different mesofaunal orders in the litter layers (47). A higher diversity of mesofaunal orders in acacia monoculture and mixed-species stands than eucalyptus monoculture is probably due to the higher quality of the litter in stands containing acacia than in pure eucalyptus counterparts (34, 37).
Seasonality is another important factor for the organisms that inhabit the soil (48). Higher mesofauna density in soil during drought periods relative to mesofauna density in the litter at the same period in forest plantations of eucalyptus and acacia highlighted the soil's importance as a refuge for the invertebrate community in periods of water scarcity in Southeastern Brazil (47). Forest plantations presented higher mesofauna diversity, but a lower mesofauna density in litter when compared to no-tillage (15) once secondary forests in regeneration accumulate organic matter of higher quality and present higher niche diversity than agricultural systems.
In Central Africa (Congolese coastal plains), probably due to stimulated microbial activity (16) and enhanced P cycling, AMF colonization, phosphatase activities (28) and nutrient dynamics (N and P) were stimulated in acacia and eucalyptus plantations in the afforested ecosystems compared with native tropical savannas (49). An enhanced N dynamics was also reported in stands containing acacia relative to eucalyptus monocultures (50, 51). In addition, nuclear magnetic resonance (solid state 13C CPMASS and NMR and 31P-NMR) spectroscopy reported higher amounts of extractable inorganic P (litter and soil) in the stands containing acacia (43), probably due to high mineralization rate and the lack of soluble leaching during the drought period. This is probably related to greater biological activity (28), which would prevent the P losses in the soil and may indicate a mechanism able to sustain forest plantation through P demand (19, 25).
Further effects have been observed through increased C stocks in stands containing acacia in the Congolese coastal plains. As in other ecosystems within the world (24, 38, 39), carbon sequestration in biomass and soil does occur in acacia and eucalyptus plantations (30, 52). Greater soil carbon stock increments were estimated in pure acacia and mixed-species stands with 0.8 t ha−1 and 1.9 t ha−1, respectively, relative to eucalyptus stands (53). This potentially may contribute to climate change mitigation, and adaptation and resilience by creating healthier soils rich in C and other nutrients (3, 53).
How Are These Interactions Impacted by Soil Properties and Pedoclimatic Conditions?
Although the three case-studies showed similar design (Table 1), there are differences in main site characteristics such as latitude, longitude, annual precipitation, soil classification, tree density, and fertilization (30). From this mini-review, it is apparent that forest management, soil type and geo-climatic zone affect the links between soil biota and nutrient and SOC dynamics. Mixed plantations on poorest soils and poor forest management i.e., those of the Congolese coastal plains responded better to the introduction of Acacia mangium relative to the control (eucalyptus), as they showed more improved soil N status (50, 51), C stocks (52, 53), and stand wood biomass (30, 45) than the trials in Southeastern Brazil.
This may illustrate the potential impact of the environment on soil functioning and productivity in forest plantations. Sustaining forest plantations strongly relies on interactions among nutrient cycling, microbial communities and environment (54). Using meta-barcoding of the 16S rRNA bacterial genes, the predominance of Actinobacteria phylum, strongly correlated to sulfur (S) at all classes, has been observed in different stands of monoculture and mixed-species plantations in the Congolese coastal plains (55). This may be due to the potential impact of H2S deposition from oil exploration activities (from the end of the 1960s) on the environment i.e., the bacterial communities (Koutika, personal communication). It was previously observed that sulfur (S) has a potential to stimulate the growth of Gram-positive bacteria, fungi and Actinobacteria in subtropical forest soil (56), while the prevalence of hydrogen sulfide (H2S) depositions from industrial areas does increase the growth of Actinobacteria alone (57).
How Can the Introduction of N2-Fixing Trees Be Used to Benefit Other Tropical Forest Plantations?
In addition to above mentioned positive effects on soil microbial functioning, other benefits of the intercropping with N2-fixing trees include its impact on ecosystems services. Most of our knowledge on the effects of N2-fixing trees on ecosystem services derives from local or regional studies, with a great influence of the environmental conditions, history and cultural background of each region on these effects (58). Therefore, it is fundamental to identify synergies (like among soil fertility, soil formation and climate regulation) and trade-offs among the effects of N2-fixing trees on different ecosystem services. For example, the introduction of N2-fixing trees was found to affect the SOC stock, and the recalcitrant carbon chemical composition in Eucalyptus urophylla plantation in subtropical China (59). N2-fixing trees may attain a higher production, especially in infertile or degraded soils, explaining their contribution to soil formation, land restoration and erosion control and water regulation (58). Also, mixed forest plantations provide a wide range of social and environmental services by mitigating future wood shortage problems and producing a huge proportion of world industrial wood and other forest products (60). When compared with monospecific plantations, mixed forest plantations have been found to give more benefits in biodiversity, economy, forest health and occasionally in productivity, producing more pulp, fuelwood and non-timber products supply (1, 2). The mixed-species forest tree stands can be beneficial for both trees and ecosystems in many regions. Therefore, we could apply findings reported in this mini-review to other similar tropical forest ecosystems in Central Africa (3, 61), and or elsewhere in the world.
Concluding Remarks
This mini-review reports how soil biota drives and enhances nutrient dynamics and C sequestration in the mixed-species acacia and eucalyptus plantations on tropical nutrient-poor soils in three case-studies in Central Africa and South America. Soil properties and pedoclimatic conditions drive this interaction. However, enhanced nutrient dynamics and C sequestration induced by the swift change in microbial communities in the acacia–eucalyptus mixed forest plantations reported in the Brazilian case-studies, may explain enhanced soil C sequestration (all case-studies), C allocation in belowground (South America), and stand wood biomass (all case-studies). The mini-review also reports other ecosystem services of the practice directly or indirectly linked to the local populations (Central Africa) such as fuelwood energy, pole for electricity network and non-timber products. Finally, the mini-review highlights the effects of anthropogenic activities and environment on soil biota of forest plantations with further benefit on soil fertility improvement, land restoration and potentiality to resilience and adaptation to climate change.
Author Contributions
All authors listed have made a substantial, direct, and intellectual contribution to the work and approved it for publication.
Funding
Part of results from the three case-studies of this mini-review has been conducted in the frame of the Project -ANR-10-STRA-0004 (Intens & fix, Agence Nationale de la Recherche, France). The two case-studies from Brazil were supported by FAPESP (The São Paulo Research Foundation) (grant n°. 10/16623-9, 16/01636-4, 16/18944-3 and 17/16610-3). AB (Annamaria Bevivino) gratefully acknowledges funding from the Italian project Creazione di un HUB italiano a supporto della partecipazione dell'Italia alla Global Soil Partnership ed alla rete di eccellenza europea sulla ricerca sul suolo–SOIL-HUB, granted by the Italian Ministry of Agricultural, Food and Forestry Policies (Decreto del Dipartimento delle Politiche Europee ed Internazionali e dello Sviluppo Rurale - DISR IV Prot. 35851 del 5/11/2019), and the European Union's Horizon 2020 research and innovation programme under grant agreement no. 652615 (European Joint Programme SOIL).
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher's Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
1. Shure J, Marien JN, de Wasseige C, Drigo R, Salbitano F, Dirou S, et al. Contribution du bois énergie à La Satisfaction des besoins énergétiques des populations d'Afrique centrale: Perspectives pour une gestion durable des ressources disponibles. Chap. (2012) 5:109–22.
2. Lescuyer G, Karsenty A, Eba'a Atyi R. A new tool for sustainable forest management in Central Africa: payments for environmental services. In: de Wasseige C, Devers D, de Marcken P, Eba'a Atyi R, Nasi R, Mayaux P, editors. The Forests of the Congo Basin: state of the Forest 2008. Publications Office of the European Union (2009). p. 131–43.
3. Koutika L.-S., Taba K, Ndongo M, Kaonga M. Nitrogen-fixing trees increase organic carbon sequestration in forest and agroforestry ecosystems in the Congo basin. Reg Environ Change. (2021) 21:109. doi: 10.1007/s10113-021-01816-9
4. Koutika L.-S., Richardson DM. Acacia mangium Willd: Benefits and threats associated with its increasing use around the world (Review). Forest Ecosystems. (2019) 6:1–13. doi: 10.1186/s40663-019-0159-1
5. Gebremikael M, Steel H, Buchan D, Bert W, De Neve S. Nematodes enhance plant growth and nutrient uptake under C and N-rich conditions. Sci Rep. (2016) 6:32862. doi: 10.1038/srep32862
6. Barthod J, Dignac M-F, Le Mer G, Bottinelli N, Watteau F, Kögel-Knabner I, et al. How do earthworms affect organic matter decomposition in the presence of clay-sized minerals? Soil Biol Biochem. (2020) 143:107730. doi: 10.1016/j.soilbio.2020.107730
7. Angst G, Mueller CW, Prater I, Angst S, Frouz J, Jílková V, et al. Earthworms act as biochemical reactors to convert labile plant compounds into stabilized soil microbial necromass. Commun Biol. (2019) 2:441. doi: 10.1038/s42003-019-0684-z
8. Olivares F,0.L., Busato JG, Paula AM, Lima LS, Aguiar NO, Canellas LP. Plant growth promoting bacteria and humic substances: crop promotion and mechanisms of action. Chem Biol Technol Agric. (2017) 4:30. doi: 10.1186/s40538-017-0112-x
9. Mercado-Blanco J, Abrantes I, Barra Caracciolo A, Bevivino A, Ciancio A, Grenni P, et al. Belowground Microbiota and the Health of Tree Crops. Front Microbiol. (2018) 9:1006. doi: 10.3389/fmicb.2018.01006
10. Maaroufi NI, De Long JR. Global change impacts on forest soils: linkage between soil biota and carbon-nitrogen-phosphorus stoichiometry. Front For Glob Change. (2020) 3:16. doi: 10.3389/ffgc.2020.00016
11. FAO., ITPS., GSBI., CBD, and EC. State of knowledge of soil biodiversity - Status, challenges and potentialities, Report 2020. Rome: FAO. (2020).
12. Bernhard-Reversat F. Dynamics of litter and organic matter at the soil-litter interface of fast-growing tree plantations on sandy ferrallitic soils (Congo). Acta Oecol. (1993) 14:179–95.
13. Santos FM, Balieiro FC, Fontes MA, Chaer GM. Understanding the enhanced litter decomposition of mixed-species plantations of Eucalyptus and Acacia mangium. Plant Soil. (2018) 423. doi: 10.1007/s11104-017-3491-7
14. Pereira APA, de Andrade PA, Bini D, Durrer A, Robin A, Bouillet J-P, et al. Shifts in the bacterial community composition along deep soil profiles in monospecific and mixed stands of Eucalyptus grandis and Acacia mangium. PLoS ONE. (2017) 12:e0180371. doi: 10.1371/journal.pone.0180371
15. Zagatto MRG, Zanão Júnior LA, Pereira APA, Estrada-Bonilla G, Cardoso EJBN. Soil mesofauna in consolidated land use systems: how management affects soil and litter invertebrates. Sci Agric. (2018) 76:165–71. doi: 10.1590/1678-992x-2017-0139
16. Bini D, Figueiredo AF, da Silva MCP, de Figueiredo Vasconcellos RL, Cardoso. E. J. B. N. Microbial biomass and activity in litter during the initial development of pure and mixed plantations of Eucalyptus grandis and Acacia mangium. Rev Bras Cienc Solo. (2012) 37:76–85. doi: 10.1590/S0100-06832013000100008
17. Bini D, dos Santos CA, Bouillet J-P, de Morais Goncalves JL, Cardosa EJBN. Eucalyptus grandis and Acacia mangium in monoculture and intercropped plantations: evolution of soil and litter microbial and chemical attributes during early stages of plant development. Appl Soil Ecol. (2013) 63:57–66. doi: 10.1016/j.apsoil.2012.09.012
18. Cui J, Yuan X, Zhang Q, Zhou J, Lin K, Xu J, et al. Nutrient availability is a dominant predictor of soil bacterial and fungal community composition after nitrogen addition in subtropical acidic forests. PLoS ONE. (2021) 16:e0246263. doi: 10.1371/journal.pone.0246263
19. Pereira APA, Santana MC, Zagatto MRG, Brandani CB. Wang, J.-T., et al. Nitrogen-fixing trees in mixed forest systems regulate the ecology of fungal community and phosphorus cycling. Sci Total Environ. (2021) 758:143711. doi: 10.1016/j.scitotenv.2020.143711
20. Santana MC, Pereira APA, Souza DJA, Zagatto MRU, Viera VLDA, Wang JT. Shifts on archaeal community structure in pure and mixed Eucalyptus grandis and Acacia mangium plantations. Forest Ecol Manage. (2021) 492:119218. doi: 10.1016/j.foreco.2021.119218
21. Masoom H, Courtier-Murias D, Farooq H, Soong R, Kelleher BP, Zhang C, et al. Soil organic matter in its native state: unravelling the most complex biomaterial on earth. Environ Sci Technol. (2016) 50:1670–80. doi: 10.1021/acs.est.5b03410
22. Paul EA. The nature and dynamics of soil organic matter: plant inputs, microbial transformations, and organic matter stabilization. Soil Biol Biochem. (2016) 98:109–26. doi: 10.1016/j.soilbio.2016.04.001
23. Van Groningen W, van Kessel C, Hungate BA, Oenema O, Powlson DS, van Groenigen KJ. Sequestering soil organic carbon: a nitrogen dilemma. Environ Sci Technol. (2017) 51:4738–9. doi: 10.1021/acs.est.7b01427
24. Mayer M, Prescott CE, Abaker WEA, Augusto L, Cecillon L, Ferreira GWD, et al. Tamm Review: Influence of forest management activities on soil organic carbon stocks: A knowledge synthesis. Forest Ecol Manage. (2020) 466:11812. doi: 10.1016/j.foreco.2020.118127
25. Koutika L-S. Afforesting tropical savannas with Acacia mangium and eucalyptus improves soil P availability in Arenosols of the Congolese coastal plains. Geoderma Reg. (2019) 16:e00207. doi: 10.1016/j.geodrs.2019.e00207
26. Voigtlaender M, Brandani C, Caldeira D, Tardy F, Bouillet J-P, Goncalves JLM, et al. Nitrogen cycling in monospecific and mixed-species plantations of Acacia mangium and Eucalyptus at 4 sites in Brazil. Forest Ecol Manage. (2019) 436:56–67. doi: 10.1016/j.foreco.2018.12.055
27. Lu X, Vitousek PM, Mao Q, Gilliam FS, Luo Y, Turner BL, et al. Nitrogen deposition accelerates soil carbon sequestration in tropical forests. PNAS. (2021) 118:e2020790118. doi: 10.1073/pnas.2020790118
28. Bini D, dos Santos CA, da Silva MCP, Bonfim JA, Cardoso EJBN. Intercropping Acacia mangium stimulates AMF colonization and soil phosphatase activity in Eucalyptus grandis. Sci Agric. (2018) 75:102–10. doi: 10.1590/1678-992x-2016-0337
29. Nouvellon Y, Laclau J, Epron D, Le Maire G, Bonnefond J, Goncalves J, et al. Carbon allocations throughout the development of monospecific and mixedspecies plantations of Eucalyptus grandis and Acacia mangium in Brazil. Tree Physiol. (2012) 32:680–95. doi: 10.1093/treephys/tps041
30. Epron D, Nouvellon Y, Mareschal L, Moreira RM, Koutika L-S, Geneste B, et al. Partitioning of net primary production in Eucalyptus and Acacia stands and in mixed-species plantations: Two case-studies in contrasting tropical environments. Forest Ecol Manage. (2013) 301:102–11. doi: 10.1016/j.foreco.2012.10.034
31. Laclau J.-P., Nouvellon Y, Reine C, de Moraes Goncalves JL, Krushe AV, et al. Mixing Eucalyptus and Acacia trees leads to fine root over-yielding and vertical segregation between species. Oecologia. (2013) 172:903–913. doi: 10.1007/s00442-012-2526-2
32. Resh SC, Binkley D, Parrotta JA. Greater soil carbon sequestration under nitrogen-fixing trees compared with Eucalyptus species. Ecosystems. (2002) 5:217–231. doi: 10.1007/s10021-001-0067-3
33. Rachid C, Balieiro F, Peixoto R, Pinheiro Y, Piccolo M, Chaer G, et al. Mixed plantations can promote microbial integration and soil nitrate increases with changes in the N cycling genes. Soil Biol Biochem. (2013) 66:146–153. doi: 10.1016/j.soilbio.2013.07.005
34. Pereira APA, Durrer A, Gumiere T, Gonçalves JLM, Robin A. Bouillet JP, et al. Mixed Eucalyptus plantations induce changes in microbial communities and increase biological functions in the soil and litter layers. Forest Ecol Manage. (2019) 433:332–42. doi: 10.1016/j.foreco.2018.11.018
35. Pereira APA, Santana MC, Bonfim JA, Mescolotti DL, Cardoso EJBN. Digging deeper to study the distribution of mycorrhizal arbuscular fungi along the soil profile in pure and mixed Eucalyptus grandis and Acacia mangium plantations. Appl Soil Ecol. (2018) 128:1–11. doi: 10.1016/j.apsoil.2018.03.015
36. Paula RR, Bouillet J.-P, Trivelin PCO, Zeller B, Gonçalves JL, Nouvellon Y, et al. Evidence of short-term belowground transfer of nitrogen from Acacia mangium to Eucalyptus grandis trees in a tropical planted forest. Soil Biol Biochem. (2015) 91:99–108. doi: 10.1016/j.soilbio.2015.08.017
37. Santos FM, Chaer GM, Diniz AR, Balieiro F. d.C. Nutrient cycling over five years of mixed-species plantations of Eucalyptus and Acacia on a sandy tropical soil. Forest Ecol Manage. (2017) 384:110–121. doi: 10.1016/j.foreco.2016.10.041
38. Binkley D. Mixtures of N2-fixing and non- N2-fixing tree species. In: Cannell MGR, Malcom DC, Robertson PA, editors. The Ecology of Mixed Species Stands of Trees. Oxford: Blackwell Scientific Publications. (1992) p. 97-123. Available online at: http://www.agris.fao.org/agris-search/search.do?recordID=XF2016029053.
39. Forrester DI, Pares A, O'Hara C, Khanna PK, Bauhus J. Soil organic carbon is increased in mixed-species plantations of eucalyptus and nitrogen-fixing acacia. Ecosystems. (2013) 16:123–32. doi: 10.1007/s10021-012-9600-9
40. Parr CL, Lehmann CE, Bond WJ, Hoffmann WA, Andersen AN. Tropical grassy biomes: misunderstood, neglected, under threat. Trends Ecol Evol. (2014) 29:205–13. doi: 10.1016/j.tree.2014.02.004
41. Bond WJ, Stevens N, Midgley GF, Lehmann CER. The Trouble with Trees: Afforestation Plans for Africa. Trends Ecol Evol. (2019) 34:963–5. doi: 10.1016/j.tree.2019.08.003
42. Silveira FAO, Arruda AJ, Bond W, Durigan G, Fidelis A. Kirkman K, et al. Myth-busting tropical grassy biome restoration. Restor Ecol. (2020) 28:1067–73. doi: 10.1111/rec.13202
43. Koutika L-S, Cafiero L, Bevivino A, Merino A. Organic matter quality of forest floor as a driver of C and P dynamics in acacia and eucalypt plantations established on a Ferralic Arenosols, Congo. For Ecosyst. (2020) 7:40. doi: 10.1186/s40663-020-00249-w
44. Laclau JP, Ranger J, Deleporte P, Nouvellon Y, Saint Andr,é L, Marlet S, et al. (2005) Nutrient cycling in a clonal stand of eucalyptus and an adjacent savanna ecosystem in Congo: 3. Input-output budget and consequences for the sustainability of the plantations. Forest Ecol Manage. 210:375–91. doi: 10.1016/j.foreco.2005.02.028
45. Bouillet J.-P., Laclau JP, Gonçalves JLM, Voigtlaender M, Gava JL, et al. Eucalyptus and Acacia tree growth over entire rotation in single- and mixed-species plantations across five sites in Brazil and Congo. Forest Ecol Manage. (2013) 301:89–101. doi: 10.1016/j.foreco.2012.09.019
46. Zagatto MRG, Pereira APA, Souza AJ, Fabri RF, Baldesin LF, Pereira CM. Interactions between mesofauna, microbiological and chemical soil attributes in pure and intercropped Eucalyptus grandis and Acacia mangium plantations. For Ecol Manag. (2019) 433:240–7. doi: 10.1016/j.foreco.2018.11.008
47. Zagatto MRG, Pereira APA, Souza J, Pereira RF, Pereira CM, Baldesin LF. Acacia mangium increases the mesofauna density and diversity in the litter layer in Eucalyptus grandis plantations. Eur J Soil Biol. (2019) 94:1–8. doi: 10.1016/j.ejsobi.2019.103100
48. Zagatto MRG, Niva CC, Thomazini MJ, Baretta D, Santos A, Nadolny H, et al. Soil Invertebrates in Different Land Use Systems: How Integrated Production Systems Seasonality Affect Soil Mesofauna Communities. J Agric Sci Technol B. (2017) 7:150–61. doi: 10.17265/2161-6264/2017.03.003
49. Koutika L-S, Mareschal L. Planting acacia and eucalypt change P, N and C concentrations in POM of arenosols in the Congolese coastal plains. Geoderma Reg. (2017) 11:37–43. doi: 10.1016/j.geodrs.2017.07.009
50. Koutika L-S, Tchichelle SV, Mareschal L, Epron D. Nitrogen dynamics in a nutrient-poor soil under mixed-species plantations of eucalypts and acacias. Soil Biol Biochem. (2017) 108:84–90. doi: 10.1016/j.soilbio.2017.01.023
51. Tchichelle SV, Epron D, Mialoundama F, Koutika L.-S, Harmand JM, Bouillet JP, et al. Differences in nitrogen cycling and soil mineralization between a eucalypt plantation and a mixed eucalypt and Acacia mangium plantation on a sandy tropical soil. South For. (2017) 79:1–8. doi: 10.2989/20702620.2016.1221702
52. Koutika L-S, Epron D, Bouillet JP, Mareschal L. Changes in N and C concentrations, soil acidity and P availability in tropical mixed acacia and eucalypt plantations on a nutrient-poor sandy soil. Plant Soil. (2014) 379:205–16. doi: 10.1007/s11104-014-2047-3
53. Koutika L.-S. Soil fertility improvement of nutrient-poor and sandy soils in the Congolese coastal plains. FAO & ITPS. 2021. Recarbonizing global soils: A technical manual of recommended management practices. Rome: Forestry, wetlands, urban soils – Case studies. (2021) p. 4–13. doi.org/10.4060/cb6605en.
54. Brockerhoff EG, Barbaro L, Castagneyrol B, Forrester DI, Gardiner B, Gonzalez-Olabarria JR, et al. Forest biodiversity, ecosystem functioning and the provision of ecosystem services. Biodivers Conserv. (2017) 26:3005–35. doi: 10.1007/s10531-017-1453-2
55. Koutika L-S, Fiore A, Tabacchioni S, Aprea G, Pereira APA, Bevivino A. Influence of Acacia mangium on soil fertility and bacterial community in Eucalyptus Plantations in the Congolese Coastal Plains. Sustainability. (2020) 12:8763. doi: 10.3390/su12218763
56. Xu Y, Fan J, Ding W, Bol R, Chen Z. Luo J, et al. Stage-specific response of litter decomposition to N and S amendments in a subtropical forest soil. Biol Fertil Soils. (2016) 52:711–24. doi: 10.1007/s00374-016-1115-7
57. Dong Q, Shi H, Liu Y. Microbial character related sulfur cycle under dynamic environmental factors based on the microbial population analysis in sewerage system. Front Microbiol. (2017) 8:64. doi: 10.3389/fmicb.2017.00064
58. Castro-Díez P, Vaz AS, Silva J. S., van Loo M, Alonso Á, Aponte C, et al. Global effects of non-native tree species on multiple ecosystem services. Biol Rev. (2019) 94:1477–1501. doi: 10.1111/brv.12511
59. Huang X, Liu S, You Y, Wen Y, Wang H, Wang Y. Microbial community and associated enzymes activity influence soil carbon chemical composition in Eucalyptus urophylla plantation with mixing N2-fixing species in subtropical China. Plant Soil. (2017) 414:199–212. doi: 10.1007/s11104-016-3117-5
60. Liu CLC, Kuchma O, Krutovsky KV. Mixed-species versus monocultures in plantation forestry: Development, benefits, ecosystem services and perspectives for the future. Glob Ecol Conserv. (2018) 15:e00419. doi: 10.1016/j.gecco.2018.e00419
Keywords: soil biota, soil nutrients, soil carbon sequestration, crop/plant growth, climate change mitigation
Citation: Koutika L-S, Zagatto MRG, Pereira APdA, Miyittah M, Tabacchioni S, Bevivino A and Rumpel C (2021) Does the Introduction of N2-Fixing Trees in Forest Plantations on Tropical Soils Ameliorate Low Fertility and Enhance Carbon Sequestration via Interactions Between Biota and Nutrient Availability? Case Studies From Central Africa and South America. Front. Soil Sci. 1:752747. doi: 10.3389/fsoil.2021.752747
Received: 03 August 2021; Accepted: 04 November 2021;
Published: 03 December 2021.
Edited by:
Benjamin N. Sulman, Oak Ridge National Laboratory (DOE), United StatesReviewed by:
Gerardo Moreno, University of Extremadura, SpainRiccardo Spaccini, University of Naples Federico II, Italy
Copyright © 2021 Koutika, Zagatto, Pereira, Miyittah, Tabacchioni, Bevivino and Rumpel. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Lydie-Stella Koutika, bHNfa291dGlrYUB5YWhvby5jb20=