 Jose Isidro Beltran-Medina
Jose Isidro Beltran-Medina Sofiane Ouazaa*Nesrine ChaaliCamilo Ignacio Jaramillo BarriosKelly Johana Pedroza Berrío
Sofiane Ouazaa*Nesrine ChaaliCamilo Ignacio Jaramillo BarriosKelly Johana Pedroza Berrío Jose Alvaro Hoyos Cartagena
Jose Alvaro Hoyos Cartagena John Edinson Calderón Carvajal
John Edinson Calderón Carvajal- Corporación Colombiana de Investigación Agropecuaria – Agrosavia, CI Nataima, Espinal, Colombia
Basil (Ocimum basilicum L.) is a high-value aromatic crop with growing global demand, and optimizing its yield under tropical conditions is critical for sustainable agriculture. This study aimed to (1) quantify basil crop coefficient (Kc) and evapotranspiration (ETc) via lysimeters and (2) characterize soil physical–chemical variability across three Tolima (Colombia) region sites: Mariquita, Honda, and El Espinal. Crop evapotranspiration, measured via lysimeters, peaked at 7.41 mm day-1 during maturity, with a total crop water requirement of 228.82 mm. Crop coefficients varied dynamically by stage, with values of 0.75, 0.98, and 0.76 during establishment, peak growth, and senescence, respectively. Historical climate analysis revealed a bimodal rainfall distribution modulated by ENSO phenomenon, with El Niño-La Niña phases, with significant impacts on crop water availability. Soil analyses showed that Mariquita soils are higher in total porosity Tp (47.80%), organic matter (2.42 g 100g-1), field capacity FC (31.62%), and available water (3.59%), whereas El Espinal showed higher bulk density (1.65 gr cm-3) and permanent wilting point PWP (21.99%), constraining water availability. Honda soils presented intermediate conditions but were notable for higher cation exchange capacity CEC (9.55 cmol kg-1) and moderate organic matter content (1.56 g 100g-1), supporting balanced nutrient retention. Cultivated plots across sites showed increased phosphorus and copper relative to adjacent natural areas, reflecting fertilization practices. These results highlight the need for precision irrigation scheduling and site-specific soil management to maximize water productivity and yield stability. Our findings provide a baseline for adapting basil production systems to climatic variability in tropical dry regions.
1 Introduction
Basil (Ocimum basilicum L.) is an aromatic herb that represents a significant component for the global market and is used as a raw material in food and non-food industrial products. Global export data indicate that basil experienced a 2% annual growth rate between 2012 and 2021, reaching a total commercialization of 675,043 t by 2021 (1). Analysis of commercial trends, applications, and value-added processes for basil and its byproducts highlights that Germany, Morocco, and Poland are leading exporters with stable production outputs for the fresh leaf market. In contrast, India dominates the essential oils sector, cultivating approximately 3,000 ha of the global 5,000 ha dedicated to basil production (1, 2).
Basil is experiencing robust demand in international markets, with the United States emerging as a particularly significant destination. Recent studies have confirmed the economic viability of basil cultivation for exports, highlighting Colombia strong potential to become a key supplier in this sector (3, 4). Basil is primarily marketed and exported in two forms: fresh and dried leaves. Additionally, the extraction of essential oils from basil presents a significant commercial opportunity, given their extensive use in the pharmaceutical, cosmetic, and food industries (21). Additionally, small-scale farmers often led by women, displaced individuals, and young entrepreneurs are increasingly adopting basil cultivation for export (5). Their involvement highlights how basil production can serve as an economic lifeline for vulnerable communities, providing them with new revenue streams and contributing to social cohesion.
Basil is a widely cultivated in central Colombia, particularly in regions like Tolima (65.87%) and Cundinamarca (32.08%), which together account for the majority of the country basil production. It grows at elevations ranging from sea level up to 1,700 m.a.s.l. Between 2008 and 2020, basil production surged from 56 t to 4,097 t, with the cultivated area expanding from 26 ha into 543 ha, and higher average yields from 1.8 t ha-1 to 6.5 t ha-1, as reported by MADR in 2019 and 2022. Basil plays a significant role in Colombia agricultural landscape, contributing to both the economy and traditional medicine practices. Colombia has established targeted agricultural policies to advance the cultivation and market integration of aromatic crops, including basil (Ocimum basilicum L.), through collaborative research frameworks and value chain optimization strategies (6).
One of the main limitations of basil varieties production is related to the water deficit, which significantly affects fresh and dry matter and essential oil yield, considered one of the main markets for basil (7). Many studies (8–11); focused on the behavior of basil genotypes and its essential oil components under water stress, fertilization, planting density, and essential oil percentage. The commercial value of fresh basil in international markets is conditioned by its color and aroma retention (12), however, its postharvest viability is constrained by rapid deterioration and susceptibility to chilling injury (13). For this reason, and to ensure the production of specific metabolites, plants need adequate amounts of nutrients and appropriate biotic and abiotic conditions to achieve optimal growth conditions and better control in fertilization. Additionally, water stress (75% and 50% FC) and storage time (up to 7 days) significantly alter the volatile profiles of basil genotypes (14). Fertilization practices, particularly imbalanced NPK ratios, further exacerbate water and nutrient inefficiencies by prioritizing vegetative growth over essential oil synthesis (15). The use of chemical fertilizers in basil cultivation results in high production costs and contributes to soil and water pollution. This needs the adoption of biofertilizers and organic substrates, which may enhance plant nutrition and growth, thereby increasing overall productivity (16). Continued research into optimized cultivation methods and sustainable practices will help ensure that basil production not only meets market demands but also supports the long-term economic and environmental sustainability of the agricultural sector in Colombia.
Implementing sustainable agronomic practices is crucial to maintain optimal production levels and preserve the quality of the crop. Precise irrigation using soil moisture sensors and lysimeter based evapotranspiration (ETc) modeling can optimize irrigation on phenological stages (17). Similarly, organic amendments and optimized fertilization (18, 19) can enhance soil structure, nutrient retention, and antioxidant capacity. Such strategies remain understudied in Tolima context, where bimodal rainfall (1200–1500 mm annually) and high evapotranspiration (4–5 mm day-1; 20) demand region specific solutions. The Tolima region, located in the west-central part of Colombia, is one of most promising areas for basil cultivation (21). Tolima diverse soil properties and climatic conditions warrant a comprehensive evaluation to determine their suitability for the optimized growth of this aromatic herb. The region has 22 basil farms registered for basil export with the Colombian Agricultural Institute, with an average of 88 ha planted and an approximate production of 1,467,482 kg of basil year-1 (22). The basil production system is managed in open fields as a monoculture and for export type (23). Detailed assessments should focus on soil fertility, moisture retention, temperature fluctuations, and other environmental factors that critically influence basil productivity and quality.
Climatic bimodal regime occurs predominantly in the Tolima region where rainfall peaks are prominent in the season March–April-May (MAM) and September–October-November (SON) (24). Several authors have pointed out the meridional migration of the Intertropical Convergence Zone (ITCZ) as the main driver of the annual cycle of Colombia hydro climatology (25). Naranjo Bedoya et al. (26) have linked the influence of the bimodal regime to the occurrence of ENSO phenomena. On the other hand, the edaphoclimatic variability encompasses the complex interactions between soil and climate factors that influence ecosystem dynamics and species distribution (27). Climate variability substantially affects agricultural production, with temperature increases leading to decreased yields, while irrigation districts show limited effectiveness as adaptation measures (28).
ENSO variability significantly modulates evapotranspiration dynamics across tropical agricultural ecosystems, exhibiting pronounced spatial heterogeneity. Current literature on Ocimum basilicum has predominantly addressed physiological responses to water deficit and evapotranspiration rather than examining direct relationships with El Niño/La Niña oscillations (29–31). Basil evapotranspiration rates range from 4.8-9.4 mm day over a 49day cycle, with crop coefficients varying from 1.5-2.8 depending on growth stage and environmental conditions (30). In contrast, ENSO research has been performed on other agricultural systems. El Niño negatively affects common bean yields, while precipitation deviations from long-term values reduce yields but increase farmer incomes due to price compensation (32). Historical land cover changes and global warming during ENSO events show complex interactions, with deforestation reducing precipitation in the Andes and Caribbean Coast, while La Niña intensifies these effects compared to El Niño (33). In northern Colombia’s warm climate, evapotranspiration variations significantly impact native pasture forage production, with simulation tools proving valuable for water resource management during dry seasons and drought events influenced by ENSO (34).
Despite extensive research on basil agronomy (7, 35), critical knowledge gaps persist for tropical dry regions like Tolima, Colombia. First, the crop coefficients (Kc) and evapotranspiration (ETc) of basil under ENSO-modulated bimodal rainfall, a hallmark of Tolima climate, remain unquantified, hindering precision irrigation. Second, while soil compaction and nutrient imbalances globally limit yields (36, 37), site specific variability in Tolima major production zones (Mariquita, Honda, El Espinal) has not been characterized, preventing targeted soil management. This study addresses these gaps by.
(1) establishing specific Kc and ETc via lysimeter under observed ENSO phases and (2) mapping soil physicochemical constraints to water and nutrient use efficiency, with implications for climate resilient basil production.
2 Materials and methods
2.1 Study area
We selected three commercial basil farms per site (nine in total) for soil surveys based on their export grade basil production and contrasting soil profiles of the Tolima region, Colombia. It is characterized by a humid tropical climate, with an average annual temperature of 26°C and an average annual precipitation of 1500 mm. Precipitation follows a bimodal pattern, with peak rainfall occurring between April–May and September–October (38).
2.2 Climate data
Auxiliary climatic data for the three localities was obtained from IDEAM (39) and Corpoica (6). Climate anomalies affecting basil cultivation were evaluated using more than 30 years of temperature and precipitation records (1980–2011). The auxiliary climatic data was first screened for outliers and temporal trends. Climate anomalies were assessed using the Oceanic Niño Index (ONI), where values above +0.5 °C for five consecutive months or more indicate El Niño conditions (dry year), while values below -0.5°C for the same duration signify La Niña conditions (wet year) (38, 40, 41). These anomalies represent the percentage difference calculated by comparing the monthly variable values during El Niño and La Niña scenarios against the historical monthly mean of the variable (38).
2.3 Basil crop water requirements
A daily basis water balance evaluation was conducted to determine the albahaca crop evapotranspiration. Three volumetric (drainage) lysimeters were installed at the Nataima Research Center of Agrosavia, located in the El Espinal study site (4°11′ 28.7″N, 74°57′39.2″ W; 323 m altitude) to measure daily crop evapotranspiration (ETc). The lysimeters consisted of reinforced fiber glass with a diameter of 0.8 m and a depth of 1.30 m. A layer of expanded clay, 0.05 m thick, was covered by a repacked soil profile of 1.20 m. All lysimeters were filled up with undisturbed soil so that the crop grows at the same field conditions. At the bottom of the lysimeter a pipe serving as drainage outlet connected the lysimeter with a drainage reservoir. The set-up was covered at a height of 4 m by a sheet of transparent plastic to protect the assembly against precipitation. Volumetric lysimeters were employed to obtain direct, physically-based measurements of basil evapotranspiration (ETc). This method was selected over relying solely on the FAO-56 Penman-Monteith equation with standard crop coefficients (Kc) because it provides an integrated and real-time measurement that captures the specific soil-plant-atmosphere interactions of the local environment (17). This approach is particularly valuable under ENSO-modulated climates, as it directly accounts for the crop physiological response to variable conditions such as water stress during El Niño-related droughts.
The approach outlined by Evett et al. (17) was adopted in this study to estimate the water balance. Crop evapotranspiration (ET) was determined through a mass balance method (Equation 1), utilizing the soil water balance model applied to a daily time period within a control volume defined by the lysimeter, characterized by a specific depth and lateral dimensions:
where I is irrigation, P is precipitation (precipitation was neglected due to the lysimeter was covered, avoiding rain), F is flux into (taken as positive) or out of the control volume considered here as the drainage volume, R is the sum of run on and runoff equating here to zero in all measurement since lysimeters used in the experiment consisted of metallic structure, so no water was infiltrated out of the tank. ΔS is the change in soil water stored in the control volume of 1 m3.
The crop coefficient Kc, which is the ratio of the evapotranspiration (ETc) to the reference crop evapotranspiration (ETo), was calculated from the estimated ETc of the three lysimeters and the ETo determined by the Penman-Monteith equation (38). Basil root depth used in calculations was 10 cm at the growth and development stage (0–15 days after transplantation, dat), 30 cm at maturity stage (16–35 dat) and 20cm at senescence stage (36–57 dat).
2.4 Soil morphological and physicochemical characterization
Soil morphological characterization was conducted across the three municipalities to assess geomorphology, climatic conditions, drainage patterns, diagnostic horizons, and taxonomic classification according to the USDA Soil Taxonomy framework. Soil profiles were described following standardized protocols (42), with trial pits (1m × 1m × 1m) excavated to evaluate horizon thickness, boundary distinctness, color (Munsell notation), texture via manual field assessment, structure (type, class, and grade), macroorganism activity, and reactivity to NaF (pH-dependent fluorescence), H2O2 (effervescence for organic matter), and HCl (carbonate detection).
Physical properties analysis was performed on soil samples collected from three commercial basil (Ocimum basilicum L.) production farms per locality (Supplementary Figure S1). Within each farm, composed soil samples were extracted from a representative production plot at two depth intervals (0–5 cm and 5–10 cm) using a randomized stratified design. Bulk density (ρa, g cm-3), real density (Rd, g cm-3), FC, PWP, available water (AW), Tp (%), and soil texture were determined in this study.
Soil chemical characterization involved sampling at three farms per location, with paired samples collected from active production plots and adjacent undisturbed relicts to assess anthropogenic impacts. Samples were air-dried, sieved (<2 mm), and analyzed at an ISO/IEC 17025:2017-accredited laboratory (Agrosavia) for standard parameters, including pH (1:2.5 H2O), electrical conductivity (EC), organic carbon (Walkley-Black), CEC (ammonium acetate), and macro/micronutrient concentrations (e.g., N, P, K, Ca, Mg, Fe, Zn) via inductively coupled plasma optical emission spectrometry (ICP-OES).
2.5 Statistical analysis
Statistical descriptive analysis of soil properties included calculations of central tendency (mean), extremes (minimum, maximum), dispersion measures (SD, CV), and skewness were calculated. Both Spearman correlation matrices and principal component analyses were performed separately for physical and chemical properties to explore relationships among variables. General linear and mixed models were used to compare physicochemical properties. Location and depth corresponded to the fixed factors, while repetition was used as a random factor. The selection of the best model was carried out through the lowest values of the Akaike information coefficient (AIC) and the Bayesian information criterion (BIC). As a comparison test, Fischer’s LSD at 5% significance was performed. Analysis was performed in the packages Car, Stats, FactoMineR and factoextra of R software v. 4.3.1. (43).
3 Results
3.1 Quantification of basil crop water requirements
3.1.1 Climate characterization
The historical distribution of rainfall in the three locations presented a bimodal behavior (Figure 1). During El Niño phases (dark yellow bars) in El Espinal locality (Figure 1a), significant negative precipitation anomalies occur, particularly in January-March (-23% to -25%) and June-August (with August showing the most extreme deficit at -48%). In contrast, La Niña phases (blue bars) demonstrate precipitation surpluses, notably in June (39%), July (56%), and August (38%). During El Niño phases for locality Honda (Figure 1b), marked rainfall deficiencies are evident, with the most pronounced reductions occurring in July (-64%), followed by substantial decreases in June (-34%) and December (-32%). This location exhibits just one significant positive deviation during El Niño conditions, which appears in January (+24%). Conversely, La Niña phases (represented by blue bars) typically bring abundant precipitation, with the most substantial excesses recorded in June (+45%), August (+44%), and December (+37%). Throughout La Niña phases, numerous months display positive anomalies that surpass 19% above normal values.
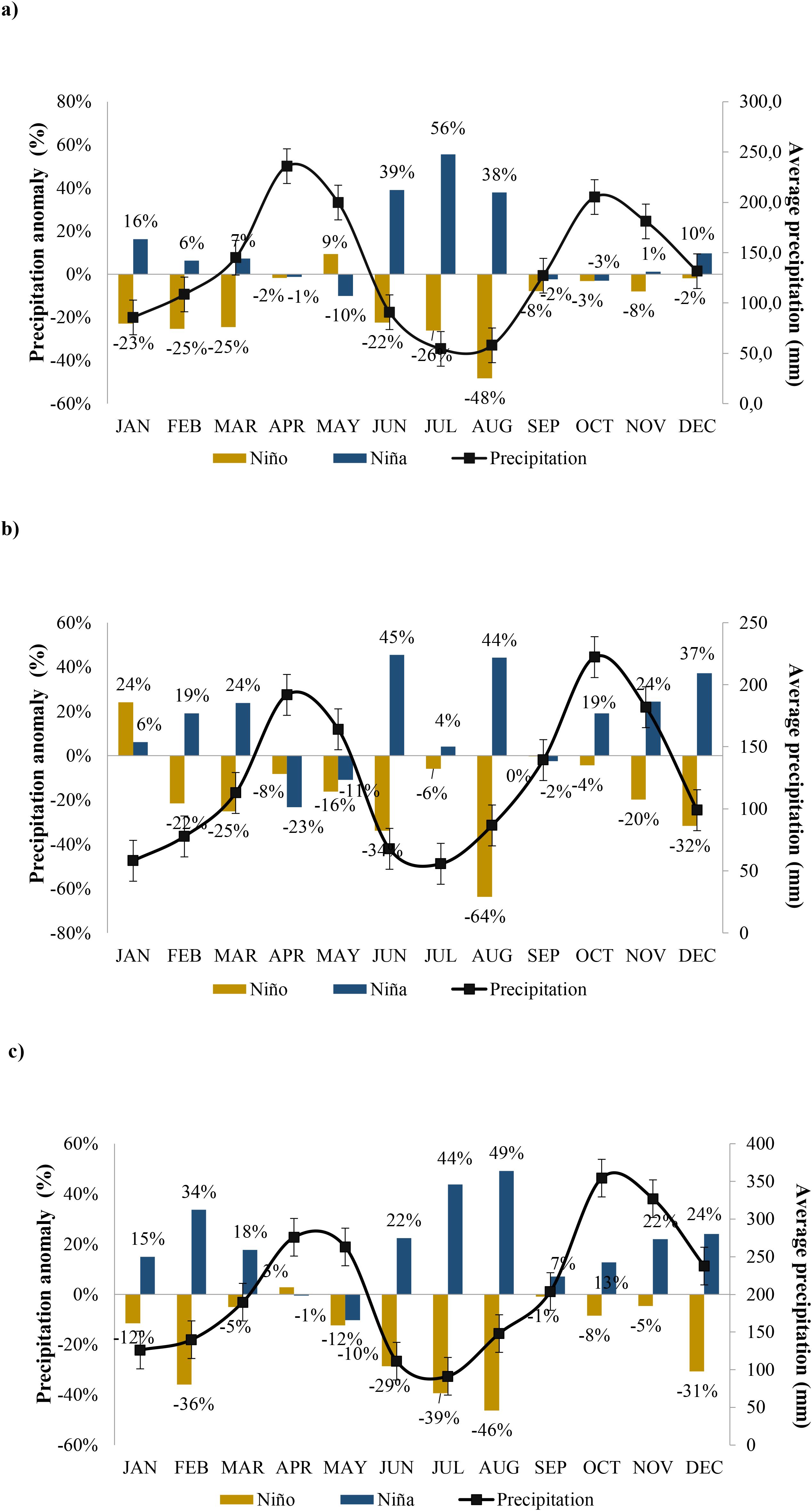
Figure 1. Relationship between El Niño/La Niña events and precipitation anomalies throughout the thirty years for (a) El Espinal, (b) Honda and, (c) Marquita.The bars represent precipitation anomalies during El Niño (dark yellow) and La Niña (blue) conditions, with percentages indicating deviations from normal precipitation patterns. The black line with square markers tracks average precipitation (mm), with error bars showing variability.
For locality Mariquita (Figure 1c), El Niño conditions consistently produce negative precipitation anomalies throughout the year, with the most severe reductions occurring in August (-46%), July (-39%), and February (-36%). Additional substantial rainfall shortfalls are observed in December (-31%) and June (-29%), creating extended dry periods. In contrast, La Niña phases (blue bars) generate significant precipitation enhancements across most months, with the most remarkable increases in August (+49%), July (+44%), and February (+34%). Several other months exhibit positive anomalies exceeding 15% during La Niña phases.
3.1.2 Daily ETc and crop coefficient
The daily crop ETc varied throughout the study period, influenced by meteorological conditions and crop development stages. The ETc values obtained from the three lysimeters showed consistent trends, with the highest water consumption recorded during peak maturity stage (average of 7.41 mm day-1). Figure 2 shows the daily ETc during the entire experimental period. The three series vary with time in a qualitatively similar manner with a few exceptions. Daily ETc showed an ample range of variation, with ETc values ranging from 1.03–7.81 mm day-1, and from 0.89–7.03 mm day-1, and from 0.43–7.40 mm day-1 for Lys1, Lys 2 and Lys 3, respectively. The maximum evapotranspiration rate, 45% of the total ETc, of basil occurred during the maturity stage, while 21% and 34% of ETc occurred in growth and senescence stages, respectively. The total ETc for the growing season was 228.82 mm. Figure 3 represents average daily ETc (mm day−1) for the different growth stages. The results indicate that the growth and development stage has the lowest daily ETc compared to other growths stages. However, the highest daily ETc was found during the maturity stage. The daily ETc is affected by the cropping period, duration, weather condition, and crop characteristics. Using the equation presented by Allen et al. (44) calculated Kc values were 0.75, 0.98 and 0.76 for basil crops at growth and development, maturity and senescence stage, respectively. The Kc values exhibited a dynamic pattern, increasing progressively from the initial establishment phase to the mid-season stage, where maximum water demand was observed. Subsequently, a decline in Kc was noted as the crop reached maturity. These findings align with expected physiological water requirements of basil, where higher transpiration rates correspond to biomass accumulation and leaf expansion.

Figure 2. Daily ETc (mm) of basil during the whole growing season for the three lysimeters.
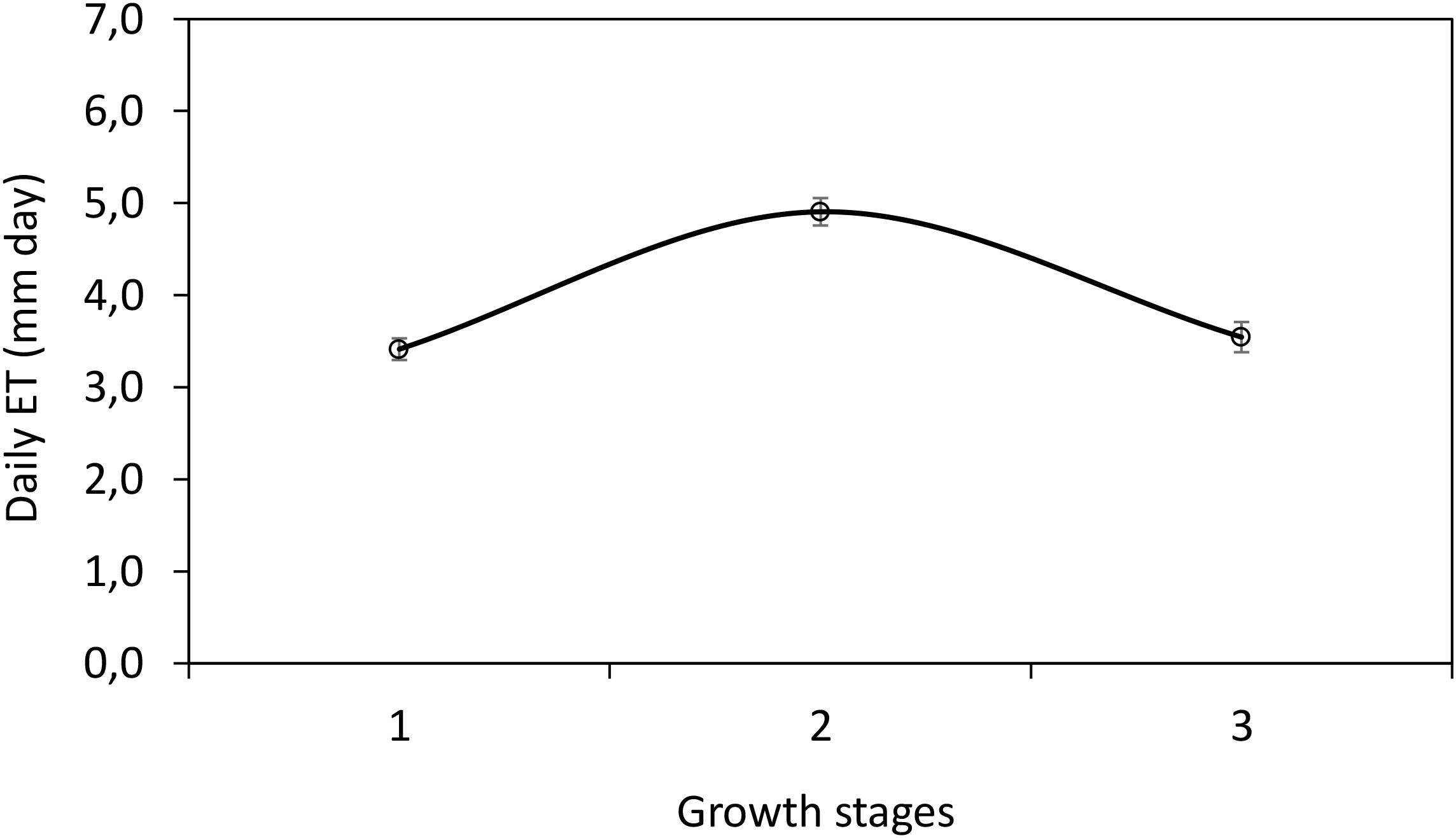
Figure 3. Average daily ETc of basil crop in different growth stages. Note: 1 represents the growth and development basil stage, 2 the maturity stage, and 3 the senescence stage. Hanging bars represent ± 95% confidence.
3.2 Soil physicochemical variability across Tolima sites
3.2.1 Soil Morphological characterization
The results of the soil morphological characterization are reported in supplementary tables (Supplementary Tables S1, S2). The soil pedological composition of the study areas of the three municipalities was highly similar (Supplementary Table S1). Results showed a presence of typical geomorphological features of alluvial plains, consisting of fluvio-volcanic sediment deposits that form terraces with particle sizes ranging from fine to coarse. The topography exhibits flat reliefs with slopes not exceeding 3%. The three study sites are classified as a tropical dry forest (bs-T), with a bimodal rainfall pattern and a warm dry climate. Taxonomically, the soils of the three municipalities share a common pedogenic origin as Typic Haplustepts (Supplementary Table S1) derived from fluvio-volcanic alluvium yet exhibit critical differences in horizonation that directly influence plant-available water and irrigation efficiency.
The edaphological characterization of the three locations revealed a similar pattern composed of altered mineral horizons (Ap) associated with shallow depths (<15 cm). This could be explained by soil loss due to anthropogenic activities (more details in Supplementary Tables S2).
3.2.2 Soil physical properties
Statistical analysis showed no significant differences between the factors of location (Mariquita, Honda, and El Espinal) and soil depth (0–5 and 5–10 cm) (p>0.05). However, significant differences were found between locations and physical properties (ρa, Rd, Tp, FC, PWP and AW) (p<0.05) (Supplementary Table S3). Results of soil physical properties per location were shown in Figure 4. El Espinal recorded the highest bulk density (1.65 ± 0.02 g cm-3), significantly surpassing Mariquita and Honda by 26.0% and 7.87%, respectively (Figure 4a). However, no significant differences were found in real density between El Espinal and Honda (p>0.05); while Mariquita showed a 6% reduction in real density compared to both locations (Figure 4b). Tp and FC were higher in Mariquita exceeding El Espinal and Honda by 30.5% and 17.5% (Figure 4c), and by 22.8% and 16.2% (Figure 4d), respectively. Finally, available water displayed this pattern, with Mariquita showing the highest value (3.59 ± 0.32%), followed by Honda (2.64 ± 0.2%) and El Espinal (2.41 ± 0.15%) (Figure 4f).
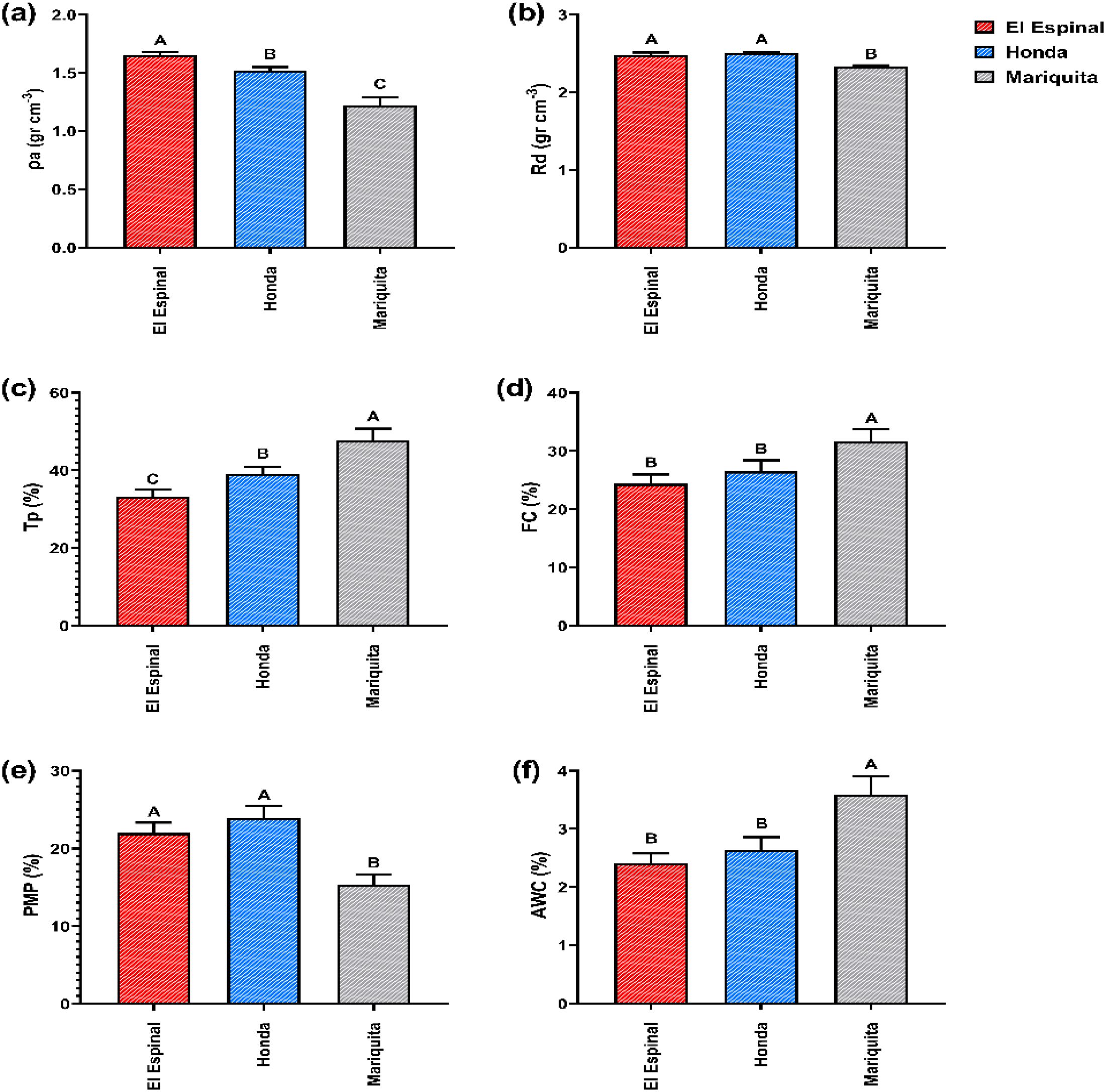
Figure 4. Soil physical properties per location: (a) Bulk density (ρa), (b) Real density (Rd), (c) Total porosity (Tp), (d) Field capacity (FC), (e) Permanent wilting point (PWP) and (f) Available water (AW) for El Espinal, Honda and Mariquita localities. Different letters indicate significant differences between treatments according to LSD test (p< 0.05) for mean ± standard error. The bars correspond to the experimental error for each treatment.
Regarding the sand, clay, and silt fractions, no significant interaction was observed between the factors of location and system (crop vs. relict) (p > 0.05) (Supplementary Table S4). However, significant differences (p < 0.05) were found for these variables across the location factor (Figure 5). The soil particle size fractions showed an inverse relationship across the localities, with higher sand content contrasting with lower silt and clay proportions. The sand fraction in Mariquita was the lowest with value of 55.90 ± 8.78%, differing significantly (p < 0.05) from the other two locations (Figure 5a). However, this location had the highest silt (25.00 ± 1.99%) and clay (19.10 ± 3.56%) fractions, with statistically significant differences (p < 0.05) compared to the other locations (Figures 5b, c).
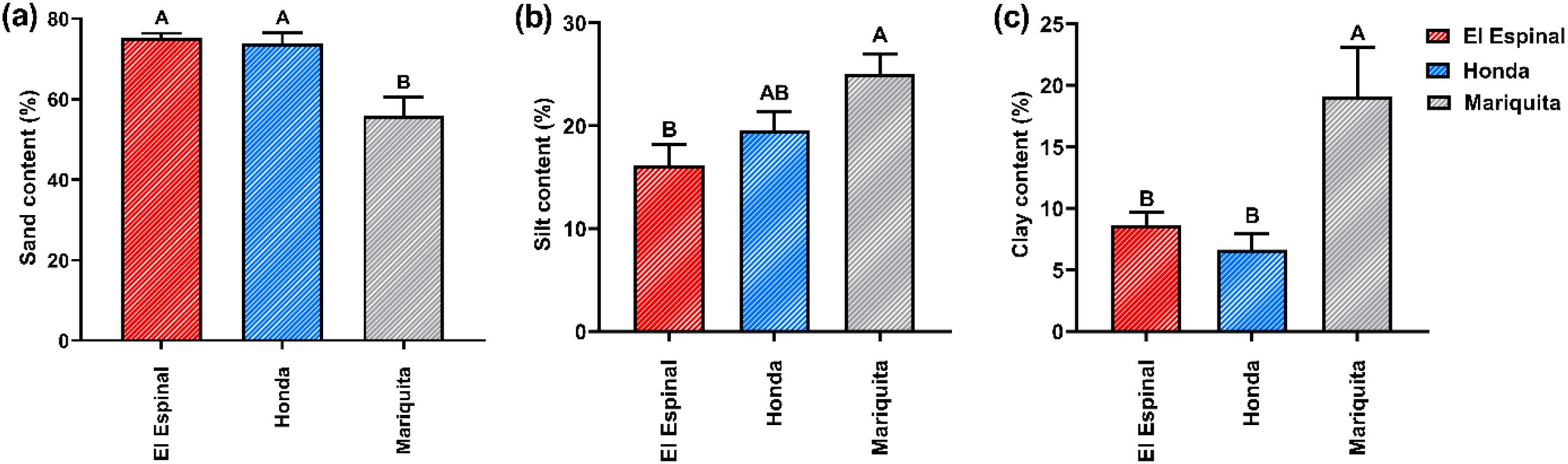
Figure 5. Sand, clay and silt fractions per interaction crop system and location factors. Different letters indicate significant differences between treatments according to LSD test (p< 0.05) for mean ± standard error. The bars correspond to the experimental error for each treatment.
3.2.3 Soil chemical properties
The statistical analysis of chemical properties revealed that pH, CE, OC, TN, SK, Na, S, B, Fe, and Zn had no significant effects (p > 0.05) at any factor level or interaction. However, ECEC, OM, K, Ca, Mg, and SMg showed significant differences (p < 0.05) at the location level. Significant differences across both factors (location and system) and their interaction (p < 0.05) were found in SMg and Mn. For the cultivation system factor, significant differences were identified in P, SCa, SNa, and Cu (Suplementary Table S5).
The LSD tests (p ≤ 0.05) for mean comparisons revealed significant differences in the factors location (Mariquita, Honda, and El Espinal) and system (crop vs. relict), as well as their interaction for the variable Mn content. The highest mean Mn levels were recorded in El Espinal, where the relict system exceeded Honda by 55% and Mariquita by 32%. The crop system showed 25% and 36% higher Mn levels compared to Honda and Mariquita, respectively (Figure 6a). For the OM variable, Mariquita exhibited the highest content, surpassing Honda by 35.5% and El Espinal by 55.3% (Figure 6b). A similar trend was observed for ECEC, with differences of 37.6% (Honda) and 50.5% (El Espinal) relative to Mariquita (Figure 6c). Finally, crop systems were 55% and 49.3% higher than relict systems, for P and Cu content, respectively (Figures 6d, e).
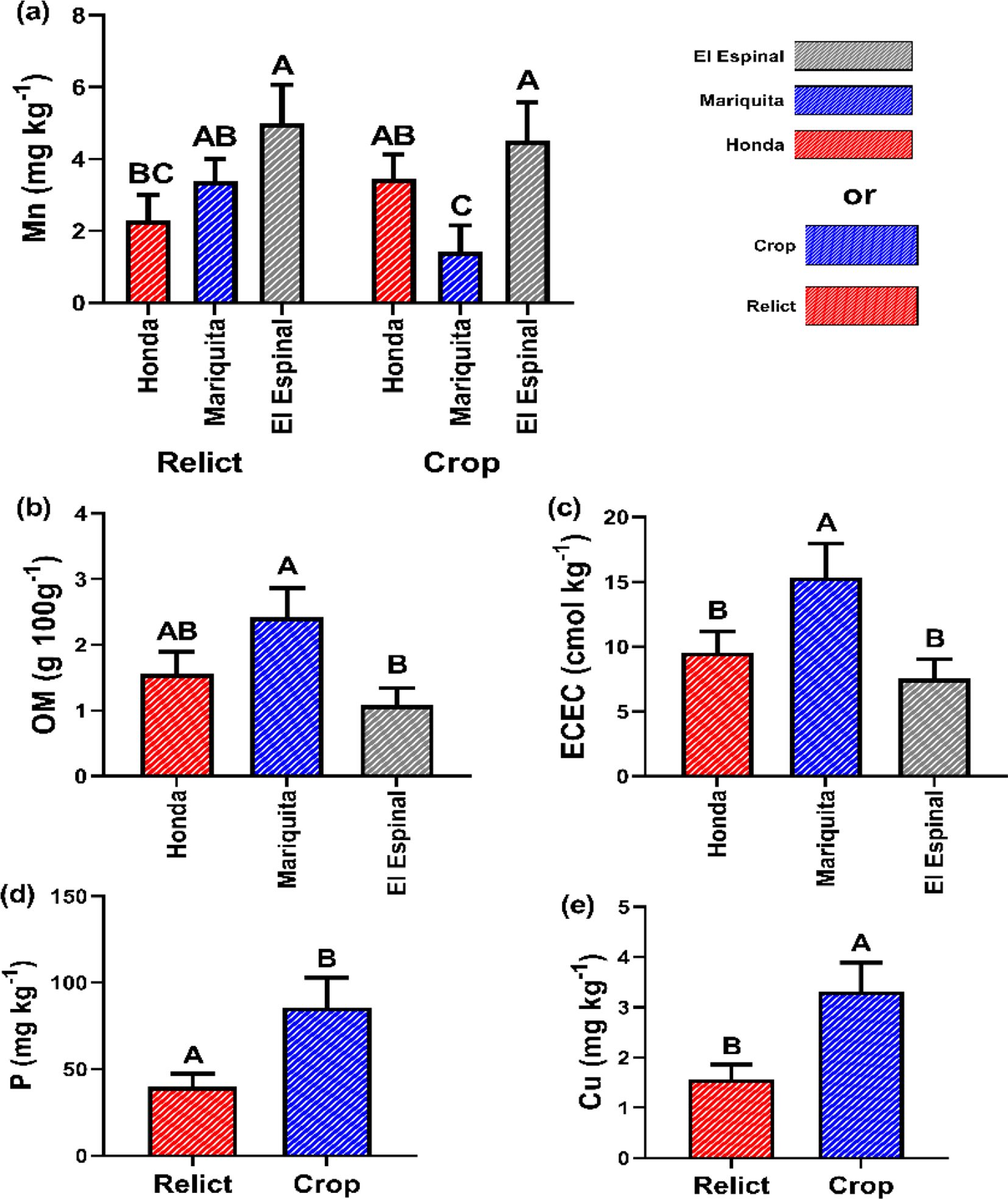
Figure 6. Chemical properties for crop and relict systems: (a) Manganese (Mn), (b) Organic material (OM), (c) Effective cation exchange capacity (ECEC), (d) Phosphorus (P) and (e) Copper (Cu). Different letters indicate significant differences between treatments according to LSD test (p< 0.05) for mean ± standard error. The bars correspond to the experimental error for each treatment.
For chemical properties related to base saturation, significant differences in K (potassium) were observed across locations. Honda and Mariquita showed higher K contents with values of 0.39 ± 0.09 and 0.36 ± 0.04 cmol kg-¹, respectively, which sharply contrasted with El Espinal (0.18 ± 0.03 cmol kg-¹) (Figure 7a). Na saturation (NaS) was significantly higher in crop system with a value of 61% compared to relict systems (Figure 7b). Calcium (Ca) concentrations were higher in Mariquita, surpassing Honda and El Espinal by 33.8% and 49%, respectively (Figure 7c). However, Ca saturation (CaS) showed no spatial variation but differed between systems, with relict systems representing a 9.47% increase over crop systems (Figure 7d). Mg content was significantly elevated in Mariquita, exceeding Honda and El Espinal by 53.3% and 55.8%, respectively (Figure 7e). Crop systems in Honda and El Espinal showed a 35.8% higher MgS compared to relict systems, while Mariquita had no significant system-level differences (Figure 7e).
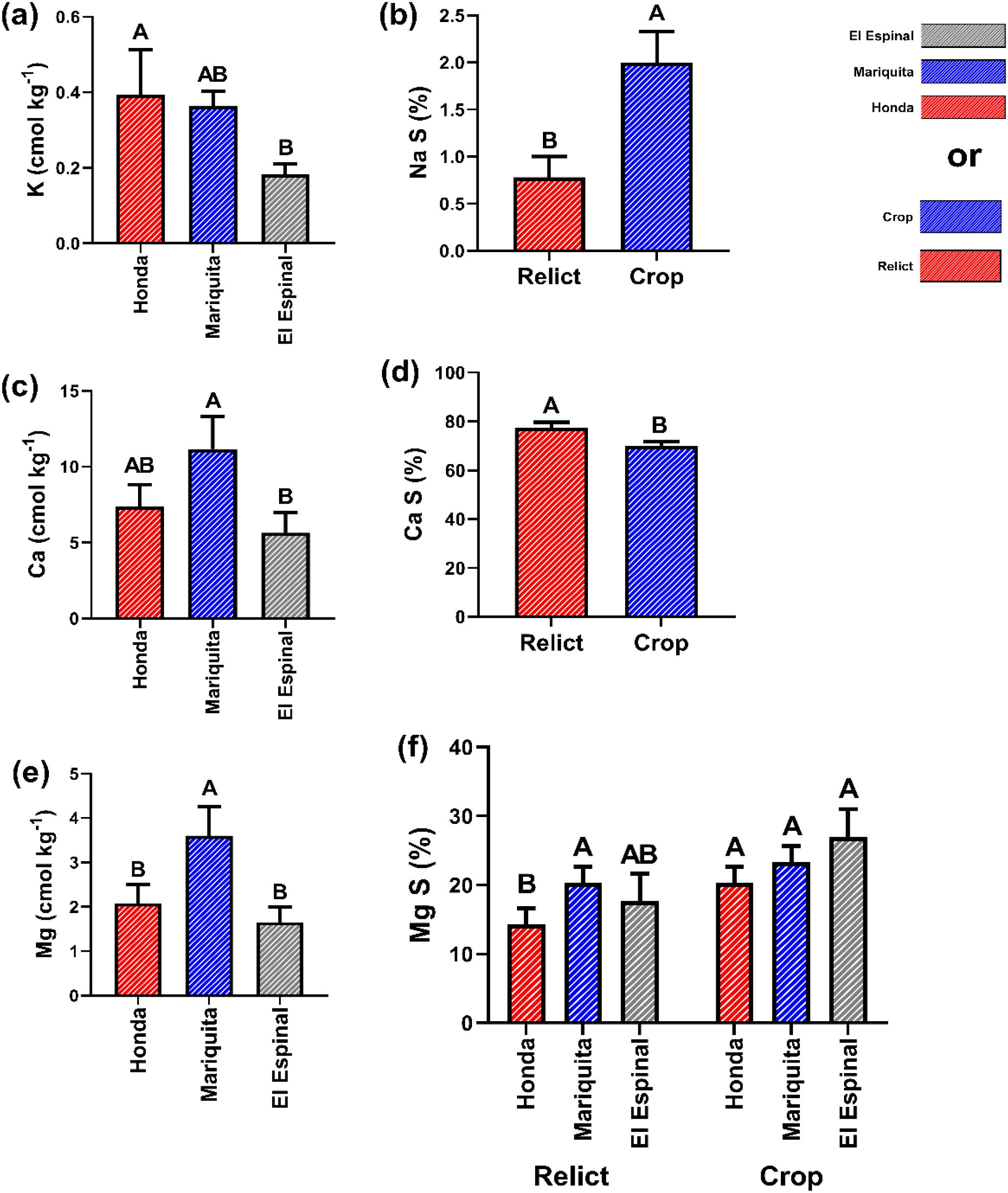
Figure 7. Chemical properties and saturation percentages: (a) Potassium K, (b) Sodium saturation (Na S), (c) Calcium, (d) Calcium saturation (Ca S), (e) Magnesium (Mg) and (f) Magnesium saturation (Mg S). Different letters indicate significant differences between treatments according to LSD test (p< 0.05) for mean ± standard error. The bars correspond to the experimental error for each treatment.
3.2.4 Correlation analysis and multivariate visualization
Figure 8 shows the correlation matrices between soil physico-chemical properties. A strong negative correlation was observed between ρa and TP (r = -0.906, p < 0.01) (Figure 8a), demonstrating an inverse relationship where increased soil bulk density corresponds to reduced total pore space due to soil compaction. Additionally, moderate positive correlations were found between ρa and PWP (rs = 0.432, p < 0.05), as well as ρa density and RD (rs = 0.397, p < 0.05). The positive relationship between ρa and RD reflects the natural influence of soil mineral particle density on overall compaction, although this relationship is moderated by factors such as structure and organic matter content. Figure 8b presents several significant associations between soil chemical and textural properties. Strong negative correlations were observed between sand and clay content (rs = -0.840, p < 0.01), as well as between sand and silt (rs = -0.827, p < 0.01), which is consistent with the complementary distribution of textural fractions. OM showed moderate positive correlations with clay (rs = 0.512, p < 0.01) and ECEC (rs = 0.559, p < 0.01), suggesting the important role of these properties in nutrient retention.
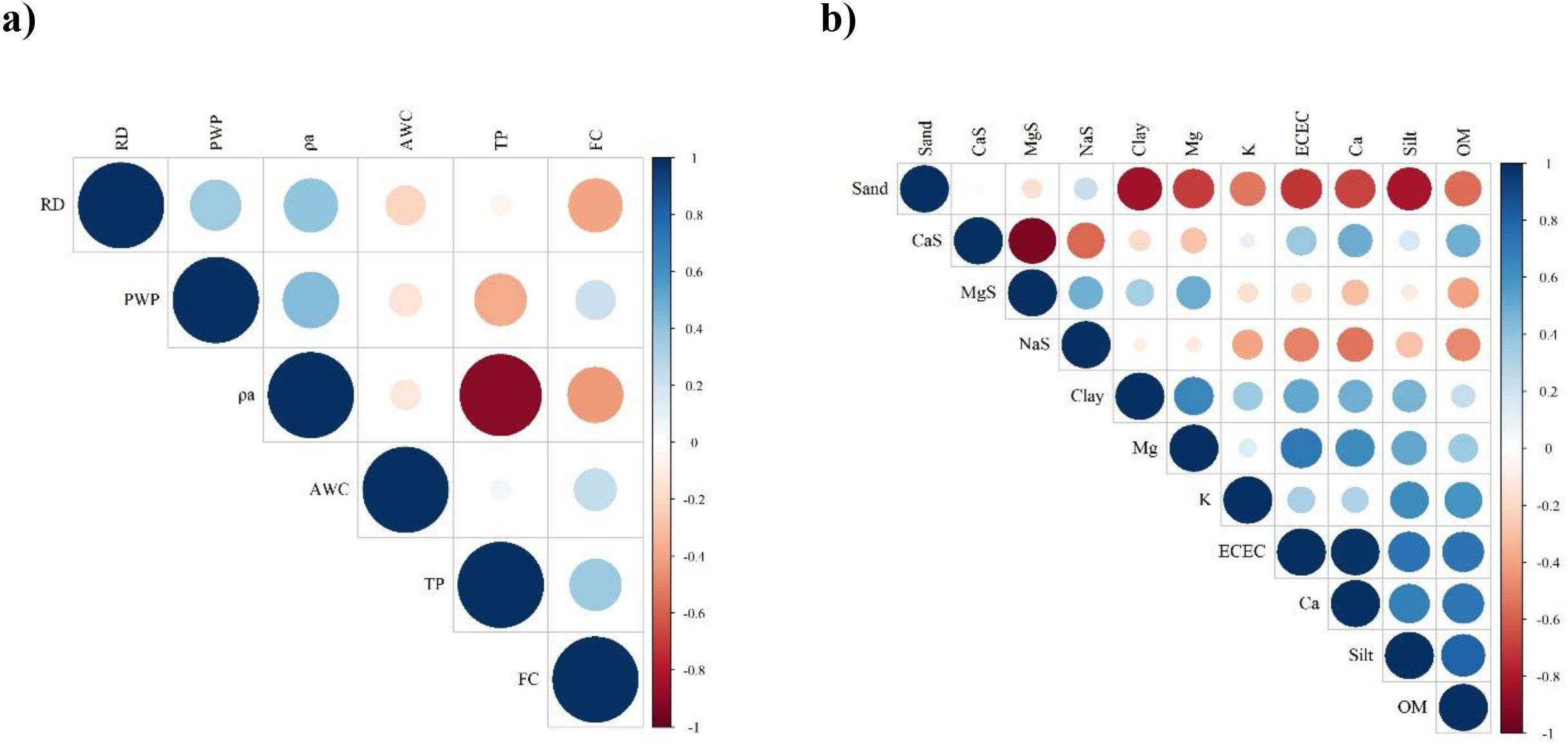
Figure 8. Correlation matrices between the soil properties evaluated. (a) Physical properties, (b) Chemical properties.
The PCA biplot analysis revealed distinct patterns in both soil physical and chemical variables. For physical properties (Figure 9a), the first principal component (Dim1) explains 48.1% of the total variability, while the second component (Dim2) contributes 20%, with both accumulating 68.1% of the variance. The Espinal locality showed a strong association with ρa and moderate correlation with PWP and RD, being predominantly located in the right quadrant of the graph. In contrast, Mariquita was mainly characterized by high HR, PT and AWC values, being positioned in the left region of the biplot. Honda presented intermediate behavior, although with greater affinity towards the variables in the right sector.
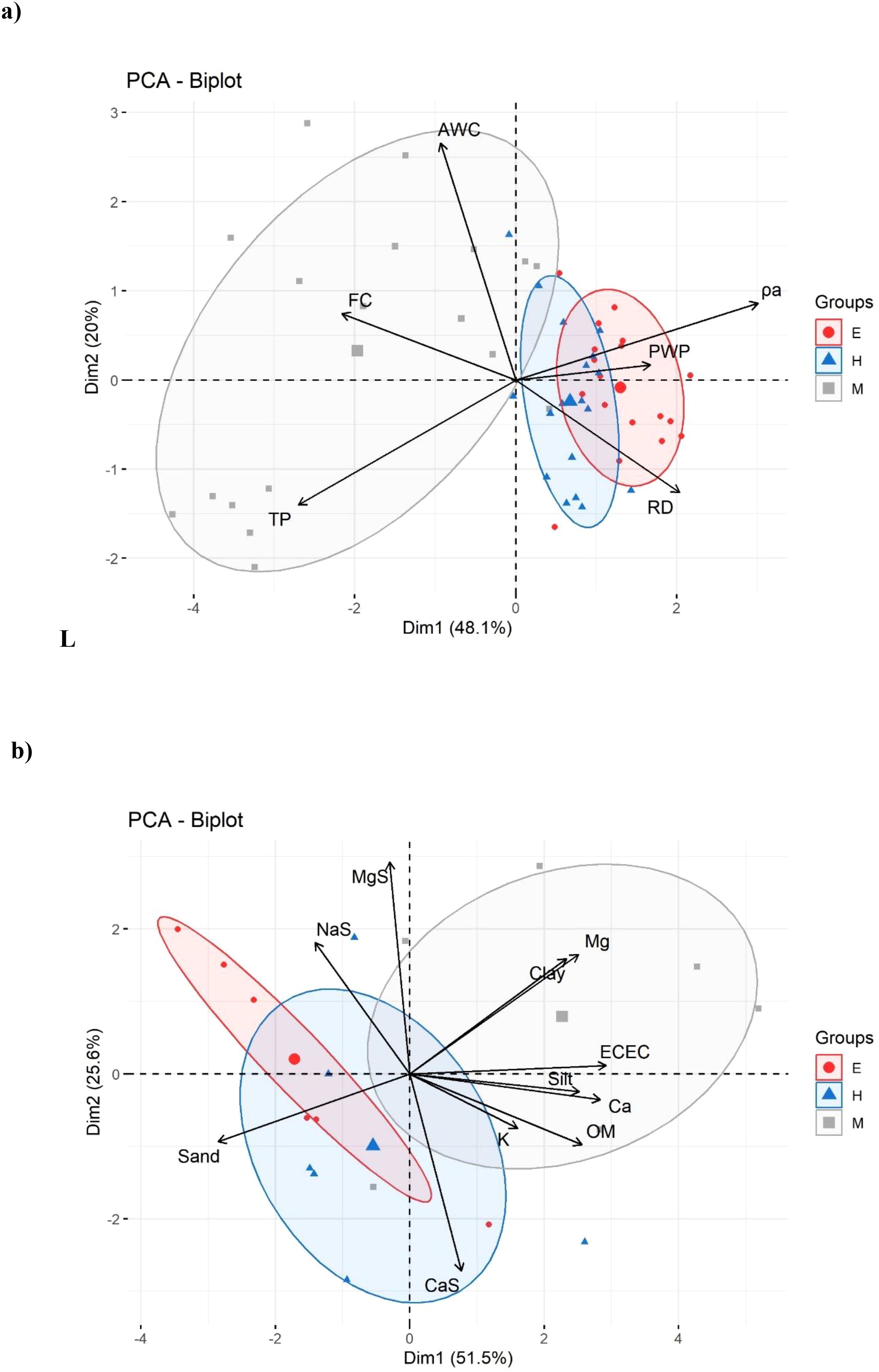
Figure 9. Principal component analysis (PCA) biplots of soil properties in Espinal, Mariquita and Honda locations: (a) physical properties and (b) chemical and textural properties.
In chemical variables (Figure 9b), the first principal component (Dim1) explains 51.5% of the total variability, while the second component (Dim2) contributes 25.6%, accumulating 77.1% of the variance. The locality Mariquita is strongly associated with the variables Clay, Mg, ECEC, Silt, Ca and OM, being predominantly located in the right sector of the biplot. In contrast, locality Espinal shows a vertical distribution along the Dim2 axis, influenced mainly by the Sand variable in the left region and with some samples affected by NaS. Locality Honda presented a more dispersed distribution, mainly in the lower left quadrant, showing affinity with the Sand and CaS variables.
4 Discussion
4.1 Climate and crop water requirements
4.1.1 Bimodal rainfall and ENSO impacts
The climatic variability observed in the Tolima region played a fundamental role in dictating the water requirements of basil. Our long-term climate characterization, based on more than 30 years of temperature and precipitation records, revealed a bimodal rainfall pattern with marked anomalies during El Niño and La Niña events. During El Niño phases, significant precipitation deficits were recorded, with reductions of up to 48% in certain months, while La Niña conditions produced substantial surpluses. Such fluctuations affect the soil moisture regime and, consequently, the crop water balance. Chaali et al. (38) highlighted that the bimodal behavior of the historical distribution of rainfall is related to the presence of the Intertropical Confluence Zone (ITCZ), which affects the annual distribution of precipitation. The latter influence the spatial distributions of precipitation, inducing a seasonality in the meteorological phenomena and regional climate. El Niño is generally associated with below average rainfall, higher temperatures and vapor-pressure deficit, which increase crop water demand and irrigation requirements and, unless compensated by targeted irrigation, tend to lower water productivity and yields. By contrast, La Niña usually brings above-average rainfall and reduced evaporative demand but increases risks of waterlogging, percolation losses and disease that can also undermine effective water productivity (32, 45). Accordingly, irrigation should be ENSO-responsive: under El Niño prioritize water conserving measures and targeted-deficit scheduling informed by site measured ETc (lysimeter-derived Kc) and seasonal forecasts, while under La Niña prioritize reduced irrigation frequency, improved drainage and disease management.
4.1.2 ETc and Kc patterns
Daily evapotranspiration (ETc) measurements obtained via volumetric lysimeters showed that basil water consumption varied markedly with its developmental stages. Peak water consumption occurred during the maturity stage (average 7.41 mm day-1), accounting for approximately 45% of the total seasonal water use. The dynamic behavior of the crop coefficient (Kc), calculated as 0.75 for the growth stage, rising to 0.98 at maturity and decreasing to 0.76 during senescence, reflected the basil physiological adjustments to changing environmental conditions. When comparing our observed Kc values (0.75, 0.98, 0.76) with existing literature, some interesting patterns emerge. In a field study of basil in Valle del Cauca (Colombia), Daza-Torres et al. (35) reported much lower Kc values: 0.45 (± 0.02) in the initial stage and 0.59 (± 0.02) in the mid-maturity stage. Meanwhile, under semi-arid conditions in Iran, Ghamarnia et al. (46) derived single and dual Kc values for basil of approximately 0.71, 1.11, 1.39 (initial, development, mid stages) in one treatment group, and 0.57, 0.97, 1.26 in another (somewhat higher mid season). More extremely, Martins et al. (30) in Brazil measured Kc values in greenhouse lysimeter settings ranging from 1.5 to 2.8 (depending on days after transplant, leaf area and coverage). These discrepancies are readily explained by methodological and environmental controls, principally canopy cover, the relative contribution of soil evaporation, irrigation regime, crop water status, and atmospheric demand that alter the transpiration and evaporation partition and thus apparent Kc. Given that our experiment was maintained under full (non-stress) irrigation and produced a dense canopy while using a lysimeter design intended to minimize edge and advection artefacts, we interpret the reported Kc as empirically calibrated, site-specific values appropriate for irrigation scheduling under the local climatic regime. Given the increasing challenges posed by climate change in Tolima region, including rising temperatures, erratic precipitation, and more frequent extreme weather events (47), the observed variability in ETc and Kc emphasizes the critical need for adaptive irrigation management. Precise, stage specific irrigation scheduling can optimize water use efficiency, ensuring that basil receives adequate moisture during periods of peak demand while minimizing water waste. Such approaches are vital for maintaining both yield and phytochemical quality, as water stress not only reduces biomass accumulation but can also alter the synthesis of key bioactive compounds (48). It was reported that basil can be grown in a soilless water-based system. It had a higher yield in the soilless system compared to the conventional system (49). According to previous studies, Rakocy et al. (50) conducted a study on basil to compare the batch and staggered cropping system. It was reported that grown under aquaponics had more yield with 1.8 kg m2 than grown under field conditions with 0.6 kg m2. These studies revealed that basil could be produced in both hydroponic and aquaponic systems together with restricted water and without soil. These systems affect the morphological and yield properties of basil genotypes positively under different growing conditions (48).
4.2 Site-specific soil constraints and management interventions
4.2.1 Physical properties: variability and solutions
Soil physical properties are fundamental factors for water availability, nutrient retention, and root development, which directly impact basil productivity. Soil characteristics are very important especially when dealing with basil oil quality, since they may also affect oil characteristics. Our study revealed significant spatial variability in these properties across the three study locations, which has important implications for basil cultivation in the Tolima region. The edaphological characterization showed different textures in the study region ranging from sandy to sandy clay loam, which could affect plant growth in different ways. In fact, plant roots grow faster in sandy soil, but water and nutrient uptake can be limited due to insufficient contact from the soil. On the other hand, the opposite situation occurs in clay soils (51), and this may lead to changes in the yield and essential oil components of plants (52). Essential oil compositions in basil plants vary based on various factors, such as, soil type, climatic conditions, growth period, harvest time, and post-harvest practices. Also, essential oil may differ in oil yield and quality according to soil type and this provides an opportunity to find out suitable soil for getting the highest essential oil yield and the best of its quality (53). Tursun (54) found that the highest essential oil yield was obtained in the basil plants grown in sandy soils while the least was obtained from clayey soils.
Bulk density was a critical parameter that reflects soil compaction and porosity. In fact, El Espinal showed the highest bulk density (1.65 ± 0.02 g cm-3), which suggests a higher degree of compaction compared to Honda and Mariquita. High bulk density is typically associated with reduced pore space, leading to diminished water infiltration and lower aeration. Such conditions can restrict basil root growth and impair the soil holding capacity to store and transmit water, which is particularly problematic during periods of water scarcity (55). Compacted soils may also exacerbate water stress during critical growth stages by limiting the movement of water into the basil root zone.
Soils in Mariquita demonstrated superior Tp and FC, with available water (AW) content reaching 3.59 ± 0.32%. High porosity enhances both water retention and air exchange, which are essential for robust basil root development and nutrient uptake (56). FC indicates the soil ability to retain water after drainage, ensuring that basil has access to moisture during dry periods. The higher available water in Mariquita soil suggested that these fields are better suited to withstand intermittent drought conditions, a crucial advantage in the context of Tolima bimodal rainfall pattern and high evapotranspiration rates. The inverse relationship between bulk density and Tp as observed in Espinal affects the root growth and also limits water retention capacity compared to Mariquita farms. The observed differences in soil physical properties among the three locations highlighted the need for location specific management practices. In areas like El Espinal, where high bulk density and low available water prevail, agronomic interventions such as the incorporation of organic amendments, reduced tillage, or the establishment of cover crops could help improve soil structure, increase porosity, and ultimately enhance water infiltration (36). These practices can reduce the negative impacts of compaction and support more effective root penetration, thereby improving water and nutrient uptake. Moreover, the physical properties of the soil act as one of the most determining factors in the essential oil composition of different aromatic plant species (57). Several researchers (53, 54, 57, 58) have focused on looking at the most suitable soil type to produce basil plants having essential oil with the best composition and concentration. Soils with inherently favorable physical properties, as observed in Mariquita, may require less intensive soil amelioration. However, even in such areas, precise irrigation scheduling is essential to align water application with the crop phenological stages. Basil exhibits significant variation in water demand during its lifecycle, low during the early growth and senescence stages, but peaking during the maturity phase when biomass accumulation is at its highest (35). Ensuring optimal soil moisture levels during these peak periods is critical not only for yield but also for maintaining the concentration of bioactive compounds that determine basil market value (59). The spatial heterogeneity observed in soil physical properties highlighted the limitations of a one size fit all irrigation strategy (60). Instead, precision agriculture approaches, such as the use of soil moisture sensors and remote sensing techniques, can facilitate real time monitoring of soil water content and compaction. This data can then be used to tailor irrigation and soil management practices to the specific conditions of each locality (17). Soils with high porosity (Mariquita) enhance drought resilience but increase nutrient leaching, while compacted soils (El Espinal) restrict root growth but potentially elevate essential oil concentration. This duality needs site specific management such as biochar amendments in El Espinal to alleviate compaction without sacrificing oil quality. Recent studies (61, 62) indicated that superabsorbent and biodegradable hydrogel soil conditioners can substantially increase soil water retention, reduce surface evaporation and percolation losses, and improve crop performance under water-limited conditions; such amendments therefore complement mulching and organic-matter additions as practical options to enhance water productivity for basil in El Espinal and other compacted soils. Further, by linking ENSO driven rainfall anomalies to real-time ETc, we enable irrigation protocols that reduce water use during El Niño, a step toward climate resilient precision agriculture. This integrated approach is novel in tropical aromatic crop systems and represents a disruptive advance in water use efficiency under ENSO driven climates.
4.2.2 Chemical properties: risks and optimization
Soil chemical properties fundamentally influence nutrient availability, root uptake, and overall crop performance, making them critical in understanding basil productivity in the Tolima region. Our analysis revealed substantial spatial variability in several chemical parameters, such as ECEC, OM, and key macronutrients (K, Ca, Mg, P) and micronutrients (Mn, Cu), across the study locations. This variability emphasizes the complex interplay between natural soil formation processes and anthropogenic influences, which are critical for optimizing basil growth and secondary metabolite synthesis.
The correlation analysis provided deep insights into the interrelationships among soil chemical properties. These relationships indicated that soils with higher clay and organic matter are inherently better at sustaining nutrient availability, a factor that is particularly beneficial for basil growth and metabolic functions (37). These soils were mainly characterized by higher clay, magnesium and ECEC contents (54, 63, 64). Thus, the soils of Mariquita presented potentially better nutrients and water retention capacity. For this locality, the higher clay content creates a structure with abundant micropores, crucial for water retention at higher tensions. This textural characteristic would allow the soil to maintain moisture available for basil plants during prolonged drought periods compared to sandy soils, which typically drain more rapidly (65).
Organic matter is a key driver of soil fertility, enhancing nutrient retention, water holding capacity, and soil structure. In our study, Mariquita soils showed the highest OM content, surpassing Honda by 35.5% and El Espinal by 55.3%. High OM is typically associated with increased microbial activity and enhanced formation of organo-mineral complexes that stabilize nutrients in the soil (Cotrufo et al., 2019). This relationship indicated that soils rich in OM are more efficient in retaining essential cations (Ca²+, Mg²+, and K+), thereby ensuring a more stable nutrient supply to basil plants. This latter is crucial for sustaining nutrient availability under the fluctuating moisture regimes characteristic of Tolima bimodal rainfall patterns.
Macronutrient balance is critical for plant growth, particularly in nutrient sensitive species such as basil. Our results revealed significant differences in P content between cultivation and relict systems, with cultivated systems exhibiting a 54.5% higher concentration. Both systems fell within the high to very high P range established for Colombian soils (66). This disparity is attributed to continuous fertigation using phosphoric acid (H3PO4 with 85% concentration) and pre-planting incorporation of phosphate rock as a soil amendment. Excessive P application through fertilizers has been shown to elevate soluble P fractions (67). Moreover, high P levels reduce Zn, Fe, and Cu uptake and translocation, inhibiting plant growth. For basil plant development, the optimal P level is ranging between 0.62 and 1.00% (68), serving as a structural component of cell membranes, DNA, RNA, and ATP (69). In medicinal plants, it further enhances essential oil synthesis and assimilation (70).
For K, Ca, and Mg, similar trends were observed across the study sites. K levels were significantly higher in Honda and Mariquita compared to El Espinal, ranging within the sufficient to high values established for Colombian soils (66). K is among the most abundant elements in basil leaves, with concentrations ranging from 1.55 to 2.05% (68). In fact, K plays a vital role in stomatal regulation and enzymatic activation, with deficiencies leading to impaired photosynthesis and reduced biomass accumulation (15). According to the Colombian soils classifications (66), the Ca content in Mariquita was very high, while in Honda and El Espinal ranged within high levels. Local agricultural practices include pre-planting dolomite (CaMg(CO3)2) amendments or calcium nitrate (Ca (NO3)2) fertigation to enhance phosphorus sorption via CaCO3 interactions (71). Normally, foliar Ca concentrations range from 1.25 to 2.00%, reflecting its structural role in the cell wall middle lamella, where it binds to pectin carboxyl groups to promote cellular elongation in shoots and roots (68). Additionally, Ca stabilizes the mitotic spindle during cell division, supports pollen tube germination, and improves postharvest quality in exported basil by reducing cold damage and decay rates (72). As regards Mg concentrations (normal range between 0.60 and 1.00%), we found in this study very high levels in Mariquita and sufficient levels in Honda and El Espinal. However, Mg deficiency manifests as interveinal chlorosis in older leaves due to its role as a constituent of chlorophyll’s tetrapyrrolic ring. Mg also acts as a cofactor for kinases and activates ribulose-1,5-bisphosphate (RuBP) carboxylase during photosynthetic light reactions (73).
Soil pH of the three localities showed a slightly acidic to neutral pH range, thus facilitating CEC dominated by Ca²+, Mg²+, K+, and Na+ (74). Calcium saturation (CaS) was higher in was higher in undisturbed (relict) areas compared to cultivated plots. Nonetheless, CaS values remained within the established reference range of 65 to 75%, while magnesium saturation (MgS) levels aligned with the proposed 10 to 15% threshold. At the cultivated system level, MgS exceeded the upper reference limit, a consequence of periodic dolomite amendments (applied every 4 to 5 months) and magnesium sulfate (MgSO4) fertigation. Although adjustments to the soil CEC complex did not translate into statistically significant yield gains, it is recommended to avoid extreme imbalances in base cation proportions to maintain soil health. Finally, sodium saturation values in both relict (0.78%) and cultivated (2%) systems remained within normal limits (0 to 5% of total CEC), indicating no sodicity concerns under current management practices (75).
Micronutrients, despite being required in lower amounts, play crucial roles in enzyme function and secondary metabolite biosynthesis (76, 77). In our study, we observed that Mn levels in soil were significantly higher in El Espinal, however, these concentrations remain below the lower critical threshold (< 5 mg kg-1) for agricultural soils. Mn deficiency is common in alkaline soils due to redox reactions and other factors such as increased OM content, microbial activity, soil temperature, and water content. Although plants absorb only a small fraction of Mn for growth and developmental processes (78), basil requires foliar Mn concentrations between 30 and 150 ppm (68). Mn is critical for redox processes in plants and acts as an activator or cofactor for numerous enzymes, including proteins essential for light-mediated water oxidation in Photosystem II (PSII) (76, 79).
Regarding Cu, cultivated soils showed elevated concentrations, primarily attributable to the use of Cu based fungicides, which accumulate in plant tissues and subsequently leach into the soil (80). Nonetheless, the measured soil Cu levels (1–3 mg kg-1) fall within the adequate range for the evaluated systems. In mineral soils, Cu concentrations should exceed 6 mg kg-1, owing to its low soluble fraction (1 × 10–8 to 60 × 10–8 mol L-1) and the fact that approximately 98% of Cu is bound to organic matter (68). In basil foliage, sufficient Cu ranges are reported between 5 and 10 mg kg-1. Like Mn, Cu is integral to plant redox systems and serves as an enzymatic activator for superoxide‐detoxifying enzymes; it also participates as a redox cofactor in PSII‐mediated electron transport and in lignin biosynthesis (81). This highlighted the need for careful monitoring and management of micronutrient levels to maintain optimal basil quality. While this study quantifies key edaphoclimatic parameters of basil productivity, the absence of direct yield measurements (e.g., biomass, essential oil yield) limits causal inference. Future work should integrate field trials with controlled irrigation and fertilization treatments to quantify basil fresh or dry weight and essential oil yield under contrasting soil water regimes. Subsequent research will couple lysimeter based soil water monitoring with harvest assessments and phytochemical profiling of essential oils to link edaphic factors directly to basil yield and aroma quality.
5 Conclusions
This study provides an empirically calibrated framework for climate-resilient basil cultivation in Tolima by establishing lysimeter derived, stage specific crop coefficients (Kc: 0.75, 0.98, 0.76) to operationalize precision irrigation. To enhance water productivity and yield stability under ENSO-modulated conditions we recommend ENSO responsive irrigation and soil management: during El Niño, implement deficit irrigation and mulching to counteract drought stress; during La Niña, reduce applications and improve drainage to prevent waterlogging. These water management practices should be coupled with site specific interventions such as organic amendments for compacted soils in El Espinal and balanced fertilization to address nutrient imbalances to collectively stabilize yield and essential oil quality. We further recommend that extension services couple seasonal ENSO forecasts with lysimeter-benchmarked Kc values to deliver actionable, site specific advisories that stabilize production in this tropical dry ecosystem.
Data availability statement
The original contributions presented in the study are included in the article/Supplementary Material. Further inquiries can be directed to the corresponding author.
Author contributions
JB-M: Conceptualization, Data curation, Formal Analysis, Funding acquisition, Investigation, Methodology, Project administration, Resources, Software, Supervision, Validation, Visualization, Writing – original draft, Writing – review & editing. SO: Conceptualization, Data curation, Formal Analysis, Investigation, Methodology, Resources, Software, Supervision, Validation, Visualization, Writing – original draft, Writing – review & editing. NC: Conceptualization, Data curation, Formal Analysis, Investigation, Methodology, Resources, Software, Supervision, Validation, Visualization, Writing – original draft, Writing – review & editing. CJ: Data curation, Formal Analysis, Investigation, Methodology, Writing – original draft. KP: Formal Analysis, Investigation, Writing – original draft. JH: Formal Analysis, Investigation, Writing – original draft. JC: Formal Analysis, Investigation, Writing – original draft.
Funding
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. This work was financially supported by the Colombian Ministry of Agriculture and Rural Development (MADR), the General System of Royalties (SGR), and the Colombian Corporation for Agricultural Research (AGROSAVIA) through the project entitled “Implementación de alternativas de manejo agronómico, postcosecha y de inocuidad en el sistema productivo de Albahaca (Ocimum basilicum L.) para el cierre de brechas en los municipios de Honda, Mariquita y Espinal en el departamento del Tolima” with BPIN code 2021000100147.
Acknowledgments
The authors would like to thank the Colombian Ministry of Agriculture and Rural Development (MADR), the Sistema General de Regalías (SGR), The Tolima government, and the Corporación Colombiana de Investigación Agropecuaria AGROSAVIA for supporting this research study and funding the Open Access publication through the project entitled “Implementación de alternativas de manejo agronómico, postcosecha y de inocuidad en el sistema productivo de Albahaca (Ocimum basilicum L.) para el cierre de brechas en los municipios de Honda, Mariquita y Espinal en el departamento del Tolima” with BPIN code 2021000100147. We also would like to thank the basil Colombian farmers of Tolima region who were involved directly and indirectly in this research study.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fsoil.2025.1621669/full#supplementary-material
References
1. Martínez DHF, Solano LJC, Cárdenas JÁ, Montes NL, Criollo J, and Medina JIB. La ruta de la albahaca (Ocimum basilicum): Aproximación a una estrategia de diversificación de usos para los excedentes de producción. Bogotá, Colombia: En Editorial AGROSAVIA (2024). doi: 10.21930/agrosavia.analisis.7407761
2. Kaur P, Verma K, Singh AK, Kumar R, and Singh S. Co-cultivation of sweet basil (Ocimum basilicum L.) with food crops for efficient resource utilization and increasing the farm profits. Ind Crops Products. (2024) 214:118495. doi: 10.1016/j.indcrop.2024.118495
3. Acevedo JA. Estrategias de competitividad para los productores de plantas medicinales en Colombia. Agexport Guatemala: tesis de pregrado publicada, Universitaria Agustiniana (2019). Available online at: https://backend.uniagustiniana.edu.co/server/api/core/bitstreams/cb43cfe4-5801-4d2c-a40e-0b31d1dd9822/contenthttps://www.export.com.gt/guias-de-cultivo.
4. Santos. JJ. Análisis de factores estratégicos para el proceso de exportación de hierbas aromáticas a los estados unidos. Colombia: [tesis de maestría publicada, Universitaria Nacional Abierta y a Distancia - UNAD (2020). Available online at: https://repository.unad.edu.co/handle/10596/34006. (Accessed January 15, 2025).
5. Esponda-Bernal M del M, Echeverri-Sánchez AF, Aguirre-González EF, and Andrade RS. A biophysical suitability model to identify best areas for the cultivation of potential cash crops: The case of basil in Valle del Cauca. Agric Syst. (2024) 216:103909. doi: 10.1016/j.agsy.2024.103909
6. Corpoica. Corporación Colombiana de Investigación Agropecuaria. In: Sistema de apoyo para la toma de decisión agroclimáticamente inteligente [SE-MAPA] V2.1. En (2016). Bogotá, Colombia: Corporación Colombiana de Investigación Agropecuaria. Available online at: https://www.agrosavia.co/productos-y-servicios/oferta-tecnol%C3%B3gica/l%C3%ADnea-agr%C3%ADcola/cultivos-transitorios-y-agroindustriales/sistemas-de-informaci%C3%B3n/723-sistema-experto-mapa (Accessed May 15, 2025).
7. Farahbakhsh M, Sarai Tabrizi M, and Babazadeh H. Determining basil production functions under simultaneous water, salinity, and nitrogen stresses. Appl Water Sci. (2023) 13:68. doi: 10.1007/s13201-022-01849-3
8. Bihter ÇE, Bintuð Ö, Özgür Ç, and Dilek A. Sweet basil (Ocimum basilicum L.) and potassium fertilization. J Plant Nutr. (2016) 39:35–44.
9. Yaldiz G, Gul F, and Kulak M. Herb yield and chemical composition of basil (Ocimum basilicum L.) essential oil in relation to the different harvest period and cultivation conditions. Afr J Traditional Complementary Altern Medicines. (2015) 12:71–6. doi: 10.4314/ajtcam.v12i6.7
10. Ekren S, Sonmez C, Özçakal E, Kurttas_ K, Bayram E, and Gürgülü H. The effect of different irrigation water levels on yield and quality characteristics of purple basil (Ocimum basilicum L.). Agric Water Manage. (2012) 109:155–61. doi: 10.1016/j.agwat.2012.03.004
11. Radácsi P, Inotai K, Sarosi S, Czovek P, Bernath J, and Nemeth E. Effect of water supply on the physiological characteristic and production of basil (Ocimum basilicum L.). Eur J Hortic Sci. (2010) 75:193–7.
12. Makri O and Kintzios S. Ocimum sp. (basil): Botany, cultivation, pharmaceutical properties, and biotechnology. J Herbs Spices Medicinal Plants. (2007) 13:123–50. doi: 10.1300/J044v13n03_10
13. Aharoni N, Kenigsbuch D, Chalupowicz D, Faura-Mlinski M, Aharon Z, Maurer D, et al. Reducing chilling injury and decay in stored sweet basil. Israel J Plant Sci. (2010) 58:167–81. doi: 10.1560/IJPS.58.3-4.167
14. Jordán MJ, Quílez M, Luna MC, Bekhradi F, Sotomayor JA, Sánchez-Gómez P, et al. Influence of water stress and storage time on preservation of the fresh volatile profile of three basil genotypes. Food Chem. (2017) 221:169–77. doi: 10.1016/j.foodchem.2016.10.059
15. Ronzón M, Hernández M, and Pérez C. Producción hidropónica y acuapónica de albahaca (Ocimum basilicum) y langostino malayo (Macrobrachium rosenbergii). Trop Subtropical Agroecosystems. (2012) 15:S63–71.
16. González-Gómez H, Juárez-López P, Quiñones-Aguilar EE, Rincón-Enríquez G, Alia-Tejacal I, Ramírez-Trujillo JA, et al. Growth of ‘Sweet Nufar’ basil (Ocimum basilicum L.) inoculated with arbuscular mycorrhizal fungi and Azospirillum brasilense. Rev. Chapingo Ser. Hortic. (2023) 29(1). doi: 10.5154/r.rchsh.2022.03.00
17. Evett SR, Schwartz RC, Casanova JJ, and Heng LK. Soil water sensing for wáter balance, ET and WUE. Agric Water Manage. (2012) 104:1–9. doi: 10.1016/j.agwat.2011.12.002
18. Yadvinder-Singh B-S, Ladha JK, Khind CS, Gupta RK, Meelu OP, and Pasuquin E. Long-term effects of organic inputs on yield and soil fertility in the rice–wheat rotation. Soil Sci Soc America J. (2004) 68:845–53. doi: 10.2136/sssaj2004.8450
19. Gavrić T, Čengić L, and Marković S. Effects of cultivars on the yield and contents of some bioactive components of basil. Works Faculty Agric Food Sciences Univ Sarajevo. (2022) 72:20–7.
20. IDEAM, Instituto de hidrología and Meteorología y Estudios Ambientales IDEAM. climatológico mensual (2021). Available online at: https://www.ideam.gov.co/web/tiempo-y-clima/climatologico-mensual/-/document_library_display/xYvlPc4uxk1Y/view/113579674 (Accessed August 30, 2025).
21. López-Hernández M, López Hernández MDP, Criollo-Núñez J, Beltrán JI, and Sandoval-Aldana AP. Chemical composition and antioxidant activity of ‘Nufar’ basil (Ocimum basilicum L.) essential oil from three municipalities of Tolima, Colombia. Food Chem Adv. (2024) 5:100819. doi: 10.1016/j.focha.2024.100819
22. Veritradecoro. La plataforma digital de comercio exterior (2021). Available online at: www.veritradecorp.com (Accessed August 30, 2025).
23. Piñeros Martínez N. Prospectiva en la producción y comercialización de la albahaca (Ocimum Basilicum l) de forma sostenible para el departamento del Tolima al 2032. Bogotá, Colombia: Master’s thesis, Maestría en Proyectos de Desarrollo Sostenible-Virtual (2022).
24. Vega J, Barco J, and Hidalgo C. Space-time analysis of the relationship between landslides occurrence, rainfall variability and ENSO in the Tropical Andean Mountain region in Colombia. Landslides. (2024) 21:1293–314. doi: 10.1007/s10346-024-02225-9
25. Urrea V, Ochoa A, and Mesa O. Seasonality of rainfall in Colombia. Water Resour Res. (2019) 55:4149–62. doi: 10.1029/2018WR023316
26. Naranjo Bedoya K, Aristizábal Giraldo EV, and Morales Rodelo JA. Influencia del ENSO en la variabilidad espacial y temporal de la ocurrencia de movimientos en masa desencadenados por lluvias en la región Andina Colombiana. Ingeniería y Ciencia. (2019) 15:11–42. doi: 10.17230/ingciencia.15.29.1
27. Spera ST, Baldoni AB, Magalhães CADS, Lulu J, Tonini H, Zolin CA, et al. Characterizing edaphoclimatic variables in sites hosting natural Brazil nut tree populations in Mato Grosso state. Nativa. (2020) 8. doi: 10.31413/nativa.v8i3.9438
28. Melo-León S, Otálora G, Buitrago D, Riveros L, and Giraldo ROS. Estimación de los efectos de la variabilidad climática en la producción agrícola en Colombia: Estimated effects of climate variability on agricultural production in Colombia. e-CUCBA. (2024) 21):133–41.
29. Ojeda-Silvera CM, Murillo-Amador B, Reynaldo-Escobar IM, Troyo-Diéguez E, Ruiz-Espinoza FH, and Nieto-Garibay A. Estrés hídrico en la germinación y crecimiento de plántulas de genotipos de albahaca Ocimum basilicum L. Rev mexicana Cienc agrícolas. (2013) 4:229–41.
30. Martins IP, Faria RTD, Palaretti LF, Santos MGD, and Fischer Filho JA. Evapotranspiration and crop coefficient of basil determined by weighing lysimeters. Horticultura Bras. (2019) 37:373–8. doi: 10.1590/S0102-053620190402
31. Mulugeta SM, Sárosi S, and Radácsi P. Physio-morphological trait and bioactive constituents of Ocimum species under drought stress. Ind Crops Products. (2023) 205:117545. doi: 10.1016/j.indcrop.2023.117545
32. Botero H and Barnes AP. The effect of ENSO on common bean production in Colombia: a time series approach. Food Secur. (2022) 14:1417–30. doi: 10.1007/s12571-022-01290-z
33. Manciu A, Rammig A, Krause A, and Quesada BR. Impacts of land cover changesand global warming on climate in Colombia during ENSO events. Climate Dynamics. (2023) 61:111–29. doi: 10.1007/s00382-022-06545-1
34. Díaz-Almanza ED, Cleves-Leguizamo JA, and Salgado-Ordosgoitia RD. Simulation of the potential effect of meteorological condition variations on forage production in native pastures in the warm climate of Colombia. Land. (2025) 14:397. doi: 10.3390/land14020397
35. Daza-Torres MC, Arias-Prado PC, Reyes-Trujillo A, and Urrutia-Cobo N. Basil (Ocimum basilicum L) water needs calculated from the crop coefficient. Ingeniería e Investigación. (2017) 37:08–16. doi: 10.15446/ing.investig.v37n3.65058
36. Lal R. Restoring soil quality to mitigate soil degradation. Sustainability. (2015) 7:5875–95. doi: 10.3390/su7055875
37. Havlin JL, Tisdale SL, Nelson WL, and Beaton JD. Soil fertility and fertilizers: an introduction to nutrient management. 9th ed. Upper Saddle River, New Jersey, USA: Pearson (2014).
38. Chaali N, Ouazaa S, Jaramillo C, Araujo G, and Ávila Pedraza E. Edaphoclimatic characterization and crop water requirement of Arracacha (Arracacia xanthorrhiza Bancroft) roots in upland production areas. Scientia Hortic. (2020) 272:109533. doi: 10.1038/s41561-019-0484-6
39. IDEAM. Atlas interactivo climatológico (2014). Available online at: https://archive.org/details/atlas-climatologico-de-Colombia/page/4/mode/2up (Accessed September 1, 2025).
40. Ávila ÁJA, Carvajal Y, and Gutiérrez SE. Análisis de la influencia de El Niño y La Niña en la oferta hídrica mensual de la cuenca del río Cali. Tecnura. (2014) 18(41):120–33. doi: 10.14483/udistrital.jour.tecnura.2014.3.a09
41. CPC-NOAA. Climatic Prediction Center, National Weather Service. Cold and Warm Episodes by Season. (2018). Available online at: http://origin.cpc.ncep.noaa.gov/products/analysis_monitoring/ensostuff/ONI_v4.shtml.
43. R Core Team. R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria (2023). Available at: https://www.r-project.org/.
44. Allen R, Pereira S, Raes D, and Smith M. Crop evapotranspiration: a guideline for computing crop water requirements. In: FAO irrigation and drainage paper no 56. FAO Water Resources, Development and Management Service, Rome, Italy (1998). p. 300.
45. Barrios-Perez C, Okada K, Garcés Varón G, Ramirez-Villegas J, Camila Rebolledo M, and Prager D:S. How does El Niño Southern Oscillation affect rice-producing environments in central Colombia? Agric For Meteorology. (2021) 306:108443. doi: 10.1016/j.agrformet.2021.108443
46. Ghamarnia H, Amirkhani D, and Arji I. Basil water use and crop coefficients in a semi-arid climate (lysimeter study). Int J Plant Prod./Journal Rep. (2014) 4(6):535–47. doi: 10.9734/IJPSS/2015/14098
47. IPCC, Pörtner H-O, Roberts DC, Poloczanska ES, Mintenbeck K, Tignor M, et al. Summary for policymakers. In: Rama B, editor. Climate change 2022: impacts, adaptation and vulnerability. Contribution of working group II to the sixth assessment report of the intergovernmental panel on climate change. Cambridge University Press, Cambridge, UK and New York, NY, USA (2022). p. 3–33. doi: 10.1017/9781009325844.001
48. Yang T and Kim H-J. Comparisons of nitrogen and phosphorus mass balance for tomato-, basil-, and lettuce-based aquaponic and hydroponic systems. J Cleaner Production. (2020) 274:122619. doi: 10.1016/j.jclepro.2020.122619
49. Medina M, Jayachandran K, Bhat MG, and Deoraj A. Assessing plant growth, water quality and economic effects from application of a plant-based aquafeed in a recirculating aquaponic system. Aquaculture Int. (2016) 24:415–27. doi: 10.1007/s10499-015-9934-3
50. Rakocy JE, Shultz RC, Bailey DS, and Thoman ES. Aquaponic production of tilapia and basil: comparing a batch and staggered cropping system. Acta Hortic. (2004) 648:63–9. doi: 10.17660/ActaHortic.2004.648.8
51. Passioura JB. Soil structure and plant growth. Soil Res. (1991) 29:717–28. doi: 10.1071/SR9910717
52. Mehalaine S and Chenchouni H. Plants of the same place do not have the same metabolic pace: soil properties affect differently essential oil yields of plants growing wild in semiarid Mediterranean lands. Arabian J Geosciences. (2020) 13:1263. doi: 10.1007/s12517-020-06219-4
53. Khalid AK, El-Gohary AE, and Ahmed AMA. Raising the efficiency of lemon trees to produce essential oil by exogenous cysteine under various soil structures. J Essen. Oil. Bear. Plant. (2020) 23:194–203. doi: 10.1080/0972060X.2020.1736646
54. Tursun AO. Impact of soil types on chemical composition of essential oil of purple basil. Saudi J Biol Sci. (2022) 29:103314. doi: 10.1016/j.sjbs.2022.103314
55. Lipiec J and Hatano R. Quantification of compaction effects on soil physical properties and crop growth. Geoderma. (2003) 116:107–36. doi: 10.1016/S0016-7061(03)00097-1
56. Bengough AG, McKenzie BM, Hallett PD, and Valentine TA. Root elongation, water stress, and mechanical impedance: A review of limiting stresses and beneficial root tip traits. J Exp Bot. (2011) 62:59–68. doi: 10.1093/jxb/erq350
57. Rioba NB, Itulya FM, Saidi M, Dudai N, and Bernstein N. Effects of nitrogen, phosphorus and irrigation frequency on essential oil content and composition of sage (Salvia officinalis L.). J Appl Res Med Arom. Plant. (2015) 2:21–9. doi: 10.1016/j.jarmap.2015.01.003
58. Naiji M and Souri MK. Nutritional value and mineral concentrations of sweet basil under organic compared to chemical fertilization. Acta Sci Pol Hortorum Cultus. (2018) 17:167–75. doi: 10.24326/asphc.2018.2.14
59. Zheljazkov VD, Cantrell CL, Tekwani B, and Khan SI. Content, composition, and bioactivity of the essential oils of three basil genotypes as a function of harvesting. J Agric Food Chem. (2008) 56:380–5. doi: 10.1021/jf0725629
60. Chaali N, Ramírez-Gómez CM, Jaramillo C, Garré S, Barrero O, Ouazaa S, et al. Enhancing irrigation management: Unsupervised machine learning coupled with geophysical and multispectral data for informed decision-making in rice production. Smart Agric Technol. (2024) 9:100635. doi: 10.1016/j.atech.2024.100635
61. Rizwan M, Gilani SR, Durani AI, and Naseem S. Materials diversity of hydrogel: Synthesis, polymerization process and soil conditioning properties in agricultural field. J Advanced Res. (2021) 33:15–40. doi: 10.1016/j.jare.2021.03.007
62. Iqbal DN, Tariq Z, Philips B, Sadiqa A, Ahmad M, Al-Ahmary KM, et al. Nanocellulose/wood ash-reinforced starch–chitosan hydrogel composites for soil conditioning and their impact on pea plant growth. RSC Adv. (2024) 14:8652–64. doi: 10.1039/D3RA08725E
63. Ćirić VI, Manojlović M, Svarc-Gajić J, and Šeremešić S. The assessment of soil organic carbon pools in different soils using four fractionation methods. Commun. Soil Sci Plant Anal. (2023) 54(14):1910–22. doi: 10.1080/00103624.2023.2211101
64. Roa-García CE, Brown SJ, Krzic M, and Lavkulich LM. Relationship of soil water retention characteristics and soil properties: a case study from the Colombian Andes. Can J Soil Sci. (2021) 101(1):147–56. doi: 10.1139/CJSS-2020-0066
65. Cai G, Carminati A, Abdalla M, and Ahmed MA. Soil textures rather than root hairs dominate water uptake and soil–plant hydraulics under drought. Plant Physiol. (2021) 187(2):858–72. doi: 10.1093/plphys/kiab271
66. Osorio N. Cómo interpretar los resultados del análisis de fertilidad del suelo. Bol Manejo Integr Suelo Nutric Veg. (2012) 1:1–3.
67. Pautler MC and Sims JT. Relationships between soil test phosphorus, soluble phosphorus, and phosphorus saturation in Delaware soils. Soil Sci Soc America J. (2000) 64:765–73. doi: 10.2136/sssaj2000.642765x
68. Bryson GM, Mills HA, Sasseville DN, Jones JB, and Barker AV. Plant analysis handbook III: A guide to sampling, preparation, analysis, interpretation and use of results of agronomic and horticultural crop plant tissue. Athens, Georgia, USA: Micro-Macro Publishing, Incorporated (2014).
69. Monib AW, Alimyar O, Mohammad MU, Akhundzada MS, and Niazi P. Macronutrients for plants growth and humans health. J Res Appl Sci Biotechnol. (2023) 2:268–79. doi: 10.55544/jrasb.2.2.38
70. Ramezani S, Rezaei MR, and Sotoudehnia P. Improved growth, yield and essential oil content of basil grown under different levels of phosphorus sprays in the field. J Appl Biol Sci. (2009) 3:105–10.
71. Solis P and Torrent J. Phosphate sorption by calcareous Vertisols and Inceptisols of Spain. Soil Sci Soc America J. (1989) 53:456–9. doi: 10.2136/sssaj1989.03615995005300020024x
72. Marschner H. Marschner’s mineral nutrition of higher plants. London, United Kingdom: Academic press (2011).
73. Song J, Yang J, and Jeong BR. Characterization of physiology, photosynthesis, and nutrition based on induced deficiencies of macro-and micronutrients in basil (Ocimum basilicum L.). Agronomy. (2024) 14:208. doi: 10.3390/agronomy14010208
74. Villar Mir P and Mir JMV. Guia de la fertilitat dels sòls i la nutrició vegetal en producció integrada. Ramaderia, Pesca i Alimentació: Departament d’Agricultura (2016).
75. Moro A. Relaciones catiónicas y su interpretación en los análisis de suelos. Obtenido de: AQM Laboratorios (2015). Available online at: http://aqmlaboratorios.com/author/alberto-moro (Accessed September 1, 2025).
76. Broadley M, Brown P, Cakmak I, Rengel Z, and Zhao F. Function of nutrients: micronutrients. In: Marschner’s mineral nutrition of higher plants. London, United Kingdom: Academic Press (2012). p. 191–248.
77. Tripathi DK, Singh S, Singh S, Mishra S, Chauhan DK, and Dubey NK. Micronutrients and their diverse role in agricultural crops: advances and future prospective. Acta Physiologiae Plantarum. (2015) 37:1–14. doi: 10.1007/s11738-015-1870-3
78. Millaleo R, Reyes-Díaz M, Ivanov AG, Mora ML, and Alberdi M. Manganese as essential and toxic element for plants: transport, accumulation and resistance mechanisms. J Soil Sci Plant Nutr. (2010) 10:470–81. doi: 10.4067/S0718-95162010000200008
79. Fischer WW, Hemp J, and Johnson JE. Manganese and the evolution of photosynthesis. Origins Life Evol Biospheres. (2015) 45:351–7. doi: 10.1007/s11084-015-9442-5
80. Londoño-Franco LF, Londoño-Muñoz PT, and Muñoz-García FG. Los riesgos de los metales pesados en la salud humana y animal. Biotecnología en el sector agropecuario y agroindustrial. (2016) 14:145–53. doi: 10.18684/BSAA(14)145-153
Keywords: soil compaction, El Niño-southern oscillation, precision agriculture, organic amendments, tropical dry ecosystems
Citation: Beltran-Medina JI, Ouazaa S, Chaali N, Jaramillo Barrios CI, Pedroza Berrío KJ, Hoyos Cartagena JA and Calderón Carvajal JE (2025) Assessing crop evapotranspiration and edaphoclimatic variability for basil (Ocimum basilicum L.) under ENSO-modulated tropical conditions in Colombia. Front. Soil Sci. 5:1621669. doi: 10.3389/fsoil.2025.1621669
Received: 01 May 2025; Accepted: 30 September 2025;
Published: 20 October 2025.
Edited by:
Tarun Belwal, Texas A and M University, United StatesReviewed by:
Ayesha Sadiqa, University of Lahore, PakistanRodrick Lepcha, Lovely Professional University, India
Copyright © 2025 Beltran-Medina, Ouazaa, Chaali, Jaramillo Barrios, Pedroza Berrío, Hoyos Cartagena and Calderón Carvajal. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Sofiane Ouazaa, c291YXphYUBhZ3Jvc2F2aWEuY28=