 Rayane Dias1,2
Rayane Dias1,2 Manuella Pereira Cerqueira Leite1,3
Manuella Pereira Cerqueira Leite1,3 Nathália Menezes de Almeida1,3
Nathália Menezes de Almeida1,3 Cecilia Ferreira de Mello1,2
Cecilia Ferreira de Mello1,2 Ana Laura Carbajal-de-la-Fuente4,5
Ana Laura Carbajal-de-la-Fuente4,5 Anderson S. Bueno6
Anderson S. Bueno6 Jeronimo Alencar1*
Jeronimo Alencar1*- 1Laboratório de Diptera, Instituto Oswaldo Cruz (Fiocruz), Rio de Janeiro, Brazil
- 2Programa de Pós-Graduação em Medicina Tropical, Instituto Oswaldo Cruz (Fiocruz), Rio de Janeiro, Brazil
- 3Programa de Pós-Graduação em Biologia Animal, Universidade Federal Rural do Rio de Janeiro, Seropédica, Brazil
- 4Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET), Ciudad Autónoma de Buenos Aires, Argentina
- 5Centro Nacional de Diagnóstico e Investigación en Endemo-Epidemias (CeNDIE), Administración Nacional de Laboratorios e Institutos de Salud “Dr. Carlos G. Malbrán” (ANLIS Malbrán), Ciudad Autónoma de Buenos Aires, Argentina
- 6Núcleo de Pesquisas em Ciências Biológicas, Instituto Federal de Educação, Ciência e Tecnologia Farroupilha, Júlio de Castilhos, Brazil
Introduction: The hatching rate of mosquito eggs can vary according to their biological characteristics and environmental adaptations. Some species produce more resistant eggs, while others are more sensitive and require particular conditions to trigger hatching. Understanding how repeated contact with water influences the mosquito life cycle is critical for the integrated management of these vectors of etiological agents.
Methods: Mosquito eggs were collected using 10 ovitraps placed in a forest remnant in the municipality of Uruaçu, state of Goiás, Brazil, during the rainy season in 2023. The egg-laden paddles were subjected to 27 weekly water-immersion cycles in the laboratory.
Results: Of the total eggs collected, 157 hatched and 136 reached the adult stage, representing four species. Aedes albopictus and Haemagogus leucocelaenus hatched after the first immersion, while Haemagogus janthinomys hatched between the 23rd and 25th immersions, and Aedes terrens hatched by the 26th immersion. The sex ratio of Ae. terrens showed a predominance of females (N = 72; 56.25%) over males (N = 56; 43.75%). Additionally, females required fewer immersion cycles to hatch compared to males.
Discussion: The impact of multiple water immersions on egg hatching varied among species, particularly for Ae. terrens and Hg. janthinomys, which required a greater number of immersions to hatch. These results provide valuable insights into the mosquito biology and carry important implications for public health, given the role of these species as vectors of pathogens.
1 Introduction
The development process of culicid eggs is subject to environmental conditions, particularly temperature and rainfall (1). The biological characteristics of a given species play a significant role in this process, enabling the development of various adaptations that increase its chances of survival and reproduction in different environments (2). These adaptations include egg diapause (3, 4), characterized by a temporary suspension of hatching after embryonic development has completed, a natural phenomenon in mosquitoes that lay their eggs outside of water. In mosquitoes of the tribe Aedini, egg diapause usually ends with the first immersion in water, representing the first contact of the egg with a larval habitat, as observed for Aedes aegypti (Linnaeus, 1762). However, for other species, the eggs may require additional water immersions to hatch (5, 6). In the case of Haemagogus janthinomys Dyar, 1921, adult densities increase up to two months after the beginning of the rainy periods, as the species’ eggs depend on rainfall but may need various immersions to hatch (7). Although the regulation of hatching deadline is known to vary across populations of the same species, as well as between sexes and among eggs from the same female (8, 9), it is understudied in mosquito species of the tribe Aedini, including those of the genera Aedes and Haemagogus.
Factors such as temperature and photoperiod can temporarily influence this suspension (10), as they are linked to the seasonality mosquitoes experience, including shorter days in certain seasons and extreme temperatures (11, 12). Therefore, it is essential to recognize that depending on the region, different species may undergo diapause induced by one or both factors. For instance, Aedes albopictus (Skuse, 1894) populations in South America, particularly in Brazil, may enter diapause in the southern part of the country (13).
The effect of multiple water immersions on egg development in immature mosquito forms can significantly impact the ecology and epidemiology of these insects, influencing their abundance and distribution (14). Insect vectors maintain a strong relationship with hydroclimatic environmental factors. Under specific temperature and rainfall conditions, these abiotic factors can alter the periodicity of blood feeding, longevity, and fecundity in Aedes females and similarly affect pathogenic agents and vectors (15, 16). Among these is Ae. albopictus, a natural vector of Zika virus (ZIKV) (17), which is also able to transmit dengue, yellow fever, and West Nile viruses in laboratory studies (18, 19). Species of the genus Haemagogus, which include vectors of the sylvatic yellow fever virus (20) such as Hg. janthinomys and Haemagogus leucocelaenus (Dyar & Shannon, 1924), can produce eggs highly resistant to desiccation (21). These eggs can hatch at the end of the rainy season, as observed for Hg. janthinomys (22); they are found at various heights, and in different natural or artificial habitats (23). Haemagogus janthinomys is the primary vector in the transmission of the wild yellow fever virus (YFV) and is almost exclusively found in rainforest environments. This species is active during the day and frequents the tree canopies to oviposit and feed on non-human primates. However, its behavior in inhabiting the tree canopy is somewhat variable (6, 21, 24). In the forest environment, the population abundance of this species is up to six times greater during the rainy season compared to the dry season (7). The ecological behavior of inhabiting the tree canopy has also been observed for Aedes terrens (Walker, 1856) (25, 26), a species that is competent at transmitting strains of the chikungunya virus (CHIKV) circulating in the Americas (28). Haemagogus janthinomys and Ae. terrens affect public health, with their activities in forests representing a potential risk factor for human populations living near forest fragments (24). Therefore, understanding the factors that influence the ecology and behavior of these mosquitoes in their natural habitat is crucial for designing strategies to control and prevent outbreaks of yellow fever and chikungunya, especially in forest regions and their transition zones to urban areas.
Considering their importance in pathogen transmission, analyzing how repeated immersions of eggs in water affect the mosquito development cycle is essential to understanding their hatching behavior. In addition to improving our understanding of the reproductive dynamics of these species, this knowledge can help implement more effective control strategies. In this context, we evaluated the effect of multiple water immersions on the hatching of male and female Aedes and Haemagogus eggs collected in a forest environment. Our study allowed us to identify possible variations in hatching behavior between species, which can help to anticipate outbreaks and implement targeted actions in risk areas.
2 Materials and methods
2.1 Ethical statement
The permanent license for the collection, capture, and transportation of zoological material was granted by the Instituto Chico Mendes de Conservação da Biodiversidade (ICMBio) through the Biodiversity Authorization and Information System (SISBIO) under the permit 84318-3. All members of the collection team were vaccinated against yellow fever.
2.2 Study area
The sampling area is located at Fazenda Pau Terra (14° 05’ 38.5” S; 48° 59’ 51.3” W) in the municipality of Uruaçu, state of Goiás, Brazil, in the Cerrado biome. The area features a gallery forest that follows small streams, forming a closed corridor over the watercourse, with tree canopies ranging between 20 and 30 meters in height. According to the Köppen classification, the climate is tropical with dry winter (Aw). The monthly temperature ranges from 21.7 to 25.8°C (mean ± standard deviation = 24.1 ± 1.4), with an annual precipitation of 1,786 mm, 94% of which occurs between October and April (28).
2.3 Mosquito egg collection and rearing
Mosquito eggs were collected from 4 January to 20 April 2023 during the rainy season (Figure 1). We used 10 ovitraps placed on tree trunks at a height of 1.5 m above the ground and spaced approximately 5 meters apart. Each ovitrap consisted of a 1-L matte black container and four plywood vanes (Eucatex® pallets, 2.5 cm x 14 cm) secured vertically with clips. Subsequently, all pallets were removed, properly labeled, and sent to the Diptera Laboratory at the Instituto Oswaldo Cruz for immersion.
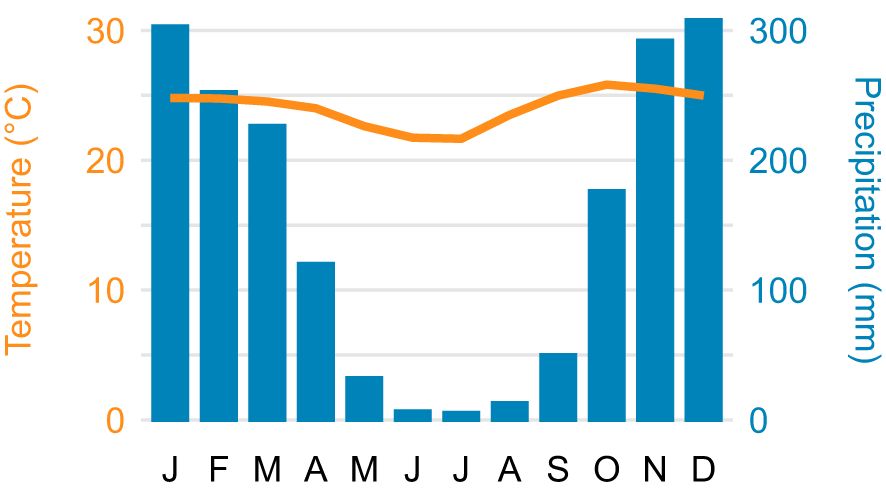
Figure 1. Climatogram depicting the monthly average temperature and the monthly total precipitation for Uruaçu, state of Goiás, Brazil, showing the marked seasonal variation in precipitation. Data from Alvares et al. (26).
In the laboratory, we counted the number of mosquito eggs on each paddle and then allowed them to dry at a room temperature of 27°C for 48 hours. The immersion process consisted of 27 separate water contacts with the eggs, each occurring on a weekly basis. The eggs were placed in 300 mL polyethylene containers, along with the paddles, and filled to one-third with dechlorinated water in a Biochemical Oxygen Demand (BOD) incubator (Eletrolab, São Paulo, Brazil) with accurate temperature and relative humidity control for three days, maintaining a temperature of 27°C, relative humidity between 75 and 90%, and a 12-hour photoperiod.
The hatched larvae during the immersions were counted and transferred to polyethylene containers kept inside adapted cages. They were fed twice a week with fish food at a ratio of 0.150 g per 10 mL of water until reaching the pupal stage, when adult emergence occurred. Containers with unhatched eggs had the water discarded and were removed from the incubator for approximately 72 hours before the next immersion cycle to allow embryonic development to progress.
Adult specimens were identified using dichotomous keys developed by Lane (29), Consoli and de Oliveira (2), and Forattini (6). Abbreviations for the generic names follow Reinert et al. (30). After species identification, all specimens were added to the Entomological Collection of the Instituto Oswaldo Cruz, Brazil.
2.4 Data analysis
We employed a descriptive analysis to quantify the minimum number of water immersions required for egg hatching of each species. Accordingly, we counted the number of eggs per species in each 3-day immersion instance from the first to the twenty-seventh.
To compare the egg hatching between female and male individuals of Ae. terrens, we employed a statistical analysis. This analysis was limited to Ae. terrens because it was the only species with a sufficient sample size. Since our data are in the form of time-to-event, where egg hatching is the ‘event’ and the number of 3-day water immersions is the ‘time’ when the event occurs, we performed a survival analysis (31). To estimate the probability of egg hatching as a function of the number of 3-day immersions, we generated Kaplan–Meier survival curves (31) without censoring because it was only possible to identify individuals to the species level that reached adulthood – that is, eggs that successfully hatched. Next, we applied the log-rank test to compare the Kaplan–Meier survival curves of female and male individuals of Ae. terrens (31). A statistical difference between the two curves (P < 0.05) indicates that the proportion of eggs hatched accumulates quicker for either female or male individuals, thereby requiring a fewer number of 3-day immersions to hatch all the eggs submitted to the experiment.
All analyses were performed in the software R version 4.3.2 (32), using the packages ggplot2 (33) and survminer (34) for data visualization, and the package survival (35) for the survival analysis and the log-rank test.
3 Results
A total of 739 eggs were counted, of which 530 had already hatched on the paddles. The remaining 209 eggs, counted on the paddles from the 10 ovitraps, underwent the immersion process, resulting in 157 larvae hatching. Of these, 136 reached adulthood and were distributed as follows: Ae. terrens (N = 128; 94.12%), Ae. albopictus (N = 4; 2.94%), Hg. janthinomys (N = 2; 1.47%), and Hg. leucocelaenus (N = 2; 1.47%). Aedes terrens was present in 6 of the 10 traps, Ae. albopictus in 2, and both Hg. leucocelaenus and Hg. janthinomys in 1 (Figure 2).
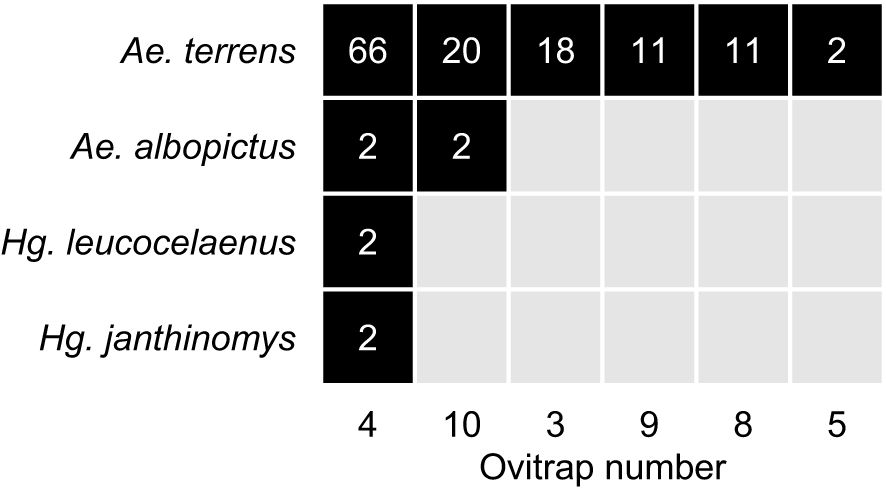
Figure 2. Species by ovitrap abundance matrix showing the number of individuals that hatched during the water immersion procedure. Ten ovitraps were placed in a forest remnant located in Uruaçu, state of Goiás, Brazil, of which six contained mosquito eggs that reached adulthood and were identified in the laboratory.
Aedes terrens had the highest number of individuals identified in the immersions where egg hatching occurred, except during the 25th immersion when only Hg. janthinomys hatched. Aedes terrens eggs hatched in 13 out of 27 immersions, while Hg. janthinomys hatched in 2 immersions, and both Ae. albopictus and Hg. leucocelaenus hatched in a single immersion (Figure 3). Aedes albopictus and Hg. leucocelaenus eggs hatched during the first immersion, whereas Ae. terrens exhibited hatching viability up to the 26th immersion. The viability of Hg. janthinomys eggs was observed between the 23rd and 25th immersions to which they were subjected (Figure 3).
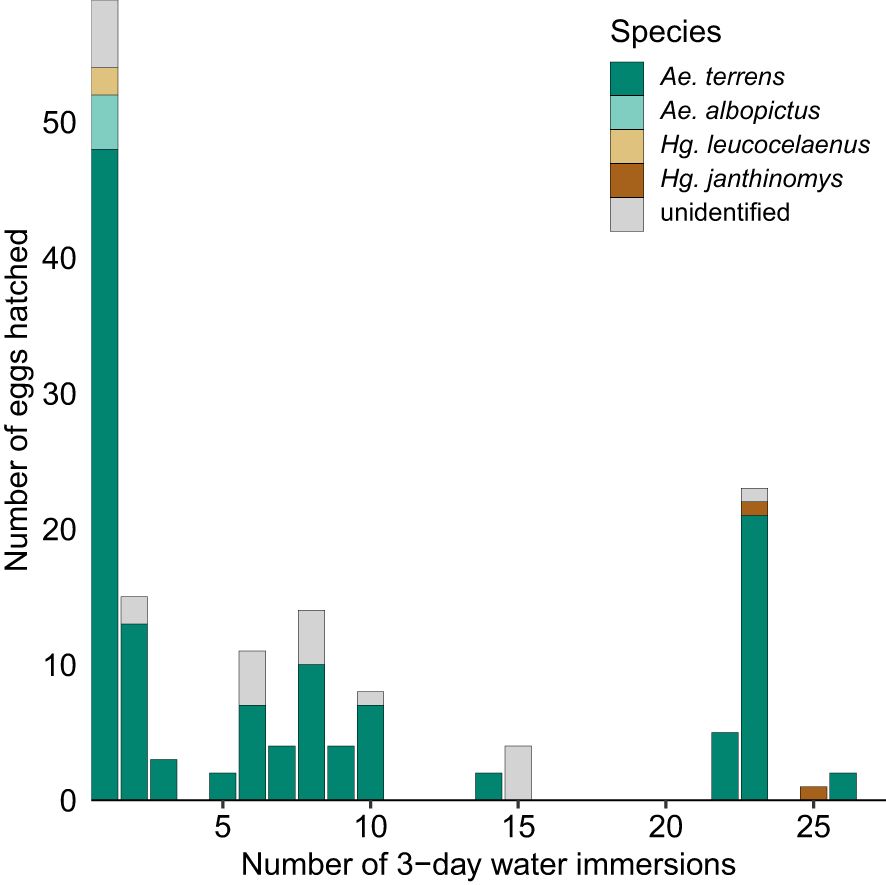
Figure 3. Effect of multiple water immersions on the hatching of eggs of Aedes terrens, Aedes albopictus, Haemagogus leucocelaenus, and Haemagogus janthinomys from a forest remnant located in Uruaçu, state of Goiás, Brazil.
The effect of multiple water immersions on egg hatching varied among species, particularly for Ae. terrens and Hg. janthinomys. These two species likely entered diapause or quiescence and required a greater number of immersions to hatch. This led to a significant increase in Ae. terrens individuals during the 23rd immersion, coinciding with the first hatching of Hg. janthinomys.
Regarding the total number of Ae. terrens eggs that hatched and reached adulthood (n = 128), 72 (56.25%) were female individuals and 56 (43.75%) were male individuals. When comparing hatching across immersion events, female eggs required a smaller number of immersions than male eggs (Log-rank: χ2 = 5.79; P = 0.016; Figure 4). Accordingly, two immersions were necessary to hatch 50% of the female eggs (95% CI = 1–7 immersions), and seven for male eggs (95% CI = 2–14 immersions).
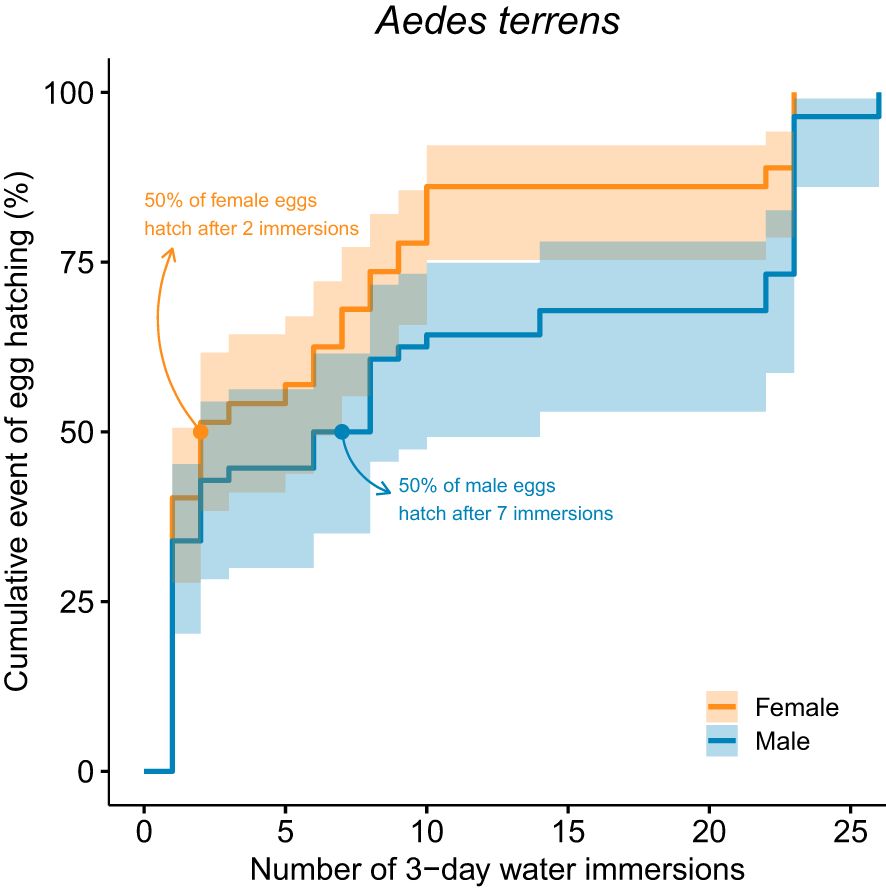
Figure 4. Kaplan–Meier survival curves for female and male eggs of Aedes terrens, showing the probability of hatching as a function of multiple water immersions. Shaded areas correspond to the 95% confidence intervals of the survival curves.
4 Discussion
Evaluating the hatching rate of mosquito eggs collected in the wild after multiple water immersions is a crucial aspect of entomological research, providing valuable insights into the ecology and reproductive biology of these insects. Additionally, it offers important epidemiological information, enabling the assessment and development of control measures for scenarios that may lead to an increase in the number of individuals of certain vector species, depending on the varying water contact requirements of their eggs (36).
Although it does not fully replicate field conditions, assessing hatching rates in a controlled environment represents a methodologically effective approach, as it allows for the isolation of the central factor under investigation in this study—namely, the repeated exposure of eggs to water—while eliminating potential interference from external environmental variables. Maintaining constant temperature (27°C), relative humidity (75–90%), and photoperiod (12:12 hours) ensures that environmental fluctuations, which could directly influence embryonic activation and hatching time, do not affect the results obtained. This experimental standardization is particularly important in studies involving species capable of entering diapause or quiescence, as observed in various representatives of Aedes and Haemagogus, for which inappropriate environmental stimuli (such as extreme temperatures, low humidity, or shortened photoperiods) may delay or inhibit hatching even after exposure to water (37, 38). By controlling these variables, the data more clearly reflect the direct effect of multiple water immersions on the breaking of egg dormancy.
Furthermore, although the hatching patterns observed in the laboratory are obtained under controlled conditions, they are consistent with behaviors documented in natural environments, provided that environmental conditions fall within the physiological tolerance range of the species studied. Therefore, the approach adopted here enables not only a more precise understanding of the hatching mechanism but also provides meaningful insights applicable to natural ecological settings.
In a study conducted in a forest remnant of the Atlantic Forest in the Brazilian state of Rio de Janeiro, Silva et al. (39) reported that eggs collected from Ae. albopictus mostly hatched during the first immersion in water, with a minority of eggs requiring a maximum of two immersions to hatch. However, despite normally requiring only a few immersions to hatch, eggs of Hg. leucocelaenus and Hg. janthinomys exhibited installment hatching up to the 22nd and 37th immersion, respectively. Similar results were found for the same three mosquito species in the same study region (9). However, in the present study, Ae. terrens did not show a continuous decrease in the number of hatchings across immersions as previously reported (9, 40). Instead, it showed unexpectedly delayed egg hatchings, with a second peak in the number of hatchings at the 23rd immersion, indicating that Ae. terrens may sustain viable eggs over extended periods.
The delayed hatching of Hg. janthinomys demonstrated the necessity for multiple water immersions for successful hatching, distinguishing it from the other three species studied. Despite Ae. terrens experiencing hatchings after multiple water immersions, its highest peak occurred after the first immersion. The unique hatching behavior of Hg. janthinomys potentially enables the prolonged maintenance of the wild yellow fever virus transmission cycle due to the robustness of its eggs and the presence of transovarial transmission.
Galindo et al. (41) reported that Hg. janthinomys eggs are resistant to desiccation and only hatch after a prolonged period of inactivity and repeated contact with water. The total duration of the development cycle varies according to abiotic factors such as temperature. The prolonged hatching viability is also documented in Alencar et al. (14), who evaluated the effect of various water immersions on the eggs of Hg. janthinomys from a forest remnant located in Linhares, state of Espírito Santo, Brazil. These authors noted that the eggs started hatching during the first immersion, with the last hatching recorded at the 16th immersion.
Likewise, Silva et al. (9, 39) observed egg hatching from the first immersion, diverging from our observations since Hg. janthinomys required a greater number of immersions to achieve egg hatching compared to what was observed in the Atlantic Forest.
Egg hatching also seems to be related to the influence of climatic variations. The presence of water, combined with suitable temperature and humidity conditions, seems to be fundamental to triggering this process. Conversely, there is evidence of specific differences between species, which supports the hypothesis that each species may respond differently to these environmental stimuli (42).
Galindo et al. (41) suggested two distinct patterns of hatching behavior in some Haemagogus species. In the case of Haemagogus equinus Theobald, 1903, eggs hatch on first contact with water and continue to hatch on subsequent contacts for a prolonged period until all viable eggs have hatched. In contrast, Hg. janthinomys and Haemagogus capricornii Lutz, 1904 show a different behavior, in which hatching occurs after the third contact with water. These differences in hatching behavior result in significant discrepancies in the abundance of specimens throughout the year. In the first case, the number of Hg. equinus specimens tend to increase at the start of the rainy season due to the rapid response to humidity. For Hg. janthinomys and Hg. capricornii, the abundance of specimens only reaches maximum values after a few months of exposure to water, indicating that there is a slower or more specific physiological response to prolonged environmental stimuli. These differences may reflect distinct adaptations to the climatic and ecological conditions of the regions in which these species live. Therefore, understanding these variations in hatching behavior is fundamental to predicting the population dynamics and spread of mosquitoes, especially concerning disease transmission.
When evaluating the sex ratio between females and males of medically important mosquito species from the Atlantic Forest, Silva et al. (9) found a sex ratio skewed toward males for Ae. terrens in relation to the total number of individuals hatched after multiple water immersions. In contrast, for the eggs we collected in the Cerrado, we found a higher proportion of females in all immersions except the last one, as all female eggs had already hatched due to the higher hatching rate of females compared to males (9). Regarding Ae. albopictus, Silva et al. (9) found more females than males, whereas Lounibos and Escher (43) reported no difference in the proportion of females and males collected in tree holes in Vero Beach, Florida, United States of America. Taken together, these results indicate that Aedes species can exhibit similar proportions of females and males (Ae. albopictus, 43), a female-biased ratio (Ae. albopictus, 9; Ae. terrens, this study), or a male-biased ratio (Ae. terrens, 9). Thus, the sex ratio appears to vary among Aedes species and, for the same species, between different collection sites. The instances where the proportion of females exceeds that of males are particularly important for public health, as only female mosquitoes engage in blood-feeding.
The species identified in our study hold significant epidemiological importance for the transmission of arboviruses. Haemagogus species are primary vectors of the yellow fever virus in Brazil (20), while Ae. albopictus is recognized as a natural vector of the Zika virus (ZIKV) (17). Additionally, Ae. terrens has been shown to transmit the chikungunya virus in experimental settings (27). Among the four species recorded in the ovitraps we placed in the Cerrado, Ae. terrens was the most pervasive, abundant, and dominant compared to the other species present in the same ovitraps. Differently, in two studies carried out in the Atlantic Forest, Ae. terrens was the third most abundant of the four species we recorded, representing only 15% of the total number of individuals (9), and it was never recorded in another yet similar study (39). This shows that the prevalence of medically important mosquito species varies across biomes, implying that the species posing greater health concerns are likely to differ even when considering both the same habitat type (i.e., forest remnants) and species pool.
Data availability statement
The data used in the study are included in the article/mosquito_data.txt. Further inquiries can be directed to the corresponding author.
Author contributions
RD: Investigation, Methodology, Writing – original draft. MC: Data curation, Investigation, Methodology, Writing – review & editing. NA: Data curation, Investigation, Methodology, Writing – review & editing. CM: Data curation, Investigation, Methodology, Writing – review & editing. AC: Writing – review & editing. AB: Formal Analysis, Visualization, Writing – review & editing. JA: Conceptualization, Funding acquisition, Methodology, Resources, Supervision, Writing – review & editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This work was carried out with the support of Conselho Nacional de Desenvolvimento Científico e Tecnológico – CNPq (grant 303286/2021-0, J.A.) and Fundação Carlos Chagas Filho de Amparo à Pesquisa do Estado do Rio de Janeiro – FAPERJ (grant E-26/200.956/2002/2022, J.A.). R.D. was funded by a PhD studentship from Coordenação de Aperfeiçoamento de Pessoal de Nível Superior – CAPES.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fitd.2025.1590603/full#supplementary-material
References
1. Silva SOF, de Mello CF, Érico Guimarães A, José Leite P, and Alencar J. Occurrence of the Sylvatic Yellow Fever Vector Mosquito Haemagogus leucocelaenus (Diptera: Culicidae) in an Atlantic Forest Fragment of the Touristic State of Rio de Janeiro, Brazil. J Med Entomol. (2022) 59:1891–7. doi: 10.1093/jme/tjac153
2. Consoli RAGB and Lourenço-De-Oliveira R. Principais Mosquitos de Importância Sanitária do Brasil. Rio de Janeiro, Brasil: Editora FIOCRUZ (1994). doi: 10.7476/9788575412909
3. Vinogradova EB. Diapause in aquatic insects, with emphasis on mosquitoes. In: Alekseev VR, de Stasio BT, and Gilbert JJ, editors. Diapause in Aquatic Invertebrates Theory and Human Use. Springer, Dordrecht (2007). p. 83–113. doi: 10.1007/978-1-4020-5680-2_5
4. Denlinger DL and Armbruster PA. Mosquito diapause. Annu Rev Entomol. (2014) 59:73–93. doi: 10.1146/annurev-ento-011613-162023
6. Forattini OP. Culicidologia Médica: Identificação, Biologia, Epidemiologia. São Paulo, Brasil: Edusp—Editora da Universidade de São Paulo (2002).
7. Chadee DD, Tikasingh ES, and Ganesh R. Seasonality, biting cycle and parity of the yellow fever vector mosquito Haemagogus janthinomys in Trinidad. Med Vet Entomol. (1992) 6:143–8. doi: 10.1111/j.1365-2915.1992.tb00592.x
8. Gillett JD. Variation in the hatching-response of Aedes eggs (Diptera: Culicidae). Bull Entomol Res. (1955) 46:241–54. doi: 10.1017/S0007485300030881
9. Silva SOF, de Mello CF, Julião GR, Dias R, and Alencar J. Sexual proportion and egg hatching of vector mosquitos in an Atlantic forest fragment in Rio de Janeiro, Brazil. Life. (2023) 13:13. doi: 10.3390/life13010013
10. Garzón M Maffey JL, Lizuain A, Soto D, Diaz PC, Leporace M, Salomón OD, et al. Temperature and photoperiod effects on dormancy status and life cycle parameters in Aedes albopictus and Aedes aegypti from subtropical Argentina. Med Vet Entomol. (2021) 35:97–105. doi: 10.1111/mve.12474
11. Vinogradova EB. Diapausa em insetos aquáticos, com ênfase em mosquitos. In: Diapausa em invertebrados aquáticos: Teoria e uso humano. Springer, Dordrecht (2007). p. 83–113.
12. Fischer S, De Majo MS, Di Battista CM, Montini P, Loetti V, and Campos RE. Adaptation to temperate climates: Evidence of photoperiod-induced embryonic dormancy in Aedes aegypti in South America. J Insect Physiol. (2019) 117:103887. doi: 10.1016/j.jinsphys.2019.05.005
13. Lounibos LP, Escher RL, and Lourenço-De-Oliveira R. Asymmetric evolution of photoperiodic diapause in temperate and tropical invasive populations of Aedes albopictus (Diptera: Culicidae). Ann Entomol Soc Am. (2003) 96:512–8. doi: 10.1603/0013-8746(2003)096[0512:AEOPDI]2.0.CO;2
14. Alencar J, De Almeida HM, Marcondes CB, and Guimarães AÉ. Effect of multiple immersions on eggs and development of immature forms of Haemagogus janthinomys from south-eastern Brazil (Diptera: Culicidae). Entomol News. (2008) 119:239–44. doi: 10.3157/0013-872X(2008)119[239:EOMIOE]2.0.CO;2
15. Calado DC and Navarro-Silva MA. Influência da temperatura sobre a longevidade, fecundidade e atividade hematofágica de Aedes (Stegomyia) albopictus Skuse, 1894 (Diptera, Culicidae) sob condições de laboratório. Rev Bras Entomol. (2002) 46:93–8. doi: 10.1590/S0085-56262002000100011
16. Confalonieri UEC. Variabilidade climática, vulnerabilidade social e saúde no Brasil. Terra Livre. (2003) 1:193–204. doi: 10.62516/terra_livre.2003.185
17. Grard G, Caron M, Mombo IM, Nkoghe D, Mboui Ondo S, Jiolle D, et al. Zika virus in Gabon (Central Africa)—2007: A new threat from Aedes albopictus? PloS Negl Trop Dis. (2014) 8:e2681. doi: 10.1371/journal.pntd.0002681
18. Lourenço de Oliveira R, Vazeille M, de Filippis AM, and Failloux AB. Large genetic differentiation and low variation in vector competence for dengue and yellow fever viruses of Aedes albopictus from Brazil, the United States, and the Cayman Islands. Am J Trop Med Hyg. (2003) 69:105–14. doi: 10.4269/ajtmh.2003.69.105
19. Rothman SE, Jones JA, LaDeau SL, and Leisnham PT. Higher West Nile virus infection in Aedes albopictus (Diptera: Culicidae) and Culex (Diptera: Culicidae) mosquitoes from lower income neighborhoods in urban Baltimore, MD. J Med Entomol. (2021) 58:1424–8. doi: 10.1093/jme/tjaa262
20. Abreu FVS, Ribeiro IP, Ferreira-de-Brito A, Santos AACD, Miranda RM, Bonelly IS, et al. Haemagogus leucocelaenus and Haemagogus janthinomys are the primary vectors in the major yellow fever outbreak in Brazil, 2016–2018. Emerg Microbes Infect. (2019) 8:218–31. doi: 10.1080/22221751.2019.1568180
21. Marcondes C and Alencar J. Revisão de mosquitos Haemagogus Williston (Diptera: Culicidae) do Brasil. Rev Biomédico. (2010) 21:221–38. Available online at: https://revistabiomedica.uady.mx/index.php/revbiomed/article/view/115 (Accessed May 16, 2025).
22. Dégallier N, Monteiro HAO, Castro FC, Silva OV, Sá Filho GC, and Elguero E. An indirect estimation of the developmental time of Haemagogus janthinomys (Diptera: Culicidae), the main vector of yellow fever in South America. Stud Neotrop Fauna Environ. (2006) 41:117–22. doi: 10.1080/01650520500398662
23. Aguiar MD, Bastos AQ, Leite PJ, Gil-Santana HR, Santos SJ, and Alencar J. Comparative analysis between sampling methods for immature mosquitoes in an Atlantic forest fragment in Brazil. J Am Mosq Control Assoc. (2020) 4:245–8. doi: 10.2987/20-6947.1
24. Vasconcelos PFC. Febre amarela. Rev Soc Bras Med Trop. (2003) 36:275–93. doi: 10.1590/S0037-86822003000200012
25. Arnell JH. Mosquito studies (Diptera, Culicidae) XXXII. A revision of the genus Haemagogus. Contrib Am Entomol Inst. (1973) 10:1–174 Available online at: https://archive.org/download/biostor-286108/biostor-286108.pdf (Accessed May 16, 2025).
26. Guimarães AÉ and Arlé M. Mosquitos no Parque Nacional da Serra dos Órgãos, estado do Rio de Janeiro, Brasil: II. Distribuição vertical. Memórias Do Inst Oswaldo Cruz. (1985) 80:171–85. doi: 10.1590/S0074-02761985000200008
27. Lourenço-de-Oliveira R and Failloux AB. High risk for chikungunya vírus to initiate an enzootic sylvatic cycle in the tropical Americas. PloS Negl Trop. (2017) 11:e0005698. doi: 10.1371/journal.pntd.0005698
28. Alvares CA, Stape JL, Sentelhas PC, De Moraes Gonçalves JL, and Sparovek G. Köppen’s climate classification map for Brazil. Meteorol Z. (2013) 22:711–28. doi: 10.1127/0941-2948/2013/0507
30. Reinert JF, Harbach RE, and Kitching IJ. Phylogeny and classification of Aedini (Diptera: Culicidae). Zool J Linn Soc. (2009) 157:700–94. doi: 10.1111/j.1096-3642.2009.00570.x
31. Rich JT, Neely JG, Paniello RC, Voelker CC, Nussenbaum B, and Wang EW. A practical guide to understanding Kaplan-Meier curves. Otolaryngol Head Neck Surg. (2010) 143:331–6. doi: 10.1016/j.otohns.2010.05.007
32. R Core Team. R: A language and environment for statistical computing. Vienna, Austria: R Foundation for Statistical Computing (2023). Available at: https://www.R-project.org (Accessed May 16, 2025).
33. Wickham H. ggplot2: Elegant Graphics for Data Analysis. New York: Springer-Verlag (2016). doi: 10.1007/978-3-319-24277-4
34. Kassambara A, Kosinski M, and Biecek P. survminer: Drawing Survival Curves using ‘ggplot2’. R package version 0.4.9 (2021). Available online at: https://CRAN.R-project.org/package=survminer (Accessed May 16, 2025).
35. Therneau T. A Package for Survival Analysis in R. R package, version 3.5-7 (2023). Available online at: https://CRAN.R-project.org/package=survival (Accessed May 16, 2025).
37. Rezende GL, Martins AJ, Gentile C, Farnesi LC, Pelajo-MaChado M, and Valle D. Embryonic desiccation resistance in Aedes aegypti: presumptive role of the chitinized serosal cuticle. BMC Dev Biol. (2008) 8:82. doi: 10.1186/1471-213X-8-82
38. Farnesi LC, Barbosa CS, Araripe LO, and Bruno RV. Influence of Aedes aEgypti egg viability on mosquito population dynamics. PloS One. (2015) 10:e0132425. doi: 10.1371/journal.pone.0132425
39. Silva SOF, de Mello CF, Gleiser RM, Oliveira AA, Maia DA, and Alencar J. Evaluation of Multiple Immersion Effects on Eggs From Haemagogus leucocelaenus, Haemagogus janthinomys, and Aedes albopictus (Diptera: Culicidae) Under Experimental Conditions. J Med Entomol. (2018) 55:1093–7. doi: 10.1093/jme/tjy080
40. Alencar J, Gleiser RM, Morone F, de Mello CF, dos Silva JS, Serra-Freire NM, et al. A comparative study of the effect of multiple immersions on Aedini (Diptera: Culicidae) mosquito eggs with emphasis on sylvan vectors of yellow fever virus. Mem Inst Oswaldo Cruz. (2014) 109:114–7. doi: 10.1590/0074-0276130168
41. Galindo P, Carpenter SJ, and Trapido HA. Contribution to the ecology and biology of tree hole breeding mosquitoes of Panama. Ann Entomological Soc America. (1955) 48:158–64. doi: 10.1093/aesa/48.3.158
42. Hovanitz W. Comparisons of mating behavior, growth rate, and factors influencing egg-hatching in South American Haemagogus mosquitoes. Physiol Zool. (1946) 19:35–53. doi: 10.1086/physzool.19.1.30151878
Keywords: Aedini, Aedes, Haemagogus, egg hatching, bioecology
Citation: Dias R, Cerqueira Leite MP, Almeida NMd, de Mello CF, Carbajal-de-la-Fuente AL, Bueno AS and Alencar J (2025) Evaluation of multiple water immersions of pathogen vector mosquito (Diptera: Culicidae) eggs. Front. Trop. Dis. 6:1590603. doi: 10.3389/fitd.2025.1590603
Received: 11 March 2025; Accepted: 05 May 2025;
Published: 30 May 2025.
Edited by:
Alessandra Gutierrez Oliveira, Universidade Federal de Mato Grosso do Sul, BrazilReviewed by:
Karina Salvatierra, Universidad Nacional de Misiones, ArgentinaDiego Alonso, São Paulo State University, Brazil
Copyright © 2025 Dias, Cerqueira Leite, Almeida, de Mello, Carbajal-de-la-Fuente, Bueno and Alencar. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Jeronimo Alencar, amFsZW5jYXJAaW9jLmZpb2NydXouYnI=