 Fabio Albanese1,2*
Fabio Albanese1,2* Matthew Harrison1
Matthew Harrison1 Michelle J. Farquhar1
Michelle J. Farquhar1 Lucy J. Holcombe1
Lucy J. Holcombe1 Jujhar Atwal1
Jujhar Atwal1 Phil Watson1
Phil Watson1 Anna M. Piccinini2
Anna M. Piccinini2- 1WALTHAM Petcare Science Institute, Melton Mowbray, Leicestershire, United Kingdom
- 2School of Pharmacy, University of Nottingham, Nottingham, United Kingdom
The immune system of kittens is less efficient at fighting pathogens compared to adult cats with kittens being more susceptible to infections. Increasing evidence shows that dietary interventions can enhance immunity in mammals and modulate the expression of microRNAs (miRNAs) with key immune functions, however research in kittens is limited. Diets that can enhance the ability of a kitten's developing immune system to successfully fight infections, and where effects can be monitored by veterinarians, are highly sought-after. Here, we utilised small RNA sequencing (RNA-Seq) to investigate the effects of an experimental (test) diet containing nucleotides and oligosaccharides on the global expression of circulating miRNAs in 23-week-old kittens (n = 9). Furthermore, we determined whether these effects were sustained up to 10 weeks post-supplementation. Kittens fed with the test diet were found to have a lower expression of a specific subset of circulating miRNAs, namely miR-1-3p, miR-133a-3p, miR-206-3p and miR-383-5p, compared to animals fed with a control diet. Notably, this effect persisted 10 weeks post-supplementation. Bioinformatics analysis indicated that these miRNAs target immune-related genes and pathways. As such, they may hold potential as biomarkers to monitor immune performance of kittens and inform the prescription of veterinary diets.
1 Introduction
Early life represents a critical stage in the development of the feline immune system, during this time kittens are more susceptible to infections than adult animals due to their inability to mount an effective immune response (1). Dietary interventions have previously shown the potential to enhance immunity in adult cats and dogs (2, 3) however, research in kittens is limited. Additional investigation in kittens is necessary for the formulation of specific diets able to support the development of the immune system, providing animals with a higher level of protection against pathogens during a critical stage of their development.
Recently, a review has shed light on the ability of nutrition to modulate gene expression through regulation of microRNAs (miRNAs) (4); endogenous ~22 nucleotide long non-coding RNAs that post-transcriptionally regulate gene expression (5) by targeting specific mRNAs (6). Since their discovery, the identification of miRNAs as immune-biomarkers in humans has increased because of interest in their ability to regulate several immune mechanisms, including the development, differentiation, and function of immune cells (6–8). Efforts are being made to use them for the early detection of infection (9) and to predict vaccine efficacy following immunisation (10). Therefore, the identification of immune-related miRNAs in kittens where expression is affected by nutrition may help veterinarians monitor immune system development and vaccination response, diagnose defective immune functions, and inform clinical interventions.
In mammals, previous work has demonstrated the ability of specific nutrients and overall nutrition to influence the expression of endogenous miRNAs (11), which in turn can regulate immune pathways. Indeed, studies conducted in cell lines revealed the ability of ingredients such as resveratrol to upregulate the anti-inflammatory miR-693, which decreased the activity of AP-1 in THP-1 cells (12). Additionally, fatty acids inhibited the expression of PTEN in hepatocytes by up-regulating miR-21 synthesis (13). Knockout experiments using cell line models in mice demonstrated the important role that miRNAs play during the maturation of immune cells. In fact, a defective interaction between dendritic cells and T lymphocytes, and the inability to mount acquired immune responses, were observed following miR-155 knockout (14, 15). Moreover, the absence of the RNA endonuclease Dicer caused an impaired development of T-helper (Th) lymphocytes and a reduced proliferation of T cells upon stimulation (16, 17). In kittens, Zhou et al. (18) showed that the expression level of miRNAs in 10-week-old animals was altered following in vivo infection with Feline parvovirus (FPV), and that several miRNAs potentially regulated the JAK-STAT pathway, thus affecting the synthesis of cytokines with antiviral effect and growth factors generally critical for immune development and haematopoiesis. Furthermore, besides endogenous miRNAs, growing evidence shows that exogenous miRNAs can regulate gene expression between different species (19). Specifically, mammals can uptake exogenous miRNAs, especially food-derived miRNAs (xeno-miRNAs), which have been detected as functional molecules in the blood and other tissues of the host, following the ingestion of vegetables, cereals and fruits (20–22).
Nucleotides are considered semi-essential nutrients that provide energy carrier molecules such as ATP and GTP for energy use and storage at the cellular level and protein synthesis, respectively (23). Although nucleotides are naturally synthesised in the body, tissues with a rapid turnover such as the intestinal epithelium and the lymphoid tissue utilise the salvage pathway and recover nucleosides and nucleobases from the degradation of RNA and DNA. Moreover, when de novo synthesis and salvage pathway do not fulfil the needs of cell nucleotides (e.g. during growth, development and inflammation), cells can utilise nucleotides from exogenous sources, including the diet (24, 25). There is evidence that dietary nucleotides positively support immunity, tissue growth and development in infants (23, 26). Nucleotide supplementation also increases bacterial resistance to several pathogens in mice (27–29), as well as supporting the maturation of murine T helper lymphocytes (30) and enhancing the antibody response following influenza and diphtheria immunisations in infants (31).
Prebiotics are non-digestible dietary carbohydrate fibres contained in cereals, fruits, and vegetables, which have been identified as being able to support the development of the gut microbiota and the synthesis of several metabolites, including short chain fatty acids (SCFA), with health benefits (32). Among several prebiotics that occur in nature, fructo-oligosaccharides (FOS) and xylo-oligosaccharides (XOS) are two well-known prebiotics that have been shown to beneficially affect immunity in mammals, including cats and dogs (33–35).
To date, although there is no evidence for the ability of dietary supplementation of nucleotides to affect the expression of endogenous miRNAs, studies have reported that carbohydrates such as FOS and XOS can influence the expression of miRNAs in mammals by epigenetic alterations (e.g. methylation or acetylation) of the promoter regions of miRNAs (36, 37). Specifically, research conducted in rats showed that dietary supplementation of XOS significantly decreased the expression of miR-192-5p and miR-221-3p in the liver, and this was linked to reduced hepatic inflammation (38). Alleviation of hepatic steatosis was also observed in mice following dietary supplementation of FOS and was attributed to decreased expression of miR-33 in the liver which may have contributed to increased fatty acid oxidation (39), as shown in an independent murine study (40). Moreover, Lim et al. (41) showed that mice fed a diet containing xylobiose, a major component of XOS, displayed decreased expression of miR-33a and increased levels of miR-122a in the liver and that this contributed to a reduced hepatic accumulation of lipids.
The potential of dietary ingredients to modulate the expression of endogenous miRNAs and the impact this could have on the immune system and the physiological response to vaccine challenge, prompted us to investigate the effects of feeding a diet fortified with nucleotides, short chain FOS (scFOS) and XOS (test diet) on circulating plasma miRNAs in 23-week-old kittens. We also determined the longer-term impact of the dietary supplements on the levels of plasma miRNAs by quantifying their levels 2 and 10 weeks after the test diet was withdrawn.
2 Materials and methods
2.1 Animals and study design
Domestic short haired kittens used in this study were bred and housed in purpose-built, environmentally enriched housing at the Waltham Petcare Science Institute. All 16 kittens (n = 5 females and n = 11 males) enrolled in this study were clinically healthy. This work was reviewed and approved by the Waltham Animal Welfare and Ethical Review Board and conducted under the authority of the Animals (Scientific Procedures) Act 1986. In a blinded parallel study, six entire litters of kittens were randomly allocated to either a control (n = 9 kittens; n = 4 litters) or test (n = 7 kittens; n = 2 litters) diet group (Figure 1). Blood samples were used to conduct analysis of miRNAs. Kittens were enrolled in this study at 3 weeks of age and fully weaned by 10 weeks. A number of male kittens (three in the control group and one in the test group) were neutered at 29 weeks of age. Following blood collection at 35-weeks of age, test diet kittens were moved onto the control diet until 45 weeks old (Test-Control; T-C kittens), whereas kittens fed with the control diet were maintained on the same diet until 45 weeks of age (Control-Control; C-C kittens). The expression of plasma miRNAs was determined at 23, 37 and 45 weeks.
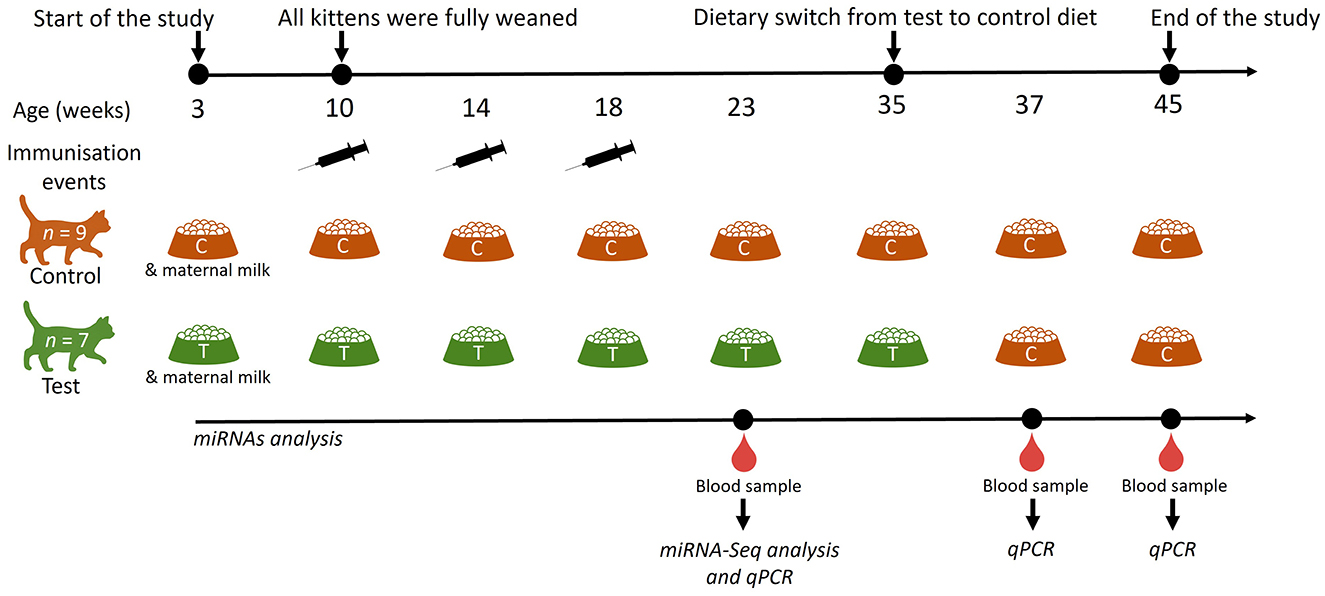
Figure 1. Schematic representation of the study design to determine the effects of dietary nucleotides and oligosaccharides on the expression of circulating miRNAs. Kittens were fed a control (C; n = 9) or a test (T; n = 7) diet from 3 weeks of age; all kittens were fully weaned at 10 weeks of age. miRNA analysis was performed at 23-, 37- and 45-weeks of age. The test diet group was moved onto a control diet at 35 weeks of age. Cats were vaccinated with Fevaxyn® Pentofel vaccine at 10, 14 and 18 weeks of age.
2.2 Vaccinations
Kittens received, as part of a standard veterinary recommended vaccination protocol, a nasal administration of Nobivac Bb® vaccine at 4 weeks of age that contained live attenuated Bordetella bronchiseptica bacteria, and a subcutaneous injection of Fevaxyn® Pentofel vaccine (Zoetis UK Ltd.) at 10, 14 and 18 weeks (± 1 week) of age that contained inactivated forms of feline herpesvirus (strain 605), parvovirus (strain CU4), calicivirus (strain 255), leukaemia virus (strain 61E) and Chlamydophila felis bacteria (strain Cello).
2.3 Diets
The two experimental diets (control and test) used in this study were extruded dry diets formulated and manufactured by Royal Canin (France). Both contained dried poultry protein, animal fat, maize flour, rice, vegetable protein isolate, hydrolysed animal protein, fish oil, soya oil, beet pulp and vegetable fibres. The test diet was also spray coated with 0.33 g yeast extract rich in nucleotides (approximately constituted by 10% GMP, 20% AMP, 40% CMP and 30% UMP; PetMOD, Prosol, Italy), 0.3 g XOS (XOS 35, Longlive, China) and 0.45 g scFOS (Profeed, Tereos, France; degree of polymerization between 3 and 5) per 100 g of diet. The concentration of nucleotides, scFOS and XOS included in the diet were chosen by Royal Canin based on literature, previous (unpublished) in vitro and in vivo studies performed by Royal Canin on the individual inclusion of the ingredients, toxicological assessment of the raw materials, and adjustments made to take into account loss in processing of the diet and bioavailability of the compounds (42–44).
The post-production analysis of the experimental diets, carried out at Eurofins Ltd (United Kingdom) and reported in Atwal et al. (45) showed that some differences in nutrient composition between the two diets existed and are likely due to the variability of raw ingredients during the manufacturing process and the analysis performed. The diets were nutritionally complete according to the Association of American Feed Control Officials (AAFCO) (43), and complied with nutritional guidelines produced by The European Pet Food Industry (FEDIAF) (44) and the National Research Council (NRC) (46). Each diet was fed to the kittens at a daily energy intake that was appropriate to their age, in accordance with the guidelines for nutrient requirements for cats (47). Amount of food (g) offered, eaten, and refused were also recorded, as reported in our previous study (45).
2.4 Blood collection and plasma preparation and storage
All the cats were fasted for at least three and a half hours before blood collection. Lidocaine-prilocaine analgesic cream was applied 45 min before 2 ml blood was drawn from the jugular vein and decanted into K3-EDTA tubes. Within 1 h of sampling, blood samples were centrifuged at 2000 x g at room temperature for 10 min, plasma collected from the upper phase, and further centrifuged at 3000 x g at 4°C for 15 min, to remove additional cellular debris and to minimise the contamination of cell-free nucleic acids by genomic DNA and RNA derived from damaged blood cells. The cleared supernatants were transferred to fresh RNase-free tubes and frozen at −80°C until required.
2.5 miRNA-Seq analysis
Plasma samples from all 23-week-old kittens were sent to QIAGEN (Germany) for next-generation sequencing (NGS) of miRNAs. RNA was isolated using the miRNeasy Serum/Plasma Kit (Cat. No. 217184, QIAGEN), according to the manufacturer's instructions. Prior to sequencing, the preparation of the libraries was performed using the QIAseq miRNA Library Kit (QIAGEN) and total RNA was converted into miRNA next-generation sequencing (NGS) libraries. Amplification of libraries was performed, and quality controlled using capillary electrophoresis (Agilent DNA 1000 Chip). Based on the quality of the inserts and their concentrations, the libraries were pooled in equimolar ratios. The library pools were then sequenced on a NextSeq instrument (Illumina®) to produce 12 million reads of 1 × 75 base pair fragments per sample. Raw data was then de-multiplexed using the bcl2fastq software (Illumina®), which also created FASTQ files for each sample. The expression of miRNAs was quantified using the Quantify miRNA tool in Genomics Workbench (QIAGEN) and normalised to counts per million reads mapped (CPM). The variance of log2CPM in expression levels was then used to conduct a principal component analysis (PCA) in QIAGEN CLC Genomics Workbench (Supplementary Figure 1) to identify the relationship between the top 300 plasma miRNAs with the highest variance across all the samples and diets offered to the kittens in this study.
2.6 Isolation of RNA from plasma and quantification
Synthetic cel-miR-2-3p and cel-miR-39-3p molecules (Applied Biosystems), used as exogenous RNA references (spike-ins) to monitor the efficiency of the RNA isolation method and the reverse transcription polymerase chain (RT) reaction, respectively, were reconstituted at 25 μM using RNase/DNase-free (no diethyl pyrocarbonate (DEPC)-treated) water and stored at −20°C until needed. To isolate RNA, plasma samples were defrosted on ice, the synthetic (spike-in) cel-miR-2-3p was diluted 1:10,000,000 and added to plasma samples, and RNA was isolated using the miRNeasy Serum/Plasma Advanced Kit (QIAGEN) and quantified using a Bioanalyzer 2100 instrument and Agilent Small RNA kit (Agilent), following manufacturer's instructions.
2.7 Synthesis of miRNA-specific cDNAs and qPCR analysis
The entire amount of RNA isolated was used for the synthesis of cDNA molecules, this was performed immediately after the isolation of RNA from plasma using TaqMan™ MicroRNA Reverse Transcription Kit and 5X TaqMan™ miRNA assays (Thermo Fisher Scientific; Supplementary Table 1). The spike-in cel-miR-39-3p (25 μM) was diluted 1:10,000,000. The synthesis of cDNA (RT reaction) was specifically performed for each of the miRNAs determined in this study, thus 10 RT reactions were performed for each RNA sample. Each RT reaction (total volume of 15 μl) was comprised of 0.15 μl of 100 mM deoxyribonucleotide triphosphate (dNTPs), 1 μl of MultiScribe RT enzyme (50 U/μl), 1.5 μl of 10X RT buffer, 0.19 μl of RNase inhibitor (20 U/μl), 3 μl of 5X RT specific primers, 5 μl of 1:5 diluted (in RNase free water) RNA, 1 μl of diluted cel-miR-39-3p and 3.16 μl of RNase-free water. RT reactions were incubated at 16°C for 30 min, then at 42°C for 30 min, and finally at 85°C for 5 min. The synthesised miRNA-specific cDNAs were immediately amplified by quantitative polymerase chain reaction (qPCR) assay.
2.8 qPCR
The qPCR reactions were specifically performed for each of the miRNAs determined in this study (Supplementary Table 1), thus one qPCR reaction was performed for each miRNA-specific cDNA sample, using the TaqMan™ Universal polymerase chain reaction (PCR) MasterMix and 20X TaqMan™ miRNA Assays (Thermo Fisher Scientific). Each qPCR reaction (total volume of 10 μl) comprised 0.5 μl of 20X TaqMan™ specific primers, 5 μl of TaqMan™ universal PCR master mix, 2 μl of miRNA-specific cDNA, and 2.5 μl of RNase free water. The qPCR reactions were incubated, using an ABI QuantStudio 7 Flex Real-Time PCR System, at 50°C for 2 min, then at 95°C for 10 min, followed by 40 cycles during which reactions were incubated at 95°C for 15 s and then at 60°C for 1 min. Amplification plots were generated using QuantStudio™ Real-Time PCR Software, and cycle threshold (Ct) values were determined for all the endogenous and exogenous (spike-ins) miRNAs analysed in this study. Let-7c-5p, miR-99a-5p, miR-146b-5p and miR-140-3p, whose expression did not significantly change between dietary groups based on results generated by the miRNA-sequencing analysis, were selected as candidate normaliser miRNAs. NormFinder algorithm was used to determine stability values and the 2−ΔΔCt method (48) to assess the relative concentration of miRNAs in feline plasma. The number of independent experiments, each of which was performed on a different day, was also reported.
2.9 Statistical analysis
The data mean ± standard error of the mean (SEM) and statistical analysis were calculated using GraphPad Prism 8 software. Two sets of hypothesis tests were performed, the difference between diet groups at each of the time points and the change from earliest timepoint to each subsequent time point within each diet group. The analysis to compare the means of expression of each plasma miRNA in 23-, 37- and 45-week-old kittens, between test and control groups, was completed performing an unpaired t-test. While a one-way ANOVA and Dunnett's multiple comparisons test were performed to compare the expression levels of miRNAs in the control group over time, a mixed-effects analysis and Dunnett's multiple comparisons test were employed to compare the expression levels of miRNAs in the test group (at 23 weeks of age) compared to T-C groups (at 37 and 45 weeks). P-values ≤ 0.05 were considered significant.
2.10 Bioinformatics analysis
The miRTarBase database (https://mirtarbase.cuhk.edu.cn/) (49) was used to identify experimentally validated targets of the miRNAs of interest, while the DAVID platform (https://david.ncifcrf.gov/tools.jsp) was used to conduct gene ontology (GO) enrichment and Kyoto Encyclopaedia of Genes and Genomes (KEGG) pathway analyses to determine the biological processes, cellular components and molecular functions potentially impacted by the miRNAs analysed in this study. Furthermore, the interactions between miR-1-3p, miR-133a-3p, miR-206-3p and miR-383-5p were investigated by creating a network map using the miRNet platform (https://www.mirnet.ca/) (50).
3 Results
3.1 Identification of differentially expressed miRNAs in kitten plasma via miRNA-sequencing
A PCA analysis, did not indicate a clear relationship between plasma miRNAs and diets fed in 23-week-old kittens, although kittens fed with the test diet clustered more than those in the control group. The miRNA sequencing data was further analysed to identify miRNAs with significant differential expression. The relationship between the log10 transformed P-values and the log2 transformed fold change expression values for the test diet compared to the control group are illustrated by volcano plot (Figure 2).
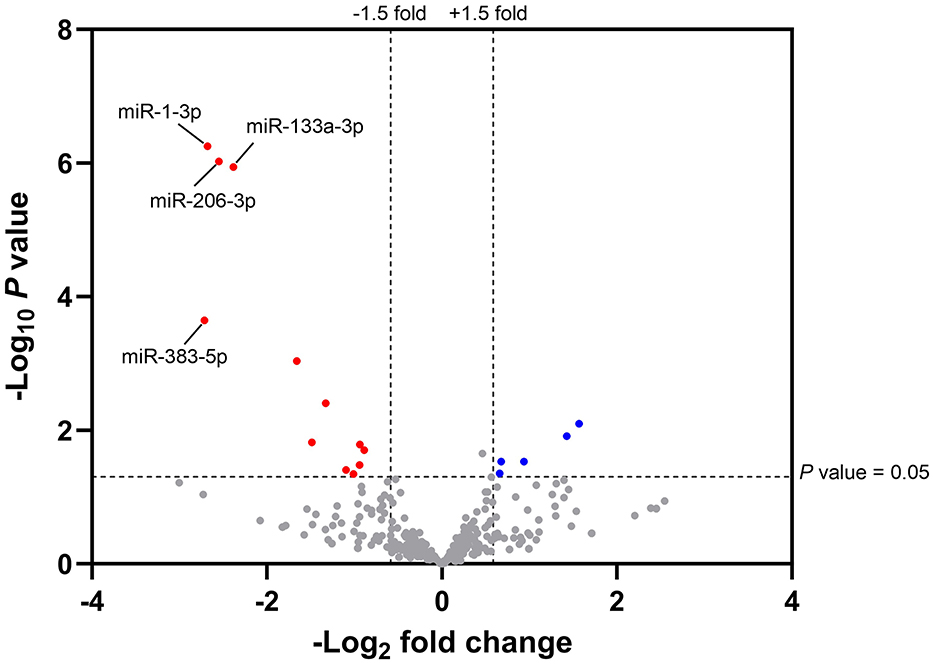
Figure 2. Volcano plot illustrating the magnitude fold change and statistical differences of plasma miRNAs from 23-week-old kittens fed with the test diet compared to control. The magnitude fold change (–Log2) is plotted on the x-axis and the significance (t-test; -Log10P value) is plotted on the y-axis. Fold change > 1.5 and P-value < 0.05 were set as the thresholds for identifying significantly differentially expressed plasma miRNAs from 23-week-old kittens fed with the test diet (n = 7) compared to the control group (n = 9). The red and blue hits represent the downregulated ( ) and upregulated (
) and upregulated ( ) miRNAs in the test group, respectively; the grey hits represent miRNAs that did not meet the threshold criteria (
) miRNAs in the test group, respectively; the grey hits represent miRNAs that did not meet the threshold criteria ( ). miRNAs ID (n = 4) with false discovery rate (FDR) adjusted P-value < 0.05 were indicated.
). miRNAs ID (n = 4) with false discovery rate (FDR) adjusted P-value < 0.05 were indicated.
The total number of miRNAs identified in feline plasma was 339. A 1.5-fold change, considered to be sufficient for miRNAs to elicit a biological response, and P < 0.05 were set as threshold criteria. In total, 17 miRNAs, 12 of which were downregulated and five upregulated compared to the control group (Supplementary Table 2), met these criteria. To exclude false positive results, P-values were adjusted using the Benjamini-Hochberg method (50) and the false discovery rate (FDR) adjusted P-values evaluated. This correction identified miR-383-5p, miR-1-3p, miR-133a-3p and miR-206-3p as significantly differentially regulated between dietary groups. These miRNAs, from now referred to as “miRNAs of interest,” had lower expression levels in the plasma of kittens fed with the test diet compared to the control group and were validated by RT-qPCR.
3.2 Validation of miRNAs of interest
RT-qPCR analysis of the miRNAs of interest was conducted to ensure that these miRNAs were not false positive results of the miRNA-sequencing analysis. Validated feline-specific TaqMan™ miRNA assays were not commercially available, therefore TaqMan™ miRNA assays validated in other species that could be used to amplify feline miRNAs were used. Alignment of the sequences of the feline miRNAs of interest identified in this study to those reported in miRBase for other mammalian species showed 100% homology in the nucleotide sequences for miR-206-3p, and 95%, 91% and 86% homology for miR-133a-3p, miR-1-3p and miR-383-5p, respectively, with a 100% homology in all seed sequences (Figure 3).
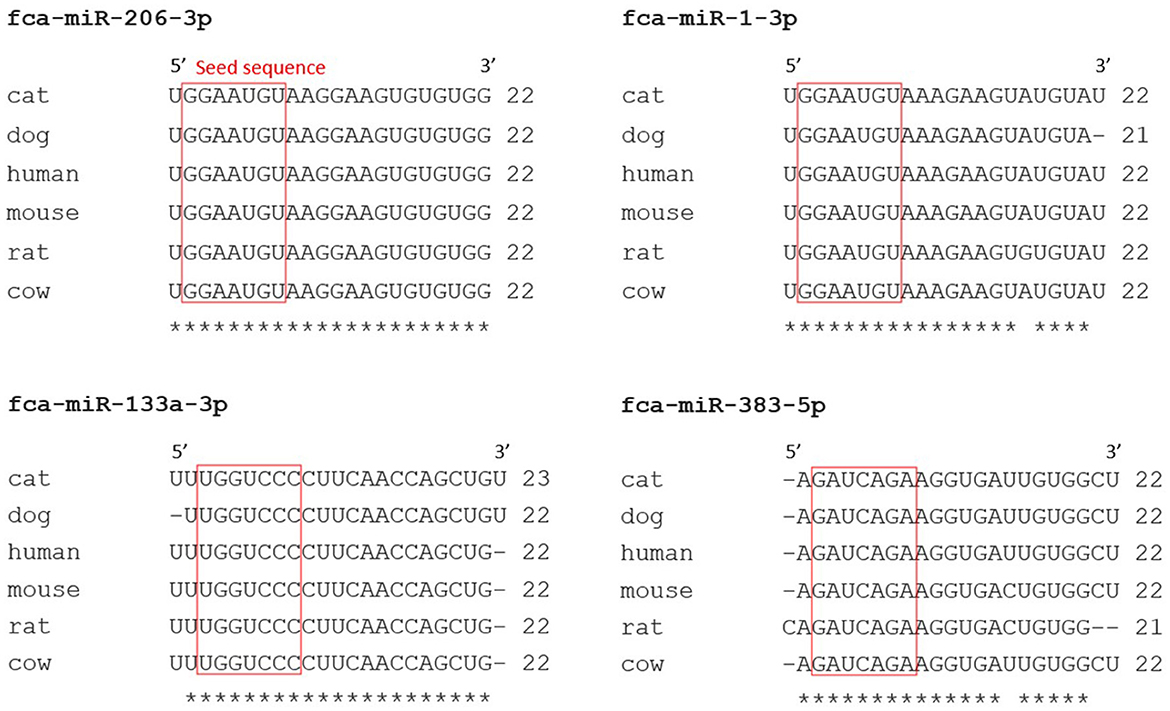
Figure 3. Nucleotide sequences of feline miRNAs with false discovery rate (FDR) <0.05 were well conserved across mammalian species. Nucleotides sequences of feline (felis catus; fca) miRNAs of interest obtained from miRNA-sequencing analysis, with FDR <0.05, were aligned (Clustal Omega) with sequences of miRNAs from other species (miRBase). A red box was drawn around miRNA seed regions and similarities between nucleotides across different species were marked with a star (*).
To validate the expression of each individual miRNA of interest, miRNA isolation and RT-qPCR experiments were performed on separate plasma aliquots collected from the same kittens analysed by miRNA-sequencing analysis. However, due to limited sample volume, this was only performed in 8 (n = 4 for each dietary group) 23-week-old kittens. Gel electrophoresis (Bioanalyzer) confirmed that the RNA isolation from plasma samples retained miRNAs, indicated by the presence of bands in the plasma sample that were similar in size to that of cel-miR-39-3p (22 nucleotides; not shown).
RT-qPCR analysis was first performed to identify the most stably expressed miRNA amongst candidate endogenous (let-7c-5p, miR-99a-5p, miR-146b-5p and miR-140-3p) and exogenous (cel-miR-2-3p and cel-miR-39-3p) miRNAs to be used as reference genes for normalisation. As a result, miRNA-146b-5p was selected as normaliser, because it showed a low variability (CV = 1.06%) across all candidate normalisers, did not cluster between diets (Supplementary Figure 2) and had a low stability value (0.005). The relative expression of miR-383-5p, miR-1-3p, miR-133a-3p and miR-206-3p in plasma from 23-week-old kittens (n = 4 per diet group) was lower in kittens fed with the test diet compared to the control group, and while their expression was more variable in the control group, the opposite was observed in kittens fed with the test diet (Figure 4).
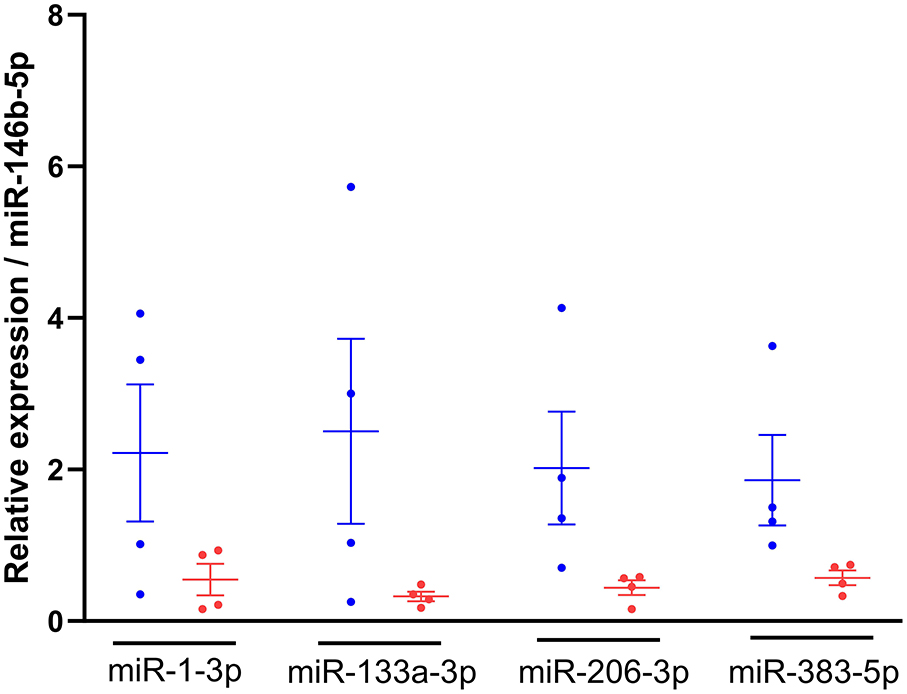
Figure 4. Relative expression of miRNAs of interest was lower in kittens fed the test diet. RNA was isolated from plasma of 23-week-old kittens fed with the control  (n = 4) or test
(n = 4) or test  (n = 4) diet, miRNA-specific cDNAs were synthesised and TaqMan qPCR performed. The 2−ΔΔCt method was used to quantify the relative expression of miR-1-3p, miR-133a-3p, miR-206-3p and miR-383-5p normalised to miR-146b-5p. Data shown as mean ± SEM (n = 6 independent experiments). Unpaired t-test control vs test diet for each miRNA.
(n = 4) diet, miRNA-specific cDNAs were synthesised and TaqMan qPCR performed. The 2−ΔΔCt method was used to quantify the relative expression of miR-1-3p, miR-133a-3p, miR-206-3p and miR-383-5p normalised to miR-146b-5p. Data shown as mean ± SEM (n = 6 independent experiments). Unpaired t-test control vs test diet for each miRNA.
3.3 miRNA target prediction
The targets of feline miRNAs were not present in miRTarBase, however, with the miRNA sequences being highly conserved between cat and human (>86%; Figure 3), the analysis was conducted using the human miRNAs. Validated miRNA-target interactions of human (hsa)-miR-1-3p (miRBase entry MIMAT0000416), hsa-miR-133a-3p (MIMAT0000427), hsa-miR-206-3p (MIMAT0000462) and hsa-miR-383-5p (MIMAT0000738) were identified (Supplementary Table 3). This indicated that a total of 127 genes could be targeted by the miRNAs of interest. Specifically, miR-1-3p, miR-133a-3p, miR-206-3p and miR-383-5p target 78, 39, 31 and 7 genes, respectively, with 28 genes targeted by more than one miRNA.
GO enrichment and KEGG pathway analyses, for all 127 validated target genes, identified 352 biological processes, 61 cellular components, and 72 molecular functions, as potentially impacted by the miRNAs of interest. Statistically significant differences were also identified. The top-20 statistically significant biological processes with the lowest P-values, together with the number of genes involved in each specific process are reported in Figure 5. The GO analysis showed that the genes targeted by the miRNAs of interest have a role in several biological functions such as positive regulation of transcription and cell proliferation, negative regulation of apoptosis, development and function of cardiac muscle cells (Figure 5A), and molecular processes, including protein and DNA binding (Supplementary Figure 3). The KEGG analysis revealed that the target genes were involved in 114 pathways, and the top-20 statistically significant pathways are reported in Figure 5B and Supplementary Table 4. Additionally, the analysis revealed that the miRNAs of interest could affect different pathways involved in cancer and other diseases, with targeted genes involved in immune mechanisms such as the PI3K-AKT (20 genes; 4.9-fold enrichment) and JAK-STAT (11 genes; 5.9-fold enrichment) pathways.
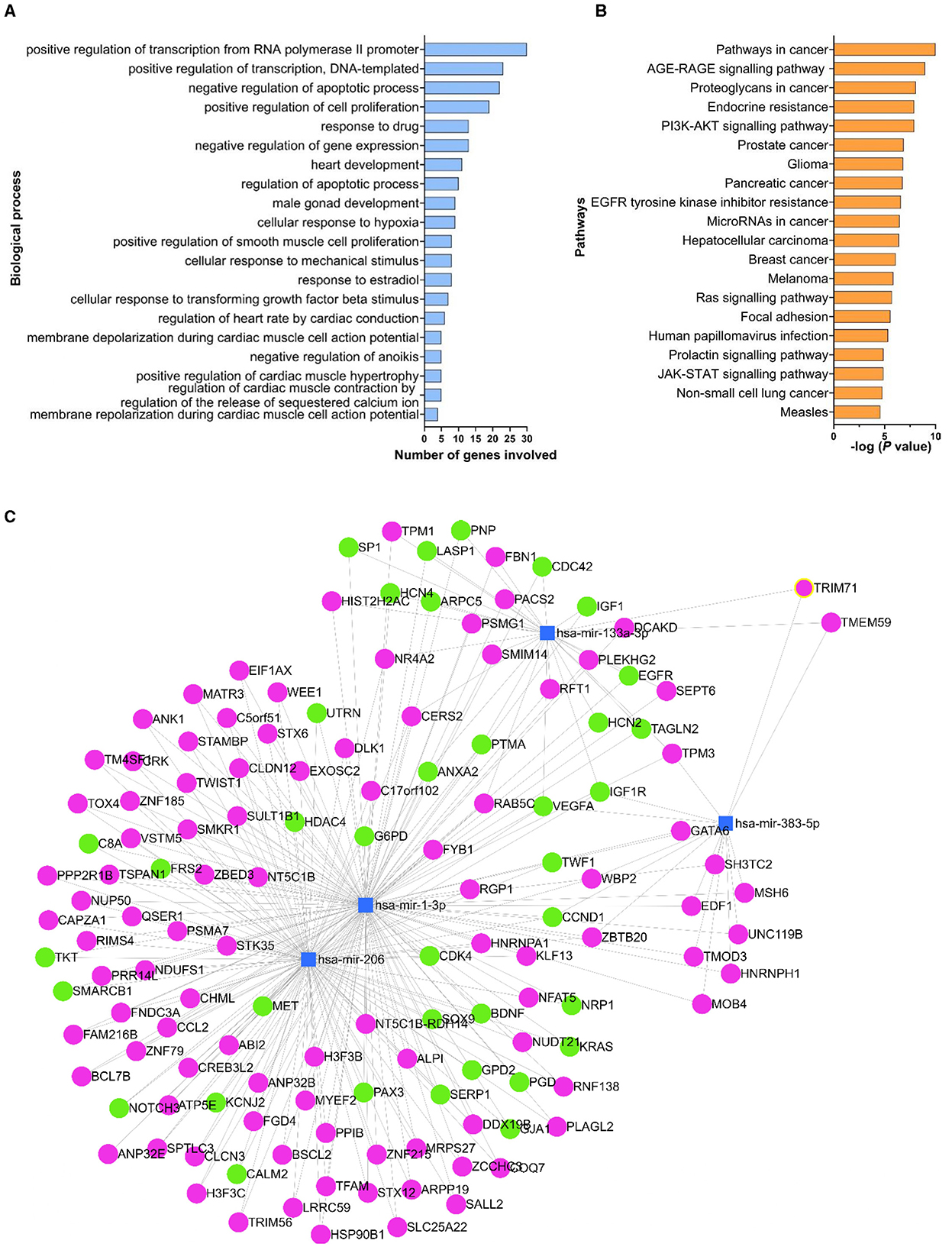
Figure 5. Biological processes, pathways and genes targeted by the miRNAs of interest. The list of experimentally validated genes (identified in miRTarBase) targeted by the miRNAs of interest was used in the DAVID database to determine the biological processes (A) and the pathways (B) affected by the miRNAs of interest. Only the top-20 statistically significant processes with the lowest P-values were reported in (A) and (B). Human (hsa)-miR-1-3p, hsa-miR-133a-3p, hsa-miR-206-3p and hsa-miR-383-5p were used to create a network map of the miRNAs of interest (blue squares) using miRNet platform (C). The target genes reported on miRTarBase as being experimentally validated by reporter assay, western blot and qPCR assay were shown in green circles; all the other genes were reported in magenta.
The network map analysis, conducted using human as reference, indicated that the miRNAs of interest target 1,138 genes in total. Of the 136 genes that were shared between at least two miRNAs of interest, 37 genes were reported on miRTarBase as being experimentally validated by reporter assay, western blot analysis and qPCR assay (Figure 5C). The highest number of common target genes was between miR-1-3p and miR-206-3p, with 96 genes in common, while only one common gene (VEGFA) was targeted by all (n = 4) the miRNAs of interest.
3.4 Long-term modulation of circulating miRNAs post-supplementation of the test diet
Following quantification of the expression of the miRNAs in plasma from 23-week-old kittens fed with the control or test diet, we also investigated the expression of plasma miRNAs 2 and 10 weeks post-supplementation. Due to limited sample volume, this investigation was conducted on 8 37- and 45-week-old non-matched kittens (n = 5 control group; n = 3 test group). The expression levels of miRNAs of interest in 37- and 45-week-old kittens were quantified (Figure 6) and compared to miRNA expression data from 23-week-old (non-matched) kittens.
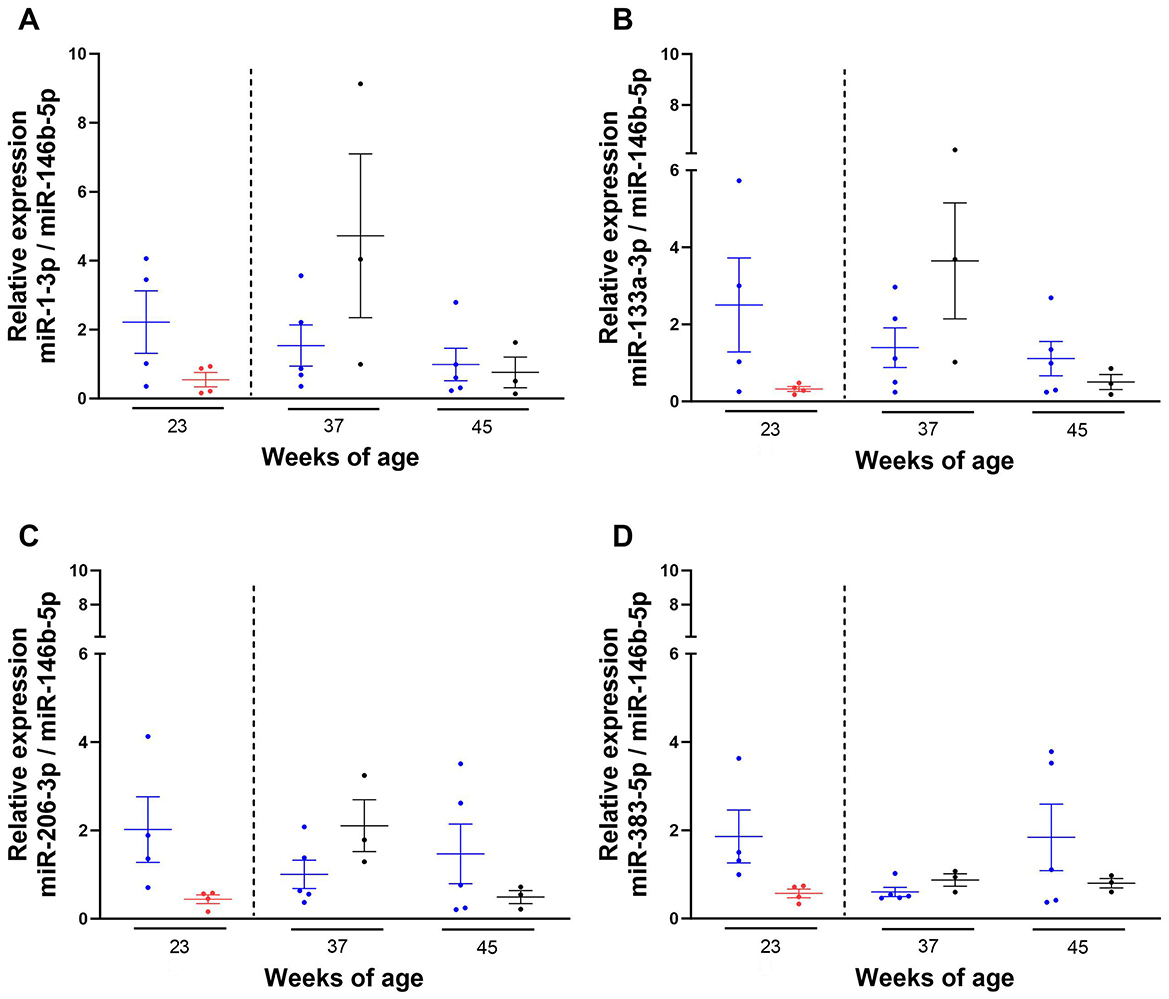
Figure 6. Expression levels of miRNAs of interest 2 and 10 weeks after the withdrawal of the test diet. Kittens 23- (n = 4), 37- (n = 5) and 45- (n = 5) week-old were maintained on the control diet (C-C) or fed with the test diet until 23-week-old (n = 4), and then moved at 35 weeks (dotted line) onto the control diet  and (blood) sampled at 37 (n = 3) and 45 (n = 3) weeks of age (T-C). RNA was isolated from plasma of kittens, miRNA-specific cDNAs were synthesised and TaqMan qPCR performed. Ct values obtained for each miRNA were normalised to the averaged Ct value of miR-146b-5p and the 2−ΔΔCt method was used to assess the relative expression of miR-1-3p (A), miR-133a-3p (B), miR-206-3p (C) and miR-383-5p (D). Final data shown as mean ± SEM (n = 6 independent experiments). Unpaired t-test between diets at each week of age. One-way ANOVA and Dunnett's multiple comparisons test vs. 23 weeks of age between control and C-C kittens. Mixed-effects analysis and Dunnett's multiple comparisons test vs. 23 weeks of age between test and T-C groups.
and (blood) sampled at 37 (n = 3) and 45 (n = 3) weeks of age (T-C). RNA was isolated from plasma of kittens, miRNA-specific cDNAs were synthesised and TaqMan qPCR performed. Ct values obtained for each miRNA were normalised to the averaged Ct value of miR-146b-5p and the 2−ΔΔCt method was used to assess the relative expression of miR-1-3p (A), miR-133a-3p (B), miR-206-3p (C) and miR-383-5p (D). Final data shown as mean ± SEM (n = 6 independent experiments). Unpaired t-test between diets at each week of age. One-way ANOVA and Dunnett's multiple comparisons test vs. 23 weeks of age between control and C-C kittens. Mixed-effects analysis and Dunnett's multiple comparisons test vs. 23 weeks of age between test and T-C groups.
While the expression levels of the miRNAs of interest in 23-week-old kittens were lower in animals fed with the test diet compared to the control group, the opposite was observed at 37 weeks of age, where the expression levels of miR-383-5p, miR-1-3p, miR-133a-3p and miR-206-3p were higher in kittens previously fed with the test diet (T-C kittens) compared to those maintained on the control diet (C-C kittens), with higher variability observed for miR-1-3p and miR-133a-3p (Figure 6).
Conversely, the expression levels of the miRNAs of interest in T-C kittens were lower than the C-C group at 45 weeks of age. None of these changes were statistically significant (P-values > 0.5).
4. Discussion
In the last decade, nutrition research has revealed the ability of dietary nucleotides and oligosaccharides to support the differentiation and function of immune cells, and the potential for nutrition to affect the expression of endogenous miRNAs in mammals. Therefore, there is a potential for miRNAs to be used as biomarkers to assess the effects of nutrition on immunity, becoming a health monitoring tool to ensure an adequate nutritional support to kittens, during their development. Previously, we found that a diet fortified with nucleotides, scFOS and XOS significantly increased antibody levels post immunisation in kittens, potentially due to an increased polarisation of T-helper cells, activators of antibody-producing B-cells (45). Here, we wanted to determine whether the same diet was also able to affect the expression level of circulating immune-related miRNAs, and if any effects were maintained post-supplementation.
Data generated through the miRNA-sequencing analysis showed a significant difference in miRNA expression between experimental diets, RT-qPCR data was in agreement with this revealing a lower expression of circulating miR-383-5p, miR-1-3p, miR-133a-3p and miR-206-3p in 23-week-old kittens fed with the test diet, compared to control, however this effect was not significant. This may be attributed to differences in sample size between the methods. Indeed, the retrospective powering analysis (not shown) performed by simulation using RT-qPCR data generated in this study, confirmed that 12 kittens were required for 80% power for an effect size of 70%−90%. Unfortunately, we could not recruit additional animals, as this was an opportunistic investigation utilising kittens on an existing study. RT-qPCR data also indicated that the expression levels of the miRNAs of interest were highly variable in kittens fed with the control diet compared to the test group. Although this variability was confirmed by the PCA analysis, further investigation should be conducted as its reason is unclear.
In accordance with previous studies reporting low amounts of cell-free RNA molecules in human plasma, the RNA yield was low in kitten plasma (51, 52). For this reason, the entire amount of unquantified RNA isolated in this study was used for the synthesis of miRNA-specific cDNAs for all the kittens, hence the input total RNA could not be standardised across samples. Nevertheless, Huggett et al. (53) demonstrated that reference genes such as the endogenous miRNA (miR-146b-5p) used in this study can control for different input RNA amounts used in the reverse transcription step.
In animals, miRNAs can regulate more than one gene (54) therefore, miRNAs that usually function in a specific tissue can also regulate the expression of genes in different body compartments. For example, miR-1-3p, miR-133a-3p and miR-206-3p are part of the “myomiRs” family and affect genes involved in skeletal and cardiac muscle development in mammals (55); an association that was confirmed by the GO and KEGG analyses we reported in this study. In addition, those miRNAs can also regulate Fas apoptosis inhibitory molecule (FAIM), an anti-apoptotic protein found in most cell types, including immune cells (56), and affect genes in murine mammary tissue (57) which have shown to affect immunity by regulating NF-κB-mediated inflammatory responses (58), NK cell-mediated killing (59), and the formation of NLRP6 inflammasomes as well as the secretion of IL-18 (60) in humans.
Literature providing evidence of the ability of nutrition to affect the expression of miRNAs in developing kittens has been lacking, potentially due to the low number of miRNA studies conducted in the cat. However, we have revealed for the first time the ability of dietary nucleotides and oligosaccharides to affect potentially immune-related miRNAs in kittens. Indeed, the GO and KEGG analyses we conducted here, indicated that the miRNAs of interest positively regulate cell proliferation and play a role in the PI3K-AKT pathway, potentially supporting the development of immune cells and increasing antibody titres following immunisation. Results suggest that in 23-week-old kittens fed with the test diet, in which a lower expression level of miR-206-3p was measured compared to the control group, the PI3K-AKT pathway was not inhibited. Our hypothesis is supported by previous results generated in a murine preadipocyte cell line, where the expression of miR-206 inhibited c-Met, an important upstream regulator of the PI3K-AKT pathway (61) that is encoded by the MET gene (62), identified here as a validated target of miR-206-3p. The low expression level of miR-206-3p measured in kittens in the test group may have led to sustained PI3K-AKT signalling, which, in turn, may have played a role in supporting the development of T cells and differentiation of T helper lymphocytes in the thymus of kittens. In fact, while the inhibition of the PI3K-AKT pathway in mice led to a reduced number of peripheral T cells (63), its activation was able to induce the differentiation of T cells (64). Moreover, our hypothesis that low miR-206-3p expression may have contributed to the development of T helper lymphocytes in 23-week-old kittens fed with the test diet agrees with data reported in our previous study, where the number of T helper cells observed in kittens fed with the same test diet was higher than that in kittens fed with the control diet, and possibly increased the seroconversion of kittens following vaccination (45).
Here, we determined a lower expression level of miR-1-3p and miR-133a-3p in the test group compared to control. Although there is no evidence showing that these miRNAs function in antibody production post-immunisation and development of T helper lymphocytes, they have been previously studied in the context of immune function. Specifically, research conducted using a murine macrophage cell line (RAW264.7) revealed that miR-1-3p suppressed the phagocytosis of E. coli by inhibiting the expression of clathrin heavy chain 1 (CLTC1) gene (65), while miR-133a-3p was able to regulate inflammasome activation through uncoupling protein-2 (UCP2) in a human monocytic cell line (THP1) (66). While UCP2 gene was identified by us as a target gene of miR-133a-3p, our analysis did not report CLTC1 as a validated target of miR-1-3p. Therefore, further investigation is needed to determine and confirm the validated targets of miR-1-3p and miR-133a-3p in the cat and their potential to affect immune functions and pathways.
The expression level of miR-383-5p in plasma of kittens fed with the test diet was also lower compared to the control group. Currently, there is no evidence showing the ability of in vivo dietary supplementations to affect the expression level of miR-383-5p, which has mainly been investigated in the context of cancer and identified as a tumour suppressor able to inhibit cell proliferation and cancer progression in several types of cancer (67). Although further investigation is needed to determine the ability of miR-383-5p to influence immune functions, a study conducted by Zhang et al., in the context of the human central nervous system (68) showed that one of the validated targets of miR-383-5p, glycoprotein 130 (gp130), which is the subunit for the receptor of IL-6 family cytokines, induces the activation of the JAK-STAT pathway (69), and its expression is not limited to the nervous system but also found in immune cells (70, 71).
The present study is the first investigating the sustained effects of a diet after its withdrawal on the expression of endogenous miRNAs in kittens. We found that, due to a potential adaptation to the new diet 2 weeks post-supplementation, the expression levels of miR-383-5p, miR-1-3p, miR-133a-3p and miR-206-3p in T-C kittens trended towards being higher than those measured in animals maintained on the control diet (C-C). On the other hand, 10 weeks post-supplementation, the expression levels of these miRNAs were comparable to their levels before the withdrawal of the test diet. One hypothesis is that the effects induced by the test diet on the expression of endogenous miRNAs persisted 10 weeks post-supplementation. Interestingly, on a similar note, a study conducted in piglets showed that the expression levels of miR-708-5p, miR-135-3p and miR-27b-3p were lower in 21-, 35- and 51-day-old piglets previously fed (from 2 to 21 days of age) with a dairy-based formula, compared to animals fed with human breastmilk (72).
The data from this study raise the question whether miR-383-5p, miR-1-3p, miR-133a-3p and miR-206-3p can affect immune functions in kittens. To answer this question, it is imperative to first validate the gene targets of the feline miRNAs of interest that we have identified. This could be achieved through bioinformatics analysis using human miRNAs as references, based on the fact that 90% of the genes in the cat are similar to humans (73). Further future studies incorporating each ingredient individually into the diet, rather than in a cocktail as in the present study, should also be conducted to determine whether the differences observed in expression of miRNAs are driven by single ingredients or their combination.
In summary, our study has found that kittens fed with a diet containing nucleotides, scFOS and XOS express lower levels of circulatory plasma miR-383-5p, miR-1-3p, miR-133a-3p and miR-206-3p compared to the control group; an expression profile that was maintained 10 weeks post-supplementation. The literature and our pathway analysis indicate that these miRNAs regulate the PI3K-AKT and JAK-STAT pathways, suggesting that their modulation may affect the animal immune response, although this was not confirmed.
Finally, in light of our previous work (45), the expression level of the circulating plasma miRNAs identified here may support the development of T cells and increase the seroconversion of kittens following vaccination. However, this cannot be confirmed. Ultimately, our findings identified nutrients able to affect potential immune-related miRNAs, the expression of which could be used to monitor feline immune performance in the future and support veterinary care to improve the healthspan of companion animals.
Data availability statement
The original contributions presented in the study are publicly available. This data can be found here: NCBI repository, PRJNA1334322.
Ethics statement
The animal study was approved by Waltham Animal Welfare and Ethical Review Board Waltham Petcare Science Institute. The study was conducted in accordance with the local legislation and institutional requirements.
Author contributions
FA: Conceptualization, Data curation, Formal analysis, Investigation, Methodology, Project administration, Software, Validation, Visualization, Writing – original draft, Writing – review & editing. MH: Conceptualization, Supervision, Visualization, Writing – review & editing. MF: Supervision, Visualization, Writing – review & editing. LH: Supervision, Writing – review & editing. JA: Writing – review & editing. PW: Writing – review & editing. AP: Supervision, Visualization, Writing – review & editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This work was funded by Mars Petcare, a manufacturer of pet food and provider of veterinary services. The Waltham Petcare Science Institute is a fundamental research centre for Mars Petcare.
Acknowledgments
The authors acknowledge the expertise (e.g. caring, training, sampling of kittens) of the WALTHAM personnel, essentials for the realisation of this project. We also acknowledge Robyn Bednall and Kimberley Slinger for the coordination of this study.
Conflict of interest
FA, MH, MF, LH, JA, and PW were employees of Mars Petcare at the time this manuscript was submitted.
The remaining author declares that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The authors declare that this study received funding from Mars Petcare. The funder had the following involvement in the study: study design, collection, analysis, interpretation of data, the writing of this article, and the decision to submit it for publication.
Publisher's note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fvets.2024.1382436/full#supplementary-material
Supplementary Figure 1 | Principal component analysis (PCA) does not show relationships between plasma miRNAs and diets offered to 23-week-old kittens. Data from plasma of kittens fed with test (n = 7) or control (n = 9) diet. Expression data was normalised to the counts per million reads mapped (CPM) and variance of the log2CPM in expression levels was calculated across all samples. The top 300 miRNAs with the highest variance across all samples were used in the principal component calculation and as input for the PCA plot.
Supplementary Figure 2 | MiR-146b was the most suitable miRNA to be used as reference gene in RT-qPCR experiments. Cycle threshold (Ct) values determined by two-step RT-qPCR for individual miRNAs were averaged across 23-week-old kittens fed with the control [[Inline Image]] (n = 4) and test [[Inline Image]] (n = 4) diets. The variation of individual Ct values from the averaged Ct were calculated together with the coefficient of variation (%CV). Data shown as means ± SEM (n = 6 independent experiments).
Supplementary Figure 3 | Molecular processes targeted by the miRNAs of interest. The list of experimentally validated genes (identified in miRTarBase) targeted by the miRNAs of interest was used in the DAVID database to determine the molecular processes affected by the miRNAs of interest. Only the top-20 statistically significant processes with the lowest P-values were reported.
Supplementary Table 1 | Target sequences of TaqMan™ miRNA Assays used in qPCR experiments.
Supplementary Table 2 | List of miRNAs that met the set threshold criteria of 1.5-fold change and P < 0.05 compared to the control group.
Supplementary Table 3 | Experimentally validated miRNA-target of miRNAs of interest reported in miRTarBase database.
Supplementary Table 4 | The miRNAs of interest target genes involved in immune signalling pathways.
References
1. Evermann JF, Wills TB. Immunologic development and immunization. Small Anim Pediatr. (2011) 14:104–12. doi: 10.1016/B978-1-4160-4889-3.00014-0
2. O'Brien T, Thomas DG, Morel PCH, Rutherfurd-Markwick KJ. Moderate dietary supplementation with vitamin E enhances lymphocyte functionality in the adult cat. Res Vet Sci. (2015) 99:63–9. doi: 10.1016/j.rvsc.2015.01.007
3. Kearns RJ, Hayek MG, Turek JJ, Meydani M, Burr JR, Greene RJ, et al. Effect of age, breed and dietary omega-6 (n-6): omega-3 (n-3) fatty acid ratio on immune function, eicosanoid production, and lipid peroxidation in young and aged dogs. Vet Immunol Immunopathol. (1999) 69:165–83. doi: 10.1016/S0165-2427(99)00052-5
4. Chodur GM, Steinberg FM. Human microRNAs modulated by diet: a scoping review. Adv Nutr. (2024) 15:100241. doi: 10.1016/j.advnut.2024.100241
5. O'Brien J, Hayder H, Zayed Y, Peng C. Overview of microRNA biogenesis, mechanisms of actions, and circulation. Front Endocrinol. (2018) 9:1–12. doi: 10.3389/fendo.2018.00402
6. Rodriguez-Galan A, Fernandez-Messina L, Sanchez-Madrid F. Control of immunoregulatory molecules by miRNAs in T cell activation. Front Immunol. (2018) 9:2148–61. doi: 10.3389/fimmu.2018.02148
7. Xu SJ, Chen JH, Chang S, Li HL. The role of miRNAs in T helper cell development, activation, fate decisions and tumor immunity. Front Immunol. (2023) 14:1320305. doi: 10.3389/fimmu.2023.1320305
8. Gaál Z. Role of microRNAs in immune regulation with translational and clinical applications. Int J Mol Sci. (2024) 25:1942. doi: 10.3390/ijms25031942
9. Biswas S, Haleyurgirisetty M, Lee S, Hewlett I, Devadas K. Development and validation of plasma miRNA biomarker signature panel for the detection of early HIV-1 infection. EBioMedicine. (2019) 43:307–16. doi: 10.1016/j.ebiom.2019.04.023
10. Oshiumi H. Circulating extracellular vesicles carry immune regulatory miRNAs and regulate vaccine efficacy and local inflammatory response after vaccination. Front. Immunol. (2019) 12:685344. doi: 10.3389/fimmu.2021.685344
11. Yu Y, Zhang J, Wang J, Sun B. MicroRNAs: the novel mediators for nutrient-modulating biological functions. Trends Food Sci Technol. (2021) 114:167–75. doi: 10.1016/j.tifs.2021.05.028
12. Tili E, Michaille JJ, Adair B, Alder H, Limagne E, Taccioli C, et al. Resveratrol decreases the levels of miR-155 by upregulating miR-663, a microRNA targeting JunB and JunD. Carcinogenesis. (2010) 31:1561–6. doi: 10.1093/carcin/bgq143
13. Vinciguerra M, Sgroi A, Veyrat-Durebex C, Rubbia-Brandt L, Buhler LH, Foti M. Unsaturated fatty acids inhibit the expression of tumor suppressor phosphatase and tensin homolog (PTEN) via microRNA-21 up-regulation in hepatocytes. Hepatology. (2009) 49:1176–84. doi: 10.1002/hep.22737
14. Rodriguez A, Vigorito E, Clare S, Warren M V, Couttet P, Soond DR, et al. Requirement of bic/microRNA-155 for normal immune function. Science. (2007) 316:608–11. doi: 10.1126/science.1139253
15. Thai TH, Calado DP, Casola S, Ansel KM, Xiao C, Xue Y, et al. Regulation of the germinal center response by microRNA-155. Science. (2007) 316:604–8. doi: 10.1126/science.1141229
16. Muljo SA, Ansel KM, Kanellopoulou C, Livingston DM, Rao A, Rajewsky K. Aberrant T cell differentiation in the absence of Dicer. J Exp Med. (2005) 202:261–9. doi: 10.1084/jem.20050678
17. Cobb BS, Hertweck A, Smith J, O'Connor E, Graf D, Cook T, et al. A role for Dicer in immune regulation. J Exp Med. (2006) 203:2519–27. doi: 10.1084/jem.20061692
18. Zhou P, Zhang X, Zeng W, Zheng Q, Hao X, Lin X, et al. MicroRNA expression analysis of feline and canine parvovirus infection in vivo (felis). PLoS ONE. (2017) 12:1–13. doi: 10.1371/journal.pone.0185698
19. Li D, Yang J, Yang Y, Liu J, Li H, Li R, et al. A timely review of cross-kingdom regulation of plant-derived microRNAs. Front Genet. (2021) 12:613197. doi: 10.3389/fgene.2021.613197
20. Liang H, Zhang S, Fu Z, Wang Y, Wang N, Liu Y, et al. Effective detection and quantification of dietetically absorbed plant microRNAs in human plasma. J Nutr Biochem. (2015) 26:505–12. doi: 10.1016/j.jnutbio.2014.12.002
21. Luo Y, Wang P, Wang X, Wang Y, Mu Z, Li Q, et al. Detection of dietetically absorbed maize-derived microRNAs in pigs. Sci Rep. (2017) 7:1–10. doi: 10.1038/s41598-017-00488-y
22. Chapado LA, Martín-Hernández R, Hernández De La Red S, Tomé-Carneiro J, Gil-Zamorano J, Ruiz-Roso MB, et al. Connection between miRNA mediation and the bioactive effects of broccoli (Brassica oleracea var. italica): exogenous miRNA resistance to food processing and GI digestion. J Agric Food Chem. (2021) 69:9326–37. doi: 10.1021/acs.jafc.1c04087
23. Ding T, Song G, Liu X, Xu M, Li Y. Nucleotides as optimal candidates for essential nutrients in living organisms: a review. J Funct Foods. (2021) 82:104498. doi: 10.1016/j.jff.2021.104498
24. Navarro J, Ruiz-Bravo A, Jiménez-Valera M, Gil A. Modulation of antibody-forming cell and mitogen-driven lymphoproliferative responses by dietary nucleotides in mice. Immunol Lett. (1996) 53:141–5. doi: 10.1016/S0165-2478(96)02624-7
25. Gil A. Modulation of the immune response mediated by dietary nucleotides. Eur J Clin Nutr. (2002) 56:1–4. doi: 10.1038/sj.ejcn.1601475
26. Sanchez-Pozo A, Gil A. Nucleotides as semiessential nutritional components. Br J Nutr. (2002) 87:135–7. doi: 10.1079/BJN2001467
27. Fanslow WC, Kulkarni AD, Buren CT, Van, Rudolph FB. Effect of nucleotide restriction and supplementation on resistance to experimental murine candidiasis. J Parenter Enter Nutr. (1988) 12:49–52. doi: 10.1177/014860718801200149
28. Kulkarni AD, Van Buren CT, Fanslow WC, Rudolph FB. Effect of dietary nucleotides on response to bacterial infections. JPEN J Parenter Enteral Nutr. (1986) 10:169–71. doi: 10.1177/0148607186010002169
29. Carver JD. Dietary nucleotides: cellular immune, intestinal and hepatic system effects. J Nutr. (1994) 124:144–8. doi: 10.1093/jn/124.suppl_1.144S
30. Buren CT, Van, Kulkarni AD, Fanslow WC, Rudolph FB. Dietary nucleotides, a requirement for helper/inducer t lymphocytes. Transplantation. (1985) 40:694–7. doi: 10.1097/00007890-198512000-00024
31. Pickering LK, Granoff DM, Erickson JR, Masor ML, Cordle CT, Schaller JP, et al. Modulation of the immune system by human milk and infant formula containing nucleotides. Pediatrics. (1998) 101:242–9. doi: 10.1542/peds.101.2.242
32. Bedu-Ferrari C, Biscarrat P, Langella P, Cherbuy C. Prebiotics and the human gut microbiota: from breakdown mechanisms to the impact on metabolic health. Nutrients. (2022) 14:2096. doi: 10.3390/nu14102096
33. Enam F, Mansell TJ. Prebiotics: tools to manipulate the gut microbiome and metabolome. J Ind Microbiol Biotechnol. (2019) 46:1445–59. doi: 10.1007/s10295-019-02203-4
34. Grześkowiak Ł, Endo A, Beasley S, Salminen S. Microbiota and probiotics in canine and feline welfare. Anaerobe. (2015) 34:14–23. doi: 10.1016/j.anaerobe.2015.04.002
35. Rochus K, Janssens GPJ, Hesta M. Dietary fibre and the importance of the gut microbiota in feline nutrition: a review. Nutr Res Rev. (2014) 27:295–307. doi: 10.1017/S0954422414000213
36. Sohel MMH. Macronutrient modulation of mRNA and microRNA function in animals: a review. Anim Nutr. (2020) 6:258. doi: 10.1016/j.aninu.2020.06.002
37. Martino E, D'Onofrio N, Balestrieri A, Colloca A, Anastasio C, Sardu C, et al. Dietary epigenetic modulators: unravelling the still-controversial benefits of miRNAs in nutrition and disease. Nutrients. (2024) 16:160. doi: 10.3390/nu16010160
38. Hintikka J, Lensu S, Mäkinen E, Karvinen S, Honkanen M, Lindén J, et al. Xylo-oligosaccharides in prevention of hepatic steatosis and adipose tissue inflammation: associating taxonomic and metabolomic patterns in fecal microbiomes with biclustering. Int J Environ Res Public Health. (2021) 18:1–24. doi: 10.20944/preprints202103.0001.v1
39. Pachikian BD, Essaghir A, Demoulin JB, Catry E, Neyrinck AM, Dewulf EM, et al. Prebiotic approach alleviates hepatic steatosis: implication of fatty acid oxidative and cholesterol synthesis pathways. Mol Nutr Food Res. (2013) 57:347–59. doi: 10.1002/mnfr.201200364
40. Gerin I, Clerbaux LA, Haumont O, Lanthier N, Das AK, Burant CF, et al. Expression of miR-33 from an SREBP2 intron inhibits cholesterol export and fatty acid oxidation. J Biol Chem. (2010) 285:33652. doi: 10.1074/jbc.M110.152090
41. Lim E, Lim JY, Kim E, Kim YS, Shin JH, Seok PR, et al. Xylobiose, an alternative sweetener, ameliorates diabetes-related metabolic changes by regulating hepatic lipogenesis and miR-122a/33a in db/db mice. Nutrients. (2016) 8:1–18. doi: 10.3390/nu8120791
42. Klein J, Altenbuchner J, Mattes R. Nucleic acid and protein elimination during the sugar manufacturing process of conventional and transgenic sugar beets. J Biotechnol. (1998) 60:145–53. doi: 10.1016/S0168-1656(98)00006-6
43. AAFCO. AAAFCO methods for substantiating nutritional adequacy of dog and cat food. In: AAFCO Dog and Cat Food Nutrient Profiles. Orlando, FL: Orlando Press (2014). p. 1–24.
44. Nutritional guidelines for complete and complementary pet food for cats and dogs. Bruxelles: The European Pet Food Industry Federation (FEDIAF) (2021). p. 1–96.
45. Atwal J, Joly W, Bednall R, Albanese F, Farquhar M, Holcombe LJ, et al. Dietary supplementation with nucleotides, short-chain fructooligosaccharides, xylooligosaccharides, beta-carotene and vitamin e influences immune function in kittens. Animals. (2023) 13:3734. doi: 10.3390/ani13233734
46. Kanakubo K, Fascetti AJ, Larsen JA. Assessment of protein and amino acid concentrations and labeling adequacy of commercial vegetarian diets formulated for dogs and cats. J Am Vet Med Assoc. (2015) 247:385–92. doi: 10.2460/javma.247.4.385
47. Nutrient Requirements of Dogs and Cats. In: Nutrient Requirements of Dogs and Cats. Washington, DC: The National Academies Press (2006). p. 13–368.
48. Schmittgen TD, Livak KJ. Analyzing real-time PCR data by the comparative CT method. Nat Protoc. (2008) 3:1101–8. doi: 10.1038/nprot.2008.73
49. Huang HY, Lin YCD Li J, Huang KY, Shrestha S, Hong HC, et al. MiRTarBase 2020: Updates to the experimentally validated microRNA-target interaction database. Nucleic Acids Res. (2020) 48:148–54. doi: 10.1093/nar/gkz896
50. Benjamini Y, Hochberg Y. Controlling the false discovery rate: a practical and powerful approach to multiple testing. J R Stat Soc. (1995) 57:289–300. doi: 10.1111/j.2517-6161.1995.tb02031.x
51. Wong RKY, MacMahon M, Woodside J V, Simpson DA. A comparison of RNA extraction and sequencing protocols for detection of small RNAs in plasma. BMC Genomics. (2019) 20:1–12. doi: 10.1186/s12864-019-5826-7
52. Li Y, Kowdley K V. Method for microRNA isolation from clinical serum samples. Anal Biochem. (2012) 431:69–75. doi: 10.1016/j.ab.2012.09.007
53. Huggett J, Dheda K, Bustin S, Zumla A. Real-time RT-PCR normalisation; strategies and considerations. Genes Immun. (2005) 6:279–84. doi: 10.1038/sj.gene.6364190
54. Peter ME. Targeting of mRNAs by multiple miRNAs: the next step. Oncogene. (2010) 29:2161–4. doi: 10.1038/onc.2010.59
55. Townley-Tilson WHD, Callis TE, Wang D. MicroRNAs 1, 133, and 206: critical factors of skeletal and cardiac muscle development, function, and disease. Int J Biochem Cell Biol. (2010) 42:1252–5. doi: 10.1016/j.biocel.2009.03.002
56. Coccia E, Masanas M, López-Soriano J, Segura MF, Comella JX, Pérez-García MJ. FAIM is regulated by miR-206, miR-1-3p and miR-133b. Front Cell Dev Biol. (2020) 8:1–9. doi: 10.3389/fcell.2020.584606
57. Zhou G, Xu Q, Wu F, Wang M, Chen L, Hu L, et al. Arginine alters miRNA expression involved in development and proliferation of rat mammary tissue. Animals. (2021) 11:1–15. doi: 10.3390/ani11020535
58. Shin C, Ito Y, Ichikawa S, Tokunaga M, Sakata-Sogawa K, Tanaka T. MKRN2 is a novel ubiquitin E3 ligase for the p65 subunit of NF-κB and negatively regulates inflammatory responses. Sci Rep. (2017) 7:1–10. doi: 10.1038/srep46097
59. Cano F, Bye H, Duncan LM, Buchet-Poyau K, Billaud M, Wills MR, et al. The RNA-binding E3 ubiquitin ligase MEX-3C links ubiquitination with MHC-I mRNA degradation. EMBO J. (2012) 31:3596–606. doi: 10.1038/emboj.2012.218
60. Xing J, Zhou X, Fang M, Zhang E, Minze LJ, Zhang Z. DHX15 is required to control RNA virus-induced intestinal inflammation. Cell Rep. (2021) 35:109–205. doi: 10.1016/j.celrep.2021.109205
61. Tang R, Ma F, Li W, Ouyang S, Liu Z, Wu J. mir-206-3p inhibits 3T3-L1 cell adipogenesis via the c-Met/Pi3k/Akt pathway. Int J Mol Sci. (2017) 18:1–10. doi: 10.3390/ijms18071510
62. Zhang Y, Xia M, Jin K, Wang S, Wei H, Fan C, et al. Function of the c-Met receptor tyrosine kinase in carcinogenesis and associated therapeutic opportunities. Mol Cancer. (2018) 17:1–14. doi: 10.1186/s12943-018-0796-y
63. Xue L, Chiang L, Kang C, Winoto A. The role of the PI3K-AKT kinase pathway in T-cell development beyond the β checkpoint. Eur J Immunol. (2008) 38:3200–7. doi: 10.1002/eji.200838614
64. So L, Fruman DA. PI3K signalling in B- and T-lymphocytes: new developments and therapeutic advances. Biochem J. (2012) 442:465–81. doi: 10.1042/BJ20112092
65. Liu C, Wang J, Zhang X. The involvement of miR-1-clathrin pathway in the regulation of phagocytosis. PLoS ONE. (2014) 9:1–8. doi: 10.1371/journal.pone.0098747
66. Bandyopadhyay S, Lane T, Venugopal R, Parthasarathy PT, Cho Y, Galam L, et al. MicroRNA-133a-1 regulates inflammasome activation through uncoupling protein-2. Biochem Biophys Res Commun. (2013) 439:407–12. doi: 10.1016/j.bbrc.2013.08.056
67. Yi Q, Xie W, Sun W, Sun W, Liao Y. A concise review of microRNA-383: Exploring the insights of its function in tumorigenesis. J Cancer. (2022) 13:313–24. doi: 10.7150/jca.64846
68. Zhang Y. Investigating targets of miR-383-5p in immune-induced regeneration (MSc thesis). McGill University (2020).
69. Lokau J, Garbers C. Activating mutations of the gp130/JAK/STAT pathway in human diseases. In:Donev R, , editor. Advances in Protein Chemistry and Structural Biology. Cambridge, MA: Academic Press (2019), p. 283–309. doi: 10.1016/bs.apcsb.2018.11.007
70. Chen YH, van Zon S, Adams A, Schmidt-Arras D, Laurence ADJ, Uhlig HH. The human GP130 cytokine receptor and its expression—an atlas and functional taxonomy of genetic variants. J Clin Immunol. (2024) 44:1–15. doi: 10.1007/s10875-023-01603-7
71. Dhuban K. Bin, Bartolucci S, D'Hennezel E, Piccirillo CA. Signaling through gp130 compromises suppressive function in human Foxp3+ regulatory T cells. Front Immunol. (2019) 10:1–13. doi: 10.3389/fimmu.2019.01532
72. Carr LE, Bowlin AK, Elolimy AA, Byrum SD, Washam CL, Randolph CE, et al. Neonatal diet impacts circulatory miRNA profile in a porcine model. Front Immunol. (2020) 11:1–10. doi: 10.3389/fimmu.2020.01240
Keywords: microRNA, nucleotide, FOS, XOS, feline, immune, development, biomarker
Citation: Albanese F, Harrison M, Farquhar MJ, Holcombe LJ, Atwal J, Watson P and Piccinini AM (2025) Dietary supplementation of nucleotides and oligosaccharides in kittens reduces the expression of circulating miR-1-3p, miR-133a-3p, miR-206-3p and miR-383-5p. Front. Vet. Sci. 11:1382436. doi: 10.3389/fvets.2024.1382436
Received: 13 March 2024; Accepted: 23 September 2024;
Published: 06 November 2025.
Edited by:
Luciana Rossi, University of Milan, ItalyReviewed by:
Marcos Edgar Herkenhoff, University of São Paulo, BrazilFrancisco Guzmán, Metropolitan Autonomous University, Mexico
Copyright This work is authored by Albanese, Harrison, Farquhar, Holcombe, Atwal, Watson, and Piccinini. © 2024 Mars Petcare UK and its affiliates for Albanese, Harrison, Farquhar, Holcombe, Atwal, and Watson, and © 2024 Anna M. Piccinini. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original authors and the copyright owner are credited and the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Fabio Albanese, YWxiYW5lc2UuZmFiQGdtYWlsLmNvbQ==