 Talita Gabriela Luna Alves1
Talita Gabriela Luna Alves1 Pedro Luiz Porfirio Xavier1
Pedro Luiz Porfirio Xavier1 Taismara Kustro Garnica1
Taismara Kustro Garnica1 Arina Lázaro Rochetti1
Arina Lázaro Rochetti1 Talal Jamil Qazi1
Talal Jamil Qazi1 Thiago Henrique Annibale Vendramini2Felipe Haddad3
Thiago Henrique Annibale Vendramini2Felipe Haddad3 Muhammad Munir4
Muhammad Munir4 Márcio Antonio Brunetto2†
Márcio Antonio Brunetto2† Heidge Fukumasu1*
Heidge Fukumasu1*- 1Laboratory of Comparative and Translational Oncology, Department of Veterinary Medicine, School of Animal Science and Food Engineering, University of São Paulo (USP), Pirassununga, Brazil
- 2Nutrition and Production Department, School of Veterinary Medicine and Animal Science, Pet Nutrology Research Center, University of São Paulo (USP), Pirassununga, Brazil
- 3Advanced Cellular Therapy Laboratory, Ribeirao Preto Blood Center Foundation, University of São Paulo (USP), Ribeirao Preto, Brazil
- 4Division of Biomedical and Life Sciences, Lancaster University, Lancaster, United Kingdom
Lymphoma is one of the most prevalent types of feline cancer. It is characterized as a group of diseases that can affect various organs, such as the gastrointestinal tract, kidneys, thymus, and skin. In feline medicine, the search for alternative treatments is of utmost importance, given the significant number of animals that relapse or are unresponsive to conventional chemotherapy treatment. As an alternative to existing modalities of treatment for a variety of cancers, oncolytic viruses have been studied in the last few years. Those viruses possess a unique ability to target and eliminate cancer cells while simultaneously stimulating an immune response against malignant cells, acting as an immunotherapy. Newcastle Disease Virus (NDV) is an avian paramyxovirus that affects both domestic and wild birds, causing symptoms that range from severe to asymptomatic, depending on the viral strain. Less virulent strains are considered safe for use as a vaccine against Newcastle Disease. In the Oncology field, those strains are also being studied to be used as oncolytic virotherapy for mammals, and several results demonstrate their efficacy in vitro and in vivo. The present study aimed to explore the oncolytic potential of Newcastle Disease Virus expressing green fluorescent protein (NDV-GFP) in feline lymphoma cells isolated from a FeLV-positive patient with thymic lymphoma. The NDV-GFP infected, replicated, and induced apoptosis in feline lymphoma cells. Therefore, these results provide preliminary evidence of the oncolytic activity of NDV in feline leukemia virus-induced lymphoma.
1 Introduction
Lymphoma is a heterogeneous group of diseases that can be classified into various subtypes, including multicentric, gastrointestinal, extranodal, and mediastinal, each with distinct treatment responses and, consequently, different prognoses (1). Mediastinal lymphoma is a type of lymphoma that involves the thymus, mediastinal, and sternal lymph nodes (2). Historically, this disease has been closely associated with feline leukemia virus (FeLV), a retrovirus capable of integrating its genome into the host cell’s native genetic material, establishing a permanent viral reservoir within the cell (3). This may result in somatically acquired insertional mutagenesis, potentially leading to FeLV-induced neoplasms (4).
However, in recent years, the incidence of mediastinal lymphoma in cats has significantly decreased, especially in developed countries, where vaccination and cat population control are more accessible (2, 5–7). Additionally, the introduction of more effective diagnostic tests has helped identify and isolate infected cats, limiting the spread of the virus. This has resulted in a reduction in the number of cats infected with FeLV and, consequently, a decrease in the number of mediastinal lymphoma cases related to the viral infection (8).
Although FeLV is still prevalent in some parts of the world (9–12) and mediastinal lymphoma remains a concern in these regions (13, 14) since cats with FeLV-associated lymphomas tend to have significantly shorter remission and survival times with available chemotherapeutic protocols (1, 13, 15). Recently, a new chemotherapeutic protocol was developed for cats with high-grade multicentric or mediastinal lymphoma in an area endemic for FeLV, known as LOPH (lomustine, vincristine, prednisolone, and doxorubicin). The protocol was well tolerated, and the Median survival time (MST) was better than similar studies with other protocols (16).
Nevertheless, new treatment strategies are still necessary for FeLV-positive cats with lymphoma since the responses to treatment can vary significantly between individual cats, with some achieving long remissions and others experiencing rapid relapses (15). Otherwise, traditional treatments also carry risks of side effects and toxicity (17). Lastly, the growing concern of drug resistance underlines the importance of developing new treatment options since the patient can stop responding to a treatment that once responded well (18). Therefore, ongoing research and the development of novel therapies are essential to improve outcomes for cats with lymphoma, enhancing both their survival and quality of life. As veterinary medicine advances, novel immunotherapies offer promising alternatives that may target tumor cells more precisely.
The use of oncolytic viruses is a therapeutic approach that has gained prominence in recent years, as these agents can directly induce oncolysis and have stimulatory effects on the immune system (19, 20). These viruses can selectively replicate in and destroy tumor cells (21). Most viruses chosen for use in this type of treatment are attenuated strains that can infect and replicate in the chosen species without causing significant side effects (22). The discovery of oncolytic viruses happened by chance, as it was observed that cancer patients infected with certain types of viruses experienced temporary remission of their disease (23). In recent years, clinical trials have demonstrated their efficacy, which can vary according to the type of virus and the delivery method that is being used (22).
In Veterinary Medicine, several viral species are being tested in vitro and in vivo for their oncolytic effects, such as vesicular stomatitis virus (24–27), adenovirus (28–31), Sendai virus (32), measles virus (33, 34), vaccinia virus (35–43), myxoma virus (44, 45), herpes virus (46, 47), reovirus (48–52), canine distemper virus (53–56) and zika virus (57). The Newcastle Disease Virus (NDV) presented oncolytic activity in various types of cancer, including canine primary and metastatic melanoma (58), canine mammary carcinoma (59–61) and several intracranial tumors (62). For human (63, 64) and canine lymphoma cells (63), NDV infected selectively the tumor cells and cell death involved apoptosis analyzed by flow-cytometry. However, more preclinical studies are necessary to explore NDV’s application in different types of lymphoma, including feline lymphomas.
The NDV is an enveloped, non-segmented avian virus with negative-sense RNA belonging to the genus Orthoavulavirus and the family Paramyxoviridae (APMV-1) (65, 66). It exhibits variable virulence, with the more virulent strains causing Newcastle disease (67). NDV strains can be classified as lentogenic, mesogenic, and velogenic according to the symptoms of NDV in chickens (no disease, moderate-to-severe disease, and severe disease with high mortality respectively).
In the poultry industry, lentogenic strains are used globally as live-attenuated vaccines (68, 69); for mammals, NDV has been studied to be used as a vaccine vector (69). There are rare documentations of NDV infections in humans, and those are mostly reported among poultry workers with self-resolving conjunctivitis and no transmission of the virus among humans (70). In cats (and dogs), a study has evaluated NDV as a virus-vectored rabies vaccine, and it has shown its safety even with repeated inoculation with high dosages with no severe clinical signs observed (71).
Hence, we assessed whether NDV is oncolytic for feline lymphoma cells by evaluating the antitumor activity of a recombinant lentogenic (low virulence) strain expressing green fluorescent protein (NDV-GFP) in a feline lymphoma cell line isolated from a cat with thymic lymphoma and positive for FeLV (FeLV3281).
2 Materials and methods
2.1 Cell line
The cell line (FeLV3281) was previously isolated, characterized (72) and acquired from the Cell Bank Riken (Japan). These cells originated from a cat (Felis catus) with thymic lymphoma and were positive for the Feline Leukemia Virus subtype A. The cells were maintained in 75 cm2 flasks at 37°C and 5% CO2 in Gibco Roswell Park Memorial Institute (RPMI) 1,640 medium supplemented with 10% fetal bovine serum (FBS) and 1% Pen-Strep antibiotic. The passage of cells was performed every 3 days, and experiments were conducted using cells between passages 10 and 20. The cells were observed daily using optical microscopy (Axio Vert A1, Zeiss, Jena, Germany). All reagents used for cell culture were purchased from Thermo Fisher Scientific (California, United States) unless otherwise specified.
2.2 Virus titration and morphological analysis
The virus used in this study was a genetically modified LaSota strain expressing GFP (NDV-GFP) (73) and was kindly provided by Dr. Muhammad Munir (Lancaster University, United Kingdom). The virus titer was obtained by calculating tissue culture infectious dose 50% method per milliliter (TCID50/mL) using the Reed and Muench method (74). Briefly, FeLV3281 cells were seeded at 4.5 × 104 cells/well in 96-well plates containing RPMI 1640 medium supplemented with 2% FBS and 1% Pen-Strep antibiotic. After being plated, the cells were immediately exposed to different concentrations of the virus (10−1 to 10−11) in octuplicate, and the cytopathic effects were monitored for 5 days as is commonly determined by the Reed and Muench method. The cytopathic effects evaluated were syncytia formation, cell lysis, vacuolization, clumping, and cell death. Images of the cells were captured in both bright-field, to evaluate the cytopathic effects, and fluorescence field, to evaluate the expression of GFP, using ZEISS—Axio Vert A1 with an Axio Can 503 camera attached using a 520 nm wavelength filter for green color (ZEISS, Germany) every 24 h for 120 h.
2.3 NDV cytotoxicity assay
The FeLV3281 cells were added to 96-well plates at a density of 1 × 104 cells/well in RPMI 1640 medium supplemented with 2% FBS, 1% Pen-Strep antibiotic, 2% Gluta-MAX, and 1% HEPES. Then, cells were immediately exposed to NDV-GFP that had undergone serial dilutions in pure RPMI 1640 with a Multiplicity of Infection (MOI) of 1 × 100, 2 × 10−1, 4 × 10−2, 8 × 10−3, 1.6 × 10−3, 3.2 × 10−4, 6.4 × 10−5, and 1.28 × 10−5 based on the TCID50/mL of 3.433 × 106. MOI was estimated by assuming a correlation between TCID₅₀/mL and the number of infectious particles per cell. The assay was conducted in triplicate, having 3 plates with the same conditions. The plates were incubated with the virus for 24 h. The cells were observed under bright-field and fluorescence conditions, and images of each dilution were captured. Differences between wells treated with different virus dilutions and cells not infected with the virus were compared by visualizing the wells under the microscope. To analyze the cytotoxicity of NDV-GFP, the half-maximal inhibitory concentration (IC50) of the virus was determined using the CellTiter-Blue® reagent (Promega, United States). At the end of the viral treatment, 20 μL of CellTiter-Blue® was added to all wells, and the cells were incubated at 37°C for more than 24 h. The plates were analyzed using a spectrophotometer at wavelengths of 540 nm and 630 nm (LMR 96, Loccus, Brazil), and cell viability was measured in terms of absorbance. The IC50 was determined using GraphPad Prism software (version 8.0; United States), using a nonlinear regression method with dose–response inhibition parameters.
2.4 NDV cell death assay
The FeLV3281 cells were added to T25 cm2 flasks at a concentration of 1 × 106 cells per flask in RPMI 1640 medium supplemented with 2% FBS, 1% Pen-Strep antibiotic, 2% glutaMAX, and 1% HEPES. Subsequently, the cells were immediately exposed to the IC50 calculated in the cytotoxicity assay. The assay was conducted in triplicate, having 3 wells with the same conditions. The flasks were maintained in a CO2 incubator at 37°C for 24 h. The cells were observed under bright-field and fluorescent conditions, and images in each field were captured. To analyze whether the observed cytopathic effects were triggered by an increase in the number of cell death, a flow cytometry assay using propidium iodide (PI) was conducted following a protocol from Riccardi et al. (75). Briefly, the cells were centrifuged at 1,200 rpm for 5 min, and the supernatant was discarded, and the cells were resuspended in 500 μL of PBS. Cells were fixed in 4.5 mL of 70% chilled ethanol and stored at −20°C for at least 24 h. The cells were centrifuged at 1,800 rpm for 5 min, the supernatant was discarded, and the cells were washed in 5 mL PBS and centrifuged at 1,800 rpm for 5 min. The cells were then resuspended in 1 mL of PI staining solution (200 μg in 10 mL PBS and 2 mg DNAse-free RNAse) and incubated for half an hour at room temperature. It is important to note that the use of propidium iodide (PI) staining alone assesses membrane integrity and does not allow us to distinguish between late apoptosis and necrosis. 20,000 events were captured using a flow cytometer (S3e Cell Sorter, BIO-RAD, Hercules, CA, United States) with a 488-nm laser line for excitation, measuring red fluorescence (4,600 nm) and side scatter. The gating strategy consisted of an initial exclusion of debris based on a side scatter area (SSC-A) vs. forward scatter area (FSC-A) plot. Singlet cells were then selected using a FSC-A versus FL2-A (PI fluorescence) plot to exclude doublets and aggregates. Finally, PI fluorescence was analyzed using a histogram (count vs. FL2-A). Statistical analyses were performed by the unpaired t-test.
3 Results
3.1 NDV-GFP infects and replicates in feline lymphoma cells
The NDV-GFP reduced the feline lymphoma cells dose-dependently (Figure 1). The titter obtained for NDV-GFP in FeLV3281 cells was 3.433 × 106 tissue culture infectious dose per milliliter (TCID50/mL) after 120 h of observation of the cytopathic effects. No syncytia were observed. Although it was observed cell lysis and cell death (Figure 1). The detection of GFP indicated the virus was viable and was able to infect and replicate in the feline lymphoma cells as observed from 24 h post-infection (p.i.) in all wells from the first to the fifth dilution (10−1 to 10−5) and persisted up to 72 h p.i., albeit with lower intensity over the days (Figure 2). The GFP was not detected from the 10−7 dilution along (Figure 2).
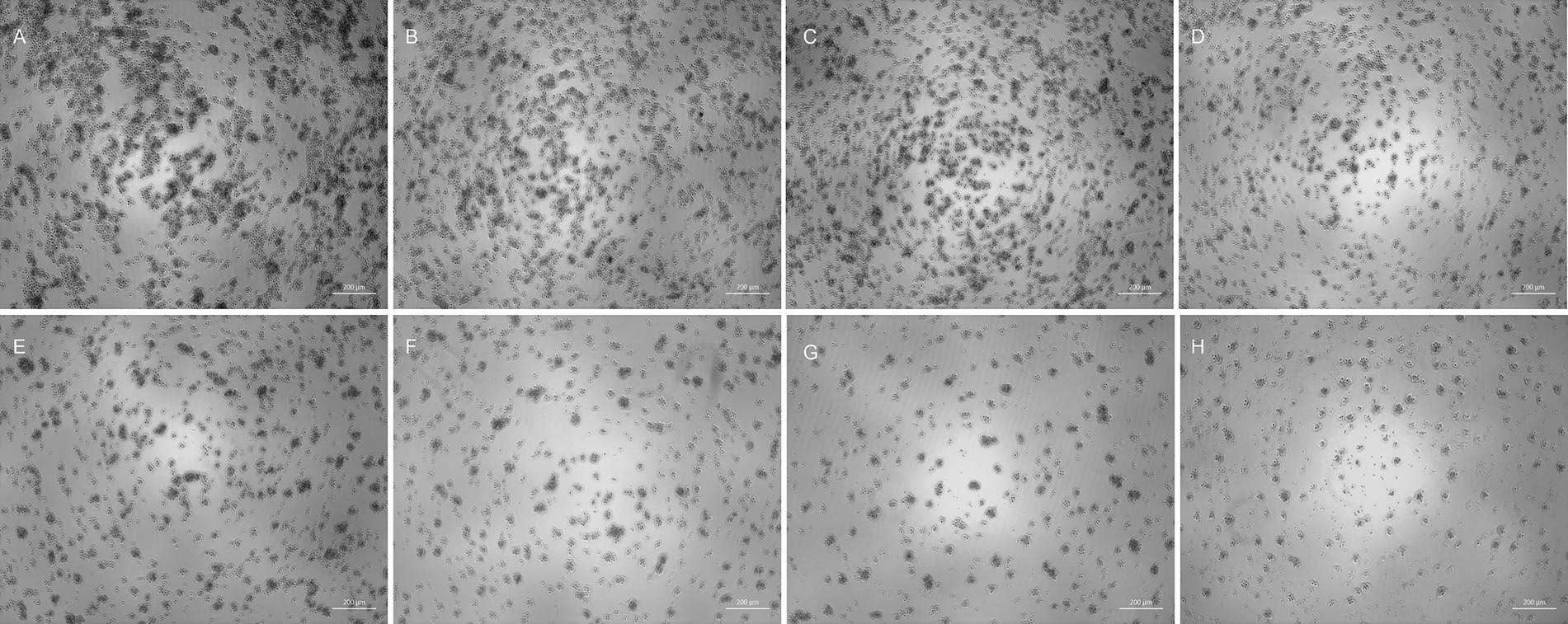
Figure 1. Viral titration by the Reed-Muench method with analysis of cytopathic effects in bright-field 120 h post-infection. Control well (A) and wells infected with viral dilution of 10−11 (B), 10−8 (C), 10−5 (D), 10−4 (E), 10−3 (F), 10−2 (G), and 10−1 (H). Arrows (H) indicate cellular debris. As the virus concentration increased, the well confluence decreased, leading to an increased number of cellular debris observed in dilutions 10−1 and 10−2. Although GFP expression can only be observed up to 72 h post-infection, the evaluation of cytopathic effects is performed up to 120 h post-infection according to the Reed-Muench method. 50x magnification.

Figure 2. Viral titration assay. Besides control (A, F and K), it is possible to observe FeLV3281 cells that did not express GFP at the concentration of 10−7 (B, G and L). At a concentration of 10−5 (C, H and M), 10−3 (D, I and N) and 10−1 (E, J and O), FeLV3281 cells express GFP. The cells were evaluated using bright-field and fluorescent microscopy at 24 (A–E), 48 (F–J), and 72 h (K–O) post-infection (p.i.), regarding GFP expression assessment and evaluation of cytopathic effects. Higher GFP expression was observed within the first 24 h of the assay. When comparing control and treatment wells, it is suggested by the observation of cells under microscopy that the confluence is lower in the treatment wells. 100x magnification.
3.2 NDV-GFP is cytotoxic and induces cell death in lymphoma cells
The NDV-GFP is oncolytic for FeLV3281 cells, as demonstrated by a dose-dependent effect on cell viability with an IC50 of MOI = 3.201 × 10−1 ± 0.04 (Supplementary Table S1). When compared to untreated cells (Figure 3A), there was observed a reduction of cell number in cells treated with the IC50 of the NDV-GFP. Besides, GFP was detected in cells treated with the IC50 of the virus (Figure 3B), unlike untreated cells (Figure 3A). In cell death analysis, it was also observed a significantly higher occurrence of cell death was observed in cells treated with the IC50 (61.95% ± 1.68%) compared to untreated cells (1.80% ± 0.39%, p < 0.0002; Figures 3C–H; Supplementary Table S2). Cell clumping was also observed (Figure 4).
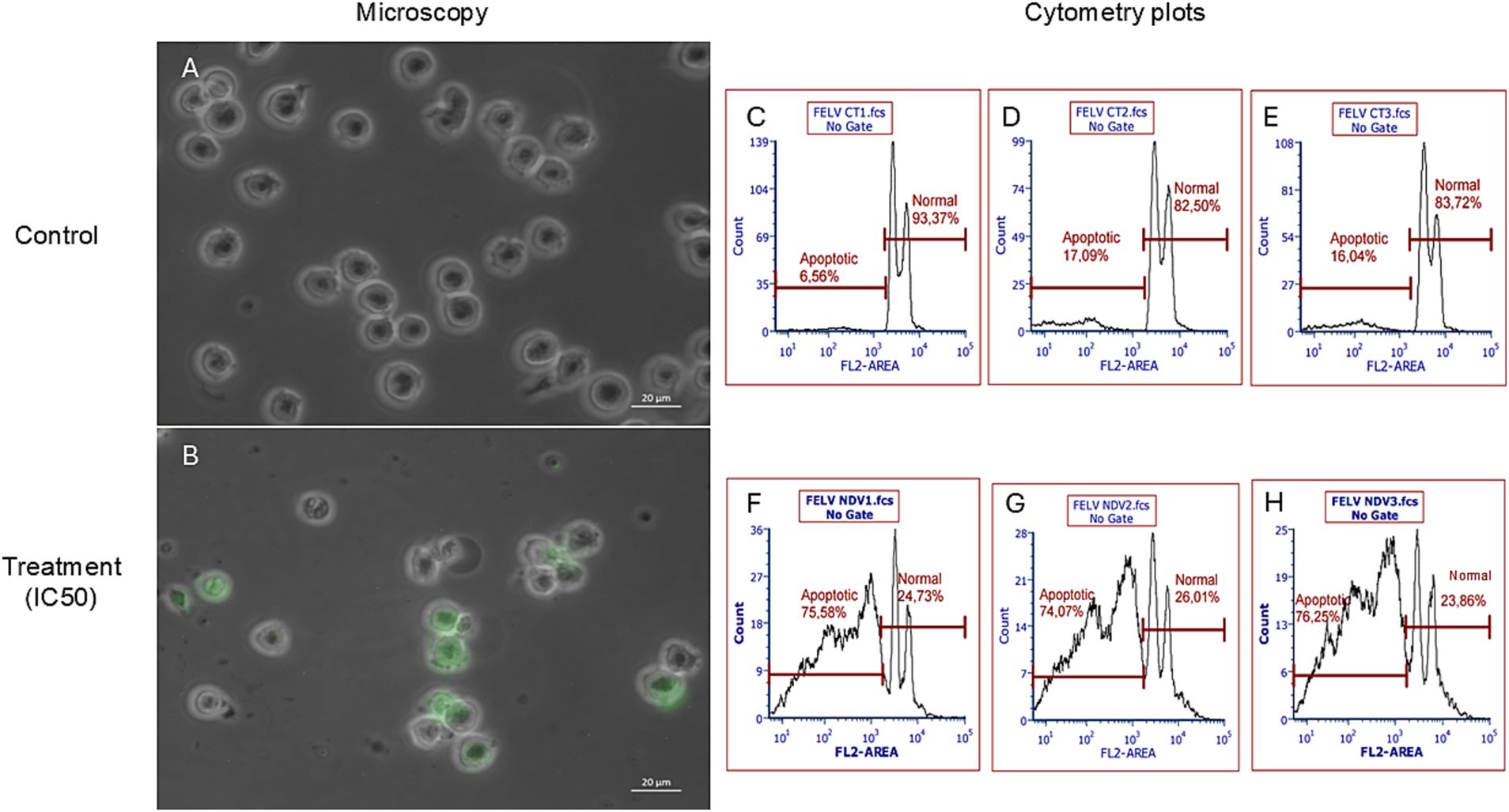
Figure 3. Evaluation of the effects of NDV-GFP on feline lymphoma cells (FeLV3281 cell line). A decrease in the number of viable cells as well as expression of GFP was observed in cells treated with the IC50 (B), but not in untreated cells (A). Bright-field and fluorescence images were superimposed (A,B) to further highlight GFP expression. The percentage of apoptotic cells and viable cells in non-treated (C–E) and treated (F–H) cells was compared by flow cytometry analysis. “FL2 area” is the measure of the total fluorescence captured in the FL2 channel.
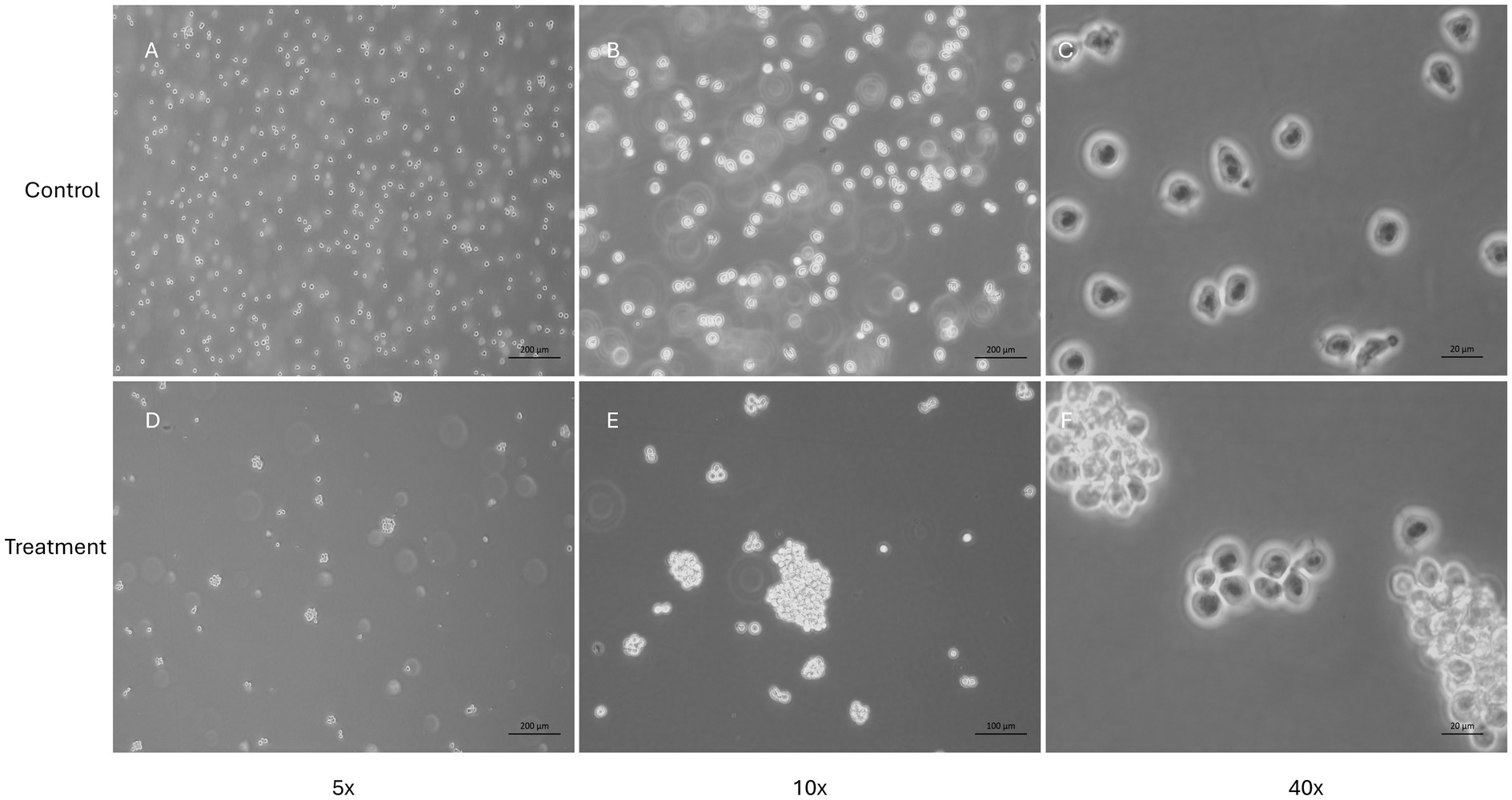
Figure 4. Evaluation of feline lymphoma cells (FeLV3281 cell line) treated with the IC50 of NDV-GFP. This figure presents the morphology of cells unexposed to NDV-GFP (A–C) and cells treated with the IC50 concentration of NDV-GFP (D–F). In virus-treated wells, a noticeable reduction in cell confluence is observed, along with the formation of distinct cell clumps, suggesting cytopathic effects induced by the oncolytic virus. In contrast, control cells maintain a typical morphology, with no cell clumps and a higher confluence.
4 Discussion
There is an urgent need for the development of new candidate therapies for feline lymphoma due to its frequency and the drawbacks of conventional chemotherapy, such as tumor relapse and the possibility of side effects. Here, we demonstrated the oncolytic effect of NDV-GFP in the feline lymphoma cell line FeLV328, as shown by its ability to infect, replicate, and induce morphological changes, culminating in cell death in cancer cells. Additional evidence demonstrated these effects as dose-dependent in the lymphoma cells.
In this study, we used a recombinant lentogenic La Sota variant of NDV-GFP that allowed the detection by fluorescent microscopy of the GFP-expressing cancer cells after virus exposure, assuring that this NDV-GFP infected and replicated in cancer cells. This virus was initially created to determine the tissue tropism of non-virulent NDV in chicken embryos (73). It became a good tool for researching new cancer therapies based on NDV since this virus could be genetically modified to express other proteins or ncRNAs of interest by reverse genetics. Another advantage of the lentogenic NDV-GFP for cancer therapy is the fact that it can be considered safe in non-natural hosts like mammals since it typically results in mild clinical signs, such as conjunctivitis and flu-like symptoms, mostly when exposed to more virulent strains or higher doses of the virus (76–78). The absence of syncytia formation in cancer cells after NDV-GFP infection supports this virus as lentogenic, non-virulent, and non-lytic, as we have already demonstrated in canine cancer cells (59).
Although the US-FDA has not approved any NDV therapy for any condition so far (April 2025), several clinical trials with NDV were performed in humans (more information found in: https://www.cancer.gov/about-cancer/treatment/cam/hp/ndv-pdq). Most human clinical trials of NDV for cancer treatment have been conducted as phase I or II clinical trials typically employing a lytic strain, an oncolysate, or vaccines derived from virus-infected tumor cells. There is only one ongoing phase I clinical trial by AstraZeneca (ID: D7880C00001) of a recombinant NDV expressing Interleukin-12 (MEDI9253) registered on the astrazenecaclinicaltrials.com website. This study was completed on May 31, 2024. On the other hand, to the best of our knowledge, this is the first description of the oncolytic effect of NDV in feline tumors.
An important limitation of this study is the absence of an evaluation of NDV’s cytotoxic effects on non-cancerous feline cells, such as peripheral blood mononuclear cells (PBMCs) or fibroblasts. Assessing these cell types is essential to determine the virus’s selectivity toward cancer cells and to characterize its safety profile. Therefore, it is highly recommended that future studies include such analyses before advancing to preclinical and clinical investigations. Nevertheless, previous studies have already demonstrated that NDV is selective for dog cancer cells (59), as well as being safe for use in dogs (60, 61), and at least one study has assessed the safety profile of NDV in cats (71), reporting no significant adverse effects, which supports its potential as a safe oncolytic agent in veterinary oncology.
We have performed cytotoxicity assays using PBMCs treated with serial dilutions of NDV-GFP (1 × 100, 2 × 10−1, 4 × 10−2, 8 × 10−3, 1.6 × 10−3, 3.2 × 10−4, 6.4 × 10−5, and 1.28 × 10−5 MOI, based on the TCID50/mL of 3.433 × 106 – the same methodology used for the lymphoma cell line, FeLV3281) and incubated for 24 h, but the results were inconclusive. In this assay, we have also used the CellTiter-Blue® with the porpuse of analyzing cell viability. CellTiter-Blue® have resazurin as the main reagent. Resazurin is a blue and non-fluorescent dye. In viable cells, it is reduced by cellular metabolic activity into resorufin, which is pink and fluorescent, resorufin. This reduction did not happen with PBMCs, even in control cells (cells that were not treated with the virus). All dilutions (and control wells) remained with the same blue dye, even after 24 h of exposition to the reagent. We hypothesize that PBMCs do not metabolize CellTiter-Blue®, because they are not metabolic active cells and need to be activated with mitogens, such as Concanavalin A (Con A). Therefore, to conduct cell viability assays with CellTiter-Blue®, it is necessary to discover the concentration of Con A that can induce PBMCs to a metabolic active state and, hence, induce the reduction of resazurin to resorufin in viable cells.
Another fact that it would be important to evaluate in the future is how immunosuppressed animals would respond to therapies based on recombinant non-lytic oncolytic viruses, with emphasis on FeLV-positive cats with lymphoma. It is known that innovative therapies need to be developed to improve the survival rate and the quality of life of those animals that are often diminished because of the interaction of both diseases (15, 79–81). Besides the poor prognosis, FeLV is still a disease that has a high prevalence in some countries (9, 13, 14, 82, 83). Therefore, treating cats with FeLV-associated lymphoma is a subject that needs to be addressed with the necessary degree of urgency.
Finally, it is important to note that our study was conducted using only a single FeLV-positive feline lymphoma cell line (FeLV3281). While our findings offer valuable preliminary evidence of NDV-GFP’s oncolytic potential in this specific model, they may not fully reflect the behavior of other feline lymphoma subtypes or FeLV-negative cases. Future studies should include additional cell lines encompassing both FeLV-associated and non-FeLV-associated models to further validate and extend these observations. We also acknowledge that the exclusive use of PI staining has limitations, as it does not differentiate between late apoptotic and necrotic cells. Nevertheless, the dose-dependent cytotoxic effects observed, along with supportive morphological and cell viability data, provide robust evidence that NDV-GFP predominantly induces apoptosis in the FeLV-positive feline lymphoma cell line. In future studies, we plan to further delineate the cell death pathways by incorporating additional markers, such as caspase activity assays.
In conclusion, these preliminary results provide support for the continuation of in vitro studies using oncolytic NDV candidates for the treatment of feline lymphoma, which is an urgent necessity due to the limitations of currently adopted treatment modalities, such as chemotherapy. Future work will extend our investigation into in vivo preclinical models to further assess the safety and therapeutic efficacy of NDV-GFP. Specifically, we plan to develop xenograft models in immunodeficient mice using FeLV-positive feline lymphoma cells to simulate the tumor microenvironment and monitor treatment response in a controlled setting. Additionally, primary tumor cultures derived from FeLV-positive cats will be explored, as these models are likely to reflect better the heterogeneity and complexity of naturally occurring feline lymphoma. These complementary approaches will help bridge our in vitro findings with clinical relevance and guide the design of subsequent clinical studies.
Data availability statement
The original contributions presented in the study are included in the article/Supplementary material, further inquiries can be directed to the corresponding author.
Ethics statement
Ethical approval was not required for the studies on animals in accordance with the local legislation and institutional requirements because only commercially available established cell lines were used.
Author contributions
TA: Conceptualization, Data curation, Formal analysis, Investigation, Methodology, Software, Visualization, Writing – original draft, Writing – review & editing. PX: Conceptualization, Data curation, Formal analysis, Investigation, Methodology, Software, Validation, Visualization, Writing – original draft, Writing – review & editing. TG: Conceptualization, Investigation, Methodology, Visualization, Writing – review & editing. AR: Data curation, Formal analysis, Methodology, Visualization, Writing – review & editing, Investigation. TQ: Writing – review & editing, Data curation, Formal analysis. TV: Writing – review & editing, Resources, Investigation. FH: Methodology, Writing – review & editing, Investigation. MM: Writing – review & editing, Resources. MB: Resources, Writing – review & editing. HF: Conceptualization, Supervision, Methodology, Funding acquisition, Writing – review & editing, Investigation, Writing – original draft, Project administration, Formal analysis, Visualization, Resources, Data curation, Validation.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This research was funded by Conselho Nacional de Desenvolvimento Científico e Tecnológico, grant number: 143954/2021-0, Fundação de Amparo à Pesquisa do Estado de São Paulo, FAPESP, Thematic grant: 2022/09378-5, Fundação de Amparo à Pesquisa do Estado de São Paulo, FAPESP, grant number: 2023/09122-3, and Fundação de Amparo à Pesquisa do Estado de São Paulo, FAPESP, Generation grant: 2022/06305-7.
Acknowledgments
The authors acknowledge Helena L. Ferreira for the initial contact between HF and MM.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be considered a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fvets.2025.1484947/full#supplementary-material
References
1. Vail, DM, Moore, AS, Ogilvie, GK, and Volk, LM. Feline lymphoma (145 cases): proliferation indices, cluster of differentiation 3 immunoreactivity, and their association with prognosis in 90 cats. J Vet Intern Med. (1998) 12:349–54. doi: 10.1111/j.1939-1676.1998.tb02134.x
2. Fabrizio, F, Calam, AE, Dobson, JM, Middleton, SA, Murphy, S, Taylor, SS, et al. Feline mediastinal lymphoma: a retrospective study of signalment, retroviral status, response to chemotherapy and prognostic indicators. J Feline Med Surg. (2014) 16:637–44. doi: 10.1177/1098612X13516621
3. Willett, BJ, and Hosie, MJ. Feline leukaemia virus: half a century since its discovery. Vet J. (2013) 195:16–23. doi: 10.1016/j.tvjl.2012.07.004
4. Fujino, Y, Ohno, K, and Tsujimoto, H. Molecular pathogenesis of feline leukemia virus-induced malignancies: insertional mutagenesis. Vet Immunol Immunopathol. (2008) 123:138–43. doi: 10.1016/j.vetimm.2008.01.019
5. Stavisky, J, Dean, RS, and Molloy, MH. Prevalence of and risk factors for FIV and FeLV infection in two shelters in the United Kingdom (2011-2012). Vet Rec. (2017) 181:451. doi: 10.1136/vr.103857
6. Gleich, SE, Krieger, S, and Hartmann, K. Prevalence of feline immunodeficiency virus and feline leukaemia virus among client-owned cats and risk factors for infection in Germany. J Feline Med Surg. (2009) 11:985–92. doi: 10.1016/j.jfms.2009.05.019
7. Studer, N, Lutz, H, Saegerman, C, Gönczi, E, Meli, ML, Boo, G, et al. Pan-European study on the prevalence of the feline leukaemia virus infection—reported by the european advisory board on cat diseases (ABCD Europe). Viruses. (2019) 11:993. doi: 10.3390/v11110993
8. Guillermo, CC. What is new on feline lymphoma? J Feline Med Surg. (2001) 3:171–6. doi: 10.1053/jfms.2001.0146
9. Biezus, G, Machado, G, Ferian, PE, da Costa, UM, Pereira, LS, Withoeft, JA, et al. Prevalence of and factors associated with feline leukemia virus (FeLV) and feline immunodeficiency virus (FIV) in cats of the state of Santa Catarina, Brazil. Comp Immunol Microbiol Infect Dis. (2019) 63:17–21. doi: 10.1016/j.cimid.2018.12.004
10. Sprißler, F, Jongwattanapisan, P, Luengyosluechakul, S, Pusoonthornthum, R, Reese, S, Bergmann, M, et al. Prevalence and risk factors of feline immunodeficiency virus and feline leukemia virus infection in healthy cats in Thailand. Front Vet Sci. (2022) 8:217. doi: 10.3389/fvets.2021.764217
11. de Almeida, NR, Danelli, MGM, da Silva, LHP, and Mazur, C. Prevalence of feline leukemia virus infection in domestic cats in Rio de Janeiro. J Feline Med Surg. (2012) 14:583–6. doi: 10.1177/1098612X12444693
12. Bande, F, Arshad, SS, Hassan, L, Zakaria, Z, Sapian, NA, Rahman, NA, et al. Prevalence and risk factors of feline leukaemia virus and feline immunodeficiency virus in peninsular Malaysia. BMC Vet Res. (2012) 8:33. doi: 10.1186/1746-6148-8-33
13. Cristo, TG, Biezus, G, Noronha, LF, Pereira, LHHS, Withoeft, JA, Furlan, LV, et al. Feline lymphoma and a high correlation with feline Leukaemia virus infection in Brazil. J Comp Pathol. (2019) 166:20–8. doi: 10.1016/j.jcpa.2018.10.171
14. Biezus, G, Grima de Cristo, T, da Silva, CM, Lovatel, M, Vavassori, M, de Souza, B, et al. Progressive and regressive infection with feline leukemia virus (FeLV) in cats in southern Brazil: prevalence, risk factors associated, clinical and hematologic alterations. Prev Vet Med. (2023) 216:105945. doi: 10.1016/j.prevetmed.2023.105945
15. Sunpongsri, S, Kovitvadhi, A, Rattanasrisomporn, J, Trisaksri, V, Jensirisak, N, and Jaroensong, T. Effectiveness and adverse events of cyclophosphamide, vincristine, and prednisolone chemotherapy in feline mediastinal lymphoma naturally infected with feline leukemia virus. Animals. (2022) 12:900. doi: 10.3390/ani12070900
16. Horta, RS, Souza, LM, Sena, BV, Almeida, IO, Jaretta, TA, Pimenta, MM, et al. LOPH: a novel chemotherapeutic protocol for feline high-grade multicentric or mediastinal lymphoma, developed in an area endemic for feline leukemia virus. J Feline Med Surg. (2021) 23:86–97. doi: 10.1177/1098612X20926893
17. Kent, MS. Cats and chemotherapy: treat as “small dogs” at your peril. J Feline Med Surg. (2013) 15:419–24. doi: 10.1177/1098612X13483240
18. Vasan, N, Baselga, J, and Hyman, DM. A view on drug resistance in cancer. Nature. (2019) 575:299–309. doi: 10.1038/s41586-019-1730-1
19. Lin, D, Shen, Y, and Liang, T. Oncolytic virotherapy: basic principles, recent advances and future directions. Signal Transduct Target Ther. (2023) 8:156. doi: 10.1038/s41392-023-01407-6
20. Ferrucci, PF, Pala, L, Conforti, F, and Cocorocchio, E. Talimogene laherparepvec (T-vec): an intralesional cancer immunotherapy for advanced melanoma. Cancers. (2021) 13:1–14. doi: 10.3390/cancers13061383
21. Chaurasiya, S, Chen, NG, and Fong, Y. Oncolytic viruses and immunity. Curr Opin Immunol. (2018) 51:83–90. doi: 10.1016/j.coi.2018.03.008
22. Mondal, M, Guo, J, He, P, and Zhou, D. Recent advances of oncolytic virus in cancer therapy. Hum Vaccin Immunother. (2020) 16:2389–402. doi: 10.1080/21645515.2020.1723363
23. Kelly, E, and Russell, SJ. History of oncolytic viruses: genesis to genetic engineering. Mol Ther. (2007) 15:651–9. doi: 10.1038/sj.mt.6300108
24. Kimpel, J, Urbiola, C, Koske, I, Tober, R, Banki, Z, Wollmann, G, et al. The oncolytic virus VSV-GP is effective against malignant melanoma. Viruses. (2018) 10:108. doi: 10.3390/v10030108
25. Makielski, KM, Sarver, AL, Henson, MS, Stuebner, KM, Borgatti, A, Suksanpaisan, L, et al. Neoadjuvant systemic oncolytic vesicular stomatitis virus is safe and may enhance long-term survivorship in dogs with naturally occurring osteosarcoma. Mol Ther Oncol. (2023) 31:100736. doi: 10.1016/j.omto.2023.100736
26. Naik, S, Galyon, GD, Jenks, NJ, Steele, MB, Miller, AC, Allstadt, SD, et al. Comparative oncology evaluation of intravenous recombinant oncolytic vesicular stomatitis virus therapy in spontaneous canine cancer. Mol Cancer Ther. (2018) 17:316–26. doi: 10.1158/1535-7163.MCT-17-0432
27. Mehrani, Y, Kakish, JE, Napoleoni, C, Thompson, JJ, Knapp, JP, Minott, JA, et al. Characterisation and sensitivity of a canine mast cell tumour line to oncolytic viruses. Vet Comp Oncol. (2024) 23:42–51. doi: 10.1111/vco.13024
28. Matsugo, H, Kitamura-Kobayashi, T, Kamiki, H, Ishida, H, Sekine, W, Takenaka-Uema, A, et al. A potential bat adenovirus-based oncolytic virus targeting canine cancers. Sci Rep. (2021) 11:16706. doi: 10.1038/s41598-021-96101-4
29. Martín-Carrasco, C, Delgado-Bonet, P, Tomeo-Martín, BD, Pastor, J, de la Riva, C, Palau-Concejo, P, et al. Safety and efficacy of an oncolytic adenovirus as an immunotherapy for canine Cancer patients. Vet Sci. (2022) 9:327. doi: 10.3390/vetsci9070327
30. Cejalvo, T, Perise-Barrios, AJ, del Portillo, I, Laborda, E, Rodriguez-Milla, MA, Cubillo, I, et al. Remission of spontaneous canine tumors after systemic cellular viroimmunotherapy. Cancer Res. (2018) 78:4891–901. doi: 10.1158/0008-5472.CAN-17-3754
31. Cloquell, A, Mateo, I, Gambera, S, Pumarola, M, Alemany, R, García-Castro, J, et al. Systemic cellular viroimmunotherapy for canine high-grade gliomas. J Immunother Cancer. (2022) 10:e005669. doi: 10.1136/jitc-2022-005669
32. Ilyinskaya, GV, Mukhina, EV, Soboleva, AV, Matveeva, OV, and Chumakov, PM. Oncolytic Sendai virus therapy of canine mast cell tumors (a pilot study). Front Vet Sci. (2018) 5:116. doi: 10.3389/fvets.2018.00116
33. Tamura, K, Fujiyuki, T, Moritoh, K, Akimoto, H, Iizuka, K, Sato, H, et al. Anti-tumor activity of a recombinant measles virus against canine lung cancer cells. Sci Rep. (2023) 13:9. doi: 10.1038/s41598-023-42305-9
34. Iizuka, K, Shoji, K, Fujiyuki, T, Moritoh, K, Tamura, K, Yoshida, A, et al. Antitumor activity of an oncolytic measles virus against canine urinary bladder transitional cell carcinoma cells. Res Vet Sci. (2020) 133:313–7. doi: 10.1016/j.rvsc.2020.10.016
35. Béguin, J, Laloy, E, Cochin, S, Gantzer, M, Farine, I, Pichon, C, et al. Oncolytic virotherapy with intratumoral injection of vaccinia virus TG6002 and 5-fluorocytosine administration in dogs with malignant tumors. Mol Ther Oncolytics. (2023) 30:103–16. doi: 10.1016/j.omto.2023.07.005
36. Béguin, J, Foloppe, J, Maurey, C, Laloy, E, Hortelano, J, Nourtier, V, et al. Preclinical evaluation of the oncolytic vaccinia virus TG6002 by translational research on canine breast Cancer. Mol Ther Oncolytics. (2020) 19:57–66. doi: 10.1016/j.omto.2020.08.020
37. Adelfinger, M, Bessler, S, Frentzen, A, Cecil, A, Langbein-Laugwitz, J, Gentschev, I, et al. Preclinical testing oncolytic vaccinia virus strain GLV-5b451 expressing an anti-VEGF single-chain antibody for canine cancer therapy. Viruses. (2015) 7:4075–92. doi: 10.3390/v7072811
38. Petrov, I, Gentschev, I, Vyalkova, A, Elashry, MI, Klymiuk, MC, Arnhold, S, et al. Canine adipose-derived mesenchymal stem cells (cAdMSCs) as a “Trojan horse” in vaccinia virus mediated oncolytic therapy against canine soft tissue sarcomas. Viruses. (2020) 12:750. doi: 10.3390/v12070750
39. Béguin, J, Gantzer, M, Farine, I, Foloppe, J, Klonjkowski, B, Maurey, C, et al. Safety, biodistribution and viral shedding of oncolytic vaccinia virus TG6002 administered intravenously in healthy beagle dogs. Sci Rep. (2021) 11:2209. doi: 10.1038/s41598-021-81831-2
40. Autio, K, Knuuttila, A, Kipar, A, Ahonen, M, Parviainen, S, Diaconu, I, et al. Anti-tumour activity of oncolytic Western reserve vaccinia viruses in canine tumour cell lines, xenografts, and fresh tumour biopsies. Vet Comp Oncol. (2016) 14:395–408. doi: 10.1111/vco.12119
41. Béguin, J, Nourtier, V, Gantzer, M, Cochin, S, Foloppe, J, Balloul, JM, et al. Safety studies and viral shedding of intramuscular administration of oncolytic vaccinia virus TG6002 in healthy beagle dogs. BMC Vet Res. (2020) 16:307. doi: 10.1186/s12917-020-02524-y
42. Cambien, B, Lebrigand, K, Baeri, A, Nottet, N, Compin, C, Lamit, A, et al. Identification of oncolytic vaccinia restriction factors in canine high-grade mammary tumor cells using single-cell transcriptomics. PLoS Pathog. (2020) 16:e1008660. doi: 10.1371/journal.ppat.1008660
43. Adelfinger, M, Gentschev, I, De Guibert, JG, Weibel, S, Langbein-Laugwitz, J, Härtl, B, et al. Evaluation of a new recombinant oncolytic vaccinia virus strain GLV-5b451 for feline mammary carcinoma therapy. PLoS One. (2014) 9:337. doi: 10.1371/journal.pone.0104337
44. Macneill, AL, Weishaar, KM, Séguin, B, and Powers, BE. Safety of an oncolytic myxoma virus in dogs with soft tissue sarcoma. Viruses. (2018) 10:398. doi: 10.3390/v10080398
45. Ashton, LV, Weishaar, KM, Séguin, B, and Mac Neill, AL. Oclacitinib and Myxoma virus therapy in dogs with high-grade soft tissue sarcoma. Biomedicines. (2023) 11:2346. doi: 10.3390/biomedicines11092346
46. Chambers, MR, Foote, JB, Bentley, RT, Botta, D, Crossman, DK, Della Manna, DL, et al. Evaluation of immunologic parameters in canine glioma patients treated with an oncolytic herpes virus. J Transl Genet Genom. (2021) 5:423–42. doi: 10.20517/jtgg.2021.31
47. Omar, NB, Bentley, RT, Crossman, DK, Foote, JB, Koehler, JW, Markert, JM, et al. Safety and interim survival data after intracranial administration of M032, a genetically engineered oncolytic HSV-1 expressing IL-12, in pet dogs with sporadic gliomas. Neurosurg Focus. (2021) 50:1–11. doi: 10.3171/2020.11.FOCUS20844
48. Igase, M, Hwang, CC, Coffey, M, Okuda, M, Noguchi, S, and Mizuno, T. The oncolytic effects of reovirus in canine solid tumor cell lines. J Vet Med Sci. (2015) 77:541–8. doi: 10.1292/jvms.14-0570
49. Igase, M, Shousu, K, Fujiki, N, Sakurai, M, Bonkobara, M, Hwang, CC, et al. Anti-tumour activity of oncolytic reovirus against canine histiocytic sarcoma cells. Vet Comp Oncol. (2019) 17:184–93. doi: 10.1111/vco.12468
50. Hwang, CC, Umeki, S, Igase, M, Coffey, M, Noguchi, S, Okuda, M, et al. The effects of oncolytic reovirus in canine lymphoma cell lines. Vet Comp Oncol. (2016) 14:61–73. doi: 10.1111/vco.12124
51. Hwang, CC, Igase, M, Sakurai, M, Haraguchi, T, Tani, K, Itamoto, K, et al. Oncolytic reovirus therapy: pilot study in dogs with spontaneously occurring tumours. Vet Comp Oncol. (2018) 16:229–38. doi: 10.1111/vco.12361
52. Igase, M, Shibutani, S, Kurogouchi, Y, Fujiki, N, Hwang, CC, Coffey, M, et al. Combination therapy with Reovirus and ATM inhibitor enhances cell death and virus replication in canine melanoma. Mol Ther Oncolytics. (2019) 15:49–59. doi: 10.1016/j.omto.2019.08.003
53. Marek, K, Armando, F, Nippold, VM, Rohn, K, Plattet, P, Brogden, G, et al. Persistent infection of a canine histiocytic sarcoma cell line with attenuated canine distemper virus expressing Vasostatin or granulocyte-macrophage Colony-stimulating factor. Int J Mol Sci. (2022) 23:156. doi: 10.3390/ijms23116156
54. Gu, C, Su, J, Wang, J, Xie, Q, Wu, J, Xiao, J, et al. Fusion protein and hemagglutinin of canine distemper virus co-induce apoptosis in canine mammary tumor cells. J Cancer Res Clin Oncol. (2023) 149:9903–18. doi: 10.1007/s00432-023-04878-w
55. Armando, F, Gambini, M, Corradi, A, Giudice, C, Pfankuche, VM, Brogden, G, et al. Oxidative stress in canine histiocytic sarcoma cells induced by an infection with canine distemper virus led to a dysregulation of HIF-1α downstream pathway resulting in a reduced expression of VEGF-B in vitro. Viruses. (2020) 12:200. doi: 10.3390/v12020200
56. Li, P, Wang, J, Chen, G, Zhang, X, Lin, D, Zhou, Y, et al. Oncolytic activity of canine distemper virus in canine mammary tubular adenocarcinoma cells. Vet Comp Oncol. (2019) 17:174–83. doi: 10.1111/vco.12466
57. Kaid, C, Madi, RA, Astray, R, Goulart, E, Caires-Junior, LC, Mitsugi, TG, et al. Safety, tumor reduction, and clinical impact of Zika virus injection in dogs with advanced-stage brain tumors. Mol Ther. (2020) 28:1276–86. doi: 10.1016/j.ymthe.2020.03.004
58. Numpadit, S, Ito, C, Nakaya, T, and Hagiwara, K. Investigation of oncolytic effect of recombinant Newcastle disease virus in primary and metastatic oral melanoma. Med Oncol. (2023) 40:138. doi: 10.1007/s12032-023-02002-z
59. Santos, MR, Xavier, PLP, Pires, PRL, Rochetti, AL, Rosim, DF, Scagion, GP, et al. Oncolytic effect of Newcastle disease virus is attributed to interferon regulation in canine mammary cancer cell lines. Vet Comp Oncol. (2021) 19:593–601. doi: 10.1111/vco.12699
60. Wang, J, Li, M, and Li, M. Newcastle disease virus LaSota strain induces apoptosis and activates the TNFα/NF-κB pathway in canine mammary carcinoma cells. Vet Comp Oncol. (2023) 21:520–32. doi: 10.1111/vco.12915
61. Sánchez, D, Cesarman-Maus, G, Romero, L, Sánchez-Verin, R, Vail, D, Guadarrama, M, et al. The NDV-MLS as an immunotherapeutic strategy for breast Cancer: proof of concept in female companion dogs with spontaneous mammary Cancer. Viruses. (2024) 16:372. doi: 10.3390/v16030372
62. Rossmeisl, JH, King, JN, Robertson, JL, Weger-Lucarelli, J, and Elankumaran, S. Phase I/II trial of Urokinase plasminogen activator-targeted oncolytic Newcastle disease virus for canine intracranial tumors. Cancers (Basel). (2024) 16:564. doi: 10.3390/cancers16030564
63. Sánchez, D, Pelayo, R, Medina, LA, Vadillo, E, Sánchez, R, Núñez, L, et al. Newcastle disease virus: potential therapeutic application for human and canine lymphoma. Viruses. (2015) 8:3. doi: 10.3390/v8010003
64. Bar-Eli, N, Giloh, H, Schlesinger, M, Zakay-Rones, Z, and Schlesinger, EM. Preferential cytotoxic effect of Newcastle disease virus on lymphoma cells. J Cancer Res Clin Oncol. (1996) 122:409–15. doi: 10.1007/BF01212880
65. Amarasinghe, GK, Bào, Y, Basler, CF, Bavari, S, Beer, M, Bejerman, N, et al. Taxonomy of the order Mononegavirales: update 2017. Arch Virol. (2017) 162:2493–504. doi: 10.1007/s00705-017-3311-7
66. Hu, H, Roth, JP, and Yu, Q. Generation of a recombinant Newcastle disease virus expressing two foreign genes for use as a multivalent vaccine and gene therapy vector. Vaccine. (2018) 36:4846–50. doi: 10.1016/j.vaccine.2018.06.055
67. Ferreira, HL, Taylor, TL, Dimitrov, KM, Sabra, M, Afonso, CL, and Suarez, DL. Virulent Newcastle disease viruses from chicken origin are more pathogenic and transmissible to chickens than viruses normally maintained in wild birds. Vet Microbiol. (2019) 235:25–34. doi: 10.1016/j.vetmic.2019.06.004
68. Alexander, DJ. Newcastle disease and other avian paramyxoviruses. Rev. Sci. Tech. (2000) 19:443–62. doi: 10.20506/rst.19.2.1231
69. De Swart, RL, and Belov, GA. Advantages and challenges of Newcastle disease virus as a vector for respiratory mucosal vaccines. Curr Opin Virol. (2023) 62:1–3. doi: 10.1016/j.coviro.2023.101348
70. Pedersden, KA, Sadasiv, EC, Chang, PW, and Yates, VJ. Detection of antibody to avian viruses in human populations. Epidemiol Infect. (1990) 104:519–25. doi: 10.1017/S095026880004752X
71. Ge, J, Wang, X, Tao, L, Wen, Z, Feng, N, Yang, S, et al. Newcastle disease virus-vectored rabies vaccine is safe, highly immunogenic, and provides long-lasting protection in dogs and cats. J Virol. (2011) 85:8241–52. doi: 10.1128/JVI.00519-11
72. Rojko, JL, Kociba, GJ, Abkowitz, JL, Hamilton, KL, Hardy, WD Jr, Ihle, JN, et al. Feline lymphomas: immunological and Cytochemical characterization 1. Cancer Res. (1989) 49:345–51.
73. Al-Garib, SO, Gielkens, ALJ, Gruys, E, Peeters, BPH, and Koch, G. Tissue tropism in the chicken embryo of non-virulent and virulent Newcastle diseases strains that express green fluorescence protein. Avian Pathol. (2003) 32:591–6. doi: 10.1080/03079450310001610695
74. Reed, LJ, and Muench, H. A simple method of estimating fifty per cent endpoints. Am J Hyg. (1938) 27:493–7.
75. Riccardi, C, and Nicoletti, I. Analysis of apoptosis by propidium iodide staining and flow cytometry. Nat Protoc. (2006) 1:1458–61. doi: 10.1038/nprot.2006.238
76. Nelson, CB, Pomeroy, BS, Schrall, K, Park, WE, and Lindeman, RJ. An outbreak of conjunctivitis due to Newcastle disease virus (NDV) occurring in poultry workers. Am J Public Health Nations Health. (1952) 42:672. doi: 10.2105/ajph.42.6.672
77. Ul-Rahman, A, Ishaq, HM, Raza, MA, and Shabbir, MZ. Zoonotic potential of Newcastle disease virus: Old and novel perspectives related to public health. Rev Med Virol. (2022) 32:e2246. doi: 10.1002/rmv.2246
78. Buijs, PRA, Van Amerongen, G, Van Nieuwkoop, S, Bestebroer, TM, Van Run, PRWA, Kuiken, T, et al. Intravenously injected Newcastle disease virus in non-human primates is safe to use for oncolytic virotherapy. Cancer Gene Ther. (2014) 21:463–71. doi: 10.1038/cgt.2014.51
79. Kristal, O, Lana, SE, Ogilvie, GK, Rand, WM, Cotter, SM, and Moore, AS. Single agent chemotherapy with doxorubicin for feline lymphoma: a retrospective study of 19 cases (1994-1997). J Vet Intern Med. (2001) 15:125–30. doi: 10.1111/j.1939-1676.2001.tb01243.x
80. Versteegh, H, Zandvliet, MMJM, Feenstra, LR, van der Steen, FEMM, and Teske, E. Feline lymphoma: patient characteristics and response outcome of the COP-protocol in cats with malignant lymphoma in the Netherlands. Animals. (2023) 13:667. doi: 10.3390/ani13162667
81. Jaroensong, T, Piamwaree, J, and Sattasathuchana, P. Effects of chemotherapy on hematological parameters and CD4+/CD8+ ratio in cats with mediastinal lymphoma and seropositive to feline leukemia virus. Animals. (2022) 12:223. doi: 10.3390/ani12030223
82. Diesel, LP, de Mello, LS, de Oliveira, SW, Ikuta, N, Fonseca, ASK, Kipper, D, et al. Epidemiological insights into feline leukemia virus infections in an urban cat (Felis catus) population from Brazil. Animals. (2024) 14:1051. doi: 10.3390/ani14071051
Keywords: cat, virotherapy, oncolytic, Newcastle Disease Virus, new therapies, cancer, in vitro , immunotherapy
Citation: Alves TGL, Xavier PLP, Garnica TK, Rochetti AL, Qazi TJ, Vendramini THA, Haddad F, Munir M, Brunetto MA and Fukumasu H (2025) Oncolytic potential of Newcastle Disease Virus in feline lymphoma cells: an in vitro evaluation. Front. Vet. Sci. 12:1484947. doi: 10.3389/fvets.2025.1484947
Edited by:
Rodrigo dos Santos Horta, Federal University of Minas Gerais, BrazilReviewed by:
Lukasz Kuryk, National Institute of Public Health, PolandCélia Lopes, University of Porto, Portugal
Copyright © 2025 Alves, Xavier, Garnica, Rochetti, Qazi, Vendramini, Haddad, Munir, Brunetto and Fukumasu. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Heidge Fukumasu, ZnVrdW1hc3VAdXNwLmJy
†Deceased