 Theophilus Odoom1,2*
Theophilus Odoom1,2* Richard Kwamena Abbiw3Joseph Kofi Abuh4Emmanuel Allegye-Cudjoe4Sherry Ama Mawuko Johnson2*William Tasiame5Daniel Arthur1Benita Anderson4
Richard Kwamena Abbiw3Joseph Kofi Abuh4Emmanuel Allegye-Cudjoe4Sherry Ama Mawuko Johnson2*William Tasiame5Daniel Arthur1Benita Anderson4 Tirumala B. K. Settypalli6
Tirumala B. K. Settypalli6 Charles E. Lamien6
Charles E. Lamien6 William G. Dundon6
William G. Dundon6- 1Accra Veterinary Laboratory, Veterinary Service Directorate, Accra, Ghana
- 2School of Veterinary Medicine, CBAS, University of Ghana, Accra, Ghana
- 3West African Centre for Cell Biology of Infectious Pathogens, University of Ghana, Accra, Ghana
- 4Veterinary Service Directorate, Accra, Ghana
- 5School of Veterinary Medicine, Kwame Nkrumah University of Science and Technology, Kumasi, Ghana
- 6Animal Production and Health Laboratory, Animal Production and Health Section, Joint FAO/IAEA Center, Department of Nuclear Sciences and Applications, International Atomic Energy Agency, Vienna, Austria
On the 24/09/2023, a video of a suspected rabid bull at a livestock market in Ghana was shared on social media and was seen by a local veterinary officer. This led to an on-site investigation by veterinary authorities on the 25/09/2023 which concluded that the bull had been bitten by a three-month old dog 4 days previously. The puppy, which was killed and buried after the bite, was subsequently exhumed, tested and confirmed positive for rabies. The bull was humanely destroyed. Brain tissue from the bull was collected and sent to the Accra Veterinary Laboratory for further analysis. RABV was confirmed by conventional RT-PCR and the full genome of the viruses from both animals were sequenced. The consensus sequences of the genomes belonging to the Africa 2 clade, were identical although sub-consensus variants in a subset of the sequences located in the RNA-dependent-RNA polymerase (L) gene of the bovine virus were observed. The implication of these findings is discussed.
1 Introduction
Rabies is a zoonotic disease caused by rabies virus (RABV) which is transmitted by the bite of infected mammals resulting in acute and generally fatal encephalitis (1). Molecular epidemiological studies have demonstrated that RABVs can be divided into dog and bat derived clusters, and subdivided into several clades, subclades and lineages (2). Dog-derived isolates belong to six well defined clades; Cosmopolitan (Africa-1 and Africa-4), Africa-2 and Africa-3 clades have been shown to be present in Africa (2, 3). Africa-1a, Africa-1b and Africa-1c are lineages of Africa-1 (4). Globally, nearly 60,000 humans die of rabies (5, 6). Dogs are integral to the transmission of RABV contributing to 9 of 10 human rabies cases. However, the virus can also infect other mammals including livestock (4–7) and wildlife (7, 8). Therefore, the impact of rabies is not limited to the loss of human lives, but poses palpable threats to livestock production and wildlife surveillance, especially in endemic areas (9–13). RABV is enzootic/endemic in Ghana and has been reported in both humans and animals including dogs, cats, pigs, goats, monkeys, sheep, bats and cattle (14–17). RABV belonging to the Africa 2 lineage and Africa 1 (a and b) sub-lineages have been identified in rabies outbreaks in Ghana (18, 19).
This case report describes the transmission of rabies virus from a puppy to a bull in a livestock market in Ghana. It provides details on the intervention of the veterinary authorities, laboratory diagnosis and molecular characterization of the causative virus. It has provided a unique opportunity to characterise and analyse epidemiologically related RABVs.
2 Case description
A video of a suspected rabid bull was seen by the Greater Accra Regional Veterinary Officer (RVO) on the 24/09/2023 (Supplementary Video S1). He immediately informed the Chief Veterinary Officer (CVO) and the Head of the Epidemiology Unit of the Veterinary Service Directorate (VSD) who later led a team of three VSD staff and the District Veterinary Officer (DVO) of the Weija-Gbawe Municipal to investigate the video taken at the Joma livestock market in Ablekuma. The team visited the site a day after the video was taken. According to the investigation, the bull (Figure 1) was found restless and profusely drooling; signs that baffled the owners and led to the videoing of the incident. It was revealed that the bull had been bitten on its nose by a three-month old unvaccinated male puppy on the 22/092023 while resting in a pen at the livestock market with 21 other cattle, 20 sheep, one goat and one horse. The dog was reported to have been overly aggressive, barking incessantly and attempted to bite people but failed. Due to the suspicion of rabies, the dog was captured, killed and buried on the 23/09/2023.

Figure 1. Photograph of the infected bull, Joma livestock market, Accra Ghana.
The buried puppy was exhumed (Supplementary Figure S1) and the brain sampled and tested for rabies using an Anigen Rapid Rabies Antigen Test Kit (Bionote Inc., Korea) on 25/09/2023. The bull was quarantined pending confirmatory laboratory diagnosis of the puppy’s brain sample. On 26/09/2024, the bull was euthanised following RT-PCR confirmation of RABV in the puppy. Contact tracing by the team identified 12 people that had come into contact with the suspected rabid animals. These individuals were directed to go for anti-rabies post exposure vaccines, while the residents and workers at the market were educated on rabies from the 25/09/20230 to 27/09/2023. The DVO was tasked by the RVO to provide regular education on rabies.
3 Diagnostic assessment
3.1 RDT
An Antigen Rapid Rabies Ag Test Kit (Bionote Inc., Korea) was used for the preliminary diagnosis of rabies in both the puppy and the bull post-mortem. The immunochromatographic assay is reported to be 96.9% sensitive and 100% specific for rabies virus. Briefly, 10% brain homogenates prepared in the PBS supplied in the kit was added to the test device and the result was read after 10 min.
3.2 RNA extraction
Two grams (2 g) of brain tissues from the bull (C2) and the puppy (D2) were homogenized in a sterile 1.5 mL Eppendorf tube. A second brain sample (D1) from a dog collected on the 28/07/2023 with reference number R/24/19 and confirmed to be positive for rabies was included for comparative purposes. Total viral RNA of the brain samples was extracted using the Qiagen RNeasy RNA mini kit according to the manufacturer’s instructions and stored at −80°C until required. The RNA extracts were used for RT-PCR at the Accra Veterinary Laboratory (AVL) and sequencing at the Animal Production and Health Laboratory, Joint FAO/IAEA Centre, Austria, for full genome sequencing.
3.3 RT-PCR
RNA samples were screened at the AVL as described by Benedictis et al. (20) using a one-step RT-PCR kit (Qiagen). Briefly, 2.5 μL of RNA was added to a master mix containing primers RabPyro Forward primer: 5′ – AACACYYCTACAATGGA – 3′ (61.4 nmol), RabPyro reverse primer 1: 5′ - TCCAATTNGCACACATTTTGTG – 3′ (66.1 nmol), RabPyro reverse primer 2: 5′ – TCCARTTAGCGCACAT YTTATG – 3′ (100.8 nmol), and RabPyro reverse primer 3: 5′ – TCCAGTTGGCRCACATCTTRTG – 3′ (97.7 nmol), 1 X RT-PCR buffer, dNTPs (10 mmol), RNase Inhibitor (1 U), enzyme mix (1 u) and RNase free water. The following cycling conditions were used: 50°C, for 30 min; 95°C for 15 min followed by 45 cycles of 94°C for 30 s, 52°C for 30 s and 72°C for 40 s with a final elongation of 72°C for 5 min. Amplicons were visualised on 1.5% (v/v) agarose gels.
3.4 Full genome sequencing by sequence-independent single primer amplification (SISPA)
RNA was reverse transcribed into cDNA using primer K-8 N (5’ GAC CAT CTA GCG ACC TCC AC -NNNNNNNN 3′) at a concentration of 2.5 μM using the SSIV First strand cDNA synthesis system Kit (Invitrogen, USA) following the manufacturer’s instructions. Residual RNA was then removed by treating with RNase H for 20 min at 37°C. Next, double stranded (ds) cDNA was synthesized using DNA polymerase I, Large (Klenow) fragment (New England Biolabs, USA) in the presence of 800 nM of K-8 N primer by polymerization at 37°C for 60 min followed by inactivation at 75°C for 10 min. PCR amplification of ds cDNA was performed using a Pfu Ultra II Fusion HS DNA polymerase Kit (Agilent Technologies, USA) with 400 nM of primer K- (5’ GAC CAT CTA GCG ACC TCC AC 3′). The thermal cycling profile was initial denaturation of 1 min at 95°C followed by 45 cycles of 20 s denaturation at 95°C, 20 s of annealing at 55°C and extension of 3 min at 72°C, ending with a final extension of 3 min at 72°C. The amplified ds cDNA was purified using 1.8X AMPure XP beads (Beckman, USA).
The purified ds cDNA was then used for library preparation to run on an S5 Next generation sequencing system (Ion Torrent, Thermo Fisher). Approximately 50-100 ng of the ds cDNA were enzymatically fragmented to a length of 200 bp using Ion shear Plus reagents (Thermo Fisher Scientific, USA) with an optimized shearing time of 13 min followed by purification using 1.8X AMPure XP beads. The purified and fragmented DNA was ligated with adapters, barcodes to prepare barcoded libraries using an Ion Xpress™ Plus Fragment Library Kit and Ion Xpress barcode adapters (Thermo Fisher Scientific, USA). The libraries were purified using 1.2X AMPure XP beads and were then size selected on a Pippin Prep platform (Sage Science, Inc., USA) followed by a further purification step using 1.5X AMPure XP beads. The purified size selected libraries were subjected to 8 cycles of amplification using a Platinum™ PCR Super Mix High Fidelity kit (Thermo Fisher Scientific, USA) and purified using 1.2X AMPure XP beads. The purified amplified barcoded libraries were pooled at a concentration of 100 pM in equal volumes and loaded on to an Ion Chef™ instrument (Thermo Fisher Scientific, USA) for automated template preparation and chip loading using the Ion 540™ Kit-Chef (Thermo Fisher Scientific, USA). With Ion Chef™ the pooled libraries were clonally amplified on Ion Spheres™ (ISPs) by emulsion PCR followed by automated loading of template enriched ISPs on to an Ion 540™ chip (Thermo Fisher Scientific, USA). The sequencing was performed on an Ion S5 next generation sequencing system with 500 flows to generate 200 bp reads.
The raw sequences were cleaned to remove low-quality reads (Phred < 20), SISPA adapters, and short reads (<50 bp) using fastq-mcf v1.04.676 (ea-utils) and cutadapt v3.5. The quality of the reads was assessed with FastQC (v0.11.5). De Novo Assemblies were performed using Megahit (v1.2.9) and Spades (v3.15.5). Using the De Novo Assembly’s contigs, BLAST searches were performed, which identified complete genome (GenBank: MW057694), as the appropriate reference. After mapping the cleaned raw reads against the reference sequence using BWA (v0.7.17), SAMtools (v1.11) was used to generate Mpileup files and perform variant calling using BCFtools (v1.9), filtering only variants with a mapping quality > 20 and a minimum coverage depth of 100. The consensus sequences, from reads with a mapping quality > 20, were produced using vcfutils.pl. (VCFtools v0.1.16) and seqtk (v1.3.106) and compared to the De Novo assemblies. The mapping quality and coverage were assessed with Qualimap v2.3 (31). Additionally, the bam files were loaded into Integrative Genomics Viewer (IGV software version 2.3; The Broad Institute, Cambridge, MA) for read visualization.
3.5 Phylogenetic analysis
Phylogenetic trees using representative sequences from GenBank were estimated using the maximum-likelihood (ML) method available in MEGA 6 (21), employing the Tamura-Nei model of nucleotide substitution and 1,000 bootstrap replications.
3.6 Analysis of sub-consensus variants
To analyse the viral subpopulations in the cattle and dog samples, reads mapping to the targeted region with evidence of a mixed population (bp position: 11315–11,565 as suggested by the IGV visualization) were extracted from the sorted BAM file using Samtools, selecting only alignments with a mapping quality higher than 50 and supplying a BED file of the targeted region. The extracted reads were converted to FASTQ using Samtools’ bam2fq and trimmed with Cutadapt to keep only reads of lengths between 170 and 250 bp. Following the conversion of the reads from FASTQ to FASTA using Seqtk, the reads were aligned using MAFFT.
The MAFFT alignment was manually inspected to keep reads that were fully overlapping a 120 bp region of the targeted area and to filter out reads that were represented less than 3 times. Sequences from a similar region in publicly available rabies sequences were also retrieved from GenBank and merged with the dataset of the current study. Following a new alignment with MAFFT, only sequence variants that were represented 2 times or more were kept. Additionally, the sequences were translated into amino acids and reads that included truncated or incorrect amino acid sequences were excluded from further analysis. The final dataset consisted of 590 sequences.
3.7 Discriminant analysis of principal components (DAPC)
After manual inspections of the MAFFT-aligned sequences, the alignment file was imported into the ‘adegenet’ package in R for SNP analysis. Initially, the alignment file was converted into a ‘genlight’ object using the ‘fasta2genlight’ function, and the SNPs were extracted. The distribution and density of the SNPs among RABVs sequences were visualized using a heatmap. Additionally, discriminant analysis of principal components (DAPC) was conducted on the extracted SNPs to identify different clusters of RABVs, which were represented through PCA scatterplots.
4 Results
4.1 RDT and RT-PCR
The brain samples from both the puppy and the bull were positive by RDT and RT-PCR (See Supplementary Figure S2).
4.2 Full genome and phylogenetic analysis
Genomes of 11,886 bp, were resolved for D1, D2 and C2. The mean coverage for D1, D2 and C2 were 1823, 9,132 and 514, respectively. All the ORFs for the nucleoprotein (N), phosphoprotein (P), matrix protein (M), glycoprotein (G), and RNA-dependent RNA polymerase (L) were identified. In the 3′ and 5′ untranslated regions (UTRs) of the genomes, 23 and 14 nucleotides were unresolved, respectively.
Phylogenetic analysis revealed that the genome sequences of D2 and C2 were identical to each other and that they clustered within the Africa-2 clade along with other RABVs from the region including Niger and Burkina Faso (Figure 2).
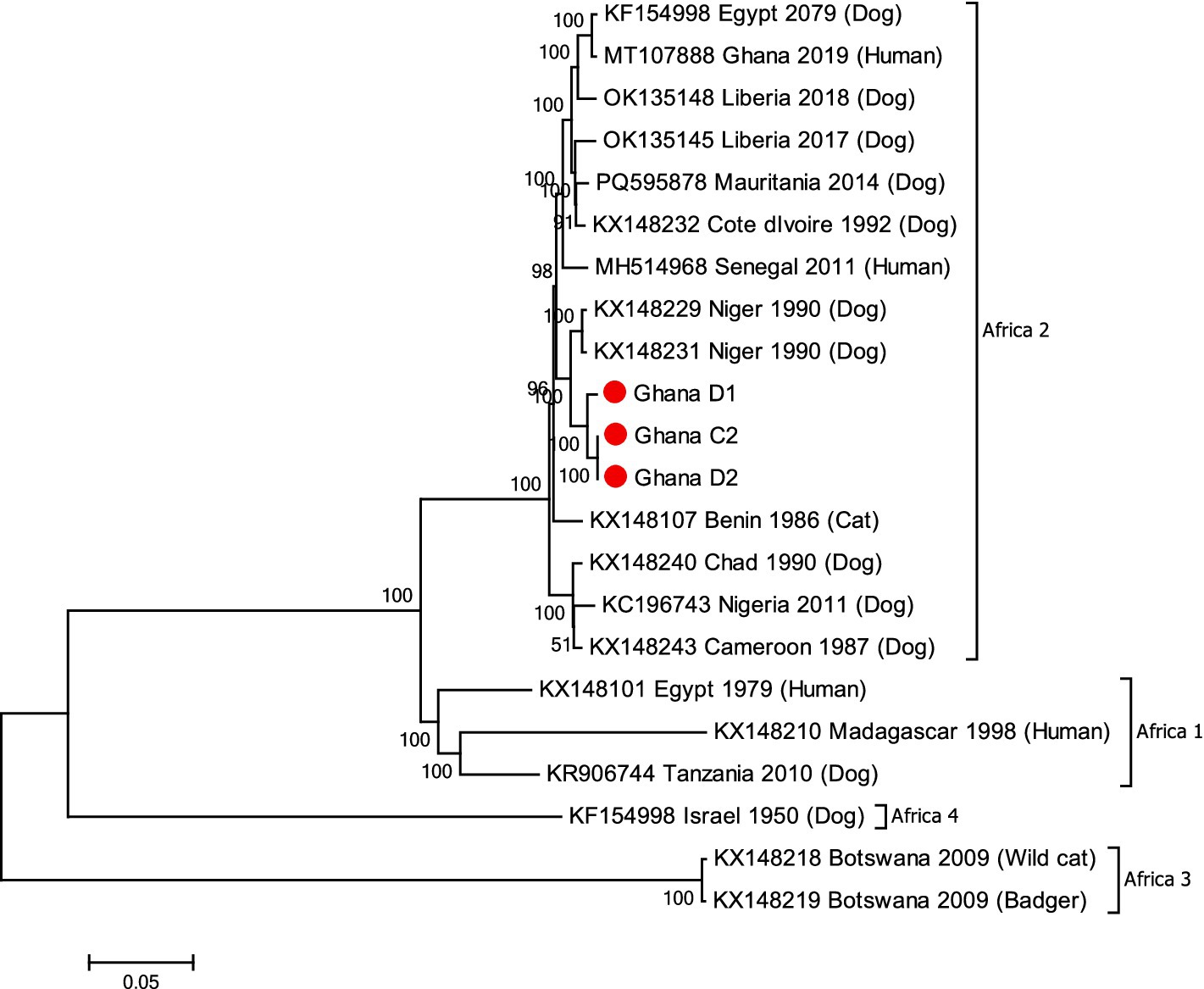
Figure 2. M-L phylogenetic tree generated using the genomes from the RABV positive samples and other RABV genomes available in GenBank (Supplementary Table S1). Comparative dog sample (D1), the puppy (D2) and bull sample (C2) are shown with red circles. Clades and bootstrap values from 1,000 replicates are indicated.
Further analysis of the sequences of C2 identified nucleotide differences in a sub-population of viral sequences (Supplementary Figure S3). Following the quality control of the region containing the SCVs there were 32 sequences available for alignment (see also Supplementary Figure S3). Of these 4 (12.5%) sequences contained the SCVs. These sub-consensus variants (SCV) were in the RNA-dependent-RNA polymerase (L) gene between region (bp 11,315–11,565) and were all synonymous mutations (Supplementary Figure S4).
4.3 PCA analysis
The PCA analysis of the genomic region where the SCVs were observed in 590 RABVs available in GenBank, revealed four clearly distinct clusters, with cluster 4 containing sequences of RABVs of African origin exclusively (Supplementary Figure S5). The consensus sequences of D2 and C2 were located in cluster 4. In contrast, the SCVs seen in the C2 sample were located in cluster 2 together with sequences from RABVs originating in Asia and North America.
5 Discussion
The devastating impact of rabies requires an expedited approach to handling outbreaks. Therefore, the speed with which an investigative team was assembled and dispatched to the site is commendable and this action averted a potentially devastating outbreak or spillover into humans. The case, however, highlights the inadequate knowledge of the public on bovine rabies. Thus, while rabies in the puppy was easily recognised, the virus in the bull was not. Perhaps, its rarity compared to canine rabies [only 1 recorded case in Ghana between 2010 and 2017 (14)] might explain the lack of public awareness of bovine rabies. The follow up public education was therefore an important aspect of the outbreak management.
Certain combinations of neurologic and generalized signs, albeit not pathognomonic, are highly suggestive of rabies. For instance, the drastic behavioral changes, butter-like hypersalivation, vocalization, confusion, gait asymmetry, dyspnoea and muscle tremors observed in the rabid bull have been associated with rabies (1, 5, 6, 11, 22–25). Depending on the site of viral introduction, the incubation of bovine rabies is between 8 and 30 days (23, 25). The short incubation observed in this outbreak could be attributed to the proximity of the puppy’s bite (on the muzzle) to the brain and, possibly, a high viral load.
The implicated RABV in this outbreak belonged to the Africa 2 lineage. The predominance of the Africa 2 lineage in West Africa had been attributed to free movement, trade and consumption of dogs within the sub-region (18). The availability of well-defined field samples in this study provided a unique opportunity to investigate the virus at the complete genomic level. The identification and analysis of variants within a viral population may identify nucleotides that have the potential to be selected if environmental conditions change (26) including adaptation to different hosts (27). Although, the consensus genome sequences of the RABV from the dog and bull samples in this current study were identical, SCVs were observed in the L gene of the RABV genome from the bull. Interpreting these SCVs is difficult given that they consisted of synonymous mutations that did not alter the amino acid sequence of the polymerase. However, PCA analysis indicated that the SCVs identified do not appear to be random or unique to the virus from this study since they have been identified in other RABVs albeit collected in different continents although there is no evidence of any epidemiological link between RABVs from Africa and those from Asia and the Americas. It is tempting to think that the SCVs might have become more dominant in the mixed viral population if the virus in the bull had more time to evolve. As it stood, the bull developed symptoms quickly due to the location of the bite and was destroyed within 4 days of being infected.
The responding team failure to screen other animals for rabies in the outbreak area was a major limitation of the on-site investigation. According to the investigators, this was due to the limited number of RDT kits available for screening. This lack of suitable diagnostic kits could have had serious public health implications due to the mixed species present in the livestock market (e.g., sheep, goat, and horse) and the limited experience of the general public with rabies presentation in these animals.
Livestock rabies has significant public health, economic and food security implications in many endemic countries; some of which have adopted livestock vaccination (10, 13, 28–30). Currently, Ghana neither vaccinates against nor prioritises livestock rabies possibly due to the cost of anti-rabies vaccines, and the relatively lower incidence of livestock rabies. Nevertheless, it will be beneficial for Ghana, like many other endemic countries, to consider an extensive public education on rabies, including bovine rabies.
Data availability statement
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found in the article/Supplementary material.
Ethics statement
Ethical approval was not required for the studies involving animals in accordance with the local legislation and institutional requirements because the animal involved was showing signs of rabies and the veterinary service directorate is mandated to control rabies. This allow Veterinary services Directorate to investigate and conduct research into rabies cases without obtaining ethical clearance from the owner. Written informed consent was not obtained from the owners for the participation of their animals in this study because the owner of the animal reported to Veterinary Services about the animal behavior and permitted Veterinary to investigate and conducted research into the behavior of his animal. Written informed consent was obtained from the participants for the publication of this case report.
Author contributions
TO: Conceptualization, Funding acquisition, Investigation, Methodology, Project administration, Resources, Supervision, Validation, Visualization, Writing – original draft, Writing – review & editing. RA: Conceptualization, Data curation, Methodology, Writing – original draft, Writing – review & editing. JA: Investigation, Resources, Supervision, Writing – review & editing. EA-C: Resources, Supervision, Project administration, Writing – review & editing. SJ: Project administration, Supervision, Writing – review & editing. WT: Writing – review & editing, Validation, Project administration, Supervision. DA: Validation, Methodology, Writing – review & editing, Investigation. BA: Writing – review & editing, Supervision, Resources. TS: Writing – review & editing, Software, Visualization, Formal analysis, Data curation. CL: Formal analysis, Validation, Methodology, Writing – review & editing, Resources, Data curation, Software, Funding acquisition. WD: Resources, Formal analysis, Writing – review & editing, Funding acquisition, Methodology, Validation, Conceptualization, Software, Data curation.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. The work reported herein was supported by the IAEA Peaceful Uses Initiative (PUI) VETLAB Network and the IAEA ZODIAC Project. Some of the sequences were generated through the Sequencing Services of the Animal Production and Health sub-programme of the Joint FAO/IAEA Division. Veterinary Services Directorate also provided funding for the outbreak investigation in terms of providing vehicle, euthanasia, proper disposal of animal, diagnostic support.
Acknowledgments
The veterinary team that attended to the case (Patrick Tetteh Ababio, Daniel Tawiah Yingar, Daniel Baah, Elisha Apatewen Akanbong, and Spencer Dugbartey), the owners of the puppy and the bull, and the videographer are acknowledged.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Gen AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fvets.2025.1524562/full#supplementary-material
References
1. Vuta, V, Picard-Meyer, E, Robardet, E, Barboi, G, Motiu, R, Barbuceanu, F, et al. Vaccine-induced rabies case in a cow (Bos taurus): molecular characterisation of vaccine strain in brain tissue. Vaccine. (2016) 34:5021–5. doi: 10.1016/j.vaccine.2016.08.013
2. Troupin, C, Dacheux, L, Tanguy, M, Sabeta, C, Blanc, H, Bouchier, C, et al. Large-scale Phylogenomic analysis reveals the complex evolutionary history of rabies virus in multiple carnivore hosts. PLoS Pathog. (2016) 12:1–20. doi: 10.1371/journal.ppat.1006041
3. Talbi, C, Holmes, EC, de Benedictis, P, Faye, O, Nakouné, E, Gamatié, D, et al. Evolutionary history and dynamics of dog rabies virus in western and Central Africa. J Gen Virol. (2009) 90:783–91. doi: 10.1099/vir.0.007765-0
4. Noel, J, Wobessi, S, Kenmoe, S, Mahamat, G, Thierry, J, Belobo, E, et al. Incidence and seroprevalence of rabies virus in humans, dogs and other animal species in Africa, a systematic review and meta-analysis. One Health. (2021) 13:100285. doi: 10.1016/j.onehlt.2021.100285
5. Sharif, M, Arhaiem, A, Giadan, O, Adam, A, Abdalla, F, Dayhum, A, et al. Rabies in bovine: first case report of rabies in Al Jabal Al Akhdar, Libya. Open Vet J. (2021) 11:96–9. doi: 10.4314/ovj.v11i1.14
6. Tang, HB, He, XX, Zhong, YZ, Liao, SH, Zhong, TZ, Xie, LJ, et al. Complete genome sequence of a rabies virus isolate from cattle in Guangxi, southern China. Genome Announc. (2013) 1:3–4. doi: 10.1128/genomeA.00137-12
7. Ma, X, Bonaparte, S, Toro, M, Orciari, LA, Gigante, CM, Kirby, JD, et al. Rabies surveillance in the United States during 2020. J Am Vet Med Assoc. (2022) 260:1157–65. doi: 10.2460/javma.22.03.0112
8. Feng, Y, Ma, J, Sun, S, Chi, L, Kou, Z, and Tu, C. Epidemiology of animal rabies — China, 2010–2020. China CDC Wkly. (2021) 3:815–8. doi: 10.46234/ccdcw2021.202
9. Beyene, TJ, Fitzpatrick, MC, Galvani, AP, Mourits, MCM, Revie, CW, Cernicchiaro, N, et al. Impact of one-health framework on vaccination cost-effectiveness: a case study of rabies in Ethiopia. One Health. (2019) 8:100103. doi: 10.1016/j.onehlt.2019.100103
10. Ulloa-Stanojlovic, FM, and Dias, RA. Spatio-temporal description of bovine rabies cases in Peru, 2003–2017. Transbound Emerg Dis. (2020) 67:1688–96. doi: 10.1111/tbed.13512
11. Lu, T, Cao, JMD, Rahman, AKMA, Islam, SS, Sufian, MA, and Martínez-López, B. Risk mapping and risk factors analysis of rabies in livestock in Bangladesh using national-level passive surveillance data. Prev Vet Med. (2023) 219:106016. doi: 10.1016/j.prevetmed.2023.106016
12. Müller, T, Hassel, R, Jago, M, Khaiseb, S, van der Westhuizen, J, Vos, A, et al. Rabies in kudu: Revisited. Adv Virus Res. (2022) 112:115–73. doi: 10.1016/bs.aivir.2022.04.001
13. Vos, A, Ün, H, Hampson, K, De Balogh, K, Aylan, O, Freuling, CM, et al. Bovine rabies in Turkey: patterns of infection and implications for costs and control. Epidemiol Infect. (2014) 142:1925–33. doi: 10.1017/S0950268813002811
14. Turkson, PK. Spatio-temporal investigation of reported cases of animal rabies in Ghana from 2010 to 2017. Vet Med Sci. (2023) 9:2559–65. doi: 10.1002/vms3.1282
15. Dsane-Aidoo, PH, Sarpong, K, Odei, E, Senya, K, Kuffour, AB, Andoh, PK, et al. Outbreak of rabies among humans, dogs and livestock, Awutu-Senya east municipality, Ghana, 2019. J Interv Epidemiol Public Health. (2022) 5:1–9. doi: 10.37432/jieph.2022.5.2.57
16. Punguyire, DT, Osei-Tutu, A, Aleser, EV, and Letsa, T. Level and pattern of human rabies and dog bites in Techiman municipality in the Middle Belt of Ghana: a six year retrospective records review. Pan Afr Med J. (2017) 28:281. doi: 10.11604/pamj.2017.28.281.14218
17. Amissah-Reynolds, PK. Zoonotic risks from domestic animals in Ghana. Int J Pathog Res. (2020) 4:17–31. doi: 10.9734/ijpr/2020/v4i330113
18. Tasiame, W, El-Duah, P, Johnson, SAM, Owiredu, EW, Bleicker, T, Veith, T, et al. Rabies virus in slaughtered dogs for meat consumption in Ghana: a potential risk for rabies transmission. Transbound Emerg Dis. (2022) 69:e71–81. doi: 10.1111/tbed.14266
19. Hayman, DTS, Johnson, N, Horton, DL, Hedge, J, Wakeley, PR, Banyard, AC, et al. Evolutionary history of rabies in Ghana. PLoS Negl Trop Dis. (2011) 5:e1001. doi: 10.1371/journal.pntd.0001001
20. De Benedictis, P, De Battisti, C, Dacheux, L, Marciano, S, Ormelli, S, Salomoni, A, et al. Lyssavirus detection and typing using pyrosequencing. J Clin Microbiol. (2011) 49:1932–8. doi: 10.1128/JCM.02015-10
21. Tamura, K, Stecher, G, Peterson, D, Filipski, A, and Kumar, S. MEGA6: molecular evolutionary genetics analysis version 6.0. Mol Biol Evol. (2013) 30:2725–9. doi: 10.1093/molbev/mst197
22. Kalthoum, S, Guesmi, K, Gharbi, R, Baccar, MN, Seghaier, C, Zrelli, M, et al. Temporal and spatial distributions of animal and human rabies cases during 2012 and 2018, in Tunisia. Vet Med Sci. (2021) 7:686–96. doi: 10.1002/vms3.438
23. Bonilla-Aldana, DK, Jimenez-Diaz, SD, Barboza, JJ, and Rodriguez-Morales, AJ. Mapping the spatiotemporal distribution of bovine rabies in Colombia, 2005–2019. Trop Med Infect Dis. (2022) 7:406. doi: 10.3390/tropicalmed7120406
24. Sharma, AK, Prashar, P, Bharti, OK, Thachamvally, R, Thakur, R, Kumar, N, et al. Diagnosis of rabies using reverse-transcription polymerase chain reaction on post-mortem skin tissue specimens of the nasolabial plate in a rabies suspected cow: a case study. J Vet Med Sci. (2023) 85:844–8. doi: 10.1292/jvms.23-0105
25. Hudson, LC, Weinstock, D, Jordan, T, and Bold-Fletcher, NO. Clinical features of experimentally induced rabies in cattle and sheep. J Veterinary Med Ser B. (1996) 43:85–95. doi: 10.1111/j.1439-0450.1996.tb00292.x
26. Fisher, CR, Streicker, DG, and Schnell, MJ. The spread and evolution of rabies virus: conquering new frontiers. Nat Rev Microbiol. (2018) 16:241–55. doi: 10.1038/nrmicro.2018.11
27. Borucki, MK, Chen-Harris, H, Lao, V, Vanier, G, Wadford, DA, Messenger, S, et al. Ultra-deep sequencing of intra-host rabies virus populations during cross-species transmission. PLoS Negl Trop Dis. (2013) 7:e2555. doi: 10.1371/journal.pntd.0002555
28. Filho, OA, Megid, J, Geronutti, L, Ratti, J, Kataoka, APAG, and Martorelli, LFA. Importance of antirabies revaccination for an adequate antirabies protection in bovine newborns. Clin Vaccine Immunol. (2010) 17:1159–61. doi: 10.1128/CVI.00119-10
29. Benavides, JA, Velasco-Villa, A, Godino, LC, Satheshkumar, PS, Ruby, N, Rojas-Paniagua, E, et al. Abortive vampire bat rabies infections in Peruvian peridomestic livestock. PLoS Negl Trop Dis. (2020) 14:1–13. doi: 10.1371/journal.pntd.0008194
30. Johnson, SAM, Odoom, T, Abubakari, RA, Oni, AA, Amoako, KK, Osei, G, et al. Rabies outbreak on a goat farm in a suburb of Cape Coast, Ghana: an epidemiologic investigation. IJID One Health. (2025) 7:100070. doi: 10.1016/j.ijidoh.2025.100070
Keywords: rabies, bull, dog, Ghana, Africa 2 lineage, phylogeny
Citation: Odoom T, Abbiw RK, Abuh JK, Allegye-Cudjoe E, Johnson SAM, Tasiame W, Arthur D, Anderson B, Settypalli TBK, Lamien CE and Dundon WG (2025) Case Report: Molecular characterization of rabies virus transmitted from a dog to a bull in a livestock market in Ghana. Front. Vet. Sci. 12:1524562. doi: 10.3389/fvets.2025.1524562
Edited by:
Francisca Di Pillo, Universidad de las Américas, ChileReviewed by:
Raul Alejandro Alegria, University of Chile, ChileWaqas Ahmad, University of Veterinary and Animal Sciences, Pakistan
Copyright © 2025 Odoom, Abbiw, Abuh, Allegye-Cudjoe, Johnson, Tasiame, Arthur, Anderson, Settypalli, Lamien and Dundon. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Sherry Ama Mawuko Johnson, c2Fqb2huc29uQHVnLmVkdS5naA==; Theophilus Odoom, dGhlb2Rvb21AeWFob28uY29t