 Emilia Radulovic1,2,3†
Emilia Radulovic1,2,3† Kemal Mehinagic1,2,3†
Kemal Mehinagic1,2,3† Tsering Monika Wüthrich3,4
Tsering Monika Wüthrich3,4 Markus Hilty4
Markus Hilty4 Artur Summerfield1,2,5
Artur Summerfield1,2,5 Nicolas Ruggli1,2‡
Nicolas Ruggli1,2‡ Charaf Benarafa1,2,5*‡
Charaf Benarafa1,2,5*‡- 1Institute of Virology and Immunology IVI, Mittelhäusern, Switzerland
- 2Department of Infectious Diseases and Pathobiology, Vetsuisse Faculty, University of Bern, Bern, Switzerland
- 3Graduate School for Cellular and Biomedical Sciences, University of Bern, Bern, Switzerland
- 4Institute for Infectious Diseases, University of Bern, Bern, Switzerland
- 5Multidisciplinary Center for Infectious Diseases (MCID), University of Bern, Bern, Switzerland
African swine fever virus (ASFV) is a major threat for pig health and meat production in many countries. The development and commercialization of vaccine candidates are complicated by efficacy and safety concerns. Improved vaccine design requires further studies to identify factors that regulate immune responses to vaccines leading to protective immunity against a virulent challenge. In a previous study, we reported that infection with the moderately virulent ASFV field strain Estonia 2014 was less severe in specific pathogen-free (SPF) pigs than in conventional farm pigs, which differ in their gut microbiome and their basal immune activation status. As shown previously using intramuscular infection, SPF pigs were more resilient to oronasal infection with the ASFV Estonia 2014 strain compared to farm pigs, which showed increased fever and clinical signs. All SPF and farm pigs nevertheless survived the infection and remained viremic for approximately 4 months. When all animals had no detectable viremia, both groups were rechallenged with the virulent ASFV Armenia 2008 strain. SPF pigs were fully protected against disease and showed little or no viremia upon re-challenge. In contrast, farm pigs developed high viremia, high proinflammatory cytokine responses, severe clinical signs, and 40% (2 of 5 pigs) reached humane endpoints. Our findings suggest that limited prior immune exposure to other pathogens and/or the microbiome composition of SPF pigs promotes resilience to infection with a moderately virulent strain such as Estonia 2014, and importantly promotes the development of a strong protective immune response against a second challenge with a virulent ASFV strain. In conclusion, testing safety and efficacy of live attenuated vaccine candidates should take into account the specific hygiene conditions and the associated changes of general immune status of pigs in clinical trials.
Introduction
African swine fever virus (ASFV) causes a severe hemorrhagic fever in domestic pigs and wild boars. ASFV was first described in Africa (1), where it continues to cause sporadic outbreaks in domestic pigs (2). Since 2007, a highly virulent genotype II strain is the cause of the largest recorded outbreak of African swine fever (ASF) that is now a global challenge (3–5). Depending on the strain virulence, the age of the animals, and the infectious dose received, infected pigs can die suddenly without showing any prior symptoms due to severe internal bleeding, organ failure, or other complications. Frequently, ASF presents as an acute disease with skin hyperemia, petechiae, and hemorrhagic diarrhea accompanying the febrile symptoms, with death occurring within 7–10 days (6).
To effectively combat ASFV and mitigate its impact, region-specific and global control strategies must be implement including robust disease surveillance, biosecurity measures in farms, implementation of control zones, movement and trade restrictions of animals and meat products, and global cooperation addressing both domestic and wild swine populations (2, 7). An effective and safe vaccine against ASFV would be an integral part of the global strategy against this disease if it were to become available. However, few experimental vaccines induce protection against an experimental challenge and the mechanisms associated with protection are not well defined. Live attenuated vaccine (LAV) candidates that are based on deletion of putative virulence factors have shown efficacy against homologous challenge, but concerns about residual pathogenicity, virulence reversion, recombination with circulating strains, and lack of a DIVA marker are major hurdles for the licensing of LAV candidates (8–10). In addition, at higher doses LAVs have been reported to induce clinical disease and, paradoxically can also be associated with a reduced rate of protection (11).
We have previously shown that pigs raised under specific pathogen-free (SPF) conditions at the Institute of Virology and Immunology (IVI) or raised in a conventional pig farm are equally highly susceptible to an intramuscular challenge with the virulent ASFV Armenia 2008 strain and succumb to the disease within 6 days. In contrast, the SPF pigs developed a markedly milder and shorter disease form of ASF than the farm pigs following intramuscular infection with the moderately virulent ASFV Estonia 2014 strain (12). Profound differences in gut microbiota composition were observed between the two groups, and the SPF pigs had a lower steady state body temperature than the farm pigs. Importantly, the baseline immune status of the SPF pigs was characterized by a lower level of inflammatory and immune activation and the SPF pigs had lower numbers of circulating immune cells, particularly neutrophils and lymphocytes. Blood cell transcriptomic profiles prior to infection revealed that the farm pigs had upregulated transcriptional modules associated with immune cell proliferation, pro-inflammatory pathways, dendritic cell activation, type I interferon signaling, and lymphocyte proliferation. These findings indicate, not surprisingly, that farm pigs are exposed to a more immune-activating environment including commensals, pathogens, and their metabolites compared to our SPF pigs. Here, we set out to investigate whether prior environmental exposure and the associated changes in the immune system in SPF and farm pigs had an impact on the development of a protective immunity induced by an oronasal infection with ASFV Estonia 2014 that was tested with a subsequent lethal oronasal challenge with ASFV Armenia 2008.
Materials and methods
Animals
Thirteen male (castrated) and female Large White domestic pigs, 10–11 week-old with a body weight of 20–25 kg were obtained from the IVI SPF breeding facility (n = 5) or a local farm (n = 8). The pigs from the IVI SPF facility are not vaccinated and are negative for the porcine viral and bacterial pathogens listed in Table 1. Access to the SPF facility is restricted to animal caretakers and includes a showering airlock at the entrance. The SPF facility is supplied with filtered air at positive pressure and pigs are fed X-Ray-irradiated (>10 kGy) pig pellet diet (Granovit AG, KLIBA NAFAG, Kaiseraugst, Switzerland) and autoclaved hay and straw. No animal has been introduced in the facility since the initial colonization in 1993 and no clinical signs of infectious disease (diarrhea, abortion, respiratory symptoms) were observed during this period. The genetic pool is maintained at a similar status as current Swiss/European Large White pig farms via artificial insemination using semen from SUISAG (Sempach, Switzerland). The commercial farm pigs were from a high standard breeding and fattening pig farm and were vaccinated against porcine circovirus 2, Escherichia coli, and Lawsonia intracellularis. The SPF and farm pigs were housed in the BSL-3Ag containment facilities for the whole duration of the study. During the trials, all animals were fed a commercial pig pellet diet with hay supplementation and water ad libitum. The animals were maintained in a 13:11 h light cycle. Infections, body temperature measurements, clinical scores, and sampling were performed at Zeitgeber time ZT 2-4. The animals were euthanized by electrical stunning followed by exsanguination.
Table 1. List of pathogens excluded from the IVI SPF pigs.
ASFV infections in vivo
The SPF and farm pigs were acclimatized for 5 days in two separate BSL-3Ag stables. The pigs were infected oronasally with ~5 × 109 genome equivalents (gEq) using a syringe with a 5 cm rubber tubing. The inoculum was sprayed equally in one nostril (2.5 ml) and in the back of the mouth (2.5 ml) while the head of the animal was held up. The ASFV Estonia 2014 strain inoculum was a pig blood sample from a previous experiment (12) containing ~1 × 109 gEq/ml determined by qPCR as described below. When viremia was no longer detectable by qPCR, five surviving pigs of each group were re-challenged oronasally with ~106 TCID50 of the highly virulent ASFV Armenia 2008 at 164 days post immunization (dpi), which corresponds to 0 day post-challenge (dpc) (Figure 1A). A larger group of farm pigs (n = 8) than SPF pigs (n = 5) was used because we anticipated some lethality in this group after the first infection. However, all animals from both groups survived and three farm pigs were euthanized prior to the second infection to match the number of SPF pigs. Due to the low number of animals, we euthanized three farm pigs (#3, #6, #8) which were representative of the group including both moderate and high clinical scores after the first infection.
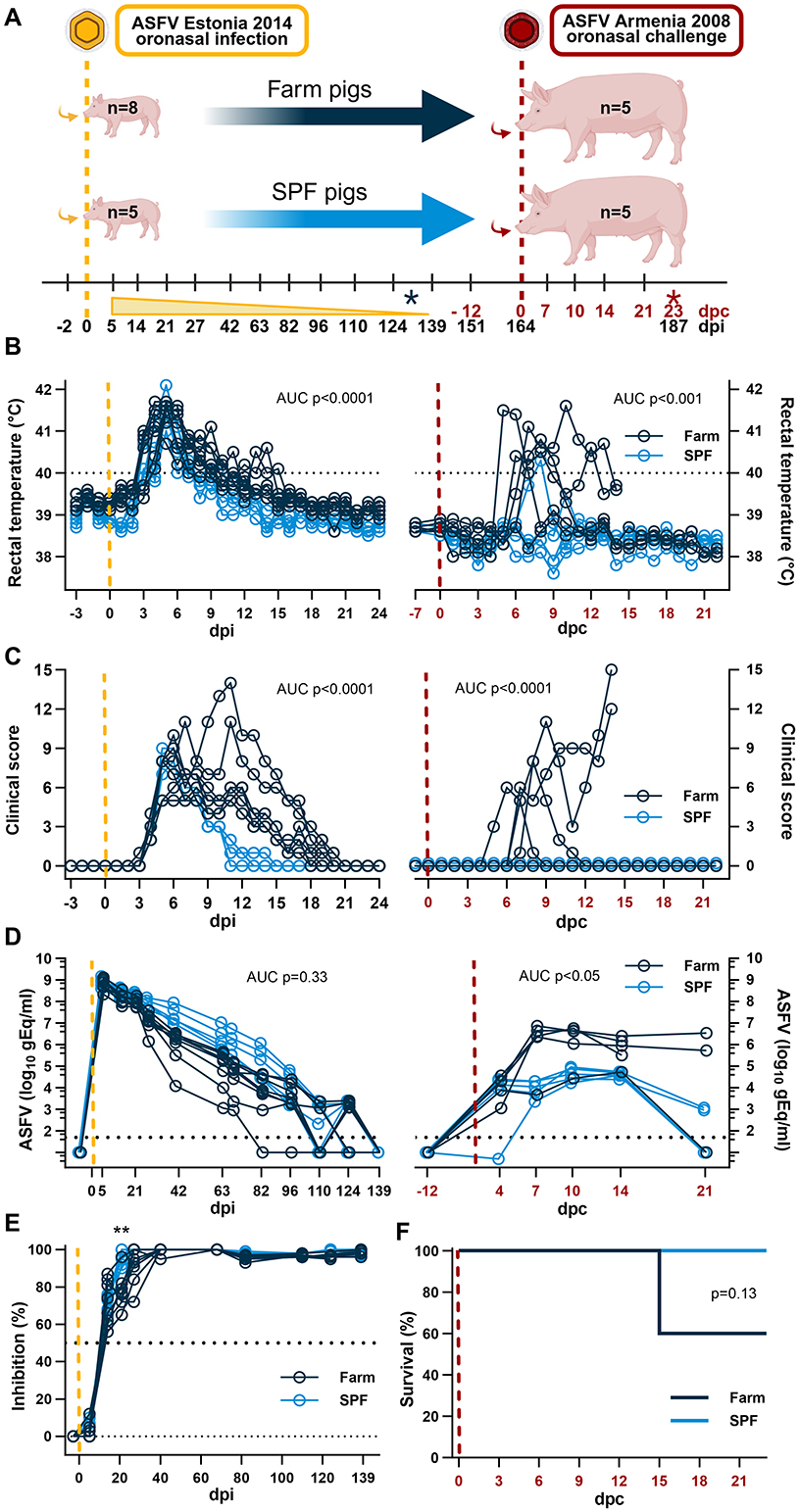
Figure 1. Experimental design and clinical parameters. (A) Diagram of experimental setup. SPF and farm pigs were inoculated oronasally with the ASFV Estonia 2014 (yellow) and re-challenged oronasally 164 days later with the Armenia 2008 strain (red). Blood samples were taken at the indicated time points and clinical parameters were measured daily in the first 3 weeks after each infection and once a week thereafter. The time points indicated in black refer to days post infection (dpi) with the Estonia 2014 strain infection; the time points in red refer to days post challenge (dpc) with the Armenia 2008 strain. The black asterisk indicates euthanasia of 3 farm pigs in excess before the second challenge. The red asterisk indicated the end of the experimentation after the second challenge. (B) Rectal temperature. (C) Clinical score. (D) Virus copy numbers determined in EDTA blood by qPCR. (E) Seroconversion determined using a competitive anti-p72 ELISA after the ASFV Estonia 2014 strain infection. (F) Survival curve after the Armenia 2008 strain challenge. (A–F) Yellow dashed lines represent the inoculation with the ASFV Estonia 2014 strain; the red dashed lines represent the inoculation with the ASFV Armenia 2008 strain. (B–D) Differences between groups were analyzed by unpaired t-test comparing the area under the curve (AUC) between the two groups after each infection. (E) Differences in antibody levels were analyzed by two-way ANOVA (mixed model) with Sidak's multiple comparisons test; **p < 0.01. (F) Differences in survival were analyzed by Log-rank (Mantel-Cox) analysis. (A) Created in BioRender. https://BioRender.com/5wxbb8e.
Body temperature and clinical parameters were assessed daily by veterinarians (KM, NR, CB) based on an adapted clinical score checklist described previously for experimental classical swine fever virus infections (13). The re-challenged pigs were euthanized at the latest on 187 dpi (corresponding to 23 dpc) unless discontinuation criteria were reached before. A cumulative score of 18 and/or a single score of 3 in any of the following parameters: liveliness, body tension, breathing, walking, or skin, were defined as the humane endpoints for discontinuation of the experiment for each pig. Blood samples were collected 1–3 days prior to infection and on 5, 14, 21, 27, 40, 63, 68, 82, 96, 110, 124, 139, 151, 168 (4 dpc), 171 (7 dpc), 174 (10 dpc), 178 (14 dpc), and 185 dpi (21 dpc), or until discontinuation criteria were reached. A full necropsy was performed on all animals; whole blood (EDTA), serum, and organs were collected for virus quantification and titration.
Virus stocks and quantification
The original stocks of genotype II ASFV strains Estonia 2014 (Genbank accession number LS478113.1) and Armenia 2008 were generously provided by Sandra Blome and Martin Beer, Friedrich-Loeffler-Institut, Greifswald-Insel Riems, Germany. Virus titration was determined by indirect immunofluorescence assay in immortalized porcine kidney macrophage (IPKM) cells (14), a generous gift of Takehiro Kokuho, National Agriculture and Food Research Organization (NARO), Tokyo, Japan. Positive cells were counted, and the titer was calculated using the Reed-Muench method.
For qPCR, DNA was extracted using the NucleoMag VET kit (Macherey-Nagel) and the KingFisher extraction platform following manufacturers' instructions. All nucleic acid extractions were performed with 200 μl of either serum, whole blood, or organ homogenates in RA1 lysis buffer adjusted to contain 5 mg of tissue. Subsequently, qPCR for the B646L gene (p72) was performed in triplicates for samples and standards to determine genome equivalents (gEq) as described previously (12).
Hematology and flow cytometry
Differential blood cell counts were determined from EDTA blood samples using an automated hematology analyzer (VetScan HM5, Abaxis). The percentage of leukocyte subsets were determined by flow cytometry and absolute subset counts were calculated using white blood cell (WBC) values from the hematology analyzer. Whole blood (100 μl) was incubated with two sets of antibodies for myeloid and lymphocyte subsets. Data acquisition (100,000 single-cell events) was done on a BD FACS Canto II (BD Bioscience), and data analyzed with FlowJo v10. The gating strategy is described in Supplementary Figure 1. In brief, single cells were determined by forward scatter (FSC) and side scatter (SSC) parameters. Neutrophils were identified as CD172+SSChi and monocytes as CD14+CD172a+. In the lymphocyte panel SCClow cells were gated as NK cells (CD16+) and T cells (CD3+). T cell subsets were first gated as cytotoxic T cells (CD4−CD8β+), then CD8β− T cells gated in four subsets based on CD4 and CD8α. CD4+CD8α− are naïve helper αβ T cells; CD4+CD8α+ are principally effector/memory helper T cells; CD4−CD8α− and CD4−CD8α+ include γδ T cells and αβ T cells with less well defined functional phenotypes. The antibodies used are listed in Table 2.
Table 2. List of antibodies.
Cytokine and antibody measurements
Serum cytokines were determined using a custom premixed Milliplex Map porcine cytokine/chemokine magnetic beads kit (Millipore, USA) for the chemokine IL-8 and nine cytokines (IL-1α, IL-1β, IL-1ra, IL-2, IL-4, IL-6, IL-10, IL-12, IL-18). Antibodies against ASFV protein p72 were detected using INgezim PPA COMPAC blocking ELISA Kit (R.11.PPA.K.3, Ingenasa, Madrid, Spain) according to manufacturer's instruction. The results are expressed in % of inhibition, with following cut-off values: < 40%, negative; 40–50%, doubtful; ≥50%, positive. Serum samples were tested in duplicate.
Fecal microbiota analysis
Fecal samples were collected from SPF and farm pigs prior to infection, 1 week after acclimatization into the biocontainment stables of the IVI. Stool collection was performed using sterile swabs and containers and stored at −80°C. DNA was extracted using the QIAamp Fast DNA Stool Mini Kit following the manufacturer's guidelines. For an optimal lysis of and separation of impurities from stool samples, the stools were first suspended and vortexed in 1 ml of InhibitEX Buffer. The V4 region of the 16S rRNA gene was amplified using forward (5'-GTGCCAGCMGCCGCGGTAA-3') and reverse (5'-GGACTACHVGGGTWTCTAAT-3') primers modified with an Illumina adaptor sequence at the 5′ end. PCR products were purified using the QIAquick PCR Purification Kit (Qiagen, Hilden, Germany). Samples were passed through to a MiSeq Illumina sequencing platform for indexing and paired-end sequencing (2 × 250 bp; reagent kit, v2). Sequencing data were analyzed using the DADA2 package (version 1.16.0) in R software (version 4.0.2) for the identification of amplicon sequence variants (ASV). The taxonomy assignment of the ASVs was done using the SILVA (version 132) database. Contaminating sequences were identified using the decontam package (version 1.8.0) in R. Contaminants were identified by their frequency of occurrence and independently within each batch. Distance matrices were calculated for the beta-diversity analyses and used as input files for the non-metric multidimensional scaling (NMDS) plots. Statistical analysis was performed by permutation test (PERMANOVA; Adonis function).
Statistical analysis
Statistical analyses were performed using GraphPad Prism version 8.0.0 for Windows unless otherwise indicated. The number of samples and statistical tests used are indicated in figure legends.
Results
We showed previously that SPF pigs develop a milder disease and have a better survival rate than farm pigs following an intramuscular inoculation of the attenuated Estonia 2014 ASFV strain (12). The current study was designed with two phases. In the first phase, 5 SPF and 8 farm pigs were inoculated oronasally with the Estonia 2014 strain to validate our previous findings where the intramuscular route was used for infection. Blood samples were taken at regular intervals to determine the duration of viremia between pigs of the two groups. In the second phase of the study, 5 pigs from each group were challenged with the highly virulent Armenia 2008 ASFV strain to determine (1) whether prior exposure to Estonia 2014 provides immunity against a challenge with Armenia 2008; and (2) whether SPF and farm pigs have a similar response to this virulent challenge (Figure 1A).
Oronasal inoculation 5 × 109 gEq of ASFV Estonia 2014 induced a first wave of clinical disease with a peak on 5–7 dpi characterized by high fever, loss of appetite, and reduced liveliness. SPF pigs presented significantly milder disease in intensity and duration than farm pigs. All SPF pigs fully recovered by 15 dpi, whereas farm pigs experienced a second peak of clinical signs and a delayed recovery until 21 dpi (Figures 1B, C, left panels). Thus, farm pigs showed a significantly extended duration of clinical signs with fever, weakness, apathy, and reduced appetite. These data concord with our previous study where the animals were inoculated intramuscularly (12). A notable difference was the absence of lethality in the farm pig group, while in the previous study we had observed 50% lethality in that group after intramuscular inoculation of the Estonia 2014 strain. The highest virus loads at ~109 gEq/ml of whole blood were measured in the first sample taken on 5 dpi and levels gradually diminished over time at a similar rate for the two groups until no virus was detectable by PCR in the blood of all the animals on 139 dpi (Figure 1D, left panel). Using a competitive anti-p72 ELISA, seroconversion (>50% inhibition) was observed in all animals at 14 dpi, and maximal antibody responses (100% inhibition) were reached sooner in SPF pigs (27–40 dpi) than in farm pigs (63 dpi) (Figure 1E).
Prior to the second phase of the study, 3 pigs from the farm group were euthanized on 131 dpi to have the same number of pigs in each group. The remaining animals (n = 5/group) were challenged oronasally with 106 TCID50 of the highly virulent Armenia 2008 ASFV strain on 164 dpi that is 0 day post challenge (dpc) (Figure 1A). Strikingly, SPF pigs showed no apparent clinical signs and maintained normal liveliness and appetite after challenge and only one SPF pig had fever for two consecutive days (39.7 and 40.3 on 7 and 8 dpc, respectively). In contrast, all farm pigs displayed severe clinical signs with high fever (>40°C), apathy, weakness, inability to stand, loss of appetite, emaciation, and skin petechiae (Figures 1B, C, right panels). Two farm pigs were euthanized on 14 dpc after reaching the predetermined maximal clinical scores (Figure 1F). Viremia was significantly higher in the EDTA blood of farm pigs compared to SPF pigs (Figure 1D, right panel). The viremia reached a significantly lower plateau at 4 dpc in SPF pigs and at 7 dpc in farm pigs. At the end of the study on 21 dpc, all five SPF pigs and one farm pig had low or no detectable virus in blood, whereas the two remaining farm pigs still had a relatively high viremia (Figure 1D). These results indicate that pigs surviving an infection with the attenuated Estonia 2014 ASFV strain can develop a protective response against a lethal challenge with the highly pathogenic ASFV Armenia 2008 strain. Furthermore, the SPF status provides stronger protection against clinical disease and better control of the viremia compared to conventional farm pigs.
The fecal microbiome of each animal was analyzed prior to the first infection with the Estonia 2014 ASFV strain. While the diversity was similar, the microbiome composition was significantly different between SPF and farm pigs: Prevotellaceae were the dominant family in farm pigs, whereas Bacteroidaceae and Muribaculaceae were the most abundant in SPF pigs (Supplementary Figure 2A). We found that the bacterial family composition remained highly consistent for each group of this study compared to our two previous studies (12) that were collected over a year before. These consistent findings were largely expected for the pigs from the IVI SPF facility where hygiene control is strict, but finding a stable microbiome was less expected for animals from a commercial pig farm (Supplementary Figure 2A).
Analysis of circulating blood cells at baseline prior to the first infection with the Estonia 2014 ASFV strain showed significantly higher red blood cell (RBC) and lower white blood cell (WBC) counts in SPF compared to farm pigs at baseline (Figures 2A, C). No significant difference in platelet (PLT) counts was noted at baseline (Figure 2B). Flow cytometry analysis at baseline revealed that the low WBC counts in SPF pigs were caused by reduced neutrophils, NK cells, and subsets of T cells such as cytotoxic CD8β+ T cells (Figures 2D, F, 3). These data are consistent with our previous study that revealed a more naïve immune system in SPF pigs with particularly low neutrophil and other cell subset counts in blood and are consistent with the different hygienic status and the stable microbiome data over time in both groups (12).
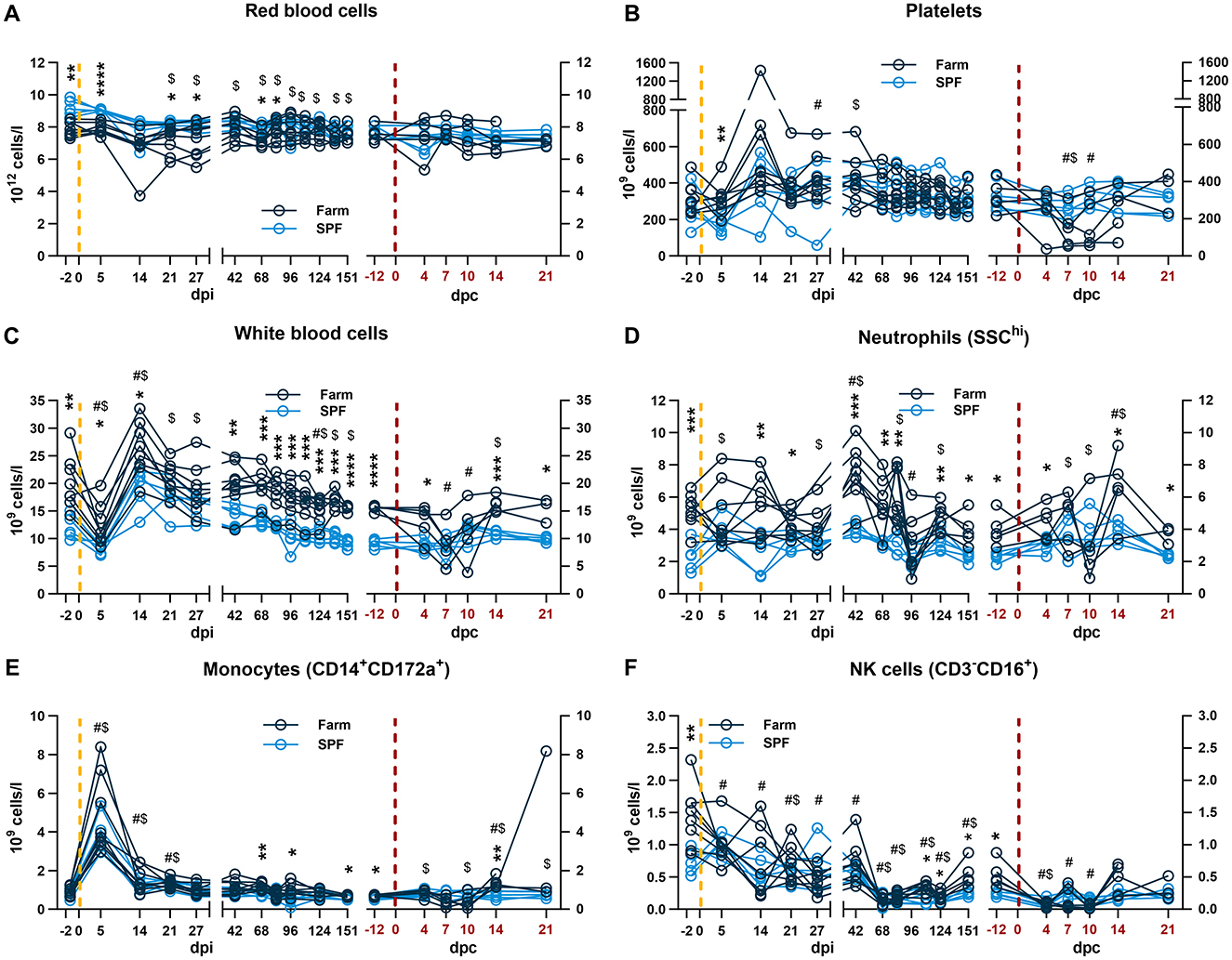
Figure 2. Blood and innate immune cell count profiles. Each panel is shown in two parts that were analyzed independently. On the left, the yellow dashed line represents the inoculation with the ASFV Estonia 2014 strain and time points are referred to as days post infection (dpi); on the right, the red dashed line represents the challenge with the ASFV Armenia 2008 strain and time points are referred to as days post challenge (dpc, in red). (A–C) Red blood cell, platelet, and white blood cell (WBC) counts were determined in whole blood. (D–F) Absolute counts of (D) neutrophils, (E) monocytes, (F) and NK cells. The percentage of each subset was determined by flow cytometry and absolute numbers were calculated using WBC counts. (A–F) Data points represent values for individual pigs. Differences between SPF and farm groups were analyzed by two-way ANOVA (mixed model) with Sidak's multiple comparisons test. Statistical differences between groups are shown as *p < 0.05; **p < 0.01; ***p < 0.001; ****p < 0.0001. Changes in cell counts within each group were also compared to their respective baseline values taken before each infection at −2 dpi or at −12 dpc (equivalent to 151 dpi). Significant differences within groups compared to their respective pre-challenge status are indicated for SPF ($, p < 0.05) and farm pigs (#, p < 0.05); data were analyzed using mixed-effects analysis.
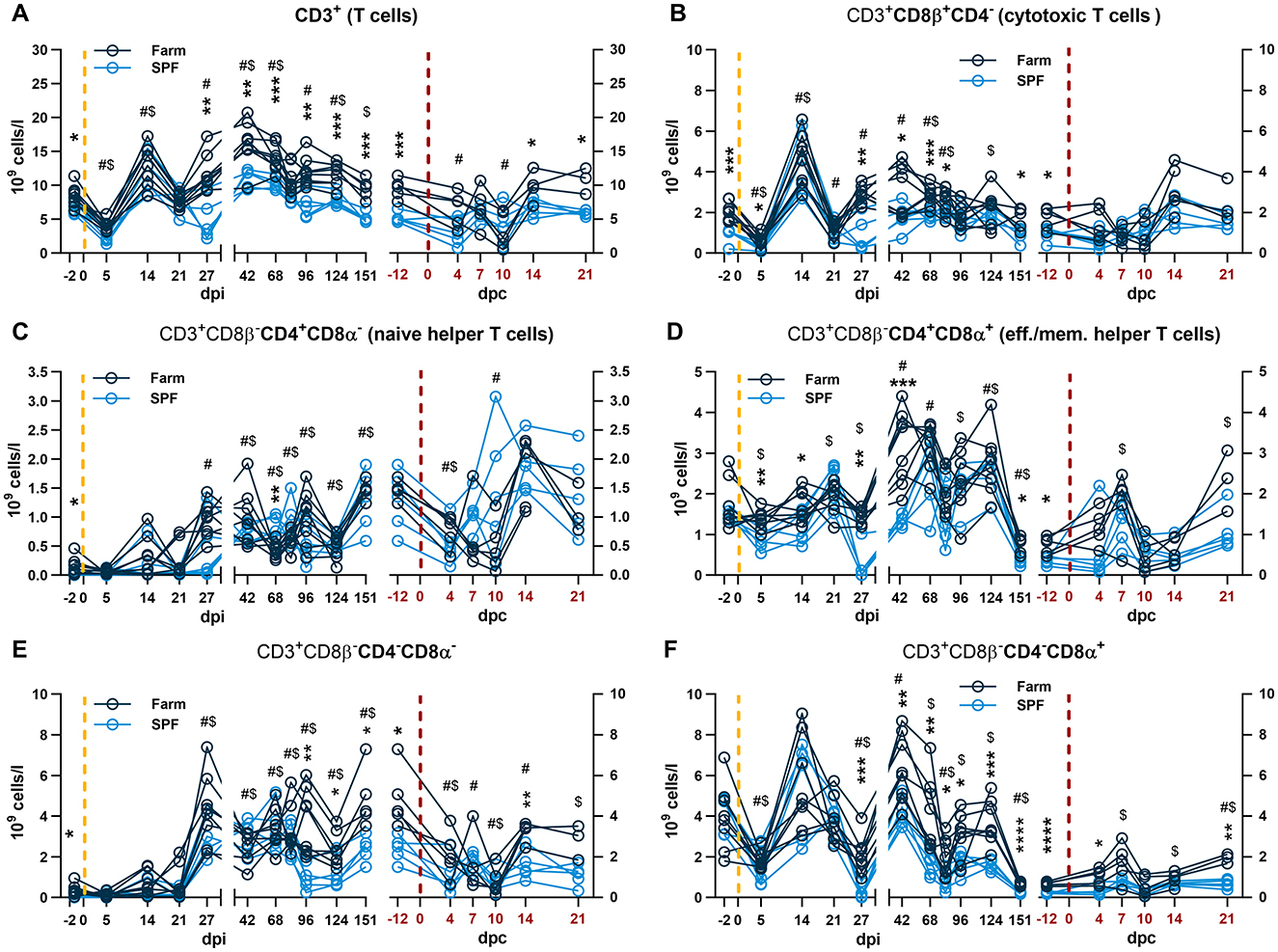
Figure 3. Blood T cell subset analysis. Each panel is shown in two parts that were analyzed independently. On the left, the yellow dashed line represents the inoculation with the ASFV Estonia 2014 strain and time points are referred to as days post infection (dpi); on the right, the red dashed line represents the challenge with the ASFV Armenia 2008 strain and time points are referred to as days post challenge (dpc, in red). (A) Total CD3+ T cell counts. (B) Cytotoxic T cells defined by CD3+CD4−CD8β+. (C–F) After gating cytotoxic T cells, the remaining T cells (CD3+ CD8β−) were identified by relative expression of CD4 and CD8α as (C) naïve helper T cells CD8β−CD4+CD8α−, (D) effector and memory (eff./mem.) T cells CD8β−CD4+CD8α−; (E) CD8β−CD4−CD8α−, and (F) CD8β−CD4−CD8α+. (A–F) The percentage of each subset was determined by flow cytometry and absolute numbers were calculated using WBC counts. (A–F) Data points represent values for individual pigs. Differences between SPF and farm groups were analyzed by two-way ANOVA (mixed model) with Sidak's multiple comparisons test. Statistical differences between groups are shown as *p < 0.05; **p < 0.01; ***p < 0.001; ****p < 0.0001. Changes in cell counts within each group were also compared to their respective baseline values taken before each infection at −2 dpi or at −12 dpc (equivalent to 151 dpi). Significant differences within groups compared to their respective pre-challenge status are indicated for SPF pigs ($, p < 0.05) and farm pigs (#, p < 0.05); data were analyzed using mixed-effects analysis.
After infection with the Estonia 2014 strain, a sharp drop in WBC counts at 5 dpi followed by a rebound at 14 dpi were observed in both groups (Figure 2C). The effect at 5 dpi was driven by lymphopenia with a reduction of total T cells and particularly CD8β+ and CD8β−CD4−CD8α+ in both groups (Figures 3A, B, F), and NK cells in farm pigs (Figure 2F). This lymphopenia was followed by a sharp increase of T cells, in particular cytotoxic CD8β+ T cells at 14 dpi (Figures 3A, B). Absolute counts of monocytes sharply peaked in both groups at 5 dpi and remained elevated on 14 and 21 dpi before returning to baseline from 27 dpi (Figure 2E).
During the recovery phase, where both groups remained viremic but asymptomatic, differences in total WBC remained comparable to the baseline prior to infection with significantly higher leukocyte counts in farm pigs compared to SPF pigs (Figure 2C). As for the baseline, neutrophil, NK cell and T cell counts significantly contributed to the reduced WBC in SPF pigs compared to farm pigs (Figures 2, 3). Prior to the challenge with the Armenia 2008 strain (-12 dpc, equivalent to 151 dpi), a second baseline was established for RBC, PLT and leukocyte subset counts (Figures 2, 3, right panels). RBC and PLT counts were similar in both groups, while WBC counts were lower in the SPF group. Neutrophil, monocyte, NK cell, and T cell subset counts were all significantly lower in the SPF pigs compared to farm pigs at −12 dpc.
After the challenge with the ASFV Armenia 2008 strain, we observed significant reduction of platelets and white blood cell counts in farm pigs, consistent with acute ASF disease (Figures 2B, C, right panels). The loss of total T cells was also prominent on 4–10 dpc in farm pigs, which was observed in several T cell subsets, but no clear pattern could be identified in relation clinical severity (Figure 3). In contrast, the challenge did not induce leukopenia in the SPF group. On the contrary, we observed increased neutrophils on 7–14 dpc, monocytes on 4–21 dpc (Figures 2D, E, right panels), and CD4+CD8α+ effector/memory helper T cells on 7 dpc (Figure 3D, right panel).
The cytokine response in serum was evaluated 5 and 14 dpi with the Estonia 2014 strain and 4, 7, 10, 14 dpc with the Armenia 2008 strain. At 5 dpi of the first infection, only the neutrophil chemokine IL-8 and the pro-inflammatory cytokine IL-12 were significantly higher in farm compared to SPF pigs (Figures 4A, B, left panels). Upon challenge with the Armenia 2008 strain, significantly higher levels of IL-8, pro-inflammatory cytokines IL-1β and IL-6, and the Th2 cytokine IL-4 were significantly increased in the farm pigs compared to SPF pigs at one or several time points after the Armenia 2008 strain challenge (Figure 4). At 7 dpc, farm pigs had significantly elevated levels of the anti-inflammatory cytokine IL-10 and the IL-1 receptor antagonist (IL-1ra) (Figures 4I, J), and this increase was particularly high in the farm pigs with the most severe clinical scores, suggesting regulatory feedback. In the SPF pig group, the cytokine levels were not altered in response to the challenge with Armenia 2008 strain. We also examined the changes in fecal microbiota of the two pig groups on 10 dpc. NMDS analysis showed that SPF and farm pigs clustered differently from each other at baseline (−2dpi) and at 10 dpc (Supplementary Figure 2B).
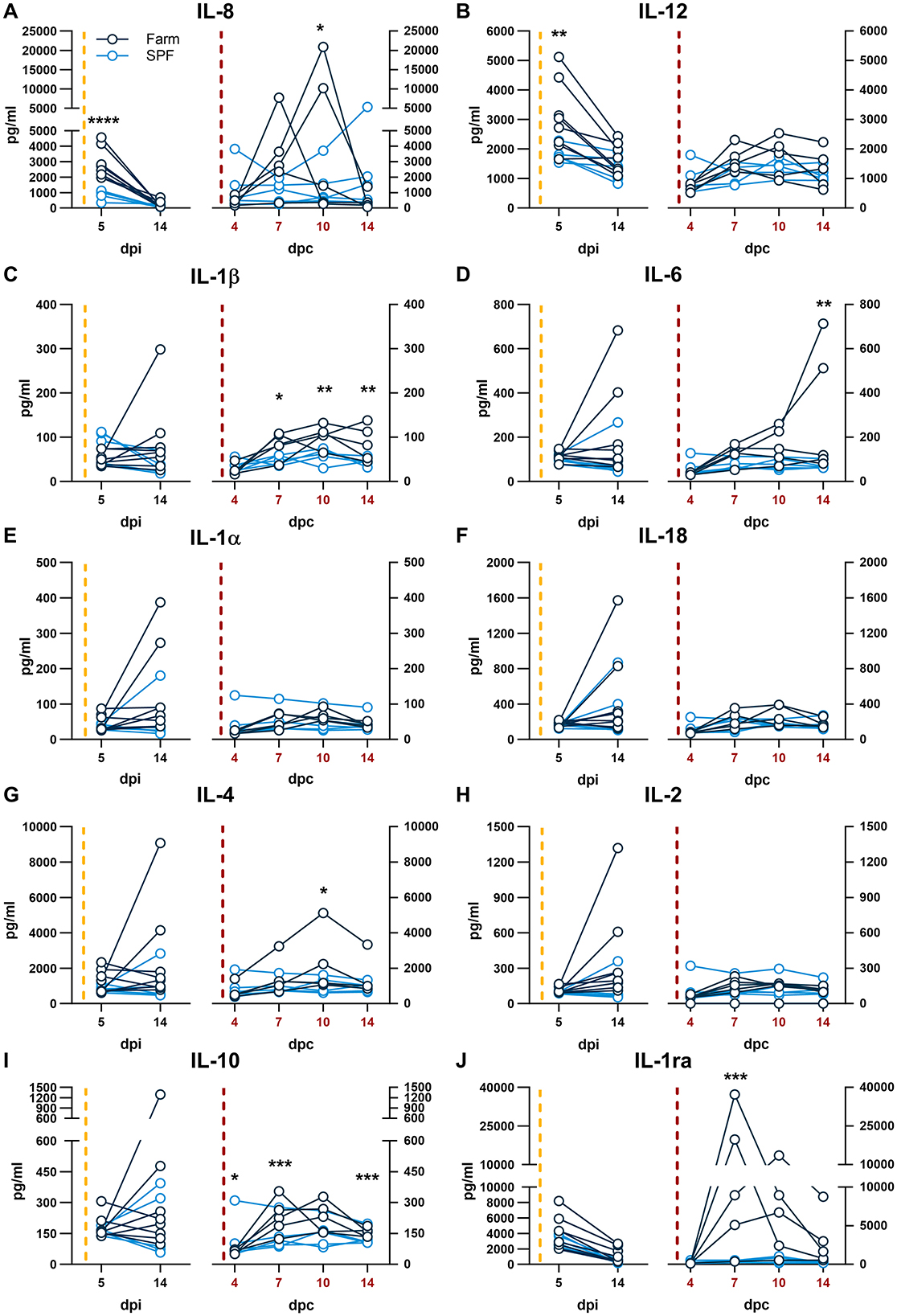
Figure 4. (A–J) Serum cytokines. Each panel is shown in two parts that were analyzed independently. On the left, the yellow dashed line represents the inoculation with the ASFV Estonia 2014 strain and time points are referred to as days post infection (dpi); on the right, the red dashed line represents the challenge with the ASFV Armenia 2008 strain and time points are referred to as days post challenge (dpc, in red). Cytokine levels were determined by multiplex ELISA. Differences between SPF and farm groups were analyzed by two-way ANOVA (mixed model); *p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001.
Overall, the lack of a strong leukocyte depletion and cytokine response in SPF pigs associated with a very low-grade viremia and clinical disease following the ASFV Armenia 2008 strain challenge demonstrates that SPF pigs have acquired a strong and almost sterilizing immunity against ASFV.
Discussion
In this new study comparing piglets born and raised in our SPF facility with those from a conventional commercial farm, we found that the gut microbiomes were strikingly different in composition as reported previously (12). The main bacterial families of the gut microbiome of pigs of the respective groups were also identical to the samples evaluated from the same facilities more than a year before. This constancy indicates that the microbiome is a stable component of the animals from a specific facility whether it is an experimental SPF facility, or a commercial pig farm, where environmental conditions may not be as strictly controlled. In addition, the farm pigs also had higher counts of circulating leukocytes and higher body temperature at baseline compared to SPF pigs as shown previously.
Furthermore, we showed a higher resilience of SPF pigs compared to farm pigs following oronasal infection with the moderately virulent ASFV Estonia 2014 strain, thus confirming our previous findings following intramuscular infection with this strain (12). In both studies, SPF pigs demonstrated significantly less severe clinical signs and a faster recovery than farm pigs. A notable difference was that we had observed 50% lethality for the farm pigs when the pigs were infected intramuscularly with ASFV Estonia 2014 strain in the previous study, whereas here, all farm pigs ultimately recovered from the infection after oronasal infection with the same inoculum dose. The improved outcome in farm pigs may be explained by the different route of infection, with more severe outcome after intramuscular infection. In addition, only two blood samples were taken in the acute phase of the disease on 5 and 14 dpi in this study, whereas seven blood samples were taken in the previous study on 1, 2, 4, 5, 7, 11, and 14 dpi, which may have compounded the effects of the virus-induced thrombocytopenia and contributed in part to the lethality.
The viremia after oronasal instillation of the ASFV Estonia 2014 strain was followed until it became undetectable by PCR in serum and whole blood. SPF and farm groups had comparable virus load at all time points. The highest viremia was measured at the first sampling at 5 dpi in both groups and the virus titers gradually reduced over several months until no virus was detectable by PCR on 139 dpi in all animals. This prolonged viremia is consistent with previous publications (15), notably the kinetics of the viremia reported for this strain (16). These findings indicate that the more severe disease observed in farm pigs is not associated with uncontrolled virus replication compared to SPF pigs and that other mechanisms linked to the host response are driving the pathology.
Upon subsequent challenge with the ASFV Armenia 2008 strain, the almost complete absence of clinical signs and the controlled viremia in SPF pigs contrasted with the more severe disease in farm pigs including the euthanasia of 2 out of 5 farm pigs. Upon challenge, farm pigs showed significantly higher viremia, increased inflammatory cytokine release, and a characteristic transient drop in NK cell and T cells counts in blood. These findings indicate that SPF pigs developed a stronger protective adaptive immune response and that host immune factors influenced by the housing conditions and potentially the microbiome are also crucially important for the development of a strong protective adaptive immunity. The timing of the seroconversion against the major capsid antigen p72 indicated a slight delay in farm pigs. Nevertheless, this parameter is unlikely an indicator of protection against a re-challenge as all animals raised a strong antibody response against this antigen.
The pattern of cytokine expression analysis indicates an association between high levels of the neutrophil chemokine IL-8 with higher clinical scores after infection with either strains. Furthermore, higher IL-8 responses at 5 dpi during the first infection with the ASFV Estonia 2014 strain also correlated with a worse outcome in farm pigs after the rechallenge months later with the Armenia 2008 strain. This suggests that the lower basal immune activation status of SPF pigs leads to lower secretion of IL-8 and milder clinical symptoms after the first infection but enhanced adaptive responses after re-challenge. The fatal outcome in the 2 farm pigs was further associated with very high levels of the inflammatory cytokine IL-6 at 14 dpc, which is consistent with previous studies (12, 17). These data are consistent with evidence indicating that high levels of serum IL-6 are associated with a bad prognosis in severe infections (18, 19).
We had previously shown that after infection with the ASFV Estonia 2014 strain, SPF pigs expressed significantly higher levels of IL-1ra at 4 dpi. IL-1ra blocks signaling of the receptor for the inflammatory cytokines IL-1α and IL-1β. We speculated that an early and high peak of IL-1ra may be a protective factor associated with a higher resilience to a moderately virulent ASFV infection. Here, the first sample taken after infection with the Estonia 2014 strain was performed only at 5 dpi, which prevented us to make a direct comparison. Like in the previous study, we found no significant difference in IL-1ra levels between the two groups at 5 dpi. The interpretation of the results of the current study is limited by the relatively low number of animals per group and the lack of cytokine data prior to each infection. Future work should include the validation of early innate markers associated with resilience.
The analysis of the blood cell subset dynamics in this longitudinal study did not provide a strong correlation between responses after the infection with Estonia 2014 that could provide a predictive outcome following the challenge with the virulent ASFV strain. However, we observed a significant increase in CD4+CD8α+ T cells in SPF pigs, suggesting the expansion of effector and/or memory T cells associated with protection. Further studies are needed to explore antigen-specific T cell responses and functional antibody assays correlating with protection.
This study highlights again the importance of environmental factors and hygiene status such as prior infections and microbiota composition on innate and adaptive immune responses in pigs. It remains unclear if the differences in resilience and adaptive responses between SPF and farm pigs are directly linked with beneficial or opportunistic bacteria and their metabolites, or if this is only a bystander correlation. Fecal microbiota transfers studies may help understand the function of the microbiota in the mechanisms of resilience and protective immune responses against ASFV.
In conclusion, controlled studies of pigs with different hygienic backgrounds provide a solid platform to explore the mechanism and correlates of protections against ASF, and how they are influenced by environmental factors.
Data availability statement
The original contributions presented in the study are included in the article and Supplementary material, further inquiries can be directed to the corresponding author. The sequencing data presented in the study were deposited in the NCBI repository, accession number PRJNA1234064.
Ethics statement
The study was performed in compliance with the Animal Welfare Act (TSchG SR 455), the Animal Welfare Ordinance (TSchV SR 455.1), and the Animal Experimentation Ordinance (TVV SR 455.163) of Switzerland. All experiments were reviewed by the committee on animal experiments of the canton of Bern and approved by the cantonal veterinary authority under the license BE18/2019.
Author contributions
ER: Data curation, Formal analysis, Investigation, Methodology, Validation, Visualization, Writing – original draft. KM: Data curation, Formal analysis, Investigation, Methodology, Validation, Writing – review & editing. TW: Data curation, Formal analysis, Investigation, Methodology, Writing – review & editing. MH: Formal analysis, Supervision, Validation, Writing – review & editing. AS: Conceptualization, Funding acquisition, Methodology, Project administration, Resources, Supervision, Validation, Writing – review & editing. NR: Conceptualization, Data curation, Formal analysis, Funding acquisition, Investigation, Methodology, Project administration, Resources, Supervision, Validation, Writing – review & editing. CB: Conceptualization, Data curation, Formal analysis, Funding acquisition, Investigation, Methodology, Project administration, Resources, Supervision, Validation, Visualization, Writing – original draft, Writing – review & editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This work was funded by grants of the Swiss Federal Food Safety and Veterinary Office (Grant Numbers 1.19.02 and 1.21.12) to NR, CB, and AS, and by internal funds of the Institute of Virology and Immunology (IVI). This research was made possible by funding from ICRAD, an ERA-NET co-funded under European Union's Horizon 2020 research and innovation program (https://ec.europa.eu/programmes/horizon2020/en), under Grant Agreement No. 862605. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript. Open access funding by University of Bern.
Acknowledgments
We thank Daniel Brechbühl, Katarzyna Sliz, Hans-Peter Lüthi, Jan Salchli, and Roman Troxler for excellent animal care. We thank Aurélie Godel and Sylvie Python for technical assistance. We thank Imbi Nurmoja and Olev Kalda from the Estonian Veterinary and Food Laboratory, Tartu, Estonia for their agreement to transfer the Estonia 2014 isolate to the IVI. We are grateful to Takehiro Kokuho (NARO, Tokyo, Japan) for generously providing the IPKM cells. We thank Nadezda Mostacci (Institute for Infectious Diseases, University of Bern) for uploading the microbiome data onto the NCBI repository.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Gen AI was used in the creation of this manuscript.
Publisher's note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fvets.2025.1553310/full#supplementary-material
References
1. Montgomery RE. On a form of swine fever occurring in British East Africa (Kenya Colony). J Comp Pathol Therap. (1921) 34:159–91. doi: 10.1016/S0368-1742(21)80031-4
2. Penrith ML, Kivaria FM. One hundred years of African swine fever in Africa: Where have we been, where are we now, where are we going? Transbound Emerg Dis. (2022) 69:e1179–200. doi: 10.1111/tbed.14466
3. Cadenas-Fernandez E, Sanchez-Vizcaino JM, Pintore A, Denurra D, Cherchi M, Jurado C, et al. Free-ranging pig and wild boar interactions in an endemic area of african swine fever. Front Vet Sci. (2019) 6:376. doi: 10.3389/fvets.2019.00376
4. Gonzales W, Moreno C, Duran U, Henao N, Bencosme M, Lora P, et al. African swine fever in the Dominican Republic. Transbound Emerg Dis. (2021) 68:3018–9. doi: 10.1111/tbed.14341
5. Zhou X, Li N, Luo Y, Liu Y, Miao F, Chen T, et al. Emergence of African Swine Fever in China, 2018. Transbound Emerg Dis. (2018) 65:1482–4. doi: 10.1111/tbed.12989
6. Huhr J, Schafer A, Schwaiger T, Zani L, Sehl J, Mettenleiter TC, et al. Impaired T-cell responses in domestic pigs and wild boar upon infection with a highly virulent African swine fever virus strain. Transbound Emerg Dis. (2020) 67:3016–32. doi: 10.1111/tbed.13678
7. Dixon LK, Sun H, Roberts H. African swine fever. Antiviral Res. (2019) 165:34–41. doi: 10.1016/j.antiviral.2019.02.018
8. Zhao D, Sun E, Huang L, Ding L, Zhu Y, Zhang J, et al. Highly lethal genotype I and II recombinant African swine fever viruses detected in pigs. Nat Commun. (2023) 14:3096. doi: 10.1038/s41467-023-38868-w
9. Wang L, Ganges L, Dixon LK, Bu Z, Zhao D, Truong QL. International African Swine fever workshop: critical issues that need to be addressed for ASF control. Viruses. (2023) 16:4. doi: 10.3390/v16010004
10. Auer A, Cattoli G, Padungtod P, Lamien CE, Oh Y, Jayme S, et al. Challenges in the application of African Swine Fever vaccines in Asia. Animals. (2024) 14:2473. doi: 10.3390/ani14172473
11. Chu X, Ge S, Zuo Y, Cui J, Sha Z, Han N, et al. Thoughts on the research of African swine fever live-attenuated vaccines. Vaccine. (2024) 42:126052. doi: 10.1016/j.vaccine.2024.06.020
12. Radulovic E, Mehinagic K, Wuthrich T, Hilty M, Posthaus H, Summerfield A, et al. The baseline immunological and hygienic status of pigs impact disease severity of African swine fever. PLoS Pathog. (2022) 18:e1010522. doi: 10.1371/journal.ppat.1010522
13. Mittelholzer C, Moser C, Tratschin JD, Hofmann MA. Analysis of classical swine fever virus replication kinetics allows differentiation of highly virulent from avirulent strains. Vet Microbiol. (2000) 74:293–308. doi: 10.1016/S0378-1135(00)00195-4
14. Masujin K, Kitamura T, Kameyama K, Okadera K, Nishi T, Takenouchi T, et al. An immortalized porcine macrophage cell line competent for the isolation of African swine fever virus. Sci Rep. (2021) 11:4759. doi: 10.1038/s41598-021-84237-2
15. Petrov A, Forth JH, Zani L, Beer M, Blome S. No evidence for long-term carrier status of pigs after African swine fever virus infection. Transbound Emerg Dis. (2018) 65:1318–28. doi: 10.1111/tbed.12881
16. Nurmoja I, Petrov A, Breidenstein C, Zani L, Forth JH, Beer M, et al. Biological characterization of African swine fever virus genotype II strains from north-eastern Estonia in European wild boar. Transbound Emerg Dis. (2017) 64:2034–41. doi: 10.1111/tbed.12614
17. Wang S, Zhang J, Zhang Y, Yang J, Wang L, Qi Y, et al. Cytokine storm in domestic pigs induced by infection of virulent African Swine Fever virus. Front Vet Sci. (2020) 7:601641. doi: 10.3389/fvets.2020.601641
18. Laing AG, Lorenc A, Del Molino Del Barrio I, Das A, Fish M, Monin L, et al. A dynamic COVID-19 immune signature includes associations with poor prognosis. Nat Med. (2020) 26:1623–35. doi: 10.1038/s41591-020-1038-6
Keywords: African swine fever, microbiota, specific pathogen-free (SPF) pigs, in vivo, immune response, resilience
Citation: Radulovic E, Mehinagic K, Wüthrich TM, Hilty M, Summerfield A, Ruggli N and Benarafa C (2025) Development of protective immunity against African swine fever depends on host-environment interactions. Front. Vet. Sci. 12:1553310. doi: 10.3389/fvets.2025.1553310
Received: 03 January 2025; Accepted: 23 April 2025;
Published: 10 June 2025.
Edited by:
Dirk Werling, Royal Veterinary College (RVC), United KingdomReviewed by:
Alexander Schäfer, Friedrich-Loeffler-Institut, GermanyGiulia Franzoni, Experimental Zooprophylactic Institute of Sardinia (IZS), Italy
Copyright © 2025 Radulovic, Mehinagic, Wüthrich, Hilty, Summerfield, Ruggli and Benarafa. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Charaf Benarafa, Y2hhcmFmLmJlbmFyYWZhQHVuaWJlLmNo
†These authors share first authorship
‡These authors share senior authorship