 Denisa Pérez Gaudio
Denisa Pérez Gaudio Mery Giantin
Mery Giantin Marianna Pauletto
Marianna Pauletto Mauro Dacasto
Mauro Dacasto- 1Laboratorio de Toxicología, Depto. de Fisiopatología, Facultad de Ciencias Veterinarias, Centro de Investigación Veterinaria de Tandil (CIVETAN), Universidad Nacional del Centro de la Provincia de Buenos Aires, Tandil, Buenos Aires, Argentina
- 2Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET), Buenos Aires, Argentina
- 3Department of Comparative Biomedicine and Food Science, Division of Veterinary Pharmacology and Toxicology, University of Padua, Padua, Italy
Introduction: Thymol, a phenolic compound present in essential oils, has shown promising pharmacological properties and potential veterinary applications. This systematic review aims to evaluate the pharmacological and toxicological effects of thymol on animals.
Materials and methods: A comprehensive search following PRISMA 2020 guidelines was conducted across databases including PubMed, Scopus, Google Scholar, Web of Science, and LILACS. Various health descriptors, medical subject headings terms, and their synonyms were used to identify studies examining thymol's effects in animals, including its in vitro, in vivo, and in silico toxicity, as well as its possible environmental impact. Only relevant experimental studies from the last 12 years were included, provided they assessed thymol itself, one of its primary phytoadditive sources, or a blend where thymol was at least as concentrated as other components. The protocol was registered in Open Science Framework (DOI: https://doi.org/10.17605/OSF.IO/B6SF9).
Results: A total of 1.472 records were identified, with 176 meeting inclusion criteria. Studies spanned from 2012 to 2024, indicating that thymol exhibits antimicrobial, antiviral, antifungal, antiparasitic, anti-inflammatory, and antioxidant properties, among others. However, thymol displays dose-dependent toxicity, especially at high levels, affecting mainly the liver and kidneys.
Discussion: Thymol holds substantial potential as a therapeutic agent in veterinary medicine due to its beneficial effects and relatively low toxicity. Nonetheless, further research is needed to establish safe and effective dosages across different animal species.
Systematic review registration: https://doi.org/10.17605/OSF.IO/B6SF9.
1 Introduction
The use of natural compounds in veterinary medicine has garnered increasing interest as alternatives to conventional pharmaceuticals, driven by concerns over antimicrobial resistance and the potential side effects of synthetic drugs. Among these natural compounds, thymol (THY; 2-isopropyl-5-methylphenol), a monoterpenoid phenol (Figure 1), has been widely studied for its pharmacological properties across various animal species, including both production and companion animals. THY is primarily found in plants of the Thymus genus, especially Thymus vulgaris (common thyme), but it is also present in Origanum (oregano), Ocimum (basil), Monarda species (bergamot, bee balm, horsemint, and Oswego tea), Lippia origanoides (Mexican oregano), and Nigella species (e.g., Nigella sativa, black cumin), among others (1). Given its presence in several essential oils (EO), THY is often incorporated into phytotherapeutic blends, where it exhibits synergistic or additive effects when combined with other active constituents, such as carvacrol (CAR), eugenol (EUG), and cinnamaldeyde (CIN), to enhance its efficacy and expand its range of biological activities (2–6).
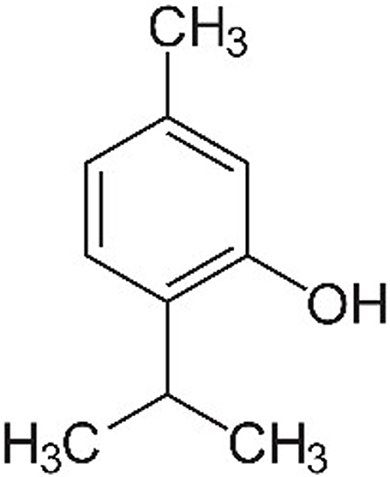
Figure 1. Thymol's chemical structure.
THY has demonstrated a range of pharmacologically beneficial activities that are highly relevant in veterinary contexts. Studies have consistently reported its antimicrobial, antifungal, anti-inflammatory, antioxidant, and analgesic properties, making it a promising candidate for managing various health conditions in animals (1). THY has also demonstrated anticancer effects in human cell lines; however, studies in animal models are scarce, and further research in broader and more specific animal systems is required to establish its potential utility in veterinary oncology (7).
Notably, THY has been shown to exert broad-spectrum antimicrobial activity against both Gram-positive and Gram-negative bacteria of production and companion animals, which is of particular importance in veterinary applications where zoonotic pathogens are common. In small animals, THY's antimicrobial effect has potential applications, particularly for oral and dermatological care. THY-containing products are widely used in pet oral care formulations, which help reduce dental plaque and prevent periodontal diseases, conditions prevalent among pets (8). In these species, THY and Thymus vulgaris essential oils (THEO) have also exhibited notable antiviral activity against a variety of viruses, including feline calicivirus (FCV), feline coronavirus (FCoV), and norovirus surrogates. Moreover, its antifungal activity offers potential in treating or preventing infections caused by pathogens such as Pythium (9) and Aspergillus species (10). In large animals, THY has also been shown to be promising as both an internal and external antiparasitic agent, effectively targeting gastrointestinal parasites as well as ectoparasites like ticks and mites, which contributes to improved animal health and productivity without relying on conventional synthetic antiparasitics (11). On the other hand, in companion animals, THY has also been investigated as an ectoparasiticide, showing potential in controlling flea and tick infestations without resorting to synthetic insecticides that may have adverse effects on pets and the environment (12). Such applications underscore the versatility of THY in veterinary care, making it an attractive option for a wide range of therapeutic interventions across multiple species.
The anti-inflammatory and antioxidant properties of THY have also been explored, particularly concerning its role in modulating immune responses and reducing oxidative stress, which can significantly impact animal health, especially in intensive production systems (13, 14). These properties suggest that THY could serve as a supportive therapy in managing chronic inflammatory conditions or as a preventive measure against stress-induced pathologies (15), common in both livestock and companion animals.
In production animals, THY has been extensively studied as a feed additive aimed at promoting growth and improving gut health. THY is known to influence gut microbiota composition positively (16, 17), enhancing the populations of beneficial microbes while inhibiting pathogenic bacteria, which results in better nutrient absorption and overall health (18). The inclusion of THY in livestock diets has been shown to improve weight gain, feed efficiency, and immune function, particularly in poultry and swine production (19, 20). These benefits have driven interest in THY as a natural growth promoter, especially as the use of antibiotic growth promoters has been increasingly restricted in various regions due to concerns over antibiotic resistance (21).
Furthermore, THY-containing blends that combine other EO demonstrated enhanced efficacy in promoting health and productivity in ruminants, such as cattle and sheep, by stimulating ruminal fermentation and enhancing fiber digestion (22). Such applications have shown promise in reducing methane emissions, a relevant environmental consideration, thereby contributing to more sustainable livestock production.
Research in laboratory animals, particularly rodents, is essential in veterinary pharmaco-toxicology for understanding compounds like THY. Rodent studies provide key data on THY's pharmacokinetics [absorption, distribution, metabolism, and excretion (ADME)] and help define dosing protocols for larger animals. Toxicological assessments in rats also clarify safety thresholds, reduce risks in veterinary applications, and reveal mechanisms of action that often translate to other species. Furthermore, these models highlight potential metabolic interactions relevant to long-term use or concurrent administration with other agents (23).
THY has also been extensively studied in cell cultures of animal origin to elucidate its pharmacological effects at the cellular level, especially regarding its antimicrobial, anti-inflammatory, and antioxidant properties. In vitro studies using animal-derived cells, such as intestinal and hepatic cell lines, demonstrated THY's ability to modulate cellular responses to oxidative stress and inflammation, mechanisms that are central to maintaining tissue integrity in both production and companion animals (24–26). Additionally, THY has shown cytotoxic effects on specific pathogenic organisms when applied to cell cultures, highlighting its potential as a natural antimicrobial agent in veterinary medicine (27). These cellular models revealed THY's dose-dependent effects, providing critical insights into its therapeutic thresholds and cytotoxic potential, which are essential for safe and effective application in animal health.
This systematic review aims to provide a comprehensive analysis of the pharmaco-toxicological properties of THY in veterinary medicine by synthesizing evidence from studies conducted on production animals, companion animals, and, where relevant, laboratory rats and mice. By evaluating the efficacy, safety, and potential limitations of THY, both as a standalone compound and in phytotherapeutic blends, this review seeks to inform veterinary practitioners about THY's applicability and contribute to evidence-based guidelines for its use. Given the increasing restrictions on synthetic antimicrobials and growth promoters, understanding the potential of THY and similar compounds is crucial for developing sustainable, effective, and safe alternatives in veterinary care.
2 Materials and methods
2.1 Question and PICOS strategy
This systematic review focuses on the toxicological and pharmacological aspects of THY and its main phytochemical sources in production and companion animals, and its purpose is to answer the following questions: Is THY toxic for animal cells and tissues? Which beneficial effect does THY exhibit on animal cells (in vitro), animal models of diseases, animal physiology or different pathologies (in vivo)? The review followed the guidelines of the Preferred Reporting Items for Systematic Reviews and Meta-Analyses (PRISMA) (28). Moreover, a PICOS strategy (patient or pathology, intervention, control, and other outcomes and type of study) was used based on: P: healthy or microorganism challenged animals; I: treatment with THY; C: no treatment, healthy cells or animals receiving placebo (vehicle) or absence of THY source; O: cytotoxic and beneficial effects; S: in vitro and in vivo studies.
2.2 Data sources and literature search
The literature search was conducted on the Web of Science, PubMed, Scopus, LILACS, and Google Scholar until December 2024. A combination of search terms was used, structured with Boolean operators to ensure comprehensive coverage. Keywords included thymol-related terms (“thymol” OR “essential oils” OR “thyme extract”), pharmacological descriptors (“pharmacological” OR “antioxidant” OR “antimicrobial” OR “bioactivity”), toxicological descriptors (“toxicity” OR “cytotoxicity” OR “safety”), pharmacokinetic-related terms (“pharmacokinetics” OR “pharmacodynamics” OR “absorption” OR “distribution” OR “metabolism” OR “excretion” OR “ADME”), and species-specific terms (“rats” OR “cattle” OR “pigs” OR “chickens” OR “rabbits”), among others. These groups of terms were combined using the AND operator to retrieve studies addressing thymol or its sources, their biological effects, and relevance to veterinary species.
Given the broad range of databases, studies, and species, the search strategy was designed to maximize coverage while maintaining relevance to the research questions. This ensured that key studies across diverse models and different animal species were included. Articles published between 2012 and 2024 were considered to ensure the inclusion of the most current and relevant research on THY's toxicological and pharmacological properties. The initial search was then followed by manual screening of reference lists from selected studies to capture any additional articles not indexed in the databases.
2.3 Study selection
The inclusion criteria for the systematic review were as follows: (i) all types of investigation design (in vitro and in vivo studies, including murine models of pathologies relevant to veterinary medicine); (ii) studies related specifically to the application of THY, or a phytoadditive THY source where THY was the main component or when the exact composition is not reported by authors but it is also known that THY is the main component of the plant, or the use of a blend where THY concentration was superior or equal to the other components; (iii) manuscripts that assessed directly or indirectly the toxicological or beneficial effects of THY in companion animals, production animals (conventional and non-conventional), and other species of veterinary interest. For article exclusion, the following specific criteria were defined: (i) restrictions on year of publication, selecting only articles published in the period 2012–2024; (ii) articles using mice, rats, or cells from murine origin were excluded if they were used as a model for exclusive human diseases and their results could not be of importance for animal treatments; (iii) studies involving THEO or blends in which THY was included but not as the primary compound. Figure 2 shows the flow diagram used for the selection of the studies included in the systematic review.
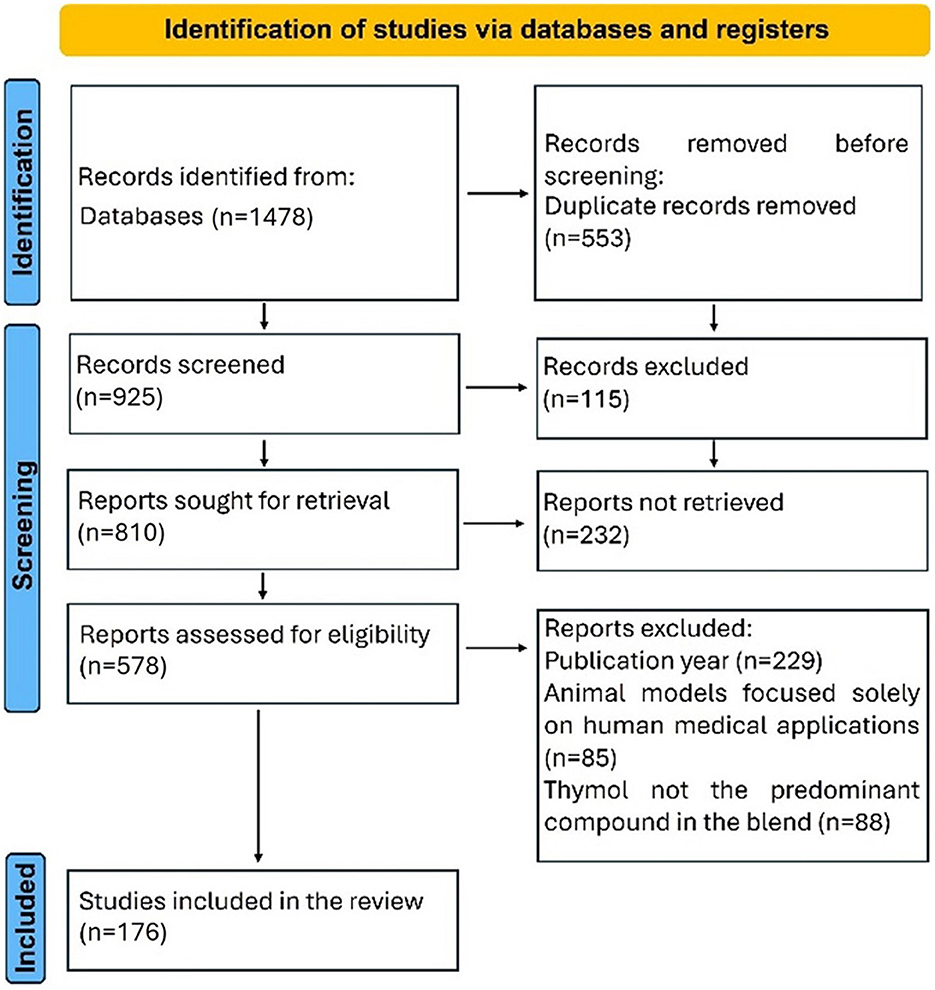
Figure 2. A flow diagram is used for the selection of the studies included in the systematic review.
2.4 Data extraction
Data extraction was carried out by two independent reviewers using a standardized form. The form collected key information, including study design, animal species, doses of THY or THY-based additives, assessment of toxicity (cytotoxicity, organ toxicity, or adverse effects), and beneficial effects (e.g., antioxidant, antimicrobial, and immunomodulatory). Discrepancies between reviewers were resolved through discussion. The data extracted from each study were then organized and transformed when necessary to ensure consistency across studies, particularly regarding units of measurement.
2.5 Quality assessment and risk of bias
The quality of the studies included in the systematic review was assessed using predefined criteria, which evaluated the methodological rigor of each study. Key factors, such as randomization, blinding, control groups, and potential sources of bias, were considered. Studies that were judged to have a high risk of bias or those that lacked sufficient methodological transparency were excluded or treated with caution in the analysis. This process ensures that the final synthesis is based on studies with reliable and robust evidence. The risk of bias and quality assessment was performed independently by two reviewers, with any discrepancies resolved by discussion.
2.6 Synthesis and presentation of results
The results of the included studies were synthesized using both tabular and narrative approaches. Key information, including study characteristics, types of interventions, outcomes measured, and major findings, was systematically organized and presented in summary tables to facilitate comparison across studies.
The consistency of the results across studies was assessed qualitatively, considering the strength, direction, and relevance of the observed effects.
Subsequently, the discussion section was developed through a comprehensive narrative integration of the findings, highlighting patterns, discrepancies, and the overall interpretation of the evidence within the context of veterinary medicine.
Due to the high degree of methodological and outcome variability among studies, no meta-analysis was performed.
2.7 Protocol registration
This systematic review was prospectively registered in the Open Science Framework (OSF) under the title Pharmaco-toxicological aspects of thymol in veterinary medicine. A systematic review, with the following DOI: https://doi.org/10.17605/OSF.IO/B6SF9. The registration includes the review protocol, eligibility criteria, and synthesis strategy to ensure transparency and reproducibility of the research process.
3 Results
A total of 1,472 records were obtained, of which 176 met the inclusion criteria for this systematic review. Figure 3 shows a bar chart illustrating the distribution of the research articles included, based on the aspects of THY that were studied. “Productive performance” was the most frequently examined aspect, accounting for 19.2% of the articles, followed by “hemato-biochemical” effects (10.2%). Other commonly researched areas included “antioxidant” properties, “digestibility, fermentation, fatty-acid profile,” and “in vitro toxicity,” each representing ~6.5% of the studies. Less studied aspects, such as “intestinal permeability,” “liver morphology,” and “halitosis” accounted for <1% of the total. This distribution highlights a primary focus on productivity and biochemical impacts of THY in animal studies, with fewer studies addressing niche areas. Figure 4 presents THY research studies grouped by species: poultry (broiler chickens, quails, laying hens, ducks), pigs, laboratory animals (rats and mice), fish (tilapia, rainbow trout, and common carp), ruminants (cattle, sheep, and goats), companion animals (dogs and cats), rabbits, and blue foxes. Studies performed with cell lines, primary cell cultures, and pathogens from a particular species were also included.
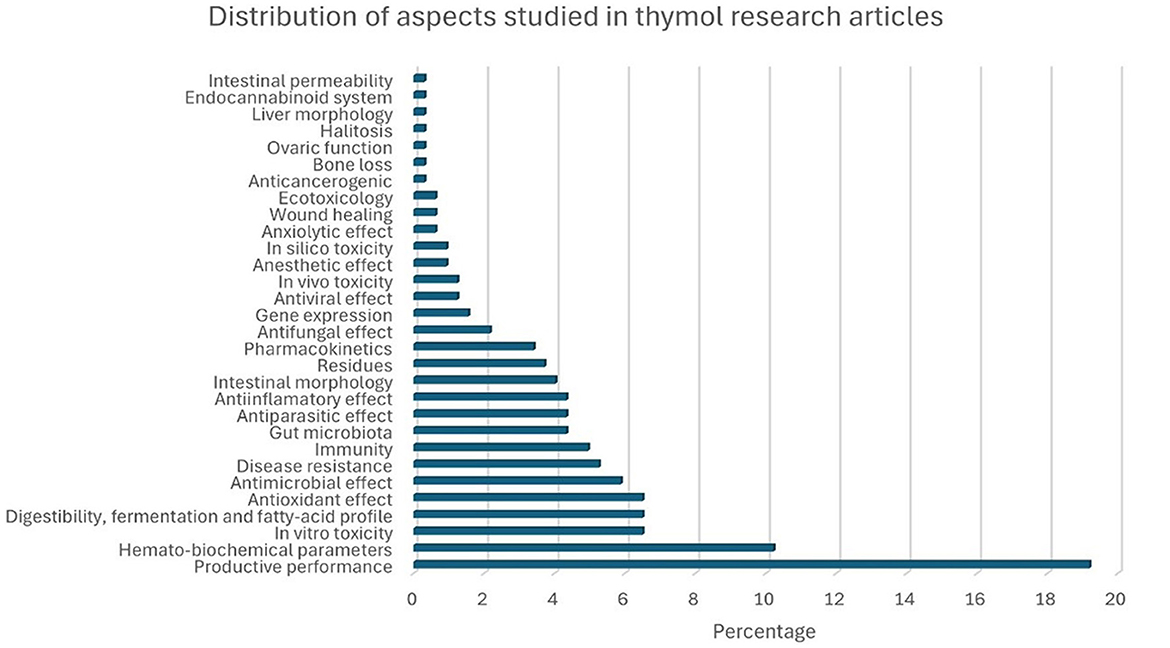
Figure 3. Percentage distribution of research articles by aspect of THY studied.
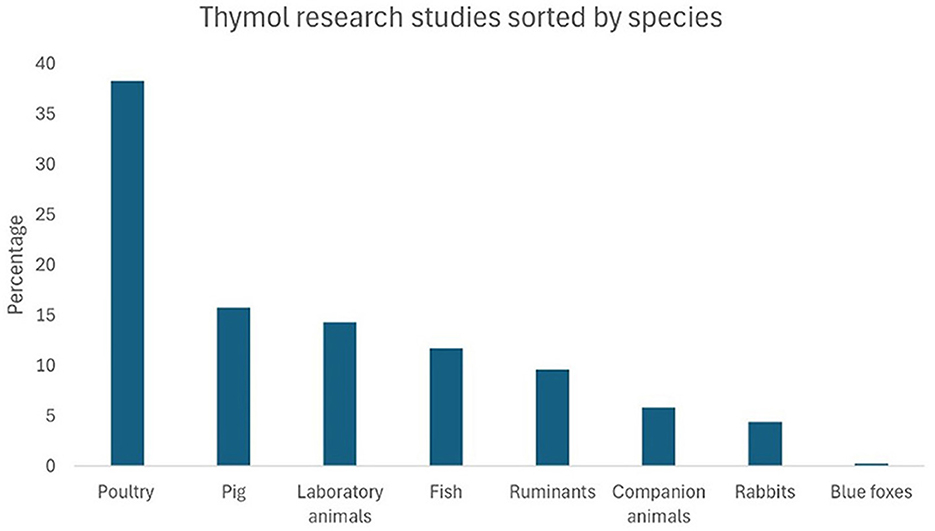
Figure 4. Percentage distribution of THY research articles by species.
3.1 THY toxicity
3.1.1 Cytotoxicity
Table 1 summarizes studies on the cytotoxicity of THY and its derivatives in different cell lines. This table presents the cell types used, the concentrations of THY or THEO to which they were exposed, the methods applied to evaluate cell viability, as well as the specific values of cytotoxicity or effects observed (e.g., LC50, IC50, CC20). The results revealed variability in the cytotoxic response, depending on the cell type and concentrations used.
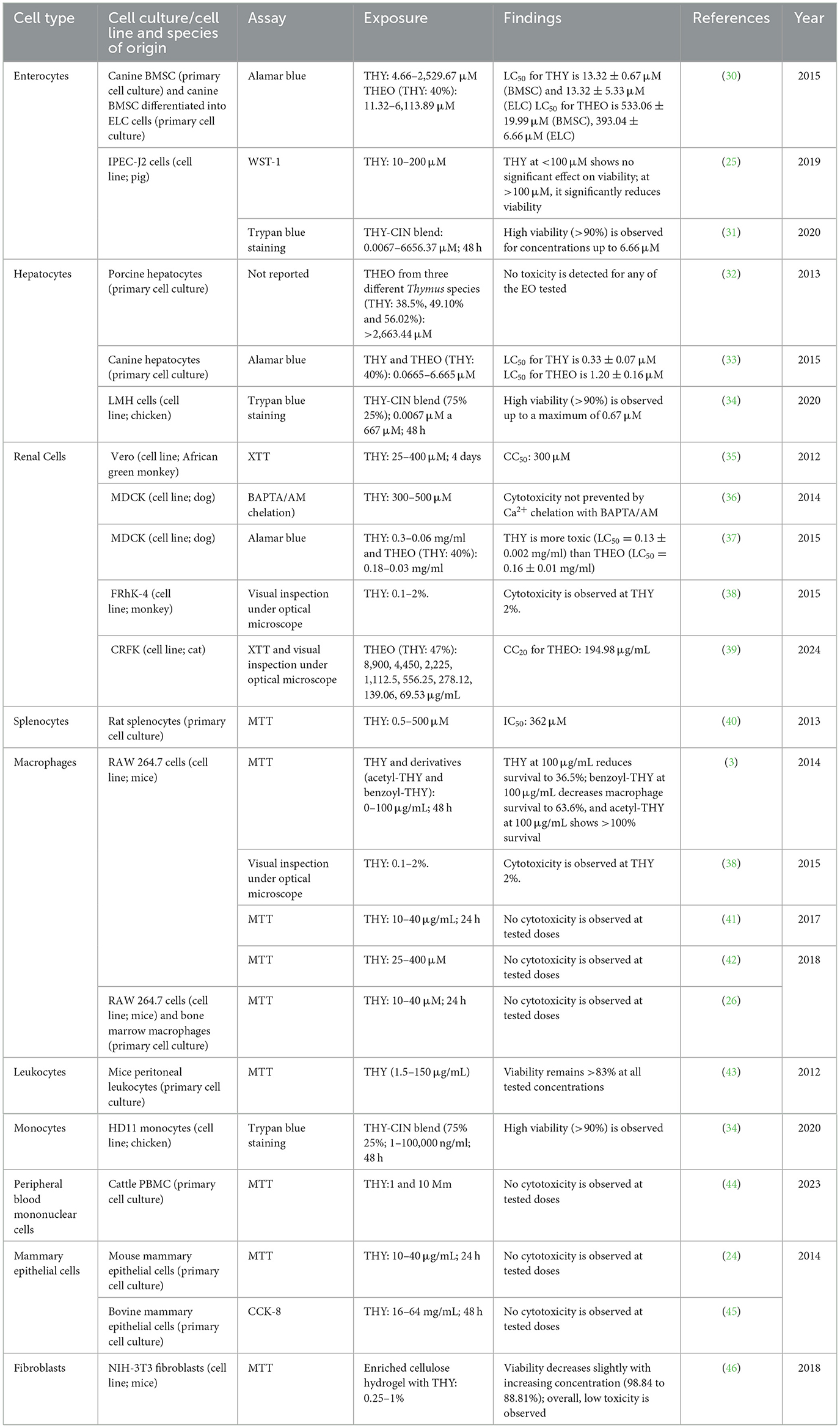
Table 1. THY toxicity in different animal cell lines.
3.1.2 In vivo toxicity
Table 2 presents findings from in vivo studies assessing the toxicity of THY and its derivatives in different animal models. The studies highlighted both acute and sub-chronic toxicity data across various dosages, providing insight into lethal dose thresholds, tissue-specific toxic effects, and potential developmental and estrogenic impacts. In murine models, THY exhibited dose-dependent lethality, with THY acetate showing reduced toxicity, suggesting a higher safety margin. Other studies reported THY's influence on immune cells and inflammatory responses, pointing to immunotoxic effects. In chicken embryos, THY exposure at higher concentrations caused specific developmental abnormalities, whereas in vitro assays revealed weak estrogenic and mutagenic activities below the established thresholds for genotoxicity.
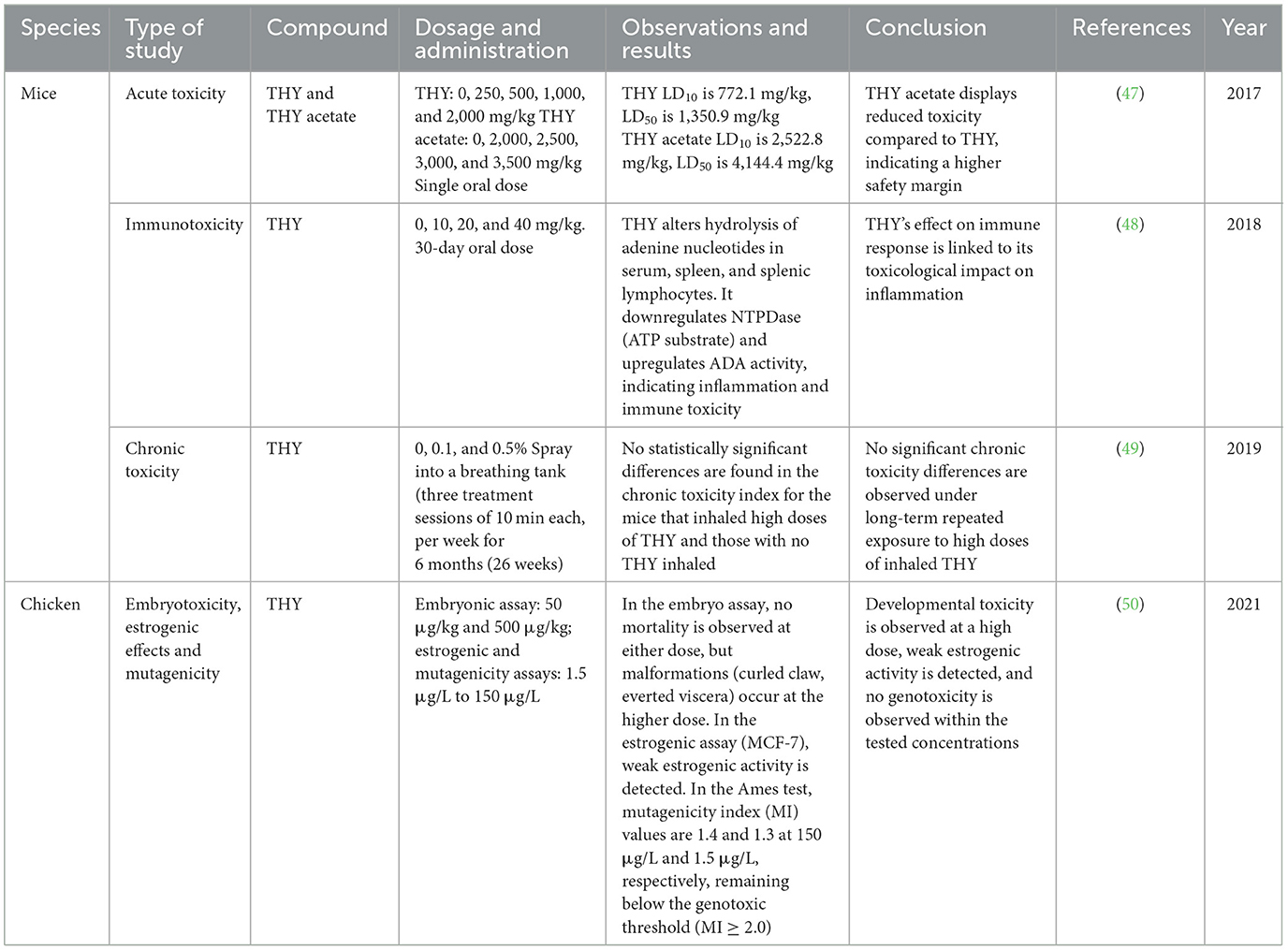
Table 2. In vivo toxicity of THY and THY derivatives in different animal models.
3.1.3 In silico toxicity
In silico studies, the theoretical toxicity and endocrine disruption properties of THY and its derivatives were evaluated. These analyses, which use advanced computational modeling, help predict potential adverse effects and interactions with biological systems without requiring in vivo or in vitro testing. Some studies conducted in silico evaluations, using software to model both general toxicity parameters and hormone receptor interactions. Their findings provided insights into THY's potential as a therapeutic compound, with specific attention to its drug-likeness, resistance-evading potential, and endocrine-disrupting properties (see Table 3).
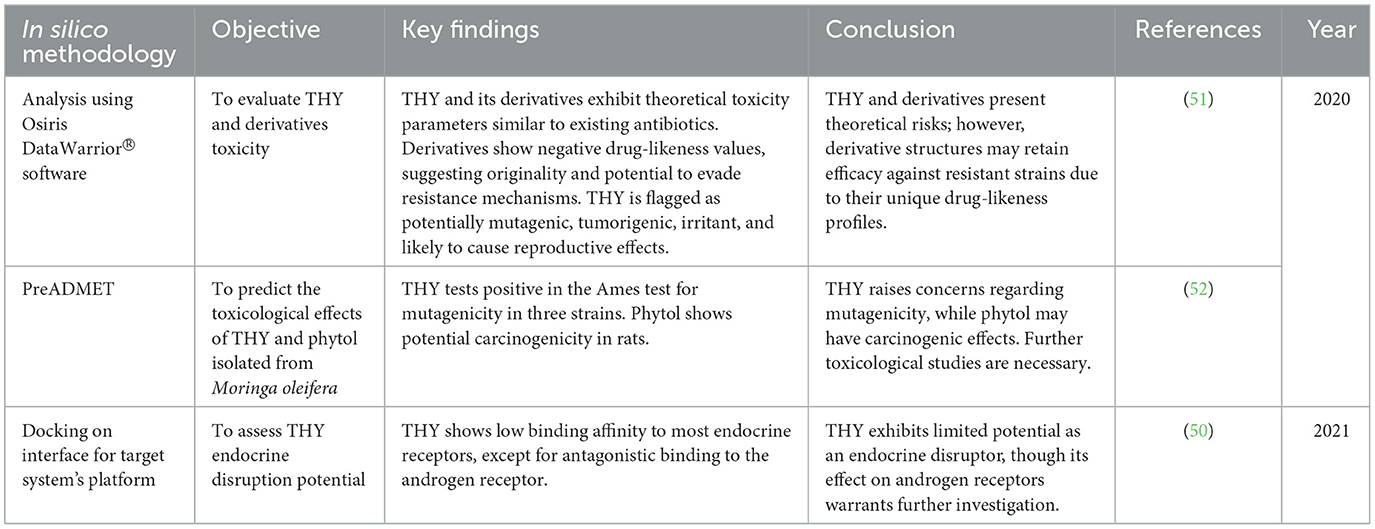
Table 3. In silico toxicity and endocrine disruption potential of THY and its derivatives.
3.1.4 Ecotoxicity
The ecotoxicity of THY has been evaluated through studies assessing its potential impact on non-target organisms, including insects, microbial communities, and bioindicator species. Research has focused on understanding its effects on insect populations involved in ecological processes like nutrient cycling, as well as its influence on soil and aquatic microbial communities, examining parameters such as biodegradation capacity and metabolic activity. Additionally, the toxicity of THY has been tested using standard bioindicators, including aquatic organisms, earthworms, and plants, to determine its environmental safety profile. These investigations aim to provide a comprehensive understanding of thymol's behavior in different ecosystems and its suitability as an alternative to conventional synthetic compounds. These findings, summarized in Table 4, supported THY's potential as a sustainable alternative to synthetic compounds.
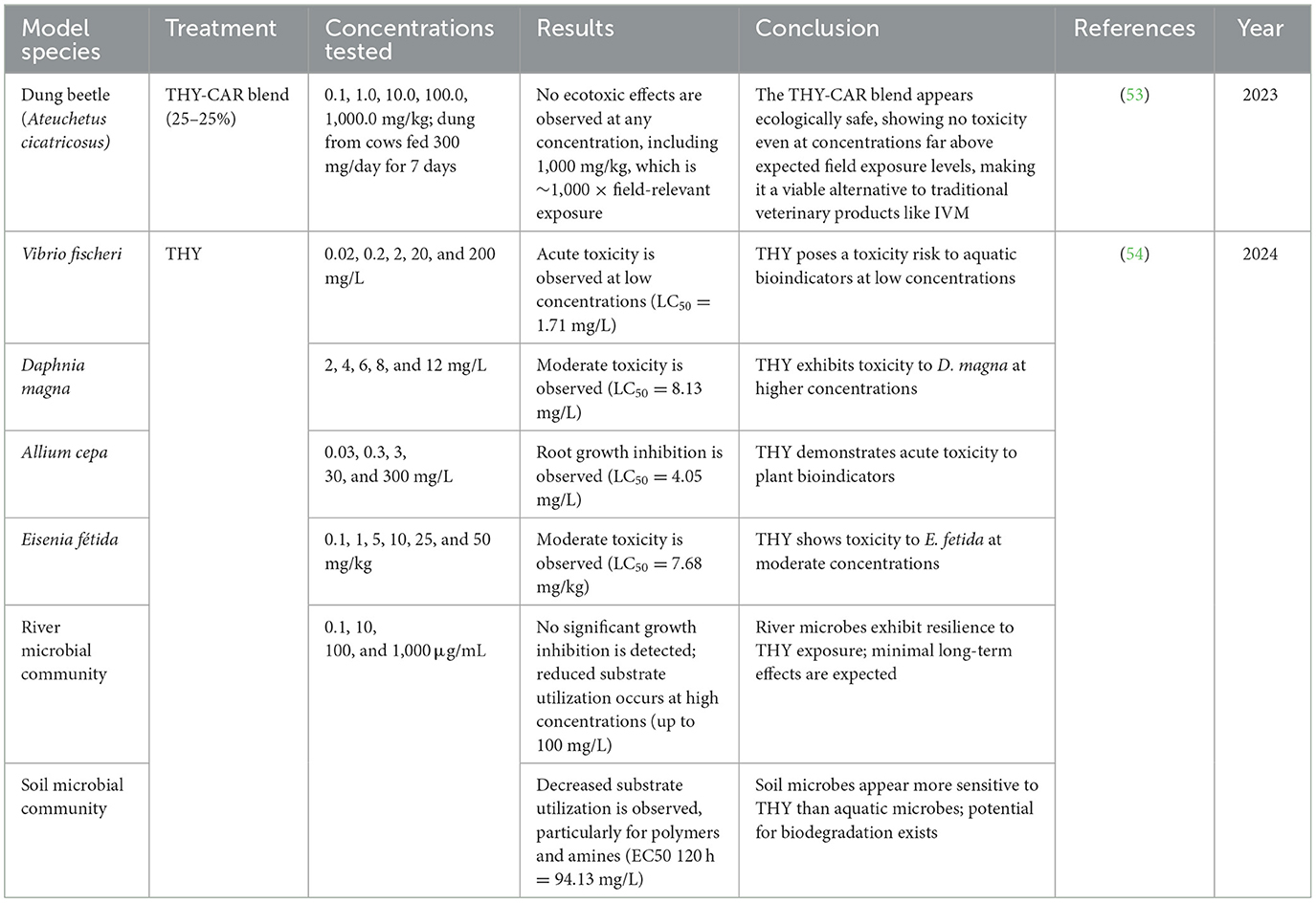
Table 4. Ecotoxicological assessment of THY- and THY-containing blends on non-target insect species.
3.2 THY pharmacokinetics
Table 5 summarizes the pharmacokinetic studies examining the ADME of THY across multiple animal species: rats, dairy cattle, broiler chickens, pigs, and rabbits, and in silico studies. Key pharmacokinetic parameters were investigated following different routes of administration, including intravenous injection, inhalation, oral, intramammary, and feed-supplemented applications.
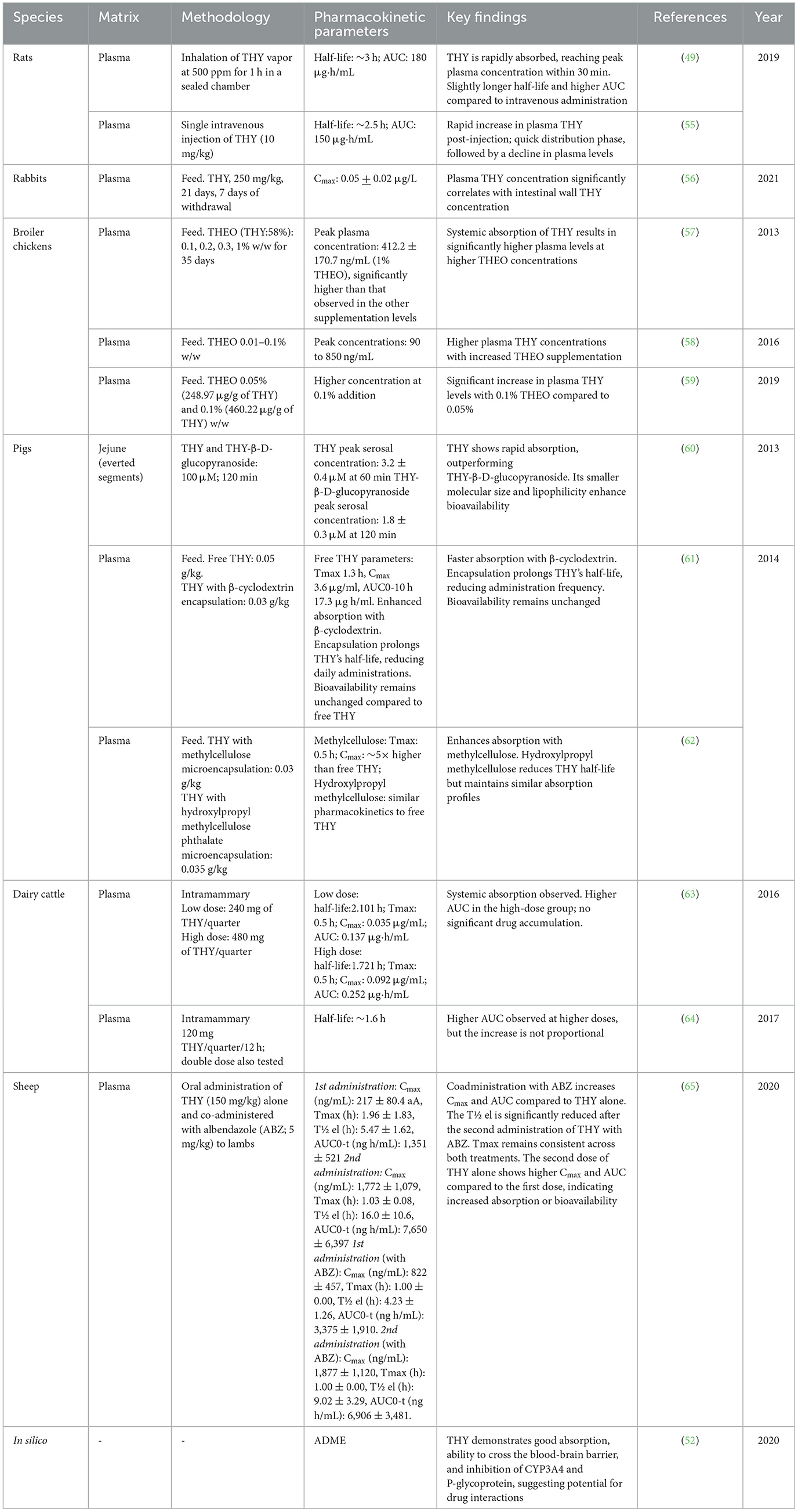
Table 5. Pharmacokinetics of THY in rats, rabbits, broiler chickens, pigs, and dairy cattle.
3.3 THY residue dynamics in dairy cattle, pigs, broiler chickens and quails
Table 6 presents a summary of THY residue studies across dairy cattle, broiler chickens, quails, and pigs, highlighting the absorption, distribution, and persistence of THY in different tissues.
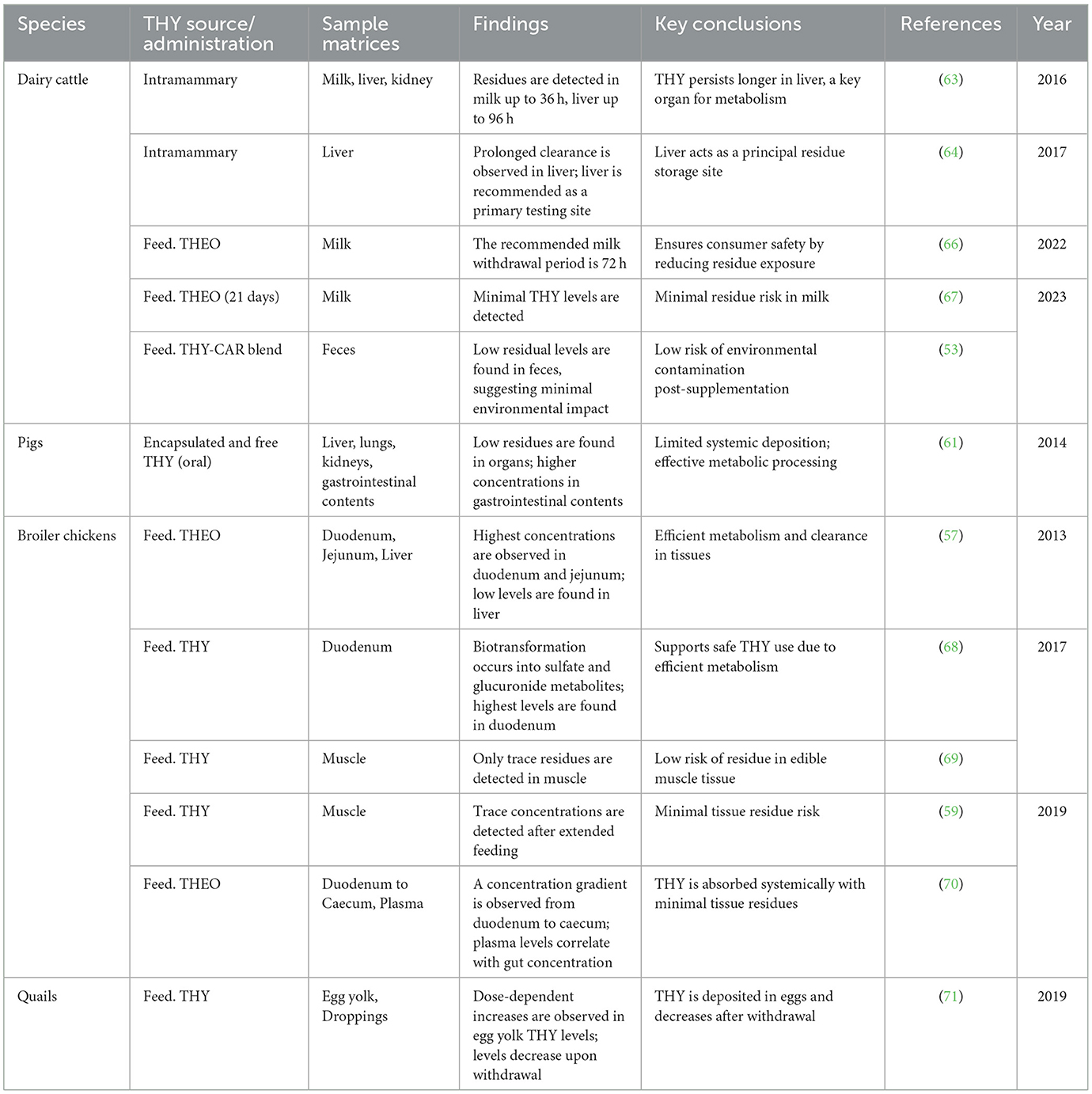
Table 6. Summary of THY residue dynamics in food-producing animals.
3.4 THY interactions with other compounds
Studies on the interactions between THY and other compounds in animal models have remained limited, particularly in the context of its pharmacokinetic and pharmacodynamic effects. The available studies have suggested that THY's interaction at the level of drug-metabolizing enzymes and related reactions, such as oxidation, reduction, and conjugation, may influence the efficacy and metabolic profile of co-administered compounds. However, understanding these interactions has remained an emerging area, with few detailed assessments, especially concerning THY's impact on ADME in production animals. In one of the few studies addressing this topic (29), the chemical-drug interaction between THY (150 mg/kg, administered two times every 24 h) and the antiparasitic agent ABZ (5 mg/kg) in lambs with resistant gastrointestinal nematodes was investigated. The study has found minimal metabolism of THY within the ruminal content and a low degree of association with the particulate phase. Notably, the pharmacokinetics of ABZ sulphoxide were unaffected by the presence of THY. However, co-administration has led to a significant reduction in the Cmax and AUC of ABZ sulfone compared to ABZ-only treatment. Additionally, the presence of THY has not enhanced ABZ's efficacy but notably inhibited the ruminal sulphoreduction and hepatic sulphonation of ABZ sulphoxide.
3.5 THY pharmacodynamics
THY exerts a broad spectrum of biological activities through multiple mechanisms of action. It disrupts cell membranes by integrating into the lipid bilayer, which increases membrane permeability and causes the leakage of essential intracellular components, ultimately leading to cell death (72). This membrane-disrupting effect is central to its antibacterial properties, as THY also inhibits quorum sensing in bacteria, reducing bacterial communication, biofilm formation, and virulence. Furthermore, THY damages bacterial DNA and RNA, impairing replication and transcription, and it inhibits key metabolic enzymes, such as adenosine triphosphatases (ATPases) and glycolytic enzymes, vital for bacterial energy production. These combined actions contribute to THY's effectiveness as an antibacterial agent. In addition to its antibacterial effects, THY demonstrates potent antiviral activity by destabilizing viral envelopes or capsids, inhibiting viral replication, and interfering with viral entry into host cells. It also exhibits antileishmanial activity through its interaction with membrane sterols, disrupting membrane integrity and fluidity, leading to oxidative stress and cellular damage in parasites. Furthermore, in parasitic cells, THY triggers chromatin condensation, DNA fragmentation, and mitochondrial dysfunction, which are hallmark features of apoptosis-like cell death. THY's antifungal effects are evident through its disruption of fungal cell membranes by altering fatty acid metabolism, reducing ergosterol content, and increasing reactive oxygen species (ROS), ultimately causing oxidative stress and fungal cell death.
THY's ability to induce apoptosis is another key feature of its biological activity. It induces apoptosis in cancer cells via mitochondrial depolarization, activation of the Bax protein, caspase activation, and increased ROS levels (36), all leading to DNA damage and cell death. This makes THY a promising candidate for anticancer applications (73). In addition to its antimicrobial and anticancer properties, THY also demonstrates significant anti-inflammatory effects. These effects are attributed to its ability to inhibit the recruitment of cytokines and chemokines and modulate NF-κB signaling. This results in the downregulation of pro-inflammatory cytokines (TNF-α, IL-1β, IL-6), cyclooxygenase-2 (COX-2), and inducible nitric oxide synthase (iNOS), thereby reducing nitric oxide production (74). THY also chelates metal ions, such as iron and magnesium, which disrupt enzymatic activity and stabilize biofilms, further contributing to its antimicrobial and anti-inflammatory actions (73). THY's antioxidant properties also play a vital role in protecting cells from oxidative stress. By scavenging free radicals and enhancing the activity of endogenous antioxidant enzymes, such as superoxide dismutase (SOD), glutathione (GSH) peroxidase (GPx), and catalase (CAT), THY supports cellular defense mechanisms and reduces cellular damage caused by oxidative stress. These antioxidant effects contribute to THY's protective role against various diseases, including inflammation and cancer (73, 74). Moreover, THY exhibits antihyperlipidemic effects by increasing high-density lipoprotein (HDL) cholesterol levels while decreasing low-density lipoprotein (LDL) cholesterol in circulation. These actions, together with THY's ability to stabilize cellular membranes and maintain ionic homeostasis, highlight its potential in managing lipid imbalances (74). In addition to its biological effects, THY also exerts anesthetic properties mediated through gamma-aminobutyric acid type A (GABA_A) receptors, though these effects are independent of benzodiazepine binding sites (75). THY also modulates intracellular calcium flux by interacting with calcium channels like TRPA1, which influences cellular signaling and physiological responses, including mast cell degranulation (73). Additionally, at low doses THY acts as an agonist of α1-, α2-, and β-adrenergic receptors; however, at higher doses, it behaves as an antagonist, thus demonstrating its efficacy in reducing ileal contractions (76) and enhancing the absorptive capacity of the intestinal mucosa as a dietary supplement (16). These diverse mechanisms of action underscore THY's potential as a natural, multifunctional agent for veterinary and therapeutic applications.
3.6 THY pharmacological effects
3.6.1 Antibacterial properties of THY and THEO
THY is widely studied for its potent antibacterial effects. Research indicates that THY disrupts bacterial cell membranes, leading to the leakage of cellular contents and eventual bacterial death. Studies on both Gram-positive and Gram-negative bacteria show that THY is effective across a range of bacterial strains, significantly reducing colony-forming units (CFUs) in a dose-dependent way. Furthermore, the structure of THY, with its phenolic groups, plays a critical role in its antibacterial activity by enhancing its hydrophobic interactions with bacterial cell membranes. Table 7 details the source of THY, the pathogenic strains tested, methods employed to determine efficacy, and minimum inhibitory concentrations (MIC) or minimum bactericidal/fungicidal concentrations (MBC/MFC) reported in each study. Additional relevant observations about THY antimicrobial mechanisms, resistance potential, and synergistic effects are noted, thereby offering insights into THY's potential applications and limitations as an alternative antimicrobial agent.
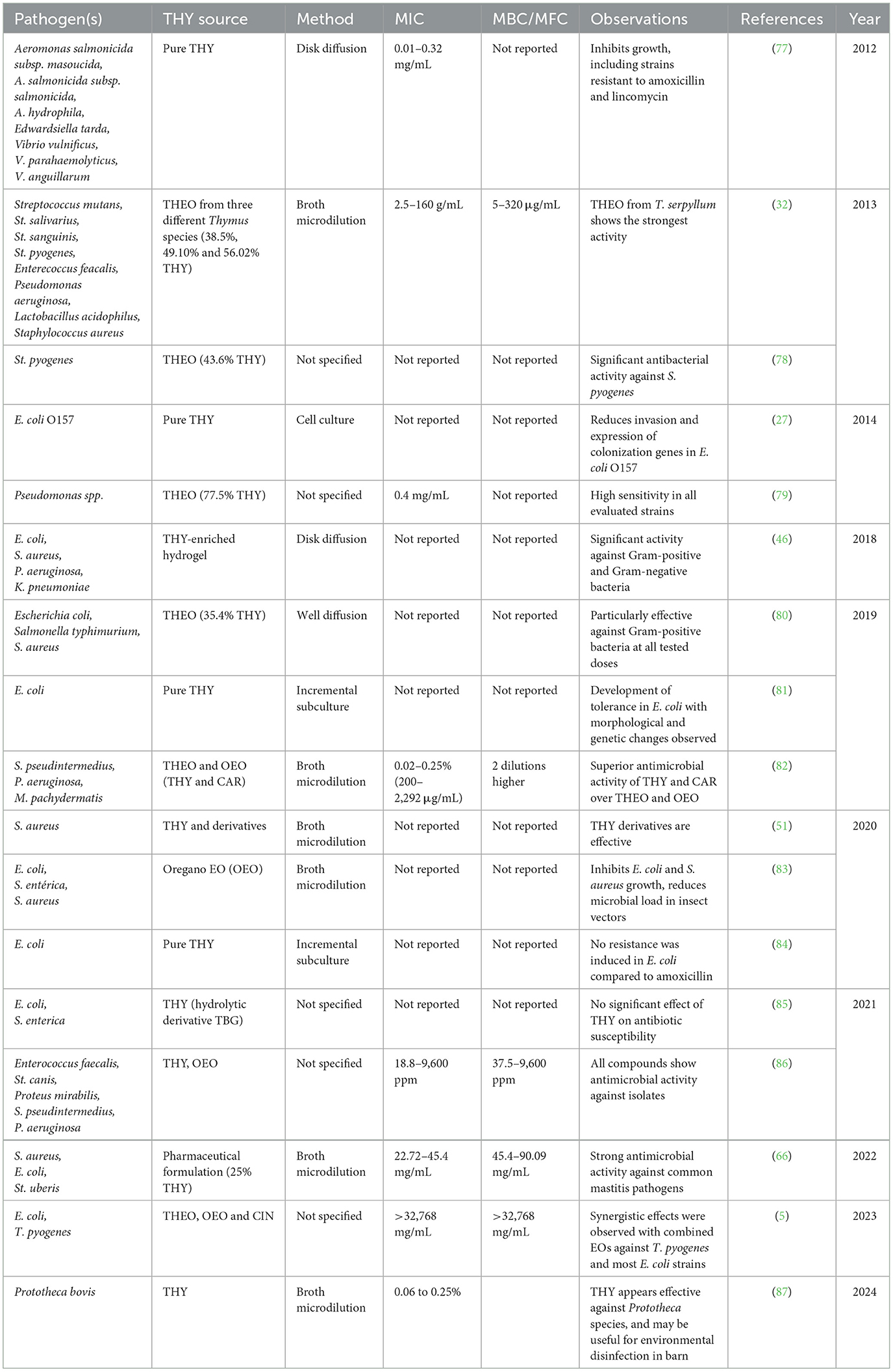
Table 7. Summary of studies on the antimicrobial efficacy of THY and THEO against various pathogens.
3.6.2 Antiviral effects of THY and THEO
THY and THEO exhibit notable antiviral activity against a variety of viruses. Table 8 summarizes studies investigating the antiviral properties of THY and THEO's against different viral pathogens. Key findings, such as the inhibitory concentration (IC50), viral titer reductions, and the specific methods employed to assess antiviral activity, are detailed for each study.
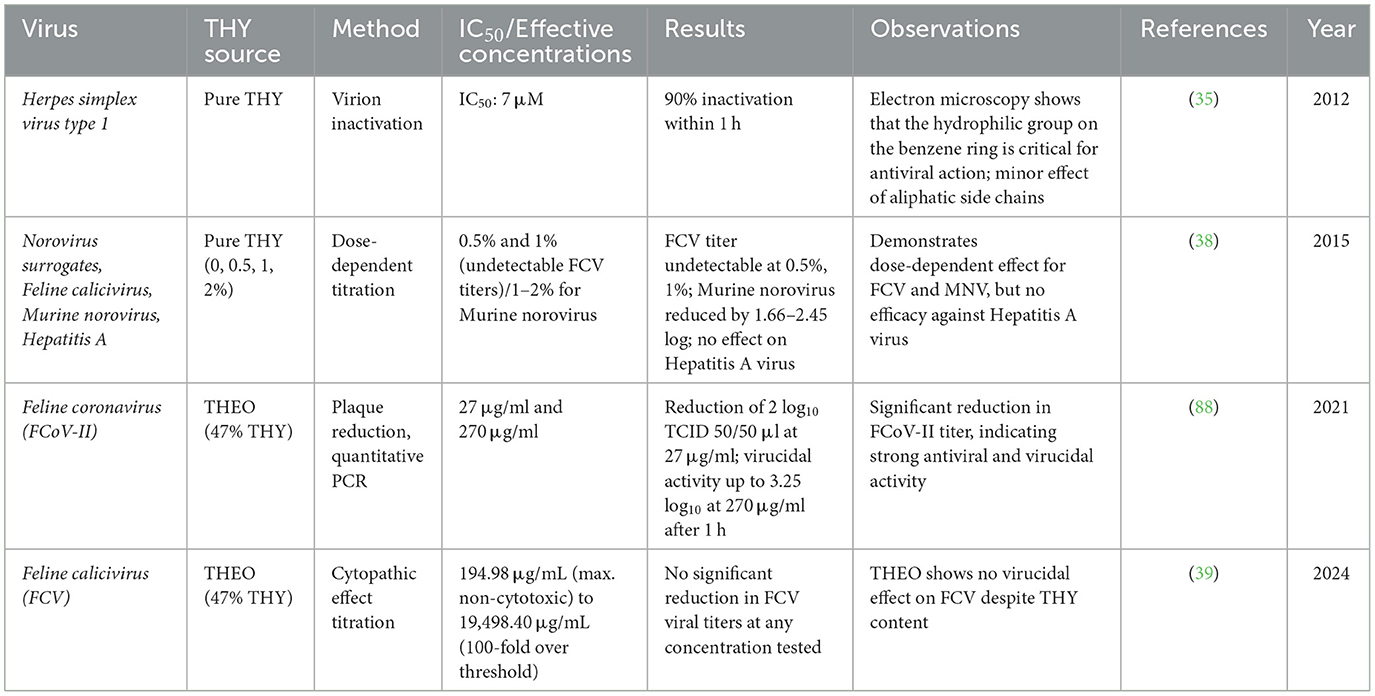
Table 8. Summary of antiviral activity of THY and THEO against various viruses.
3.6.3 Antifungal effect of THY and THEO
The antifungal properties of THY and THEO have been documented against a range of fungal pathogens. Studies report effective inhibition of mycelial growth and spore germination. Table 9 presents data on minimum inhibitory concentrations (MICs) and minimum fungicidal concentrations (MFCs) against various fungal strains, as well as outcomes in studies involving alternative delivery methods, such as vapor-phase application, and in vivo testing on fish survival.
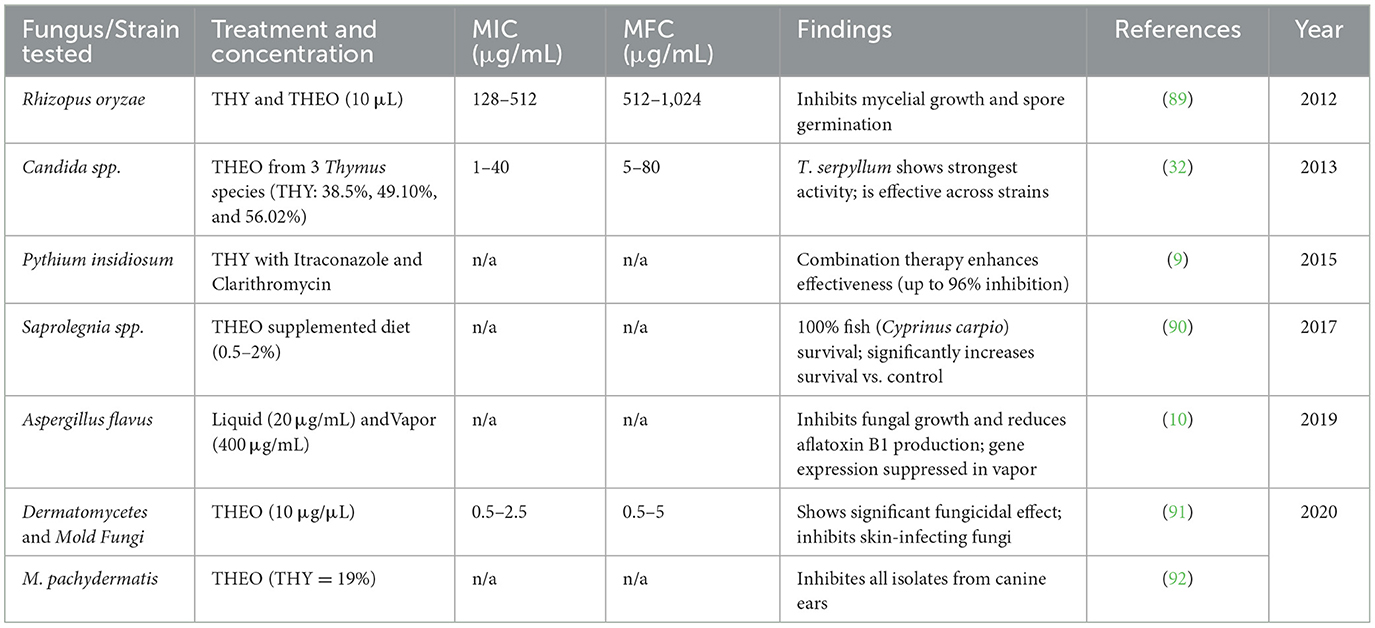
Table 9. Antifungal activity of THY and THEO against various fungal strains.
3.6.4 Antiparasitic effect of THY and THEOs
3.6.4.1 Ectoparasites
THY and THEOs are effective against a range of ectoparasites, including tick species like Rhipicephalus microplus and Ixodes Ricinus and poultry mites. Additionally, THY is proven effective as a repellent and insecticide for pests such as the lesser mealworm (Alphitobius diaperinus) and houseflies (Musca domestica), supporting its role as a versatile natural antiparasitic.
3.6.4.2 Endoparasites
THY shows strong antiparasitic activity against protozoans like Eimeria spp. and Cryptosporidium spp., where it disrupts oocyst walls, suggesting its potential as a disinfectant for poultry. For nematodes, particularly Haemonchus contortus, THY effectively inhibits egg hatching and larval motility, with near-complete suppression in vitro. THY also induces structural damage and mortality in Echinococcus spp. and Mesocestoides corti (cestodes) larvae. Studies on Leishmania spp. further highlight the THY's ability to reduce parasite loads and enhance host survival, indicating its promise in treating leishmaniasis. THY's interaction with drugs like ABZ suggests a potential role as a co-treatment, though its metabolic impact warrants further study.
Table 10 summarizes THY's antiparasitic effects across multiple studies, covering both ecto- and endoparasites, including flies, ticks, mites, and various parasite stages and species like coccidia, nematodes, cestodes, and Leishmania.
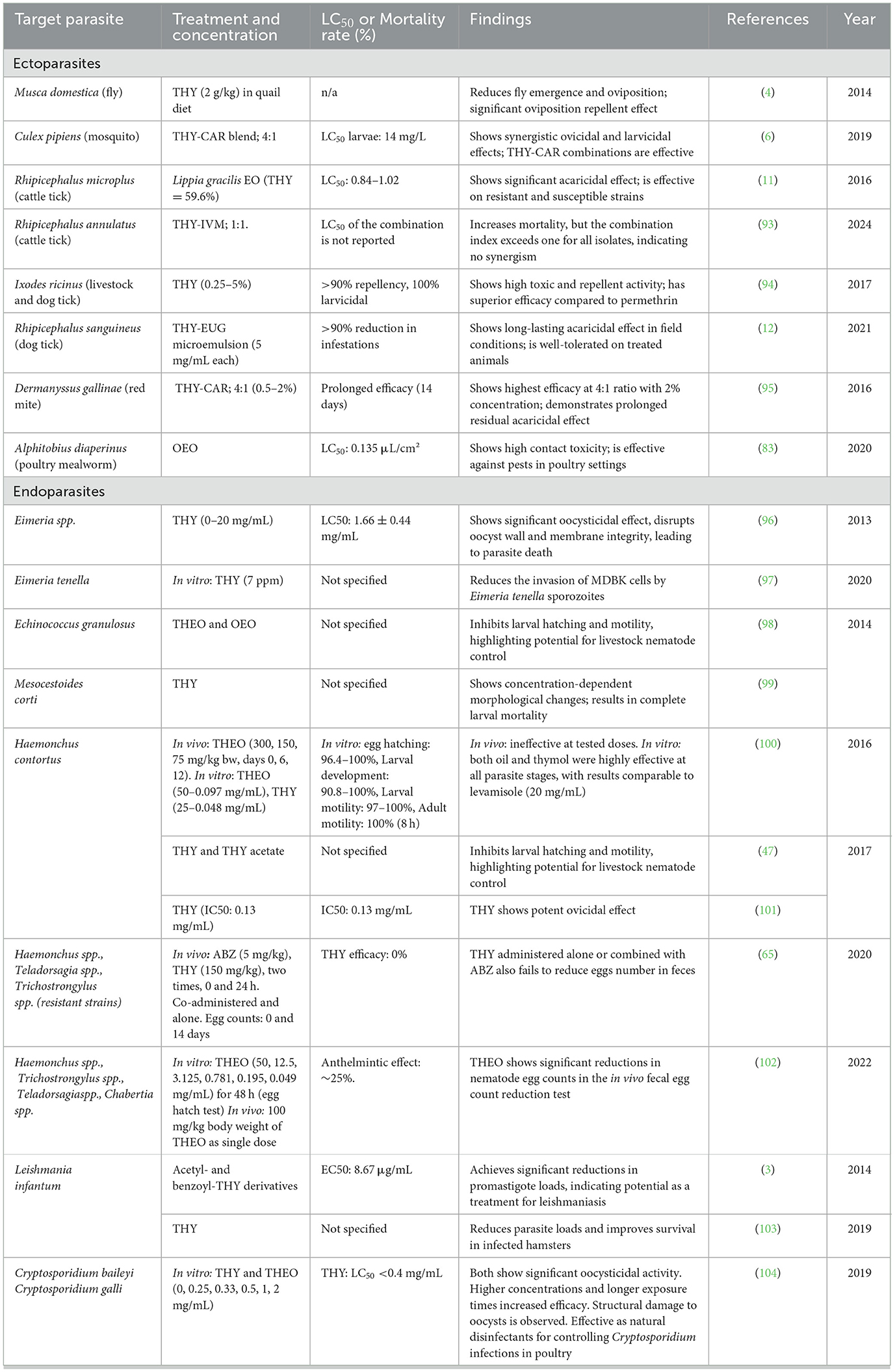
Table 10. Antiparasitic activity of THY and THEOs against various ectoparasites and endoparasites.
3.6.5 Antiinflammatory effects of THY
Data reported in Table 11 highlight THY's anti-inflammatory effects across a variety of animal models and in vitro systems, demonstrating its potential to modulate inflammation through multiple pathways. THY's anti-inflammatory properties are evident in rodent models, avian species, and cellular assays, with studies reporting reductions in edema, leukocyte infiltration, and pro-inflammatory cytokine expression. The studies utilized diverse methods, including THY alone or in combination with CAR, administered through various doses and routes. Key indicators of inflammation, such as cytokine levels (e.g., TNF-α, IL-1β), myeloperoxidase (MPO) activity, and oxidative stress markers, were measured to assess THY's effects.
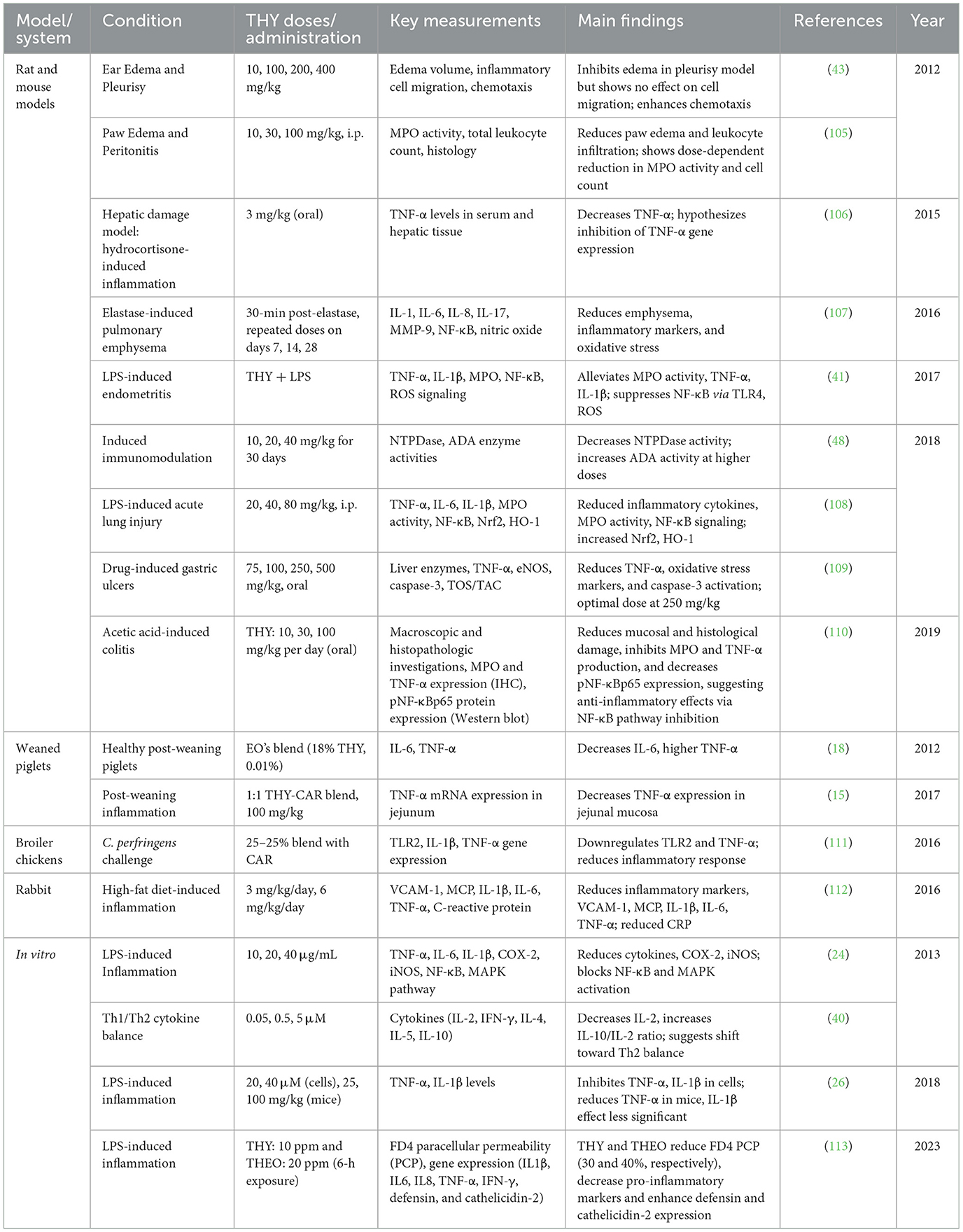
Table 11. Summary of studies evaluating the anti-inflammatory effects of THY in veterinary species.
3.6.6 Antioxidant effects of THY across animal models and in vitro systems
Table 12 provides a summary of studies that evaluated the antioxidant effects of THY across multiple species and models. THY consistently enhances antioxidant activity through reductions in lipid peroxidation markers, such as malondialdehyde (MDA), and increases in enzymatic antioxidants like SOD and GPx. These effects are observed across aquatic, mammalian, and avian species, as well as in in vitro studies, highlighting THY's broad-spectrum potential to mitigate oxidative stress.
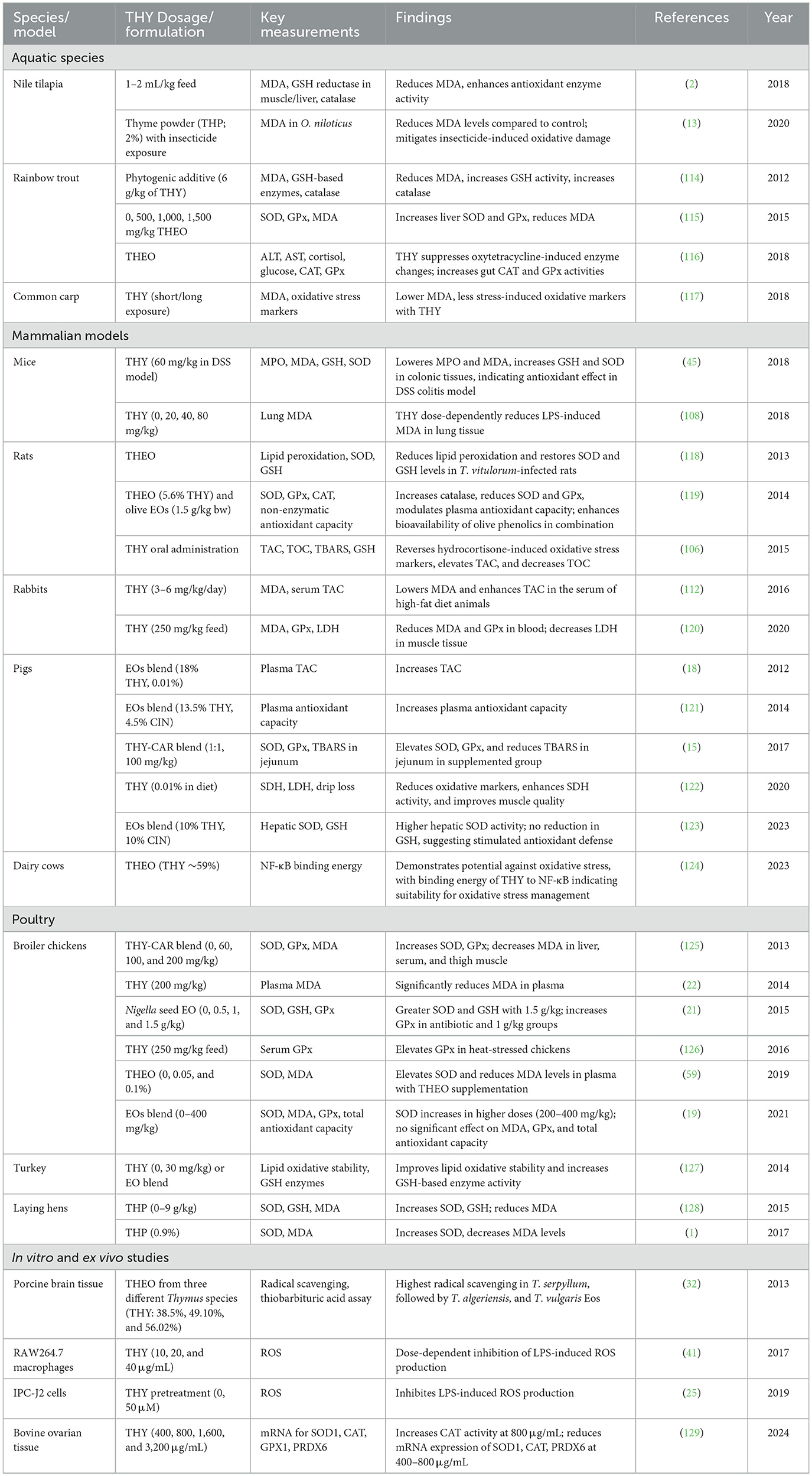
Table 12. Antioxidant effects of THY across multiple species and models.
3.6.7 Hemato-biochemical effects of THY, THEO, and other EOs containing THY
The hemato-biochemical effects of THY and THEO have been widely studied across various animal models. Across species, THY and THEO consistently show beneficial effects. Table 13 summarizes findings from studies on their effects in multiple species, including mammals, poultry, fish, and other livestock animals.
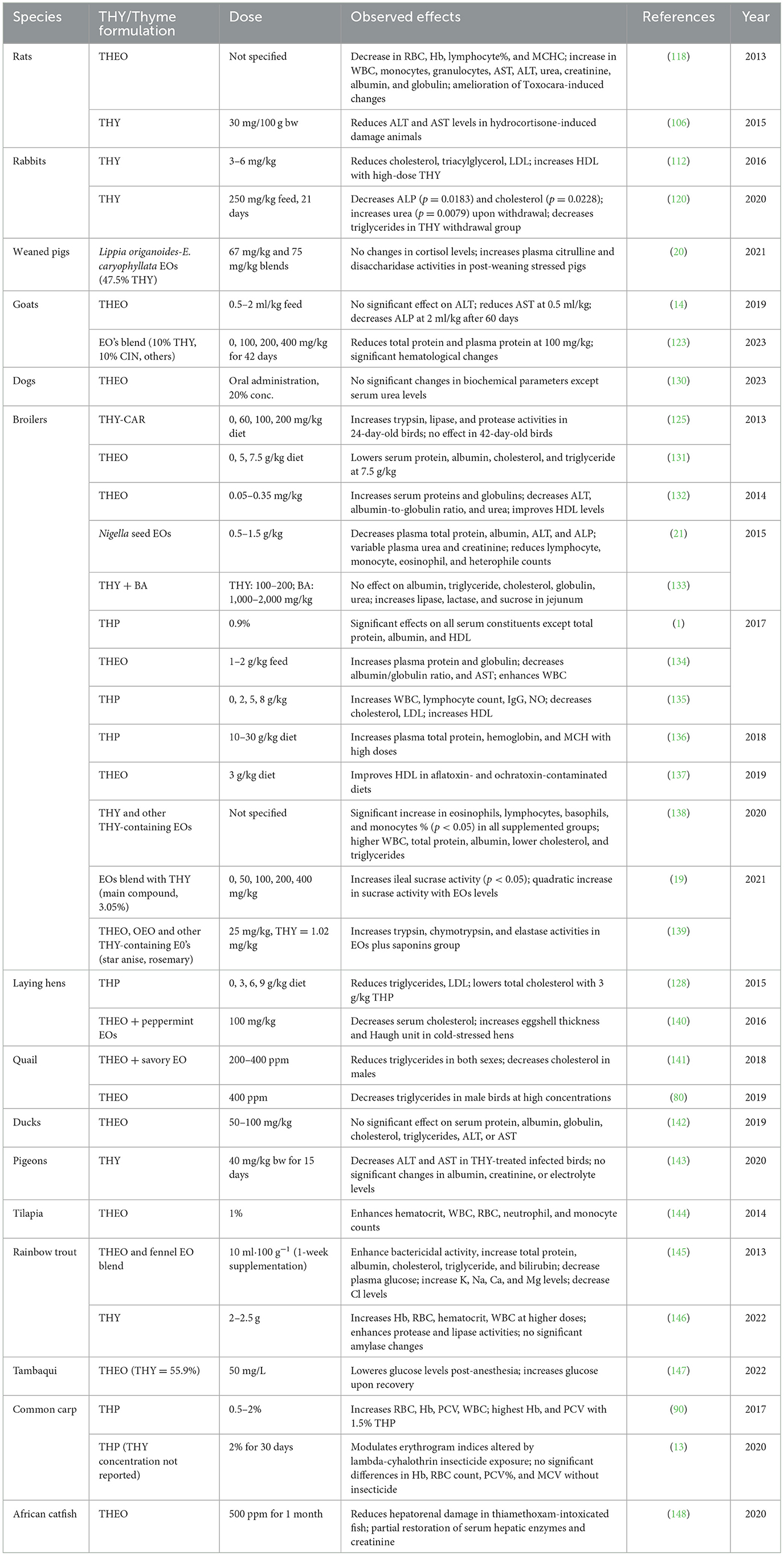
Table 13. Effects of THY, THEO, and other THY-containing EOs on hemato-biochemical indices in animal models.
3.6.8 THY and THEO supplementation effects on animal immunity
THY and THEO supplementation have shown positive impacts on immune function across multiple species, including poultry, fish, and mammals. Together, these findings underscore the potential of THY and THEO as dietary additives to support immune resilience in both livestock and aquaculture, though effects are dosage- and species-dependent. Table 14 summarizes findings from studies that evaluated immune responses to dietary THY or THEO supplementation in various animal species.
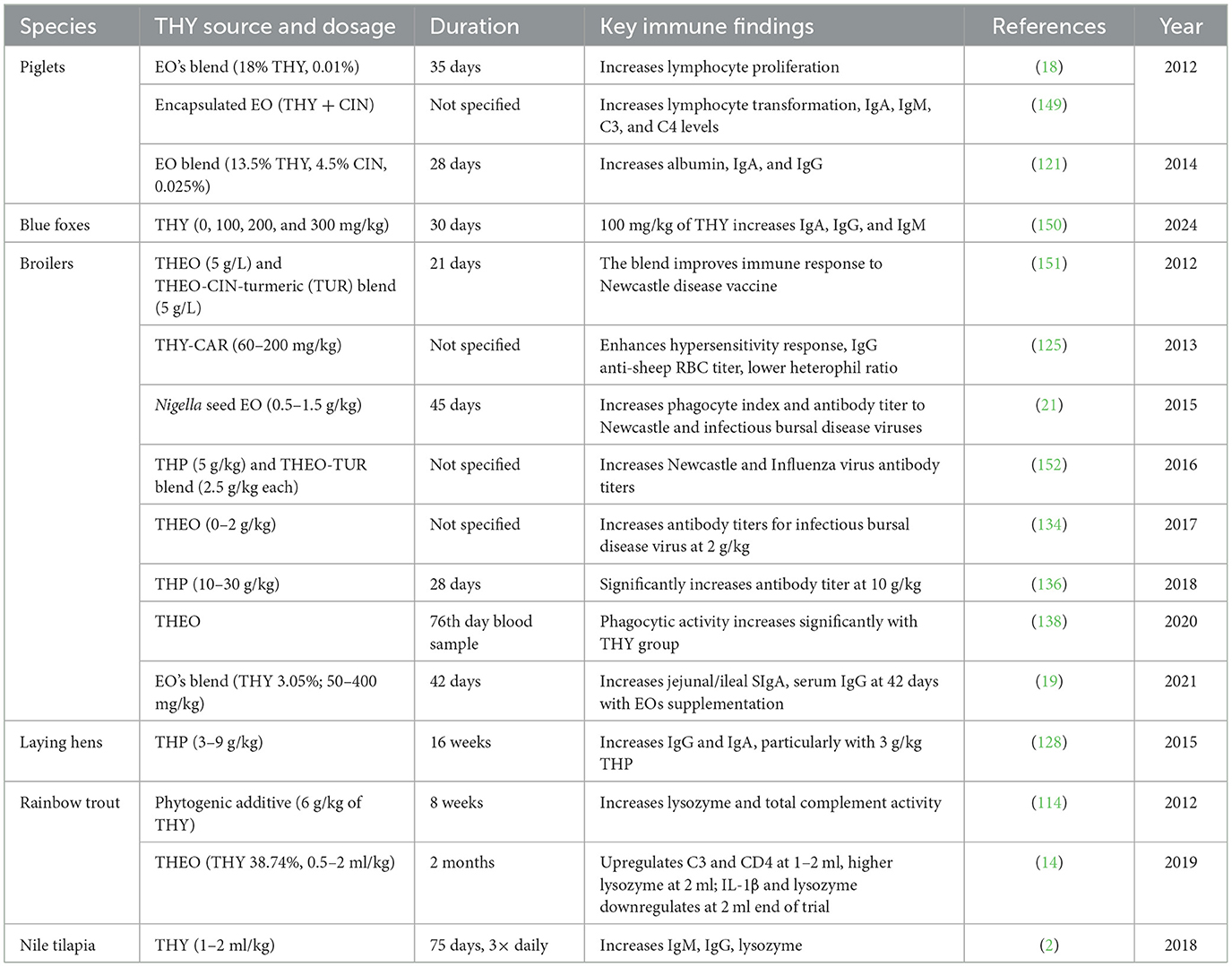
Table 14. Summary of immune responses to THY, THEO, and different blends of supplementation across animal species.
3.6.9 THY effects on disease resistance
Several studies examined the pharmaco-toxicological properties of THY, with a particular focus on its effects on disease resistance, immune modulation, and cellular response across various species, including poultry, livestock, fish, and rabbits. Researchers have explored the role of THY in combatting bacterial pathogens such as Campylobacter spp., Salmonella spp., S. aureus, and St. iniae, as well as its potential in reducing the impact of environmental stressors, including mycotoxin contamination, pesticide exposure, and heat-induced stress. Many studies underscored THY's ability to modulate immune responses, demonstrating increased antibody production, enhanced leukocyte activity, and decreased pathogen colonization in animal models, as previously mentioned. These findings offer promising insights into the therapeutic applications of THY as a natural alternative for enhancing disease resistance and mitigating toxicological impacts in veterinary medicine. Table 15 presents a comprehensive overview of studies evaluating the effects of THY and THEO on disease resistance in various animal species.
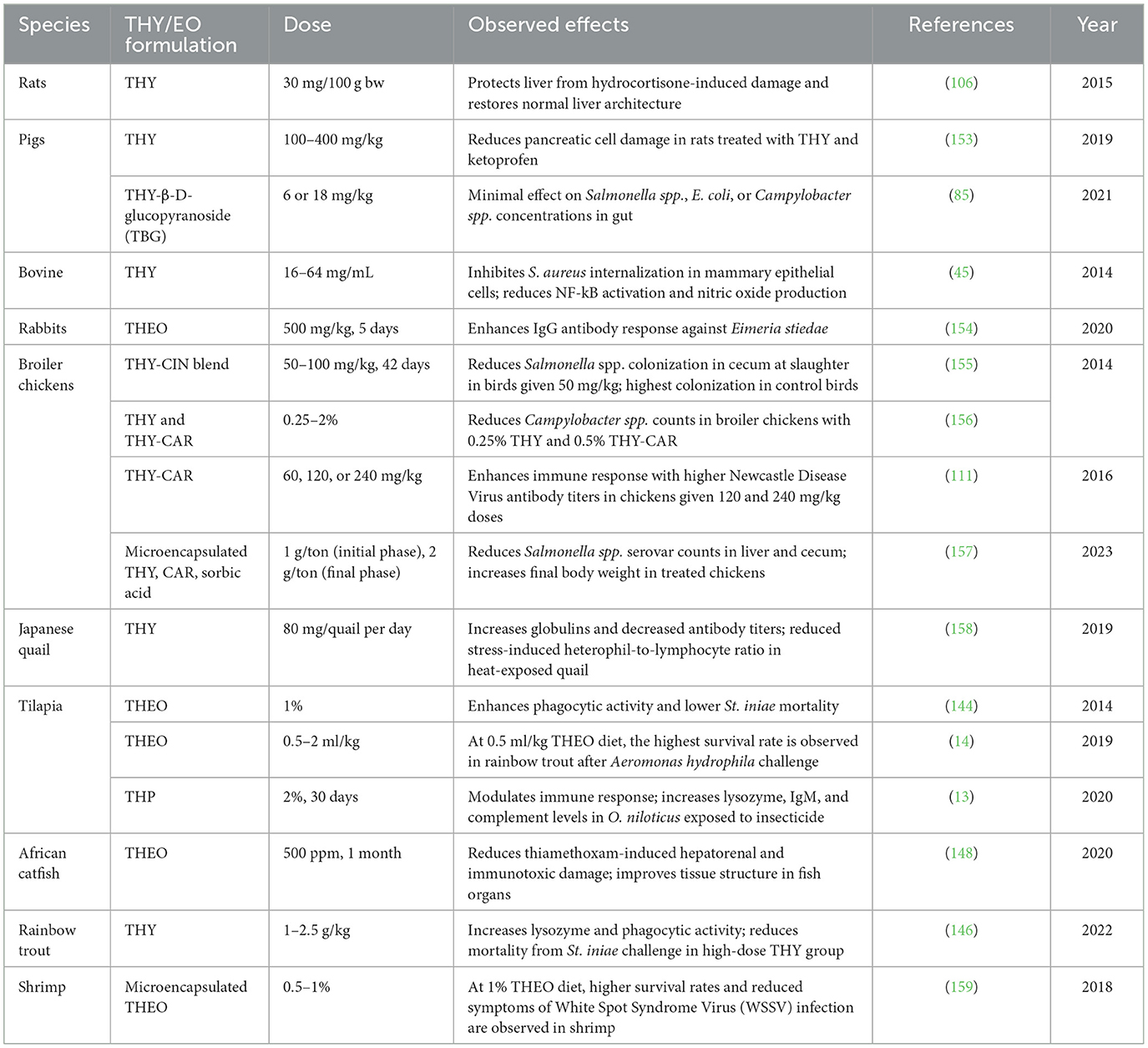
Table 15. Disease resistance effects of THY, THEO, and different blend supplementation in animal models.
3.6.10 THY effects on productive performance parameters
The presented tables summarize studies investigating the impact of THY and THEO-based dietary supplements across various animal species, including poultry (Table 16), mammals (Table 17), and fish (Table 18). These studies are focused on evaluating the effects of different sources, concentrations, and durations of THY or THEO supplementation on productive parameters such as body weight, feed conversion ratio (FCR), growth rate, and other relevant metrics.
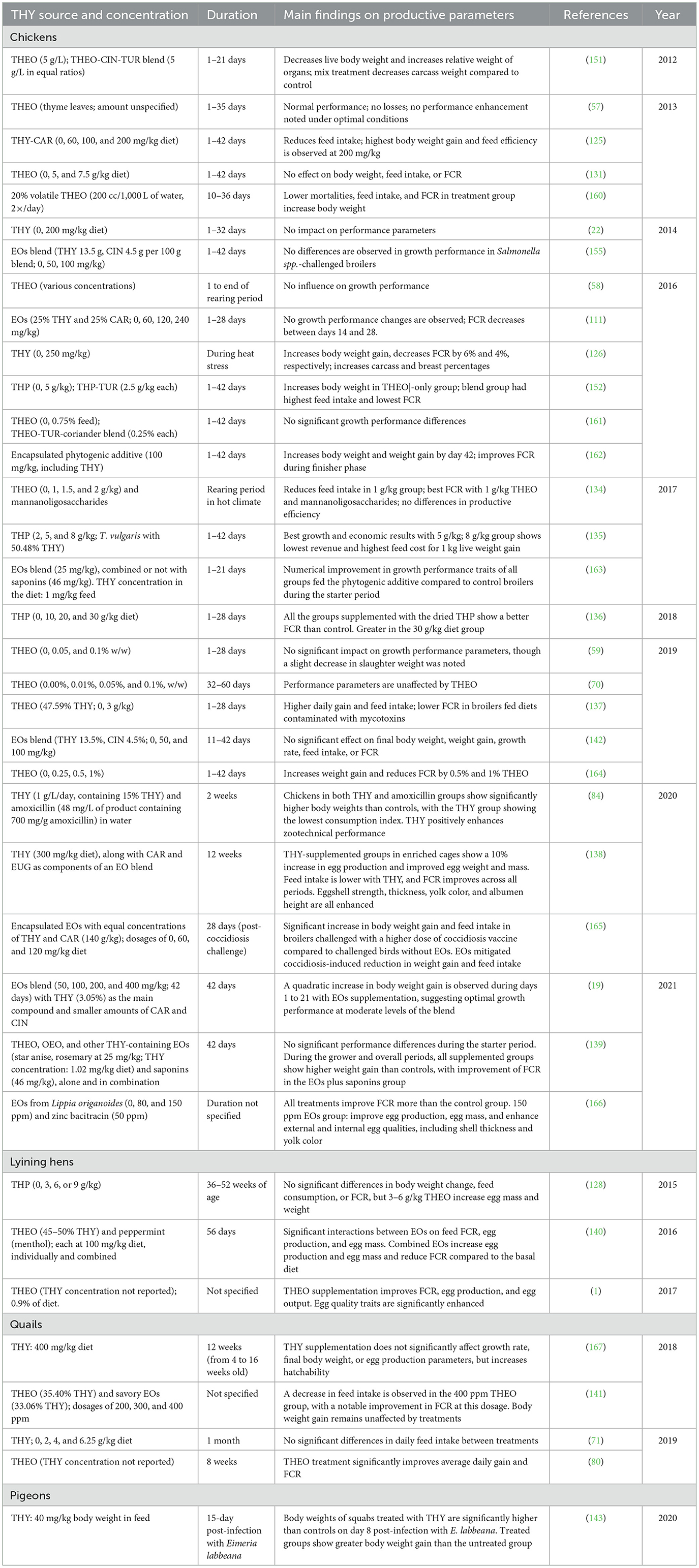
Table 16. Productive performance in birds fed with a diet supplemented with a THY source.
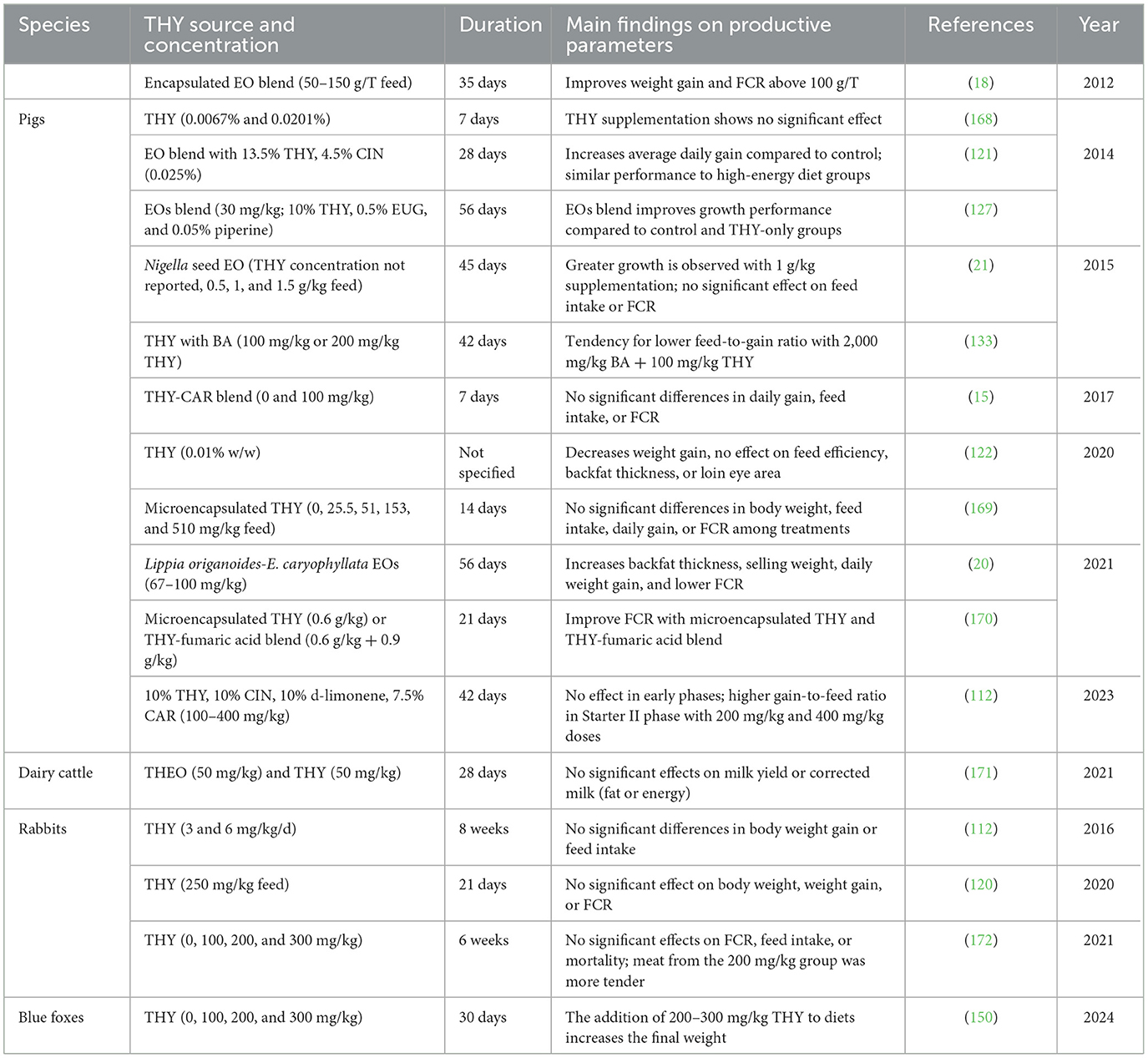
Table 17. Productive performance in mammals fed with a diet supplemented with a THY source.
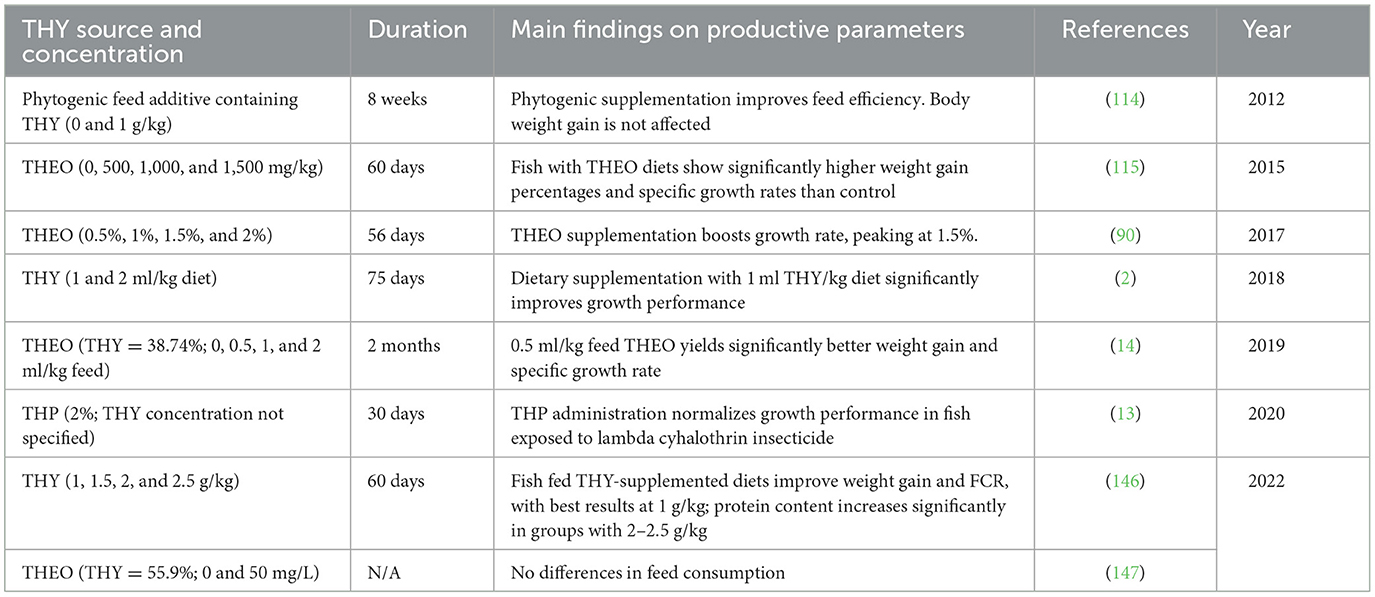
Table 18. Productive performance in fish fed with a diet supplemented with a THY source.
3.6.11 Effects of THY on digestibility, fermentation, and fatty acid profile
Studies consistently show that THY and THEO-based additives positively affect these parameters in certain animal species, though results vary based on dose and the target animal species. Table 19 synthesizes recent research findings on the impact of THY and THEO-based supplements on nutrient digestibility, microbial fermentation, and fatty acid composition. This summary includes studies on various livestock animals and pets, detailing each study's design, THY dosage, treatment duration, and key results.
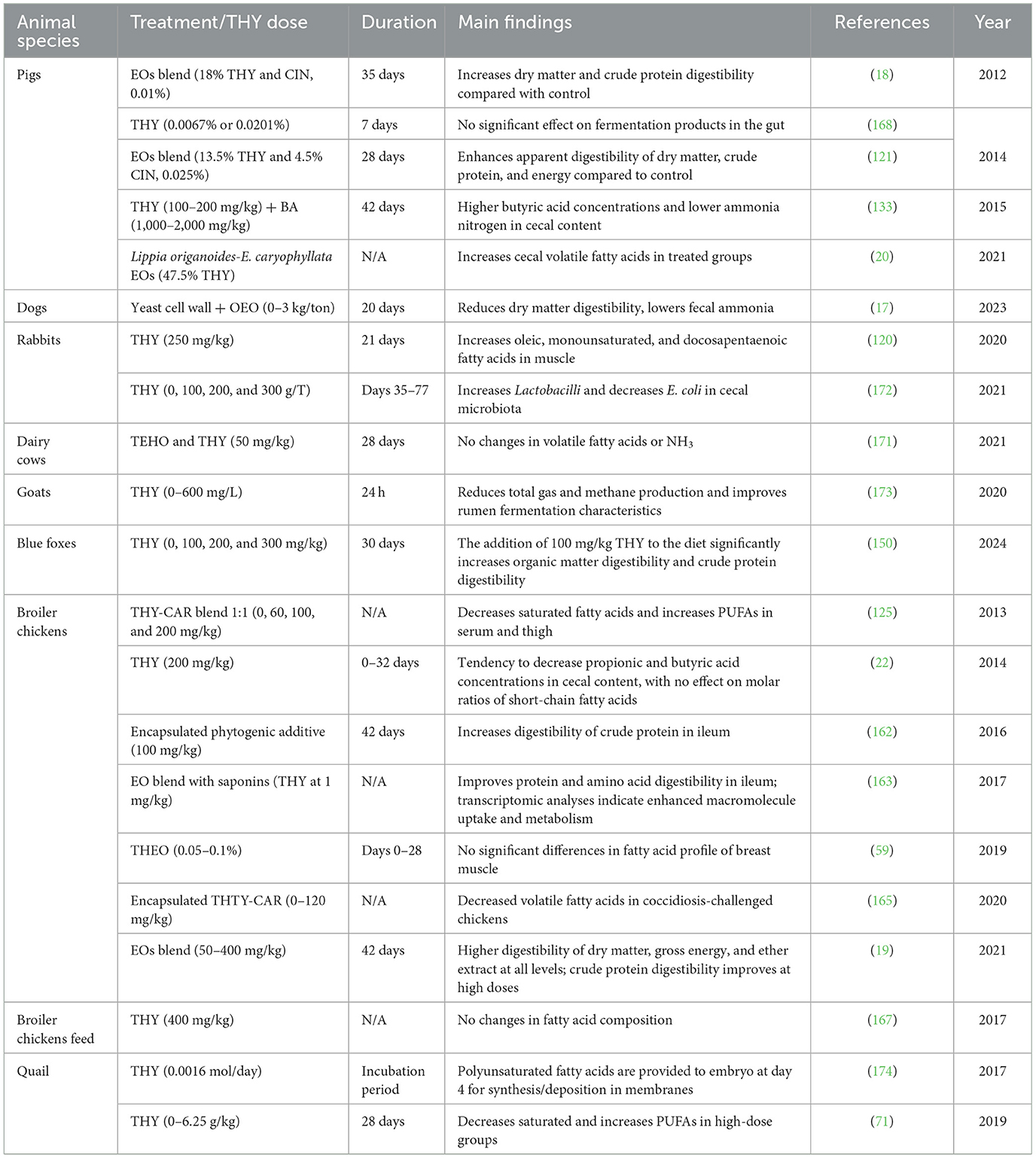
Table 19. Effects of THY- and THEO-based supplements on digestibility, fermentation, and fatty acid profile in various animal species.
3.6.12 THY effects on gut microbiota
THY supplementation has been shown to positively impact gut health by modulating microbial populations, often reducing pathogenic bacteria, such as E. coli and coliforms, while supporting the growth of beneficial microorganisms like Lactobacillus spp. and other lactic acid bacteria. Changes in microbial diversity have also been observed, with certain studies reporting an increased abundance of beneficial taxa following THY supplementation. However, the effects of thymol can vary depending on factors such as dosage, species, and the specific gut region studied, with some reports showing no significant shifts in microbiota composition. Overall, evidence suggests that THY's influence on intestinal microbiota is context-dependent and may differ across animal species, including fish, pigs, poultry, and dogs. Table 20 summarizes findings from studies evaluating these effects.
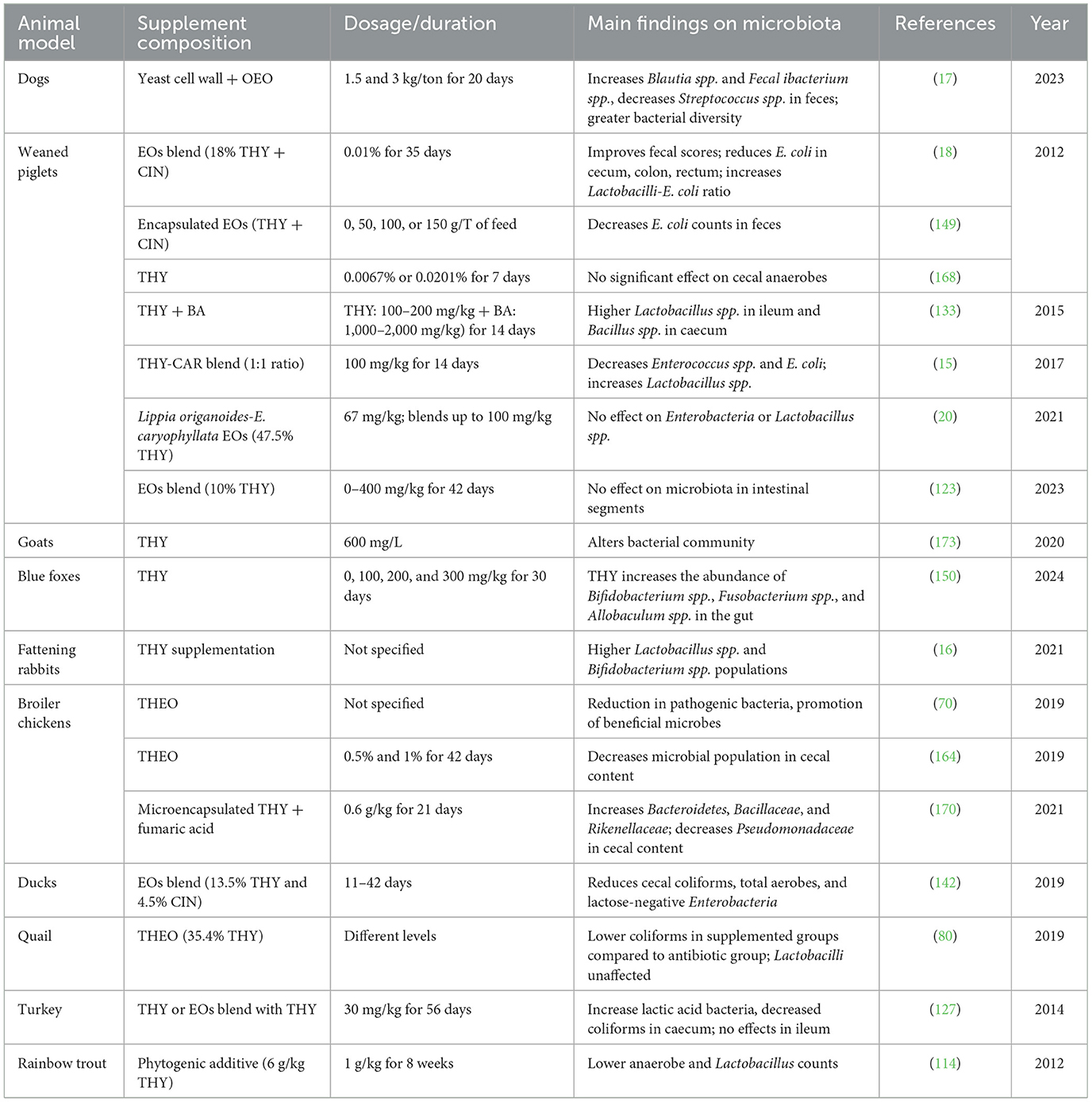
Table 20. Summary of THY's effects on gut microbiota across animal models.
3.6.13 THY effects on intestinal permeability, tight junctions and gastric mucosa gene expression and modulation of the endocannabinoid system in various animal models
In various animal models and in vitro systems, researchers assessed how THY influences tight junction proteins, nutrient transport genes, markers of epithelial integrity, and components of the endocannabinoid and chemosensing systems under both normal and inflammatory conditions. These studies collectively investigated THY's potential for enhancing gut function, maintaining epithelial resilience, and modulating signaling pathways involved in gastrointestinal homeostasis. Table 21 provides a comprehensive overview of key studies, detailing the effects of THY on target gene expression profiles related to barrier function (e.g., claudin-1, occludin), ion transport, gastrointestinal maturation (e.g., somatostatin, peptide transporter 1), and the endocannabinoid system, including cannabinoid receptors and enzymes involved in endocannabinoid biosynthesis and degradation.
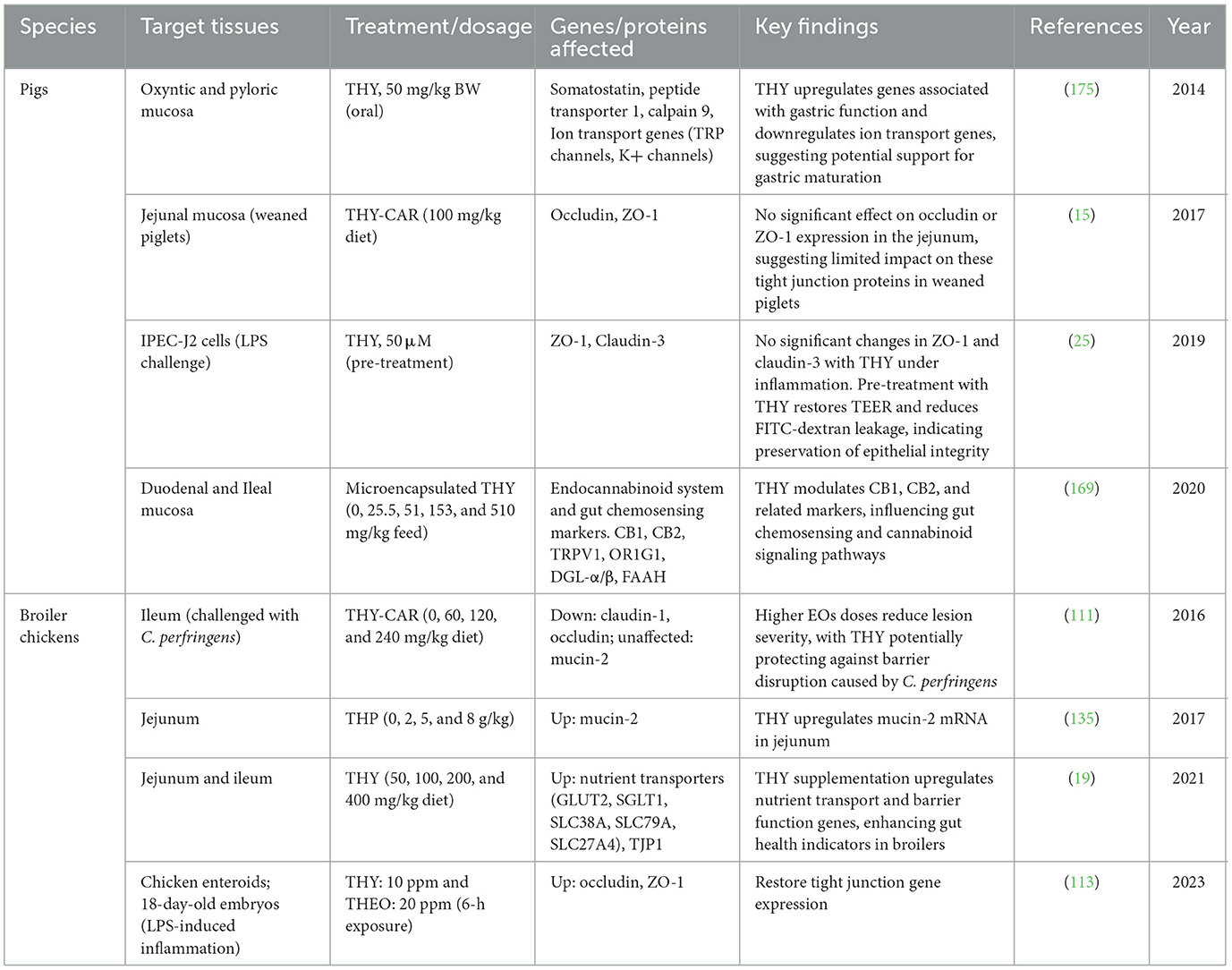
Table 21. Effects of THY on intestinal permeability, tight junctions, gastric mucosa gene expression, and modulation of the endocannabinoid system in various animal models.
3.6.14 THY effects on intestinal and hepatic morphological parameters
Data revealed consistent benefits of THY supplementation on gut morphology, as indicated by increased villus height and villus-to-crypt ratios in most animal models. These morphological improvements suggest enhanced absorptive capacity, which could contribute to better nutrient uptake and overall gut health. Concerning the liver, published data indicate that higher THY doses or its prolonged exposure may cause mild, reversible changes. Table 22 provides an overview of studies investigating the effects of THY on intestinal and hepatic morphological parameters across various animal models, including weaned piglets, broiler chickens, quail, pigeons, and rabbits.
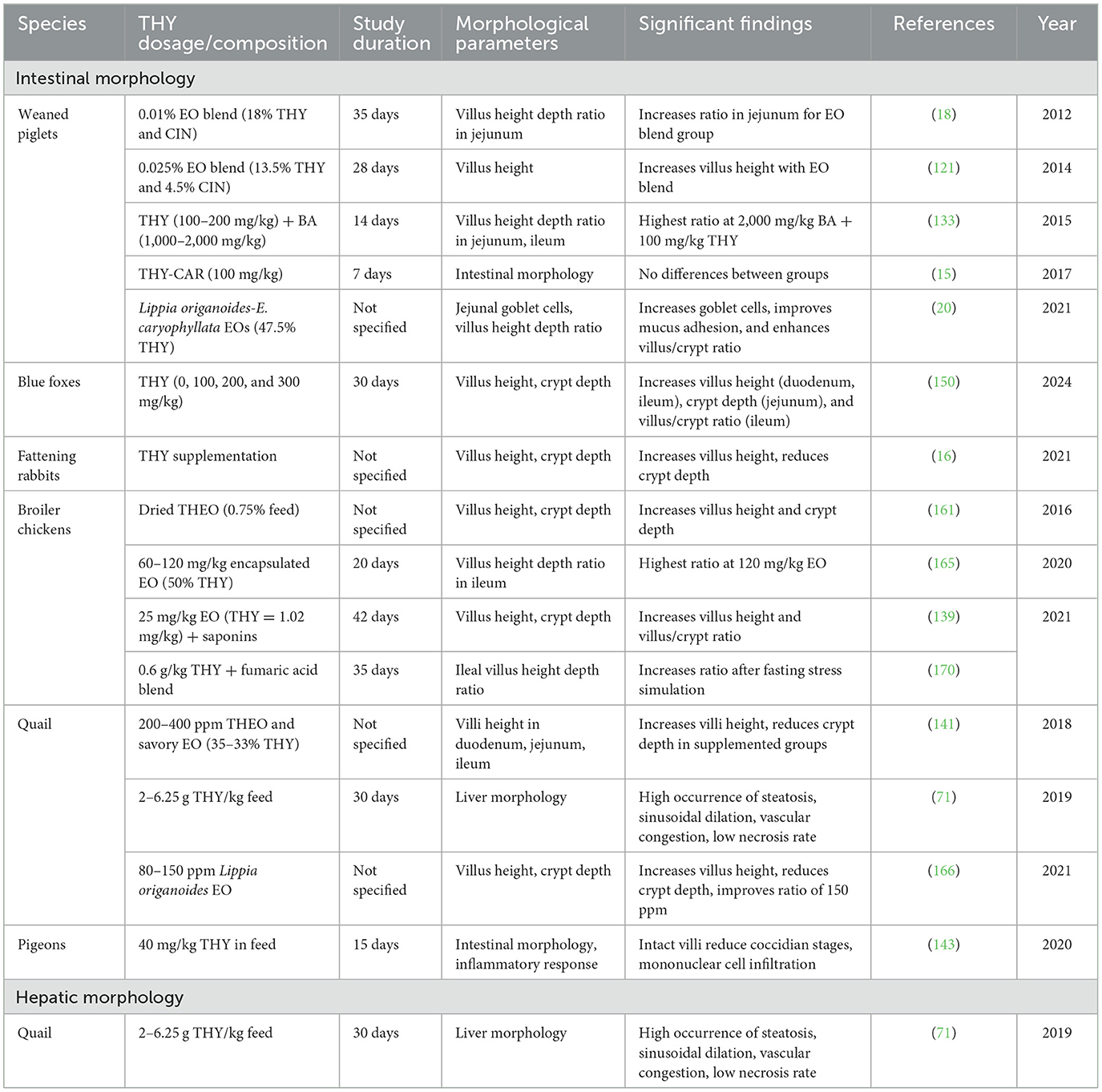
Table 22. Summary of studies evaluating the effects of THY on intestinal morphology in various animal models and hepatic morphology in quails.
3.6.15 Other effects of THY and THEO
Recent studies have explored various additional effects of THY across diverse biological systems, though research remains limited in most cases. These effects are summarized below:
3.6.15.1 Anticarcinogenic effects
Thymol (THY) has demonstrated anticancer activity in human cell lines and, in preclinical models, has shown protective effects against colon cancer. A 2021 study (7) in rats exposed to 1,2-dimethylhydrazine and a high-fat diet found that THY reduced serum levels of tumor markers (CEA, CA 19-9) and caspase-3, attenuated oxidative stress and inflammation in colonic tissue, and improved histopathological alterations, supporting its potential as a chemopreventive agent. Although these findings come from experimental models, they may have particular relevance for companion animals, such as dogs and cats, where colorectal cancer represents a clinical concern and research remains limited.
3.6.15.2 Behavioral effects
Several studies have investigated the potential anxiolytic properties of thymol (THY) in experimental animal models. THY appears to modulate stress- and fear-related behaviors, positioning it as a promising natural compound for managing anxiety. Although limited in number, the available studies, conducted primarily in avian (176) and rodent models (23), have assessed behavioral responses to experimentally induced stress, reporting reductions in anxiety-like behaviors such as excessive struggle and altered exploratory activity. Notably, these effects occurred without compromising general locomotor function, suggesting a selective action on anxiety-related pathways.
3.6.15.3 Anesthetic properties
THY and THEO exhibit dose-dependent anesthetic properties in various fish species, such as silver catfish, common carp, and tambaqui, making them promising alternatives to conventional anesthetics in aquaculture, as their efficacy depends on both concentration and species-specific sensitivity. It has been shown that THY acts via GABAA_AA receptors, although through mechanisms independent of benzodiazepine binding sites (75). The anesthetic potential of THY in common carp has been confirmed, although with slower induction and prolonged recovery times (117). Similarly, THEO has been reported to induce anesthesia in tambaqui, with higher doses associated with faster induction but extended recovery periods (147). These findings support the utility of THY as a natural anesthetic agent, although interspecies variability and concentration-dependent effects must be taken into account.
3.6.15.4 Wound healing
In rodent models, THY has demonstrated significant wound healing properties across different formulations and wound types. A study using collagen-based dressing films infused with THY reported enhanced wound retraction at days 7 and 14, along with improved granulation tissue and increased collagen density, suggesting accelerated and higher-quality tissue regeneration (105). Similarly, a THY-enriched bacterial cellulose hydrogel (1%) applied to third-degree burn wounds resulted in faster re-epithelialization, reduced inflammation, and improved collagen deposition over a 25-day evaluation period (46). These findings highlight the therapeutic potential of THY in promoting wound healing in preclinical models.
3.6.15.5 Bone loss
THY has been shown to exert significant anti-osteoclastogenic and bone-protective effects. In vitro, THY inhibits osteoclast differentiation in a dose-dependent manner, as evidenced by a reduction in both the number and size of TRAP-positive multinucleated cells. In vivo, THY administration in mice effectively prevents lipopolysaccharide (LPS)-induced bone loss, improving bone microarchitecture and reducing osteoclast counts (26). These findings underscore THY's potential in protecting bone integrity and modulating osteoclastogenesis.
3.6.15.6 Ovarian function
THY was evaluated for its effects on follicular activation, stromal cell protection, and collagen fibers in bovine ovarian cortical tissues. THY increased the percentage of normal follicles and improved follicular activation, collagen fiber density, and stromal cell density (129). These findings suggest that THY promotes follicular activation and helps maintain ovarian tissue integrity, indicating a potential protective effect on ovarian function.
3.6.15.7 Halitosis management
A clinical trial in dogs diagnosed with oral malodor shows that a THY- and menthol-containing gel effectively reduces bad breath by targeting malodor-producing bacteria (8).
4 Discussion
This systematic review highlights THY's potential as a bioactive compound in veterinary medicine due to its diverse pharmacological properties and comparatively low toxicity across various animal models. Key areas of THY's application include its antimicrobial, anti-inflammatory, antioxidant, and therapeutic roles in managing parasitic infections, gut health, and productive performance in livestock. The results presented in this review support the potential of THY as a bioactive compound with diverse health and productivity benefits across animal models and cellular studies. Researchers have demonstrated THY's efficacy in enhancing health outcomes, improving productivity, and mitigating the adverse effects of environmental and metabolic stress in different species. However, the effectiveness of THY appears to be contingent upon factors such as dosage, form of administration, and the specific physiological context of the target species.
THY's cytotoxicity varies widely across cell types, with LC50 or IC50 values ranging from 0.002 mg/mL (13.32 μM) in canine bone marrow stromal cells and enterocyte-like cells (30) to 362 μM in rat splenocytes (40). This range underscores the need for cell-specific considerations in therapeutic applications. Canine and porcine enterocytes are moderately sensitive to THY, with toxic concentrations above 100 μM in pig IPEC-J2 cells (25) and 0.05 mg/mL (333 μM) in canine hepatocytes (33), while EOs from various species show no toxicity in porcine hepatocytes (32), suggesting possible mitigating effects from other constituents. THY is relatively safe in immune cells and fibroblasts at low concentrations, with no significant cytotoxicity observed in murine macrophages up to 40 μg/mL (266.5 μg/mL); however, higher doses (665.94 μM) reduce cell survival (3). Fibroblasts (NIH-3T3) show only a slightly reduced viability at increasing THY doses (46), suggesting the obtainment and use of effective doses may be feasible. Tolerance to THY also varies in renal and mammary cells, with moderate toxicity (IC50 = 300 μM) in Vero cells (35) and no significant toxicity in mammary epithelial cells from mice and cattle, even at high concentrations (24, 28), thereby highlighting THY's potential for low-dose veterinary applications prioritizing renal and mammary health. In vivo toxicity data show species-specific, dose-dependent effects, corroborating the in vitro cytotoxicity findings. For instance, THY's LD50 in rodents is 1350.9 mg/kg, while its derivative THY acetate has a much higher LD50 (4,144.4 mg/kg) (47), suggesting how structural modifications can reduce acute toxicity. Extended THY exposure at lower doses (10–40 mg/kg over 30 days) induces immunotoxic effects, affecting ATP hydrolysis pathways and triggering inflammation (48), linking back to in vitro data showing stress responses in immune cells. Further in vivo studies on embryotoxicity and estrogenicity indicate that THY (0.5 mg/kg) in chickens induces developmental abnormalities such as curled claws and everted viscera, alongside weak estrogenic activity, although without evident mutagenic effects (50). These findings suggest that THY could interfere with organogenesis and hormonal regulation at higher concentrations. The alignment between in vitro and in vivo results demonstrates that while in vitro assays highlight potential risks, in vivo data provide a more comprehensive view of THY's systemic effects, including inflammation and developmental impacts, emphasizing the need for cautious application in sensitive contexts.
THY pharmacokinetics show species-specific variations in absorption, half-life and bioavailability, significantly influenced by administration route, encapsulation, and dosage. Understanding these profiles is essential for effective therapeutic use and minimizing adverse effects. Rodent studies indicate a rapid systemic absorption with short half-lives (~2.5–3 h) via intravenous and inhalation routes (49, 55). In rabbits fed with THY-containing diets, THY plasma levels correlate with concentrations found in the intestinal wall, suggesting a focus on gastrointestinal distribution (56). In broiler chickens, THY absorption increases in a dose-dependent manner with THEO supplementation, highlighting the avian gut's efficiency in absorbing lipid-soluble compounds such as essential oils and their components (57, 59). For pigs, encapsulated THY formulations extend half-life and bioavailability, enhancing intestinal targeting while reducing systemic exposure (60, 61). In dairy cattle, intramammary THY shows limited systemic absorption, supporting its use in mastitis with low milk residue risks (63, 64). These findings underline the need for tailored dosing and encapsulation to optimize THY's efficacy and safety across species.
Residue analysis also reveals species-specific THY dynamics. In dairy cattle, liver and milk retain THY residues post-treatment, suggesting a 72-h withdrawal period (63, 64). Broiler chickens show THY localization in the gut with minimal muscle residue, supporting its safe use as a feed additive (57, 70). In quails, THY dose-dependently accumulates in eggs, thus necessitating a withdrawal period to prevent residual concentrations (71). Pigs demonstrate limited systemic deposition, primarily in the gut, enhancing safety as a dietary supplement (61). Collectively, these findings suggest that THY, used within regulated doses, minimizes residue risks across animal products, making it a safe additive with appropriate withdrawal guidelines.
THY's interaction with xenobiotics, particularly veterinary drugs, remains underexplored but may significantly influence drug efficacy and metabolism in production animals. Known for its therapeutic benefits, THY may impact oxidative, reductive, and conjugative drug metabolism, potentially altering co-administered drugs' pharmacokinetics and pharmacodynamics. For example, THY altered ABZ's metabolism in lambs, reducing the Cmax and AUC of ABZ sulfone. Notably, THY did not enhance ABZ's efficacy, but instead inhibited sulphonation and sulphoreduction processes, suggesting enzyme competition that may affect drug clearance (29). This highlights the need for research into THY's effects within multi-drug regimens, given its potential to influence therapeutic outcomes in production animals.
THY exhibits broad-spectrum antimicrobial activity, being effective against both Gram-positive and Gram-negative bacteria and making it valuable in addressing antibiotic resistance in veterinary contexts. THY exerts its antibacterial effects through multiple mechanisms, primarily by disrupting cell membranes. It integrates into the lipid bilayer of bacterial membranes, increasing permeability and leading to the leakage of essential intracellular components, ultimately causing cell death (72). Furthermore, THY inhibits bacterial quorum sensing, thereby reducing biofilm formation and virulence. Additionally, it damages bacterial DNA and RNA, impairing replication and transcription, and inhibits key metabolic enzymes such as ATPases and glycolytic enzymes, vital for bacterial energy production. These combined actions contribute to THY's effectiveness as an antibacterial agent. THY's antibacterial properties are thus not only due to membrane disruption but also to its interference with essential cellular processes, making it a potent antimicrobial compound. Effective concentrations depend on pathogen type, THY form, and application. Studies show that THY inhibits a range of pathogens, including those resistant to common antibiotics, at MICs of 0.01–0.32 mg/mL (77). THY demonstrates stronger antimicrobial effects when combined with CAR or CIN essential oils (5, 86), and its derivatives may be more effective than alternatives like EUG against S. aureus (51). The variability in MIC values reported across studies, such as 0.4 mg/mL for Thymus numidicus essential oils against Pseudomonas spp. (79) and 2.5–160 μg/mL for Thymus spp. against various pathogens (32), reflects differences in Thymus species and microbial species, with formulations and study conditions also potentially influencing the results. Concerns about resistance are limited with THY; studies found no resistance in E. coli or mesophilic gut flora post-treatment (84), though tolerance was observed in some cases (81). THY's efficacy against pathogens causing bovine mastitis (S. aureus, E. coli, St. uberis, and the algae Prototheca bovis) suggests it could serve as an alternative or adjunct to antibiotics (66, 87). While THY's antimicrobial utility is clear, the variability in MIC/MBC values underscores the need for standardized protocols in veterinary chemotherapy.
THY and THEO demonstrate varied antiviral effects, showing efficacy against certain viruses while presenting limitations with other ones. Studies reveal strong inhibitory effects on enveloped viruses such as HSV-1, where a 90% virion inactivation was noticed at an IC50 value of 7 μM (35). THY's antiviral action likely stems from its ability to destabilize viral envelopes or capsids, inhibiting viral replication and interfering with viral entry into host cells. In contrast, THY showed mixed results against non-enveloped viruses. For instance, it dose-dependently inactivates norovirus surrogates, while it shows minimal effects on HAV, thereby suggesting that non-enveloped viruses may resist THY's mechanism or require higher doses to be effective (38). The structural disruption of viral envelopes by THY supports its selective efficacy against enveloped viruses, while higher concentrations may be necessary for non-enveloped viruses. THY-enriched EOs demonstrated potent virucidal effects against feline coronavirus (FCoV-II), reducing titers by up to 3.25 log10 (88), indicating potential veterinary applications. However, there is a lack of effect on FCV and its effectiveness against lipid-enveloped viruses. THY's concentration-dependent antiviral effect and structural interaction with viral envelopes further emphasize its selective efficacy.
THY and THEO also exhibit broad-spectrum antifungal efficacy, impacting pathogenic, dermatophytic, and environmental fungi. Studies demonstrate consistent antifungal effects, with a significant inhibition of Rhizopus oryzae growth at 128 μg/mL (89); in addition, a strong growth inhibition was observed with various Thymus species (32). THY's antifungal action is primarily due to the disruption of fungal cell membranes. It alters fatty acid metabolism, reduces ergosterol content, and induces oxidative stress through the generation of ROS, leading to fungal cell death. THY's effectiveness in combination therapies is noteworthy; for instance, a THY-itraconazole-clarithromycin blend achieved 96% inhibition against Pythium insidiosum, suggesting synergy that could enhance conventional treatments and reduce dosage (9). This synergy could be beneficial for treating complex fungal infections, particularly in veterinary settings. Additionally, THY vapor has been shown to inhibit fungal growth and toxin production; as an example, it suppressed A. flavus growth and reduced aflatoxin B1 production through gene downregulation (10), thereby suggesting potential for THY in agricultural food safety to control both fungal growth and toxin contamination in stored grains.
THY's antifungal potential extends to veterinary contexts, particularly against dermatophytes and fungi linked to infections. THEO is effective at low concentrations (0.5–2.5 μg/μL) against skin-infecting fungi (91), while THY-rich THEO inhibited Malassezia pachydermatis, common in canine ear infections (92). This supports THY's suitability in topical treatments for dermatophytic infections in animals. Additionally, species-specific sensitivity to THY was observed; for instance, dietary THEO (0.5–2%) protected carp from Saprolegnia spp. (90), suggesting that dietary supplementation could protect against fungal infections in aquaculture. The versatility of THY application methods, including liquid and vapor forms, offers flexibility for use in diverse veterinary and agricultural practices, as noted for A. flavus (10).
THY and THY-containing EOs demonstrate strong antiparasitic properties against various ectoparasites, such as flies, ticks, mites, and mosquitoes, indicating high potential for pest control in veterinary and agricultural settings. Significant acaricidal effects have been noticed on ticks like Rhipicephalus microplus and Ixodes ricinus, with Lippia gracilis EOs displaying high efficacy (LC50 between 0.84 and 1.02) even on resistant strains (11, 94). THY's larvicidal and repellent actions were similarly effective, achieving over 90% repellency and 100% larvicidal activity against ticks (94), which supports its utility as a natural acaricide. Additionally, prolonged residual effects were observed in certain applications; for example, a THY-CAR blend provided 14-day protection against red mites (Dermanyssus gallinae) in poultry (95). Such sustained effects make THY viable for long-term pest control, reducing treatment frequency. THY has also shown ovicidal and larvicidal activity across ectoparasite life stages. For example, including THY in quail diets reduces housefly (Musca domestica) oviposition (4), which is beneficial for livestock environments where flies proliferate. THY's effects were enhanced when combined with other natural compounds like CAR and EUG, suggesting synergistic potential. Significant reductions in mosquito and tick populations were observed when using THY-based blends (6, 12), highlighting lower dosage requirements and improved animal tolerance compared to standalone treatments. Moreover, THY demonstrated comparable or superior efficacy to synthetic acaricides like permethrin against ticks (94), showing promise as an organic alternative for pest management in contexts where chemical residues are a concern. Beyond ectoparasites, THY exhibits broad-spectrum efficacy against protozoan and helminth parasites, with a potential for use as a natural anthelmintic in veterinary practices. THY has shown strong oocysticidal effects against protozoa such as Eimeria spp. and Cryptosporidium spp., which are key pathogens in poultry. Specifically, THY disrupts oocyst wall integrity, causing parasite death with LC50 values of 1.66 mg/mL for Eimeria (96) and <0.4 mg/mL for Cryptosporidium (104). These findings support THY's potential as a natural disinfectant in poultry farming, reducing reliance on chemical agents that may leave residues in food products. Significant anthelmintic activity was also observed against parasitic nematodes, especially Haemonchus contortus, a prevalent gastrointestinal parasite in ruminants. THY's efficacy extends to cestodes like Echinococcus granulosus, with a reduction of cyst infectivity following a THY-induced structural damage in protoscoleces (98), suggesting THY's utility in treating cestode infections, especially where drug resistance is a concern. THY has also been proven effective against Leishmania spp., which may have implications for treating leishmaniasis in animals. In particular, THY derivatives showed a significant activity against Leishmania infantum chagasi promastigotes (3), while THY alone reduced parasite loads in infected hamsters (103). These findings suggest THY's therapeutic potential for zoonotic diseases, possibly offering a safer, cost-effective alternative to traditional antileishmanial drugs. Finally, THY interacts with synthetic anthelmintics such as ABZ; indeed, THY inhibits hepatic ABZ metabolism, thereby affecting its activation (65). While this interaction could support co-treatment strategies, it underscores the need for research into THY's pharmacokinetic interactions to prevent unintended efficacy reduction or toxicity, particularly in livestock where drug combinations are prevalent.
In terms of mechanisms of action, THY's anti-inflammatory and antioxidant effects are multifaceted and involve various molecular pathways and cellular processes. As to anti-inflammatory effects, THY exerts its influence through several mechanisms. The inhibition of MPO activity (105) is one key mechanism, preventing leukocyte infiltration and reducing the oxidative damage associated with inflammation. THY also reduced inflammation in pleurisy models (43), although its effect on cell migration varied and hints at concentration-specific responses. Additionally, THY modulates cytokine production, notably increasing the ratio of IL-10 to IL-2, which supports a Th2-dominant anti-inflammatory response (40). This modulation of immune responses is likely mediated by THY's ability to interfere with NF-κB and MAPK signaling pathways, which are crucial in the regulation of pro-inflammatory cytokines, such as TNF-α, IL-6, and IL-1β (24). Systemically, THY effectively reduced inflammation markers in liver and vascular tissue models. For example, it decreased TNF-α in a liver inflammation model (106), while reductions in VCAM-1, MCP, and C-reactive protein were noticed in a high-fat diet-induced inflammation model in rabbits (112), highlighting THY's broad efficacy in both tissue and systemic inflammation. Studies in broilers and piglets (15, 111) revealed a reduction of pro-inflammatory markers, showing potential applications of THY in livestock where bacterial infections and stress-induced inflammation are relatively common. Additionally, THY reduced lung inflammation markers and enhanced antioxidant defenses through Nrf2 and HO-1 pathways (108), while a dose-dependent reduction of the inflammatory response was observed in a model of gastric ulcer (109), underscoring THY's therapeutic versatility. Overall, these findings suggest that THY can effectively manage both acute and chronic inflammatory conditions, with evidence supporting dose-dependent optimization for enhanced therapeutic outcomes.
In terms of oxidative stress, THY's antioxidant activity is attributed to its ability to scavenge free radicals and enhance endogenous antioxidant defense systems. THY reduces MDA, a marker of lipid peroxidation, while simultaneously boosting the activity of antioxidant enzymes such as SOD and GPx (114, 117). This dual effect helps to reduce oxidative damage to cellular structures, particularly in tissues exposed to environmental stressors or inflammatory responses. In aquatic species, such as rainbow trout and Nile tilapia, THY supplementation demonstrated significant protection against lipid oxidation (2), suggesting its potential use in aquaculture to enhance fish health and stability. Similarly, THY supplementation in pigs and rodents has been linked to improved oxidative balance, with THY mitigating the oxidative damage caused by infections and pharmacological treatments, thus supporting its potential use in livestock management (106, 118). THY's antioxidant properties encompass its ability to reduce oxidative stress markers in high-fat diet-induced models, indicating its broad applicability in managing diet-induced oxidative conditions (112). Furthermore, THY's influence on oxidative stress pathways is not only limited to lipid peroxidation; it also affects reactive oxygen species (ROS) production. In vitro, THY's ability to reduce ROS in LPS-stimulated macrophages and intestinal cells confirms its direct role in modulating oxidative bursts, which are common in inflammatory responses. This further strengthens its potential as a therapeutic agent in conditions where both inflammation and oxidative stress are prevalent, such as colitis and gastrointestinal disorders (42). In poultry, THY increased antioxidant defenses (125), with dose-responsive trends improving resilience in broiler chickens and laying hens (1). THY's efficacy depends on botanical source, with some Thymus species offering higher radical scavenging activity than other ones (32). Overall, THY's ability to reduce MDA and increase antioxidant enzyme activity supports its role as a natural antioxidant in veterinary applications, making it a promising option for managing oxidative stress-related conditions across species.
Studies examining THY's impact on blood biochemical markers across animal models demonstrate beneficial yet variable responses, which are influenced by species, dosage, and formulation. Generally, THY has shown potential in enhancing blood lipid profiles and modulating immune functions, but optimized dosing is crucial given the variability observed among species. In terms of lipid profiles, THY supplementation reduced serum cholesterol, triglycerides, and LDL levels in rabbits and quail, while increasing HDL levels (112, 141). Similar lipid-lowering effects were observed in broilers, suggesting THY's potential role in managing lipid profiles and possibly supporting cardiovascular health in high-stress environments common to intensive farming (126). As far as blood biomarkers are concerned, in broilers and fish, THY showed immunostimulatory effects by enhancing hematological markers, particularly leukocyte and lymphocyte counts (90, 144). In broilers given THY, increased eosinophil, lymphocyte, and monocyte percentages have been noticed (138), which could enhance resilience to infections and reduce the need for antibiotics. In rainbow trout fed with a THY-supplemented diet, an increase in hematocrit, hemoglobin, and WBC counts was observed (146), suggesting an enhanced oxygen transport and immune function in fish. However, certain studies reported minimal changes in hemato-biochemical parameters, indicating that THY's physiological effects may vary across species and contexts. In ducks given THY supplementation, no significant impact on serum proteins or cholesterol levels was recorded (142), while minimal biochemical changes occurred with a 20% THEO (130), suggesting that specific formulations or lower concentrations may not elicit strong physiological responses. As mentioned above, species differences in the response to THY and THY-based EOs have been recorded, too. For instance, THY elicited dose-dependent effects on goat liver enzymes, where higher doses induced hepatic strain (14), underscoring the importance of cautious dosing to avoid liver stress with long-term use.
In poultry, THY and THY-based EOs showed indirect benefits on animal digestion; indeed, increasing trypsin, lipase, and protease activities were noticed in broilers (125, 139), indicating THY's potential to improve nutrient absorption, feed efficiency and the overall digestive process. Interestingly, THEO is beneficial in regulating glucose levels post-anesthesia in fish, which could help mitigate stress during handling and transport (147). However, contradictory results on lipid metabolism markers have also been observed, suggesting that age or health condition could influence digestive benefits (133).
THY's immune-modulating effects also demonstrate its broad applicability across different species. It is evident that THY can enhance immune function by increasing antibody titers, modulating cytokine responses, and improving immune cell function. A number of studies made in piglets (18, 121, 149) are an example of THY's ability to boost immune resilience through improvements in lymphocyte proliferation and elevated levels of immunoglobulins (IgA, IgM) and complement proteins. This is particularly valuable during growth phases or periods of stress, reducing vulnerability to infections. Similarly, poultry studies [e.g., (125) and (151)] showed that THY improves responses to vaccinations (e.g., against Newcastle disease) and reduces stress markers in broilers, enhancing overall health and vaccine efficacy. Additional research showed increased antibody titers for viral pathogens (21, 152), suggesting that THY could support vaccine efficacy and overall health. In aquaculture, THY's effects on immune health have also been demonstrated through an enhancement of lysozyme and catalase activities, which are critical components of innate immunity in species like rainbow trout (114) and Nile tilapia (2). THY also reduces bacterial load, including pathogens such as Campylobacter spp. and Salmonella spp., which are relatively common in poultry farming (155, 156). This reduction in bacterial infections is a key consideration in veterinary applications, especially in reducing antibiotic use in intensive farming systems. THY's potential to reduce mastitis in dairy cows, by reducing S. aureus adhesion (45), further exemplifies its antimicrobial benefits, especially in high-stress and high-density farming environments. In terms of dose-dependent effects, it is crucial to note that THY's efficacy in enhancing immune responses and reducing stress-related immune suppression is contingent on the optimal dosing. Studies with hens (128) and trout (14) show that lower or higher doses can either reduce efficacy or even suppress immune function, indicating the need for careful dose optimization to maximize its benefits. Furthermore, THY is promising in mitigating immunosuppression caused by environmental stressors. THY supplementation improved immune responses in broilers exposed to mycotoxins (137) and protected African catfish from pesticide-induced immunotoxicity (148). Additionally, THY's anti-inflammatory properties provide protective effects in models of induced tissue damage, such as reduced colonic inflammation in colitis models (42) and pancreatic protection in conjunction with non-steroidal anti-inflammatory drugs (153). In summary, THY's diverse immune-modulating properties, spanning from enhancing resistance to infections to alleviating stress-related immune suppression, underline its potential as a natural additive for improving animal health. This broad-spectrum efficacy, particularly in high-stress production environments, suggests that THY could reduce dependency on antibiotics, making it a promising agent for sustainable animal health management.
Data indicate that THY and THEO supplementation yield mixed results on productivity in various animal species, with effects largely dependent on dosage, species, and environmental conditions. For instance, 200 mg/kg of THY and CAR improved poultry feed efficiency and weight gain (125), possibly due to enhanced digestive enzyme activity and gut health. In broilers under heat stress, THY improved weight gain and feed FCR, underscoring its capacity to mitigate stress-related performance losses (126). However, other studies [e.g., (58) and (155)] found no significant impact on performance, thus suggesting that THY may be more effective in challenging environments than in optimal conditions. In egg-laying hens, THY combined with other EOs improved FCR, enhanced egg production and quality, and shell strength, maybe due to THY's antioxidant properties (1, 138, 140). For meat and dairy production, the findings were inconsistent. While some authors reported no effect on milk yield or composition in dairy cattle (171), other ones (18) observed weight gain improvements in pigs when using encapsulated THY (18), which may enhance nutrient absorption by targeting specific gut regions. In aquaculture, THY supplementation under stressful or pathogen-challenged conditions resulted in improved survival rates and growth. Its antimicrobial and immune-boosting properties appear beneficial under high pathogen loads, though effects on FCR and growth in unstressed conditions remain inconsistent. THY also enhances nutrient digestibility and gut microbiota composition across species. In monogastric animals, studies on piglets (18, 121), blue foxes (150), and broiler chickens (125, 163) showed increased digestibility of dry matter, protein, and energy, likely resulting from improved gut health. THY also affects fatty acid profiles: it provoked a reduction in saturated fatty acids and an increase in polyunsaturated fatty acids (PUFAs) in broiler meat (125), with similar trends observed in eggs, too (174). In ruminants, THY's potential to alter fermentation profiles is notable for sustainability. It is capable to Yu et al. (173) reduce methane production in an in vitro rumen model (173) and protozoa density in dairy cows without changes in pH or volatile fatty acids (171). Variable responses in volatile fatty acid profiles across species suggest a need for targeted research to optimize dosing and application methods for maximum productivity and environmental benefits. Overall, THY's effects on productivity, nutrient digestibility, and gut microbiota underline its potential as a natural additive in animal production systems. However, species-specific responses and mixed findings on fatty acid and methane profiles highlight the need for further studies to refine its application across livestock sectors.
Studies on THY supplementation reveal generally positive effects on gut microbiota across animal species, frequently reducing pathogenic bacteria like E. coli and coliforms while enhancing beneficial populations, such as Lactobacillus spp. This suggests THY may play a key role in gut health and microbiota balance, though outcomes depend on dosage, delivery method, and existing microbial communities. For example, in canines, THY with yeast cell wall increased beneficial bacteria (17), including Blautia and Faecalibacterium, while reducing Streptococcus, underscoring THY's potential to boost gut health and immune resilience. In piglets, Li et al. (18) and Diao et al. (133) reported reduced E. coli and increased Lactobacillus, supporting gut barrier function and nutrient absorption, which are especially crucial post-weaning. In poultry, THY consistently improved microbiota, reducing harmful bacteria while boosting beneficial ones, which can enhance growth and feed efficiency. In pigs, Abdelli et al. (170) observed that microencapsulated THY increased beneficial bacterial families (e.g., Bacteroidetes and Bacillaceae), while reducing pathogenic Pseudomonadaceae. These findings indicate THY's potential to foster a balanced gut microbiome. Studies in blue foxes, rabbits and quail also show increases in beneficial bacteria, suggesting THY's utility as a natural alternative to antibiotics. However, its effects on aquatic species, such as trout, are inconsistent and require further research to understand species-specific responses.
THY also appears to support gut barrier integrity, especially in inflammatory contexts. For instance, it protects IPEC-J2 cells from lipopolysaccharide-induced permeability disruptions (25), suggesting it stabilizes tight junctions and limits inflammatory damage to epithelial cells. Although changes in proteins like ZO-1 and claudin-3 were not observed, THY's ability to improve barrier function may involve other pathways. In pig models, THY upregulated gastric mucosa genes related to gut defense (175), suggesting benefits that extend beyond the small intestine. In broiler chickens, THY also alleviated bacterial damage to gut integrity. Moreover, a THY-CAR blend reduced ileum lesion severity during Clostridium perfringens challenges while maintaining mucin-2 levels, a protective gut lining component (111). In weaned piglets, THY had limited effects on tight junction proteins under normal conditions (15), suggesting it may be most beneficial under stress or inflammation. On the other hand, in broilers, THY enhances the expression of GLUT2 and SGLT1 nutrient transporters, indicating potential to support nutrient absorption and gut health, which are critical for productivity in animal production systems (19). Overall, THY supplementation shows promise for improving microbiota balance, nutrient absorption, and gut barrier integrity in animals. Its benefits appear most pronounced under inflammatory or stressful conditions, where it may help maintain gut health and productivity, though species-specific effects warrant further investigation.
Additionally, a study suggests that THY may support gut health and development in weaning piglets by modulating gene expression linked to the endocannabinoid system and gut chemosensing pathways (169). THY's influence on CB1 and CB2 cannabinoid receptors, TRPV1 chemosensory receptor, OR1G1 olfactory receptor, and key enzymes in endocannabinoid synthesis and degradation (DGL-α, DGL-β, FAAH) points to a complex regulatory mechanism of gut motility, inflammation, and permeability. The CB1 receptor is primarily involved in regulating gut motility and appetite, whereas CB2 helps modulate immune responses, which may reduce gut inflammation and support overall intestinal health during stress, such as weaning. These effects suggest THY may help pigs to adapt to dietary and microbial changes associated with weaning. On the other hand, THY's modulation of TRPV1 and OR1G1 mRNA levels suggests additional roles in chemosensing and mucosal health. TRPV1, a receptor involved in inflammation and immune responses, may reduce gut irritation caused by dietary shifts or microbiota changes, while OR1G1 aids in nutrient sensing and could optimize nutrient absorption by the gut. By influencing enzymes for endocannabinoid biosynthesis and degradation, THY may balance endocannabinoid tone, potentially enhancing stress resilience and reducing gut inflammation during weaning. Overall, these findings indicate that THY could facilitate a smoother transition to solid food, improving gut health and growth. Furthermore, THY's impact on the balance of endocannabinoids suggests a modulation of the gut-brain axis, potentially reducing anxiety and promoting a more balanced immune response. Clearly, further research is needed to validate these effects across different diets and environmental conditions.
THY supplementation shows strong potential to improve intestinal morphology and gut health across animal models. Studies consistently report increases in villus height, crypt depth, and villus-to-crypt ratios, all markers of nutrient absorption and digestive efficiency, which ultimately support growth and productivity in livestock. In weaned piglets, THY significantly enhances intestinal structure, i.e., resulting in higher villi and improved villus-to-crypt ratios in the jejunum and small intestine (121, 149). These latter changes increase mucosal surface area and improve the absorption of nutrients. Worth mentioning, THY's efficacy can be optimized when combined with other gut health-promoting compounds, such as benzoic acid (BA) (133). Also, in broiler chickens, THY supplementation improves gut morphology, as seen in increased villus height and villus-to-crypt ratios, especially at moderate to high doses (161, 165). As the combination of THY and saponin produced similar enhancements, it is conceivable to hypothesize a synergism between THY and other natural additives (139). Additionally, THY helps maintain intestinal structure under fasting stress, potentially supporting gut integrity in challenging conditions (170). In rabbits and Japanese quail, THY shows comparable effects, promoting villus height and reducing crypt depth, which supports a more efficient digestion (16, 141). Overall, this evidence suggests that THY may enhance nutrient absorption and reduce pathogen colonization in avian species and potentially other target species. In contrast, THY's effects on liver morphology vary. Signs of hepatic stress, such as steatosis and vascular congestion, were observed at high doses in quail, though low necrosis rates suggest a dose-dependent sensitivity (71). Therefore, careful dosing is recommended to balance THY's gut health benefits with potential liver impacts. Finally, THY supplementation in pigeons preserved gut integrity and reduced coccidian stages, supporting its use as a natural alternative to antibiotics in managing infections like coccidiosis (143).
While the effects of THY on intestinal health are well-documented, its potential as a cancer therapeutic, especially for colon cancer, has garnered increasing attention. Although less frequently diagnosed in animals than in humans, colon cancer remains a significant concern in veterinary oncology, particularly among species such as dogs and cats. This malignancy is typically associated with genetic predispositions, chronic inflammation, or exposure to carcinogens. Research on colon cancer in animals is critical not only for improving diagnostic and therapeutic approaches in veterinary medicine but also for providing translational models that enhance our understanding of the disease in humans. A recent study (7) on THY suggests that the compound may significantly reduce tumor markers, such as CA 19-9 and CEA, which are used in clinical settings to monitor colon cancer progression. Additionally, THY's anticancer potential is linked to its ability to induce apoptosis in cancer cells through mitochondrial depolarization, activation of the Bax protein, caspase activation (including caspase-3), and increased ROS levels (36, 73), all of which lead to DNA damage and cell death. This mechanistic action highlights its role as a natural compound capable of modulating cancer cell survival and proliferation. Moreover, THY's effects on oxidative stress and inflammation in colonic tissue further support its role in cancer prevention, as oxidative damage and chronic inflammation are key contributors to carcinogenesis. Histopathological analysis from the same study reveals THY's impact on reducing aberrant crypt foci (ACF) and improving tissue architecture, reinforcing its potential as an agent for preventive cancer therapy.
THY has been shown to exhibit anxiolytic properties in avian and rodent models, effectively reducing stress-related behaviors without impairing motor activity. In quails exposed to restraint stress, THY selectively reduced anxiety without causing motor suppression (176). Similar anxiolytic effects have been observed in rats administered THEO (23). These findings highlight THY's potential as a natural anxiolytic, making it particularly valuable in livestock settings where managing stress is crucial. The anxiolytic effects of THY may be linked to its interaction with the central nervous system, where it likely modulates GABAergic and serotoninergic pathways to reduce anxiety-like behavior. Because THY reduces anxiety without inducing sedation, it could serve as a preferable altersnative to synthetic anxiolytics for stress-sensitive species during transport, weaning, or confinement. Further studies are needed to fully understand its mechanisms and determine optimal dosages across species for broader use in animal care.
Studies have reported THY's dose-dependent anesthetic efficacy in fish, with variations in induction and recovery times. THY's interaction with GABAA receptors suggests unique anesthetic mechanisms that minimize side effects like dependence or tolerance, making it a potentially safer alternative for routine anesthesia (75). In common carp, Yousefi et al. (117) found that THY had slower induction and longer recovery than EUG, suggesting that higher doses or modified protocols may be necessary for quicker effects. Similarly, Boaventura et al. (147) reported that higher doses of THY in tambaqui led to quicker induction but prolonged recovery, emphasizing the need for species-specific dosing protocols. THY's potential as an anesthetic is promising for aquaculture, especially for reducing handling stress during extended procedures. Optimizing its formulation, such as microencapsulation, could enhance THY's bioavailability and accelerate its anesthetic action across diverse fish species. Further studies on THY's neuroreceptor interactions might uncover safer anesthetic pathways for broader veterinary use.
THY also seems to be effective as a wound healing agent in rodents. Incorporated into collagen films or hydrogels, THY enhances wound retraction, re-epithelialization, inflammation control, and collagen deposition, making it a promising natural compound for wound care. In addition, the antioxidant properties of THY are likely to reduce oxidative stress at the wound site, accelerating the healing process. THY-enriched collagen films improved wound retraction and increased collagen density, supporting stronger tissue regeneration (105). Furthermore, a 1% THY-enriched hydrogel accelerated healing in third-degree burns, with faster re-epithelialization and enhanced collagen deposition, suggesting value in treating complex wounds (46). THY's inclusion in collagen-based and hydrogel formulations illustrates its flexibility in wound care applications. These results encourage further testing in larger animal models and clinical settings to confirm its therapeutic efficacy across species. Given its antimicrobial and anti-inflammatory properties, THY holds promise as a multifunctional agent for wound care, especially in complex cases such as burns.
The limited research on THY's effects on bone health includes a study (26) demonstrating its potential as a natural treatment for bone degeneration caused by excessive osteoclast activity, highlighting the need for further investigation. This study shows that THY inhibits osteoclast differentiation and bone loss. In vitro, RAW264.7 cells and bone marrow macrophages exposed to THY at concentrations of 10–40 μM exhibited a decrease in tartrate-resistant acid phosphatase (TRAP)-positive cells, a marker of osteoclast differentiation, suggesting interference with osteoclastogenesis. THY also modulates key signaling pathways such as NF-κB, known for its pivotal role in osteoclastogenesis and inflammation. In vivo, THY reduced osteoclast numbers and improved bone structure in LPS-induced bone loss in mice treated with 25 and 100 mg/kg. Computer tomography and histological analysis confirmed enhanced bone microarchitecture. These results, though promising, underline the need for more comprehensive research to validate THY's effectiveness in preventing bone resorption. Nonetheless, its potential makes it a promising natural alternative for maintaining bone health in veterinary applications, including aging pets or animals prone to bone disorders.
The only available study on THY's effects on bovine ovarian tissue (129) highlights its potential in supporting follicular activation, maintaining tissue integrity, and enhancing collagen density in in vitro cultures. THY's modulation of collagen synthesis and its effect on follicular activation are likely mediated through its interaction with signaling pathways that regulate cell proliferation and differentiation, such as the Wnt/β-catenin pathway. In this study, THY at 400 and 800 μg/mL increased the proportion of morphologically intact follicles, with 400 μg/mL notably promoting primordial follicle activation, essential for early folliculogenesis. Additionally, this dose enhanced stromal cell density and collagen fiber support, suggesting THY's potential involvement in modulating the extracellular matrix (ECM), possibly by affecting the activity of matrix metalloproteinases (MMPs) that regulate tissue remodeling. These changes are crucial for ovarian structure and function. THY's role in follicular activation and structural preservation suggests it could improve the success of assisted reproductive technologies (ART) in cattle, with potential benefits in breeding, conservation, and fertility preservation efforts. Future studies should examine the molecular mechanisms underlying these effects and explore THY's interactions with other reproductive technology compounds.
Finally, evidence from a single study (8) demonstrates that a THY- and menthol-containing gel effectively reduces halitosis in dogs. In a crossover design where each dog received both active and placebo treatments, the gel significantly improved oral odor during the active treatment phase. The antimicrobial and deodorizing properties of THY and menthol are likely to contribute to these results by targeting bacterial populations responsible for malodor, particularly those involved in sulfur compound production. The reduction in halitosis scores, reported by both owners and clinicians, supports the gel's efficacy. These findings suggest that THY-menthol gels could be a valuable addition to canine oral hygiene routines, helping manage persistent bad breath and potentially improving oral health in pets prone to periodontal issues.
In conclusion, this systematic review contributes to the growing body of evidence on THY's multifaceted bioactivity in veterinary medicine, attributed to its broad-spectrum antimicrobial, anti-inflammatory, antioxidant, and antiparasitic properties, among others. THY's efficacy across various animal models demonstrates its capacity to support immune resilience, enhance gut health, and improve overall productivity in livestock. Nevertheless, its safety and effectiveness are contingent upon species-specific dosing and administration forms, as THY's cytotoxicity and pharmacokinetics vary notably across different cell types and animal models. Encapsulation strategies and co-formulations with other bioactive compounds may optimize its therapeutic potential while mitigating risks of toxicity and environmental residues. Given its efficacy in supporting animal health and its potential to reduce antibiotic dependency, THY represents a promising candidate for sustainable disease management in animal husbandry. However, further research into its pharmacodynamic interactions with veterinary drugs and long-term safety in food-producing animals remains essential to maximize its utility as a safe, natural additive.
Data availability statement
The original contributions presented in the study are included in the article/supplementary material, further inquiries can be directed to the corresponding author.
Author contributions
DP: Conceptualization, Data curation, Formal analysis, Investigation, Methodology, Supervision, Validation, Visualization, Writing – original draft, Writing – review & editing. MG: Data curation, Funding acquisition, Project administration, Resources, Supervision, Validation, Writing – review & editing. MP: Data curation, Funding acquisition, Project administration, Resources, Supervision, Validation, Writing – review & editing. MD: Conceptualization, Data curation, Formal analysis, Funding acquisition, Project administration, Resources, Supervision, Validation, Writing – review & editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. DP was a visiting scientist at the Department of Comparative Biomedicine and Food Science, supported by a grant from the project “animalS and ENvironmenT: toward a sustaINablE Life (SENTINEL)” financed by the italian Ministry of University and Research (MUR) for the period 2023–2027 under the funding scheme Department of Excellence. Open Access funding provided by Università degli Studi di Padova | University of Padua, Open Science Committee.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declared that they were an editorial board member of Frontiers, at the time of submission. This had no impact on the peer review process and the final decision.
Generative AI statement
The author(s) declare that no Gen AI was used in the creation of this manuscript.
Publisher's note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
1. Alagawany M, Abd El-Hack ME, Saeed M, Arain MA, Bhutto ZA, Fazlani SA, et al. Effect of some phytogenic additives as dietary supplements on performance, egg quality, serum biochemical parameters and oxidative status in laying hens. Indian J Anim Sci. (2017) 87:100. doi: 10.56093/ijans.v87i7.72300
2. Amer SA, Metwally AE, Ahmed SA. The influence of dietary supplementation of CIN and thymol on the growth performance, immunity and antioxidant status of monosex Nile tilapia fingerlings (Oreochromis niloticus). Egypt J Aquat Res. (2018) 44:251–6. doi: 10.1016/j.ejar.2018.07.004
3. de Morais SM, Vila-Nova NS, Bevilaqua CML, Rondon FC, Lobo CH, Moura ADAAN, et al. Thymol and eugenol derivatives as potential antileishmanial agents. Bioorg Med Chem. (2014) 22:6250–5. doi: 10.1016/j.bmc.2014.08.020
4. Ianniello IL, Horenstein MB, Labaque MC, Luna A, Marin RH, Gleiser RM. Fly emergence from manure of Japanese quail fed thymol-or isoeugenol-supplemented diets. Poult Sci. (2014) 93:2449–56. doi: 10.3382/ps.2014-03951
5. Paiano RB, de Sousa RL, Bonilla J, Moreno LZ, de Souza ED, Baruselli PS, et al. In vitro effects of cinnamon, oregano, and thyme essential oils against Escherichia coli and Trueperella pyogenes isolated from dairy cows with clinical endometritis. Theriogenology. (2023) 196:106–11. doi: 10.1016/j.theriogenology.2022.11.010
6. Youssefi MR, Tabari MA, Esfandiari A, Kazemi S, Moghadamnia AA, Sut S, et al. Efficacy of two monoterpenoids, carvacrol and thymol, and their combinations against eggs and larvae of the West Nile vector Culex pipiens. Molecules. (2019) 24:1867. doi: 10.3390/molecules24101867
7. Hassan FH, Mansour AM, Salama SA, El-Sayed EM. The chemopreventive effect of thymol against dimethylhydrazine and/or high fat diet-induced colon cancer in rats: relevance to NF-κB. Life Sci. (2021) 1:119335. doi: 10.1016/j.lfs.2021.119335
8. Low SB, Peak RM, Smithson CW, Perrone J, Gaddis B, Kontogiorgos E. Evaluation of a topical gel containing a novel combination of essential oils and antioxidants for reducing oral malodor in dogs. Am J Vet Res. (2014) 75:653–7. doi: 10.2460/ajvr.75.7.653
9. Jesus FPK, Ferreiro L, Bizzi KS, Loreto ÉS, Pilotto MB, Ludwig A, et al. In vitro activity of carvacrol and thymol combined with antifungals or antibacterials against Pythium insidiosum. J Mycol Med. (2015) 25:e89–93. doi: 10.1016/j.mycmed.2014.10.023
10. Tian F, Lee SY, Chun HS. Comparison of the antifungal and antiaflatoxigenic potential of liquid and vapor phase of Thymus vulgaris essential oil against Aspergillus flavus. J Food Prot. (2019) 82:2044–8. doi: 10.4315/0362-028X.JFP-19-016
11. Costa-Júnior LM, Miller RJ, Alves PB, Blank AF, Li AY, de León AAP. Acaricidal efficacies of Lippia gracilis essential oil and its phytochemicals against organophosphate-resistant and susceptible strains of Rhipicephalus (Boophilus) microplus. Vet Parasitol. (2016) 228:60–4. doi: 10.1016/j.vetpar.2016.05.028
12. Monteiro C, Ferreira LL, de Paula LGF, de Oliveira Filho JG, de Oliveira Silva F, Muniz ER, et al. Thymol and eugenol microemulsion for Rhiphicephalus sanguineus sensu lato control: Formulation development, field efficacy, and safety on dogs. Vet Parasitol. (2021) 296:109501. doi: 10.1016/j.vetpar.2021.109501
13. Khalil SR, Abd Elhakim Y, Abd El-fattah AH, Farag MR, Abd El-Hameed NE, Abd Elhakeem EM. Dual immunological and oxidative responses in Oreochromis niloticus fish exposed to lambda cyhalothrin and concurrently fed with Thyme powder (Thymus vulgaris L.): Stress and immune encoding gene expression. Fish Shellfish Immunol. (2020) 100:208–18. doi: 10.1016/j.fsi.2020.03.009
14. Zargar A, Rahimi-Afzal Z, Soltani E, Taheri Mirghaed A, Ebrahimzadeh-Mousavi HA, Soltani M, et al. Growth performance, immune response and disease resistance of rainbow trout (Oncorhynchus mykiss) fed Thymus vulgaris essential oils. Aquac Res. (2019) 50:3097–106. doi: 10.1111/are.14243
15. Wei HK, Xue HX, Zhou ZX, Peng J. A carvacrol-thymol blend decreased intestinal oxidative stress and influenced selected microbes without changing the messenger RNA levels of tight junction proteins in jejunal mucosa of weaning piglets. Animal. (2017) 11:193–201. doi: 10.1017/S1751731116001397
16. Placha I, Bacova K, Zitterl-Eglseer K, Laukova A, Chrastinova L, Madarova M, et al. Thymol in fattening rabbit diet, its bioavailability and effects on intestinal morphology, microbiota from caecal content and immunity. J Anim Physiol Anim Nutr. (2021) 106:368–77. doi: 10.1111/jpn.13595
17. Soares NMM, Bastos TS, Kaelle GCB, de Souza RBMDS, de Oliveira SG, Félix AP. Digestibility and palatability of the diet and intestinal functionality of dogs fed a blend of yeast cell wall and oregano essential oil. Animals. (2023) 13:2527. doi: 10.3390/ani13152527
18. Li SY, Ru YJ, Liu M, Xu B, Péron A, Shi XG. The effect of essential oils on performance, immunity and gut microbial population in weaner pigs. Livest Sci. (2012) 145:119–23. doi: 10.1016/j.livsci.2012.01.005
19. Su G, Wang L, Zhou X, Wu X, Chen D, Yu B, et al. Effects of essential oil on growth performance, digestibility, immunity, and intestinal health in broilers. Poult Sci. (2021) 100:101242. doi: 10.1016/j.psj.2021.101242
20. Dieguez SN, Decundo JM, Martínez G, Amanto FA, Bianchi CP, Pérez Gaudio DS, et al. Effect of dietary oregano (Lippia origanoides) and clover (Eugenia caryophillata) essential oils' formulations on intestinal health and performance of pigs. Planta Med. (2022) 88:324–35. doi: 10.1055/a-1698-8469
21. Attia YA, Al-Harthi MA. Nigella seed oil as an alternative to antibiotic growth promoters for broiler chickens. Eur Poult Sci. (2015) 79:1–13. doi: 10.1399/eps.2015.80
22. Mašek T, Starčević K, Mikulec Ž. The influence of the addition of thymol, tannic acid or gallic acid to broiler diet on growth performance, serum malondialdehyde value and cecal fermentation. Eur Poult Sci. (2014) 78:1–8. doi: 10.1399/eps.2014.64
23. Komaki A, Hoseini F, Shahidi S, Baharlouei N. Study of the effect of extract of Thymus vulgaris on anxiety in male rats. J Tradit Complement Med. (2016) 6:257–61. doi: 10.1016/j.jtcme.2015.01.001
24. Liang D, Li F, Fu Y, Cao Y, Song X, Wang T, et al. Thymol inhibits LPS-stimulated inflammatory response via down-regulation of NF-κB and MAPK signaling pathways in mouse mammary epithelial cells. Inflammation. (2014) 37:214–22. doi: 10.1007/s10753-013-9732-x
25. Omonijo FA, Kim S, Guo T, Wang Q, Gong J, Lahaye L, et al. Development of novel microparticles for effective delivery of thymol and lauric acid to pig intestinal tract. J Agric Food Chem. (2018) 66:9608–15. doi: 10.1021/acs.jafc.8b02808
26. Sapkota M, Li L, Kim SW, Soh Y. Thymol inhibits RANKL-induced osteoclastogenesis in RAW264.7 and BMM cells and LPS-induced bone loss in mice. Food Chem Toxicol. (2018) 120:418–29. doi: 10.1016/j.fct.2018.07.032
27. Ananda Baskaran S, Venkitanarayanan K. Plant-derived antimicrobials reduce Escherichia coli O157:H7 virulence factors critical for colonization in cattle gastrointestinal tract in vitro. Biomed Res Int. (2014) 2014:212395. doi: 10.1155/2014/212395
28. Page MJ, McKenzie JE, Bossuyt PM, Boutron I, Hoffmann TC, Mulrow CD, et al. The PRISMA 2020 statement: an updated guideline for reporting systematic reviews. PLoS Med. (2021) 18:e1003583. doi: 10.1371/journal.pmed.1003583
29. Miró MV, Lifschitz A, Viviani P, Rocha C, Lanusse C, Costa Jr L, et al. In vitro inhibition of the hepatic S-oxygenation of the anthelmintic albendazole by the natural monoterpene thymol in sheep. Xenobiotica. (2020) 50:408–14. doi: 10.1080/00498254.2019.1644390
30. Ortega MT, Jeffery B, Riviere JE, Monteiro-Riviere NA. Toxicological effects of pet food ingredients on canine bone marrow-derived mesenchymal stem cells and enterocyte-like cells. J Appl Toxicol. (2016) 36:189–98. doi: 10.1002/jat.3158
31. Shen C, Christensen LG, Rasmussen PB, Kragh KM. In vitro immunomodulatory effects of THY and CIN in a pig intestinal epithelial cell line (IPEC-J2). J Appl Anim Nutr. (2020) 8:127–34. doi: 10.3920/JAAN2020.0010
32. Nikolić M, Glamočlija J, Ferreira IC, Calhelha RC, Fernandes Â, Marković T, et al. Chemical composition, antimicrobial, antioxidant and antitumor activity of Thymus serpyllum L., Thymus algeriensis Boiss. & Reut., and Thymus vulgaris L. essential oils. Ind Crops Prod. (2014) 52:183–90. doi: 10.1016/j.indcrop.2013.10.006
33. Zhang LW, Koci J, Jeffery B, Riviere JE, Monteiro-Riviere NA. Safety assessment of potential food ingredients in canine hepatocytes. Food Chem Toxicol. (2015) 78:105–15. doi: 10.1016/j.fct.2015.02.003
34. Shen C, Christensen LG, Bak SY, Christensen N, Kragh K. Immunomodulatory effects of thymol and CIN in chicken cell lines. J Appl Anim Nutr. (2020) 8:21–30. doi: 10.3920/JAAN2020.0001
35. Lai WL, Chuang HS, Lee MH, Wei CL, Lin CF, Tsai YC. Inhibition of herpes simplex virus type 1 by thymol-related monoterpenoids. Planta Med. (2012) 78:1636–8. doi: 10.1055/s-0032-1315208
36. Chang HT, Chou CT, Liang WZ, Lu T, Kuo DH, Shieh P, et al. Effects of thymol on Ca2+ homeostasis and apoptosis in MDCK renal tubular cells. Chin J Physiol. (2014) 57:90–8. doi: 10.4077/CJP.2014.BAB192
37. Koči J, Jeffery B, Riviere JE, Monteiro-Riviere NA. In vitro safety assessment of food ingredients in canine renal proximal tubule cells. Toxicol In Vitro. (2015) 29:289–98. doi: 10.1016/j.tiv.2014.11.002
38. Sánchez G, Aznar R. Evaluation of natural compounds of plant origin for inactivation of enteric viruses. Food Environ Virol. (2015) 7:183–7. doi: 10.1007/s12560-015-9181-9
39. Lanave G, Catella C, Catalano A, Lucente MS, Pellegrini F, Fracchiolla G, et al. Assessing the virucidal activity of essential oils against feline calicivirus, a non-enveloped virus used as surrogate of norovirus. Heliyon. (2024) 10:e30492. doi: 10.1016/j.heliyon.2024.e30492
40. Ku CM, Lin JY. Anti-inflammatory effects of 27 selected terpenoid compounds tested through modulating Th1/Th2 cytokine secretion profiles using murine primary splenocytes. Food Chem. (2013) 141:1104–13. doi: 10.1016/j.foodchem.2013.04.044
41. Wu H, Jiang K, Yin N, Ma X, Zhao G, Qiu C, et al. Thymol mitigates lipopolysaccharide-induced endometritis by regulating the TLR4-and ROS-mediated NF-κB signaling pathways. Oncotarget. (2017) 8:20042. doi: 10.18632/oncotarget.15373
42. Liu DM, Zhou CY, Meng XL, Wang P, Li W. Thymol exerts anti-inflammatory effect in dextran sulfate sodium-induced experimental murine colitis. Trop J Pharm Res. (2018) 17:1803–10. doi: 10.4314/tjpr.v17i9.18
43. Fachini-Queiroz FC, Kummer R, Estevao-Silva CF, Carvalho MDDB, Cunha JM, Grespan R, et al. Effects of thymol and carvacrol, constituents of Thymus vulgaris L. essential oil, on the inflammatory response. Evid Based Complement Alternat Med. (2012) 2012:657026. doi: 10.1155/2012/657026
44. Titus EE, Palavesam A, Rajaram SM, Perumal P, Darwin SS, Sanmugapriya NK, et al. In vitro efficacy of plumbagin and thymol against Theileria annulata. J Parasitol Dis. (2023) 47:152–60. doi: 10.1007/s12639-022-01550-x
45. Wei Z, Zhou E, Guo C, Fu Y, Yu Y, Li Y, et al. Thymol inhibits Staphylococcus aureus internalization into bovine mammary epithelial cells by inhibiting NF-κB activation. Microb Pathog. (2014) 71–2:15–9. doi: 10.1016/j.micpath.2014.01.004
46. Jiji S, Udhayakumar S, Rose C, Muralidharan C, Kadirvelu K. Thymol enriched bacterial cellulose hydrogel as effective material for third degree burn wound repair. Int J Biol Macromol. (2019) 122:452–60. doi: 10.1016/j.ijbiomac.2018.10.192
47. Pinheiro André WP, Cavalcante GS, Ribeiro WLC, Santos JMLD, Macedo ITF, Paula HCB, et al. Anthelmintic effect of thymol and thymol acetate on sheep gastrointestinal nematodes and their toxicity in mice. Rev Bras Parasitol Vet. (2017) 26:323–30. doi: 10.1590/s1984-29612017056
48. Baldissera MD, Souza CF, De Matos AFIM, Baldisserotto B, Stefani LM, da Silva AS. Purinergic system as a potential target for inflammation and toxicity induced by thymol in immune cells and tissues. Mol Cell Biochem. (2019) 452:105–10. doi: 10.1007/s11010-018-3416-7
49. Xie K, Tashkin DP, Luo MZ, Zhang JY. Chronic toxicity of inhaled thymol in lungs and respiratory tracts in mouse model. Pharmacol Res Perspect. (2019) 7:e00516. doi: 10.1002/prp2.516
50. Zhang X, Peng Y, Wu C. Chicken embryonic toxicity and potential in vitro estrogenic and mutagenic activity of carvacrol and thymol in low dose/concentration. Food Chem Toxicol. (2021) 150:112038. doi: 10.1016/j.fct.2021.112038
51. Nunes DO, Vinturelle R, Martins FJ, Dos Santos TF, Valverde AL, Ribeiro CMR, et al. Biotechnological potential of eugenol and thymol derivatives against Staphylococcus aureus from bovine mastitis. Curr Microbiol. (2021) 78:1846–55. doi: 10.1007/s00284-021-02344-9
52. Santana LEGS, Miranda IKI, Sousa JA. In silico analysis of the pharmacokinetics, pharmacodynamics and toxicity of two compounds isolated from Moringa oleifera. Res Soc Dev. (2020) 9:e81991110469. doi: 10.33448/rsd-v9i11.10469
53. Verdú JR, Cortez V, Rosa-García R, Ortiz AJ, García-Prieto U, Lumaret JP, et al. Nontoxic effects of thymol, carvacrol, CIN, and garlic oil on dung beetles: a potential alternative to ecotoxic anthelmintics. PLoS ONE. (2023) 18:e0295753. doi: 10.1371/journal.pone.0295753
54. Gan C, Langa E, Ballestero D, Pino-Otín MR. Comparative ecotoxicity assessment of highly bioactive isomeric monoterpenes carvacrol and thymol on aquatic and edaphic indicators and communities. Chemosphere. (2024) 368:143666. doi: 10.1016/j.chemosphere.2024.143666
55. Xie K, Tashkin DP, Luo MZ, Zhang JY. Pharmacokinetic study of thymol after intravenous injection and high-dose inhalation in mouse model. Pharmacol Res Perspect. (2019) 7:e00515. doi: 10.1002/prp2.515
56. Bacova K, Eglseer KZ, Räuber GK, Chrastinova L, Laukova A, Takacsova M, et al. Effect of sustained administration of thymol on its bioaccessibility and bioavailability in rabbits. Animals. (2021) 11:2595. doi: 10.3390/ani11092595
57. Haselmeyer A, Zentek J, Chizzola R. Effects of thyme as a feed additive in broiler chickens on thymol in gut contents, blood plasma, liver and muscle. J Sci Food Agric. (2015) 95:504–8. doi: 10.1002/jsfa.6758
58. Ocel'ova V, Chizzola R, Pisarčíková J, Novak J, Ivanišinová O, Faix Š, et al. Effect of thyme essential oil supplementation on thymol content in blood plasma, liver, kidney and muscle in broiler chickens. Nat Prod Commun. (2016) 11:1934578X1601101031. doi: 10.1177/1934578X1601101031
59. Placha I, Ocelova V, Chizzola R, Battelli G, Gai F, Bacova K, et al. Effect of thymol on the broiler chicken antioxidative defence system after sustained dietary thyme oil application. Br Poult Sci. (2019) 60:589–96. doi: 10.1080/00071668.2019.1631445
60. Petrujkić BT, Sedej I, Beier RC, Anderson RC, Harvey RB, Epps SV, et al. Ex vivo absorption of thymol and thymol-β-D-glucopyranoside in piglet everted jejunal segments. J Agric Food Chem. (2013) 61:3757–62. doi: 10.1021/jf401013a
61. Nieddu M, Rassu G, Boatto G, Bosi P, Trevisi P, Giunchedi P, et al. Improvement of thymol properties by complexation with cyclodextrins: in vitro and in vivo studies. Carbohydr Polym. (2014) 15:102:393-9. doi: 10.1016/j.carbpol.2013.10.084
62. Rassu G, Nieddu M, Bosi P, Trevisi P, Colombo M, Priori D, et al. Encapsulation and modified-release of thymol from oral microparticles as adjuvant or substitute to current medications. Phytomedicine. (2014) 21:1627–32. doi: 10.1016/j.phymed.2014.07.017
63. Armorini S, Yeatts JE, Mullen KA, Mason SE, Mehmeti E, Anderson KL, et al. Development of a HS-SPME-GC-MS/MS method for the quantitation of thymol and carvacrol in bovine matrices and to determine residue depletion in milk and tissues. J Agric Food Chem. (2016) 64:7856–65. doi: 10.1021/acs.jafc.6b02899
64. Mason SE, Mullen KA, Anderson KL, Washburn SP, Yeatts JL, Baynes RE. Pharmacokinetic analysis of thymol, carvacrol and diallyl disulfide after intramammary and topical applications in healthy organic dairy cattle. Food Addit Contam Part A. (2017) 34:740–9. doi: 10.1080/19440049.2017.1285056
65. Miró MV, Silva CR, Viviani P, Luque S, Lloberas M, Costa-Júnior LM, et al. Combination of bioactive phytochemicals and synthetic anthelmintics: in vivo and in vitro assessment of the albendazole-thymol association. Vet Parasitol. (2020) 281:109–21. doi: 10.1016/j.vetpar.2020.109121
66. Kovačević Z, Tomanić D, Cabarkapa I, Šarić L, Stanojević J, Bijelić K, et al. Chemical composition, antimicrobial activity, and withdrawal period of essential oil-based pharmaceutical formulation in bovine mastitis treatment. Int J Environ Res Public Health. (2022) 19:16643. doi: 10.3390/ijerph192416643
67. Castelani L, Pfrimer K, Giglioti R, van Cleef EHCB, Salles MSV, Júnior LCR. Effects of thyme (Thymus vulgaris L.) essential oil supplementation on the microbiological quality of raw milk of lactating dairy cows. Res Vet Sci. (2023) 161:118–121. doi: 10.1016/j.rvsc.2023.06.013
68. Pisarčíková J, Ocelová V, Faix Š, Placha I, Calderon AI. Identification and quantification of thymol metabolites in plasma, liver and duodenal wall of broiler chickens using UHPLC-ESI-QTOF-MS. Biomed Chromatogr. (2017) 31:e3881. doi: 10.1002/bmc.3881
69. Avila Ramos F, Pro Martínez A, Sosa Montes E, Narciso Gaytán C, Hernández Cázarez AS, Cibrián Tovar J, et al. Oregano oil use in broiler diet increases accumulation of carvacrol and thymol in breast meat. Acta Univ. (2017) 27:34–9. doi: 10.15174/au.2017.1074
70. Ocelová V, Chizzola R, Battelli G, Pisarcikova J, Faix S, Gai F, et al. Thymol in the intestinal tract of broiler chickens after sustained administration of thyme essential oil in feed. J Anim Physiol Anim Nutr. (2019) 103:204–9. doi: 10.1111/jpn.12995
71. Fernandez ME, Kembro JM, Ballesteros ML, Caliva JM, Marin RH, Labaque MC. Dynamics of thymol dietary supplementation in quail (Coturnix japonica): Linking bioavailability, effects on egg yolk total fatty acids and performance traits. PLoS ONE. (2019) 14:e0216623. doi: 10.1371/journal.pone.0216623
72. Farhadi K, Rajabi E, Varpaei HA, Iranzadasl M, Khodaparast S, Salehi M. Thymol and carvacrol against Klebsiella: anti-bacterial, anti-biofilm, and synergistic activities—a systematic review. Front Pharmacol. (2024) 15:1487083. doi: 10.3389/fphar.2024.1487083
73. Kowalczyk A, Przychodna M, Sopata S, Bodalska A, Fecka I. Thymol and thyme essential oil—New insights into selected therapeutic applications. Molecules. (2020) 25:4125. doi: 10.3390/molecules25184125
74. Nagoor Meeran MF, Javed H, Al Taee H, Azimullah S, Ojha SK. Pharmacological properties and molecular mechanisms of thymol: prospects for its therapeutic potential and pharmaceutical development. Front Pharmacol. (2017) 8:380. doi: 10.3389/fphar.2017.00380
75. Bianchini AE, Garlet QI, Da Cunha JA, Bandeira Junior G, Brusque ICM, Salbego J, et al. Monoterpenoids (thymol, carvacrol and S-(+)-linalool) with anesthetic activity in silver catfish (Rhamdia quelen): evaluation of acetylcholinesterase and GABAergic activity. Braz J Med Biol Res. (2017) 50:e6346. doi: 10.1590/1414-431x20176346
76. Premrov Bajuk B, Prem L, Vake T, Žnidaršič N, Snoj T. The effect of thymol on acetylcholine-induced contractions of the rat ileum and uterus under ex vivo conditions. Front Pharmacol. (2022) 13:990654. doi: 10.3389/fphar.2022.990654
77. Heo GJ, Kim CH, Park SC, De Zoysa M, Shin GW. Antimicrobial activity of thymol against pathogenic gram-negative bacteria of fishes. Philipp J Vet Med. (2012) 49:103–6.
78. Sfeir J, Lefrançois C, Baudoux D, Derbré S, Licznar P. In vitro antibacterial activity of essential oils against Streptococcus pyogenes. Evid Based Complement Alternat Med. (2013) 2013:269161. doi: 10.1155/2013/269161
79. Mina K, Salima B, Abdelghani D, Saoud A. Antipseudomonal activity of the essential oil of Thymus numidicus Poiret. Int J Pharm Sci Rev Res. (2014) 25:49–153. Available online at: http://www.globalresearchonline.net
80. Dehghani N, Afsharmanesh M, Salarmoini M, Ebrahimnejad H. In vitro and in vivo evaluation of thyme (Thymus vulgaris) essential oil as an alternative for antibiotic in quail diet. J Anim Sci. (2019) 97:2901–13. doi: 10.1093/jas/skz179
81. Al-Kandari F, Al-Temaimi R, van Vliet AH, Woodward MJ. Thymol tolerance in Escherichia coli induces morphological, metabolic and genetic changes. BMC Microbiol. (2019) 19:1–11. doi: 10.1186/s12866-019-1663-8
82. Sim JX, Khazandi M, Chan WY, Trott DJ, Deo P. Antimicrobial activity of thyme oil, oregano oil, thymol and carvacrol against sensitive and resistant microbial isolates from dogs with otitis externa. Vet Dermatol. (2019) 30:524–e159. doi: 10.1111/vde.12794
83. Arena JS, Merlo C, Defagó MT, Zygadlo JA. Insecticidal and antibacterial effects of some essential oils against the poultry pest Alphitobius diaperinus and its associated microorganisms. J Pest Sci. (2020) 93:403–14. doi: 10.1007/s10340-019-01141-5
84. Hriouech S, Akhmouch AA, Tanghort M, Chefchaou H, Mzabi A, Chami N, et al. In vitro and in vivo comparison of changes in antibiotics susceptibility of E. coli and chicken's intestinal flora after exposure to amoxicillin or thymol. Vet Med Int. (2020) 2020:8824008. doi: 10.1155/2020/8824008
85. Levent G, Anderson RC, Petrujkić B, Poole TL, He H, Genovese KJ, et al. Evaluation of thymol-β-d-glucopyranoside as a potential prebiotic intervention to reduce carriage of zoonotic pathogens in weaned and feeder pigs. Microorganisms. (2021) 9:860. doi: 10.3390/microorganisms9040860
86. Gómez-García M, Madrigal I, Puente H, Mencía-Ares Ó, Argüello H, Carvajal A, et al. In vitro activity of essential oils against microbial isolates from otitis externa cases in dogs. Nat Prod Res. (2021) 36:4546–50. doi: 10.1080/14786419.2021.1993217
87. Nojo H, Ishijima SA, Morikawa M, Ito T, Kano R. In vitro susceptibility testing of phytochemicals from essential oils against Prototheca species. J Vet Med Sci. (2024) 86:847–9. doi: 10.1292/jvms.24-0032
88. Catella C, Camero M, Lucente MS, Fracchiolla G, Sblano S, Tempesta M, et al. Virucidal and antiviral effects of Thymus vulgaris essential oil on feline coronavirus. Res Vet Sci. (2021) 137:44–7. doi: 10.1016/j.rvsc.2021.04.024
89. De Lira Mota KSDL, Pereira FDO, De Oliveira WA, Lima IO, Lima EDO. Antifungal activity of Thymus vulgaris L. essential oil and its constituent phytochemicals against Rhizopus oryzae: interaction with ergosterol. Molecules. (2012) 17:14418–33. doi: 10.3390/molecules171214418
90. Alsafah AH, Al-Faragi JK. Influence of thyme (Thymus vulgaris) as feed additives on growth performance and antifungal activity on Saprolegnia spp. in Cyprinus carpio L. J Entomol Zool Stud. (2017) 5:1598–602. Available online at: http://www.entomoljournal.com
91. Michalczyk A, Ostrowska P. Essential oils and their components in combating fungal pathogens of animal and human skin. J Med Mycol. (2021) 31:101118. doi: 10.1016/j.mycmed.2021.101118
92. Bismarck D, Dusold A, Heusinger A, Müller E. Antifungal in vitro activity of essential oils against clinical isolates of Malassezia pachydermatis from canine ears: a report from a practice laboratory. Complement Med Res. (2020) 27:143–54. doi: 10.1159/000504316
93. Aboelhadid SM, Abdel-Baki AAS, Ibrahium SM, Al-Quraishy S, Gadelhaq SM, Arafa WM, et al. The efficacy of essential oil components with ivermectin against Rhipicephalus annulatus: an in-vitro study. Vet Parasitol. (2024) 332:110335. doi: 10.1016/j.vetpar.2024.110335
94. Tabari MA, Youssefi MR, Maggi F, Benelli G. Toxic and repellent activity of selected monoterpenoids (thymol, carvacrol and linalool) against the castor bean tick, Ixodes ricinus (Acari: Ixodidae). Vet Parasitol. (2017) 245:86–91. doi: 10.1016/j.vetpar.2017.08.012
95. Masoumi F, Youssefi MR, Tabari MA. Combination of carvacrol and thymol against the poultry red mite (Dermanyssus gallinae). Parasitol Res. (2016) 115:4239–43. doi: 10.1007/s00436-016-5201-4
96. Remmal A, Achahbar S, Bouddine L, Chami F, Chami N. Oocysticidal effect of essential oil components against chicken Eimeria oocysts. Int J Vet Med. (2013) 2:133–9. doi: 10.5171/2013.599816
97. Felici M, Tugnoli B, Ghiselli F, Massi P, Tosi G, Fiorentini L, et al. In vitro anticoccidial activity of thymol, carvacrol, and saponins. Poultry Sci. (2020) 99:5350–5. doi: 10.1016/j.psj.2020.07.035
98. Pensel PE, Maggiore MA, Gende LB, Eguaras MJ, Denegri MG, Elissondo MC. Efficacy of essential oils of Thymus vulgaris and Origanum vulgare on Echinococcus granulosus. Interdiscip Perspect Infect Dis. (2014) 2014:693289. doi: 10.1155/2014/693289
99. Maggiore M, Elissondo MC. In vitro cestocidal activity of thymol on Mesocestoides corti tetrathyridia and adult worms. Interdiscip Perspect Infect Dis. (2014) 2014:268135. doi: 10.1155/2014/268135
100. Ferreira LE, Benincasa BI, Fachin AL, Franca SC, Contini SS, Chagas AC, et al. Thymus vulgaris L. essential oil and its main component thymol: anthelmintic effects against Haemonchus contortus from sheep. Vet Parasitol. (2016) 228:70–6. doi: 10.1016/j.vetpar.2016.08.011
101. Katiki LM, Barbieri AME, Araujo RC, Veríssimo CJ, Louvandini H, Ferreira JFS. Synergistic interaction of ten essential oils against Haemonchus contortus in vitro. Vet Parasitol. (2017) 243:47–51. doi: 10.1016/j.vetpar.2017.06.008
102. Štrbac F, Bosco A, Maurelli MP, Ratajac R, Stojanović D, Simin N, et al. Anthelmintic properties of essential oils to control gastrointestinal nematodes in sheep-In vitro and in vivo studies. Vet Sci. (2022) 9:93. doi: 10.3390/vetsci9020093
103. Youssefi MR, Moghaddas E, Tabari MA, Moghadamnia AA, Hosseini SM, Farash BRH, et al. In vitro and in vivo effectiveness of carvacrol, thymol and linalool against Leishmania infantum. Molecules. (2019) 24:2072. doi: 10.3390/molecules24112072
104. Tanghort M, Chefchaou H, Mzabi A, Moussa H, Chami N, Chami F, et al. Oocysticidal effect of essential oils (EOs) and their major components on Cryptosporidium baileyi and Cryptosporidium galli. Int J Poult Sci. (2019) 18:475–82. doi: 10.3923/ijps.2019.475.482
105. Riella KR, Marinho RR, Santos JS, Pereira-Filho RN, Cardoso JC, Albuquerque-Junior RLC, et al. Anti-inflammatory and cicatrizing activities of THY, a monoterpene of the essential oil from Lippia gracilis, in rodents. J Ethnopharmacol. (2012) 143:656–63. doi: 10.1016/j.jep.2012.07.028
106. Aboelwafa HR, Yousef HN. The ameliorative effect of thymol against hydrocortisone-induced hepatic oxidative stress injury in adult male rats. Biochem Cell Biol. (2015) 93:282–9. doi: 10.1139/bcb-2014-0154
107. Games E, Guerreiro M, Santana FR, Pinheiro NM, De Oliveira EA, Lopes FD, et al. Structurally related monoterpenes p-cymene, carvacrol and thymol isolated from essential oil from leaves of Lippia sidoides Cham (Verbenaceae) protect mice against elastase-induced emphysema. Molecules. (2016) 21:1390. doi: 10.3390/molecules21101390
108. Yao L, Hou G, Wang L, Zuo XS, Liu Z. Protective effects of thymol on LPS-induced acute lung injury in mice. Microb Pathog. (2018) 116:8–12. doi: 10.1016/j.micpath.2017.12.065
109. Geyikoglu F, Yilmaz EG, Erol HS, Koc K, Cerig S, Ozek NS, et al. Hepatoprotective role of THY in drug-induced gastric ulcer model. Ann Hepatol. (2019) 17:980–91. doi: 10.5604/01.3001.0012.7198
110. Chamanara M, Abdollahi A, Rezayat SM, Ghazi-Khansari M, Dehpour A, Nassireslami E, et al. Thymol reduces acetic acid-induced inflammatory response through inhibition of NF-kB signaling pathway in rat colon tissue. Inflammopharmacology. (2019) 27:1275–83. doi: 10.1007/s10787-019-00583-8
111. Du E, Wang W, Gan L, Li Z, Guo S, Guo Y. Effects of thymol and carvacrol supplementation on intestinal integrity and immune responses of broiler chickens challenged with Clostridium perfringens. J Anim Sci Biotechnol. (2016) 7:1–10. doi: 10.1186/s40104-016-0079-7
112. Yu YM, Chao TY, Chang WC, Chang MJ, Lee MF. Thymol reduces oxidative stress, aortic intimal thickening, and inflammation-related gene expression in hyperlipidemic rabbits. J Food Drug Anal. (2016) 24:556–63. doi: 10.1016/j.jfda.2016.02.004
113. Ghiselli F, Yihang L, Liang-en Y, Tugnoli B, Piva A, Grilli E. Thymol and thyme essential oil can reduce LPS-induced inflammation and disruptive effects on apical-out chicken enteroids. Poult Sci Assoc Annu Meet Abstr. (2023) 307:148–9.
114. Giannenas I, Triantafillou E, Stavrakakis S, Margaroni M, Mavridis S, Steiner T, et al. Assessment of dietary supplementation with carvacrol or thymol containing feed additives on performance, intestinal microbiota and antioxidant status of rainbow trout (Oncorhynchus mykiss). Aquaculture. (2012) 350:26–32. doi: 10.1016/j.aquaculture.2012.04.027
115. Sönmez AY, Bilen S, Alak G, Hisar O, Yanik T, Biswas G. Growth performance and antioxidant enzyme activities in rainbow trout (Oncorhynchus mykiss) juveniles fed diets supplemented with sage, mint and thyme oils. Fish Physiol Biochem. (2015) 41:165–75. doi: 10.1007/s10695-014-0014-9
116. Hoseini SM, Yousefi M. Beneficial effects of thyme (Thymus vulgaris) extract on oxytetracycline-induced stress response, immunosuppression, oxidative stress and enzymatic changes in rainbow trout (Oncorhynchus mykiss). Aquac Nutr. (2019) 25:298–309. doi: 10.1111/anu.12853
117. Yousefi M, Hoseini SM, Vatnikov YA, Nikishov AA, Kulikov EV. Thymol as a new anesthetic in common carp (Cyprinus carpio): efficacy and physiological effects in comparison with eugenol. Aquaculture. (2018) 495:376–83. doi: 10.1016/j.aquaculture.2018.06.022
118. Amin MM, El-Kabany H. Evaluation of protective and treatment of thyme (Thymus vulgaris) oil on Toxocara vitulorum infected rats. J Radiat Res Appl Sci. (2013) 6:209–32.
119. Rubió L, Serra A, Chen CYO, Macia A, Romero MP, Covas MI, et al. Effect of the co-occurring components from olive oil and thyme extracts on the antioxidant status and its bioavailability in an acute ingestion in rats. Food Funct. (2014) 5:740–7. doi: 10.1039/c3fo60446b
120. Bacova K, Zitterl-Eglseer K, Chrastinova L, Laukova A, Madarova M, Gancarcikova S, et al. Effect of thymol addition and withdrawal on some blood parameters, antioxidative defence system and fatty acid profile in rabbit muscle. Animals. (2020) 10:1248. doi: 10.3390/ani10081248
121. Zeng Z, Xu X, Zhang Q, Li P, Zhao P, Li Q, et al. Effects of essential oil supplementation of a low-energy diet on performance, intestinal morphology and microflora, immune properties and antioxidant activities in weaned pigs. Anim Sci J. (2015) 86:279–85. doi: 10.1111/asj.12277
122. Luo P, Luo L, Zhao W, Wang L, Sun L, Wu H, et al. Dietary thymol supplementation promotes skeletal muscle fibre type switch in longissimus dorsi of finishing pigs. J Anim Physiol Anim Nutr. (2020) 104:570–8. doi: 10.1111/jpn.13269
123. Grando MA, Costa V, Genova JL, Rupolo PE, de Azevedo LB, Costa LB, et al. Blend of essential oils can reduce diarrheal disorders and improve liver antioxidant status in weaning piglets. Anim Biosci. (2023) 36:119. doi: 10.5713/ab.22.0072
124. Azhar J, Nadeem A, Javed M, Ahmad HI, Hassan FU, Shah FS. Evaluation of phytochemicals from Thymus serpyllum as potential drug candidates to manage oxidative stress in transition dairy cows. J Biomol Struct Dyn. (2024) 42:2897–912. doi: 10.1080/07391102.2023.2209190
125. Hashemipour H, Kermanshahi H, Golian A, Veldkamp T. Effect of thymol and carvacrol feed supplementation on performance, antioxidant enzyme activities, fatty acid composition, digestive enzyme activities, and immune response in broiler chickens. Poult Sci. (2013) 92:2059–69. doi: 10.3382/ps.2012-02685
126. Saadat Shad H, Mazhari M, Esmaeilipour O, Khosravinia H. Effects of thymol and carvacrol on productive performance, antioxidant enzyme activity and certain blood metabolites in heat stressed broilers. Iran J Appl Anim Sci. (2016) 6:195–202.
127. Giannenas I, Papaneophytou CP, Tsalie E, Pappas I, Triantafillou E, Tontis D, et al. Dietary supplementation of benzoic acid and essential oil compounds affects buffering capacity of the feeds, performance of turkey poults and their antioxidant status, pH in the digestive tract, intestinal microbiota and morphology. Asian-Australas J Anim Sci. (2014) 27:225. doi: 10.5713/ajas.2013.13376
128. Abd El-Hack ME, Alagawany M. Performance, egg quality, blood profile, immune function, and antioxidant enzyme activities in laying hens fed diets with thyme powder. J Anim Feed Sci. (2015) 24:127–33. doi: 10.22358/jafs/65638/2015
129. Caetano Filho FF, Paulino LR, Bezerra VS, Azevedo VA, Barroso PA, Costa FC, et al. Thymol increases primordial follicle activation, protects stromal cells, collagen fibers and down-regulates expression of mRNA for superoxide dismutase 1, catalase and periredoxin 6 in cultured bovine ovarian tissues. Anim Reprod Sci. (2024) 266:107514. doi: 10.1016/j.anireprosci.2024.107514
130. Duru Ö, Gökpinar S, Duru SY, Akanbong E, Kara E. The therapeutic effectiveness of thyme extract in naturally infected puppies with ascariasis. Turk J Vet Res. (2023) 7:39–45. doi: 10.47748/tjvr.1226644
131. Hosseini SA, Meimandipour A, Alami F, Mahdavi A, Mohiti-Asli M, Lotfollahian H, et al. Effects of ground thyme and probiotic supplements in diets on broiler performance, blood biochemistry and immunological response to sheep red blood cells. Ital J Anim Sci. (2013) 12:e19. doi: 10.4081/ijas.2013.e19
132. Zhu X, Liu W, Yuan S, Chen H. The effect of different dietary levels of thyme essential oil on serum biochemical indices in Mahua broiler chickens. Ital J Anim Sci. (2014) 13:3238. doi: 10.4081/ijas.2014.3238
133. Diao H, Zheng P, Yu B, He J, Mao X, Yu J, et al. Effects of benzoic acid and thymol on growth performance and gut characteristics of weaned piglets. Asian-Australas J Anim Sci. (2015) 28:827. doi: 10.5713/ajas.14.0704
134. Attia YA, Bakhashwain AA, Bertu NK. Thyme oil (Thymus vulgaris L) as a natural growth promoter for broiler chickens reared under hot climate. Ital J Anim Sci. (2017) 16:275–82. doi: 10.1080/1828051X.2016.1245594
135. Hassan FA, Awad A. Impact of thyme powder (Thymus vulgaris L.) supplementation on gene expression profiles of cytokines and economic efficiency of broiler diets. Environ Sci Pollut Res Int. (2017) 24:15816–26. doi: 10.1007/s11356-017-9251-7
136. Attia YA, Bakhashwain AA, Bertu NK. Utilisation of thyme powder (Thymus vulgaris L.) as a growth promoter alternative to antibiotics for broiler chickens raised in a hot climate. Eur Poult Sci. (2018) 82:238. doi: 10.1399/eps.2018.238
137. Nazarizadeh H, Mohammad Hosseini S, Pourreza J. Effect of plant extracts derived from thyme and chamomile on the growth performance, gut morphology and immune system of broilers fed aflatoxin B1 and ochratoxin A contaminated diets. Ital J Anim Sci. (2019) 18:1073–81. doi: 10.1080/1828051X.2019.1615851
138. Ghanima MMA, Alagawany M, Abd El-Hack ME, Taha A, Elnesr SS, Ajarem J, et al. Consequences of various housing systems and dietary supplementation of thymol, carvacrol, and euganol on performance, egg quality, blood chemistry, and antioxidant parameters. Poult Sci. (2020) 99:4384–97. doi: 10.1016/j.psj.2020.05.028
139. Youssef IM, Männer K, Zentek J. Effect of essential oils or saponins alone or in combination on productive performance, intestinal morphology and digestive enzymes' activity of broiler chickens. J Anim Physiol Anim Nutr. (2021) 105:99–107. doi: 10.1111/jpn.13431
140. Akbari M, Torki M, Kaviani K. Single and combined effects of peppermint and thyme essential oils on productive performance, egg quality traits, and blood parameters of laying hens reared under cold stress condition (6.8±3 °C). Int J Biometeorol. (2016) 60:447–54. doi: 10.1007/s00484-015-1042-6
141. Dehghani N, Afsharmanesh M, Salarmoini M, Ebrahimnejad H, Bitaraf A. Effect of pennyroyal, savory and thyme essential oils on Japanese quail physiology. Heliyon. (2018) 4:e00881. doi: 10.1016/j.heliyon.2018.e00881
142. Abouelezz K, Abou-Hadied M, Yuan J, Elokil AA, Wang G, Wang S, et al. Nutritional impacts of dietary oregano and enviva essential oils on the performance, gut microbiota and blood biochemicals of growing ducks. Animal. (2019) 13:2216–22. doi: 10.1017/S1751731119000508
143. Arafa WM, Abolhadid SM, Moawad A, Abdelaty AS, Moawad UK, Shokier KA, et al. Thymol efficacy against coccidiosis in pigeon (Columba livia domestica). Prev Vet Med. (2020) 176:104914. doi: 10.1016/j.prevetmed.2020.104914
144. Gültepe N, Bilen S, Yilmaz S, Güroy D, Aydin S. Effects of herbs and spice on health status of tilapia (Oreochromis mossambicus) challenged with Streptococcus iniae. Acta Vet Brno. (2014) 83:125–31. doi: 10.2754/avb201483020125
145. Gulec AK, Danabas D, Ural M, Seker E, Arslan A, Serdar O. Effect of mixed use of thyme and fennel oils on biochemical properties and electrolytes in rainbow trout as a response to Yersinia ruckeri infection. Acta Vet Brno. (2013) 82:297–302. doi: 10.2754/avb201382030297
146. Hafsan H, Saleh MM, Zabibah RS, Obaid RF, Jabbar HS, Mustafa YF, et al. Dietary thymol improved growth, body composition, digestive enzyme activities, hematology, immunity, antioxidant defense, and resistance to Streptococcus iniae in the rainbow trout (Oncorhynchus mykiss). Aquac Nutr. (2022) 2022:3288139. doi: 10.1155/2022/3288139
147. Boaventura TP, dos Santos FAC, de Sena Souza A, Batista FS, Júlio GSC, Luz RK. Thymol and linalool chemotypes of the essential oil of Thymus vulgaris (thyme) as anesthetic for Colossoma macropomum: physiology and feed consumption. Aquaculture. (2022) 554:738161. doi: 10.1016/j.aquaculture.2022.738161
148. El Euony OI, Elblehi SS, Abdel-Latif HM, Abdel-Daim MM, El-Sayed YS. Modulatory role of dietary Thymus vulgaris essential oil and Bacillus subtilis against thiamethoxam-induced hepatorenal damage, oxidative stress, and immunotoxicity in African catfish (Clarias garipenus). Environ Sci Pollut Res. (2020) 27:23108–28 doi: 10.1007/s11356-020-08588-5
149. Li P, Piao X, Ru Y, Han X, Xue L, Zhang H. Effects of adding essential oil to the diet of weaned pigs on performance, nutrient utilization, immune response and intestinal health. Asian-Australas J Anim Sci. (2012) 25:1617. doi: 10.5713/ajas.2012.12292
150. Yuan C, Chen S, Sun R, Ren L, Zhao T, Wu M, et al. Thymol improves the growth performance of blue foxes by regulating the gut microbiota. Front Microbiol. (2024) 15:1368293. doi: 10.3389/fmicb.2024.1368293
151. Sadeghi GH, Karimi A, Padidar Jahromi SH, Azizi T, Daneshmand A. Effects of cinnamon, thyme and turmeric infusions on the performance and immune response in 1-to 21-day-old male broilers. Braz J Poult Sci. (2012) 14:15–20. doi: 10.1590/S1516-635X2012000100003
152. Fallah R, Mirzaei E. Effect of dietary inclusion of turmeric and thyme powders on performance, blood parameters and immune system of broiler chickens. J Livest Sci. (2016) 7:180–6.
153. Deniz GY. The protective effects of thymol against ketoprofen-induced damages on pancreatic acinar and islet of langerhans cells in rats. J Essent Oil Bear Plants. (2019) 22:604–13. doi: 10.1080/0972060X.2019.1625814
154. Abu El Ezz NMT, Aboelsoued D, Hassan SE, Abdel Megeed KN, El-Metenawy TM. Therapeutic effect of Moringa oleifera and Thymus vulgaris oils against hepatic coccidiosis in experimentally infected rabbits. Trop Biomed. (2020) 37:1018–28. doi: 10.47665/tb.37.4.1018
155. Cerisuelo A, Marín C, Sánchez-Vizcaino F, Gómez EA, De La Fuente JM, Durán R, et al. The impact of a specific blend of essential oil components and sodium butyrate in feed on growth performance and Salmonella counts in experimentally challenged broilers. Poult Sci. (2014) 93:599–606. doi: 10.3382/ps.2013-03528
156. Arsi K, Donoghue AM, Venkitanarayanan K, Kollanoor-Johny A, Fanatico AC, Blore PJ, et al. The efficacy of the natural plant extracts, thymol and carvacrol against Campylobacter colonization in broiler chickens. J Food Saf. (2014) 34:321–5. doi: 10.1111/jfs.12129
157. Stingelin GM, Scherer RS, Machado AC, Piva A, Grilli E, Penha Filho RC. The use of THY, carvacrol and sorbic acid in microencapsules to control Salmonella Heidelberg, S. Minnesota and S. Typhimurium in broilers. Front Vet Sci. (2023) 9:1046395. doi: 10.3389/fvets.2022.1046395
158. Nazar FN, Videla EA, Marin RH. Thymol supplementation effects on adrenocortical, immune and biochemical variables recovery in Japanese quail after exposure to chronic heat stress. Animal. (2019) 13:318–25. doi: 10.1017/S175173111800157X
159. Tomazelli Júnior O, Kuhn F, Mendonça Padilha PJ, Nunes Nesi C, Mestres M, Dal Magro J, et al. Effect of microencapsulated thyme essential oil on white spot virus-infected Litopenaeus vannamei. Aquac Int. (2018) 26:1459–68. doi: 10.1007/s10499-018-0296-5
160. Feizi A, Bijanzad P, Kaboli K. Effects of thyme volatile oils on performance of broiler chickens. Eur J Exp Biol. (2013) 3:250–4.
161. Hady MM, Zaki MM, Abd El-Ghany W, Korany Reda MS. Assessment of the broilers performance, gut healthiness and carcass characteristics in response to dietary inclusion of dried coriander, turmeric and thyme. Int J Environ Agric Res. (2016) 2:153–9.
162. Hafeez A, Männer K, Schieder C, Zentek J. Effect of supplementation of phytogenic feed additives (powdered vs. encapsulated) on performance and nutrient digestibility in broiler chickens. Poult Sci. (2016) 95:622–9. doi: 10.3382/ps/pev368
163. Reyer H, Zentek J, Manner K, Youssef IM, Aumiller T, Weghuber J, et al. Possible molecular mechanisms by which an essential oil blend from star anise, rosemary, thyme, and oregano and saponins increase the performance and ileal protein digestibility of growing broilers. J Agric Food Chem. (2017) 65:6821–30. doi: 10.1021/acs.jafc.7b01925
164. Gholami-Ahangaran M, Peimani N, Ahmadi-Dastgerdi A. The effect of thyme (Thymus daenensis) supplement on growth and hygienic parameters of broilers meat. Iraqi J Vet Sci. (2019) 33:1048. doi: 10.33899/ijvs.2019.125526.1048
165. Lee JW, Kim DH, Kim YB, Jeong SB, Oh ST, Cho SY, et al. Dietary encapsulated essential oils improve production performance of coccidiosis-vaccine-challenged broiler chickens. Animals. (2020) 10:481. doi: 10.3390/ani10030481
166. Ramirez SY, Peñuela-Sierra LM, Ospina MA. Effects of oregano (Lippia origanoides) essential oil supplementation on the performance, egg quality, and intestinal morphometry of Isa Brown laying hens. Vet World. (2021) 14:595. doi: 10.14202/vetworld.2021.595-602
167. Luna A, Dambolena JS, Zygadlo JA, Marin RH, Labaque MC. Effects of thymol and isoeugenol feed supplementation on quail adult performance, egg characteristics and hatching success. Br Poult Sci. (2012) 53:631–9. doi: 10.1080/00071668.2012.721536
168. Anderson RC, Krueger NA, Genovese KJ, Stanton TB, MacKinnon KM, Harvey RB, et al. Effect of thymol or diphenyliodonium chloride on performance, gut fermentation characteristics, and Campylobacter colonization in growing swine. J Food Prot. (2012) 75:758–61. doi: 10.4315/0362-028X.JFP-11-390
169. Toschi A, Tugnoli B, Rossi B, Piva A, Grilli E. Thymol modulates the endocannabinoid system and gut chemosensing of weaning pigs. BMC Vet Res. (2020) 16:1–11. doi: 10.1186/s12917-020-02516-y
170. Abdelli N, Francisco Pérez J, Vilarrasa E, Melo-Duran D, Cabeza Luna I, Karimirad R, et al. Microencapsulation improved fumaric acid and thymol effects on broiler chickens challenged with a short-term fasting period. Front Vet Sci. (2021) 8:686143. doi: 10.3389/fvets.2021.686143
171. Benchaar C. Diet supplementation with thyme oil and its main component thymol failed to favorably alter rumen fermentation, improve nutrient utilization, or enhance milk production in dairy cows. J Dairy Sci. (2021) 104:324–36. doi: 10.3168/jds.2020-18401
172. Rhouma Raouia B, Boubaker B, Chouaibi M, Aziza G. Effect of supplementing diets with thymol on growth performance and meat quality of growing rabbit under heat season. Egypt J Nutr Feeds. (2021) 24:75–80. doi: 10.21608/ejnf.2021.210782
173. Yu J, Cai L, Zhang J, Yang A, Wang Y, Zhang L, et al. Effects of thymol supplementation on goat rumen fermentation and rumen microbiota in vitro. Microorganisms. (2020) 8:1160. doi: 10.3390/microorganisms8081160
174. Fernandez ME, Marin RH, Luna A, Zunino MP, Labaque MC. Thymol feed supplementation in quail alters the percentages of nutritionally relevant egg yolk fatty acids: effects throughout incubation. J Sci Food Agric. (2017) 97:5233–40. doi: 10.1002/jsfa.8407
175. Colombo M, Priori D, Trevisi P, Bosi P. Differential gene expression in the oxyntic and pyloric mucosa of the young pig. PLoS ONE. (2014) 9:e111447. doi: 10.1371/journal.pone.0111447
Keywords: thymol, pharmacological effects, toxicological effects, veterinary applications, in vivo, in vitro and in silico studies
Citation: Pérez Gaudio D, Giantin M, Pauletto M and Dacasto M (2025) Pharmaco-toxicological aspects of thymol in veterinary medicine. A systematic review. Front. Vet. Sci. 12:1562641. doi: 10.3389/fvets.2025.1562641
Received: 17 January 2025; Accepted: 30 June 2025;
Published: 12 August 2025.
Edited by:
Narda Gail Robinson, CuraCore Integrative Medicine & Education Center, United StatesReviewed by:
Raquel Díez Laiz, University of León, SpainAna M. Sahagun Prieto, University of León, Spain
Copyright © 2025 Pérez Gaudio, Giantin, Pauletto and Dacasto. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Denisa Pérez Gaudio, ZGVuaXNhQHZldC51bmljZW4uZWR1LmFy