 Tomáš Husák1
Tomáš Husák1 Zuzana Čadková1
Zuzana Čadková1 Ondřej Máca1,2*
Ondřej Máca1,2* Marek Kouba3Zdeňka Klimková1
Marek Kouba3Zdeňka Klimková1 Richard Sehnal1
Richard Sehnal1 Jana Nápravníková1
Jana Nápravníková1 Václava Hrabětová1Ivana Jankovská1
Václava Hrabětová1Ivana Jankovská1 Jaroslav Vadlejch1
Jaroslav Vadlejch1 Iva Langrová1
Iva Langrová1- 1Department of Zoology and Fisheries, Faculty of Agrobiology, Food and Natural Resources, Czech University of Life Sciences Prague, Prague, Czechia
- 2Department of Pathology and Parasitology, State Veterinary Institute Prague, Prague, Czechia
- 3Department of Ethology and Companion Animal Science, Faculty of Agrobiology, Food and Natural Resources, Czech University of Life Sciences Prague, Prague, Czechia
Background: Larval stages of carnivore cestodes in rodents have been extensively studied for decades, primarily based on morphological indicators. Numerous datasets, particularly from Central Europe, exist on this topic. Traditionally, the shape, number, and size of hooks have been key distinguishing features. However, recent findings suggest that morphology alone may not provide accurate identification. In this study, rodent livers from various habitats across five regions of the Czech Republic were examined for the presence of taeniid larval cestodes.
Methods: A total of 2,934 rodent specimens were collected using snap traps between 2014 and 2022. Taeniid metacestodes detected in these intermediate hosts were subsequently subjected to species determination through molecular (targeting the cytochrome oxidase subunit I gene) and morphological examinations.
Results: The following cestodes were identified using molecular methods: Hydatigera kamiyai (found in Apodemus flavicollis, Apodemus sylvaticus, Microtus arvalis, and Microtus agrestis), Taenia martis (in A. flavicollis, M. arvalis, and Myodes glareolus), and Versteria mustelae (in M. arvalis and My. glareolus). Sequencing of 55 PCR-positive isolates revealed 13 haplotypes of H. kamiyai, one of T. martis and four of V. mustelae. Further, we also identified co-infection by more metacestode species or haplotypes. No rodent liver samples tested were positive for Echinococcus multilocularis or Hydatigera taeniaeformis sensu stricto, either through morphological or molecular identification methods. Furthermore, no other cryptic species were detected during this study.
Conclusion: This study presents the first comprehensive molecular data on the aforementioned zoonotic cestode species in the Czech Republic. The findings demonstrate the importance of using molecular techniques for Taeniidae species determination, as morphological methods may lead to erroneous classifications. Furthermore, this research highlights the importance of accurate diagnostic techniques, enabling the development of effective prevention and control strategies by verifying host–parasite relationships.
1 Introduction
The cestode family Taeniidae comprises approximately 50 endoparasitic species across four genera: Echinococcus Rudolphi, 1801; Hydatigera Lamarck, 1816; Taenia Linnaeus, 1758; and Versteria Gmelin, 1790. Members of this parasite group have an indirect life cycle, with herbivores and insectivores primarily serving as intermediate hosts (IH), while carnivores act as definitive hosts (DH). Metacestodes, the larval stages of tapeworms, can infect a wide range of host species of veterinary and/or public health importance, potentially resulting in significant economic losses (1, 2).
Rodents (Rodentia) frequently serve as IH for taeniid cestodes (1–3). As the most abundant and successful group of mammals (Mammal Diversity Database), rodents possess a wide capacity to spread various pathogens, making them ideal indicators for epidemiological studies. Numerous researchers have investigated the occurrence of cestode larval stages in rodents. However, most of these studies have relied solely on morphological identification (4–9), while investigations employing molecular-level determinations remain limited (3, 10–19).
In this context, rodents can serve as important indicators of the prevalence of dangerous tapeworms affecting humans. It should be emphasized that while many cestode genera contain zoonotic species, Echinococcus multilocularis is particularly noteworthy as it causes one of the deadliest parasitic diseases in humans, proving fatal if left untreated (20). In Europe, dozens of cases are diagnosed annually, with nine cases reported in the Czech Republic alone in 2023 (National Reference Centre for Epidemiological Data Analysis, State Institute of Health). The prevalence of E. multilocularis in rodents varies according to numerous factors, and the roles of IH have been studied across various European countries (21). However, this topic still requires attention due to limited data, especially from the Czech Republic. Moreover, other tapeworms involving rodents in their life cycles, including Taenia or Versteria species, can also pose risks to humans (22).
Despite the medical relevance of rodent-borne helminths, knowledge of the extraintestinal helminth fauna in rodents from the Czech Republic remains limited. Previous epidemiological studies have documented several cestode species (with mammalian DHs) in Czech rodents, including: Taenia pisiformis, T. crassiceps, T. polyacantha, T. martis, Hydatigera taeniaeformis sensu lato, Versteria mustelae, and Mesocestoides lineatus (4, 6, 23–27).
To our knowledge, comprehensive data on the prevalence and molecular identification of taeniid metacestodes in Czech rodents, including genetic variation determination, are entirely lacking. The recent reclassification of H. taeniaeformis s.l. into H. taeniaeformis sensu stricto, H. kamiyai, and Hydatigera sp. have highlighted the need for more focused research on these species worldwide (11). Particularly, there is a need to verify the occurrence of H. taeniaeformis s.s. and H. kamiyai strobilocerci in the Central European region and identify their haplotypes.
In the present study, we sought to identify metacestodes in rodents captured between 2014 and 2022. It was aimed to provide proper identification supplemented by genetic variation descriptions, addressing the current lack of data from the studied region.
2 Materials and methods
2.1 Sample locations
The study was conducted in five regions of the Czech Republic: Sokolov area, Mostecko area, Krušné hory area, Příbram area, and Křižanovská vrchovina area. The Sokolov area is a reclaimed post-coal mining landscape, featuring mowed meadows, pastures (clover-grass mixtures), fields, and occasional forests (various coniferous and deciduous tree species) and wetlands. Samples were collected between 2014 and 2022. The Mostecko area is also a reclaimed post-coal mining landscape, including grasslands, pastures, and fields. Sampling was obtained from 2017 to 2020. In the Příbram area, sites were in the vicinity of ore mines, comprising grasslands and meadows. Samples were collected between 2014 and 2016. The Krušné hory area is predominantly forested, featuring blue spruce, larch, and occasional rowan. Sampling took place in 2019. The Křižanovská vrchovina area is also predominantly forested and dominated by spruce and beech. Samples were collected in 2022 (Figure 1).

Figure 1. Map of localities investigated. Forest covers are represented in green and mountains are presented in brown.
2.2 Sample collection
A total of 2,934 rodent specimens were collected using snap traps across all study sites. Five, 25 and 70 localities were repeatedly monitored in Příbram, Mostecko and Sokolov areas, respectively. Based on the characteristics of each trapping plot, traps were established in 2 lines at a distance of 25 m between the lines with 5 m intervals between traps or in quadrates with 5 m grid. The number of traps per site ranged from 25 to 81 according to the habitat size monitored. Eight localities were monitored during 2019 in the Krušné hory area. Animals were sampled on 8 quadrate grids (10 × 10 traps, span 10 m) and along 10 trap-lines (50 trapping points, span 5 m). The quadrate grids and trap lines were situated in deforested habitats with dense growth of the reed-grass and a new plantation of tree species. Six localities were monitored during spring 2022 in the Křižanovská vrchovina area. The trapping plots were in three habitat types: the Norway spruce forest, the mixed forest with a dominance of European beech, and permanent grassland (meadow), two plots each. The 40 traps were set up with 10 m spacing (4 × 10 traps) at each quadrate/site.
Small terrestrial rodents were trapped using simple snap traps baited with cotton candlewicks soaked with mixture of plant oil, flour and rendered bacon. The trapped specimens were processed according to standard mammalogical techniques comprising species, sex, age determination, and basic morphological measurements (body weight, tail length, hind foot, and ear).
All experimental procedures were conducted in compliance with Czech legislation (section 29 of Act No. 246/1992 Coll. on the protection of animals against cruelty, as amended by Act No. 77/2004 Coll.).
2.3 Parasitological examination
Each captured individual was subjected to necropsy, with a particular focus on the liver. In addition to macroscopic examination, livers were held over and palpated to search for parasitic lesions within the liver parenchyma. Any visible cysts, spots, lesions, or other abnormal formations on the livers were isolated for further analysis. The initial selection of worms was based on the characteristic scolex morphology of Taeniidae. Fully developed cestode larval stages were morphologically identified to species level based on cyst shape, size, and location in the host as well as on the size, number, and shape of rostellar hooks (3, 7, 11). In either case, all worms were analyzed individually. Cysts and any visible spots or lesions were preserved in 70% ethanol until further examination. Small pieces of the host’s infected tissue (cysts/lesions) were used for molecular identification.
2.4 Molecular analysis
Individual specimens (fixed in 70% (v/v) ethanol or thawed material) were transferred to Eppendorf tubes for DNA extraction. Genomic DNA (n = 89) was extracted using the NucleoSpin tissue XS kit (Macherey-Nagel, Düren, Germany), following the manufacturer’s instructions. DNA was stored at −20°C until use in polymerase chain reaction (PCR) assays targeting the mitochondrial cox1 gene, selected for characterization of species and haplotypes. PCR amplifications were performed in reaction mixtures consisting of 12.5 μL of GoTaq® G2 Hot Start Green Master Mix (Promega, Madison, WI, United States), 0.4 μM of each primer, and 5 μL DNA template. PCRs were carried out using primers JB3 (forward): 5′-TTTTTTGGGCATCCTGAGGTTTAT-3′ and JB4.5 (reverse): 5′-TAAAGAAAGAACATAATGAAAATG-3′ (28), to amplify a part of the gene under the following conditions: 1 cycle of 3 min at 95°C as an initial hot start step, followed by 36 cycles of 30 s at 94°C, 45 s at 55°C, 30 s at 72°C, and a final extension step at 72°C for 10 min. A negative control using RNase/DNase-free water was included in each reaction. PCR products were assessed by gel electrophoresis on a 1% (w/v) agarose gel and purified using the ExoSAP-IT™ Express PCR Product Cleanup Reagent Kit (Thermo Fisher Scientific), as per the manufacturer’s protocol. Purified amplicons were sequenced by Eurofins Genomics (Ebersberg, Germany) using forward and reverse primers. Nucleotide sequences derived in this study have been deposited in GenBank (accession numbers: PQ868574; PQ868575; PQ868896; PQ868998; PQ869002; PQ869008; PQ869162; PQ869198; PQ869199; PQ869203; PQ869225; PQ869280; PQ869284; PQ869303; PQ869304; PQ870821; PQ870820; PQ870818; PQ870819; PQ870824; PQ870826; PQ870817; PQ870825; PQ870822; PQ870823; PQ870827). Sequences were bidirectionally manually edited using FinchTV software (Geospiza Inc., Seattle, WA, United States) and compared using the NCBI Basic Local Alignment Search Tool (BLASTn)1. Alignment of sequences was performed using an online version of MAFFT Alignment v7 (29). Phylogenetic relationships of obtained nucleotide sequences were analyzed using the MEGA11 software (30). Phylogenetic relationships were reconstructed using the maximum likelihood (ML) method, based on the Hasegawa-Kishino-Yano parameter model (31) with a gamma distribution rate and a proportion of invariant sites (HKY + G + I), bootstrapped at 1,000 replicates. Sequences of Echinococcus granulosus sensu lato (GenBank accession number: MH300987) and E. multilocularis (GenBank accession number: OR911432) were used as outgroups.
3 Results
3.1 Morphological assessment
A total of 2,934 individuals representing 9 rodent species were captured and examined by necropsy. Cysts, visible spots, or lesions were isolated from the livers of 89 rodents. Among these abnormalities, 55 were confirmed as cestode infections through DNA sequencing and, in some cases, by morphological features. Only the cysts or lesions that were molecularly identified to the species level were used for morphological description of the larvae. These cysts or macroscopically visible spots or lesions were found in only five out of nine examined rodent species (listed in Table 1).
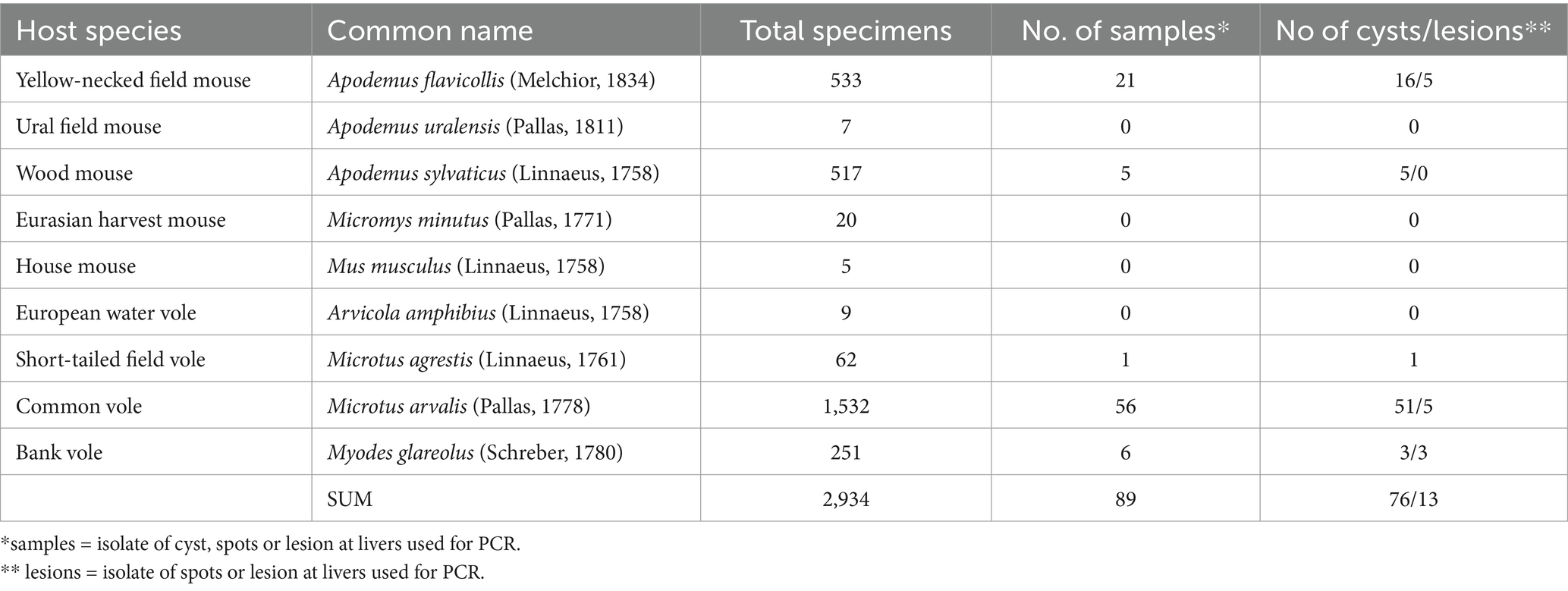
Table 1. List of rodent specimens obtained using snap traps and examined during the study period 2014–2022.
Larval cestodes were found in only 5 species: A. flavicollis, A. sylvaticus, M. arvalis, M. agrestis, and My. glareolus. No metacestodes were detected in A. uralensis, A. amphibius, Micromys minutus, or Mus musculus (Mu. m. domesticus and Mu. m. musculus). Among 55 cestodes the following species were identified: Hydatigera kamiyai, Versteria mustelae and Taenia martis. The prevalence of individual rodent species is given in Table 2; the occurrence of metacestode species in individual areas is shown in Table 3.
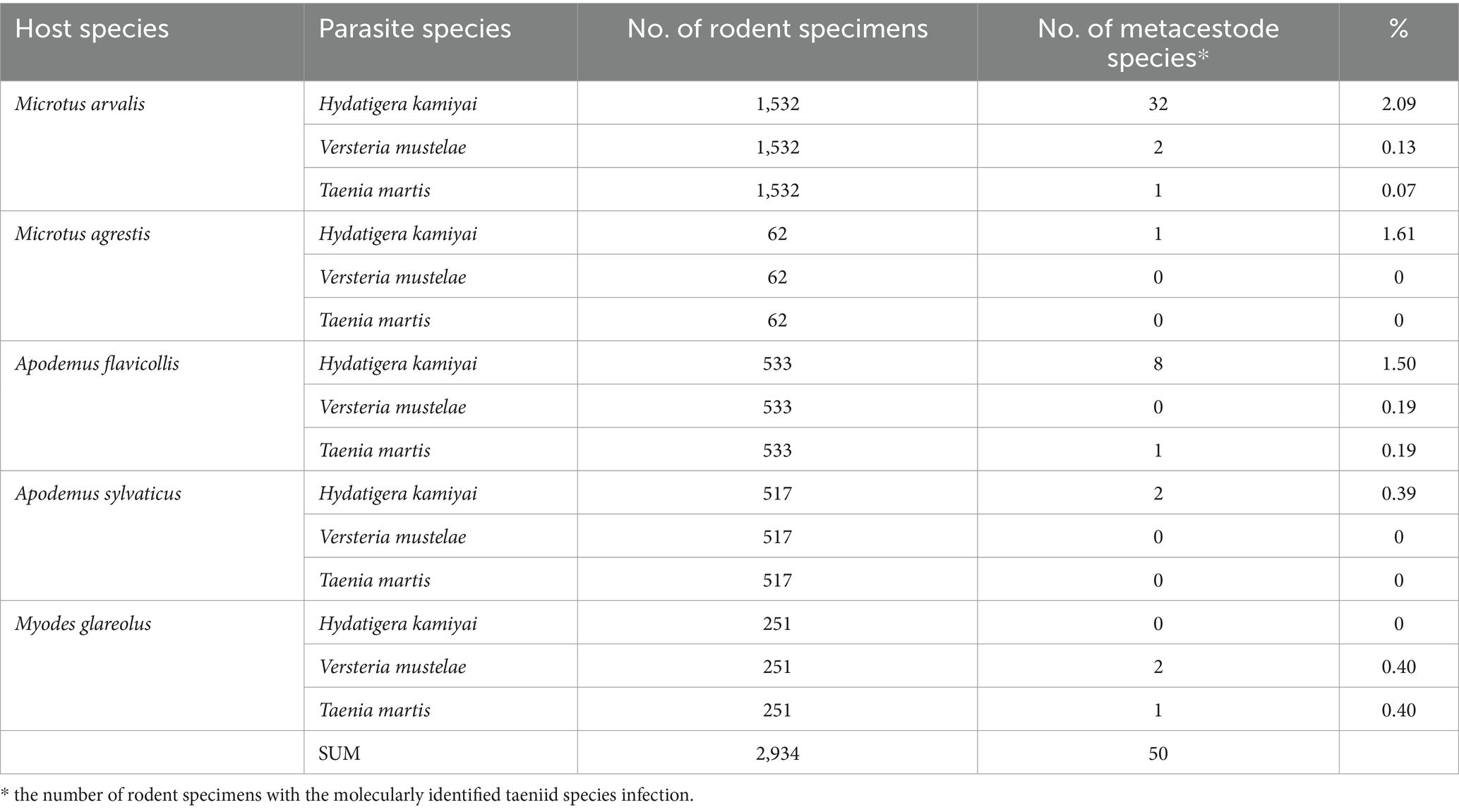
Table 2. The prevalence of the infection (%) of metacestodes in rodent species from five areas of the Czech Republic.
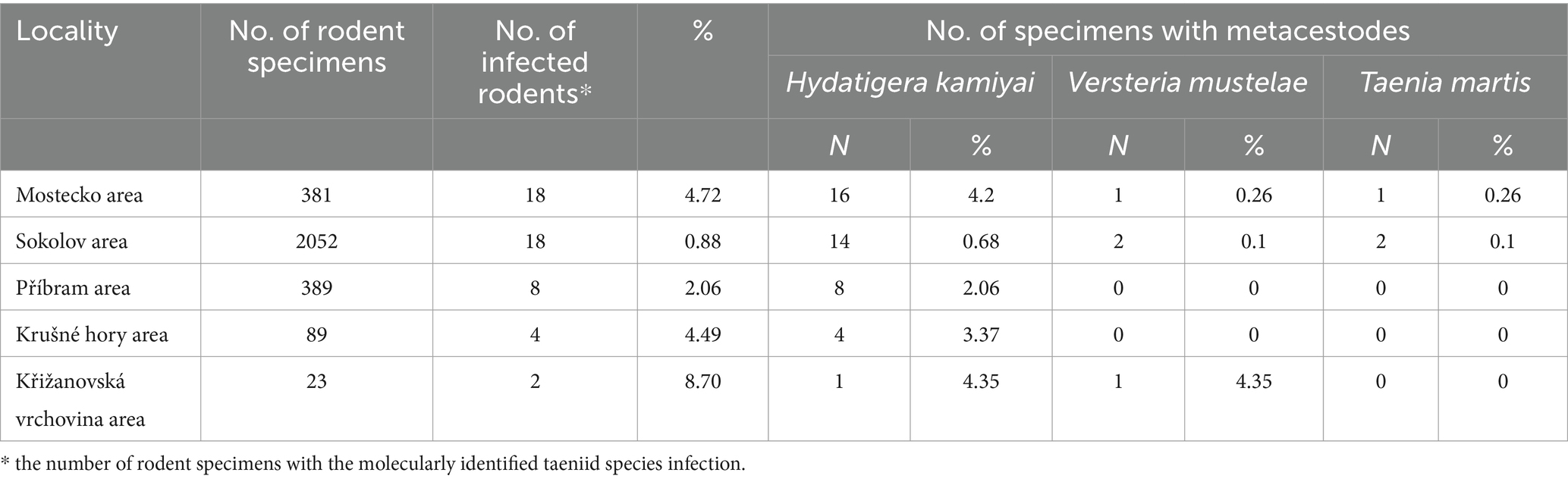
Table 3. The prevalence of the infection (%) of taeniid species in rodents from five areas of the Czech Republic.
Six animals were infected by more than one developmental stage of the cestode parasite, including co-infections involving morphologically distinct parasite species in two cases. Microtus arvalis exhibited the highest number of cysts per host, with 2–4 cysts or strobilocerci found per individual (in Sokolov and Mostecko areas). The second most prevalent species was A. flavicollis, with up to three cysts/strobilocerci per animal (in Krušné hory and Mostecko areas). Lesions and cysts morphologically resembling any metacestodes were subjected to molecular identification.
Hydatigera kamiyai Iwaki, 2016 (larval stages, Figures 2, 3) was identified in 43 rodents from the Sokolov, Mostecko, Příbram, and Krušné hory areas. The hosts included A. flavicollis, A. sylvaticus, M. arvalis, and M. agrestis, with the liver serving as the primary predilection site. The infection intensity ranged from 1 to 4 cysts. Morphological examination revealed ovular or globular cysts, whitish to yellow, measuring 3.24–15.14 mm on the liver. These cysts contained white pseudosegmented strobilocerci with terminal bladders, with individual strobilocerci varying in length from 6.27–7.02 mm. Scolices with hooks were observed only in cysts with a diameter of at least 0.5 mm. The scolex diameter ranged from 1.1–1.3 mm (without suckers), with an average of 1.2 mm, and featured 4 suckers measuring 0.331–0.497 mm in diameter. The prominent suckers ranged in height from 60 to 198 μm. Two hook crowns were on the scolex, with a crown diameter of 0.792–1.016 mm. The total hook count varied from 26 to 36, comprising 11–18 large hooks and 14–18 small hooks. Large hooks measured 352–466 μm, while small hooks ranged from 245 to 268 μm. Detailed hook measurements are provided in Table 4.
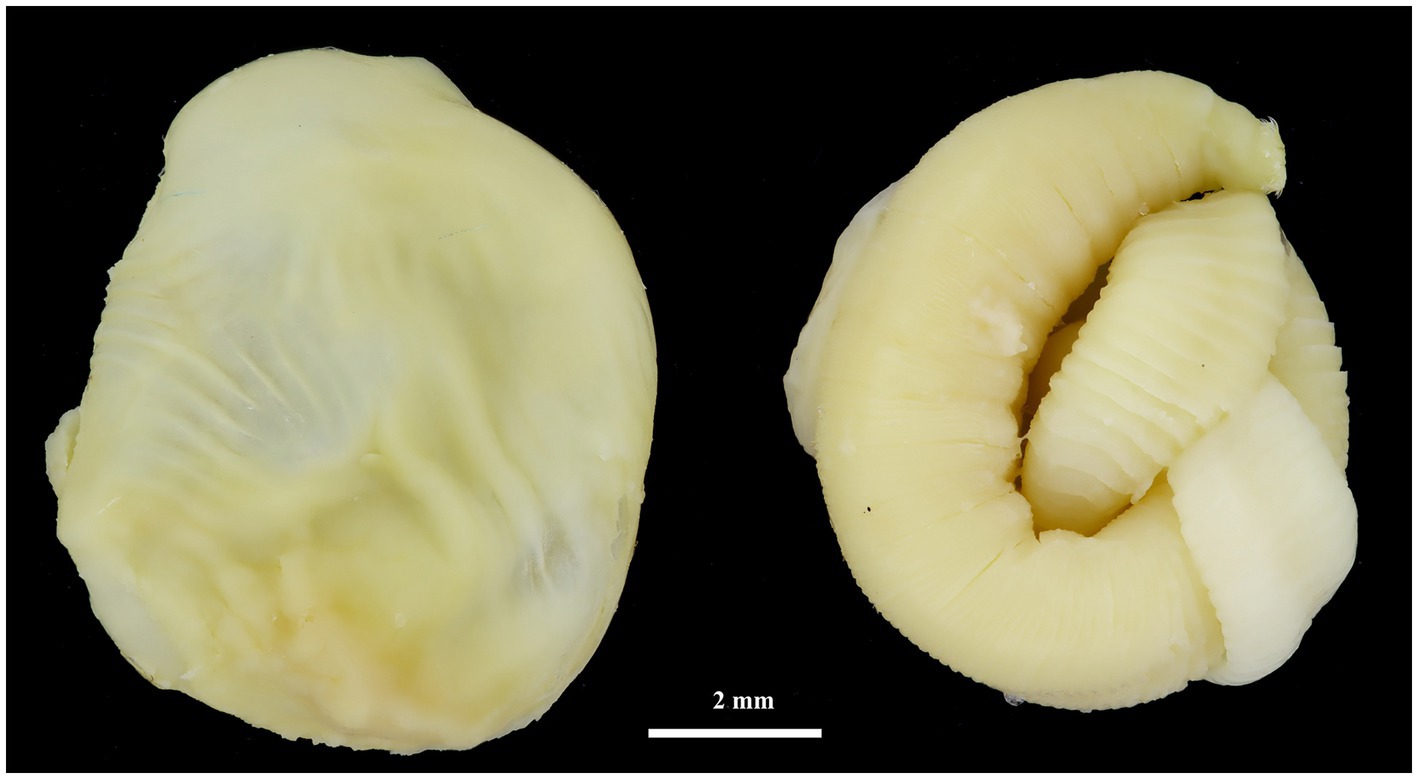
Figure 2. Cyst (left) and strobilocercus (right) of H. kamiyai removed from the liver of M. arvalis (Mostecko area).
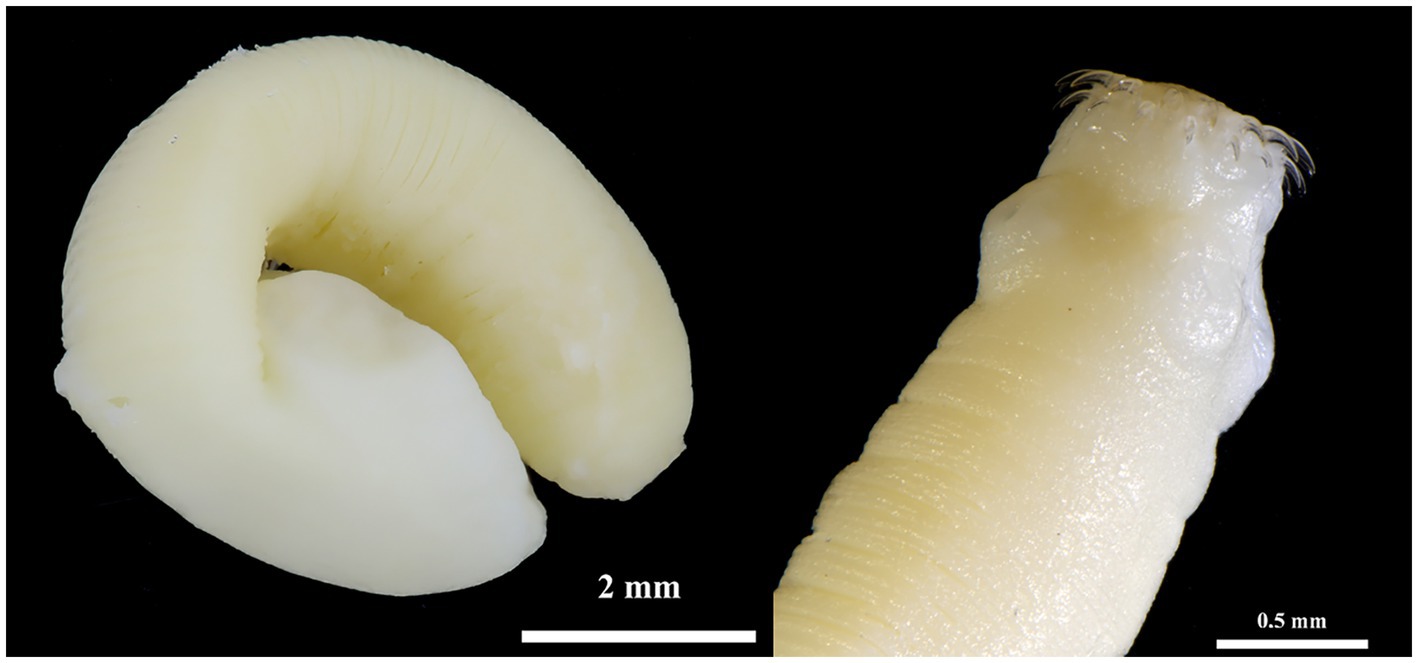
Figure 3. Hydatigera kamiyai. Strobilocercus (left) and detail of the scolex of another individual (right) obtained from two specimens of M. arvalis (Příbram area).
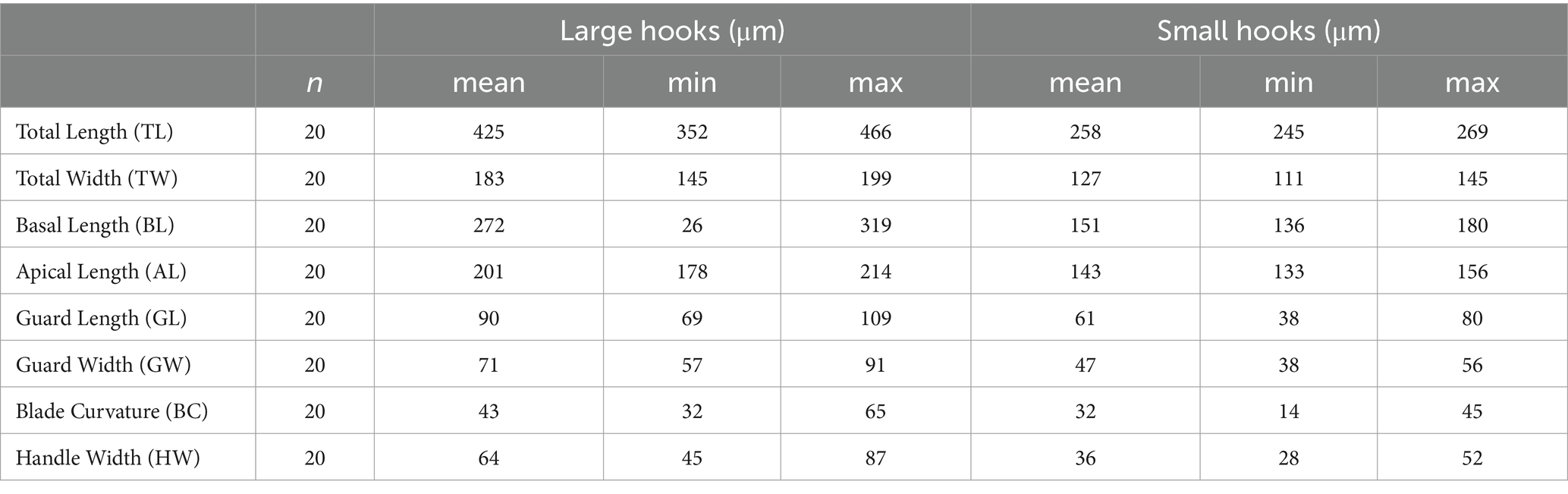
Table 4. Various large and small hook dimensions of metacestodes of H. kamiyai.
Taenia martis (Zeder, 1803) larval stages (Figure 4) were also identified in three rodents in the Sokolov and Mostecko areas. The hosts included A. flavicollis, M. arvalis, and My. glareolus, with the larvae found on the liver. The infection intensity ranged from 1 to 3 larval stages per host. The pseudosegmented metacestodes measured 2.9–10.1 mm in length and 2–4.2 mm in width, with some specimens exhibiting a slender tail-like structure at the posterior end. Despite careful examination, no scolices or hooks were observed in these larval stages.
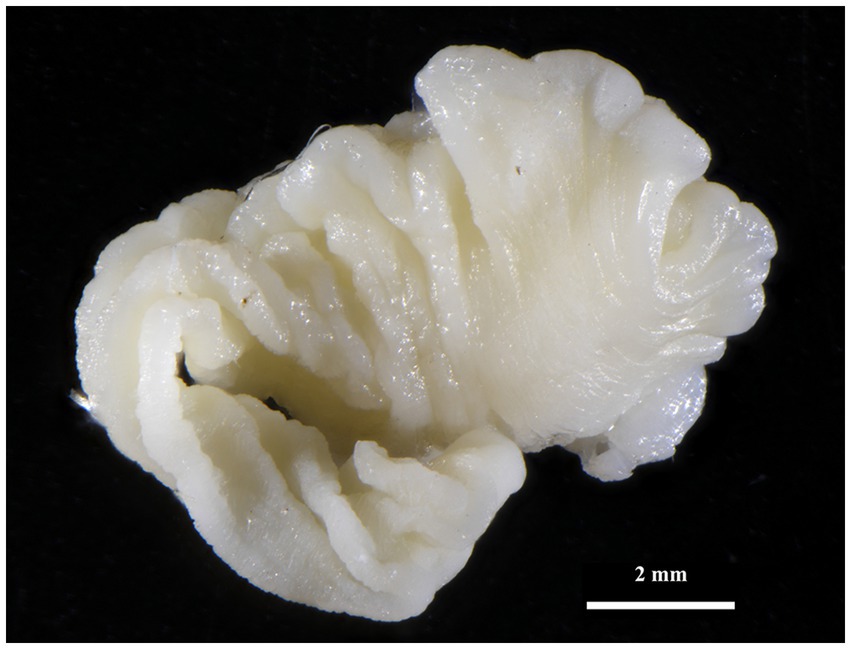
Figure 4. Strobilocercus of T. martis removed from liver of My. glareolus (Sokolov area).
Versteria mustelae (Gmelin, 1970) (syn. Taenia tenuicollis Rudolphi, 1919) larval stages were identified in the Sokolov, Mostecko, and Křižanovská vrchovina areas. The hosts included M. arvalis (two animals) and My. glareolus (two animals), with the liver serving as the primary site of infection. The infection intensity was consistently one metacestode per host. It’s worth noting that Nakao et al. (2) proposed the creation of a new genus, Versteria, for Taenia mustelae. In all cases, only spots and lesions measuring 1 to 2 mm were observed in the liver. No scolices or hooks were found during the examination of these lesions; the identification of this species was carried out using molecular methods.
3.2 Molecular identification
In this work, we detected the following cestodes using molecular methods: H. kamiyai, T. martis, and V. mustelae. Among the 89 DNA isolates from cysts, spots, and lesions, 55 sequences were successfully amplified for the partial cox1 gene. These comprised 13 haplotypes of H. kamiyai, one of T. martis, and four of V. mustelae (Table 5). No PCR product was obtained from the remaining isolates.
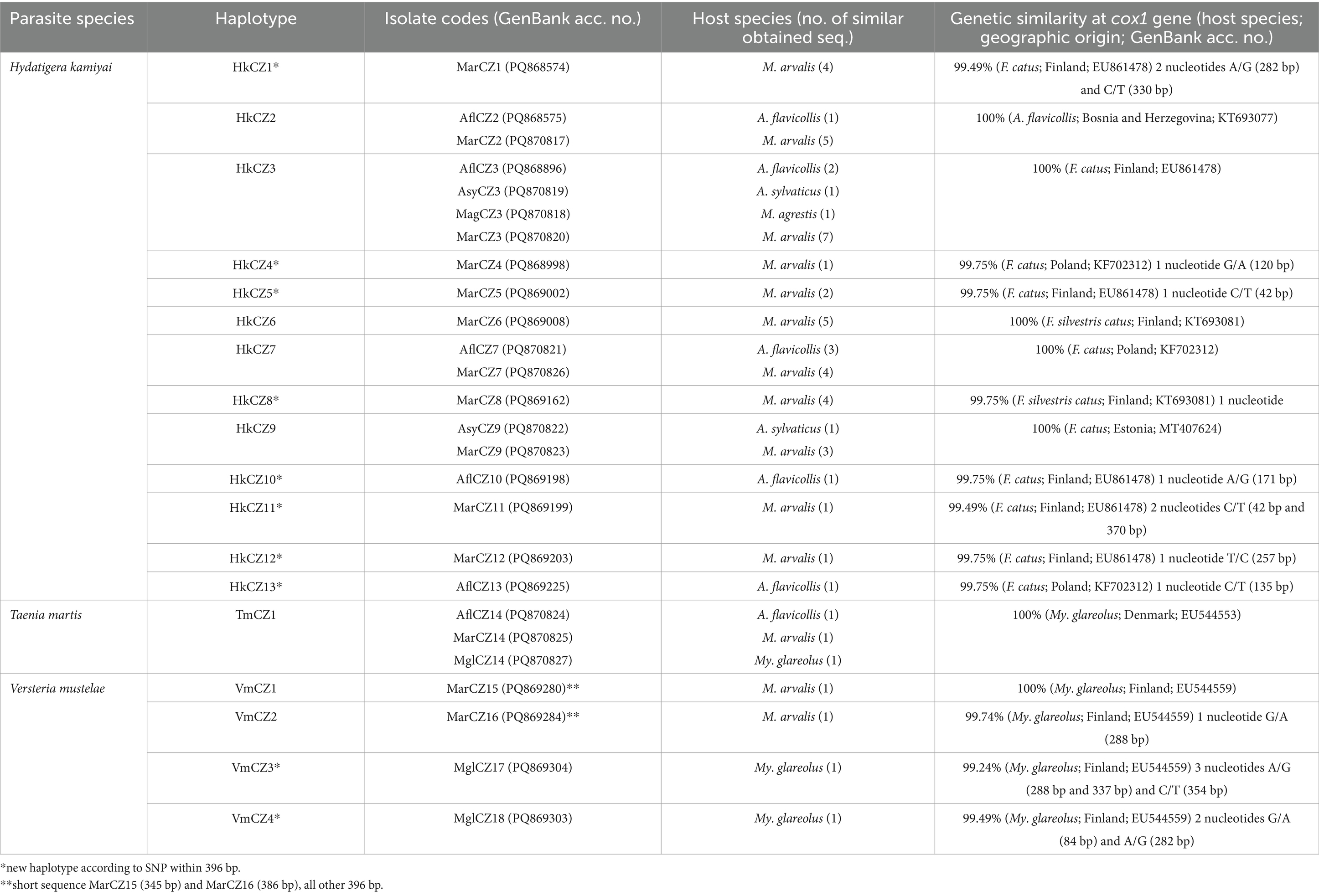
Table 5. Metacestodes characterization with haplotype identification supplemented with the degree of similarity among data with other sequences retrieved from GenBank.
Hydatigera kamiyai was the most frequent finding, with 48 sequences obtained from 4 host species comprising 13 haplotypes (HkCZ1-HkCZ13). These haplotypes differed by single nucleotide polymorphisms (SNPs). Five haplotypes were 100% identical to published cox1 sequences of H. kamiyai described from small mammal hosts, for example, A. flavicollis from Bosnia and Herzegovina (GenBank no. KT693077) or Felis catus from Estonia, Finland, and Poland (GenBank nos: MT407624, EU861478, KF702312) (see Table 5 for details). The remaining sequenced isolates differed from each other by 1–2 SNPs (99.75–99.49% identity). The phylogenetic tree based on cox1 for representatives of H. kamiyai (Figure 5) showed that sequences obtained in the present study are grouped in a single clade with the same species from other hosts and countries. The subclades within this single clade are formed by species sharing the same haplotypes (e.g., sequences PQ868575, PQ870817 and KT693077 shared haplotype) or new haplotypes.

Figure 5. Maximum likelihood tree based on cox1 gene for representatives of H. kamiyai and two related species of Hydatigera from various hosts and countries. Sequences obtained in the present study are shown in blue. Sequences of E. granulosus s.l. and E. multilocularis were used as outgroups.
Five animals were positive for multiple H. kamiyai strobilocerci. In two A. flavicollis and three M. arvalis, different haplotypes were identified within the same host. Co-infection of one M. arvalis by two cestode species, H. kamiyai and T. martis, was identified in one case. Notably, H. taeniaeformis s.s. and no other cryptic species were found in this study.
The T. martis metacestode was molecularly determined for the first time in rodents from Central Europe. We detected metacestodes of T. martis using cox1 gene PCR in M. arvalis, My. glareolus, and A. flavicollis. All three isolate sequences (TmCZ1 haplotype) were 100% identical to My. glareolus from Denmark (GenBank no.: EU544553). Resulted phylogenetic tree based on cox1 for representatives of T. martis (Figure 6) showed that sequences obtained in the present study are grouped in a clade with the same species from other hosts and countries.
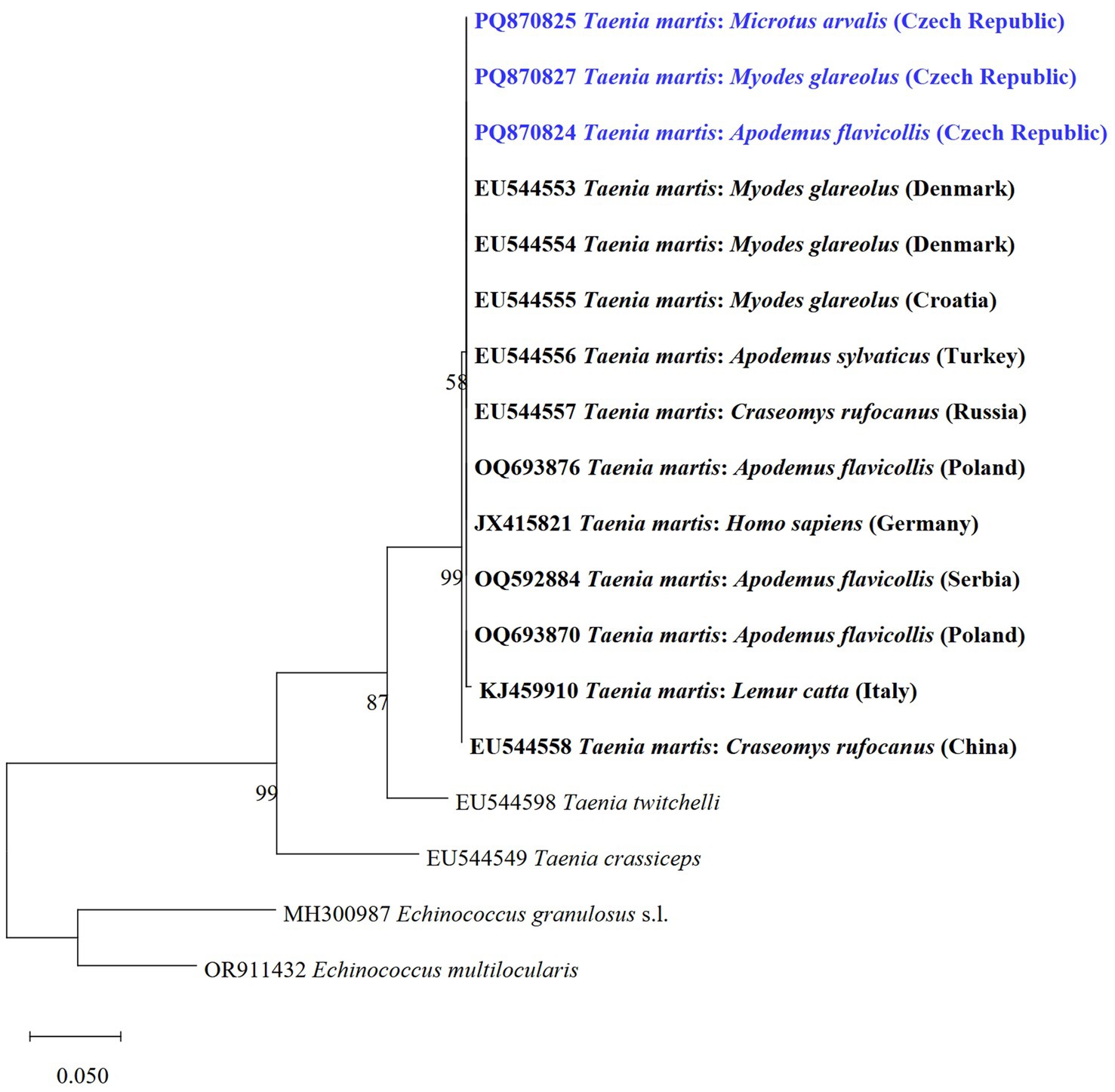
Figure 6. Phylogenetic tree of the cox1 gene of T. martis and two related species of Taenia from various host species and countries using the maximum likelihood method. Sequences of E. granulosus s.l. and E. multilocularis were used as outgroups. Sequences obtained in this study are marked in blue.
We also identified DNA of V. mustelae metacestodes (VmCZ1-VmCZ4) in four voles solely through molecular analyses, detecting them twice in M. arvalis and twice in My. glareolus. This included two newly identified haplotypes. Our sequences showed 99.24–100% similarity to a haplotype from Finland (GenBank no. EU544559). This represents the first molecular finding of V. mustelae in Central Europe. The phylogenetic tree analysis based on cox1 revealed that our isolates grouped with V. mustelae isolates (Figure 7). All other lesions or cysts isolated for molecular determination were negative for E. multilocularis.
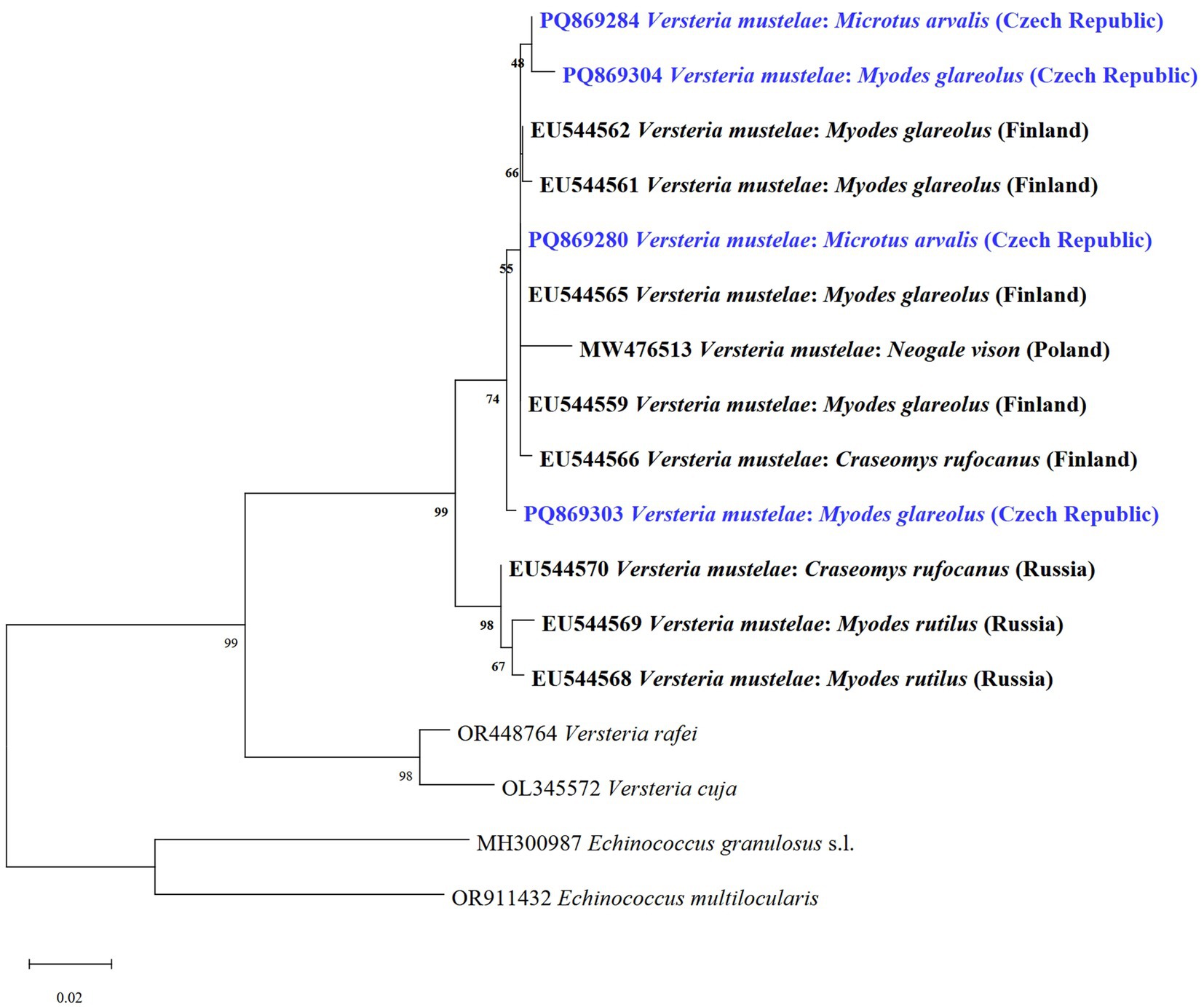
Figure 7. Phylogenetic tree of V. mustelae and two related species of Versteria from various hosts and countries based on of the cox1 gene using the maximum likelihood method. Sequences of E. granulosus s.l. and E. multilocularis were used as outgroups. Sequences obtained in this study are marked in blue.
4 Discussion
Due to a lack of data concerning the current prevalence, species diversity, and genetic variability of rodent-borne metacestodes in Central Europe, the present study aimed to address these gaps. Previous studies on the presence of taeniid metacestodes lacked molecular information, making this study the first to provide valuable data on the molecular identity of some species. All species reported herein were previously known (4, 7, 26, 27, 32), although all species (H. kamiyai, T. martis and V. mustelae) represent new geographical records in Central Europe.
We found a maximum of four metacestodes in a single host. All co-infections of one individual by one or more larvae were identified individually for the first time, highlighting the need for further studies on molecular diversity from individual stages in whole animals, similar to research on other tapeworm species such as Echinococcus granulosus s.s. (33). Multiple parasite stages in one animal are commonly recorded; for example, T. martis has been found in multiple infections. Murai (32) reported five cysts per animal, Schmidt (9) eight cysts, Prokopič and Mahnert (26) up to 12 cysts, and Frank and Zeyhkle (34) even detected 43 cysts of T. martis in one common muskrat (Ondatra zibethicus). Similarly, V. mustelae is often detected as a multi-infection, most often up to 10 cysts per animal (9, 35). Prokopič and Mahnert (26) found 18 cysts per animal, while Murai (32) found up to 20. Unfortunately, none of these previous results were supported by molecular analyses and characterization of all metacestodes, as was done in the present study.
Our study also includes a morphological description of H. kamiyai metacestodes, which were determined using the cox1 gene. Hydatigera kamiyai and H. taeniaeformis s.s. differ molecularly, though less distinctly, and most features overlap (11). However, these authors showed that both congeners differed significantly in seven large and small hooks characters. Unlike molecular analyses, no other morphological studies of these species have been conducted to date, unlike molecular analyses (18).
The present study revealed a slight difference in the morphology of the large hooks of H. kamiyai metacestodes compared to those described by Lavikainen et al. (11), which measured 421–461 μm, while our samples ranged from 352 to 466 μm. This variability in hook length may be related to the amount of material or the age of the larvae.
Finally, we found that the hook numbers differed for large and small hooks on most tapeworm larvae specimens measured. This validates molecular methods as important tools that greatly facilitate taxonomic identification of organisms globally. However, the results of measuring the metacestodes of H. kamiyai in this study indicate that this tapeworm has considerable morphological variability, and molecular identification will be necessary for all future studies.
Additional studies are needed, particularly to elucidate whether H. kamiyai is also common in cats and other potential domestic or wildlife DH species present in the Czech Republic, as observed in other countries (see Table 1 in reference 11). Apart from Lavikainen et al. (11) H. kamiyai has been recorded in the following studies: by Bajer et al. (16) in M. arvalis and My. glareolus (Poland), by Martini et al. (17) in O. zibethicus (Luxembourg), by Miljeviš et al. (18) in A. flavicollis, A. agrarius, M. arvalis and Crocidrua leucodon (Serbia). Outside of Europe, it has been molecularly determined recently only from the Tibetan Plateau from Neodon fuscus (19). None of these reports contain morphological characteristics. This implies that the haplotype occurrence and diversity remain unknown, and a study on DHs is necessary to obtain data specific to the Czech Republic. More samples, particularly from murine rodents or DHs, are required to reliably determine the possible occurrence of H. taeniaeformis s.s. or Hydatigera sp. in Central Europe.
Studies on intra-species variations and differences in the distribution of H. kamiyai are very limited. Its diversity across various IHs in our samples and elsewhere (18) is an interesting topic for future research, similar to studies on E. multilocularis (36). The present data contain newly described haplotypes alongside those typical for Europe, adding value to other studies focused on haplotype network analysis or predominant variant determination. In future studies, these data could be used for DH identifications. Previously, 22 haplotypes (BB1-B22; 396 bp) were described by Lavikainen et al. (11), with additional haplotypes later reported by other authors (18), collectively supporting a high level of genetic diversity worldwide. The next species we found in the examined rodents was T. martis, one of the most widely distributed helminths of Mustelidae occurring throughout the Palearctic region in the Martes genus. In recent years, metacestodes have been frequently found in various primates and humans (37–39), indicating considerable zoonotic potential for this tapeworm.
The importance of T. martis is underscored by the fact that martens are common predators in urban and suburban areas (40, 41). Thus, monitoring the occurrence of this tapeworm would be appropriate and reasonable. Its metacestodes have been found in immunocompetent individuals worldwide, including recent cases in Europe. Examples include brain infections reported by Steinsiepe et al. (42) in Switzerland and Eggink et al. (39) in the Netherlands; eye infections documented by Eberwein et al. (43), Koch et al. (44), and Tappe et al. (45) in Germany; and an infection in the pouch of Douglas reported by Mueller et al. (37), also in Germany. Most of these patients lived in rural villages, were involved in agriculture, and reported frequent marten sightings around their homes.
Metacestodes of T. martis are found in Arvicolinae, Murinae, Cricetinae, Sciurinae, and also in Myocastor coypus, Castor fiber, and Sorex araneus (5, 46–48). The current study is based on molecular identification, and many studies are available for comparison with our molecular data. Taenia martis has been further identified by Al-Sabi and Kapel in Denmark (49); Umhang et al. (47) in Myocastor coypus in France; Krücken et al. (50) in My. glareolus, A. agrarius, and A. flavicollis in Germany; Meyer et al. (51) in pigs in Switzerland; Martini et al. (17) in O. zibethicus in Luxembourg; and Miljevič et al. (18) in A. flavicollis, A. sylvaticus, and My. glareolus in Serbia. Additionally, Reinhardt et al. (52) used PCR sequencing to identify T. martis adult tapeworms in raccoons (Procyon lotor) from Germany.
Versteria mustelae, the third recorded tapeworm species in our study, was molecularly determined for the first time in Central Europe. Versteria mustelae is widespread throughout the world; the DHs are mainly mustelids, whereas IHs include many species of Arvicolinae, Murinae, Cricetinae, Sciurinae, as well as Myocastor coypus, Ochotona alpina, Eospalax baileyi, Talpa europaea, S. araneus, and Crocidura russula (5–7, 22, 26, 53–55). Members of the genus Versteria have been detected in primates (56) and humans (57). It is commonly reported using morphometric characteristics, especially in voles, and was recently found in My. glareolus (58–60), M. agrestis (61), and A. amphibius (62). An experimental study showed limited development of V. mustelae in laboratory mice (54). Our four identifications of V. mustelae were based solely on sequencing results, thus adding valuable data for further studies/findings. Only several records worldwide have used molecular methods, such as V. mustelae determined by Al-Sabi and Kapel in Denmark (49); Al-Sabi et al. (10) in My. glareolus in Denmark; Umhang et al. (47) in O. zibethicus and Myocastor coypus in France; Miller et al. (13) in M. agrestis and A. terrestris in Sweden; Martini et al. (17) in O. zibethicus in Luxembourg; and Zhao et al. (63) in E. baileyi on the Qinghai-Tibet Plateau.
We further sought to verify the occurrence of E. multilocularis in rodents, a widespread species among the main DH, red foxes (Vulpes vulpes), in the Czech Republic. The pooled prevalence in foxes has long been high, reaching around 25% (e.g., 25.32%; 254 positive/1003 examined red foxes in 2023; Máca, unpublished data), which is considerably higher compared to other European countries (21). However, we found no E. multilocularis infection among cysts/lesions on the rodents’ livers, similar to findings from Serbia (18). In fact, E. multilocularis has only been found once in rodents in the Czech Republic, specifically in My. glareolus (1 positive/36 examined) based on histological examination, but without finding protoscoleces (64). Thus, further studies are needed to fully elucidate the role of rodents in the Czech Republic regarding E. multilocularis and its distribution in specific areas.
To conclude, we used molecular biology-based methods to show the occurrence of three tapeworms with zoonotic potential in five common rodent species of Central Europe. Microtus arvalis was the most frequently trapped rodent species and the most commonly infected with larval cestodes. The metacestodes of H. kamiyai were also morphologically described and discussed based on molecularly characterized individuals. New findings with described European mitochondrial haplotypes enrich the present study, offering the potential for new research focusing on transmission pathways or variant distribution in broader geographical contexts. Our new data on T. martis and V. mustelae represent the only recent records from the Czech Republic supplemented by molecular characterization. These findings can aid in epidemiological studies and advance our understanding of infection transmission pathways in cases of human infections. Future research should focus on urban areas and, regions with high E. multilocularis prevalence to evaluate the contribution of rodent hosts in spreading this causative agent.
Data availability statement
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found in the article/supplementary material.
Ethics statement
The animal study was approved by all experimental procedures were conducted in compliance with Czech legislation (section 29 of Act No. 246/1992 Coll. on the protection of animals against cruelty, as amended by Act No. 77/2004 Coll.). The study was conducted in accordance with the local legislation and institutional requirements.
Author contributions
TH: Investigation, Writing – review & editing. ZČ: Investigation, Methodology, Writing – review & editing. OM: Conceptualization, Data curation, Formal analysis, Funding acquisition, Investigation, Methodology, Supervision, Validation, Writing – original draft, Writing – review & editing. MK: Investigation, Methodology, Writing – review & editing. ZK: Investigation, Methodology, Writing – review & editing. RS: Data curation, Formal analysis, Investigation, Methodology, Visualization, Writing – review & editing. JN: Investigation, Methodology, Visualization, Writing – review & editing. VH: Data curation, Formal analysis, Investigation, Writing – review & editing. IJ: Investigation, Methodology, Writing – review & editing. JV: Investigation, Methodology, Writing – review & editing. IL: Conceptualization, Data curation, Formal analysis, Funding acquisition, Investigation, Methodology, Project administration, Supervision, Validation, Visualization, Writing – original draft, Writing – review & editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This study was supported by the Czech Science Foundation (Grant No. 23-07811S). Open access funding was provided by the Faculty of Agrobiology, Food and Natural Resources, Czech University of Life Sciences Prague.
Acknowledgments
The authors thank I. Křičková from the Parasitology laboratory (SVI Prague) for technical support.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The authors declare that no Gen AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Footnotes
References
1. Deplazes, P, Eckert, J, Mathis, A, Samson-Himmelstjerna, GV, and Zahner, H. Parasitology in veterinary medicine. Wageningen: Wageningen Academic Publishers (2016).
2. Nakao, M, Lavikainen, A, Iwaki, T, Haukisalmi, V, Konyaev, S, Oku, Y, et al. Molecular phylogeny of the genus Taenia (Cestoda: Taeniidae): proposals for the resurrection of Hydatigera Lamarck, 1816 and the creation of a new genus Versteria. Int J Parasitol. (2013) 43:427–37. doi: 10.1016/j.ijpara.2012.11.014
3. Loos-Frank, B. An up-date of Verster's (1969) "taxonomic revision of the genus Taenia Linnaeus" (Cestoda) in table format. Syst Parasitol. (2000) 45:155–84. doi: 10.1023/A:1006219625792
5. Abuladze, KI. Taeniata of animals and man and diseases caused by them. Moskva: Akademiia Nauk SSSR, Gel'mintologicheskaia Laboratoriia, Izdatel'stvo Nauka (1964). 554 p.
6. Prokopič, J. Biocenotical study on cestodes of small mammals in various biotopes. Acta Sc Nat Brno. (1972) 6:1–68.
8. Pétavy, AF, Tenora, F, and Deblock, S. Contributions to knowledge on the helminths parasitizing several Arvicolidae (Rodentia) in Auvergne (France). Helminthologia. (1996) 33:51–8.
9. Schmidt, S. Untersuchungen zum Vorkommen von Capillaria hepatica und Metazestoden der Cyclophyllida bei Wildmäusen in Deutschland. Leipzig: der Universität Leipzig (2001).
10. Al-Sabi, MNS, Jensen, PM, Christensen, MU, and Kapel, CMO. Morphological and molecular analyses of larval taeniid species in small mammals from contrasting habitats in Denmark. J Helminthol. (2013) 89:112–7. doi: 10.1017/S0022149X13000680
11. Lavikainen, A, Iwaki, T, Haukisalmi, V, Konyaev, SV, Casiraghi, M, Dokuchaev, NE, et al. Reappraisal of Hydatigera taeniaeformis (Batsch, 1786) (Cestoda: Taeniidae) sensu lato with description of Hydatigera kamiyai n. sp. Int J Parasitol. (2016) 46:361–74. doi: 10.1016/j.ijpara.2016.01.009
12. Miller, AL, Olsson, GE, Walburg, MR, Sollenberg, S, Skarin, M, Ley, C, et al. First identification of Echinococcus multilocularis in rodent intermediate hosts in Sweden. Int J Parasitol Parasites Wildl. (2016) 5:56–63. doi: 10.1016/j.ijppaw.2016.03.001
13. Miller, AL, Olsson, GE, Sollenberg, S, Walburg, MR, Skarin, M, and Höglund, J. Transmission ecology of taeniid larval cestodes in rodents in Sweden, a low endemic area for Echinococcus multilocularis. Parasitology. (2017) 144:1041–51. doi: 10.1017/S0031182017000257
14. Catalano, S, Bâ, K, Diouf, ND, Léger, E, Verocai, GG, and Webster, JP. Rodents of Senegal and their role as intermediate hosts of Hydatigera spp. (Cestoda: Taeniidae). Parasitology. (2018) 146:299–304. doi: 10.1017/S0031182018001427
15. Panti-May, JA, Hernández-Betancourt, SF, and García-Prieto, L. Hydatigera taeniaeformis (Cestoda: Taeniidae) in the Yucatán squirrel Sciurus yucatanensis (Rodentia: Sciuridae), Mexico. Therya. (2019) 10:179–82. doi: 10.12933/therya-19-740
16. Bajer, A, Alsarraf, M, Dwużnik, D, Mierzejewska, EJ, Kołodziej-Sobocińska, M, Behnke-Borowczyk, J, et al. Rodents as intermediate hosts of cestode parasites of mammalian carnivores and birds of prey in Poland, with the first data on the life-cycle of Mesocestoides melesi. Parasit Vectors. (2020) 13:95. doi: 10.1186/s13071-020-3961-2
17. Martini, M, Dumendiak, S, Gagliardo, A, Ragazzini, F, La Rosa, L, Giunchi, D, et al. Echinococcus multilocularis and other taeniid metacestodes of muskrats in Luxembourg: prevalence, risk factors, parasite reproduction, and genetic diversity. Pathogens. (2022) 11:1414. doi: 10.3390/pathogens11121414
18. Miljević, M, Rajičić, M, Umhang, G, Bajić, B, Čabrilo, OB, Budinski, I, et al. Cryptic species Hydatigera kamiyai and other taeniid metacestodes in the populations of small mammals in Serbia. Parasit Vectors. (2023) 16:250. doi: 10.1186/s13071-023-05879-x
19. Zhou, G, Zhang, H, Chen, W, Li, Z, Zhang, X, and Fu, Y. Morphological observation, molecular identification and evolutionary analysis of Hydatigera kamiyai found in Neodon fuscus from the Qinghai-Tibetan plateau. Infect Genet Evol. (2024) 123:105629. doi: 10.1016/j.meegid.2024.105629
20. Torgerson, PR, Schweiger, A, Deplazes, P, Pohar, M, Reichen, J, Ammann, RW, et al. Alveolar echinococcosis: from a deadly disease to a well-controlled infection. Relative survival and economic analysis in Switzerland over the last 35 years. J Hepatol. (2008) 49:72–7. doi: 10.1016/j.jhep.2008.03.023
21. Oksanen, A, Siles-Lucas, M, Karamon, J, Possenti, A, Conraths, FJ, Romig, T, et al. The geographical distribution and prevalence of Echinococcus multilocularis in animals in the European Union and adjacent countries: a systematic review and meta-analysis. Parasit Vectors. (2016) 9:519. doi: 10.1186/s13071-016-1746-4
22. Deplazes, P, Eichenberger, RM, and Grimm, F. Wildlife-transmitted Taenia and Versteria cysticercosis and coenurosis in humans and other primates. Int J Parasitol Parasites Wildl. (2019) 9:342–58. doi: 10.1016/j.ijppaw.2019.03.013
25. Prokopič, J. The ecological division of micromammalian cestodes. Helminthologia. (1970) 11:207–12.
26. Prokopič, J, and Mahnert, V. Über Helminthen der Kleinsäuger (Insectivora, Rodentia) Tirols (Österreichs). Ber Nat-Med Ver Innsbruck. (1970) 58:143–54.
27. Tenora, F, and Staněk, M. Changes of the helminthfauna in several Muridae and Arvicolidae at Lednice in Moravia. I. Systematics and taxonomy. Acta Univ Agri. (1994) 42:237–47.
28. Bowles, J, Blair, D, and McManus, DP. Genetic variants within the genus Echinococcus identified by mitochondrial DNA sequencing. Mol Biochem Parasitol. (1992) 54:165–73. doi: 10.1016/0166-6851(92)90109-W
29. Katoh, K, Rozewicki, J, and Yamada, KD. MAFFT online service: multiple sequence alignment, interactive sequence choice and visualization. Brief Bioinform. (2019) 20:1160–6. doi: 10.1093/bib/bbx108
30. Tamura, K, Stecher, G, and Kumar, S. MEGA11: molecular evolutionary genetics analysis version 11. Mol Biol Evol. (2021) 38:3022–7. doi: 10.1093/molbev/msab120
31. Hasegawa, M, Kishino, H, and Yano, T. Dating the human-ape split by a molecular clock of mitochondrial DNA. J Mol Evol. (1985) 22:160–74. doi: 10.1007/BF02101694
32. Murai, É. Taeniid species in Hungary (Cestoda: Taeniidae). II. Larval stages of taeniids parasitizing rodents and lagomorphs. Miscnea Zool Hung. (1982) 1:27–44.
33. Hidalgo, C, Stoore, C, Pereira, I, Paredes, R, and Alvarez Rojas, CA. Multiple haplotypes of Echinococcus granulosus sensu stricto in single naturally infected intermediate hosts. Parasitol Res. (2020) 119:763–70. doi: 10.1007/s00436-019-06578-2
34. Frank, B, and Zeyhle, E. Echinokokken – und andere Bandwurmlarven im Bisam (Ondatra zibethicus). Nachr Deut Pflanzenschutz. (1981) 33:166–70.
35. Memaran, M. Die Helminthenfauna der Waldmaus (Apodemus sylvaticus L.), der Gelbhalsmaus (Apodemus flavicollis Melchior) und der Rötelmaus (Clethrionomys glareolus Schreber) im Naturpark Hoher Vogelsberg. Gießen: der Justus Liebig-Universität Gießen (1970).
36. Guo, B, Cairen, WJ, Wang, W, Wang, B, Ren, Y, et al. The A2 haplotype of Echinococcus multilocularis is the predominant variant infecting humans and dogs in Yili prefecture, Xinjiang. Infect Genet Evol. (2024) 119:105581. doi: 10.1016/j.meegid.2024.105581
37. Mueller, A, Förch, G, Zustin, J, Muntau, B, Schuldt, G, and Tappe, D. Case report: molecular identification of larval Taenia martis infection in the pouch of Douglas. Am J Trop Med Hyg. (2020) 103:2315–7. doi: 10.4269/ajtmh.20-0782
38. Peters, M, Mormann, S, Gies, N, and Rentería-Solís, Z. Taenia martis in a white-headed lemur (Eulemur albifrons) from a zoological park in North Rhine-Westphalia, Germany. Vet Parasitol Reg Stud Rep. (2023) 44:100913. doi: 10.1016/j.vprsr.2023.100913
39. Eggink, H, Maas, M, van den Brand, JMA, Dekker, J, Franssen, F, Hoving, EW, et al. Taenia martis neurocysticercosis-like lesion in child, associated with local source, the Netherlands. Emerg Infect Dis. (2024) 30:555–9. doi: 10.3201/eid3003.231402
40. Krauze-Gryz, D, Jackowiak, M, Klich, D, Gryz, J, and Jasińska, KD. Following urban predators – long-term snow-tracking data reveal changes in their abundance and habitat use. J Zool. (2024) 323:213–24. doi: 10.1111/jzo.13170
41. Capon, M, Lysaniuk, B, Godard, V, Clauzel, C, and Simon, L. Characterising the landscape compositions of urban wildlife encounters: the case of the stone marten (Martes foina), the red fox (Vulpes vulpes), and the hedgehog (Erinaceus europaeus) in the greater Paris area. Urban Ecosyst. (2021) 24:885–903. doi: 10.1007/s11252-020-01071-6
42. Steinsiepe, VK, Ruf, MT, Rossi, M, Fricker-Feer, C, Kolenc, D, Suter Buser, B, et al. Human Taenia martis neurocysticercosis, Switzerland. Emerg Infect Dis. (2023) 29:2569–72. doi: 10.3201/eid2912.230697
43. Eberwein, P, Haeupler, A, Kuepper, F, Wagner, D, Kern, WV, Muntau, B, et al. Human infection with marten tapeworm (Taenia martis). Emerg Infect Dis. (2013) 19:1152–4. doi: 10.3201/eid1907.121114
44. Koch, T, Schoen, C, Muntau, B, Addo, M, Ostertag, H, Wiechens, B, et al. Molecular diagnosis of human Taenia martis eye infection. Am J Trop Med Hyg. (2016) 94:1055–7. doi: 10.4269/ajtmh.15-0881
45. Tappe, D, Berkholz, J, Mahlke, U, Lobeck, H, Nagel, T, Haeupler, A, et al. Molecular identification of zoonotic tissue-invasive tapeworm larvae other than Taenia solium in suspected human cysticercosis cases. J Clin Microbiol. (2016) 54:172–4. doi: 10.1128/JCM.02171-15
46. Yushkov, VY. Helminth fauna of mammals (insectivores, predators, lagomorphs, and rodents) of the Komi ASSR. Tr Gel’mintologicheskoi Lab. (1971) 22:232–48.
47. Umhang, G, Demerson, JM, Legras, L, Boucher, JM, Peytavin de Garam, C, Bastid, V, et al. Rodent control programmes can integrate Echinococcus multilocularis surveillance by facilitating parasite genotyping: the case of Arvicola terrestris voles screening in France. Parasitol Res. (2021) 120:1903–8. doi: 10.1007/s00436-021-07126-7
48. Campbell-Palmer, R, Del Pozo, J, Gottstein, B, Girling, S, Cracknell, J, Schwab, G, et al. Echinococcus multilocularis detection in live Eurasian beavers (Castor fiber) using a combination of laparoscopy and abdominal ultrasound under field conditions. PLoS One. (2015) 10:e0130842. doi: 10.1371/journal.pone.0130842
49. Al-Sabi, MNS, and Kapel, CMO. Multiplex PCR identification of Taenia spp. in rodents and carnivores. Parasitol Res. (2011) 109:1293–8. doi: 10.1007/s00436-011-2373-9
50. Krücken, J, Blümke, J, Maaz, D, Demeler, J, Ramünke, S, Antolová, D, et al. Small rodents as paratenic or intermediate hosts of carnivore parasites in Berlin, Germany. PLoS One. (2017) 12:e0172829. doi: 10.1371/journal.pone.0172829
51. Meyer, A, Olias, P, Schüpbach, G, Henzi, M, Barmettler, T, Hentrich, B, et al. Combined cross-sectional and case-control study on Echinococcus multilocularis infection in pigs in Switzerland. Vet Parasitol. (2020) 277:100031. doi: 10.1016/j.vpoa.2020.100031
52. Reinhardt, NP, Wassermann, M, Härle, J, Romig, T, Kurzrock, L, Arnold, J, et al. Helminths in invasive raccoons (Procyon lotor) from Southwest Germany. Pathogens. (2023) 12:919. doi: 10.3390/pathogens12070919
53. Prochaska, S. Oekologische beziehungen im wirt – parasiten – system der wirtspopulation von Apodemus sylvaticus L. und der parasitenpopulationen der darmhelminthen Syphacia stroma (von Linstow, 1844) und Nematospiroides dubius (Baylis, 1926). Bern: der Universität Bern (1984).
54. Iwaki, T, Abe, N, Shibahara, T, Oku, Y, and Kamiya, M. Developmental study of Taenia mustelae in the intermediate and definitive hosts, with a note on the life cycle of T. mustelae in Hokkaido, Japan. J Parasitol. (1996) 82:840–2. doi: 10.2307/3283903
55. Tinnin, DS, Ganzorig, S, and Gardner, SL. Helminths of small mammals (Erinaceomorpha, Soricomorpha, Chiroptera, Rodentia, and Lagomorpha) of Mongolia. Lubbock: Museum of Texas Tech University (2011). 56 p.
56. Goldberg, TL, Gendron-Fitzpatrick, A, Deering, KM, Wallace, RS, Clyde, VL, Lauck, M, et al. Fatal metacestode infection in Bornean orangutan caused by unknown Versteria species. Emerg Infect Dis. (2014) 20:109–13. doi: 10.3201/eid2001.131191
57. Barkati, S, Gottstein, B, Müller, N, Sheitoyan-Pesant, C, Metrakos, P, Chen, T, et al. First human case of metacestode infection caused by Versteria sp. in a kidney transplant recipient. Clin Infect Dis. (2019) 68:680–3. doi: 10.1093/cid/ciy602
58. Behnke, JM, Bajer, A, Harris, PD, Newington, L, Pidgeon, E, Rowlands, G, et al. Temporal and between-site variation in helminth communities of bank voles (Myodes glareolus) from N.E. Poland. 1. Regional fauna and component community levels. Parasitology. (2008) 135:985–97. doi: 10.1017/S0031182008004393
59. Pétavy, AF, Tenora, F, and Deblock, S. Co-occurrence of metacestodes of Echinococcus multilocularis and Taenia taeniaeformis (Cestoda) in Arvicola terrestris (Rodentia) in France. Folia Parasitol. (2003) 50:157–8. doi: 10.14411/fp.2003.028
60. Le Pasteur, MH, Giraudoux, P, Delattre, P, Damange, JP, and Quéré, JP. Spatiotemporal distribution of four species of cestodes in a landscape of mid-altitude mountains (Jura, France). Ann Parasitol Hum Comp. (1992) 67:155–60. doi: 10.1051/parasite/1992675155
61. Soveri, T, Henttonen, H, Rudbäck, E, Schildt, R, Tanskanen, R, Husu-Kallio, J, et al. Disease patterns in field and bank vole populations during a cyclic decline in Central Finland. Comp Immunol Microbiol Infect Dis. (2000) 23:73–89. doi: 10.1016/S0147-9571(99)00057-0
62. Chechulin, AI, Zav’yalov, EL, Gerlinskaya, LA, Krivopalov, AV, Makarikov, AA, and Muzyka, VY. Helminthes of the water vole Arvicola terrestris Linnaeus of forest-bog biocenoses in the north of the Baraba lowland. Contemp Probl Ecol. (2010) 3:4–9. doi: 10.1134/S1995425510010029
63. Zhao, F, Ma, JY, Cai, HX, Su, JP, Hou, ZB, Zhang, TZ, et al. Molecular identification of Taenia mustelae cysts in subterranean rodent plateau zokors (Eospalax baileyi). Zool Res. (2014) 35:313–8. doi: 10.13918/j.issn.2095-8137.2014.4.313
Keywords: Hydatigera, Taenia, Versteria, haplotype, liver, molecular epidemiology, small mammals, Czech Republic
Citation: Husák T, Čadková Z, Máca O, Kouba M, Klimková Z, Sehnal R, Nápravníková J, Hrabětová V, Jankovská I, Vadlejch J and Langrová I (2025) Molecular identification of zoonotic taeniids metacestodes in several rodent species trapped in Central Europe. Front. Vet. Sci. 12:1571082. doi: 10.3389/fvets.2025.1571082
Edited by:
Simona Gabrielli, Sapienza University of Rome, ItalyReviewed by:
Ali Tumay Gurler, Ondokuz Mayıs University, TürkiyeJeba Jesudoss Chelladurai, Auburn University College of Veterinary Medicine, United States
Copyright © 2025 Husák, Čadková, Máca, Kouba, Klimková, Sehnal, Nápravníková, Hrabětová, Jankovská, Vadlejch and Langrová. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Ondřej Máca, bWFjYW9AYWYuY3p1LmN6