 Sylvia Cheptoo1,2*
Sylvia Cheptoo1,2* Erhan Yalcindag3
Erhan Yalcindag3 Lina González Gordon3
Lina González Gordon3 Benson Rukwaro2,4
Benson Rukwaro2,4 Joseph Samuel Kimatu2,5
Joseph Samuel Kimatu2,5 Joseph Wasonga2,6
Joseph Wasonga2,6 Benedict E. Karani2
Benedict E. Karani2 Gideon Ndambuki2
Gideon Ndambuki2 Susan Migeni2
Susan Migeni2 Jesse Kagai7
Jesse Kagai7 Linus Eric Kiprotich7Nelson Saya7
Linus Eric Kiprotich7Nelson Saya7 Deepali Vasoya8
Deepali Vasoya8 Getrude Nangekhe2
Getrude Nangekhe2 Justus Onguso1
Justus Onguso1 Grace Mungai1
Grace Mungai1 Barend Mark Bronsvoort3
Barend Mark Bronsvoort3 Elizabeth Anne Jessie Cook2*
Elizabeth Anne Jessie Cook2*- 1Jomo Kenyatta University of Agriculture and Technology (JKUAT), Nairobi, Kenya
- 2Centre for Tropical Livestock Genetics and Health (CTLGH), International Livestock Research Institute (ILRI), Nairobi, Kenya
- 3Centre for Tropical Livestock Genetics and Health (CTLGH), Roslin Institute, University of Edinburgh, Easter Bush Campus, Edinburgh, United Kingdom
- 4Department of Veterinary Pathology, Microbiology and Parasitology, The University of Nairobi, Nairobi, Kenya
- 5Department of Public Health, Pharmacology and Toxicology, The University of Nairobi, Nairobi, Kenya
- 6Department of Community Health, Amref International University (AMIU), Nairobi, Kenya
- 7Mazingira Centre, International Livestock Research Institute (ILRI), Nairobi, Kenya
- 8UK Dementia Research Institute at University of Edinburgh, Edinburgh, United Kingdom
Gastrointestinal nematodes (GIN) are of major concern in dairy farming, particularly in smallholder systems, because of their impact on the health of the calves and later on their productivity. These infections often occur as co-infections, which can complicate their prevention and treatment. The aim of this study was to conduct fecal egg counts (FEC), genetically identify GIN species, assess species diversity, and identify associated risk factors for GIN infections in dairy calves. Fecal samples were collected from 532 dairy calves across 289 small holder dairy farms. Species identification was achieved through deep amplicon sequencing of the Internal Transcribed Spacer-2 rDNA locus (ITS-2) of first-stage larvae (L1). The mean eggs per gram (EPG) was 62.0 ± 93.0. Most of the calves 64.2% had low-intensity infections (<50 EPG), 28.6% had medium-intensity infections (50–200 EPG), and 7.2% had high-intensity infections (>200 EPG). Next Generation Sequencing analysis identified nine GIN species, with Cooperia punctata (27.8%), Haemonchus placei (26.3%), and Haemonchus contortus (23.6%) being the most prevalent. Co-infections were common, accounting for 69.5% of all infections, with two (40.1%), three (26.9%), and four-species combinations (19.8%) predominating. Male calves showed a significant association with both increased FEC and smaller heart girth, while FEC decreased with age. H. placei and C. punctata were associated with increased FEC, whereas Ostertagia ostertagi (14.5%) and Trichostrongylus colubriformis (8.0%) were associated with decreased heart girth. Calves managed under pasture systems had higher odds of co-infection. This study reveals that GIN infections are highly prevalent in dairy calves, with co-infections being common, and that GIN burden is significantly influenced by calf age, sex, and management system. The Nemabiome tool offers a promising approach to assessing GIN burden and guiding the selection of anthelmintic protocols as part of sustainable farming strategies in tropical regions.
1 Introduction
Gastrointestinal nematodes (GIN) infections are a global concern for livestock, particularly in pasture-based ruminants (1, 2). These infections often result in morbidity and productivity losses (3). First-season grazing calves are predominantly affected (4), showing clinical signs such as diarrhea, stunted growth and weight loss (5). Heavily parasitized calves suffer extensive damage to the intestinal lining and can develop hematological changes including severe anemia and hypoproteinaemia, which can lead to mortality (6). However, in dairy cattle, the most important impact of GIN infections are the subclinical, chronic production loses such as decreased milk production, with an estimated loss of 1.4 liter per cow per day (7), as well as lower milk protein content (8).
GIN exist in complex communities comprising several co-infecting species (9, 10), and co-infection is widespread in cattle globally (11, 12). Co-infections are associated with more severe pathology than single infections and can influence the host immune response by increasing susceptibility to other microparasitic infections, as well as changing the transmission patterns and dynamics of these infections within populations (13–15). Accurate species identification of co-infections is essential for guiding treatment decisions, particularly considering the rapid development of resistance to available anthelminthic drugs (16–18).
Conventional diagnostic methods rely on coproscopy using microscopic examination (19). GIN eggs have similar morphology, making it necessary to culture them to identify the third-stage larvae (L3) based on their distinct morphological features (20). However, some GIN species L3 have overlapping morphological features, making their identification harder (21). This has led to the development of molecular assays to allow accurate identification of GIN species in ruminants (22). Such assays include real-time polymerase chain reaction (qPCR), droplet digital PCR (ddPCR), PCR-linked Restriction Fragment Length Polymorphism (RFLP), multiplex PCR and multiplexed-tandem PCR (23–26). Although these methods can identify species-level infections, they are limited to predetermined species, potentially overlooking novel or non-targeted species (12).
A novel deep amplicon sequencing method, termed “Nemabiome,” was recently developed to examine nematode populations using the ITS-2 rDNA region. The locus of choice reliably differentiates between Clade V nematodes to the species level, most of which infect cattle (10). Within this clade is the Trichostrongyloidea family, including important genera such as Haemonchus, Ostertagia, Cooperia, Trichostrongylus, and Teladorsagia (27, 28). L3 have been commonly used for Nemabiome analysis (10, 12), however first-stage larvae (L1) are becoming an increasingly popular alternative due to improved larval recovery, a more labor-friendly process, and significantly shorter turnaround time of approximately 48 h compared to 21 days for L3 (3, 21).
The Nemabiome technique has been effectively used to identify GIN infections across various species and geographical areas, including beef cattle (12, 29) and heifers in Canada (30), sheep in Sweden and the United Kingdom (21, 31), wildlife in France (32), and horses from Thailand, United States, and Canada (33, 34). However, in Sub-Saharan Africa, studies on GIN infection among dairy calves are scarce, and rely on conventional methods (7, 35–43). To address this gap, this study investigates the occurrence and genetic diversity of GIN species using deep amplicon sequencing and explores the risk factors associated with GIN co-infections among smallholder dairy calves in a key dairy farming region in Kenya.
2 Materials and methods
2.1 Ethics statement
Scientific and ethical approval for the study was obtained from the International Livestock Research Institute (ILRI) Animal Care and Use Committee (ILRI-IACUC2023-10), Institutional Research Ethics Committee (ILRI-IREC2023-44) and The National Commission for Science and Technology (NACOSTI). Written consent forms were signed by farmers willing to take part in the study. Sampling did not pose any long-term risk for the calves and was conducted by qualified veterinary professionals.
2.2 Study setting
The study was conducted in Nandi county, situated in the North Rift region of Kenya within Agroecological Zone II (upper highlands, lower highlands and upper midlands) (44), covering an area of 2855.8 square kilometers, and including neighboring Kesses and Ainabkoi Sub-counties within the Uasin Gishu County (Figure 1). In this area, the alternating dry and wet seasons, with heavy rains in April (~200 mm) and low rainfall between January and February (~50 mm) (44), provide favorable conditions for dairy farming. Significant investments have been made in local dairy genetic resources and toward controlling the transmission of infectious diseases among this population. Locally, dairy farming accounts for 10% of the national dairy herd (309,038 dairy cattle as of the 2019 National Census) (45), generating an estimated 121.5 million kilograms of milk annually, with a market value of approximately 3.6 billion Kenyan shillings (44). The combination of a favorable climate, widespread grazing-based management, and lack of robust diagnostic methods for GIN make this region particularly suitable for studying nematode co-infections in calves.
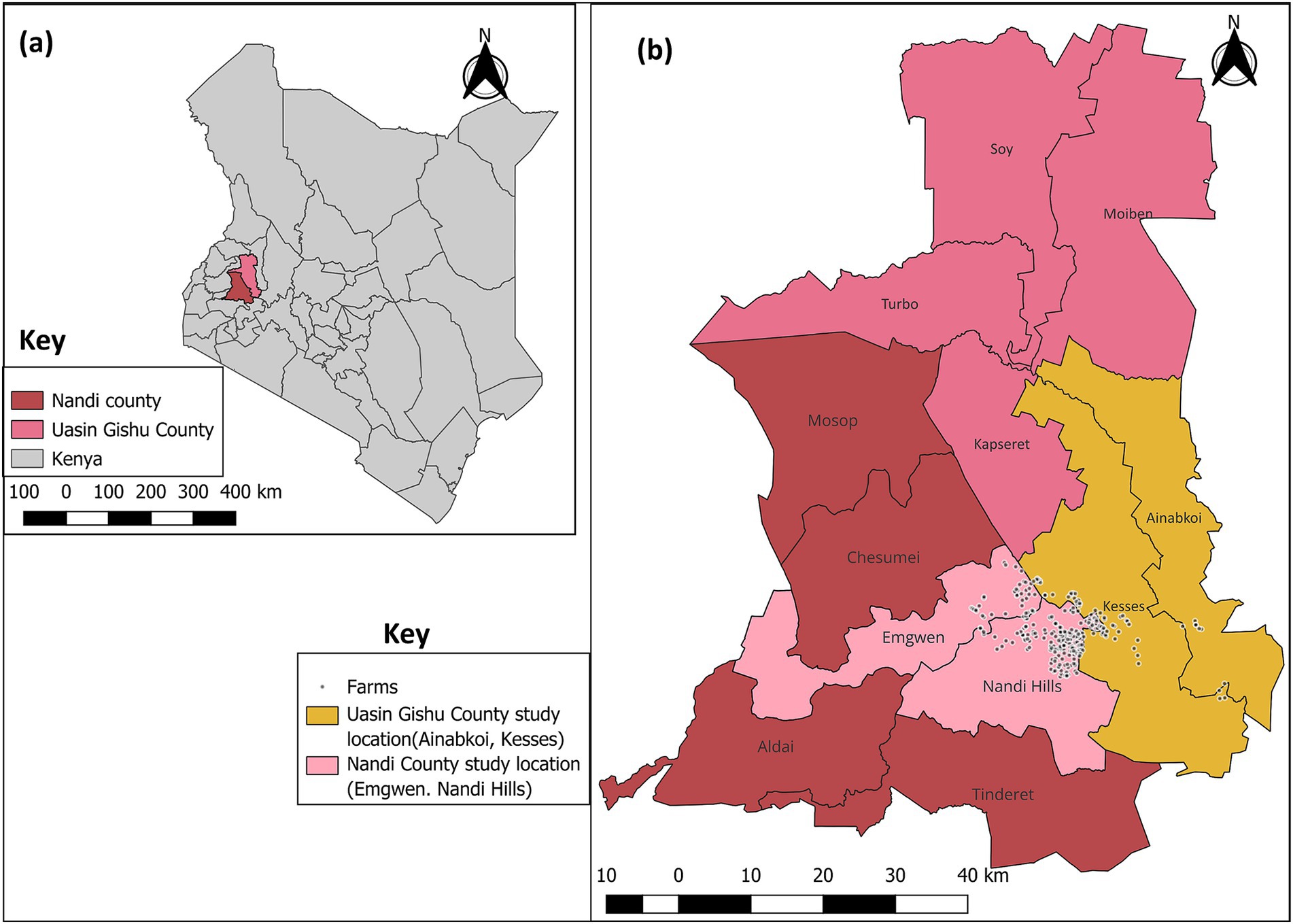
Figure 1. Map of Kenya showing study locations in Nandi and Uasin Gishu County. (a) Nandi and Uasin Gishu County and (b) Sub-counties sampled (Nandi County: Emgwen, Nandi hills. Uasin Gishu County: Ainabkoi and Kesses). Source. Map was drawn using QGIS version 3.36.0 with the shape files downloaded from Humanitarian Data Exchange database (https://data.humdata.org).
2.3 Study design and sampling
This study was designed as a cross-sectional study. The sampling frame, collated from the Africa Dairy Genetic Gain (ADGG) database at ILRI,1 included 448 farms from the Lessos Farmer Cooperative enrolled in the Centre for Tropical Livestock Genetics and Health (CTLGH) abortion cohort, of which 289 farms with eligible dairy calves were visited for sample collection between September and December 2023. The sample size was calculated as described by Audigé (46), which suggested a minimum of 375 animals.
While the definition of “calf” varies across sources, ranging from age-based cut-offs to criteria focused on the timing of weaning, this study defined calves as animals up to 12-month-old. This broader definition, consistent with the latest FAO guidelines for the World Agricultural Census (47), was adopted to account for local variations in weaning practices, calf growth rates, and their overall maturity, often lagging behind international standards in East Africa (48, 49). Across farms, all calves aged 3 to 12 months were sampled, for a total of 532 calves included in the study. This age range was chosen because it corresponds with a period of increased grazing activity, raising the exposure to pastures potentially contaminated with infective larvae (50). Calves younger than 3 months were not sampled as they primarily depend on nursing, have minimal pasture exposure, and benefit from short-term immunity acquired through colostrum (51).
Fecal samples were collected from each calf using a rectal grab approach to avoid environmental contamination. Samples were placed in an airtight glove, labeled with the calf ID, and transported to the Kapsabet Veterinary Laboratory in cool boxes with ice packs to avoid GIN egg hatching.
2.4 Data collection
A structured questionnaire was administered to a consenting adult member of the household (≥18 years) familiar with the herd; all responses were recorded using the Open Data Kit (ODK) system (52). Data collected included several factors at both the animal and farm levels. At the farm level, the type of management system was recorded. Animal-level factors comprised sex, age (estimated in months by the farmer at the time of sampling), heart girth (measured by the animal health assistant), breed (assessed based on phenotype by the animal health assistant), weaning status at the time of sampling, and deworming status within the past 4 weeks, including the type of dewormers used.
2.5 Fecal egg count
Fecal Egg Count (FEC) was performed using the modified McMaster technique as described previously by Christie and Jackson (53). Briefly, 1 g of feces was weighed and placed in a sieve bowl, where 15 mL of water was added and mixed. The resulting solution was transferred to a 15 mL centrifuge tube and spun at 1,000 revolutions per minute (rpm) for 2 min. The supernatant was discarded, and the pellet was resuspended in 12 mL of saturated salt solution and centrifuged at 1,000 rpm for 2 min. The supernatant was then transferred to a cuvette and viewed using a microscope at a 10X magnification lens (53). Samples were done in duplicates and the mean FEC was calculated by averaging the results from both runs. The mean eggs per gram (EPG) was then grouped into low intensity (<50), medium intensity (50–200) and high intensity (>200) (20).
2.6 First stage larvae (L1) coproculture and isolation
Larval coproculture and isolation was performed as explained by Burgess et al. (54). Briefly, larval coproculture was set on a sample of fresh feces from each calf. This process involved the use of stacked filters with pore sizes of 150 μm, 53 μm, and 30 μm. The filtrate collected from the final 30 μm filter was transferred to a 15 mL tube, and a flotation technique was used to recover the eggs. The recovered eggs were rinsed three times with water, and 3 mL of the suspension was then incubated in 6-well cell culture plates for 48 h.
The incubated solution containing L1 larvae was passed through the filter directly into a 6-well plate and incubated for 2 h at room temperature. The solution was then centrifuged, and the resulting pellet was resuspended in 2 mL of water and stored at −20°C until DNA extraction was performed. After DNA extraction, the L1 was stored in 70% Ethanol (Sigma-Aldrich) and frozen at −80°C for long term storage.
2.7 DNA extraction
DNA extraction was performed via the TANBead® nucleic acid extraction system model Maelstrom 9,600 series using the TANBead Nucleic acid Extraction kit (W6T2A46),2 which was performed according to the manufacturer’s recommendations and DNA was eluted in 100 μL of elution buffer.
2.8 PCR and deep amplicon sequencing of the ITS-2 rDNA locus
PCR and deep amplicon sequencing were performed as described by Avramenko et al. (10). It involved two PCR reactions. The first PCR used NC1 and NC2 primers (Supplementary Table S1). L3 stages of Haemonchus contortus and Teladorsagia circumcincta (provided by Professor Neil Sargison, Royal (Dick) School of Veterinary Studies, University of Edinburgh), and 9 GIN samples (Roslin Institute, University of Edinburgh), were used as positive controls. Nuclease free water was subsequently used as a negative control to test for contamination. The PCR products were purified using AMPure XP Magnetic Beads (Beckman Coulter, Inc). The second PCR, with limited cycles, was done to add Illumina barcoding indices and P5/P7 tags combinations to each sample (Supplementary Table S2). An aliquot of 10 μL from the second PCR products from each sample were pooled to make a master sequencing library pool and were purified using AMPure XP Magnetic Beads (Beckman Coulter, Inc) according to manufacturer’s recommendation. Finally, 70 μL of the pooled libraries were submitted to Genewiz Genomics, Germany3 for Illumina MiSeq sequencing. This was done using a 500-cycle paired-end reagent kit (MiSeq Reagent Kits v2, MS-103-2003) with an addition of 10% PhiX Control v3 (Illumina, FC-110-2003) to generate 250 bp paired-end sequencing reads. Each resequencing step followed Illumina’s standard protocol.
2.9 Bioinformatics analysis
The amplicons sequence data was analyzed using the Snakemake pipeline developed for analysis of amplicon sequencing data from MiSeq4 which was adapted from Silwamba et al. (55). The raw sequencing reads were trimmed at Phred score of 28 using Sickle (56). The high-quality paired end reads were overlapped and extended using Flash (57) followed by identification of PCR primers and those with exact match with forward and reverse primers were retained. The sequences with 100% identity were clustered to form unique variants present in the data. The variants were then searched against the custom Nematode database and NCBI nr database using BLAST (58). Subsequently, manual curation of the data was performed to confirm the infection. A read number threshold of 500 reads was used to differentiate between true positives and background noise, with read counts below 500 considered negative.
To validate the assay, 36 negative controls were included, two from the first PCR, two from the indexing PCR, and two from DNA extraction for each plate. Of these, 31 showed no reads confirming clear results. The remaining five exhibited very low read numbers, all of which were below the established threshold of 500 reads attached as a Supplementary Table S3. The raw sequencing data was submitted to the European Nucleotide Archive and is available under the project accession number PRJEB85916.
2.10 Phylogenetic analysis
The DNA sequences for each species were grouped separately, and duplicate sequences appearing at least twice were merged into a single representative sequence using the FaBox DNA Collapser tool.5 Amplicon Sequence Variants (ASV) were aligned using Multiple Sequence Alignment Fast Fourier Transform (MAFFT) with the maximum number of iterations set to 1,000 using the local pairwise alignment algorithm and visualized using SEAVIEW (59, 60). A maximum likelihood phylogenetic tree was then constructed for the ASV of the study samples utilizing IQ-TREE software6 with 1,000 bootstraps. TPM2 model was chosen based on the Bayesian Information Criterion (BIC), with a gamma shape parameter of 0.765. The tree was then visualized using the iTOL (Interactive Tree of Life) online tool (61).
2.11 Statistical analysis
All data analyses were conducted using R software version 4.3.37 in R Studio (Boston, MA8). A descriptive summary of the study population, FEC and heart girth was generated. The survey package (62), was used to estimate the design-adjusted true species prevalence, and all estimates were given with a 95% confidence intervals (CI) with adjustment done at the farm level.
Generalized mixed-effects models using the lme4 package (63) were employed to evaluate risk factors for FEC, heart girth, and co-infections, while accounting for clustering at the farm level. Variables with a p-value ≤ 0.2 in the univariable analysis against each outcome were selected for further multivariable analysis. Age was included as a continuous variable in the models.
Three models were developed (Figure 2): (1) a linear regression model for FEC, to identify associated epidemiological factors. FEC data was log-transformed to meet linear model assumptions. This model included six GIN species identified through deep amplicon sequencing to explore species-specific associations with FEC (Supplementary Table S4); (2) a logistic regression model aimed to identify risk factors for nematode co-infections. Animals with single or no infection were categorized as negative, and those with multiple infections were classified as positive (Supplementary Table S5). Finally, (3) a linear regression model for heart girth, a proxy for weight, to assess potential health and productivity impacts of GIN infections, based on the Nemabiome results (Supplementary Table S6). Age, deworming status, and sex were included as key confounding variables in the FEC and co-infection models. Variable selection was conducted using a backward stepwise approach, and Akaike Information Criterion (AIC) was assessed for each model (64).
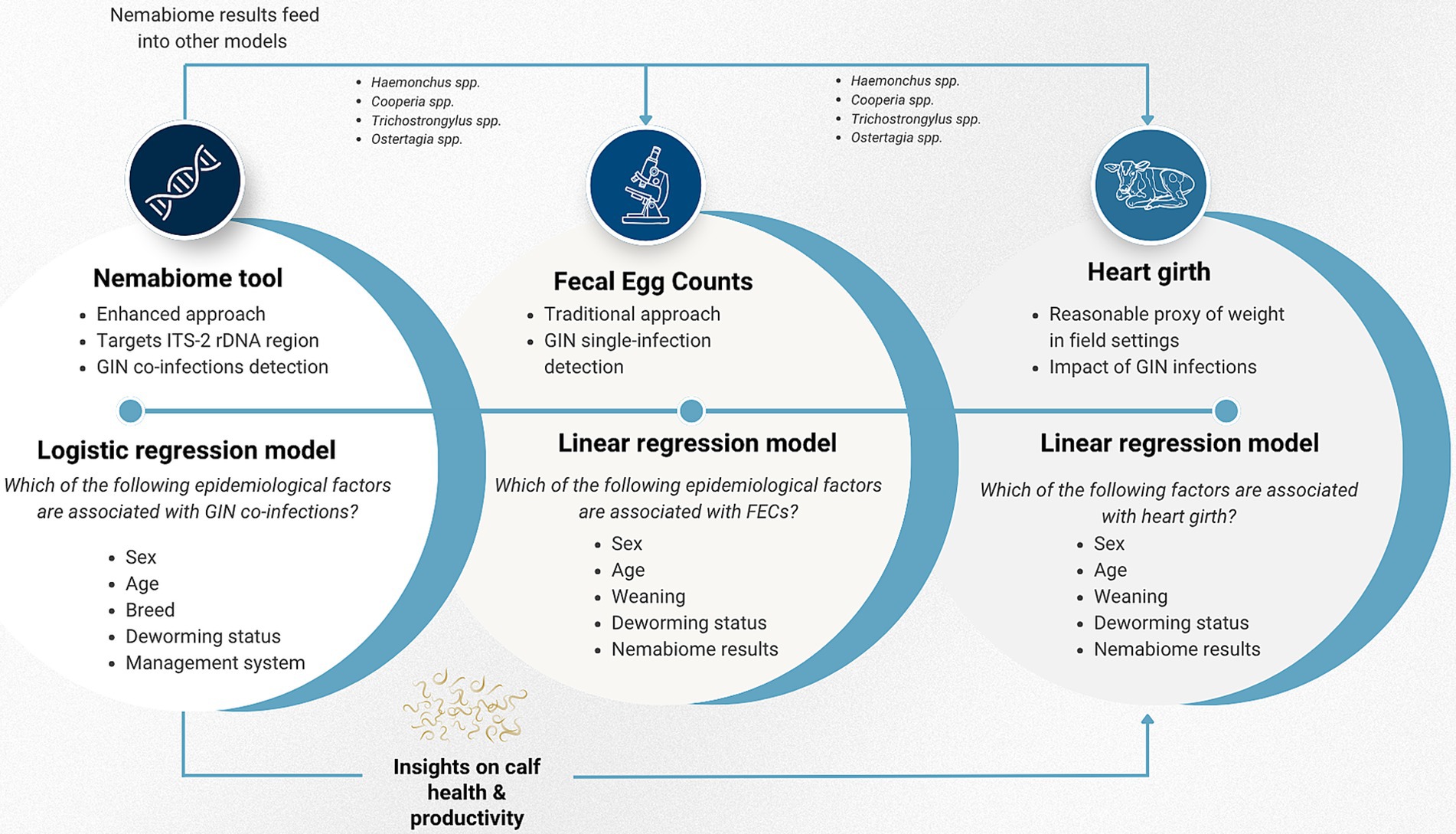
Figure 2. Conceptual framework of the statistical modeling. Three interconnected models were developed. The Nemabiome data feeds into the heart girth model, which assesses health and productivity impacts by evaluating the potential role of specific GIN species on heart girth. Simultaneously, recognizing that FEC is an imperfect diagnostic tool, the Nemabiome results are used as epidemiological factors to explore whether certain species are relevant when interpreting FEC findings.
To measure Alpha diversity among calves, the Shannon-Wiener (H) and Simpson indices (D) were calculated using the vegan package (version 2.6–8) (65). Simpson’s diversity index assesses the evenness of species in a community, its value varies from 0 to 1, with values close to zero suggesting low community diversity. In contrast, the Shannon diversity index considers both richness and evenness within a community, the higher the value, the more diverse the community (66).
3 Results
3.1 Description of the study population
The study population consisted of 532 dairy calves from 289 farms. Over half of the calves were females (57%) and most were kept under a pasture management system (85.5%). Most calves were young, aged 3–4 months (32.7%) and had been weaned (45.7%) at the time of sampling. Holstein-Friesian crosses (61.3%) were the predominant breed, followed by Ayrshire (36.3%) and Channel Island crosses (2.4%). Most of the calves had been dewormed in the 4 weeks prior to sample collection (63.5%), primarily with benzimidazoles (45.5%) (Supplementary Table S7).
3.2 Fecal egg count results
Among the 532 sampled calves, 98.7% (n = 525) had at least one egg per gram (EPG), with a mean of 62.0 ± 93.0. Most calves, 64.2% (n = 337) had low-intensity FEC (<50 EPG), 28.6% (n = 150) had medium intensity (50–200 EPG), and 7.2% (n = 38) had high intensity (>200 EPG). The highest mean FEC was observed in male (72.9 ± 104.3 EPG), non-weaned calves (71.0 ± 105.7 EPG), aged 3–4 months (77.5 ± 123.8 EPG), and those that had not been dewormed (67.4 ± 88.5 EPG) (Supplementary Table S8).
Univariable analysis results are provided in Supplementary Table S9. The final multivariable linear regression model for FEC showed that male calves had significantly higher FEC compared to females (estimate = 0.23, 95% CI: 0.02–0.43), while FEC decreased with age (estimate = −0.07, 95% CI −0.11 to −0.03). Calves that were not dewormed had a higher FEC compared to the dewormed calves (estimate = 0.3, 95% CI: 0.04–0.56). H. placei (estimate = 0.36, 95% CI: 0.09–0.62) and C. punctata (estimate = 0.32, 95% CI: 0.05–0.58) were positively associated with higher FEC (Table 1).
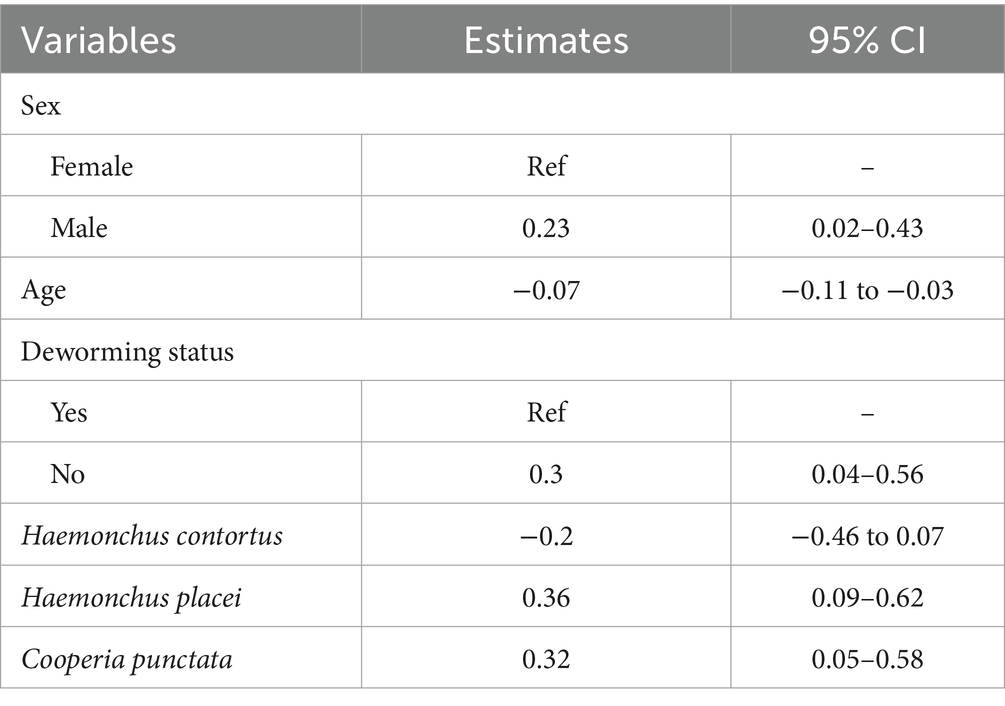
Table 1. Generalized mixed effects linear regression model for fecal egg count (FEC) in dairy calves including species associated with FEC.
3.3 Nemabiome of Kenyan dairy calves
A total of 528 samples were submitted for sequencing; out of these, 525 samples were analyzed, three were excluded due to missing linkage to the field IDs. The amplicon sequences had an average read number of 45,181, ranging from 503 to 189,339 reads. Based on sequences analysis we identified 214 Amplicon Sequence Variants (ASV) which were distributed across four genera: Cooperia, Haemonchus, Trichostrongylus and Ostertagia. Distinct clustering of ASVs was observed, with each major clade corresponding to a distinct trichostrongyloid GIN species (Figure 3). Haemonchus contortus was the most diverse species with more ASV, which reflect a high level of intraspecific variation. This was followed by Cooperia punctata, Cooperia curticei, Trichostrongylus axei, Trichostrongylus colubriformis, Haemonchus placei and Ostertagia ostertagi which constituted smaller but distinct clusters, likely indicating lower sequences diversity within these species. Cooperia pectinata and Cooperia oncophora exhibited the lowest ASV counts with 4 and 1 ASVs, respectively.
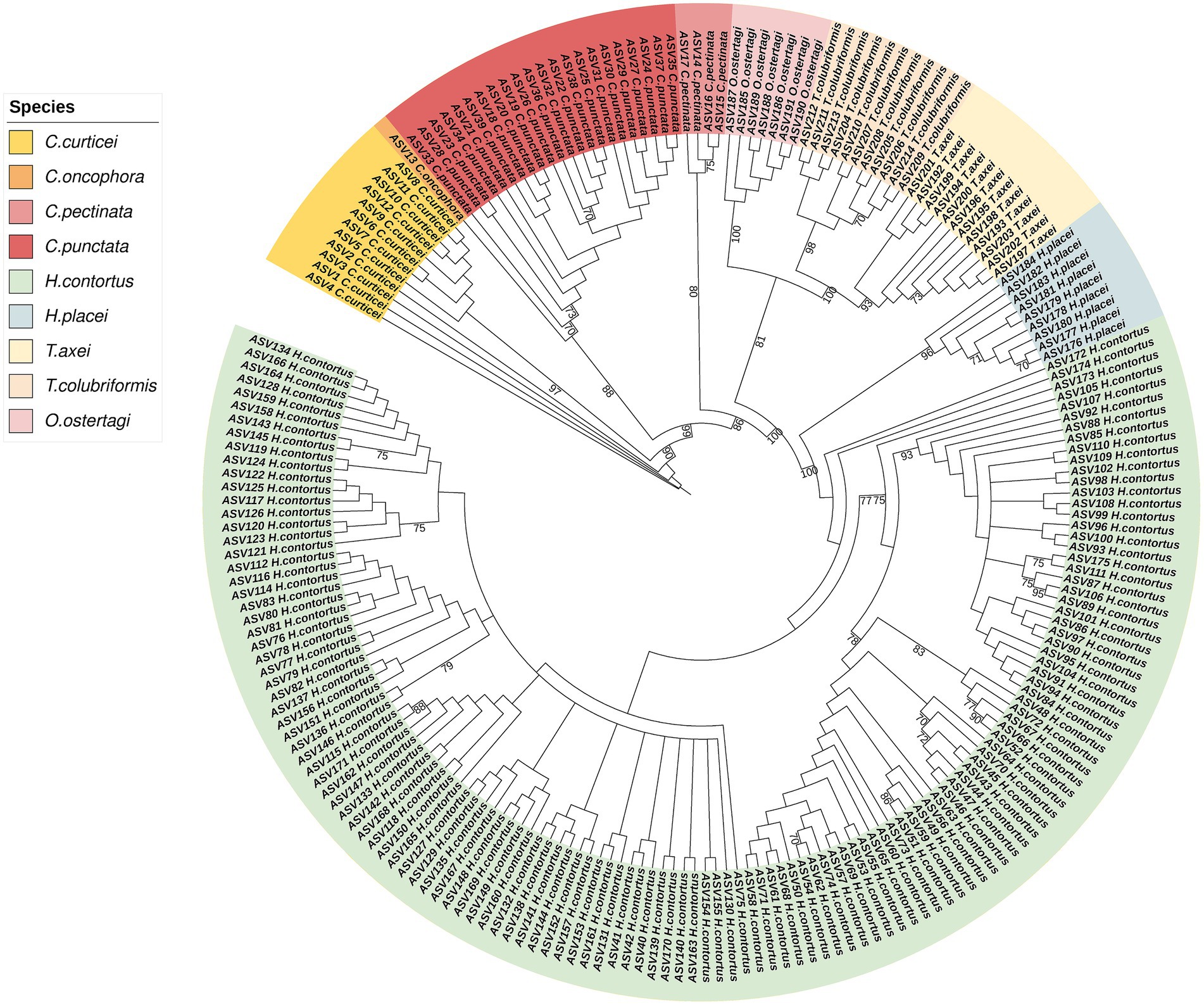
Figure 3. Maximum likelihood phylogenetic tree of ASV from the study samples. ITS-2 rDNA amplicons were sequenced using Illumina MiSeq sequencing. The sequences are differentiated by color, each representing their respective species. The numerical annotations on the branch are bootstrap values and >70 bootstrap value is shown.
The prevalence of GIN from deep amplicon sequencing results was 49.9% (95% CI: 44.8–55.0; n = 262). Among the species identified, C. punctata was the most prevalent at 27.8% (95% CI: 23.5–32.5), followed by H. placei 26.3% (95% CI: 22.2–30.8), H. contortus 23.6% (95% CI: 19.6–28.1), T. axei 16.4% (95% CI: 13.1–20.3), O. ostertagi 14.5% (95% CI: 11.5–18.1), T. colubriformis 8.0% (95% CI: 5.8–10.9), C. curticei 2.9% (95% CI: 1.7–4.8), C. pectinata 2.5% (95% CI: 1.3–4.7), and C. oncophora 0.2% (95% CI: 0.0–1.3). The proportion of unique nematode species and read abundance per animal is provided in Supplementary Figure S1.
Nemabiome results showed that, 30.5% of calves (n = 80) had single infections, while 69.5% (n = 182) were infected with two or more GIN species. Among these, two-species combinations were the most common (n = 73), followed by three-species combinations (n = 49), four-species combinations (n = 36), five-species combinations (n = 20), and finally, six-species combination (n = 4). For two-species infections, C. punctata frequently paired with H. contortus and H. placei (n = 15) whereas C. punctata, H. contortus, and H. placei were the most common three-species infections (n = 7). In four-species infections, the dominant group consisted of C. punctata, H. placei, O. ostertagi, and T. axei (n = 12). For five-species infections, C. punctata, H. contortus, H. placei, O. ostertagi, and T. axei dominated (n = 10) (Figure 4).
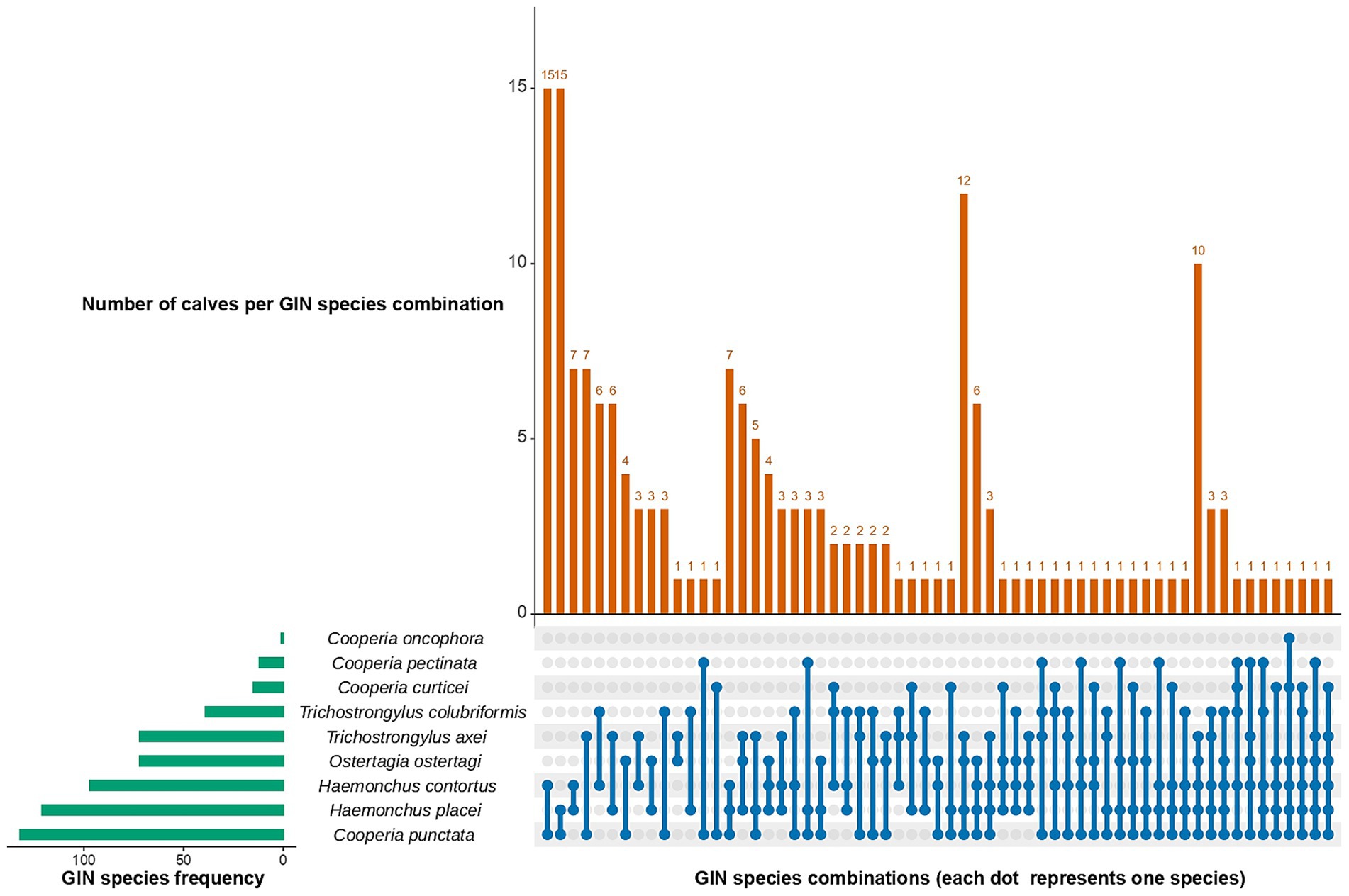
Figure 4. UpSet plot showing co-infection patterns for gastrointestinal nematodes (GIN) in dairy calves. The horizontal bars (left) represent the total number of samples positive for each species, while the vertical bars (top) indicate the number of calves with specific species combinations. Dots connected by lines at the bottom indicate species combinations.
Univariable analysis results for the co-infection model are provided in Supplementary Table S10. The final multivariable logistic regression model for co-infections showed that calves managed under a pasture system had twice the odds (OR = 2.52, 95% CI: 1.17–5.40) of experiencing mixed infections compared to those in zero-grazing systems. Age, deworming status, management system and sex were not significantly associated with co-infections (Table 2).
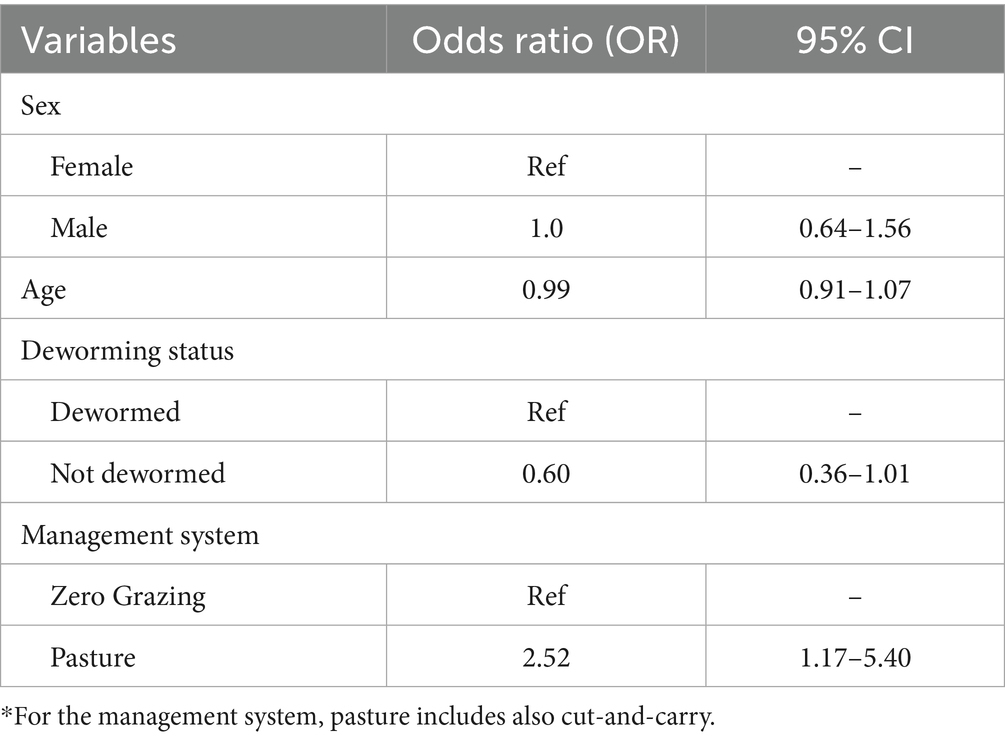
Table 2. Generalized mixed effects logistic regression model for co-infections in dairy calves.
3.4 Effect of GIN species and other factors on heart girth in dairy calves
The mean heart girth for all calves was 99.2 ± 14 cm (Supplementary Table S11). Univariable analysis results for the heart girth model are provided in Supplementary Table S12. The final multivariable linear regression model for heart girth showed that male calves had a significantly smaller girth compared to females (estimate = −2.61, 95% CI: −4.64 to −0.58). Age was positively associated with heart girth (estimate = 2.47, 95% CI: 2.06–2.89). Infections with O. ostertagi (estimate = −4.43, 95% CI: −7.65 to −1.20) and T. colubriformis (estimate = −3.96, 95% CI: −7.68 to −0.24) were associated with a decrease in girth (Table 3).
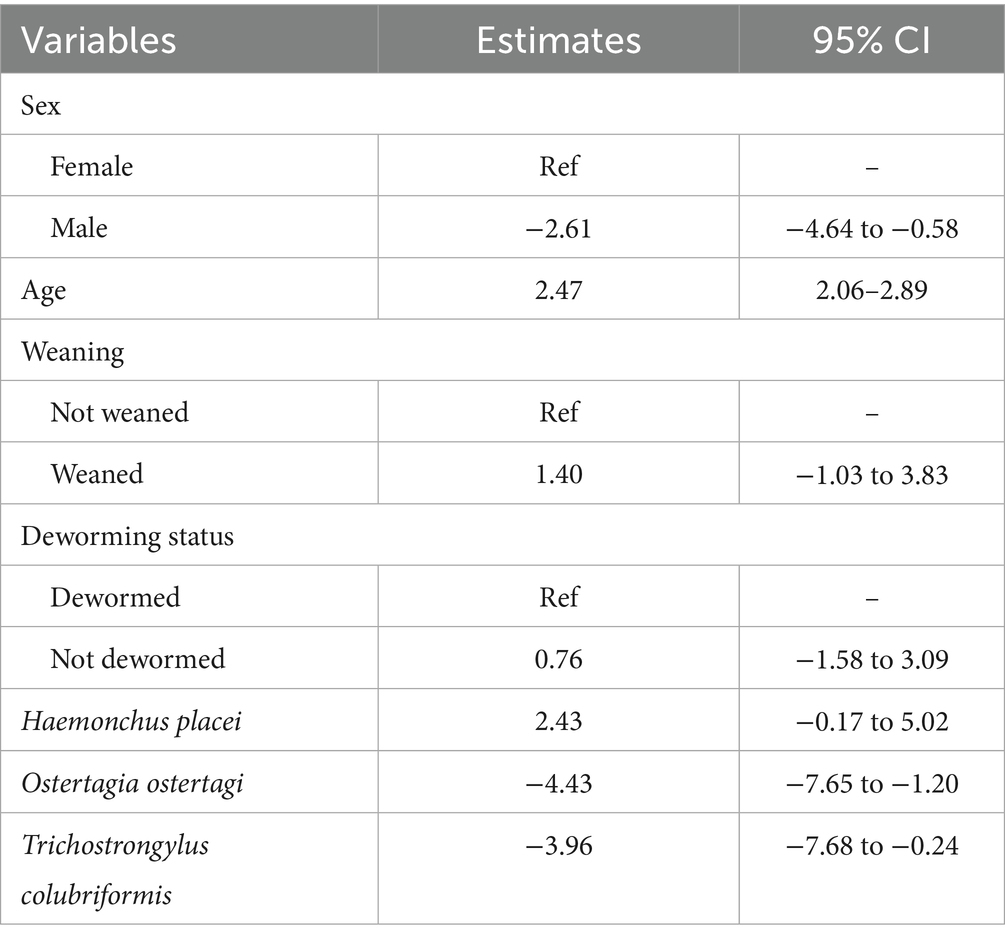
Table 3. Generalized mixed effects linear regression model for heart girth in dairy calves.
3.5 Alpha diversity indices
The Shannon Index ranged from 0 to 1.5, with a mean of 0.49 ± 0.43. In contrast, the Simpson Index ranged from 0 to 0.7 and a mean of 0.3 ± 0.25. Figure 5 illustrates Shannon and Simpson index values in relation to sex (Figure 5A), age (Figure 5B), deworming status (Figure 5C) and management system (Figure 5D). While the results were not statistically significant (p > 0.05) for any of the variables assessed, notable patterns were observed across several variables. Specifically, the 10–12 months age group presented slightly higher Shannon index values compared to the younger calves (3–4 months and 5–6 months), suggesting a tendency to increased GIN diversity with age. Female animals exhibited marginally greater GIN diversity than males. A modest increase in diversity was also observed in calves managed under pasture-grazing systems compared to those under zero-grazing management. Interestingly, calves that had been dewormed showed slightly higher GIN diversity than those that had not been dewormed.
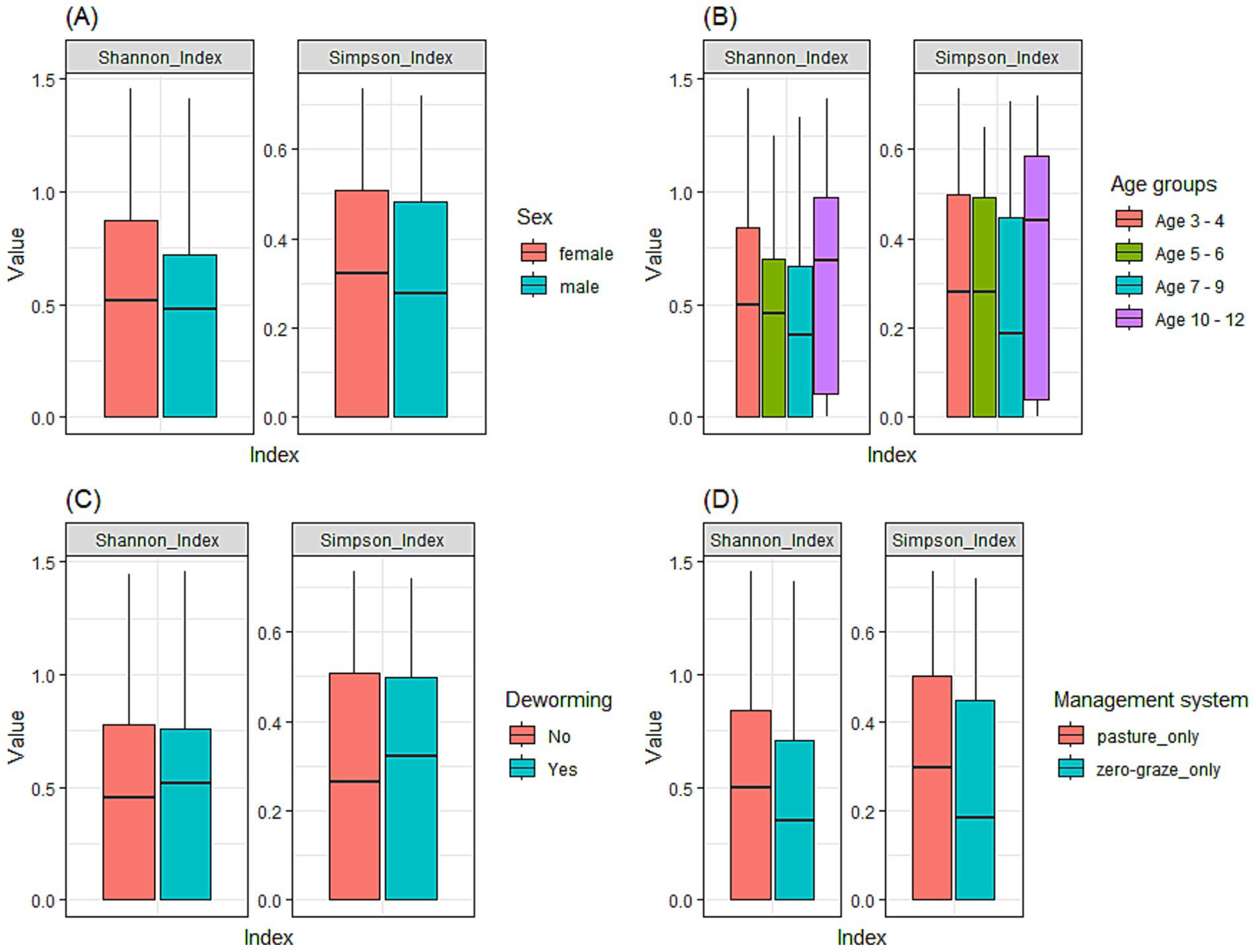
Figure 5. Alpha diversity of GIN communities in dairy calves using the Shannon and Simpson indices to compare diversity across various factors. (A) sex; (B) age groups (3–4, 5–6, 7–9, and 10–12 months); (C) deworming status; (D) management system.
4 Discussion
Microscopy-based diagnostic tests, remain essential for detecting GIN infections and estimating infection intensity, particularly for everyday diagnostic purposes. However, high-throughput molecular methods such as the Nemabiome allow for a robust and precise identification of individual GIN species and capture the complexity of co-infections, often undetected by conventional methods like FEC (10).
The mean FEC in this study was higher than the FEC reported in smallholder dairy farms in Nakuru and Nyeri Counties (48.0 EPG) (7), but lower than a study conducted in smallholder farms in Kirinyaga County (88.0 EPG) (37) in Kenya. In general, while the prevalence of FEC (i.e., the proportion of calves with at least one EPG) was high, the intensity of infection was generally low, with most of the calves in our study having less than 200 EPG. The different findings across FEC-based studies could be due to variations on the study design, test method, analyst experience, study population, host genetics, management system, and geoclimatic conditions (20, 67, 68).
Earlier studies have demonstrated that age significantly influences FEC (35, 69, 70), with young calves representing the highest-risk subpopulation for GIN infection (67), consistent with the findings of this study. Lower immunity among younger calves increases their susceptibility to various infections. Primary exposure to GIN species typically triggers a strong protective immune response, which significantly reduces the risk of GIN reinfection over time and contributes to reduced parasite transmission within pasture-based dairy herds (67, 69). In contrast, literature reports mixed findings regarding FEC differences between male and female calves. Some studies indicate males have a higher GIN burden as detected through FEC (36, 71), while others report no significant difference in infection rates between male and female, such as in pasture-kept indigenous cattle (39). In this study, the observed sex-related difference in FEC may be partially attributed to management practices in dairy farms, where male calves are usually weaned earlier and may experience undernutrition, leading to increased susceptibility to infections and stunted growth compared to their female counterparts (48, 49, 72).
Microscopic examination has previously identified multiple GIN species in Kenyan livestock (7, 37, 43). Using these traditional methods, the most common species belonged to Haemonchus, Cooperia, Trichostrongylus, Oesophagostomum, and Nematodirus genera. In contrast, through the nemabiome approach, this study identified nine different GIN species isolated from the L1, four Cooperia species (C. curticei, C. oncophora, C. pectinata, and C. punctata), two Haemonchus species (H. contortus and H. placei), two Trichostrongylus species (T. axei and T. colubriformis) and one Ostertagia species (O. ostertagi). Unlike previous studies in Kenya (7, 37, 42, 43, 73, 74), our study detected O. ostertagi, which had not been reported earlier. Prevalent in temperate regions, O. ostertagi has been documented in tropical cattle in Nigeria (36), South Africa (38) and Tanzania (40). We did not detect any Oesophagostomum species such as Oesophagostomum radiatum, which has been documented in past studies (7, 37, 43). Discrepancies across studies may be attributed to dissemination of parasites through animal movements (75), differential host susceptibility linked to genetics (67), pasture management strategies, anthelmintic protocols, and ecological factors such as climate change, that affect the environmental suitability for GIN survival across ecological niches (76). It also reflects the diagnostic performance of the approach used, with the nemabiome being a more reliable, high-resolution tool for species identification compared to traditional morphological and coproculture methods (10).
Haemonchus species are recognized for their pathogenicity in livestock, causing lesions in the abomasum and severe anaemia due to its blood-feeding behavior (77). Among these, H. placei and H. contortus emerge as the most pathogenic species (78). H. placei is typically found in cattle, although mixed infections with H. contortus can also occur (79). While H. contortus is regarded as the most significant internal parasite of sheep and goats, it can also infect cattle (80), with calves being highly susceptible (77). The co-rearing of cattle with small ruminants, a common practice in dairy farms within the study region, may have played a role in this local transmission dynamic.
T. colubriformis and T. axei species are known to infect both humans and livestock (81). In humans, adult worms of Trichostrongylus species inhabit the mucosa of the small intestine and in large numbers, can induce physical trauma leading to mucosal shedding, and hemorrhages in severe cases (82). In contrast, in cattle it lives in the abomasum, typically part of a mixed helminth infection, with clinical symptoms that include diarrhea, anorexia, progressive weight loss, and general weakness (83). Studies in Iran, Australia, and Thailand have reported infections in humans living near animals (84–86). In Kenya, T. colubriformis has been recorded in baboons in the Amboseli region, suggesting its presence in various hosts (87). Since GIN are soil-transmitted, they can contaminate the environment, leading to increased exposure among the human population (88). Using cow or sheep manure as fertilizer can further contribute to the spread of infection in farming communities, elevating the risk of zoonotic transmission (89). This can result in a higher incidence of infections, particularly among rural communities with poor hygiene standards and limited access to health services (81). The occurrence of Trichostrongylus species in livestock and wildlife raises public health concerns in Kenya, where little information is available about the burden of nematode infections among farming communities.
FEC findings are influenced by the burden of infection, species composition and fecundity of GIN, and individual differences in the immune-response (90), as well as the time of sampling (91). For instance, Haemonchus and Cooperia are highly fecund species compared to Ostertagia (20). Despite producing fewer eggs, Ostertagia is still highly pathogenic highlighting a key limitation of the FEC as a diagnostic method. In cases of co-infection, FEC results may be further misleading because different nematode eggs can appear similar under a microscope. If the person analysing the sample lacks expertise, they may misidentify the species, potentially leading to inaccurate identification and impacting the therapeutic plan.
Morphological measurements, such as heart girth, are valuable proxies for estimating livestock live weight, particularly in field settings where weighing scales are impractical (92–94). Heart girth has been validated in various contexts, including crossbred dairy cattle in Sub-Saharan Africa, and is consistently presented as an accurate tool across age and breed groups (93, 94). However, animal genetics, husbandry practices, and nutrition around weaning can influence growth performance. Along with individual immune responses and GIN parasitic loads, these factors collectively impact the type and severity of clinical signs and girth measurements (72, 95, 96).
Heavy or chronic parasitism can impair growth and weight gain by exacerbating the physiological stress associated with GIN infestation (48), which may partly explain the observed reduction in heart girth among male calves. For instance, O. ostertagi infection in cattle is often associated with chronic diarrhea and weight loss, especially during the first grazing season (97, 98). Similarly, infection with T. colubriformis is known to impair feed efficiency and nutrient absorption, leading to growth retardation, even in cases of subclinical GIN parasitism (99). Differences in management practices may also play a role. In dairy-oriented regions, farmers tend to provide more care to female calves due to their value as replacement stock and their importance to the dairy enterprise (96). These findings underscore the importance of effective, integrated GIN control in the farm to interrupt transmission cycles of GIN shared by calves, adult animals, and other hosts.
Mixed parasitic infections are common in pasture-managed cattle (36, 100, 101). Favorable climatic conditions provides a conducive environment for GIN to mature (102), leading to pasture contamination and increasing the likelihood of calves being exposed to infective larvae. Co-infection with multiple parasites can influence both the spread and severity of diseases; parasite interactions may alter the immune response of the host, making them more susceptible to other infections (103). For instance, Thumbi et al. (104) highlighted that the risk of death from the infection by Theileria parva (causative agent of East Coast fever) significantly increased with a high helminth burden measured as strongyle EPG. Nematode infections in buffalos have also been linked to increased vulnerability to diseases like bovine tuberculosis (bTB), associated with a reduction in the Th1 immune response (105).
The alpha diversity indices revealed no statistically significant differences in GIN species among calves in the study region. This suggests that the overall diversity of GIN in dairy calves remains relatively stable, regardless of features such as sex or age. Increased GIN exposure and continuous reinfection cycles occur with prolonged grazing periods, after maternal immunity wanes, allowing for colonization by multiple parasite species (67). The slightly higher diversity observed in dewormed animals is a counterintuitive finding, potentially arising from combination of recall bias among smallholders regarding their anthelmintic schedules, or alternatively indicating reinfection from contaminated pasture, where deworming without proper pasture management facilitates GIN host repopulation (106). This finding also highlights the advantages of using the Nemabiome tool as the next-generation approach to GIN characterization compared to traditional methods. Further research is needed to better understand the observed patterns, particularly integrating emerging anthelmintic resistance with ecological and husbandry factors.
Our study offers valuable insights into the prevalence, predominant GIN species, and co-infections affecting calves in one of the most important dairy regions in Kenya. The increasing use of the Nemabiome tool to monitor GIN diversity and anthelmintic resistance in high-income countries (HICs) (3, 10, 16, 30, 31, 107, 108), highlights emerging opportunities for its application in tropical regions, where the burden of infection is typically higher. This aligns with the growing need for sustainable and safe strategies to manage GIN, particularly considering the cost of anthelmintics, the increase in drug resistance, and consumer concerns about animal welfare and organic animal products. Although the Nemabiome technique remains relatively costly for widespread implementation in resource-limited settings, it becomes more cost-effective with larger sample sizes and serves as a powerful tool for evidence-based practice (10).
Future research should focus on assessing the GIN burden in calves under 3 months of age, who were not included in this study, and conduct longitudinal surveys to capture seasonal variations and long-term trends related to the impact of GIN infections on calf and heifer health, welfare, and productivity.
5 Conclusion
This study is the first to the assess the burden and diversity of gastrointestinal nematodes in dairy calves in Africa using a combination of fecal egg count (FEC) analysis and the Nemabiome assay using L1. In the study region, age, deworming, and management practices were found to influence the nematode burden, which in turn had an impact on heart girth, a proxy for weight. Significant associations between infections with O. ostertagi and T. colubriformis and reduced heart girth were identified, underscoring the clinical significance of the findings. Due to the complex epidemiology of GIN in calves in the region, characterized by frequent co-infections, this study also highlights the need to evaluate the efficacy of commonly used anthelminthics to support the design of a sustainable GIN control strategy.
Data availability statement
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found at: https://www.ebi.ac.uk/ena, PRJEB85916 and the animal metadata and test results are available at https://doi.org/10.3389/fvets.2025.1588350.
Ethics statement
The animal studies were approved by International Livestock Research Institute (ILRI) Animal Care and Use Committee International Livestock Research Institute (ILRI) Institutional Research Ethics Committee. The studies were conducted in accordance with the local legislation and institutional requirements. Written informed consent was obtained from the owners for the participation of their animals in this study.
Author contributions
SC: Conceptualization, Data curation, Formal analysis, Investigation, Methodology, Validation, Visualization, Writing – original draft, Writing – review & editing. EY: Conceptualization, Data curation, Formal analysis, Methodology, Writing – original draft, Writing – review & editing, Investigation, Validation, Visualization, Software. LG: Data curation, Formal analysis, Writing – original draft, Writing – review & editing, Validation, Visualization, Methodology. BR: Data curation, Formal analysis, Investigation, Writing – review & editing. JSK: Data curation, Formal analysis, Investigation, Writing – review & editing. JW: Data curation, Formal analysis, Investigation, Writing – review & editing. BK: Conceptualization, Formal analysis, Investigation, Methodology, Writing – review & editing. GiN: Investigation, Writing – review & editing. SM: Investigation, Writing – review & editing. JK: Investigation, Writing – review & editing. LK: Investigation, Writing – review & editing. NS: Investigation, Writing – review & editing. DV: Data curation, Software, Validation, Writing – review & editing. GeN: Investigation, Writing – review & editing. JO: Conceptualization, Formal analysis, Supervision, Validation, Writing – review & editing. GM: Conceptualization, Formal analysis, Supervision, Validation, Writing – review & editing. BB: Conceptualization, Data curation, Formal analysis, Funding acquisition, Investigation, Methodology, Project administration, Resources, Software, Validation, Visualization, Writing – review & editing, Supervision. EC: Conceptualization, Data curation, Formal analysis, Funding acquisition, Investigation, Methodology, Project administration, Resources, Software, Supervision, Validation, Visualization, Writing – review & editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. The research project was funded in part by Gates Foundation and with UK aid from the UK Foreign, Commonwealth and Development Office (Grant Agreement INV-040641) under the auspices of the Centre for Tropical Livestock Genetics and Health (CTLGH), established jointly by the University of Edinburgh, SRUC (Scotland’s Rural College), and the International Livestock Research Institute. The findings and conclusions contained within are those of the authors and do not necessarily reflect the positions or policies of the Bill & Melinda Gates Foundation nor the UK Government. Research at the Roslin Institute was funded through support from the UK Biotechnology and Biological Sciences Research Council (BBS/E/RL/230002C; BBS/E/RL/230002D - BB, EY).
Acknowledgments
The authors would like to thank Lessos Cooperative and farmers from Nandi and Uasin Gishu County and Kapsabet Veterinary Laboratory, under the Ministry of Agriculture in Nandi County. We also thank the cooperative dairy farm assistants (DFAs) and animal health assistants (AHA) for their help in sample collection. We express our gratitude to Professor Neil Sargison for the positive controls used in this study.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declared that they were an editorial board member of Frontiers, at the time of submission. This had no impact on the peer review process and the final decision.
Generative AI statement
The authors declare that Gen AI was used in the creation of this manuscript. Generative AI was used for editing grammar and avoid conciseness.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fvets.2025.1588350/full#supplementary-material
Footnotes
1. ^https://portal.adgg.ilri.org/
2. ^https://www.tanbead.com/en
4. ^https://github.com/deepalivasoya/MiAmp
5. ^https://users-birc.au.dk/palle/php/fabox/dnacollapser.php
References
1. Adamou Karimou, I, Abdou, H, and Issa, M. Prevalence and risk factors for gastrointestinal parasitosis in small ruminant farms in the Sahelian zone of Niger. Vet Parasitol Reg Stud Reports. (2024) 53:101070. doi: 10.1016/j.vprsr.2024.101070
2. Oliveira Macedo, L, Souza Silva, S, Alves, LC, Carvalho, GA, Antonio, R, and Ramos, N. An overview of anthelmintic resistance in domestic ruminants in Brazil. Ruminants. (2023) 3:214–32. doi: 10.3390/ruminants3030020
3. Borkowski, EA, Redman, EM, Chant, R, Avula, J, Menzies, PI, Karrow, NA, et al. Comparison of ITS-2 rDNA nemabiome sequencing with morphological identification to quantify gastrointestinal nematode community species composition in small ruminant feces. Vet Parasitol. (2020) 282:109104. doi: 10.1016/j.vetpar.2020.109104
4. Charlier, J, Höglund, J, von Samson-Himmelstjerna, G, Dorny, P, and Vercruysse, J. Gastrointestinal nematode infections in adult dairy cattle: Impact on production, diagnosis and control. Vet Parasitol. (2009) 164:70–9. doi: 10.1016/j.vetpar.2009.04.012
5. Vande Velde, F, Charlier, J, and Claerebout, E. Farmer Behavior and Gastrointestinal Nematodes in Ruminant Livestock—Uptake of Sustainable Control Approaches. Front Vet Sci. (2018) 5:255. doi: 10.3389/fvets.2018.00255
6. Thumbi, SM, De, C, Bronsvoort, BM, Poole, EJ, Kiara, H, Toye, P, et al. Parasite co-infections show synergistic and antagonistic interactions on growth performance of East African zebu cattle under one year. Parasitology. (2013) 140:1789–98. doi: 10.1017/S0031182013001261
7. Kabaka, W, Gitau, G, Kitala, P, Maingi, N, and Van Leeuwen, J. The prevalence of gastrointestinal nematode infection and their impact on cattle in Nakuru and Mukurweini districts of Kenya. Ethiop Vet J. (2014) 17:95. doi: 10.4314/evj.v17i1.8
8. May, K, Brügemann, K, König, S, and Strube, C. Patent gastrointestinal nematode infections in organically and conventionally pastured dairy cows and their impact on individual milk and fertility parameters. Vet Parasitol. (2017) 245:119–27. doi: 10.1016/j.vetpar.2017.08.024
9. Zahid, O, Butler, M, Hopker, A, Freeman, E, Costa Júnior, LM, Chaudhry, U, et al. Nemabiome metabarcoding shows a high prevalence of Haemonchus contortus and predominance of Camelostrongylus mentulatus in alpaca herds in the northern UK. Parasitol Res. (2024) 123:201–10. doi: 10.1007/s00436-024-08226-w
10. Avramenko, RW, Redman, EM, Lewis, R, Yazwinski, TA, Wasmuth, JD, and Gilleard, JS. Exploring the Gastrointestinal “Nemabiome”: Deep Amplicon Sequencing to Quantify the Species Composition of Parasitic Nematode Communities. PLoS One. (2015) 10:e0143559. doi: 10.1371/journal.pone.0143559
11. Wang, T, Redman, EM, Morosetti, A, Chen, R, Kulle, S, Morden, N, et al. Seasonal epidemiology of gastrointestinal nematodes of cattle in the northern continental climate zone of western Canada as revealed by internal transcribed spacer-2 ribosomal DNA nemabiome barcoding. Parasit Vectors. (2021) 14:604. doi: 10.1186/S13071-021-05101-W
12. Avramenko, RW, Redman, EM, Lewis, R, Bichuette, MA, Palmeira, BM, Yazwinski, TA, et al. The use of nemabiome metabarcoding to explore gastro-intestinal nematode species diversity and anthelmintic treatment effectiveness in beef calves. Int J Parasitol. (2017) 47:893–902. doi: 10.1016/j.ijpara.2017.06.006
13. Ezenwa, VO, and Jolles, AE. From Host Immunity to Pathogen Invasion: The Effects of Helminth Coinfection on the Dynamics of Microparasites. Integr Comp Biol. (2011) 51:540–51. doi: 10.1093/icb/icr058
14. Supali, T, Verweij, JJ, Wiria, AE, Djuardi, Y, Hamid, F, Kaisar, MMM, et al. Polyparasitism and its impact on the immune system. Int J Parasitol. (2010) 40:1171–6. doi: 10.1016/j.ijpara.2010.05.003
15. Wimmer, B, Craig, BH, Pilkington, JG, and Pemberton, JM. Non-invasive assessment of parasitic nematode species diversity in wild Soay sheep using molecular markers. Int J Parasitol. (2004) 34:625–31. doi: 10.1016/j.ijpara.2003.11.022
16. Queiroz, C, Levy, M, Avramenko, R, Redman, E, Kearns, K, Swain, L, et al. The use of ITS-2 rDNA nemabiome metabarcoding to enhance anthelmintic resistance diagnosis and surveillance of ovine gastrointestinal nematodes. Int J Parasitol Drugs Drug Resist. (2020) 14:105–17. doi: 10.1016/j.ijpddr.2020.09.003
17. Baiak, BHB, Lehnen, CR, and da Rocha, RA. Anthelmintic resistance in cattle: A systematic review and meta-analysis. Livest Sci. (2018) 217:127–35. doi: 10.1016/j.livsci.2018.09.022
18. Tsotetsi, AM, Njiro, S, Katsande, TC, Moyo, G, Baloyi, F, and Mpofu, J. Prevalence of gastrointestinal helminths and anthelmintic resistance on small-scale farms in Gauteng Province, South Africa. Trop Anim Health Prod. (2013) 45:751–61. doi: 10.1007/s11250-012-0285-z
19. Demeler, J, Ramünke, S, Wolken, S, Ianiello, D, Rinaldi, L, Gahutu, JB, et al. Discrimination of Gastrointestinal Nematode Eggs from Crude Fecal Egg Preparations by Inhibitor-Resistant Conventional and Real-Time PCR. PLoS One. (2013) 8:e61285. doi: 10.1371/journal.pone.0061285
20. Sabatini, GA, de Almeida, BF, Claerebout, E, Gianechini, LS, Höglund, J, Kaplan, RM, et al. Practical guide to the diagnostics of ruminant gastrointestinal nematodes, liver fluke and lungworm infection: interpretation and usability of results. Parasit Vectors. (2023) 16:1–17. doi: 10.1186/s13071-023-05680-w
21. Redman, E, Queiroz, C, Bartley, DJ, Levy, M, Avramenko, RW, and Gilleard, JS. Validation of ITS-2 rDNA nemabiome sequencing for ovine gastrointestinal nematodes and its application to a large scale survey of UK sheep farms. Vet Parasitol. (2019) 275:108933. doi: 10.1016/j.vetpar.2019.108933
22. Yan, X, He, S, Liu, Y, Han, B, Zhang, N, Deng, H, et al. Molecular identification and phylogenetic analysis of gastrointestinal nematodes in different populations of Kazakh sheep. Exp Parasitol. (2023) 254:108625. doi: 10.1016/j.exppara.2023.108625
23. Baltrušis, P, Halvarsson, P, and Höglund, J. Molecular detection of two major gastrointestinal parasite genera in cattle using a novel droplet digital PCR approach. Parasitol Res. (2019) 118:2901–7. doi: 10.1007/s00436-019-06414-7
24. Rashid, MH, Gebrekidan, H, and Jabbar, A. Multiplexed-tandem PCR (MT-PCR) assay to detect and differentiate gastrointestinal nematodes of alpacas. Parasit Vectors. (2018) 11:370–6. doi: 10.1186/s13071-018-2963-9
25. Roeber, F, Morrison, A, Casaert, S, Smith, L, Claerebout, E, and Skuce, P. Multiplexed-tandem PCR for the specific diagnosis of gastrointestinal nematode infections in sheep: an European validation study. Parasit Vectors. (2017) 10:226–11. doi: 10.1186/s13071-017-2165-x
26. Zarlenga, DS, Barry Chute, M, Gasbarre, LC, and Boyd, PC. A multiplex PCR assay for differentiating economically important gastrointestinal nematodes of cattle. Vet Parasitol. (2001) 97:201–11. doi: 10.1016/s0304-4017(01)00410-1
27. Blaxter, M. Nematodes: The Worm and Its Relatives. PLoS Biol. (2011) 9:e1001050. doi: 10.1371/journal.pbio.1001050
28. Parkinson, J, Mitreva, M, Whitton, C, Thomson, M, Daub, J, Martin, J, et al. A transcriptomic analysis of the phylum Nematoda. Nat Genet. (2004) 36:1259–67. doi: 10.1038/ng1472
29. De Seram, EL, Uehlinger, FD, de Queiroz, C, Redman, EM, Campbell, JR, Nooyen, D, et al. Integration of ITS-2 rDNA nemabiome metabarcoding with Fecal Egg Count Reduction Testing (FECRT) reveals ivermectin resistance in multiple gastrointestinal nematode species, including hypobiotic Ostertagia ostertagi, in western Canadian beef cattle. Int J Parasitol Drugs Drug Resist. (2023) 22:27–35. doi: 10.1016/j.ijpddr.2023.04.002
30. Scott, H, Gilleard, JS, Jelinski, M, Barkema, HW, Redman, EM, Avramenko, RW, et al. Prevalence, fecal egg counts, and species identification of gastrointestinal nematodes in replacement dairy heifers in Canada. J Dairy Sci. (2019) 102:8251–63. doi: 10.3168/jds.2018-16115
31. Halvarsson, P, and Höglund, J. Sheep nemabiome diversity and its response to anthelmintic treatment in Swedish sheep herds. Parasit Vectors. (2021) 14:1–12. doi: 10.1186/s13071-021-04602-y
32. Beaumelle, C, Redman, EM, de Rijke, J, Wit, J, Benabed, S, Debias, F, et al. Metabarcoding in two isolated populations of wild roe deer (Capreolus capreolus) reveals variation in gastrointestinal nematode community composition between regions and among age classes. Parasit Vectors. (2021) 14:594. doi: 10.1186/S13071-021-05087-5
33. Hamad, MH, Islam, SI, Jitsamai, W, Chinkangsadarn, T, Naraporn, D, Ouisuwan, S, et al. Metabarcoding study to reveal the structural community of strongylid nematodes in domesticated horses in Thailand. BMC Vet Res. (2024) 20:70. doi: 10.1186/s12917-024-03934-y
34. Poissant, J, Gavriliuc, S, Bellaw, J, Redman, EM, Avramenko, RW, Robinson, D, et al. A repeatable and quantitative DNA metabarcoding assay to characterize mixed strongyle infections in horses. Int J Parasitol. (2021) 51:183–92. doi: 10.1016/j.ijpara.2020.09.003
35. Tiele, D, Sebro, E, Meskel, DH, and Mathewos, M. Epidemiology of Gastrointestinal Parasites of Cattle in and Around Hosanna Town, Southern Ethiopia. Vet Med Res Rep. (2023) 14:1–9. doi: 10.2147/VMRR.S389787
36. Ola-Fadunsin, SD, Ganiyu, IA, Rabiu, M, Hussain, K, Sanda, IM, Baba, AY, et al. Helminth infections of great concern among cattle in Nigeria: Insight to its prevalence, species diversity, patterns of infections and risk factors. Vet World. (2020) 13:338–44. doi: 10.14202/vetworld.2020.338-344
37. Njonge, FK. Epidemiology of Helminths in Cattle of Kirinyaga County, Kenya. IOSR J Agric Vet Sci. (2017) 10:106–9. doi: 10.9790/2380-100501106109
38. Horak, IG, Evans, U, and Purnell, RE. Parasites of domestic and wild animals in South Africa. XLV. Helminths of dairy calves on dry-land Kikuyu grass pastures in the Eastern Cape Province. Onderstepoort J Vet Res. (2004) 71:291–306. doi: 10.4102/ojvr.v71i4.249
39. Keyyu, JD, Kassuku, AA, Kyvsgaard, NC, and Willingham, AL. Gastrointestinal nematodes in indigenous Zebu cattle under pastoral and nomadic management systems in the lower plain of the Southern Highlands of Tanzania. Vet Res Commun. (2003) 27:371–80. doi: 10.1023/A:1024706120270
40. Keyyu, JD, Kyvsgaard, NC, Monrad, J, and Kassuku, AA. Epidemiology of gastrointestinal nematodes in cattle on traditional, small-scale dairy and large-scale dairy farms in Iringa district, Tanzania. Vet Parasitol. (2005) 127:285–94. doi: 10.1016/j.vetpar.2004.10.014
41. Waruiru, RM, Weda, EH, Otieno, RO, and Ngotho, JW. Seasonal availability of gastrointestinal nematode larvae to cattle on pasture in the central highlands of Kenya. Onderstepoort J Vet Res. (2002) 69:141–6.
42. Waruiru, RM, Thamsborg, SM, Nansen, P, Kyvsgaard, NC, Bogh, HO, Munyua, WK, et al. The epidemiology of gastrointestinal nematodes of dairy cattle in Central Kenya. Trop Anim Health Prod. (2001) 33:173–87. doi: 10.1023/a:1010322703790
43. Waruiru, RM, Nansen, P, Kyvsgaard, NC, Thamsborg, SM, Munyua, WK, Gathuma, JM, et al. An abattoir survey of gastrointestinal nematode infections in cattle in the Central Highlands of Kenya. Vet Res Commun. (1998) 22:325–34. doi: 10.1023/A:1006164805185
44. MoALFC. Climate risk profile for nandi county. Kenya county climate risk profile series. Nairobi, Kenya: The Ministry of Agriculture, Livestock, Fisheries and Co-operatives (MoALFC). CGspace. (2021) 1–33.
45. Erickson, PS, and Kalscheur, KF. Nutrition and feeding of dairy cattle In: Animal Agriculture Sustainability, Challenges and Innovations (2020). 157–80.
46. Audigé, L. Veterinary Epidemiologic Research: I. Dohoo, W. Martin, H. Stryhn, Atlantic Veterinary College, Charlottetown, PE, Canada, 2003. Prev Vet Med. (2005) 68:289–92. doi: 10.1016/j.prevetmed.2004.11.001
47. FAO. Fao Statistical Developement Series 20 World Programme for the Census of Agriculture 2030 Programme, Concepts and Definitions. Food and Agricultural Organization of The United Nations Rome.
48. Chang'a, JS, Mdegela, RH, Ryoba, R, Løken, T, and Reksen, O. Calf health and management in smallholder dairy farms in Tanzania. Trop Anim Health Prod. (2010) 42:1669–76. doi: 10.1007/s11250-010-9619-x
49. Welk, A, Neave, HW, and Jensen, MB. Invited review: The effect of weaning practices on dairy calf performance, behavior, and health—A systematic review. J Dairy Sci. (2024) 107:5237–58. doi: 10.3168/jds.2024-24521
50. Molento, MB, Buzatti, A, and Sprenger, LK. Pasture larval count as a supporting method for parasite epidemiology, population dynamic and control in ruminants. Livest Sci. (2016) 192:48–54. doi: 10.1016/j.livsci.2016.08.013
51. Chase, CCL, Hurley, DJ, and Reber, AJ. Neonatal Immune Development in the Calf and Its Impact on Vaccine Response. Vet Clin North Am Food Anim Pract. (2008) 24:87–104. doi: 10.1016/j.cvfa.2007.11.001
52. Hartung, C, Anokwa, Y, Brunette, W, Lerer, A, Tseng, C, and Borriello, G. Open data kit: Tools to build information services for developing regions. ACM International Conference Proceeding Series (2010).
53. Christie, M, and Jackson, F. Specific identification of strongyle eggs in small samples of sheep faeces. Res Vet Sci. (1982) 32:113–7. doi: 10.1016/S0034-5288(18)32448-2
54. Burgess, CGS, Bartley, Y, Redman, E, Skuce, PJ, Nath, M, Whitelaw, F, et al. A survey of the trichostrongylid nematode species present on UK sheep farms and associated anthelmintic control practices. Vet Parasitol. (2012) 189:299–307. doi: 10.1016/j.vetpar.2012.04.009
55. Silwamba, I, Vasoya, D, Simuunza, M, Tzelos, T, Nalubamba, KS, Simulundu, E, et al. High throughput analysis of MHC class I and class II diversity of Zambian indigenous cattle populations. HLA. (2023) 101:458–83. doi: 10.1111/TAN.14976
56. Joshi, NA, and Fass, JN. Git Hub - najoshi/sickle: Windowed Adaptive Trimming for fastq files using quality. (2021). Available online at: https://github.com/najoshi/sickle.
57. Magoč, T, and Salzberg, SL. FLASH: fast length adjustment of short reads to improve genome assemblies. Bioinformatics. (2011) 27:2957–63. doi: 10.1093/bioinformatics/btr507
58. Camacho, C, Coulouris, G, Avagyan, V, Ma, N, Papadopoulos, J, Bealer, K, et al. BLAST+: Architecture and applications. BMC Bioinformatics. (2009) 10:1–9. doi: 10.1186/1471-2105-10-421
59. Gouy, M, Tannier, E, Comte, N, and Parsons, DP. Seaview Version 5: A Multiplatform Software for Multiple Sequence Alignment, Molecular Phylogenetic Analyses, and Tree Reconciliation. Methods Mol Biol. (2021) 2231:241–60. doi: 10.1007/978-1-0716-1036-7_15
60. Katoh, K, Misawa, K, Kuma, KI, and Miyata, T. MAFFT: a novel method for rapid multiple sequence alignment based on fast Fourier transform. Nucleic Acids Res. (2002) 30:3059–66. doi: 10.1093/NAR/GKF436
61. Letunic, I, and Bork, P. Interactive Tree of Life (iTOL) v6: recent updates to the phylogenetic tree display and annotation tool. Nucleic Acids Res. (2024) 52:W78–82. doi: 10.1093/NAR/GKAE268
62. Thomas, L. Complex surveys: a guide to analysis using R In: Wiley Series in Survey Methodology, vol. 296 (2010)
63. Bates, D, Mächler, M, Bolker, BM, and Walker, SC. Fitting linear mixed-effects models using lme 4. J Stat Softw. (2015) 67:1–48. doi: 10.18637/jss.v067.i01
64. Ratner, B. Variable selection methods in regression: Ignorable problem, outing notable solution. J Target Meas Anal Mark. (2010) 18:65–75. doi: 10.1057/jt.2009.26
65. Oksanen, J, Simpson, GL, Blanchet, FG, Kindt, R, Legendre, P, Minchin, PR, et al. vegan: Community Ecology Package. CRAN: Contributed Packages (2001).
66. Finn, DR. A metagenomic alpha-diversity index for microbial functional biodiversity. FEMS Microbiol Ecol. (2024) 100:fiae019. doi: 10.1093/FEMSEC/FIAE019
67. Gasbarre, LC, Leighton, EA, and Sonstegard, T. Role of the bovine immune system and genome in resistance to gastrointestinal nematodes. Vet Parasitol. (2001) 98:51–64. doi: 10.1016/S0304-4017(01)00423-X
68. Moussouni, L, Benhanifia, M, Saidi, M, and Ayad, A. Prevalence of gastrointestinal parasitism infections in cattle of bass Kabylie area: Case of Bejaia province. Algeria Maced Vet Rev. (2018) 41:73–82. doi: 10.2478/macvetrev-2018-0010
69. da Silva, JB, Fagundes, GM, Soares, JPG, and da Fonseca, AH. Parasitism level by helminths and weight gain of calves kept in organic and conventional grazing. Pesqui Veterinária Bras. (2013) 33:586–90. doi: 10.1590/S0100-736X2013000500006
70. Pfukenyi, DM, Mukaratirwa, S, Willingham, AL, and Monrad, J. Epidemiological studies of parasitic gastrointestinal nematodes, cestodes and coccidia infections in cattle in the highveld and lowveld communal grazing areas of Zimbabwe. Onderstepoort J Vet Res. (2007) 74:129–42. doi: 10.4102/ojvr.v74i2.132
71. Gunathilaka, N, Niroshana, D, Amarasinghe, D, and Udayanga, L. Prevalence of Gastrointestinal Parasitic Infections and Assessment of Deworming Program among Cattle and Buffaloes in Gampaha District, Sri Lanka. Biomed Res Int. (2018) 2018:3048373–10. doi: 10.1155/2018/3048373
72. Makau, DN, Van Leeuwen, JA, Gitau, GK, Muraya, J, McKenna, SL, Walton, C, et al. Animal and management factors associated with weight gain in dairy calves and heifers on smallholder dairy farms in Kenya. Prev Vet Med. (2018) 161:60–8. doi: 10.1016/j.prevetmed.2018.10.017
73. Waruiru, RM, Kyvsgaard, NC, Thamsborg, SM, Nansen, P, Bøgh, HO, Munyua, WK, et al. The Prevalence and Intensity of Helminth and Coccidial Infections in Dairy Cattle in Central Kenya. Vet Res Commun. (2000) 24:39–53. doi: 10.1023/A:1006325405239
74. Maichomo, MW, Kagira, JM, and Walker, T. The point prevalence of gastro-intestinal parasites in calves, sheep and goats in Magadi division, south-western Kenya. Onderstepoort J Vet Res. (2004) 71:257–61. doi: 10.4102/ojvr.v71i4.229
75. Vasileiou, NGC, Fthenakis, GC, and Papadopoulos, E. Dissemination of parasites by animal movements in small ruminant farms. Vet Parasitol. (2015) 213:56–60. doi: 10.1016/j.vetpar.2015.04.031
76. Rose, H, Hoar, B, Kutz, SJ, and Morgan, ER. Exploiting parallels between livestock and wildlife: Predicting the impact of climate change on gastrointestinal nematodes in ruminants. Int J Parasitol Parasites Wildl. (2014) 3:209–19. doi: 10.1016/j.ijppaw.2014.01.001
77. Craig, TM. Helminth parasites of the ruminant gastrointestinal tract In: Current Veterinary Therapy: Food Animal Practice (2009). 78–91.
78. Fávero, FC, Buzzulini, C, Cruz, BC, Felippelli, G, Maciel, WG, Salatta, B, et al. Experimental infection of calves with Haemonchus placei and Haemonchus contortus: Assessment of parasitological parameters. Vet Parasitol. (2016) 217:25–8. doi: 10.1016/J.VETPAR.2015.12.017
79. Mohammedsalih, KM, Krücken, J, Bashar, A, Juma, FR, Abdalmalaik, AAH, Khalafalla, A, et al. Susceptible trichostrongyloid species mask presence of benzimidazole-resistant Haemonchus contortus in cattle. Parasit Vectors. (2021) 14:101–16. doi: 10.1186/s13071-021-04593-w
80. Delano, ML, Mischler, SA, and Underwood, WJ. Biology and diseases of ruminants: sheep, goats, and cattle In: Laboratory Animal Medicine (2002). 519–614.
81. Bhat, AH, Tak, H, Malik, IM, Ganai, BA, and Zehbi, N. Trichostrongylosis: A zoonotic disease of small ruminants. J Helminthol. (2023) 97:e26–11. doi: 10.1017/S0022149X2300007X
82. Gutierrez, Y. Other Tissue Nematode Infections. Tropical Infectious Diseases: Principles, Pathogens, & Practice, 2-Volume Set with CD-ROM (2005):1231–47. doi: 10.1016/B978-0-443-06668-9.50111-3,
83. Mehlhorn, H. Trichostrongylosis In: Animals BT-Encyclopedic Reference of Parasitology: Diseases, Treatment, Therapy. Berlin, Heidelberg: Springer Berlin Heidelberg (2001). 616–7.
84. Gholami, S, Babamahmoodi, F, Abedian, R, Sharif, M, Shahbazi, A, Pagheh, A, et al. Trichostrongylus colubriformis: possible most common cause of human infection in Mazandaran Province, North of Iran. Iran J Parasitol. (2015) 10:110–5.
85. Phosuk, I, Intapan, PM, Sanpool, O, Janwan, P, Thanchomnang, T, Sawanyawisuth, K, et al. Molecular Evidence of Trichostrongylus colubriformis and Trichostrongylus axei Infections in Humans from Thailand and Lao PDR. Am J Trop Med Hyg. (2013) 89:376–9. doi: 10.4269/AJTMH.13-0113
86. Boreham, RE, McCowan, MJ, Ryan, AE, Allworth, AM, and Robson, JMB. Human trichostrongyliasis in Queensland. Pathology. (1995) 27:182–5. doi: 10.1080/00313029500169842
87. Obanda, V, Maingi, N, Muchemi, G, Ng’Ang’A, CJ, Angelone, S, and Archie, EA. Infection dynamics of gastrointestinal helminths in sympatric non-human primates, livestock and wild ruminants in Kenya. PLoS One. (2019) 14:e0217929. doi: 10.1371/journal.pone.0217929
88. Betson, M, Alonte, AJI, Ancog, RC, Aquino, AMO, Belizario, VY Jr, Bordado, AMD, et al. Zoonotic transmission of intestinal helminths in southeast Asia: Implications for control and elimination. Adv Parasitol. (2020) 108:47–131. doi: 10.1016/bs.apar.2020.01.036
89. Bundy, DAP, Appleby, LJ, and Brooker, SJ. Nematodes limited to the intestinal tract (Enterobius vermicularis, Trichuris trichiura, Capillaria philippinensis, and Trichostrongylus spp.) In: Hunter's Tropical Medicine and Emerging Infectious Diseases (2020). 834–9.
90. Ngongeh, LA. Variation in faecal worm egg counts of experimentally infected goats and mice with time of day and its implications in diagnosis of helminthosis. J Parasit Dis Off Organ Indian Soc Parasitol. (2017) 41:997–1000. doi: 10.1007/s12639-017-0924-4
91. Odoi, A, Gathuma, JM, Gachuiri, CK, and Omore, A. Risk factors of gastrointestinal nematode parasite infections in small ruminants kept in smallholder mixed farms in Kenya. BMC Vet Res. (2007) 3:6–11. doi: 10.1186/1746-6148-3-6
92. Sherwin, V, Hyde, R, Green, M, Remnant, J, Payne, E, and Down, P. Accuracy of heart girth tapes in the estimation of weights of pre-weaned calves. Vet Rec Open. (2021) 8:e16. doi: 10.1002/vro2.16
93. Lukuyu, MN, Gibson, JP, Savage, DB, Duncan, AJ, Mujibi, FDN, and Okeyo, AM. Use of body linear measurements to estimate liveweight of crossbred dairy cattle in smallholder farms in Kenya. Springerplus. (2016) 5:63–14. doi: 10.1186/s40064-016-1698-3
94. Ndung’u, PW, Bebe, BO, Ondiek, JO, Butterbach-Bahl, K, Merbold, L, and Goopy, JP. Improved region-specific emission factors for enteric methane emissions from cattle in smallholder mixed crop: livestock systems of Nandi County, Kenya. Anim Prod Sci. (2018) 59:1136–46. doi: 10.1071/an17809
95. Stear, M, Preston, S, Piedrafita, D, and Donskow-Łysoniewska, K. The Immune Response to Nematode Infection. Int J Mol Sci. (2023) 24:2283. doi: 10.3390/ijms24032283
96. Creutzinger, KC, Pempek, JA, Locke, SR, Renaud, DL, Proudfoot, KL, George, K, et al. Dairy producer perceptions toward male dairy calves in the Midwestern United States. Frontiers. Anim Sci. (2022) 3:1–13. doi: 10.3389/fanim.2022.1000897
97. Berk, Z, Bishop, SC, Forbes, AB, and Kyriazakis, I. A simulation model to investigate interactions between first season grazing calves and Ostertagia ostertagi. Vet Parasitol. (2016) 226:198–209. doi: 10.1016/j.vetpar.2016.05.001
98. Blowey, RW, and Weaver, AD. Alimentary disorders In: Color Atlas of Diseases and Disorders of Cattle (2011). 53–82.
99. Can-Celis, A, Mancilla-Montelongo, G, Castañeda-Ramírez, GS, Chan-Pérez, JI, and de Torres-Acosta, JF. Isolation of pure Trichostrongylus colubriformis strains from naturally infected sheep using two methodologies. Vet Parasitol Reg Stud Reports. (2020) 22:100474. doi: 10.1016/j.vprsr.2020.100474
100. Wangboon, C, Martviset, P, Jamklang, M, Chumkiew, S, Penkhrue, W, Rangdist, S, et al. Microscopic and molecular epidemiology of gastrointestinal nematodes in dairy and beef cattle in Pak Chong district, Nakhon Ratchasima province, Thailand. Vet World. (2024) 17:1035–43. doi: 10.14202/vetworld.2024.1035-1043
101. Kouam, MK, Fokom, GT, Luogbou, DN, and Kantzoura, V. Gastro-intestinal parasitism and control practices in dairy cattle in North-west Cameroon (Central Africa). Acta Parasitol. (2021) 66:947–53. doi: 10.1007/s11686-021-00343-1
102. Tibebu, A, Tamiru, Y, and Abdeta, D. Prevalence of Major Gastrointestinal Nematode and Degree of Parasite Infestation in Sheep of Bako Agricultural Research Center Community Based Breeding Program Project Small Holder Farms at Horro District. J Dairy Vet Sci. (2018) 8:1–12. doi: 10.19080/jdvs.2018.08.555740
103. Karvonen, A, Jokela, J, and Laine, AL. Importance of sequence and timing in parasite coinfections. Trends Parasitol. (2019) 35:109–18. doi: 10.1016/J.PT.2018.11.007
104. Thumbi, SM, Bronsvoort, BMDC, Poole, EJ, Kiara, H, Toye, PG, Mbole-Kariuki, MN, et al. parasite co-infections and their impact on survival of indigenous cattle. PLoS One. (2014) 9:e76324. doi: 10.1371/journal.pone.0076324
105. Hananeh, WM, Radhi, A, Mukbel, RM, and Ismail, ZB. Effects of parasites coinfection with other pathogens on animal host: A literature review. Vet World. (2022) 15:2414–24. doi: 10.14202/vetworld.2022.2414-2424
106. Kumar, N, Rao, TKS, Varghese, A, and Rathor, VS. Internal parasite management in grazing livestock. J Parasit Dis Off Organ Indian Soc Parasitol. (2012) 37:151–7. doi: 10.1007/s12639-012-0215-z
107. Avramenko, RW, Redman, EM, Windeyer, C, and Gilleard, JS. Assessing anthelmintic resistance risk in the post-genomic era: a proof-of-concept study assessing the potential for widespread benzimidazole-resistant gastrointestinal nematodes in North American cattle and bison. Parasitology. (2020) 147:897–906. doi: 10.1017/S0031182020000426
108. Avramenko, RW, Redman, EM, Melville, L, Bartley, Y, Wit, J, Queiroz, C, et al. Deep amplicon sequencing as a powerful new tool to screen for sequence polymorphisms associated with anthelmintic resistance in parasitic nematode populations. Int J Parasitol. (2019) 49:13–26. doi: 10.1016/j.ijpara.2018.10.005
Keywords: gastrointestinal nematodes, deep amplicon sequencing, risk factor, co-infections, dairy calves, Kenya
Citation: Cheptoo S, Yalcindag E, González Gordon L, Rukwaro B, Kimatu JS, Wasonga J, Karani BE, Ndambuki G, Migeni S, Kagai J, Kiprotich LE, Saya N, Vasoya D, Nangekhe G, Onguso J, Mungai G, Bronsvoort BM and Cook EAJ (2025) Species diversity and risk factors of gastrointestinal nematodes in smallholder dairy calves in Kenya. Front. Vet. Sci. 12:1588350. doi: 10.3389/fvets.2025.1588350
Edited by:
Rudi Cassini, University of Padua, ItalyReviewed by:
Bersissa Kumsa, Addis Ababa University, EthiopiaQingxia Wu, Tibet Agricultural and Animal Husbandry University, China
Copyright © 2025 Cheptoo, Yalcindag, González Gordon, Rukwaro, Kimatu, Wasonga, Karani, Ndambuki, Migeni, Kagai, Kiprotich, Saya, Vasoya, Nangekhe, Onguso, Mungai, Bronsvoort and Cook. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Sylvia Cheptoo, c3lsdmlhY2hlcHRvb2tvcmlyQGdtYWlsLmNvbQ==; Elizabeth Anne Jessie Cook, ZS5jb29rQGNnaWFyLm9yZw==