 Gábor Csitári1
Gábor Csitári1 Nikoletta Such1
Nikoletta Such1 László Menyhárt2Kornél Schermann1Kornél Humpok3
László Menyhárt2Kornél Schermann1Kornél Humpok3 Valéria Farkas1
Valéria Farkas1 László Pál1
László Pál1 Károly Dublecz1*
Károly Dublecz1*- 1Department of Animal Nutrition and Nutritional Physiology, Institute of Physiology and Nutrition, Georgikon Campus, Hungarian University of Agriculture and Life Sciences, Keszthely, Hungary
- 2Department of Applied Statistics, Institute of Mathematics and Basic Sciences, Budai Campus, Hungarian University of Agriculture and Life Sciences, Budapest, Hungary
- 3Gallus Ltd. of Poultry Breeding and Hatching, Devecser, Hungary
Floor eggs represent a significant loss for broiler breeder farms and hatcheries due to the increased risk of bacterial contamination of embryos, the hatchery environment, and newly hatched chicks. In this trial, the effects of litter exposure duration (3, 6, and 16 h) and breeder flock age (22–23 weeks vs. 65–66 weeks) were evaluated in terms of hatchability and the microbial composition of different egg components (eggshell, egg membrane, and egg content). The number of total culturable aerobic microbes on the eggshell surface increased following litter exposure and decreased after 3 h. Hatchability, however, was significantly reduced only after 16 h of exposure, primarily due to increased embryonic mortality. Litter exposure and flock age led to significant differences in microbiota composition, but only on the eggshell surface. At the phylum level, 16 h of litter exposure significantly increased the abundance of Firmicutes and decreased that of Proteobacteria. At the genus level, litter exposure resulted in increased relative abundances of Lactobacillus and Ruminococcus torques group and a decreased abundance of Staphylococcus. The microbiota of the egg membrane and egg content were similar at the phylum level; however, notable differences were observed at the genus level. Pseudomonas was dominant in the egg membrane but underrepresented in the egg content, leading to a significantly higher abundance of spoilage-associated bacterial genera in the membrane than in the content. Interestingly, the genus Flexivirga (phylum Actinobacteria) was detected in high abundance in both the egg membrane and egg content, despite not having been previously reported inside eggs. According to the results, no measurable bacterial translocation from the litter into the internal egg structures was observed. However, the frequency of floor egg collection may represent a critical factor if such eggs are intended for hatching.
1 Introduction
In hatching egg production, floor eggs represent a significant source of loss for broiler breeder farms and pose considerable risks of bacterial contamination within the hatchery. A floor egg refers to an egg laid outside the designated nest area, typically in manure. Several recent studies have confirmed that floor eggs generally exhibit reduced hatchability compared to conventional nest eggs (1–3). However, in some cases, increased microbial contamination has not resulted in lower hatchability (4), and the eggshell microbiota may even serve as a protective barrier against pathogens (5).
The eggshell microbiota typically comprises microorganisms originating from both the hen and the surrounding litter (6). Commonly represented bacterial phyla on the eggshell include Firmicutes, Bacteroidetes, and Actinobacteria, while Proteobacteria are often underrepresented. Dominant genera include Bacteroides, Blautia, Clostridium, Faecalibacterium, Lactobacillus, Megamonas, and Oscillibacter (5–9).
Poultry litter is a complex matrix comprising bedding materials, chicken excreta, spoiled feed, feathers, and other environmental elements (10–12). It harbors a dynamic microbiome influenced by various factors, such as the animals’ gut microbiota, manure dry matter content, ventilation, and environmental temperature and humidity (10, 13, 14). Litter amendments, such as acidifiers, alkalizers, charcoal, and gypsum, can also modify the microbiome (15). Several studies have explored interactions between litter and the gastrointestinal microbiome, as well as the occurrence of pathogens (14, 16). This interaction is relevant in both broilers and layers, as the gut microbiota can affect meat and egg safety. Ingestion of manure by young chicks is considered to play a key role in the development of the gut microbiota (10). Despite ongoing research, the interaction between the litter microbiome and the host remains largely unexplored (17).
Egg spoilage can result from microbial contamination, particularly when bacteria penetrate the eggshell and proliferate in the albumen or yolk. Spoilage-associated genera reported in the literature include Alcaligenes, Enterococcus, Escherichia/Shigella, Proteus, Pseudomonas, Staphylococcus, and Streptococcus (18–20). Freshly laid eggs are particularly vulnerable due to their moist surface. Although eggs possess natural defense mechanisms, Gram-negative bacteria such as Pseudomonas, Alcaligenes, and Salmonella are among the most common invaders (21). To prevent trans-shell bacterial penetration, hatching eggs are typically sanitized, significantly reducing microbial loads (4, 7, 22). Post-sanitization, the remaining eggshell microbiota has minimal impact on the developing embryo. However, some spore-forming bacteria, resistant to disinfection and oxygen stress, may survive and colonize chicks during hatching (7, 23).
Lower hatchability of floor eggs may also result from prolonged contact with unsuitable litter conditions. These eggs are typically collected after a delay and cooled later, exposing them to inappropriate temperature and moisture, which negatively affects quality. Hen age is another important factor influencing egg weight and hatchability (24, 25). Older hens tend to lay larger eggs with thinner shells, and their eggs often exhibit reduced hatchability (26–28). However, some studies report conflicting results; for example, Roque and Soares (29) found that hatchability and viability were lower in younger flocks due to increased early and late embryonic mortality. Pre-hatch storage time and environmental temperature also impair hatchability in an age-dependent manner (30, 31).
Eggshell microbial integrity is similarly influenced by hen age. As shell quality declines with age, the risk of microbial penetration increases (32). The total bacterial load on the eggshell surface has been shown to increase with flock age (33). Moreover, the intestinal microbiota of layer hens has been experimentally associated with egg quality (34). Despite controlling for breed, age, and environment, large variations in hatchability between flocks are frequently observed (35), suggesting the existence of other, currently unknown influencing factors.
Microorganisms can also enter the egg internally via the hen’s reproductive tract (36, 37), which is not sterile (37–39) and shares similarities with the gut microbiome (39). Dominant phyla in the reproductive tract include Proteobacteria, Firmicutes, Actinobacteria, and Bacteroidetes, with their abundances influenced by factors such as growth rate (37, 38, 40–42). It is hypothesized that bacteria from the reproductive tract may translocate into the forming egg, becoming localized between the chorion and inner membrane or even within the developing embryo (37, 43).
The present study aimed to investigate whether the time eggs spend in the litter and the age of breeder flocks influence hatchability and the microbial composition of various egg components (eggshell, inner membrane, and egg content). Special attention was given to the inner egg membrane, as it may serve as the first barrier to bacterial invasion through the shell. Our findings aim to provide practical insights for broiler breeder farms and hatcheries regarding the risks and viability of using floor eggs for hatching.
2 Materials and methods
2.1 Egg collection and hatching
A total of 1,440 hatching eggs (720 eggs per flock) were collected from two Ross 308 parent flocks of different ages—22–23 weeks and 65–66 weeks—from separate farms operated by Gallus Ltd. (Bakonypölöske and Oroszi, Hungary). Eggs were transported to the Gallus Ltd. hatchery in Devecser, Hungary, using air-conditioned vehicles to maintain egg quality during transit.
To simulate floor egg conditions and assess the impact of litter exposure duration, four treatment groups were established: Control group: Conventional nest eggs collected from the collecting belt during the second egg collection of the day; these eggs were freshly laid and not contaminated with litter. Litter treatment groups: Eggs subjected to 3-, 6-, or 16-h exposure to poultry litter. The eggs used for litter treatments were originally nest eggs and manually placed in litter within different areas of the barn. Each egg was gently mixed with the litter once per hour using sterile gloves to mimic natural floor egg conditions. To prevent hen contact, plastic baskets were placed over the eggs. After the designated exposure periods (3, 6, or 16 h), eggs were collected and stored at 16–18°C and 75–80% relative humidity until hatching.
The distribution of eggs across treatment groups was as follows: control and 16-h treatment: 185 eggs each, 3-h and 6-h treatments: 180 eggs each. The discrepancy in sample size reflects the design of the microbiota analysis: only the control and 16-h groups were subjected to next-generation sequencing (NGS). For microbiological analysis, five eggs from each treatment group (per farm) were transported to the laboratory for culturable microbiota assessment. An additional five eggs from the control and 16-h groups (per farm) were sent for NGS-based microbial community analysis, focusing on the eggshell, inner membrane, and egg content. The remaining 175 eggs per treatment group were incubated at the Gallus Ltd. hatchery. All eggs were incubated under identical conditions according to the standard procedures of the hatchery. Prior to incubation, eggs were sanitized using formaldehyde fumigation at a concentration of 7 g/m3 paraformaldehyde.
Hatching commenced on day 7 post-laying in Petersime BioStreamer 24S pre-hatching machines. Environmental parameters, including temperature, relative humidity, CO₂ concentration, and egg turning, were automatically regulated based on the manufacturer’s standard hatching program.
On day 18 of incubation, candling was performed to identify and remove infertile, dead, damaged, or rotten eggs. Fertile eggs were then vaccinated in ovo against infectious bursal disease (IBD) using an automated injection system (Embrex Inovoject, Zoetis Inc., New York, United States). Post-vaccination, eggs were automatically transferred to hatcher trays, and incubation continued under controlled conditions. Hatchability was calculated based on both total and fertile eggs. Embryonic mortality was expressed as a percentage of fertile eggs.
2.2 Microbiological evaluation of eggshells by culturing
Microbiological analysis of the eggshells was conducted at the Microbiological Laboratory of Gallus Ltd. (Devecser, Hungary). A total of 40 eggs were analyzed, comprising 20 eggs from the younger flock and 20 from the older flock. Each litter treatment group (0, 3, 6, and 16 h) included five replicate eggs per age group. All microbiological assessments were performed prior to hatchery sanitization.
Sample preparation followed the ISO 6887-4:2017 standard. The following parameters were assessed: total aerobic bacterial counts, determined using ISO 4833-1:2014, and coliform counts, determined using ISO 21528-2:2017. For culturing, plate count agar (PCA) was used for total aerobic bacteria, while violet Red Bile Glucose (VRBG) agar was used for coliform enumeration.
2.3 DNA sequencing and microbial community analysis
For DNA sequencing, a total of 20 eggs were collected, 10 from the younger and 10 from the older flock. Only two treatment groups were analyzed: control nest eggs and eggs exposed to 16-h litter treatment, each in five replicates per age group.
2.3.1 Sample preparation
Eggshells were washed using 1.2 mL DNA/RNA Shield solution (Zymo Research, CA, USA), and then transferred into DNA/RNA shield lysis tubes for DNA extraction. For the inner egg membrane, the eggs were washed with detergent, surface-sterilized three times by flaming with ethanol, and then aseptically broken. The inner membrane was collected using a sterile scalpel.
For the egg content, a 150 μL mixture of egg white and yolk was sampled using a sterile pipette.
2.3.2 DNA extraction and quantification
DNA was extracted from the eggshell, egg membrane and egg content samples using the ZymboBIOMICS 96 MagBead DNA Kit with ZR BashingBed Lysis Tubes (Zymo Research, CA, USA). DNA concentration was measured with a Qubit 3.0 Fluorometer using the Qubit dsDNA HS Assay Kit (Thermo Fisher Scientific, Waltham, MA, United States).
2.3.3 Library preparation and sequencing
The bacterial 16S rRNA V3–V4 region was amplified using tagged primers. PCR amplification and library purification followed Illumina’s 16S Metagenomic Sequencing Library Preparation protocol (44). Libraries were validated using High Sensitivity D1000 ScreenTape on a TapeStation 2,200 system (Agilent Technologies, Santa Clara, CA, United States). Equimolar pooled libraries were sequenced on the Illumina MiSeq platform with a MiSeq Reagent Kit v3 (600 cycles) using a 300-bp paired-end read format.
The raw sequence data were deposited in the NCBI Sequence Read Archive (SRA) under BioProject ID PRJNA1012860.
2.3.4 Data analysis
Using next-generation sequencing results, the relative abundance of egg spoilage-associated bacterial genera was calculated. This included the sum of reads assigned to the following genera: Alcaligenes, Enterococcus, Escherichia/Shigella, Proteus, Pseudomonas, Staphylococcus, and Streptococcus.
2.4 Bioinformatics and statistical analyses
Bacterial communities were identified by analyzing the V3–V4 region of the 16S rRNA gene using the Illumina MiSeq platform. Sequence data were processed using QIIME2 (Quantitative Insights Into Microbial Ecology 2), version 2020.2 (45).
2.4.1 Sequence processing and OTU clustering
Raw sequences were quality-filtered based on Phred quality scores and the presence of ambiguous bases using the quality-filter q-score plugin. Denoising was performed using the Deblur method (deblur denoise-16S), and representative sequences were identified using a 16S reference as a positive filter. Sequences were clustered into operational taxonomic units (OTUs) using the VSEARCH open-reference algorithm, with a 97% similarity threshold against the SILVA reference database (release 132) (46).
2.4.2 Diversity analyses
Alpha diversity metrics—including species richness, Chao1, Shannon, and Simpson indices—were calculated using the qiime2-diversity plugin. Beta diversity was assessed using Bray–Curtis dissimilarity. All diversity analyses were conducted after rarefaction to 1,000 sequences per sample. Supplementary analysis was performed using MicrobiomeAnalyst (47).
2.4.3 Statistical analysis
A two-way ANOVA was conducted to assess the effects of litter treatment and flock age on microbiota culturing and hatchability. A three-way ANOVA was used to analyze OTU numbers, alpha diversity metrics, and the relative abundance of spoilage-associated bacterial genera, with egg part, litter treatment, and flock age as the main factors.
All ANOVA analyses were performed using R software (48).
2.4.4 Taxonomic abundance testing
Relative abundance data at different taxonomic levels were analyzed using the Aligned Rank Transform (ART) for non-parametric factorial analysis, implemented via the ARTool R package (49). This method enables valid testing of main effects and interactions in non-parametric factorial designs by aligning and rank-transforming the data, followed by standard ANOVA on the aligned ranks.
For each taxon, the primary effects of litter treatment, flock age, and their interaction were tested separately for each egg part (eggshell, egg membrane, and egg content). To correct for multiple testing, p-values were adjusted using the Benjamini–Hochberg False Discovery Rate (BH-FDR) procedure (50). A BH-adjusted p-value < 0.05 was considered statistically significant.
The ART procedure has been validated as a robust and suitable alternative for microbiome data analysis in designs with complex interactions and non-normal distributions (51, 52).
3 Results
3.1 Egg weight and culturable microbes of the eggshell
The weight of the eggs ranged from 49.2 to 77.0 g, with eggs from the older breeder flock being significantly heavier, as expected (Table 1). The total aerobic bacterial counts on the eggshells increased proportionally with the duration of litter exposure, and all litter-treated groups showed significantly higher counts compared to the control group (Table 1). The age of the breeder flock also influenced bacterial load: eggs from the younger hens exhibited higher aerobic bacterial counts on their shells across all treatments. In contrast, coliform bacteria were not detected on any of the eggshells, remaining below the detection limit of 1 × 102 CFU/egg in all cases.
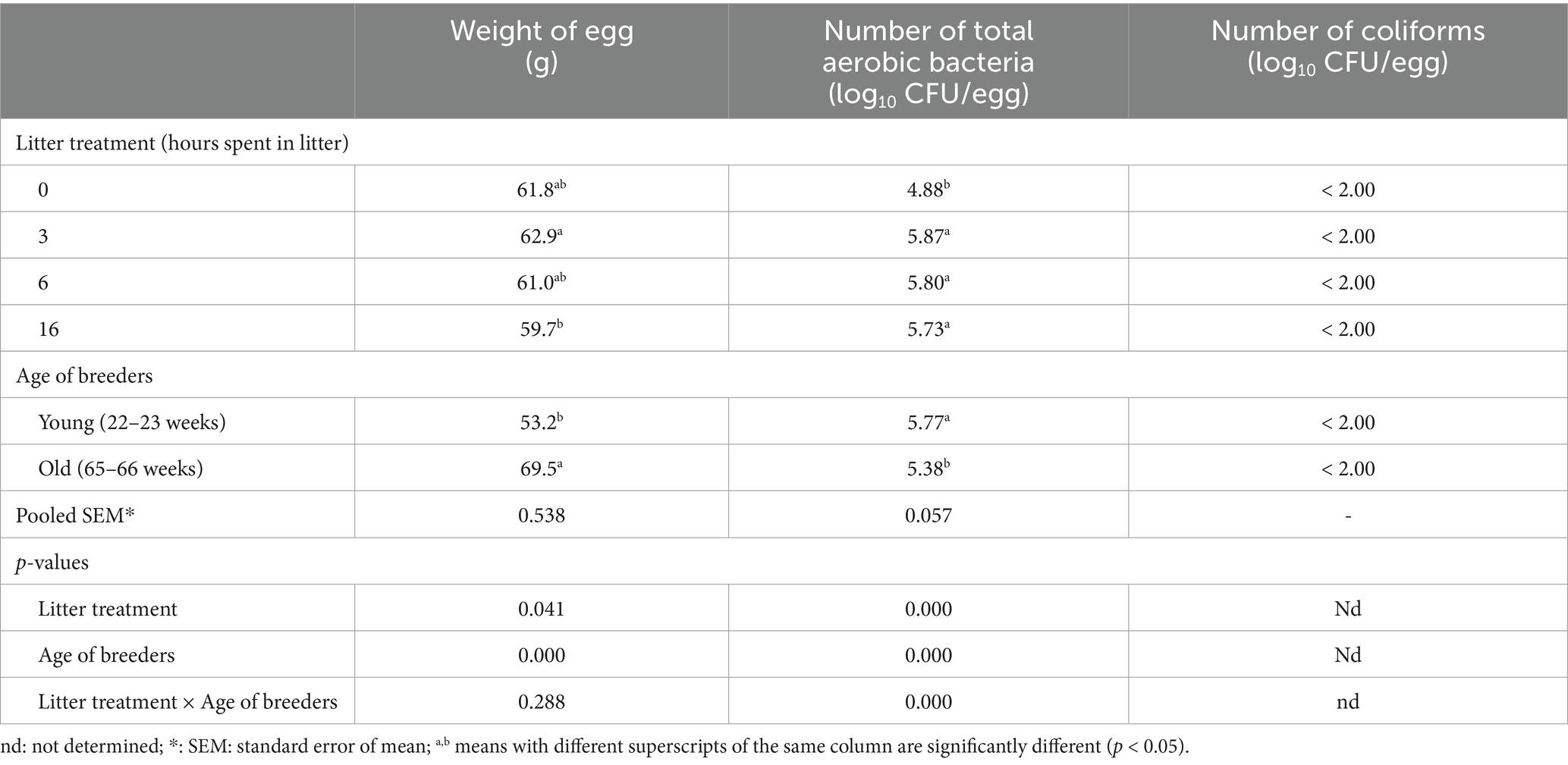
Table 1. The effect of litter treatments and the age of laying hens on the egg weight and the number of culturable microbes of eggshell (n = 10).
3.2 Hatching parameters
Fertility rates and the proportion of damaged eggs were not significantly affected by litter treatment duration (Table 2). However, exposure to litter for 16 h resulted in a significant increase in embryonic mortality and a decrease in hatchability compared to the 3- and 6-h litter exposure groups. Additionally, eggs from the younger breeder flock exhibited a significantly higher hatchability rate relative to the total number of incubated eggs (Table 2).
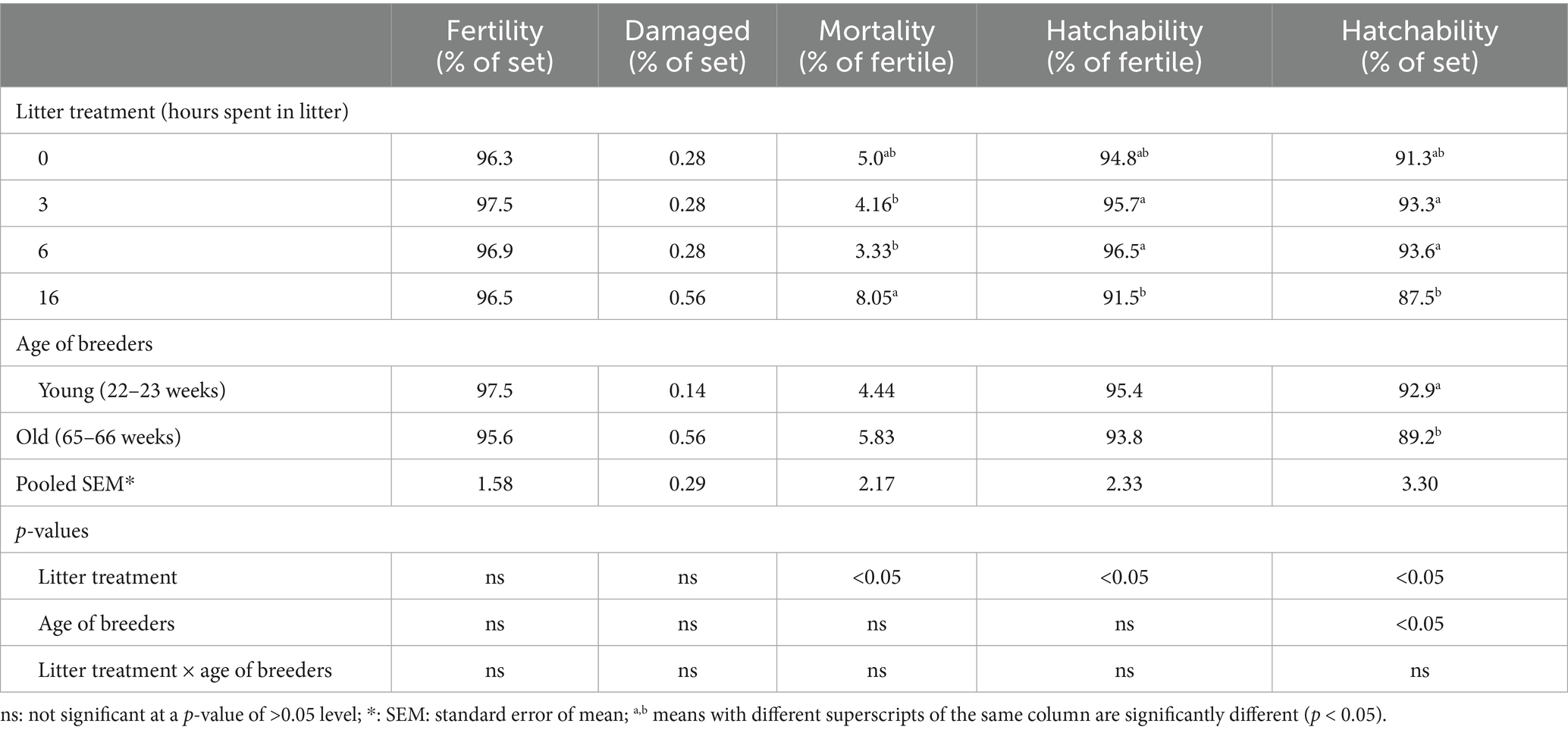
Table 2. The effect of litter treatments and the age of laying hens on the hatching parameters (n = 175).
3.3 Results of next-generation sequencing
In this study, a total of 1,207,766 good-quality 16S rRNA reads were obtained for analysis from all 40 samples after quality filtering. The overall average sequence numbers were 20,129 (minimum: 10,845; maximum: 31,174). When separated by egg parts, the average sequence numbers were 19,020 (minimum: 15,605; maximum: 26,592) on the eggshell, 19,197 (minimum: 15,151; maximum: 24,610) in the inner membrane, and 22,172 (minimum: 10,845; maximum: 31,174) in the egg content. These sequences were assigned to 2,643 OTUs at 97% similarity using the open approach. A total of 1,498 OTUs remained after the data filtering step in MicobiomeAnalyst.
3.3.1 Alpha and beta diversities
All alpha diversity indices were significantly influenced by the egg component (eggshell, inner membrane, and egg content). The eggshell microbiome exhibited the highest OTU count and Chao1 index, followed by the inner membrane and then the egg content (Table 3). For the Shannon and Simpson indices, the 16-h litter exposure significantly reduced microbial alpha diversity. However, due to significant interactions between the main factors, these effects cannot be interpreted independently. Breeder age had no significant impact on any of the diversity indices.
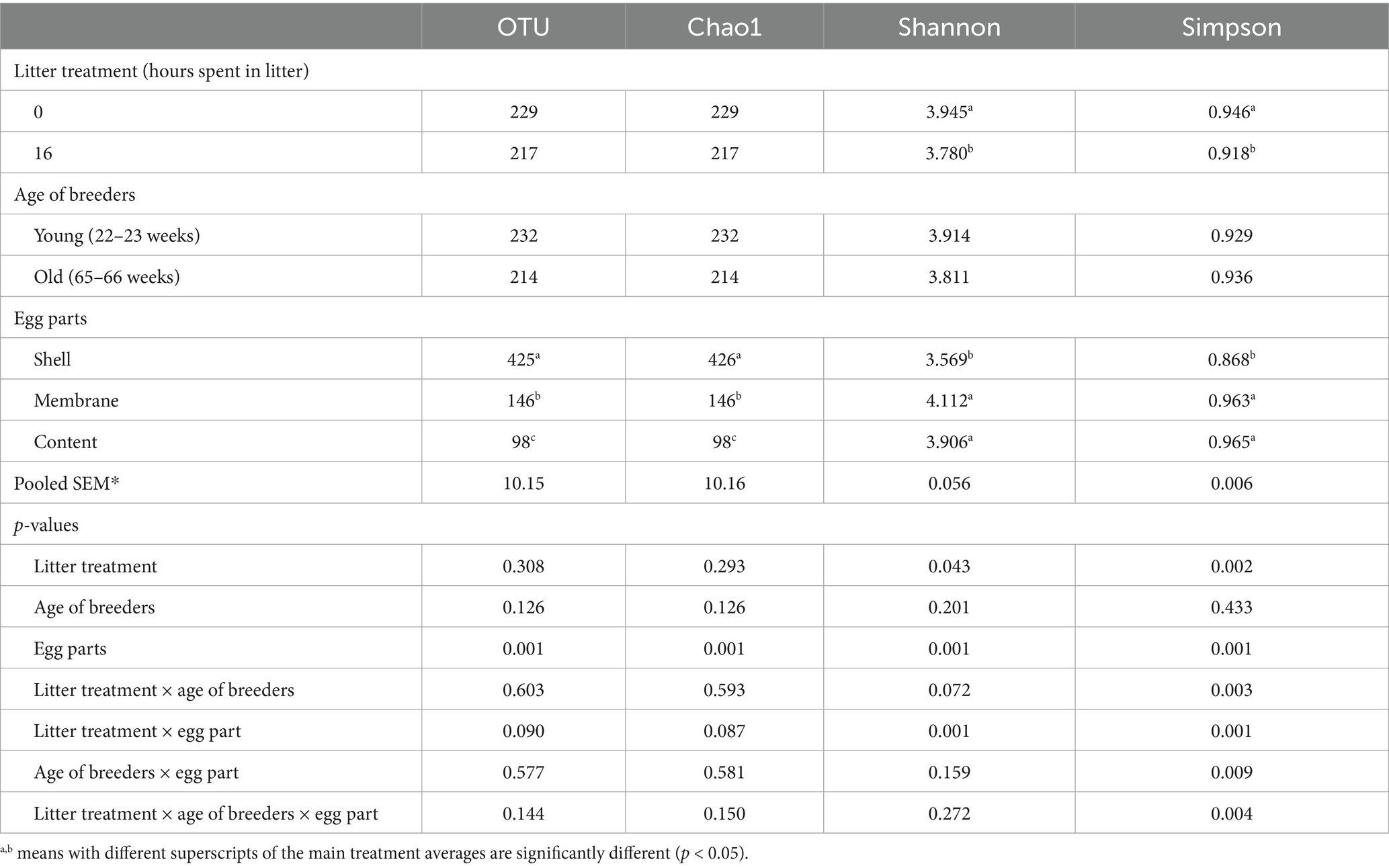
Table 3. Treatment effects on the OTU numbers and bacterial alpha diversity indices (n = 15 for litter treatment; n = 5).
Beta diversity analysis was performed to assess differences in microbial community composition among the samples (Figure 1). Principal coordinate analysis (PCoA) based on the Bray–Curtis dissimilarity matrix revealed significant clustering by egg part (PERMANOVA, R2 = 0.39, p = 0.001), with the microbial communities of eggshells distinctly separated from those of the inner membrane and egg content (Figure 1A). The inner membrane and egg content microbiota exhibited high similarity. The effect of hen age showed a trend toward separation (R2 = 0.03, p = 0.058) (Figure 1B), whereas litter treatment had no significant impact on microbial community structure, as indicated by a considerable overlap between litter-treated and untreated eggs (Figure 1C).
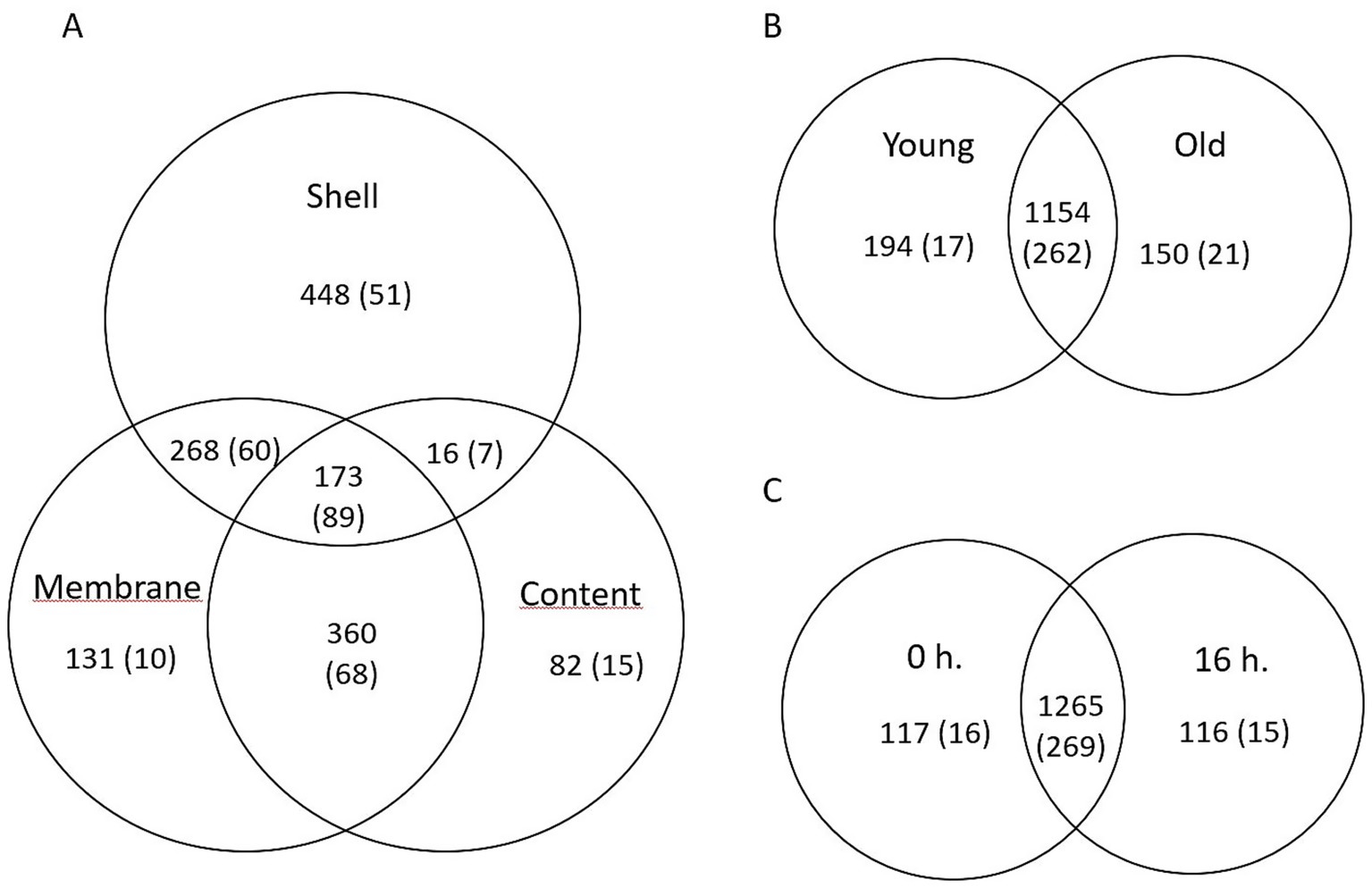
Figure 1. Principal coordinate analysis (PCoA) based on the Bray–Curtis dissimilarity matrix depicting bacterial community composition as influenced by (A) egg parts, (B) breeder age, and (C) litter treatment. Statistical differences in beta diversity were assessed using permutational multivariate analysis of variance (PERMANOVA).
1A: [PERMANOVA] F-value: 17.964; R-squared: 0.38662; p-value: 0.001.
1B: [PERMANOVA] F-value: 1.8807; R-squared: 0.031407; p-value: 0.058.
1C: [PERMANOVA] F-value: 0.77102; R-squared: 0.013119; p-value: 0.607.
3.3.2 Taxonomic microbiota composition of eggshell
Four bacterial phyla—Firmicutes, Bacteroidetes, Actinobacteria, and Proteobacteria—were detected on the eggshell at relative abundances exceeding 1%, with Firmicutes being the dominant phylum. The relative abundance of these phyla was significantly influenced by both litter treatment and the age of the hens (Table 4). Specifically, litter treatment increased the abundance of Firmicutes and decreased that of Proteobacteria on the eggshell compared to nest eggs. Eggs from older flocks exhibited significantly lower frequencies of Firmicutes and Bacteroidetes, while Actinobacteria abundance was approximately two-fold higher compared to eggs from younger flocks. At the genus level, Staphylococcus, Lactobacillus, and Salinicoccus were the most abundant on the eggshell surface (Supplementary Table S1). Litter treatment significantly reduced the abundance of Staphylococcus and increased that of Lactobacillus and the Ruminococcus_torques_group. Additionally, breeder flock age affected genus distribution; eggshells from older flocks had significantly lower proportions of Staphylococcus and Lactobacillus, but higher abundances of Salinicoccus, Brachybacterium, and Brevibacterium.
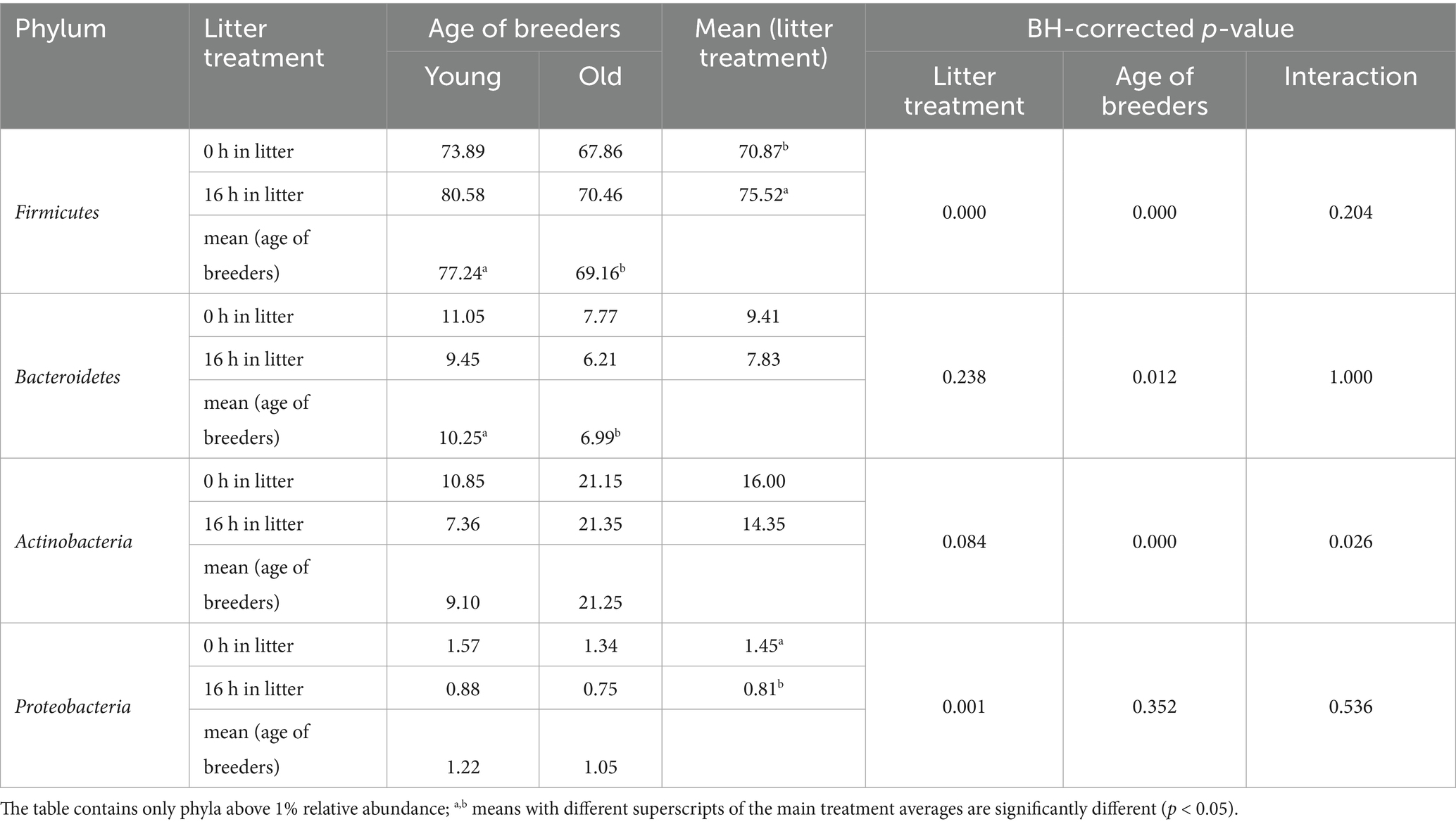
Table 4. Treatment effects on the relative abundances of bacterial phyla of the eggshell (%) (n = 5).
3.3.3 Taxonomic microbiota composition of egg membranes
Eight bacterial phyla were detected at relative abundances exceeding 1% in the egg membranes. Among these phyla, Proteobacteria and Firmicutes were dominant, while Actinobacteria, Cyanobacteria, and Bacteroidetes each comprised more than 10% of the community (Table 5). No significant effects of litter treatment or breeder age were observed at either the phylum or genus level. The 10 most abundant genera identified in the egg membrane included Pseudomonas, Flexivirga, Staphylococcus, Paracoccus, Rhodanobacter, Enhydrobacter, Candidimonas, Chujaibacter, Ruminococcaceae_NK4A214_group, and Salinicoccus (Supplementary Table S2).
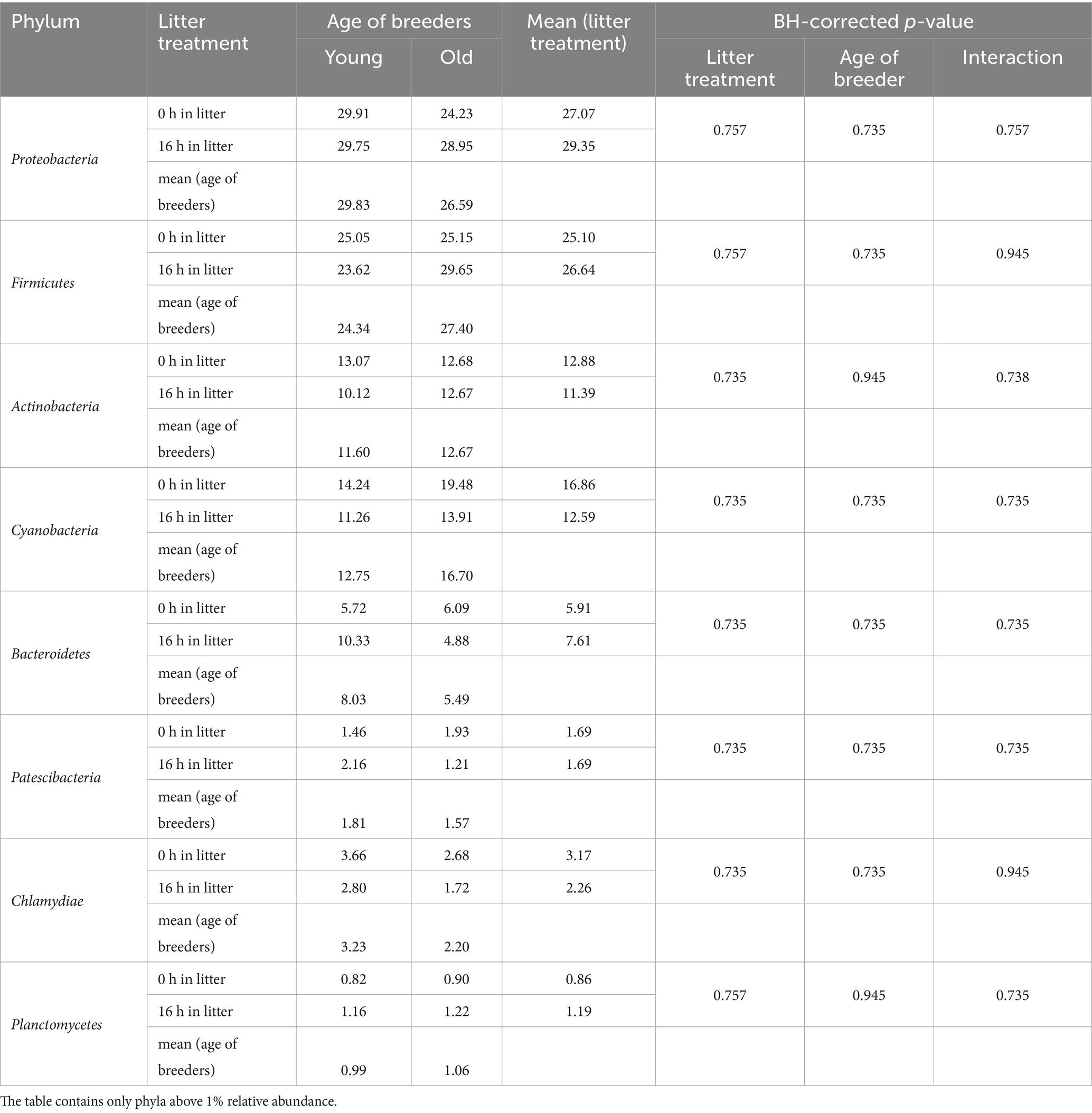
Table 5. Changes in the relative abundances of bacterial phyla in the egg membrane (%) (n = 5).
3.3.4 Taxonomic microbiota composition of egg contents
The egg content, comprising a mixture of egg yolk and egg white, comprised eight bacterial phyla with relative abundances above 1% (Table 6). Alongside the dominant Proteobacteria and Firmicutes, Actinobacteria and Cyanobacteria were present at frequencies exceeding 10%. Similar to the egg membrane, no significant differences were observed at any taxonomic level. The 10 most abundant genera in the egg content, based on relative abundance, were Flexivirga (9.39%), Rhodanobacter (3.90%), Paracoccus (3.17%), Rhodococcus (2.95%), Chujaibacter (2.62%), Alkanibacter (2.37%), Mycobacterium (1.97%), Ruminococcaceae_NK4A214_group (1.86%), Enhydrobacter (1.80%), and Lactobacillus (1.58%) (Supplementary Table S3).
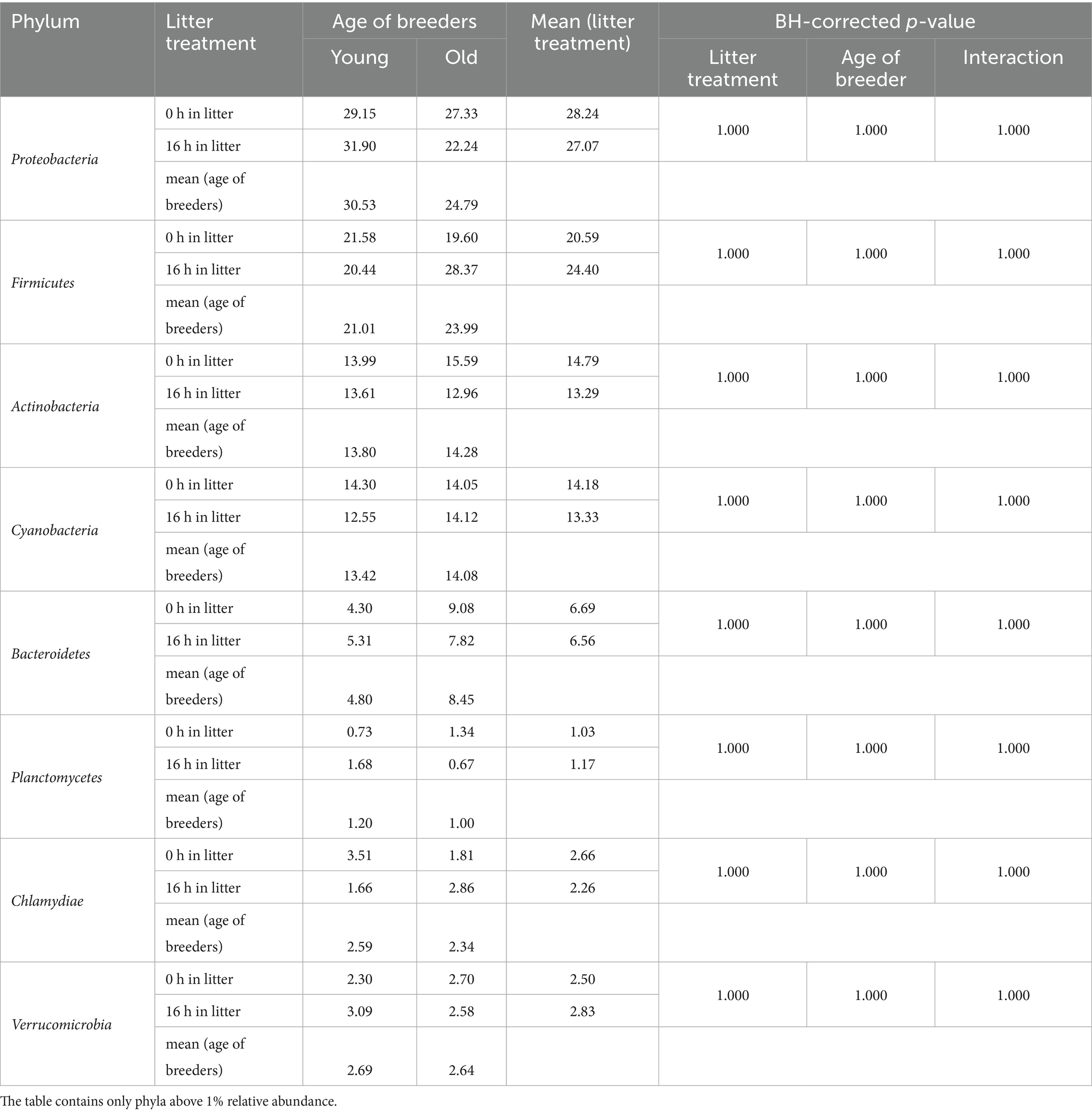
Table 6. Changes in the relative abundances of bacterial phyla in the egg content (%) (n = 5).
3.3.5 Common OTUs among eggshell, egg membrane, and egg contents
Venn diagrams were constructed to analyze the common and unique OTUs and genera across the different egg parts (Figure 2). A total of 173 OTUs and 89 genera were shared among the three egg parts (eggshell, inner membrane, and egg content) (Figure 2A). The 10 most frequent genera within these shared groups were Staphylococcus, Lactobacillus, Flexivirga, Salinicoccus, Pseudomonas, Rhodanobacter, Bacteroides, Paracoccus, Chujaibacter, and Rhodococcus. Compared to the egg parts, litter treatments (Figure 2B) and the age of the flocks (Figure 2C) resulted in fewer unique OTUs and genera.
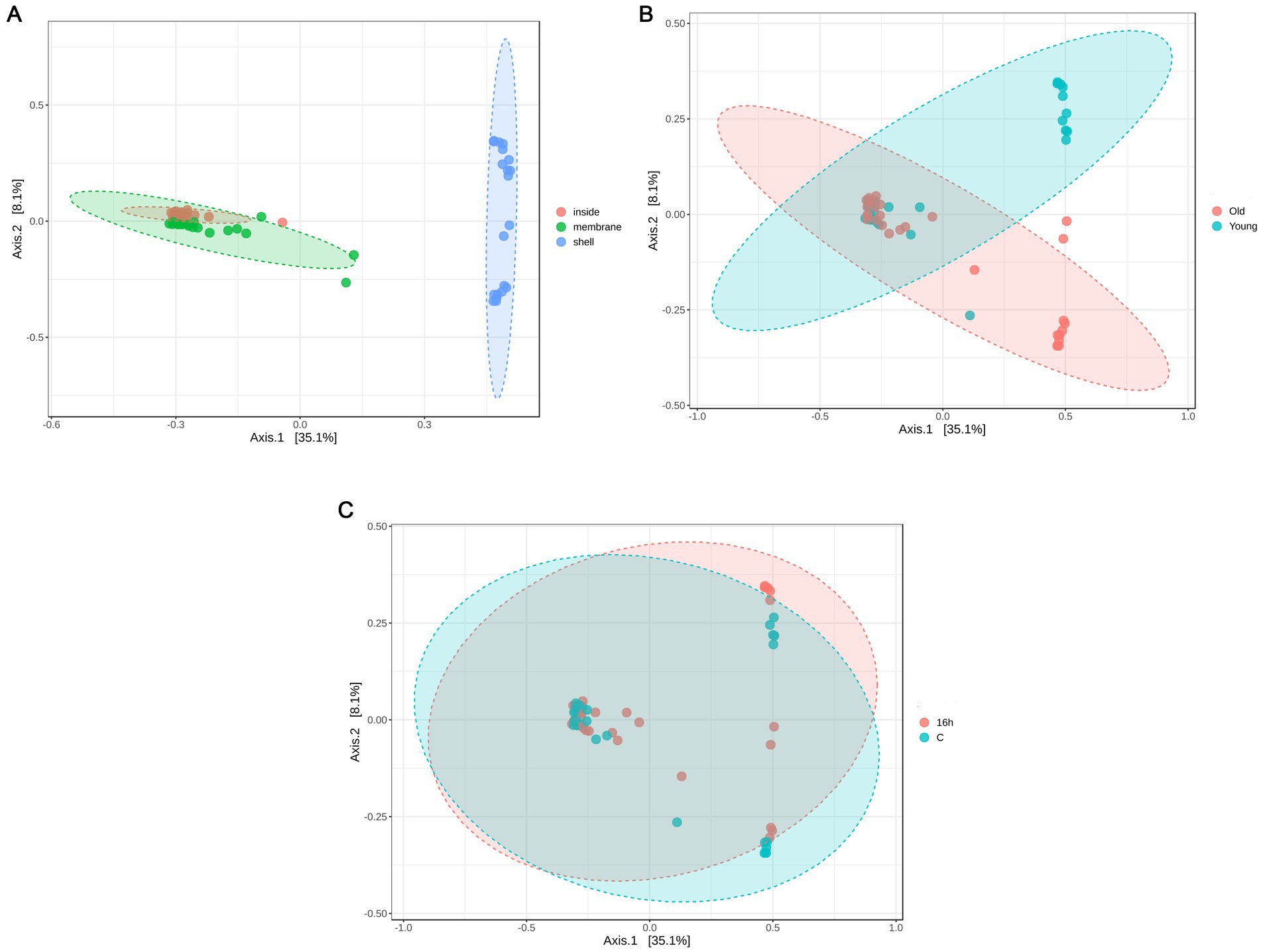
Figure 2. Venn diagrams showing the numbers of unique and shared OTUs and bacterial genera (numbers in parentheses) based on (A) egg parts, (B) age of breeder flocks, and (C) litter treatments.
3.3.6 Distribution of spoilage bacteria
The sum of relative abundances of eggshell spoilage-associated bacterial genera (Alcaligenes, Enterococcus, Escherichia/Shigella, Proteus, Pseudomonas, Staphylococcus, and Streptococcus) significantly increased after 16 h of litter treatment (Supplementary Table S4). Eggs from the younger flock contained a significantly higher ratio of these bacteria. The ratio of spoilage bacteria was highest on the eggshell surface, but interestingly, the difference between the egg membrane and egg content was also significant (Supplementary Table S4). This suggests some bacterial diffusion through the eggshell, although this could not be confirmed from the results of 16S rRNA sequencing.
4 Discussion
Floor eggs result in significant losses for broiler breeder farms and breeding companies due to their higher risk of bacterial contamination, which threatens both embryo viability and hatchery hygiene. If these eggs are excluded from incubation, the breeding farm incurs substantial losses. A floor egg ratio below 2% is considered good, and a range between 2 and 4% is generally acceptable. Floor egg ratios are typically higher in younger flocks. Cooper and Appleby (53) reported a 5% floor egg rate in 27–28-week-old broiler breeder hens, while other studies have recorded even higher percentages—up to 13.3% in 35-44-week-old flocks (54).
Although the hatchability of floor eggs is lower, the underlying causes are not yet fully understood. Two key, often interacting, factors have been identified: microbial infection of the embryo and ambient temperature (35). In our study, microbial contamination of the eggshell increased significantly, and hatchability decreased markedly after 16 h of contact with litter. While the number of culturable aerobic microbes increased on the surface of the floor eggs, no further increase was observed after 3 h. This finding indicates that time spent in the litter is not the sole determinant of aerobic bacterial load on the eggshell.
From a practical standpoint, this finding emphasizes the importance of frequent floor egg collection to preserve hatchability. We found no significant differences in coliform counts between litter treatments, and their numbers remained low across all samples. Our findings on microbial contamination are consistent with previous studies (1–3) and align with field observations. Peralta-Sánchez et al. (55) reported a negative relationship between aerobic mesophilic bacterial density on the eggshell and hatchability across 17 bird species. In contrast, other studies have reported no significant differences in hatchability between floor and nest eggs despite higher contamination levels in the former (4).
While the majority of studies on this topic focus on floor eggs, some have also investigated the effects of litter treatments (2, 3). In our study, hatchability decreased significantly only after 16 h of litter exposure, which correlated with increased embryonic mortality. Since aerobic bacterial counts on the eggshell plateaued after 3 h, the observed decline in hatchability was likely due to delayed egg cooling rather than microbial load. Prolonged exposure to warm litter delays temperature reduction, which is detrimental to embryo viability (53).
Interestingly, the number of aerobic bacteria on the eggshell decreased with hen age in our study, a result that contrasts with Moyle et al. (33), who reported increasing bacterial counts with hen age. However, their study was conducted on free-range laying hens, which are not directly comparable to our broiler breeder setup.
Egg quality is also influenced by hen age (56). As expected, the eggs of older hens (65–66 weeks) were heavier than those of younger hens (22–23 weeks), consistent with prior findings (26, 28, 57). Hatchability is generally higher in eggs from younger hens (3, 24, 27, 28, 30, 58), although some studies found no age-related differences (25) or even reported better hatchability in older hens (29). The discrepancies may be attributed to breed differences, flock age, or egg storage conditions before incubation (30). Optimal storage conditions must be tailored to the specific breeder flock to maximize embryo survival (30). These inconsistencies highlight the importance of standardizing hen age, breed, and storage conditions in hatchability studies.
Microbial diversity indices and OTU counts differed between egg components. Chao1 reflects species richness, Shannon accounts for both richness and evenness with sensitivity to rare OTUs, while Simpson emphasizes dominant OTUs (59). As expected, the eggshell exhibited the highest OTU counts and Chao1 diversity, followed by the membrane and then the egg content. Notably, the membrane had significantly higher OTU numbers and Chao1 indices than the egg content—a finding not previously reported, warranting further investigation.
The Shannon and Simpson indices also showed significant effects of litter treatment, although interactions among factors complicate interpretation. Our Shannon and Simpson values for egg content are consistent with those reported for egg whites in previous studies (40, 41).
Neither the membrane nor the egg content showed significant differences in OTU numbers or diversity indices across treatments, suggesting that the internal egg microbiota remain unaffected as long as the eggshell structure is intact.
Litter treatment significantly altered the microbial community composition on the eggshell at both the phylum and genus levels. The dominant phyla—Firmicutes, Bacteroidetes, Actinobacteria, and Proteobacteria—were consistent with earlier reports (5, 9, 60). In our study, litter exposure treatment increased the relative abundance of Firmicutes while reducing Proteobacteria. At the genus level, Staphylococcus abundance increased, and Lactobacillus decreased after 16 h of litter exposure. Older flocks had a lower abundance of Staphylococcus and Lactobacillus but a higher abundance of Salinicoccus. Although the litter microbiota was not characterized in our study, we attribute these differences to variations in litter microbial composition between flocks.
The eggshell microbiota originates from the hen’s reproductive and digestive tracts, as well as from the litter environment (6, 37). Litter is predominantly aerobic, which limits the survival of strictly anaerobic fecal bacteria. However, facultative anaerobes, aerotolerant, or spore-forming species can persist (12). While the majority of data on gut microbiota dynamics comes from broilers, some studies on laying phases indicate significant changes before laying and relative stability during the laying period (61, 62). Firmicutes dominate early laying phases (weeks 18–46), while Bacteroidetes increase during late phases (weeks 58–75) (34, 61). Litter microbiota composition is dynamic and influenced by factors such as diet, excreta, and litter moisture content.
Few studies have investigated temporal shifts in poultry manure microbiota (12, 63). Crippen et al. (13) and Zwirzitz et al. (14) reported that Firmicutes and Actinobacteria dominate mature litter, with Actinobacteria increasing over time—consistent with our eggshell microbiota results. Although eggshell microbiota has been studied (7, 60, 64, 65), the effect of flock or litter age on the eggshell microbial community has not been previously reported.
Neither litter treatment nor hen age significantly affected the microbial composition of the egg membrane. To the best of our knowledge, this is the first study to characterize the microbial composition of broiler hatching egg membranes. Dominant phyla included Proteobacteria and Firmicutes, with high abundances of Actinobacteria, Cyanobacteria, and Bacteroidetes. Minor phyla such as Patescibacteria, Chlamydiae, and Planctomycetes also exceeded 1% relative abundance. The genera Pseudomonas and Flexivirga together represented over 10% of the membrane microbiota. While Pseudomonas presence is expected, the detection of Flexivirga—a Gram-positive, aerobic, non-spore-forming genus in Actinobacteria—is novel in poultry-related research. Flexivirga has been previously isolated from soil (66), pine species (67), and mosquitoes (68), but not from poultry. Its relatively high abundance in the membrane (5.95%) and egg content (9.39%) compared to the eggshell (0.015%) suggests an oviduct origin. Previous studies have shown significant correlations between the microbial communities of the oviduct and egg white, as well as between the embryo gut and egg yolk microbiota (37, 41).
Our study examined the total egg content (yolk and white combined). Neither litter treatment nor breeder age significantly influenced its microbiome. Only a few studies have characterized yolk or white microbiota (37, 40, 41), but these findings generally indicate that Proteobacteria, Firmicutes, Bacteroidetes, and Actinobacteria are predominant. Despite analyzing the mixed content, our results are consistent, with Proteobacteria and Firmicutes as the dominant phyla, and minor phyla in similar proportions. Flexivirga was the most abundant genus in the egg content (9.4%), not previously reported in this context.
In litter-treated eggs, the relative abundance of spoilage-associated genera on the eggshell was nearly double that of untreated eggs. Although their abundance was lower internally, it was surprisingly higher in the membrane than in the egg content. This finding suggests that some spoilage organisms may penetrate the shell but remain below detection limits, even when sequencing-based methods are applied.
5 Conclusion
Litter treatment of hatching eggs increased the number of culturable aerobic microbes on the eggshell, with microbial penetration of the shell surface occurring after only 3 h. However, hatchability declined significantly only after 16 h of litter exposure. Since no bacterial translocation into the egg contents was detected, the reduced hatchability is likely due to prolonged exposure to uncontrolled temperatures, which may trigger premature embryonic development or compromise embryo viability. The primary cause of reduced hatchability was increased embryonic mortality. Litter treatment and flock age influenced only the composition of the eggshell microbiota; no significant effects were observed in the microbiota of the egg membrane or egg contents. The cumulative relative abundance of spoilage-associated bacterial genera—Alcaligenes, Enterococcus, Escherichia/Shigella, Proteus, Pseudomonas, Staphylococcus, and Streptococcus—increased significantly on the eggshell following litter exposure. Interestingly, the combined relative abundance of these genera was significantly higher in the egg membrane than in the egg content. A particularly novel finding was the high relative abundance of the genus Flexivirga in both the egg membrane and content, which has not been reported in previous studies. Further research is needed to clarify its origin, role, and potential impact on egg quality and embryonic development.
Data availability statement
The raw sequence data were deposited in the NCBI Sequence Read Archive (SRA) under BioProject ID PRJNA1012860.
Ethics statement
The animal study was approved by Animal Welfare Committee, Georgikon Campus, Hungarian University of Agriculture and Life Sciences. The study was conducted in accordance with the local legislation and institutional requirements.
Author contributions
GC: Formal analysis, Writing – original draft. NS: Data curation, Formal analysis, Investigation, Writing – review & editing. LM: Data curation, Writing – original draft. KS: Investigation, Writing – review & editing. KH: Investigation, Writing – original draft. VF: Data curation, Investigation, Writing – original draft. LP: Investigation, Writing – original draft. KD: Conceptualization, Resources, Supervision, Writing – review & editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This study was supported by the Flagship Research Groups Programme of the Hungarian University of Agriculture and Life Sciences.
Conflict of interest
KH was employed by Gallus Ltd. of Poultry Breeding and Hatching.
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Gen AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fvets.2025.1589607/full#supplementary-material
References
1. Van Den Brand, H, Sosef, MP, Lourens, A, and Van Harn, J. Effects of floor eggs on hatchability and later life performance in broiler chickens. Poult Sci. (2016) 95:1025–32. doi: 10.3382/ps/pew008
2. Vekić, M, Bijelonić, B, Perić, L, Savić, Đ, Jotanović, S, and Mitraković, M. Effect of nest and floor eggs with visually clean shells on hatchability and chick quality in broiler breeders. Agro Knowl J. (2021) 22:77–85. doi: 10.7251/agren2103077v
3. Perić, L, Mitraković, M, Tomić, B, Orehovački, I, and Meijerhof, R. Effects of flock age, place of oviposition and cleaning treatments of hatching eggs on hatchability in broiler breeders. J Appl Poult Res. (2022) 31:100279. doi: 10.1016/j.japr.2022.100279
4. Costa, BTA, Lopes, TSB, Sousa, LS, Barbosa, HJS, Gonçalves, TF, Franco, CMA, et al. Floor eggs: hatchability, microbiological analysis, and effects of post-hatch use of ceftiofur on performance of broiler chicks. Res Vet Sci. (2022) 152:610–7. doi: 10.1016/j.rvsc.2022.09.020
5. Shi, Y, Wu, S, Li, W, Liu, D, Ma, G, Zhang, Z, et al. Microbiota analysis of eggshells in different areas and during different storage time by non-cultural methods. Curr Microbiol. (2020) 77:3842–50. doi: 10.1007/s00284-020-02212-y
6. Rychlik, I, Karasova, D, and Crhanova, M. Microbiota of chickens and their environment in commercial production. Avian Dis. (2023) 67:1–9. doi: 10.1637/aviandiseases-d-22-00048
7. Olsen, R, Kudirkiene, E, Thøfner, I, Pors, S, Karlskov-Mortensen, P, Li, L, et al. Impact of egg disinfection of hatching eggs on the eggshell microbiome and bacterial load. Poult Sci. (2017) 96:3901–11. doi: 10.3382/ps/pex182
8. Neira, C, Laca, A, Laca, A, and Díaz, M. Microbial diversity on commercial eggs as affected by the production system. A first approach using PGM. Int J Food Microbiol. (2017) 262:3–7. doi: 10.1016/j.ijfoodmicro.2017.09.008
9. Maki, JJ, Bobeck, EA, Sylte, MJ, and Looft, T. Eggshell and environmental bacteria contribute to the intestinal microbiota of growing chickens. J Anim Sci Biotechnol. (2020) 11:60. doi: 10.1186/s40104-020-00459-w
10. Xu, X, Rothrock, MJ, Mishra, A, Kumar, GD, and Mishra, A. Relationship of the poultry microbiome to pathogen colonization, farm management, poultry production, and foodborne illness risk assessment. J Food Prot. (2023) 86:100169. doi: 10.1016/j.jfp.2023.100169
11. Lu, J, Sanchez, S, Hofacre, C, Maurer, JJ, Harmon, BG, and Lee, MD. Evaluation of broiler litter with reference to the microbial composition as assessed by using 16S rRNA and functional gene markers. Appl Environ Microbiol. (2003) 69:901–8. doi: 10.1128/AEM.69.2.901-908.2003
12. Kubasova, T, Faldynova, M, Crhanova, M, Karasova, D, Zeman, M, Babak, V, et al. Succession, replacement, and modification of chicken litter microbiota. Appl Environ Microbiol. (2022) 88:e0180922. doi: 10.1128/aem.01809-22
13. Crippen, TL, Sheffield, CL, Singh, B, Byrd, JA, Beier, RC, and Anderson, RC. Poultry litter and the environment: microbial profile of litter during successive flock rotations and after spreading on pastureland. Sci Total Environ. (2021) 780:146413. doi: 10.1016/j.scitotenv.2021.146413
14. Zwirzitz, B, Oladeinde, A, Johnson, J, Zock, G, Milfort, MC, Fuller, AL, et al. Temporal dynamics of the cecal and litter microbiome of chickens raised in two separate broiler houses. Front Physiol. (2023) 14:14. doi: 10.3389/fphys.2023.1083192
15. de Toledo, TDS, Roll, AAP, Rutz, F, Dallmann, HM, Prá, MAD, Leite, FPL, et al. An assessment of the impacts of litter treatments on the litter quality and broiler performance: a systematic review and metaanalysis. PLoS One. (2020) 15:e0232853. doi: 10.1371/journal.pone.0232853
16. Wang, L, Lilburn, M, and Yu, Z. Intestinal microbiota of broiler chickens as affected by litter management regimens. Front Microbiol. (2016) 7. doi: 10.3389/fmicb.2016.00593
17. Valeris-Chacin, R, Pieters, M, Hwang, H, Johnson, TJ, and Singer, RS. Association of broiler litter microbiome composition and Campylobacter isolation. Front Vet Sci. (2021) 8:654927. doi: 10.3389/fvets.2021.654927
18. De Reu, K, Messens, W, Heyndrickx, M, Rodenburg, TB, Uyttendaele, M, and Herman, L. Bacterial contamination of table eggs and the influence of housing systems. Worlds Poult Sci J. (2008) 64:5–19. doi: 10.1017/S0043933907001687
19. Liu, W, Zhang, J, Guo, A, Chen, Q, Gu, L, Ruan, Y, et al. The specific biological characteristics of spoilage microorganisms in eggs. LWT. (2021) 135:110069. doi: 10.1016/j.lwt.2020.110069
20. Svobodová, J, and To̊mová, E. Factors affecting microbial contamination of market eggs: a review. Sci Agric Bohem. (2014) 45:226–37. doi: 10.1515/sab-2015-0003
21. De Reu, K, Grijspeerdt, K, Messens, W, Heyndrickx, M, Uyttendaele, M, Debevere, J, et al. Eggshell factors influencing eggshell penetration and whole egg contamination by different bacteria, including Salmonella enteritidis. Int J Food Microbiol. (2006) 112:253–60. doi: 10.1016/j.ijfoodmicro.2006.04.011
22. Oliveira, G d S, McManus, C, Salgado, CB, and dos Santos, VM. Effects of sanitizers on microbiological control of hatching eggshells and poultry health during embryogenesis and early stages after hatching in the last decade. Animals. (2022) 12:2826. doi: 10.3390/ani12202826
23. Mohammadi-Aragh, MK, Linhoss, JE, and Evans, JD. Effects of various disinfectants on the bacterial load and microbiome of broiler hatching eggs using electrostatic spray. J Appl Poult Res. (2022) 31:100278. doi: 10.1016/j.japr.2022.100278
24. Tona, K, Bamelis, F, Coucke, W, Bruggeman, V, and Decuypere, E. Relationship between broiler breeder’s age and egg weight loss and embryonic mortality during incubation in large-scale conditions. J Appl Poult Res. (2001) 10:221–7. doi: 10.1093/japr/10.3.221
25. Joseph, NS, and Moran, ET. Effect of flock age and postemergent holding in the hatcher on broiler live performance and further-processing yield. J Appl Poult Res. (2005) 14:512–20. doi: 10.1093/japr/14.3.512
26. Damaziak, K, Koznaka-Lipka, M, Gozdowski, D, Gołębiowska, A, and Kędziorek, E. Effects of broiler breeder strain, age, and eggs preheating profile in single-stage systems on the hatchability of eggs and quality of chicks. Animal. (2021) 15:100057. doi: 10.1016/j.animal.2020.100057
27. Iqbal, J, Khan, SH, Mukhtar, N, Ahmed, T, and Pasha, RA. Effects of egg size (weight) and age on hatching performance and chick quality of broiler breeder. J Appl Anim Res. (2016) 44:54–64. doi: 10.1080/09712119.2014.987294
28. Abudabos, A. The effect of broiler breeder strain and parent flock age on hatchability and fertile hatchability. Int J Poult Sci. (2010) 9:231–5. doi: 10.3923/ijps.2010.231.235
29. Roque, L, and Soares, MC. Effects of eggshell quality and broiler breeder age on hatchability. Poult Sci. (1994) 73:1838–45. doi: 10.3382/ps.0731838
30. Nasri, H, van den Brand, H, Najjar, T, and Bouzouaia, M. Egg storage and breeder age impact on egg quality and embryo development. J Anim Physiol Anim Nutr. (2020) 104:257–68. doi: 10.1111/jpn.13240
31. Pokhrel, N, Cohen, EBT, Genin, O, Ruzal, M, Sela-Donenfeld, D, and Cinnamon, Y. Physiology and reproduction: effects of storage conditions on hatchability, embryonic survival and cytoarchitectural properties in broiler from young and old flocks. Poult Sci. (2018) 97:1429–40. doi: 10.3382/ps/pex393
32. Jones, DR, Anderson, KE, Curtis, PA, and Jones, FT. Microbial contamination in inoculated shell eggs: I. Effects of layer strain and hen age. Poult Sci. (2002) 81:715–20. doi: 10.1093/ps/81.5.715
33. Moyle, T, Drake, K, Gole, V, Chousalkar, K, and Hazel, S. Bacterial contamination of eggs and behaviour of poultry flocks in the free range environment. Comp Immunol Microbiol Infect Dis. (2016) 49:88–94. doi: 10.1016/j.cimid.2016.10.005
34. Dai, D, Qi, G h, Wang, J, Zhang, H j, Qiu, K, and Wu, S g. Intestinal microbiota of layer hens and its association with egg quality and safety. Poult Sci. (2022) 101:102008. doi: 10.1016/j.psj.2022.102008
35. Wang, JM, Firestone, MK, and Beissinger, SR. Microbial and environmental effects on avian egg viability: do tropical mechanisms act in a temperate environment? Ecology. (2011) 92:1137–45. doi: 10.1890/10-0986.1
36. Ding, J, Dai, R, Yang, L, He, C, Xu, K, Liu, S, et al. Inheritance and establishment of gut microbiota in chickens. Front Microbiol. (2017) 8:1967. doi: 10.3389/fmicb.2017.01967
37. Lee, SW, La, TM, Lee, HJ, Choi, IS, Song, CS, Park, SY, et al. Characterization of microbial communities in the chicken oviduct and the origin of chicken embryo gut microbiota. Sci Rep. (2019) 9:6838–11. doi: 10.1038/s41598-019-43280-w
38. Wen, C, Li, Q, Lan, F, Li, X, Li, G, Yan, Y, et al. Microbiota continuum along the chicken oviduct and its association with host genetics and egg formation. Poult Sci. (2021) 100:101104. doi: 10.1016/j.psj.2021.101104
39. Shterzer, N, Rothschild, N, Sbehat, Y, Stern, E, Nazarov, A, and Mills, E. Large overlap between the intestinal and reproductive tract microbiomes of chickens. Front Microbiol. (2020) 11. doi: 10.3389/fmicb.2020.01508
40. Ding, P, Liu, H, Tong, Y, He, X, Yin, X, Yin, Y, et al. Developmental change of yolk microbiota and its role on early colonization of intestinal microbiota in chicken embryo. Animals. (2022) 12:16. doi: 10.3390/ani12010016
41. Jin, J, Zhou, Q, Lan, F, Li, J, Yang, N, and Sun, C. Microbial composition of egg component and its association with hatchability of laying hens. Front Microbiol. (2022) 13:943097. doi: 10.3389/fmicb.2022.943097
42. Shterzer, N, Sbehat, Y, Poudel, B, Rothschild, N, Oloko, OE, Druyan, S, et al. Comparative analysis of reproductive tract microbiomes in modern and slower-growing broiler breeder lines. Front Vet Sci. (2024) 11:1–9. doi: 10.3389/fvets.2024.1386410
43. Akinyemi, FT, Ding, J, Zhou, H, Xu, K, He, C, Han, C, et al. Dynamic distribution of gut microbiota during embryonic development in chicken. Poult Sci. (2020) 99:5079–90. doi: 10.1016/j.psj.2020.06.016
44. Illumina Inc. 16S Metagenomic sequencing library preparation-preparing 16S Ribosomal RNA gene amplicons for the illumina MiSeq system. San Diego, CA, USA: Illumina Inc. (2013).
45. Bolyen, E, Rideout, JR, Dillon, MR, Bokulich, NA, Abnet, CC, Al-Ghalith, GA, et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat Biotechnol. (2019) 37:852–7. doi: 10.1038/s41587-019-0209-9
46. Quast, C, Pruesse, E, Yilmaz, P, Gerken, J, Schweer, T, Yarza, P, et al. The SILVA ribosomal RNA gene database project: improved data processing and web-based tools. Nucleic Acids Res. (2013) 41:590–6. doi: 10.1093/nar/gks1219
47. Chong, J, Liu, P, Zhou, G, and Xia, J. Using microbiome analyst for comprehensive statistical, functional, and meta-analysis of microbiome data. Nat Protoc. (2020) 15:799–821. doi: 10.1038/s41596-019-0264-1
48. R Core Team. R: a language and environment for statistical computing. R Found Stat Comput (2020) 2. Available online at: https://www.R-project.org.
49. Kay, M, and Wobbrock, JO. ARTool: aligned rank transform for nonparametric factorial ANOVAs, R package version 0.10.0 (2015) 8.
50. Benjamini, Y, and Hochberg, Y. Controlling the false discovery rate: a practical and powerful approach to multiple testing. J R Stat Soc Ser B Stat Methodol. (1995) 57:289–300. doi: 10.1111/j.2517-6161.1995.tb02031.x
51. Wobbrock, JO, Findlater, L, Gergle, D, and Higgins, JJ. (2011). The aligned rank transform for nonparametric factorial analyses using only ANOVA procedures. In: Proceedings of the ACM conference on human factors in computing systems (CHI ‘11). Vancouver, British Columbia. New York: ACM Press, pp. 143–146.
52. la Rosa, PS, Brooks, JP, Deych, E, Boone, EL, Edwards, DJ, Wang, Q, et al. Hypothesis testing and power calculations for taxonomic-based human microbiome data. PLoS One. (2012) 7:e52078. doi: 10.1371/journal.pone.0052078
53. Cooper, JJ, and Appleby, MC. Individual variation in prelaying behaviour and the incidence of floor eggs. Br Poult Sci. (1996) 37:245–53. doi: 10.1080/00071669608417856
54. Sheppard, KC, and Duncan, IJH. Feeding motivation on the incidence of floor eggs and extraneously calcified eggs laid by broiler breeder hens. Br Poult Sci. (2011) 52:20–9. doi: 10.1080/00071668.2010.550600
55. Peralta-Sánchez, JM, Martín-Platero, AM, Wegener-Parfrey, L, Martínez-Bueno, M, Rodríguez-Ruano, S, Navas-Molina, JA, et al. Bacterial density rather than diversity correlates with hatching success across different avian species. FEMS Microbiol Ecol. (2018) 94:fiy022. doi: 10.1093/femsec/fiy022
56. Manyeula, F, Sebolai, B, Sempule, G, and Moreki, JC. Effects of broiler breeders’ age on egg quality characteristics and their correlation coefficients. J World's Poult Res. (2021) 11:368–75. doi: 10.36380/JWPR.2021.44
57. Tona, K, Onagbesan, O, De Ketelaere, B, Decuypere, E, and Bruggeman, V. Effects of age of broiler breeders and egg storage on egg quality, hatchability, chick quality, chick weight, and chick posthatch growth to forty-two days. J Appl Poult Res. (2004) 13:10–8. doi: 10.1093/japr/13.1.10
58. Ipek, A, and Sozcu, A. The effects of broiler breeder age on intestinal development during hatch window, chick quality and first week broiler performance. J Appl Anim Res. (2015) 43:402–8. doi: 10.1080/09712119.2014.978783
59. Nagendra, H. Opposite trends in response for the Shannon and Simpson indices of landscape diversity. Appl Geogr. (2002) 22:175–86. doi: 10.1016/S0143-6228(02)00002-4
60. Trudeau, S, Thibodeau, A, Côté, JC, Gaucher, M, Lou, M, and Fravalo, P. Contribution of the broiler breeders’ fecal microbiota to the establishment of the eggshell microbiota. Front Microbiol. (2020) 11:666. doi: 10.3389/fmicb.2020.00666
61. Joat, N, Van, TTH, Stanley, D, Moore, RJ, and Chousalkar, K. Temporal dynamics of gut microbiota in caged laying hens: a field observation from hatching to end of lay. Appl Microbiol Biotechnol. (2021) 105:4719–30. doi: 10.1007/s00253-021-11333-8
62. Videnska, P, Sedlar, K, Lukac, M, Faldynova, M, Gerzova, L, Cejkova, D, et al. Succession and replacement of bacterial populations in the caecum of egg laying hens over their whole life. PLoS One. (2014) 9:e115142. doi: 10.1371/journal.pone.0115142
63. Johnson, J, Zwirzitz, B, Oladeinde, A, Milfort, M, Looft, T, Chai, L, et al. Succession patterns of the bacterial community in poultry litter after bird removal and sodium bisulfate application. J Environ Qual. (2021) 50:923–33. doi: 10.1002/jeq2.20248
64. Rychlik, I. Composition and function of chicken gut microbiota. Animals. (2020) 10:103. doi: 10.3390/ani10010103
65. Volf, J, Crhanova, M, Karasova, D, Faldynova, M, Kubasova, T, Seidlerova, Z, et al. Eggshell and feed microbiota do not represent major sources of gut anaerobes for chickens in commercial production. Microorganisms. (2021) 9:1480. doi: 10.3390/microorganisms9071480
66. Anzai, K, Sugiyama, T, Sukisaki, M, Sakiyama, Y, Otoguro, M, and Ando, K. Flexivirga alba gen. Nov., sp. nov., an actinobacterial taxon in the family Dermacoccaceae. J Antibiot. (2011) 64:613–6. doi: 10.1038/ja.2011.62
67. Ponpandian, LN, Rim, SO, Shanmugam, G, Jeon, J, Park, YH, Lee, SK, et al. Phylogenetic characterization of bacterial endophytes from four Pinus species and their nematicidal activity against the pine wood nematode. Sci Rep. (2019) 9:12457. doi: 10.1038/s41598-019-48745-6
Keywords: floor egg, breeders’ age, broiler breeders, hatchability, microbiome, eggshell, egg membrane, egg content
Citation: Csitári G, Such N, Menyhárt L, Schermann K, Humpok K, Farkas V, Pál L and Dublecz K (2025) Effects of litter exposure and flock age of broiler breeders on hatchability and the microbial composition of eggshells, egg membranes, and egg contents. Front. Vet. Sci. 12:1589607. doi: 10.3389/fvets.2025.1589607
Edited by:
Regiane R. Santos, Schothorst Feed Research, NetherlandsReviewed by:
Monika Proszkowiec-Weglarz, United States Department of Agriculture, United StatesOyegunle Emmanuel Oke, Federal University of Agriculture, Nigeria
Copyright © 2025 Csitári, Such, Menyhárt, Schermann, Humpok, Farkas, Pál and Dublecz. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Károly Dublecz, ZHVibGVjei5rYXJvbHlAdW5pLW1hdGUuaHU=