 Peiyao Liu1†
Peiyao Liu1† Chunna Cao1,2†Xiangding Wang1Kewei Li1
Chunna Cao1,2†Xiangding Wang1Kewei Li1 Ebadu Areb1
Ebadu Areb1 Ran Li1Qingfeng Zhang3
Ran Li1Qingfeng Zhang3 Chuanying Pan1*
Chuanying Pan1* Xianyong Lan1*
Xianyong Lan1*- 1Key Laboratory of Animal Genetics, Breeding and Reproduction of Shaanxi Province, College of Animal Science and Technology, Northwest A&F University, Yangling, Shaanxi, China
- 2Yangling Vocational and Technical College, Yangling, Shaanxi, China
- 3Tianjin Aoqun Sheep Industry Academy Company, Tianjin, China
Introduction: Bone morphogenetic protein 15 (BMP15) is a pivotal regulator of reproductive performance. This study aimed to investigate genetic variations in the sheep BMP15 gene and their role in lambing traits.
Methods: Whole-genome sequencing was performed on 2,409 individuals from 75 global sheep breeds to identify functional variations in BMP15. Variant annotation, protein structure prediction, haplotype analysis, and association studies with lambing traits were conducted.
Results: 139 SNPs were detected, including 10 exonic variants (6 missense, 2 synonymous, 2 premature stop codons), 9 of which were novel. The high-frequency missense mutation g.54284258A>G (L251P) showed predicted localized conformational changes. Low-frequency stop-gain mutations (e.g., g.54284551C>T) were rare (<0.005%) in domestic breeds. Haplotype H6 (AAAA) predominated in domestic breeds (48.4%), diverging significantly from the wild ancestral H1 (AAGA), indicating artificial selection. The g.54284258A>G variant was significantly associated with live lamb number at first (p < 0.01) and second parity (p < 0.05). AG genotypes had more live lambs than AA (1.29 vs. 1.12; p < 0.01). No significant associations were found with total lambing number, lamb survival rate, stillbirth count, or stillbirth rate.
Discussion: BMP15 exhibits substantial genetic variability shaped by selection. The g.54284258A>G variant significantly influences live lamb numbers and may serve as a genetic marker for improving this trait in sheep breeding programs, though its impact is parity-specific and unrelated to stillbirth or overall survival metrics.
1 Introduction
Sheep populations have dispersed across diverse ecology worldwide after domestication, encountering heterogeneous environmental pressures and selective constraints that resulted adaptive diversification through genetic variation (1). This evolutionary process, underpinned by the emergence of advantageous alleles, has shaped the species’ global genetic diversity, highlighting the importance of characterizing genetic variability for both conservation of unique breeds and optimization of breeding programs (2). While advances in reproducible genotyping tools have enabled cross-border collaborations for in-depth study of genetic diversity (3).
Reproductive traits, particularly litter size (total lambs born including stillbirths) and number of live lambs (viable neonates surviving >24 h postpartum), are polygenic in nature, modulated by intricate interactions among environmental factors (e.g., nutrition and management), epigenetic modifications, and multi-gene regulatory networks. Their low heritability (h2 = 0.1–0.3) renders conventional phenotypic selection strategies ineffective for achieving genetic gains (4). This limitation underscores the urgency to decipher the genetic regulatory networks governing these traits and identify causal functional mutations. It might be prerequisite for developing precision breeding technologies aimed at enhancing ovine reproductive efficiency (including litter size, live lambs and others).
Bone morphogenetic protein 15 (BMP15), an X-linked oocyte-specific member of the TGF-β superfamily, has emerged as a pivotal regulator of reproductive performance. Expressed from the early secondary follicular stage through ovulation (5). BMP15 influences folliculogenesis and embryo survival via conserved signaling pathways. Although mutations such as FecXI and FecXR are associated with prolificacy in select sheep breeds (6). On the other hand, BMP15 variants show no significant association with litter size in Indian Black Bengal goats (7) and Tibetan cashmere goats (8). These discrepancies suggest species- and breed-specific functional roles for BMP15, potentially modulated by genetic background or environmental covariates. Therefore, systematic validation across globally diverse populations is essential.
This study integrated high-throughput sequencing with functional gene annotation to systematically identify genetic variants associated with litter size and number of live lambs, while evaluating their distribution patterns and potential effects across diverse sheep breeds. Through comprehensive analyses, we aim to screen candidate genes and loci significantly associated with lambing traits, providing novel insights into the genetic basis for reproductive performance. These findings contribute to understanding of the genetic regulation of complex traits and lay scientific foundation for molecular breeding strategy to enhance sheep reproductive efficiency.
2 Materials and methods
All experimental procedures were performed in accordance with the Faculty of Animal Policy and Welfare Committee of Northwest A&F University (protocol no. NWAFU-314020038) for the use and care of animals in research.
2.1 Samples collection
This study utilized whole-genome sequencing data from 2,409 sheep individuals publicly available in the NCBI SRA database1 (Supplementary Table S1), with an average sequencing depth of 13.25×. These samples represent 75 global sheep breeds, including 361 individuals from Africa, 339 from America, 1,021 from East Asia, 587 from Europe, 52 from the Middle East, and 49 wild sheep (Ovis orientalis) (Supplementary Table S1).
Lambing traits such as litter size, number of live lambs, live lamb rate, stillbirth count, still birth rate, and lamb survival rate was collected from Australian White (AUW) sheep (n = 576) in Tianjin, China. Approximately 8 mL blood sample was collected from each ewe into EDTA-coated tubes from jugular vein and stored at −80°C for subsequent DNA extraction. Phenol-chloroform extraction method was used to extract genomic DNA from blood. The concentrations were measured by a Nanodrop 2000 Spectrophotometer to assess DNA purity (A260/280 ratio) and quality, and were diluted to 20 ng/μL and frozen at −40°C for further experiments. To minimize genetic confounding effect, unrelated individuals were selected based on pedigree records. Additionally, fixed effects such as age and parity were collected.
2.2 Read alignment and SNP detection
Raw FASTQ files underwent initial quality filtering using fastp v0.20.0. The processed reads were then aligned to the Ovis aries reference genome, ARS-UI_Ramb_v22 using BWA-MEM v0.7.13-r1126, generating binary alignment (BAM) files. Variant calling was performed using GATK v3.6-0-g89b7209, leveraging the “HaplotypeCaller” and “GenotypeGVCFs” modules to identify single nucleotide polymorphisms (SNPs). To ensure data quality, subsequent filtering was conducted with bcftools-1.13, applying stringent thresholds to remove artifacts: QD < 2.0, QUAL < 30.0, SOR > 3.0, FS > 60.0, MQ < 40.0, MQRankSum < −12.5, and ReadPosRankSum < −8.0. Further filtering retained only biallelic SNPs (-m2 -M2 -i) with a minor allele frequency (MAF) ≥ 0.05 and a genotype missing rate (F_MISSING) < 0.2.
2.3 Variant annotation
Gene annotation was performed using SnpEff v4.2 (9), categorizing variants into intronic, exonic, intergenic, splice-site, upstream, and downstream regulatory regions. SNPs located in exonic regions were further classified as synonymous or nonsynonymous variants. Finally, VCFtools v0.1.16 (10) was employed to calculate allele frequencies for all SNPs across different sheep breeds.
2.4 Linkage disequilibrium analysis
Linkage disequilibrium (LD) analysis was performed for all SNPs within the coding regions of the BMP15 gene using the LDheatmap R package (11).
2.5 Protein structure prediction
The amino acid sequences of the ovine BMP15 gene (NP_001108239.1) were retrieved from the NCBI Protein database.3 Three-dimensional protein structures were predicted using the AlphaFold2 online tool,4 with structural modeling performed for both wild-type and missense mutation variants. Finally, PyMOL v7.0.1 (12) was used to visualize and compare the protein structures before and after mutation.
2.6 Haplotype analysis
VCF files for the coding regions of the BMP15 gene were first filtered using bcftools v1.13 (10) with criteria of minor allele frequency (MAF) > 0.1 and genotype missing rate < 0.1. The filtered VCF files were subsequently converted to FASTA format. Multiple sequence alignment was then performed using MAFFT (13). Haplotype identification was streamlined by extracting haplotype data (.rdf files) from the aligned FASTA sequences using DnaSP v6.0 (14). Finally, low-frequency haplotypes (frequency < 5%) were excluded, and haplotype networks were visualized using Network v10.2 (15).
2.7 KASP genotyping
KASP (Kompetitive Allele-Specific PCR) primers were designed for the identified missense SNP 54284258 A>G loci. KASP primer set included a common reverse primer and two allele-specific forward primers differentiated by their 3′-terminal nucleotides to target alternate SNP alleles (16). Primers were synthesized by Sangon Biotech (Shanghai, China) Co., Ltd.; sequences are detailed in Table 1.
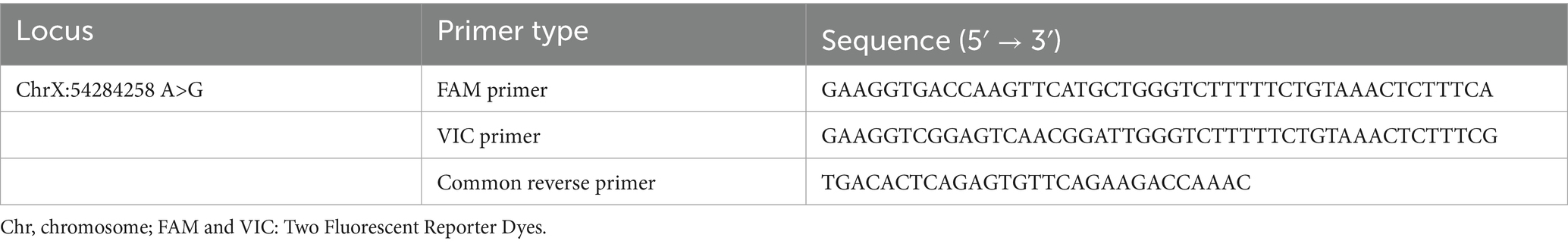
Table 1. KASP primers pairs for BMP15-54284258 genotyping.
The KASP reaction comprises two primary components, Primer Mix and Master Mix. The Primer Mix consists of forward and reverse primers pooled at specified concentrations. The Master Mix contains universal detection primers labeled with distinct fluorophores (e.g., FAM and VIC). PCR amplification for genotyping used a reaction mixture with a total volume of 10 μL with 2.5 μL ddH2O, 0.5 μL of primer mix (FAM/VIC/R primers), 2 μL of DNA, and 5 μL of master mix (17). The cycling temperature adjustment with the respective number of cycles was described in Table 2.
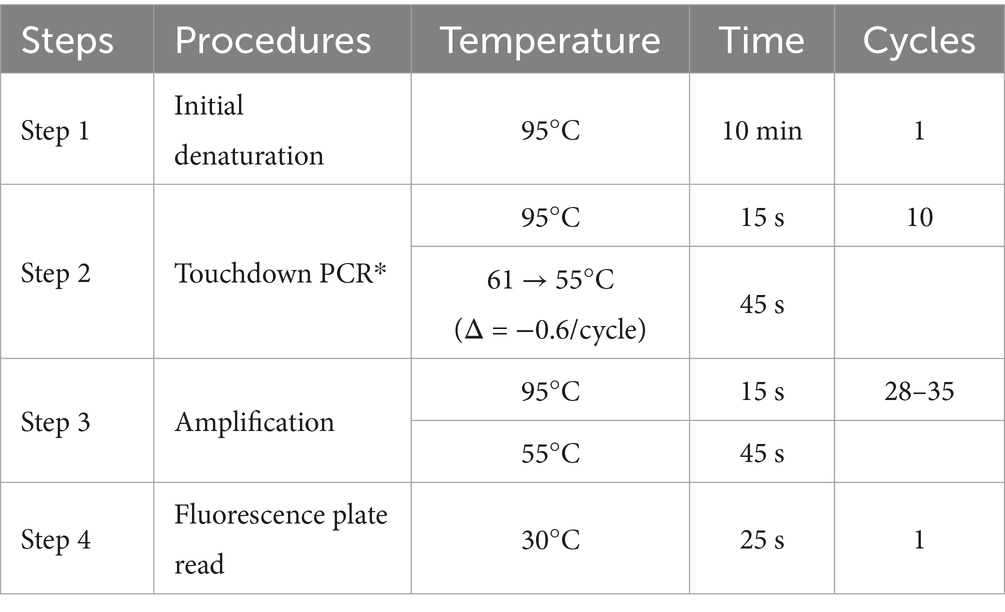
Table 2. KASP reaction protocol.
2.8 Data analysis
The association analysis between lambing traits and the g.54284258A>G SNP of BMP15 gene was analyzed by using the general linear model of SPSS (IBM, 2019). The fixed effects considered were parities and genotype of the ewe. The linear model was Yjkl = μ + Aj + Gk + ejkl, where Yjkl = lambing trait, μ = population mean, Aj = effect of parities of ewe (5 levels; from party 1 to parity 5), Gk = effect of genotype (3 levels), and ejkl = random error. Statistical significance is defined as p < 0.05, and high significance is defined as p < 0.01.
3 Results
3.1 Variant annotation of BMP15
The BMP15 gene is located on the sheep X chromosome (positions 54,283,635–54,290,315 bp). Whole-genome sequencing data from 2,409 sheep were analyzed, identifying 139 SNPs within the BMP15 locus (Supplementary Table S2). Of these, 129 SNPs were localized to intronic regions, while 10 SNPs were exonic (Table 3; Figure 1). Among the exonic variants, 6 were missense mutations, 2 were synonymous mutations, and 2 introduced premature termination codons (PTCs).
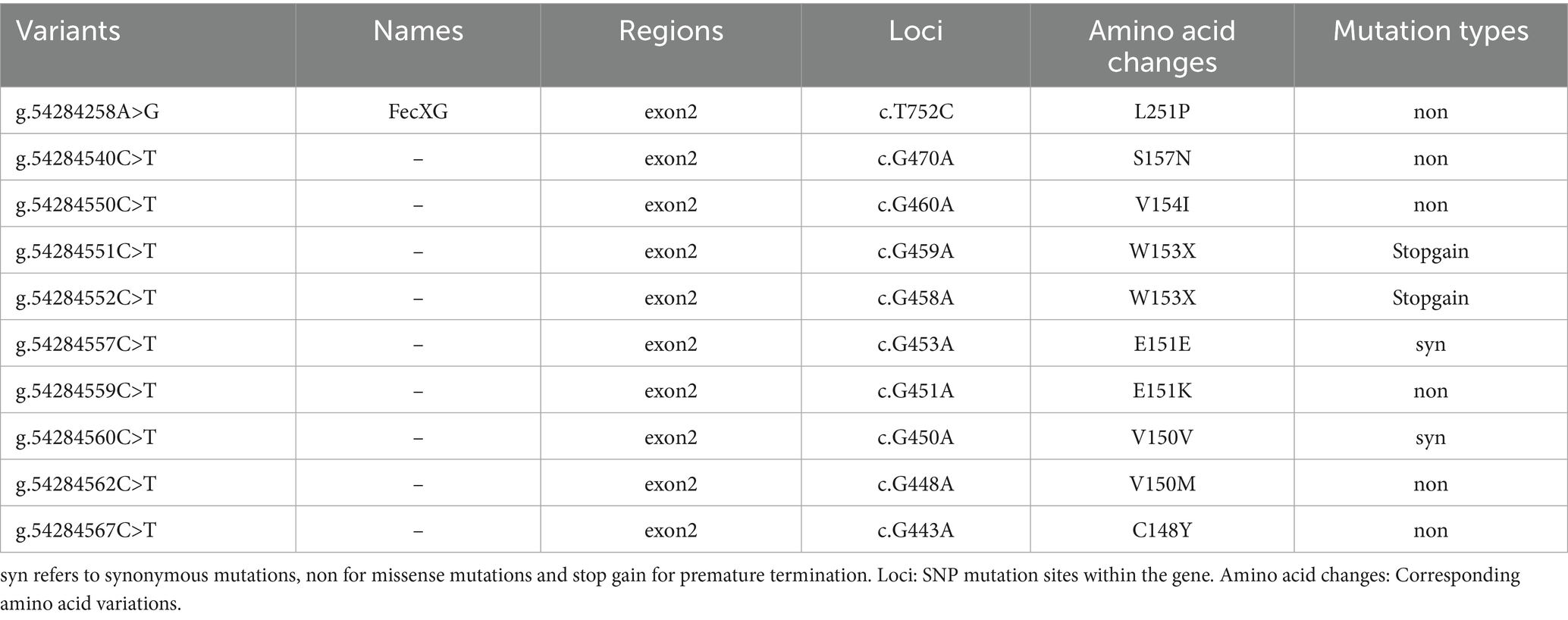
Table 3. SNP variation information in the coding region of sheep BMP15 gene.
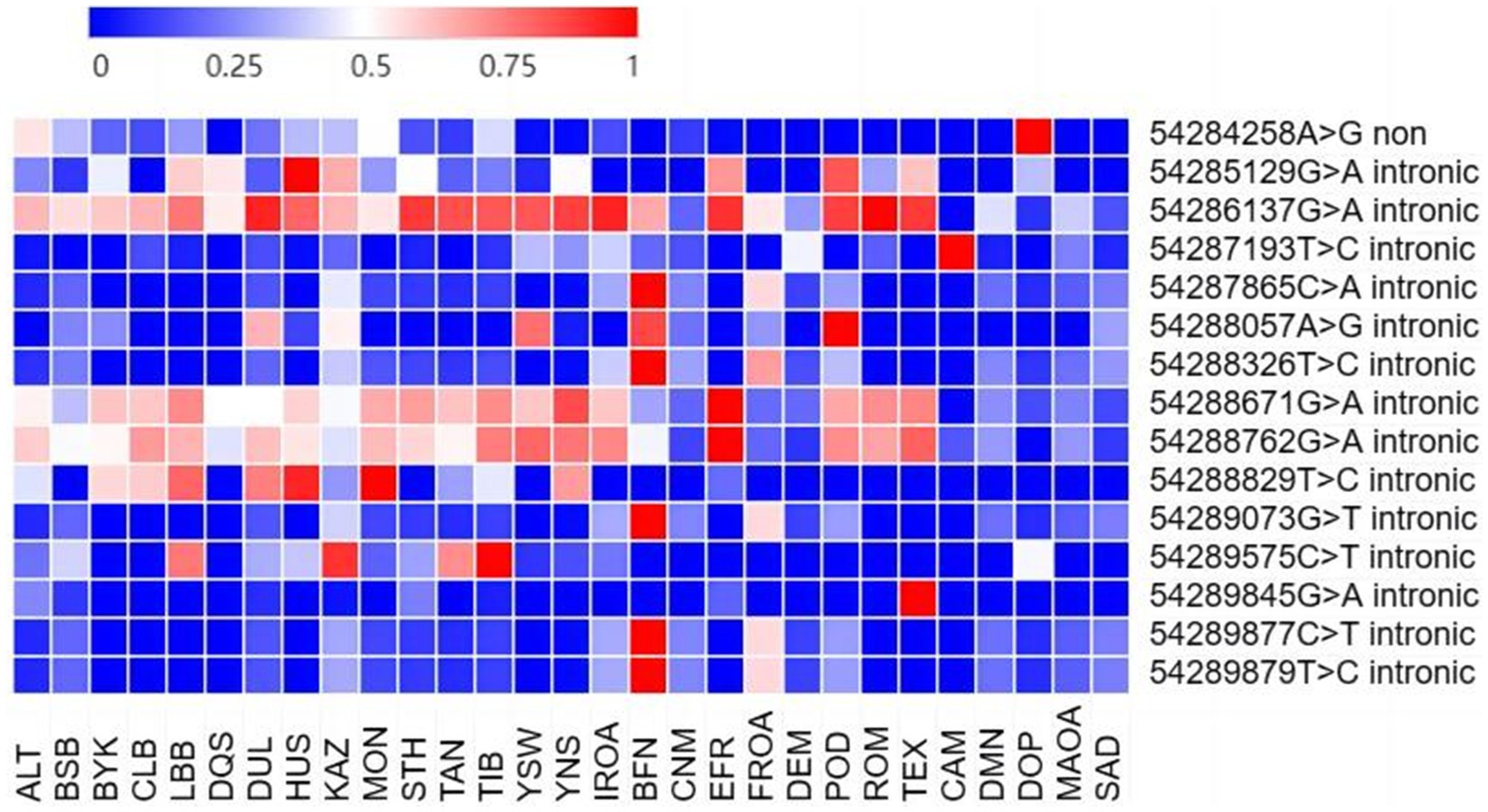
Figure 1. Frequency of SNP variation in BMP15 gene in different sheep populations. Only those populations with sample size >20 and MAF > 0.01 are displayed in figure; ALT, Altay sheep; BSB, Bashibai sheep; BYK, Bayinbuluke sheep; CLB, Cele Black sheep; LBB, Liangshan black sheep; DQS, Diqing sheep; DUL, Duolang sheep; HUS, Hu sheep; KAZ, Kazakh sheep; MON, Mongolian sheep; STH, Small Tail Han sheep; TAN, Tan sheep; TIB, Tibetan sheep; YSW, Yunnan semi-fine wool sheep; YNS, Yunnan sheep; IROA, Iran Local breed sheep; BFN, Baikal Fine-Fleeced sheep; CNM, Chinese Merino sheep; EFR, East Friesian Dairy sheep; FROA, France local populations sheep; DEM, German merino sheep; POD, Poll Dorset sheep; ROM, Romney sheep; TEX, Texel sheep; CAM, Cameroon; DMN, D’man; DOP, Dorper sheep; MAOA, Morocco local populations; SAD, Sardi sheep.
Notably, the SNPs g.54286137G>A, g.54288671G>A, and g.54288762G>A exhibited allele frequencies exceeding 0.5 in 75% of the studied populations. In contrast, g.54289877C>T was not detected (frequency = 0) in most breeds but was present at a frequency exceeding 0.1 in the Altay, Valley Tibetan, and Baikal Fine-Fleeced sheep.
Notably, 9 out of 10 exonic SNPs identified in this study are novel, with only g.54284258A>G having been previously reported (Table 3). The potential functional implications of these newly identified variants need further investigation.
3.2 Prediction of key mutation impact on protein structure
A high-frequency missense mutation (MAF > 0.05) was identified within the BMP15 gene. To evaluate its potential biological impact, the L251P amino acid substitution (leucine to proline at position 251) was modeled using PyMOL v7.0.1, and the resulting protein tertiary structure was compared to the wild-type (Figure 2). Structural predictions revealed detectable alterations in the three-dimensional arrangement of residues near the mutation site, suggesting that this substitution may affect BMP15 function through interference of local folding.
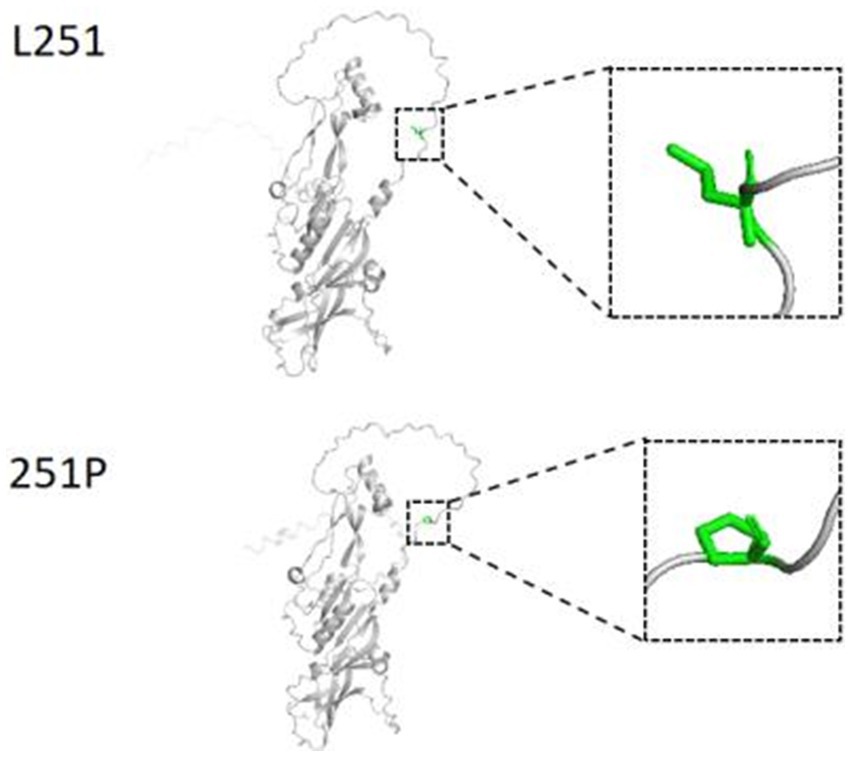
Figure 2. Three-dimensional protein structure changes of L251P before and after mutation. L251, leucine amino acid position at 251 of BMP15; 251P, proline amino acid position at 251 after mutation.
3.3 Haplotype analysis
Four SNPs in the BMP15 gene with MAF > 0.05 such as g.54284258A>G, g.54286137G>A, g.54288671G>A, and g.54288762G>A were used to reconstruct haplotypes. Analysis revealed nine haplotypes (H1–H9), with H6 exhibiting the highest global frequency (0.484) across domestic sheep populations (Table 4; Figure 3). The H1 haplotype was exclusively observed in wild sheep populations (e.g., Argali and Mouflon), strongly supporting its designation as the ancestral haplotype, while H2–H9 represent derived haplotypes arising through mutational events during domestication.
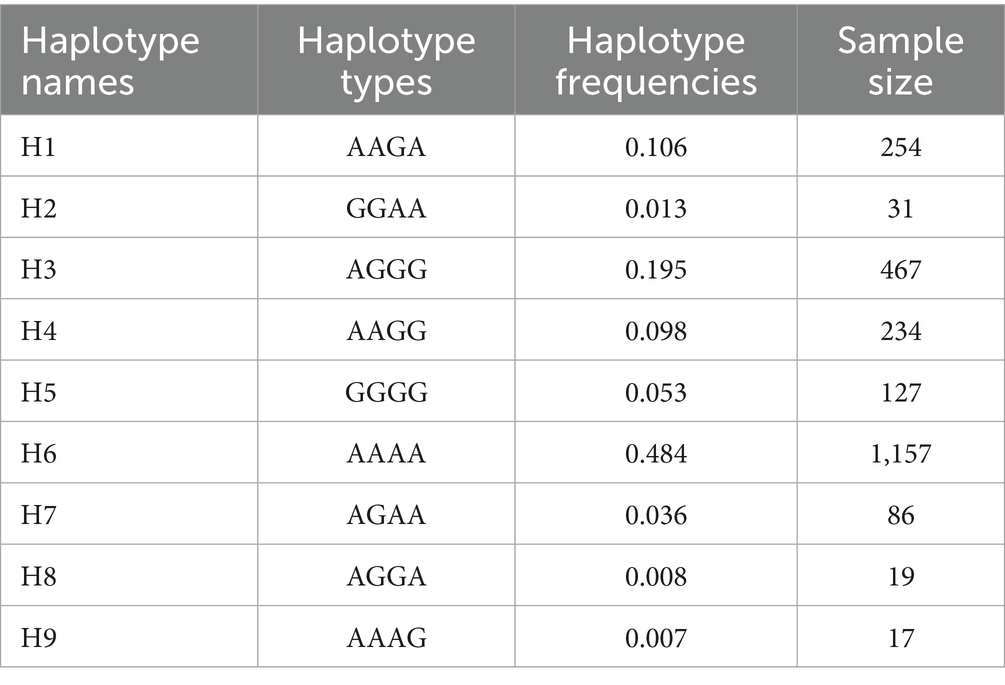
Table 4. Inferred haplotype frequency of BMP15 gene.
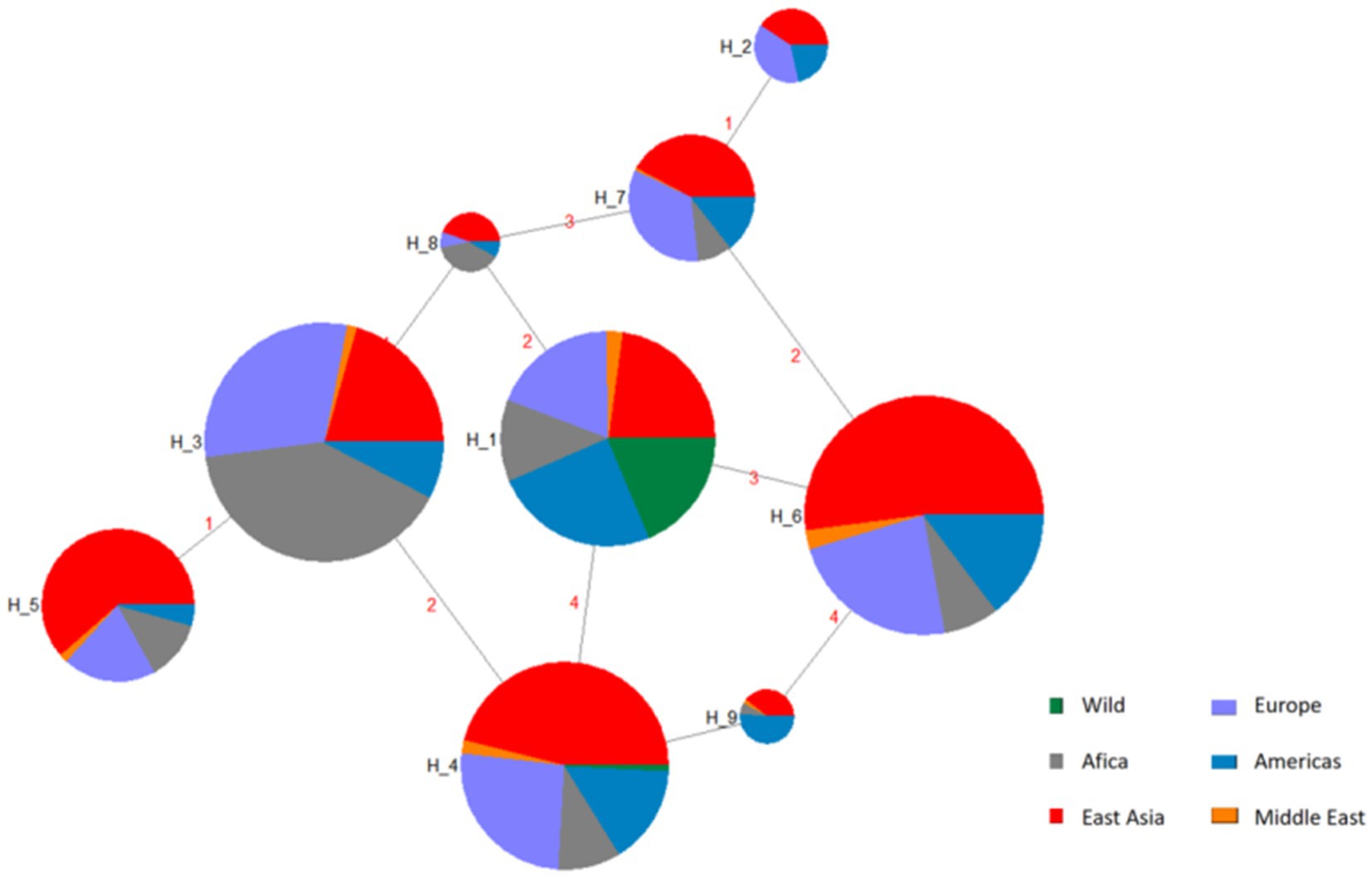
Figure 3. Haplotype networks inferred from SNP mutations in the BMP15 gene, H1 = AAGA, H2 = GGAA, H3 = AGGG, H4 = AAGG, H5 = GGGG, H6 = AAAA, H7 = AGAA, H8 = AGGA, H9 = AAAG. The network connects the haplotypes with lines which indicate their relationship. On the other hand, size of circles indicates frequency of each haplotype.
3.4 Association analysis of BMP15 g.54284258A>G with lambing traits in Australian white sheep
Genotyping of the Australian White sheep population at the BMP15 locus g.54284258A>G was performed using fluorescence signal scanning. The three genotypes (AA, AG, and GG) formed three distinct clusters in the scatter plot (Figure 4), demonstrating high-resolution discrimination and validating the accuracy of the KASP assay.
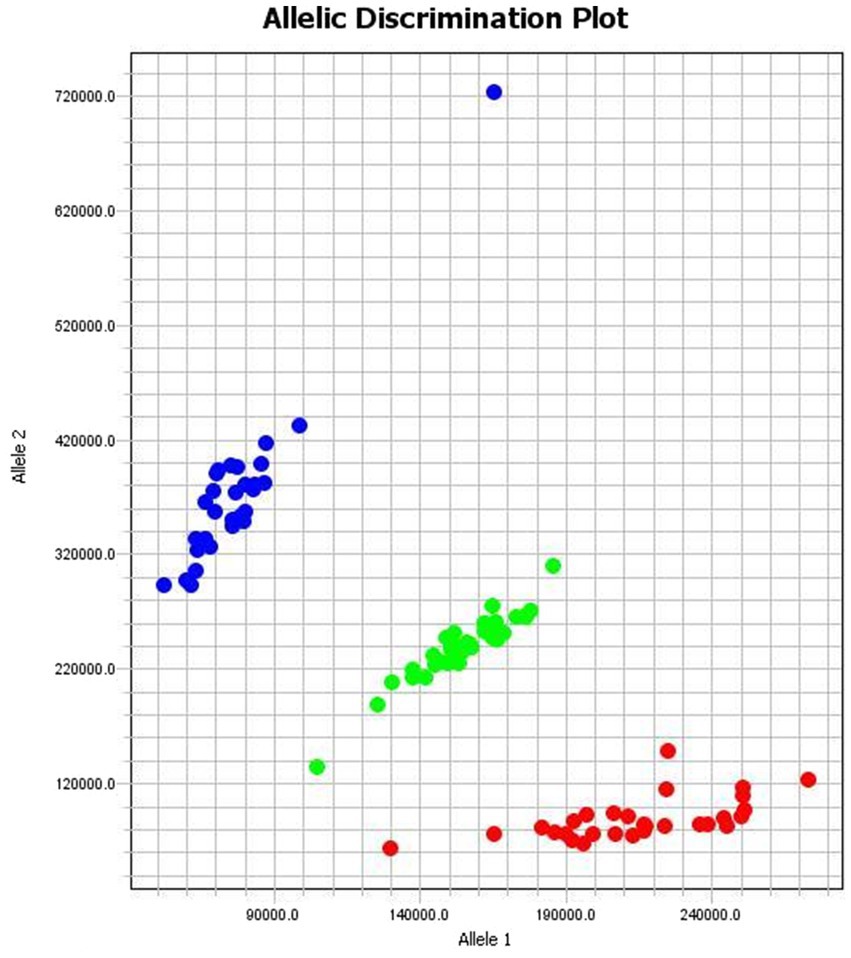
Figure 4. The KASP genotyping results of the g.54284258A>G locus of the sheep BMP15 gene GG in red, AG in green and AA in blue color. Allele 1 at X-axis and Allele 2 at Y-axis represent fluorescence intensities corresponding to two different alleles for each sample. The clear separation between the three clusters indicates that the genotyping has successfully distinguished between the different genotypes.
The BMP15 g.54284258A>G locus showed a significant association of number of live lambs with first and second parities at (p < 0.01) and (p < 0.05), respectively (Table 3). For most parities, the G allele (in both AG and GG genotypes) results in a better number of live lambs than the A allele, except in the fourth parity (Supplementary Table S3). Therefore, the BMP15 g.54284258 A>G variant could be used as a marker for marker-assisted selection to improve the reproductive efficiency of sheep.
4 Discussion
This study identified 139 SNPs within the BMP15 locus in sheep, including 10 exonic variants which comprised 6 missense mutations, 2 synonymous mutations, and 2 premature termination codon (PTC)-introducing mutations. Notably, 9 out of 10 exonic SNPs were novel discoveries, with only g.54284258A>G (FecXG) having been previously reported. The PTC mutations g.54284551C>T (p.W153X) and g.54284552C>T (p.W153X) are predicted to result in complete loss of BMP15 function, analogous to the fertility-impairing effects of known FecX mutations (e.g., FecXI, FecXL) (18). However, these novel truncating mutations exhibited extremely low frequencies in domestic sheep (MAF < 0.005), suggesting potential negative impacts on reproductive efficiency and subsequent purging via natural or artificial selection.
The high-frequency missense mutation g.54284258A>G (p.L251P) was predicted to alter local protein conformation. This discrepancy may be due to two potential mechanisms. First, X-linked dosage effects could lead to partial compensation in heterozygous (AG) females compared to homozygous (AA/GG) individuals, a phenomenon mediated by X-chromosome inactivation (19). Second, indirect mechanisms might operate, where the mutation modulates follicular atresia to influence fetal survival rather than directly increasing ovulation rate—a hypothesis supported by its significant association with first-parity live lamb number (p < 0.01) (20).
Haplotype analysis of four high-frequency SNPs delineated nine haplotypes (H1–H9), with H6 (AAAA) dominating global domestic sheep populations (48.4% frequency), while wild mouflon sheep retained the ancestral haplotype H1 (AAGA). The prevalence of H6 likely reflected long-term artificial selection for enhanced fecundity: BMP15 in wild populations balances ovulation rates to avoid maternal energy depletion, whereas domestication favored haplotypes promoting higher prolificacy. Interestingly, H3 (AGGG) showed elevated frequency in East Asian breeds (19.5%), suggesting potential adaptive advantages in local environments, though functional validation is required.
While g.54284258A>G showed no association with total litter size, its significant effect on number of live lambs (1.29 vs. 1.12 lambs for AG vs. AA genotypes in first parity, p < 0.01) (Table 5) suggested BMP15’s regulatory role in embryonic survival. This dual functionality might originate from two mechanisms. First, BMP15-mediated inhibition of granulosa cell apoptosis, which promotes follicular maturation (21). Second, BMP15 signaling optimization of placental angiogenesis, thereby enhancing fetal nutrient supply and postnatal viability (22). Thus, AG/GG genotypes may improve lamb survival through enhanced placental efficiency rather than direct increases in ovulation. Notably, the enhancing effect of AG/GG genotypes on number live lambs was confined to early parities, with no statistically significant differences observed in subsequent parities. This divergence might be explained by two combined mechanisms. First, maternal life-history trade-offs between reproductive efficiency and survival may lead primiparous individuals to prioritize nutrient allocation to fetal development over self-maintenance, adopting a ‘high-care strategy’ to maximize offspring survival (23). Second, diminished statistical power caused by reduced sample sizes in later parities likely obscured subtle genotypic effect. Therefore, future research should employ large-scale, age-stratified cohort designs with sufficient sample sizes to elucidate the complex interactions among genetic, epigenetic, and environmental factors driving parity-specific phenotypic outcomes.
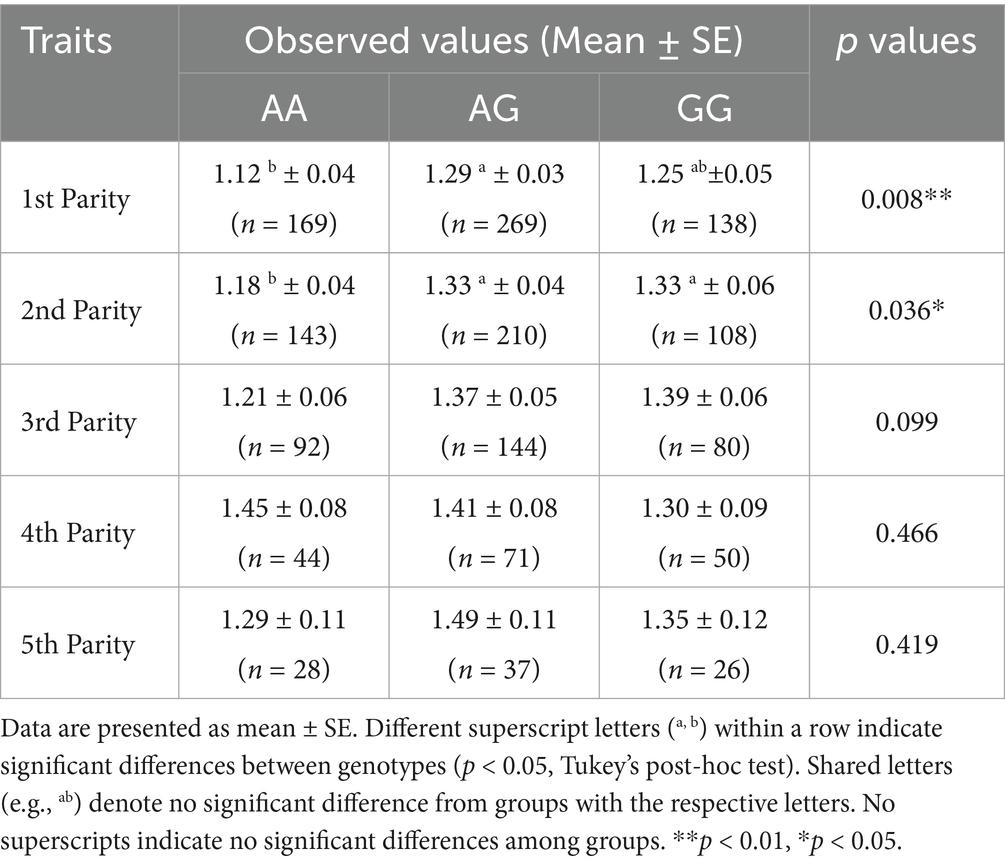
Table 5. Association analysis between the g.54284258A>G locus of the BMP15 gene and the number of live lambs in Australian white sheep.
These findings underscore that functional variants in highly conserved reproductive genes like BMP15 may exist as rare alleles, necessitating large-scale cross-breed meta-analyses for detection. On the other hand, the current research limitations and due attention for future related study included the sample size should be as large as possible, a lack of functional validations like in vitro assays for p.W153X effects on protein secretion, and unable to address epigenetic factors.
5 Conclusion
This comprehensive study elucidates the significant genetic variation landscape within the BMP15 gene across 75 global sheep breeds and its pivotal role in lambing performance, particularly live lamb number. Whole-genome sequencing revealed 139 SNPs, including novel functional variants (6 missense, 2 synonymous, and 2 stop-gain).
Haplotype analysis demonstrated a pronounced divergence between domestic sheep and their wild ancestors, with the H6 (AAAA) haplotype dominating global domestic populations (48.4%), suggesting strong artificial selection for enhanced reproductive efficiency during domestication. Critically, the association analysis established the missense mutation g.54284258A>G (L251P) as a significant genetic marker, specifically linked to increased number of live lambs in both first (p < 0.01) and second (p < 0.05) parities. Ewes carrying the AG genotype produced significantly more live lambs (1.29 vs. 1.12) compared to the AA genotype in the first parity.
While not associated with total litter size, this finding underscores BMP15’s crucial role in influencing postnatal viability, potentially through mechanisms supporting embryonic/placental development. Collectively, the identified genetic variation, ancestral haplotype shift, and the validated association of the L251P variant with key lambing traits highlight BMP15’s considerable potential as a candidate gene for marker-assisted selection to augment reproductive efficiency and live lamb output in sheep breeding programs. Future studies should focus on functional validation of novel variants, expanding association analyses with larger cohorts encompassing diverse breeds and parities, and exploring potential epistatic or epigenetic interactions.
Data availability statement
The original contributions presented in the study are included in the article/Supplementary material, further inquiries can be directed to the corresponding authors.
Ethics statement
All experimental procedures were performed in accordance with the guidelines of the Faculty Animal Policy and Welfare Committee of Northwest A&F University (protocol no. NWAFU-314020038) for the use and care of animals in research. The study was conducted in accordance with the local legislation and institutional requirements.
Author contributions
PL: Investigation, Writing – review & editing, Software, Formal analysis. CC: Formal analysis, Investigation, Software, Writing – review & editing. XW: Software, Writing – review & editing, Formal analysis. KL: Formal analysis, Software, Writing – review & editing. EA: Writing – review & editing. RL: Conceptualization, Resources, Project administration, Writing – review & editing. QZ: Writing – review & editing, Resources. CP: Writing – review & editing, Funding acquisition, Conceptualization, Project administration. XL: Funding acquisition, Validation, Writing – review & editing, Supervision, Project administration, Conceptualization.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This work was funded by the Science and Technology Innovation 2030 Major Project of China (Grant No. 2022ZD040130207).
Acknowledgments
We sincerely thank the Tianjin Aoqun Sheep Industry Academy Company for providing us with AUW sheep samples and litter size data. We thank the High-Performance Computing platform of Northwest A&F University and the Hefei Advanced Computing Center for providing computing resources.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The authors declare that no Gen AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fvets.2025.1612263/full#supplementary-material
Footnotes
1. ^https://www.ncbi.nlm.nih.gov/sra/
2. ^https://www.ncbi.nlm.nih.gov/datasets/genome/GCA_016772045.1/
3. ^https://www.ncbi.nlm.nih.gov/protein/NP_001108239.1/
4. ^https://colab.research.google.com/github/sokrypton/ColabFold/blob/main/AlphaFold2.ipynb
References
1. Li, C, Chen, B, Langda, S, Pu, P, Zhu, X, Zhou, S, et al. Multi-omic analyses shed light on the genetic control of high-altitude adaptation in sheep. Genomics Proteomics Bioinformatics. (2024) 22:qzae030. doi: 10.1093/gpbjnl/qzae030
2. Meyermans, R, Gorssen, W, Aerts, N, Hooyberghs, K, Chakkingal Bhaskaran, B, Chapard, L, et al. Genomic characterisation and diversity assessment of eight endangered Belgian sheep breeds. Animal. (2024) 18:101315. doi: 10.1016/j.animal.2024.101315
3. Baazaoui, I, Bedhiaf-Romdhani, S, Mastrangelo, S, Lenstra, JA, Da Silva, A, Benjelloun, B, et al. Refining the genomic profiles of north African sheep breeds through meta-analysis of worldwide genomic SNP data. Front Vet Sci. (2024) 11:1339321. doi: 10.3389/fvets.2024.1339321
4. Abdoli, R, Ziaeddin Mirhoseini, S, Ghavi Hossein-Zadeh, N, and Zamani, P. Screening for causative mutations of major prolificacy genes in Iranian fat-tailed sheep. Int J Fertil Steril. (2018) 12:51–5. doi: 10.22074/ijfs.2018.5247
5. Zhang, Y, Yan, Z, Qin, Q, Nisenblat, V, Chang, H-M, Yu, Y, et al. Transcriptome landscape of human folliculogenesis reveals oocyte and granulosa cell interactions. Mol Cell. (2018) 72:1021–1034.e4. doi: 10.1016/j.molcel.2018.10.029
6. Gholizadeh, M, and Esmaeili-Fard, SM. Meta-analysis of genome-wide association studies for litter size in sheep. Theriogenology. (2022) 180:103–12. doi: 10.1016/j.theriogenology.2021.12.025
7. Ahlawat, S, Sharma, R, Roy, M, Mandakmale, S, Prakash, V, and Tantia, MS. Genotyping of novel SNPs in BMPR1B, BMP15, and GDF9 genes for association with prolificacy in seven Indian goat breeds. Anim Biotechnol. (2016) 27:199–207. doi: 10.1080/10495398.2016.1167706
8. Song, T, Liu, Y, Cuomu, R, Tan, Y, A Wang, C, De, J, et al. Polymorphisms analysis of BMP15, GDF9 and BMPR1B in Tibetan cashmere goat (Capra hircus). Genes (Basel). (2023) 14:1102. doi: 10.3390/genes14051102
9. Lv, F-H, Cao, Y-H, Liu, G-J, Luo, L-Y, Lu, R, Liu, M-J, et al. Whole-genome resequencing of worldwide wild and domestic sheep elucidates genetic diversity, introgression, and agronomically important loci. Mol Biol Evol. (2022) 39:msab353. doi: 10.1093/molbev/msab353
10. Danecek, P, Auton, A, Abecasis, G, Albers, CA, Banks, E, DePristo, MA, et al. The variant call format and VCFtools. Bioinformatics. (2011) 27:2156–8. doi: 10.1093/bioinformatics/btr330
11. Vens, M, and Ziegler, A. Estimating disequilibrium coefficients. Methods Mol Biol (Clifton, NJ). (2017) 1666:117–32. doi: 10.1007/978-1-4939-7274-6_7
12. Mooers, BHM. Shortcuts for faster image creation in PyMOL. Protein Sci. (2019) 29:268–76. doi: 10.1002/pro.3781
13. Katoh, K, and Standley, DM. MAFFT multiple sequence alignment software version 7: improvements in performance and usability. Mol Biol Evol. (2013) 30:772–80. doi: 10.1093/molbev/mst010
14. Rozas, J, Ferrer-Mata, A, Sánchez-DelBarrio, JC, Guirao-Rico, S, Librado, P, Ramos-Onsins, SE, et al. DnaSP 6: DNA sequence polymorphism analysis of large data sets. Mol Biol Evol. (2017) 34:3299–302. doi: 10.1093/molbev/msx248
15. Bandelt, HJ, Forster, P, and Röhl, A. Median-joining networks for inferring intraspecific phylogenies. Mol Biol Evol. (1999) 16:37–48. doi: 10.1093/oxfordjournals.molbev.a026036
16. Li, Y, Liu, Q, Pan, C-Y, and Lan, X-Y. The free fatty acid receptor 2 (FFA2): mechanisms of action, biased signaling, and clinical prospects. Pharmacol Ther. (2025) 272:108878. doi: 10.1016/j.pharmthera.2025.108878
17. Jiang, E, Zhang, C, He, Z, Zhang, Y, Yang, Y, Pan, C, et al. A novel A-to-G mutation in circBDP1 alters adipocyte proliferation and differentiation and affects bovine carcass traits. J Zhejiang Univ Sci B. (2025). doi: 10.1631/jzus.B2500084
18. Bodin, L, Di Pasquale, E, Fabre, S, Bontoux, M, Monget, P, Persani, L, et al. A novel mutation in the bone morphogenetic protein 15 gene causing defective protein secretion is associated with both increased ovulation rate and sterility in Lacaune sheep. Endocrinology. (2007) 148:393–400. doi: 10.1210/en.2006-0764
19. Liu, M, Zhang, K, and Xu, T. The role of BMP15 and GDF9 in the pathogenesis of primary ovarian insufficiency. Hum Fertil. (2021) 24:325–32. doi: 10.1080/14647273.2019.1672107
20. Peng, J, Li, Q, Wigglesworth, K, Rangarajan, A, Kattamuri, C, Peterson, RT, et al. Growth differentiation factor 9:bone morphogenetic protein 15 heterodimers are potent regulators of ovarian functions. Proc Natl Acad Sci. (2013) 110:E776–85. doi: 10.1073/pnas.1218020110
21. Zhai, B, Liu, H, Li, X, Dai, L, Gao, Y, Li, C, et al. BMP15 prevents cumulus cell apoptosis through CCL2 and FBN1 in porcine ovaries. Cell Physiol Biochem. (2013) 32:264–78. doi: 10.1159/000354435
22. Peng, C. The TGF-beta superfamily and its roles in the human ovary and placenta. J Obstet Gynaecol Can. (2003) 25:834–44. doi: 10.1016/s1701-2163(16)30674-0
Keywords: sheep, BMP15 gene, live lamb number, whole genome sequencing, SNP
Citation: Liu P, Cao C, Wang X, Li K, Areb E, Li R, Zhang Q, Pan C and Lan X (2025) SNP variation landscape of BMP15 gene from 75 global sheep breeds and their genetic effect on lambing traits. Front. Vet. Sci. 12:1612263. doi: 10.3389/fvets.2025.1612263
Edited by:
Maslyn Greene, Clemson University, United StatesReviewed by:
Quratulain Hanif, University of Management and Technology, PakistanLiu Chunjie, Tarim University, China
Copyright © 2025 Liu, Cao, Wang, Li, Areb, Li, Zhang, Pan and Lan. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Xianyong Lan, bGFueGlhbnlvbmc3OUAxMjYuY29t; Chuanying Pan, cGFueXUxOTgwQDEyNi5jb20=
†These authors have contributed equally to this work