 Stefany Barrientos-Villegas
Stefany Barrientos-Villegas María Isabel García-Álvarez
María Isabel García-Álvarez Juana L. VidalLuis M. Gómez-Osorio
Juana L. VidalLuis M. Gómez-Osorio Sara López-Osorio
Sara López-Osorio Jenny J. Chaparro-Gutiérrez*
Jenny J. Chaparro-Gutiérrez*- CIVAB Research Group, Faculty of Agricultural Sciences, University of Antioquia (UdeA), Medellín, Colombia
Objective: To determine the frequency of phenotypic and genotypic resistance to quinolones and fluoroquinolones in Salmonella spp. isolated from production animals (pigs, poultry, cattle) and rodents in South America between 2020 and 2024, with the goal of providing key information on resistance in these countries for public health and food safety.
Methods: A systematic review was conducted following the PRISMA guidelines, using databases such as Scopus, PubMed, SciELO, and Latindex. Studies on Salmonella spp. resistant to quinolones and fluoroquinolones in production animals, meat products, and rodents in South America during 2020–2024 were included.
Results: Of the 83 initial results, 27 studies were selected. 70.4% of the studies were conducted in Brazil. 88% of the studies (n = 24/27) used phenotypic methods, with the disk diffusion technique being the most common. Ciprofloxacin was the most studied antibiotic, with an overall resistance of 32.5%, followed by nalidixic acid (60.6%) and enrofloxacin (23.7%). The average multidrug resistance (MDR) was 62%. 44% of the studies (n = 12/27) employed genotypic methods, with whole genome sequencing (WGS) being the most notable technique. Mutations were reported in parC (58%), gyrA (50%), gyrB (8%), and the presence of qnr genes (75%) and aac(6′)-Ib-cr (8%). No studies on rodents were found.
Conclusion: Resistance to quinolones and fluoroquinolones in Salmonella spp. in South America endangers public health and food safety. To address antimicrobial resistance, monitoring and control measures must be implemented, regional research should be promoted, and stronger restrictions should be enforced.
1 Introduction
Salmonella spp. belongs to the Enterobacteriaceae family, with over 2,600 serotypes reported, affecting a wide range of animals, including humans (1). The Salmonella genus consists of two species: enterica and bongori (2). S. enterica is classified into six subspecies: enterica (subsp. I), salamae (subsp. II), arizonae (subsp. IIIa), diarizonae (subsp. IIIb), houtenae (subsp. IV), and indica (subsp. VI). Subspecies I is associated with more than 99% of the diseases caused by Salmonella in warm-blooded animals, including gastroenteritis and enteric fever (3).
Nontyphoidal salmonellosis has been the most commonly reported zoonotic disease in humans (4). It is acquired through three main routes: food, animal handling on farms or carcass handling in slaughterhouses, contact with pets (dogs and cats), and exotic animals (birds, ferrets, lagomorphs, mustelids, reptiles, and rodents) (5). Salmonella transmission has been primarily linked to contaminated water and food sources, including eggs, meats, and vegetables (6). In this context, the prevention and control of pathogens are ongoing challenges, which is why antimicrobials are frequently used in veterinary medicine to treat and prevent diseases (7). However, there is growing concern that the use of these in animal production may compromise human health through the zoonotic transfer of resistant bacteria via contaminated animal-derived food, direct contact, and their spread in the environment (8).
Nontyphoidal salmonellosis mainly causes self-limiting gastroenteritis in both humans and animals (9). However, when this infection becomes invasive, it requires antibiotic treatment. If the pathogens show resistance, it limits the therapeutic options available for the patient (10). Fluoroquinolones (FQ) have been widely used in clinical practice for the treatment of salmonellosis in both humans and animals (11) and Ciprofloxacin is the first-line antibiotic used to treat both typhoidal and nontyphoidal salmonellosis in humans (9, 12). However, the emergence of resistance or multidrug resistance (MDR) to these antibiotics has become a critical issue in the clinical treatment of the disease (13). This is why the World Health Organization (WHO) classifies fluoroquinolone-resistant Salmonella as a high-priority pathogen to support research and the development of new antibiotics (12, 14).
Resistance to fluoroquinolones in Salmonella can occur due to mutations in the quinolone resistance-determining regions (QRDR) of the chromosomal gyr and par genes, resulting in a reduced binding affinity of the topoisomerase enzymes to quinolones (15). Secondly, plasmid-mediated quinolone resistance (PMQR) involves the acquisition of (i) qnr genes (qnrA, qnrB, qnrS, qnrC, qnrD), which encode topoisomerase-binding proteins that provide physical protection against quinolones, (ii) the aac(6′)-Ib-cr gene, which encodes a modifying enzyme that reduces the activity of fluoroquinolones, and (iii) oqxAB and qepA, which encode quinolone efflux pumps. Finally, the negative and positive regulation of porins encoded by chromosomal genes or the efflux pumps of multiple drugs (AcrAB-TolC), respectively, reduce intracellular concentrations of fluoroquinolones (14).
In South America, the dynamics of antimicrobial resistance (AMR) in Salmonella within the animal sector—particularly in production animals and across the food supply chain—are poorly characterized. Countries such as Venezuela, Guyana, French Guiana, and Suriname reported very limited research between 2020 and 2024, highlighting significant gaps in AMR surveillance and data. Understanding the current status of fluoroquinolone-resistant Salmonella is especially critical, given the scarcity and fragmentation of existing evidence. This study aims to consolidate available data on phenotypic and genotypic resistance to quinolones and fluoroquinolones in Salmonella isolated from pigs, poultry, cattle, and rodents. The inclusion of rodents is supported by their established role as reservoirs and amplifiers of zoonotic pathogens in agricultural environments. By providing a unified analysis, this review contributes to a clearer understanding of the regional AMR landscape and supports the development of targeted public health strategies, including improved biosecurity and responsible antibiotic use on farms.
2 Materials and methods
2.1 Study search
The study was conducted following the guidelines established in the Preferred Reporting Items for Systematic Reviews and Meta-Analyses (PRISMA)(16). The study populations included Salmonella isolates from production animals such as cattle, poultry, pigs, meat products from these animals, and rodents. The primary outcome of interest was the reported frequencies of phenotypic and genotypic resistance to FQ: non-susceptibility to nalidixic acid (Nal ns), non-susceptibility to ciprofloxacin (Cip ns), non-susceptibility to enrofloxacin (Enr ns), frequency of mutations in QRDR genes, and the presence or absence of PMQR genes. The secondary outcomes included MDR, reported serotypes and sequence types (ST), amino acid substitutions in mutated genes, and the phenotypic and genotypic techniques used. MDR was defined as resistance to three or more drugs.
A literature search was conducted in English, Spanish, Portuguese, and French using Boolean logic tools with the operators “AND” and “OR” to search for relevant articles in the PubMed, Scopus, SciELO, and Latindex databases. The search aimed to identify pertinent articles published from January 1, 2020, to August 24, 2024. The search string that allowed for the identification of most studies was as follows: Salmonella AND (quinolone OR fluoroquinolone OR ciprofloxacin OR nalidixic acid OR enrofloxacin) AND (livestock OR cattle OR swine OR pig OR poultry OR rodent OR rat OR beef OR chicken OR pork OR meat) AND (Peru OR Brazil OR Colombia OR Ecuador OR Chile OR Venezuela OR Argentina OR Uruguay OR Bolivia OR Guyana OR Paraguay OR French Guiana OR Suriname). The search was conducted on August 24, 2024. Additional articles were also included, manually located in the Scopus, PubMed, SciELO, and Latindex databases. Additional articles were also included through manual searches of reference lists from selected studies and relevant journals.
2.2 Study selection
The study selection was carried out by two independent reviewers (SBV and MIG), and the references were exported to the Rayyan online application software for screening and selection. In the first phase of review (screening), titles and abstracts were evaluated to identify studies related to the primary outcomes of interest. At this stage, exclusion criteria were applied to discard studies whose titles and/or abstracts were not relevant. Discrepancies between reviewers were resolved through discussion or, if necessary, by consulting a third reviewer.
In the second phase (eligibility), a full-text review of the selected articles was conducted, with detailed assessment based on the following eligibility criteria: (i) publication in English, Spanish, Portuguese, or French; (ii) inclusion of phenotypic and/or genotypic determinants of fluoroquinolone resistance; (iii) isolation of Salmonella from production animals, meat products, or rodents; (iv) exclusion of incomplete or unclear studies; (v) exclusion of studies conducted outside South America; and (vi) inclusion of studies published before August 24, 2024.
Duplicate references were identified and removed using EndNote software prior to the screening process. Additionally, data extraction was performed by one reviewer and independently validated by a second reviewer to minimize errors or inconsistencies. To assess the risk of bias and methodological quality of the included studies, we used the Joanna Briggs Institute (JBI) Critical Appraisal Checklist for prevalence studies. Two independent reviewers (SBV and MIG) performed the quality assessment, and disagreements were resolved by consensus or by involving a third reviewer. Studies were not excluded based on quality, but the appraisal results were considered when interpreting the findings.
2.3 Data extraction
The following data were considered and extracted: (i) study identifier: Title, authors, year of publication, country, species (production animals, meat animals, or rodents); (ii) Methods: sample type, sample size, identification method, antimicrobial susceptibility testing (fluoroquinolones or quinolones), breakpoint/interpretive standard level, and gene detection (phenotype-based/genotype-based); (iii) Results: number of isolates, number of isolates tested for susceptibility, number of MDR strains, number of strains resistant to nalidixic acid (Nal ns), number of strains resistant to ciprofloxacin (Cip ns), number of strains resistant to enrofloxacin (Enr ns), number of strains examined for mutation detection (gyrA, gyrB, parC y parE), number of mutants, mutation positions, substituted amino acids, number of strains examined for plasmid-mediated quinolone resistance genes (PMQR) (qnrA, qnrB, qnrC, qnrD, qnrS, aac(6′)-Ib-cr, qepA, oqxA/B) and number of strains with PMQR genes.
2.4 Data analysis
Data were analyzed using descriptive statistics in Microsoft Excel (Microsoft 365®). Additionally, GIS software (QGIS 3.16.15) was used to generate maps illustrating the distribution of resistance patterns across South America.
Inferential statistical analyses were not applied in this study due to the high heterogeneity among the included articles in terms of study design, sample sizes, animal species, sampling matrices, and laboratory methodologies. As a result, quantitative synthesis through meta-analysis was not feasible. A descriptive approach was used instead, in line with the exploratory nature of this review.
3 Results
Our search strategy yielded a total of 83 results, with 57 found in PubMed, 18 in Scopus, and 8 in SciELO. After excluding 46 articles based on their title and abstract, 37 were selected for full-text reading, and of these, only 10 were excluded, resulting in a total of 27 articles included in this study. The main reason for excluding articles during the selection process was that they did not analyze the Salmonella agent or did not focus on relevant animal matrices of interest (Figure 1).
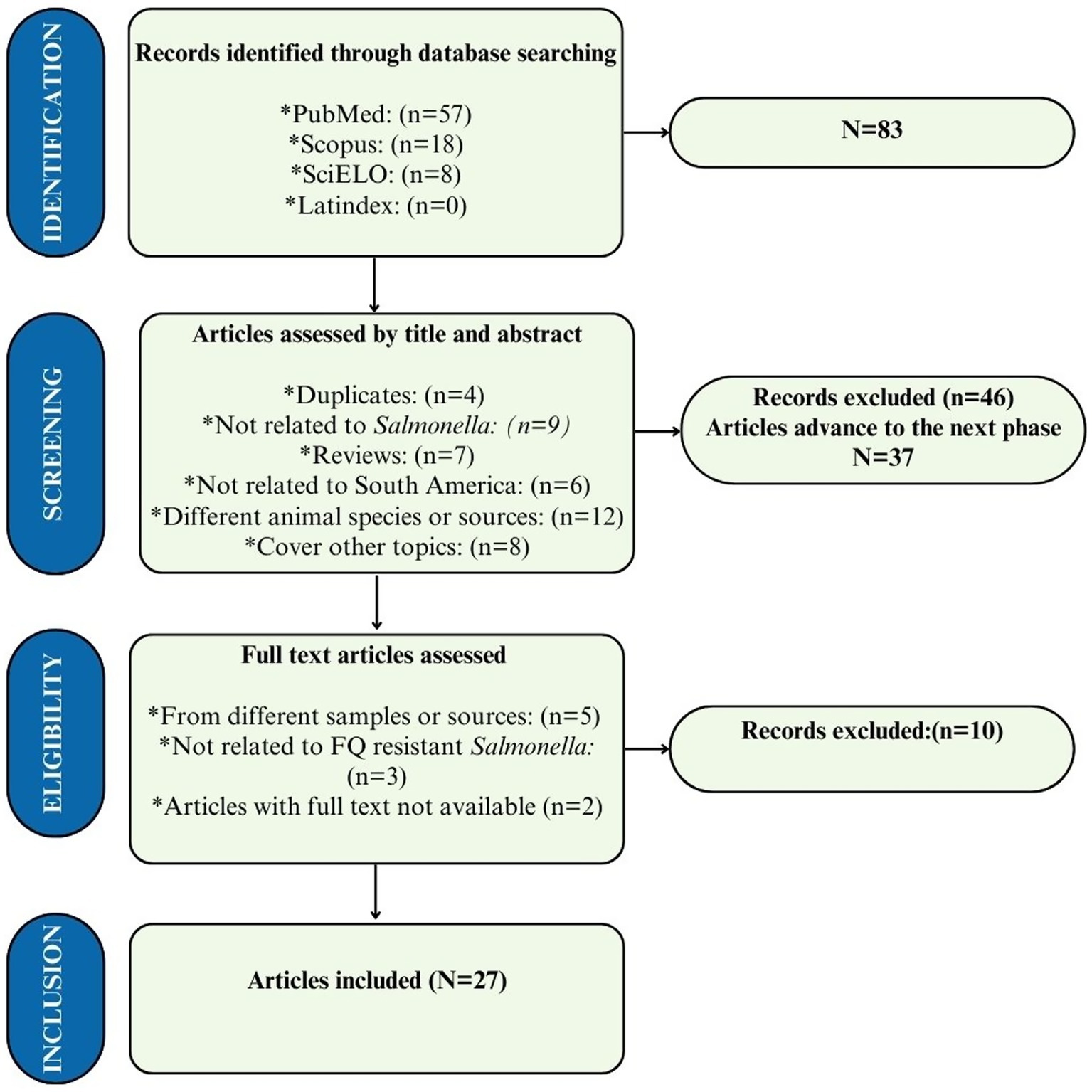
Figure 1. PRISMA flow diagram for study categorization and selection of the 27 studies included in this systematic review. Data came from databases between 2020 and 2024.
According to our results, 70.4% (n = 19/27) of the studies were conducted in Brazil, followed by Argentina and Colombia with 7.4% (n = 2/27) each. Next, Paraguay, Uruguay, Chile, and Ecuador each represented 3.7% (n = 1/27).
Regarding the species studied, 40.74% of the articles focused on swine and its products (n = 11/27), a percentage equal to that of poultry and its products (n = 11/27). Cattle and their products were investigated in 3.7% (n = 1/27), while 14% (n = 4/27) involved a combination of matrices from different species (swine, cattle, and poultry). No published research was found regarding rodents.
3.1 Phenotypic resistance to quinolone and fluoroquinolone in Salmonella
Of the 27 studies, 88% (n = 24/27) employed phenotypic methods to detect fluoroquinolone resistance in Salmonella isolates the most commonly method used to assess AMR was disk diffusion (17), in 75% (n = 18/24) of the studies, followed by broth microdilution (18) in 25% (n = 6/24) of the studies, according to CLSI guidelines. The most studied fluoroquinolone was ciprofloxacin, present in 91.6% (n = 22/24) of the studies, with an overall resistance of 32.53%. It was followed by nalidixic acid, studied in 15 of the 24 studies, which showed an overall resistance of 60.6%. In contrast, enrofloxacin was the least evaluated, being analyzed in 10 of the 24 studies, with an overall resistance of 23.74%. MDR was assessed in 17 of the 24 studies (n = 17/24), with an overall percentage of 62%.
3.1.1 Poultry isolates
The most used sample type in poultry studies was a combination of matrices from various sources, accounting for 37.5% (n = 3/8), followed by feces and chicken meat. The most frequently isolated serotype was Salmonella Heidelberg, reported in 62.5% of the studies (n = 5/8), with average resistances in chicken meat of 96.3, 38.9% to nalidixic acid and ciprofloxacin, respectively. In feces, resistances of 54% to ciprofloxacin were found (Table 1).
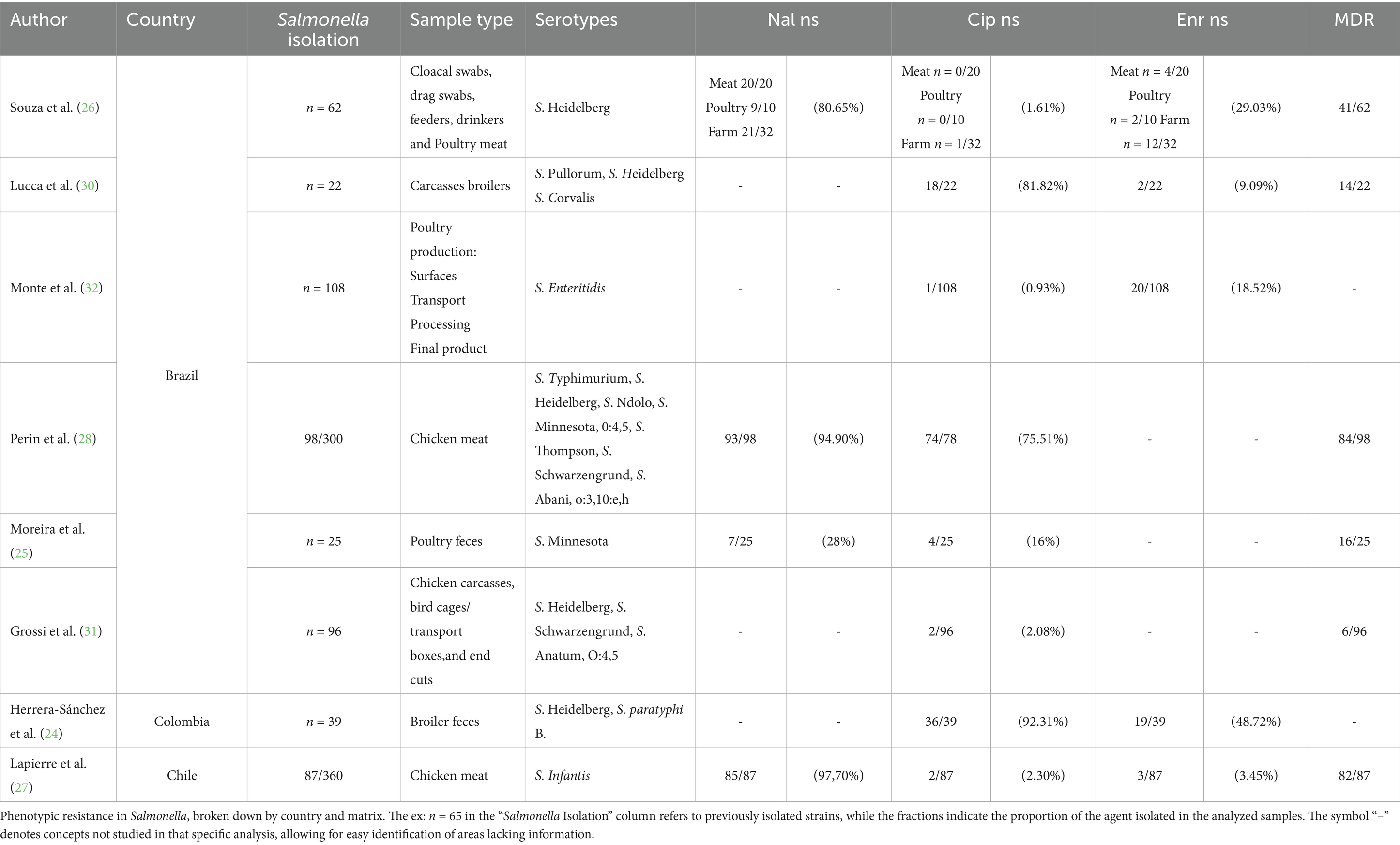
Table 1. Phenotypic resistance to FQ into Salmonella in poultry isolates in South America (2020–2024).
3.1.2 Pigs isolates
The most commonly used sample type in swine studies was a combination of matrices from various sources, representing 45% (n = 5/11) of the cases. The most frequently reported serotype was S. typhimurium, found in 81.8% (n = 9/11) of the isolates, followed by S. Derby, which appeared in 63.6% (n = 7/11). On average, in the mesenteric lymph nodes, the most studied individual matrix, resistances of 44, 40, and 37% were observed for nalidixic acid, ciprofloxacin, and enrofloxacin, respectively (Table 2).
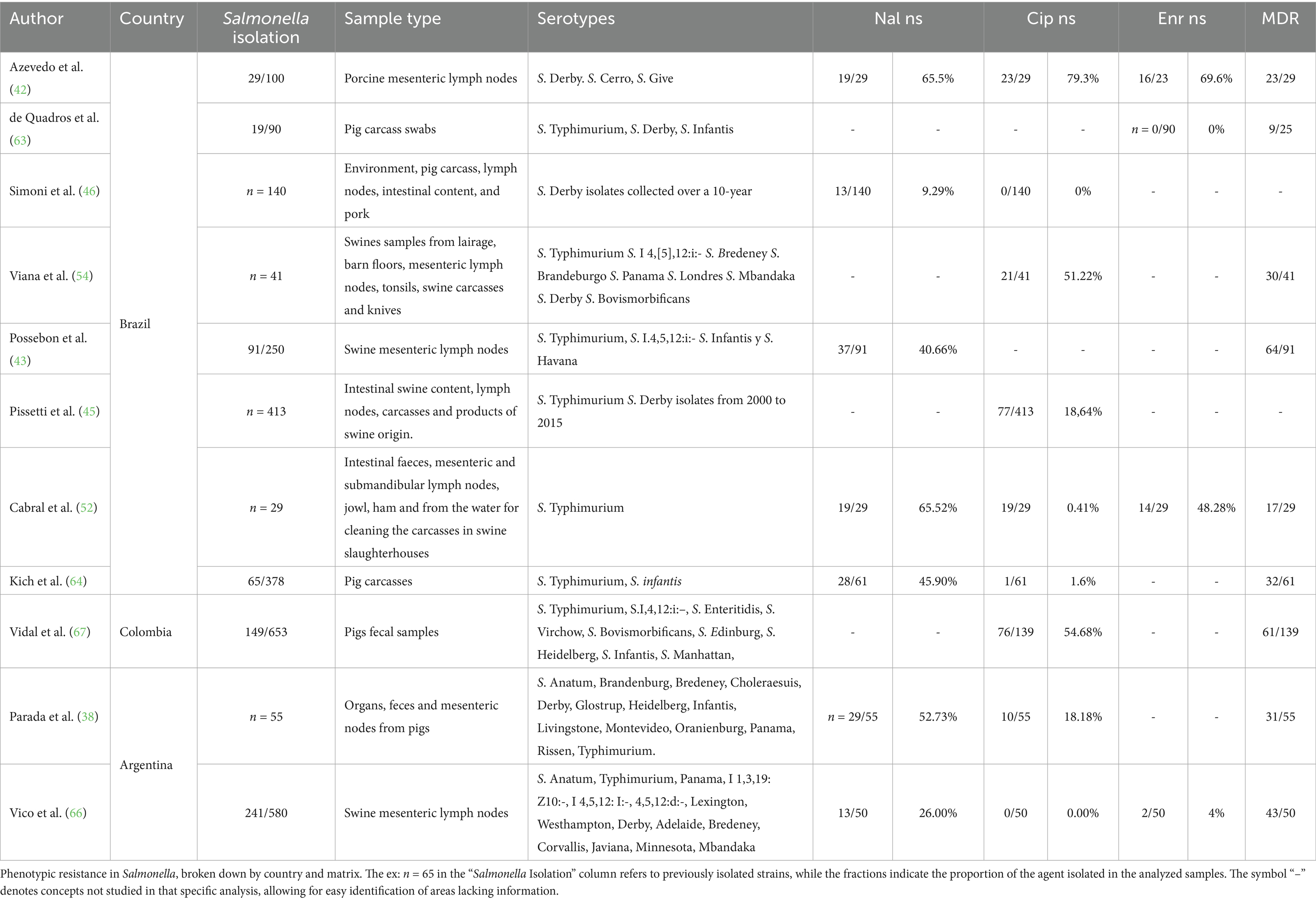
Table 2. Phenotypic resistance to FQ into Salmonella in pig isolates in South America (2020–2024).
3.1.3 Cattle isolates
According to a study conducted in Uruguay isolates with non-susceptibility to fluoroquinolones were reported, with 77.3% (n = 58/75) of the isolates being non-susceptible to ciprofloxacin, which is a second-generation fluoroquinolone. Additionally, in this group, 6.6% (n = 5/75) were non-susceptible to enrofloxacin. 56% (n = 42/75) were MDR (Table 3).
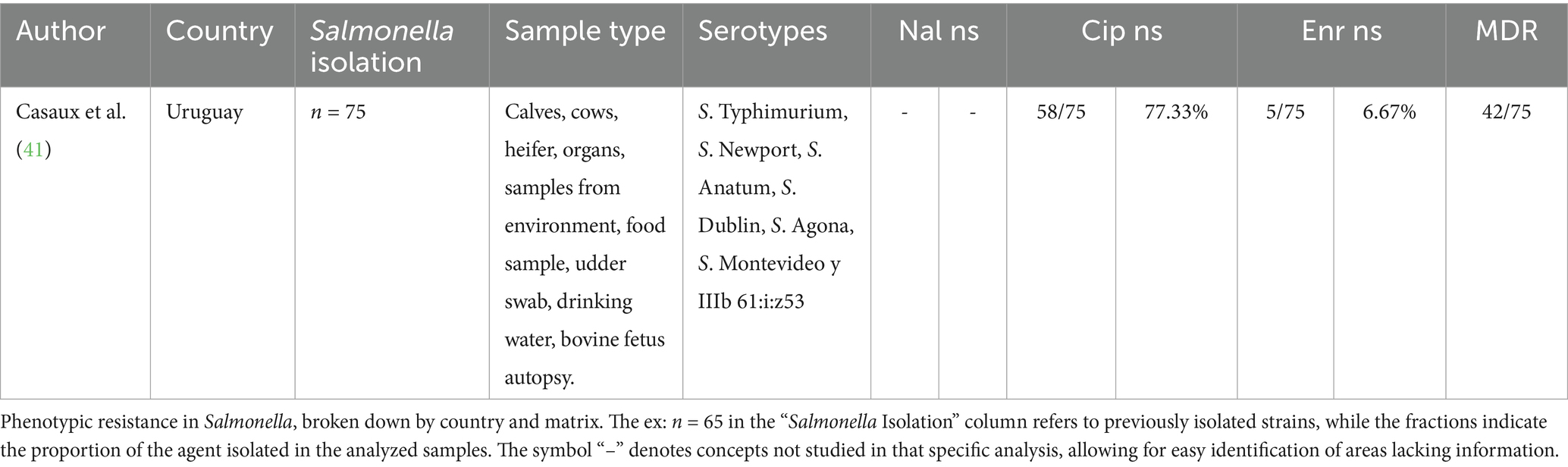
Table 3. Phenotypic resistance to FQ Salmonella in cattle isolates in South America (2020–2024).
3.1.4 Combined matrices
Of the 24 studies that used phenotypic methods, 4 of them used matrices involving a mixture of different species, primarily from their meat products. The average resistance to nalidixic acid was 75%, and to ciprofloxacin was 35% (Table 4).
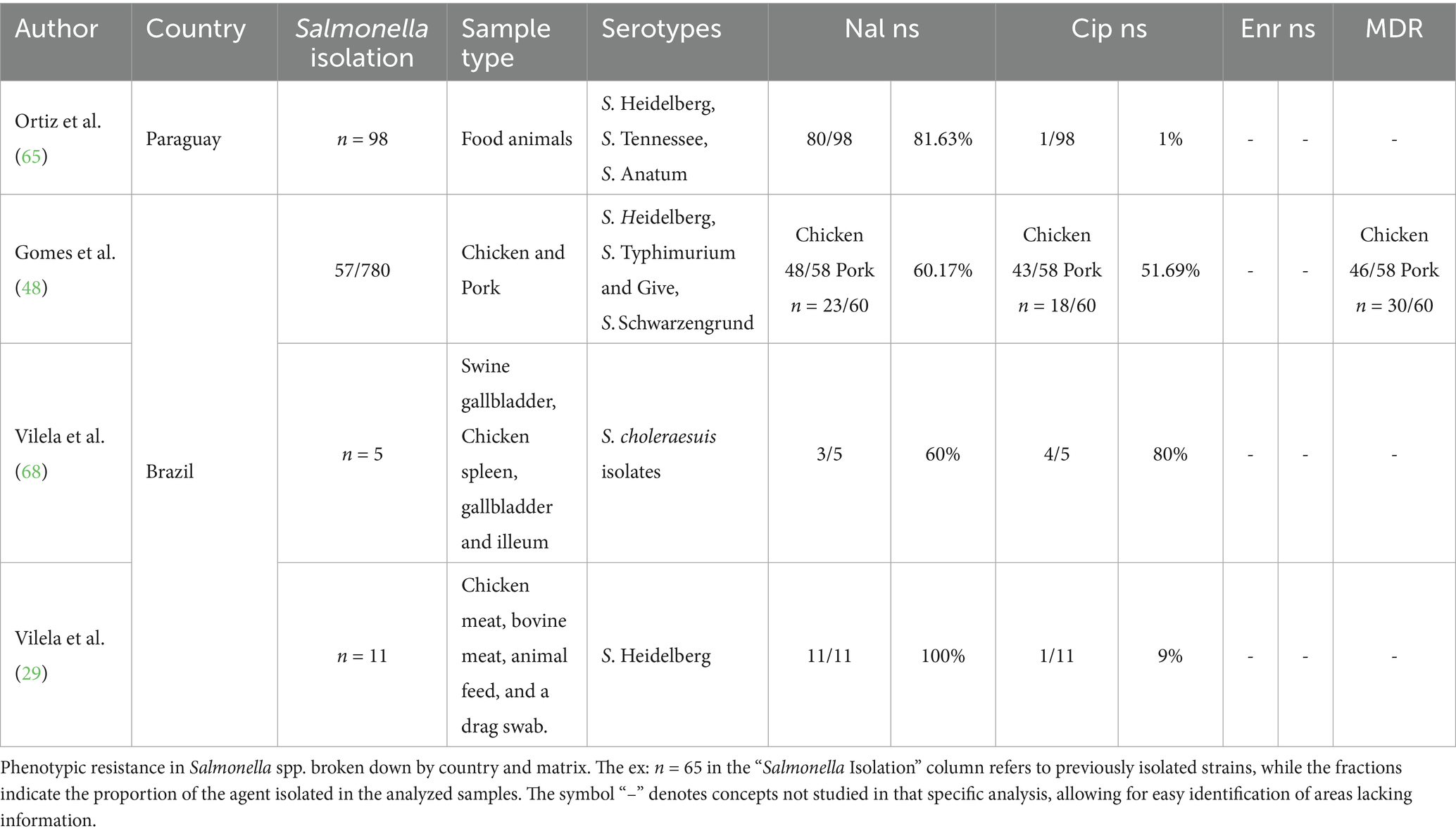
Table 4. Phenotypic resistance in Salmonella in combined matrices in South America (2020–2024).
3.2 Genotypic resistance to quinolone and fluoroquinolone in Salmonella
Of the 27 studies reviewed, 44% (n = 12/27) employed genotypic techniques to detect resistance genes in Salmonella. The most commonly used methodology was WGS, with the Illumina MiSeq platform being the most widely used, employed in 71% (n = 5/7) of the studies that applied Whole Genome Sequencing (WGS), followed by Illumina HiSeq. In comparison, the PCR technique was used in a smaller number of studies, as shown below. In all of the studies that used these techniques, mutations were found in gyrA 50% (n = 6/12), parC 58% (n = 7/12), gyrB 8% (n = 1/12), qnr genes 75% (n = 9/12), and aac(6′)-Ib-cr 8% (n = 1/12).
3.2.1 Polymerase chain reaction (PCR) techniques
PCR involves DNA extraction, amplification with specific primers, and thermal cycling, followed by detection via gel electrophoresis or real-time PCR (19). This rapid and sensitive technique enables precise detection of microorganisms and resistance genes, even in low-DNA samples (20). Of the 27 studies, only 19% (n = 5/27) used PCR techniques or molecular identification of resistance genes, and all of them correspond to different South American countries. Among the most commonly investigated genes are PMQR genes, highlighting qnrB, which was present in 100% of the studies that employed molecular methods, where the most commonly used primer sequences were: qnrb-F GATCGTGAAAGCCAGAAAGG and qnrb-R ACGATGCCTGGTAGTTGTCC (21).
In two of these five studies, QRDRs (quinolone resistance-determining regions) genes were identified, which are chromosomal and result from mutations in the gyr and par genes. Of these mutations, the most frequently investigated was in the gyrA gene, which was analyzed in two of the five studies that used these techniques and primarily presented mutations at position 83. The parC gene was only investigated in one study, where mutations T57S were found (Table 5).
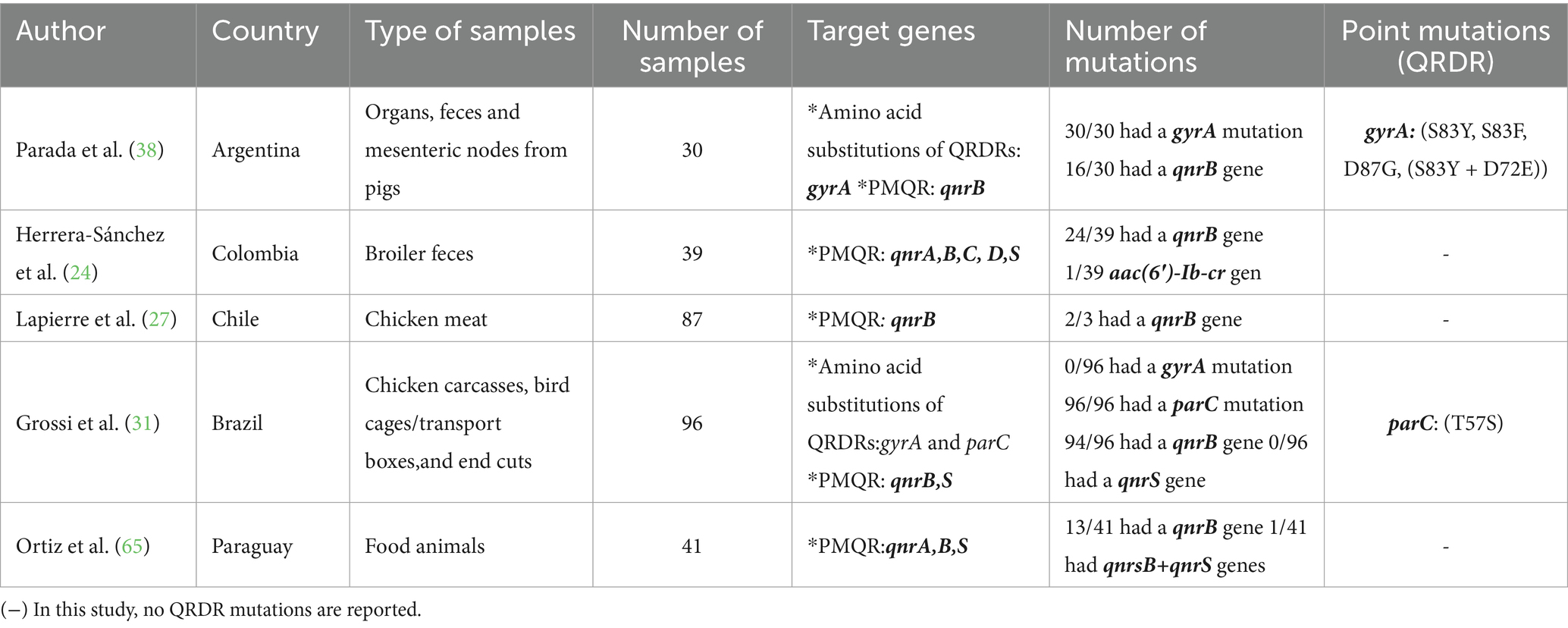
Table 5. Genes conferring resistance to quinolones and fluoroquinolones identified in Salmonella spp. using PCR techniques in South America between 2020 and 2024.
3.2.2 Whole genome sequencing (WGS)
WGS involves DNA extraction, library preparation, sequencing (Illumina/Nanopore), genome assembly, and bioinformatics analysis to detect genetic variants and resistance genes (22). This high-resolution technique enhances epidemiological surveillance and pathogen control by identifying resistance mechanisms and phylogenetic relationships (23).
Of the 27 studies analyzed, only 25.9% (n = 7/27) implemented WGS, mostly in poultry isolates. Mutations in QRDR genes were identified in all the studies, with the gyrA gene being involved in 71% of the cases. The most frequent mutation in this gene occurred at position 83, reported in 57% of the studies, with changes in various amino acids. On the other hand, mutations in the parC gene were observed in 71% of the studies, with the most common being the one at position 57, where in all cases, a threonine to serine change was detected. Regarding PMQR-mediated resistance genes, the qnrB19 gene was the most reported, present in 43% of the studies. Additionally, different associated plasmid replicons were identified (Table 6).
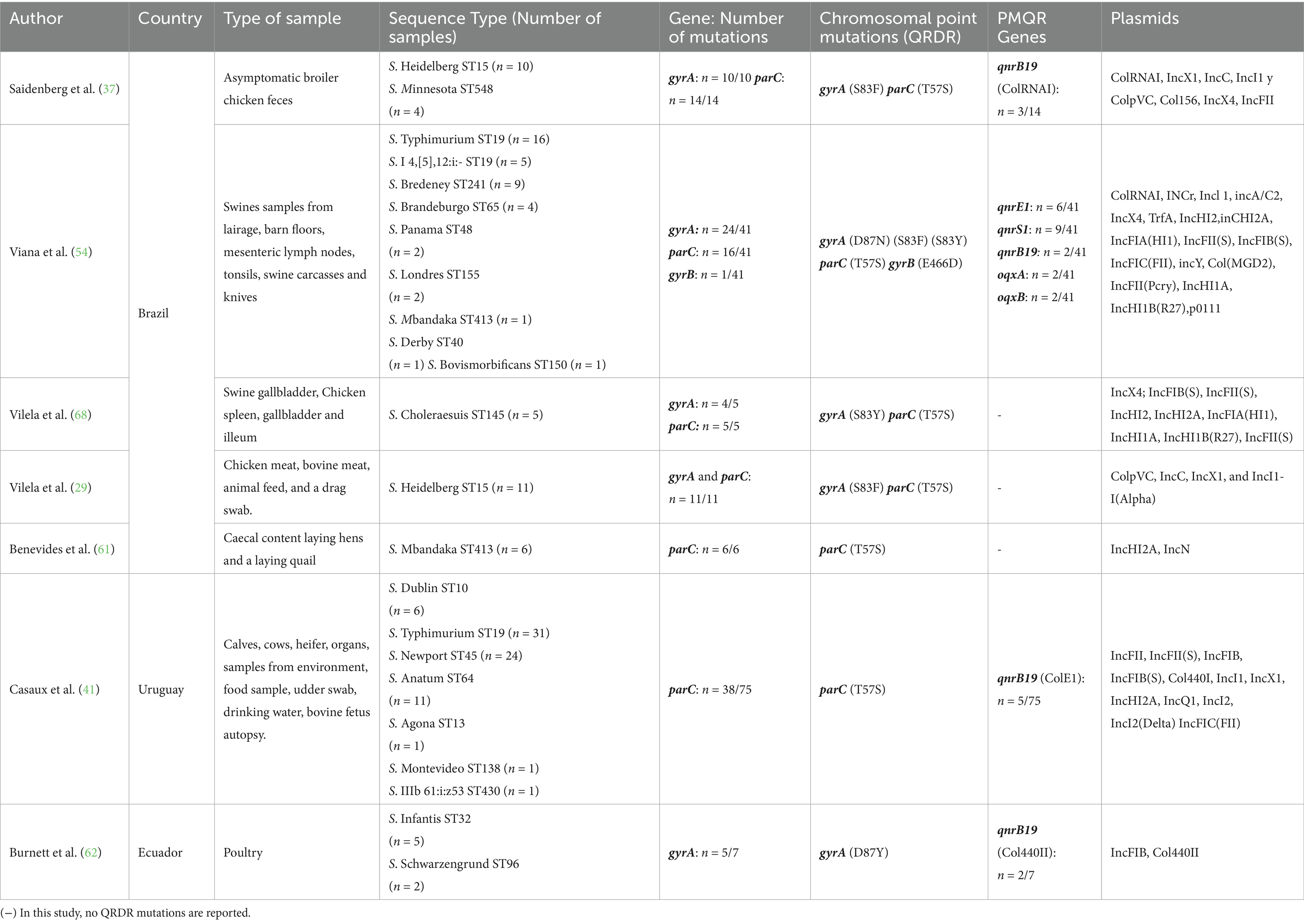
Table 6. Genes conferring resistance to quinolones and fluoroquinolones identified in Salmonella spp. using WGS Techniques in South American countries between 2020 and 2024.
These findings reveal a recurring pattern of specific chromosomal mutations (gyrA S83 and parC T57) and plasmid-mediated resistance (qnrB19) in isolates, which may indicate clonal spread or horizontal gene transfer in the food production chain. The frequent detection of these markers underscores the need for routine WGS-based surveillance in high-risk reservoirs to guide more targeted interventions in antimicrobial resistance control.
As previously mentioned, resistance to ciprofloxacin is of great importance, as it is the most studied antibiotic in this study and the first-line treatment for both typhoidal and non-typhoidal Salmonella spp. infections in humans. Between 2020 and 2024 in South America, a total of 490 Salmonella strains were found to be resistant to the antibiotic through phenotypic antibiogram testing, out of 1,781 tested strains, resulting in an overall resistance rate of 27.5%. Specifically, 307 resistant strains were reported in Brazil, 112 in Colombia, 58 in Uruguay, 10 in Argentina, 2 in Chile, and 1 in Paraguay (Figure 2).
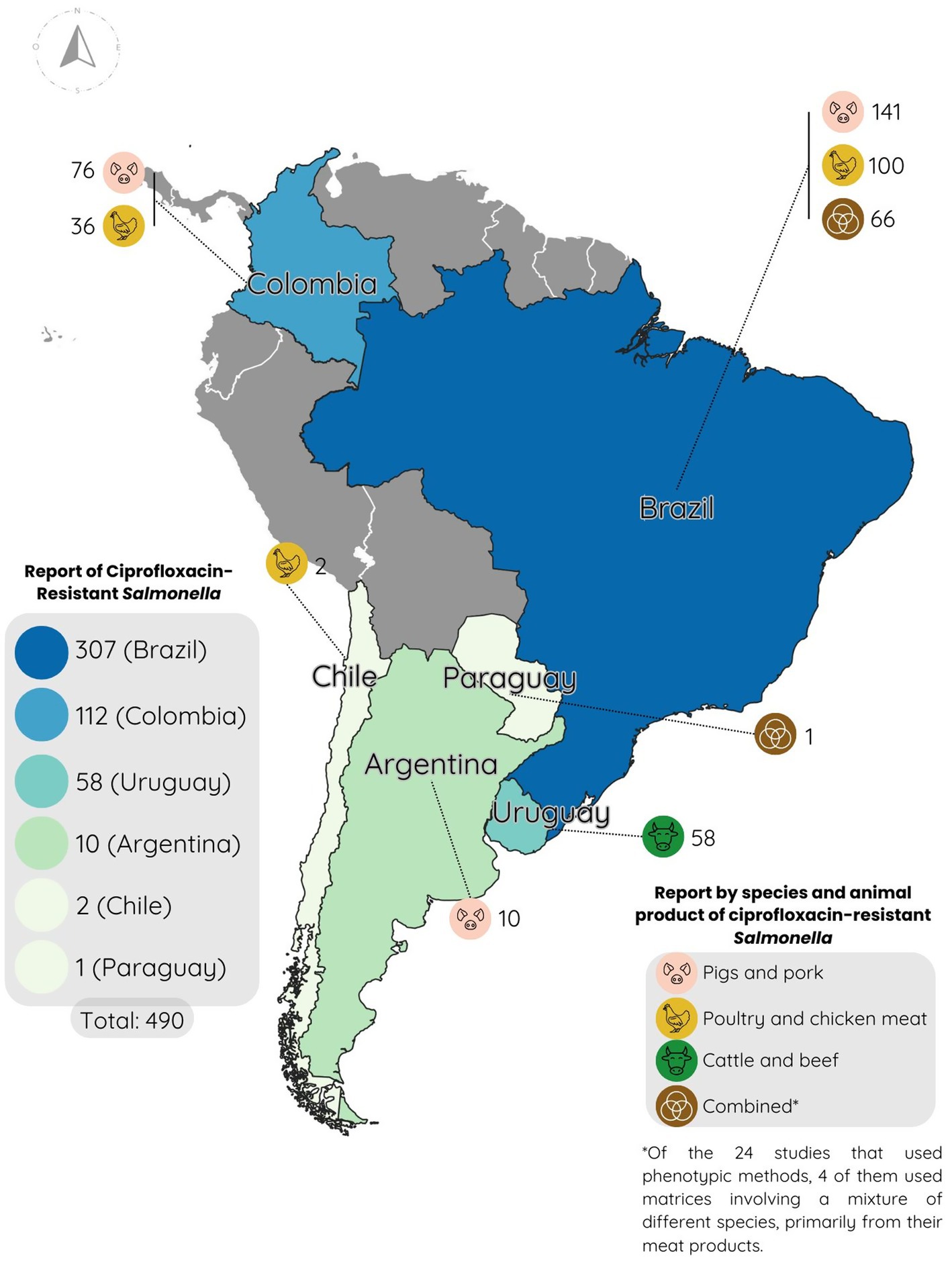
Figure 2. Map of South America with reports of ciprofloxacin resistance percentages reported between 2020 and 2024.
4 Discussion
4.1 Phenotypic resistance and variability between countries and species
Phenotypic resistance to quinolones and fluoroquinolones in Salmonella from poultry and swine in South America reveals an alarming trend of antimicrobial resistance that varies between countries and production systems. This is particularly concerning, as fluoroquinolones are classified as “highest priority” by the World Health Organization (WHO) (12), they play a crucial role in the treatment of serious bacterial infections in humans and animals. Their prioritization is due to the fact that, in many cases, they are the only or few therapies available to treat severe non-human-origin infections (24).
Phenotypic resistance in Salmonella from poultry fecal samples is widely documented. In Brazil, resistance to nalidixic acid was reported at 90% in cloacal swabs and 28% in other samples, while ciprofloxacin resistance ranged from 0 to 16% (25, 26). In Colombia, resistance to ciprofloxacin (92.3%), levofloxacin (57%), and enrofloxacin (48.7%) was observed in poultry feces (24). Serotypes such as S. Paratyphi B, S. Minnesota, and S. Heidelberg have been identified, with the latter two showing multidrug resistance (MDR) rates of 64–66% (24–26).
In Chile, Salmonella isolates from chicken meat show 97% resistance to nalidixic acid, with 94% classified as MDR (27). Similar trends are observed in Brazil, where resistance to nalidixic acid ranges from 94.9 to 100%, likely due to the selective pressure from extensive antibiotic use in poultry production (28, 29). In Brazil, chicken carcass isolates also exhibit high FQ resistence, with ciprofloxacin resistance reaching 81.82% and MDR rates at 63.64% (30). However, other studies report significantly lower ciprofloxacin resistance (0.93–2.08%), suggesting possible methodological differences or evolving resistance patterns over time (31, 32).
In Colombia, older studies reported low resistance in poultry farms to levofloxacin, 2.3% in Cundinamarca, 0% in Santander (33), but more recent data show a sharp increase in resistance, likely due to continued antibiotic use (24). Genetic studies highlight Salmonella Heidelberg (ST15) as a major concern due to its high morbidity, resistance, and outbreak potential, posing a significant public health risk (34–37).
4.2 Genotypic resistance and key mutations
Genotypic studies identified key mutations in gyrA and parC, along with the qnrB19 gene, as major contributors to fluoroquinolone resistance (34, 35). Many ciprofloxacin-resistant strains carried both gyrA mutations and the qnrB gene, enhancing resistance (38). A study on S. Heidelberg from Brazilian poultry meat imported to the Netherlands found parC mutations in all 122 isolates, with 96.7% also carrying gyrA mutations (39). Similarly, qnrB19 was detected in S. Minnesota from Brazilian poultry meat (35, 37). The aac(6′)-Ib-cr gene, linked to fluoroquinolone resistance, was found in one S. Heidelberg isolate from Colombian broiler farms, which showed resistance to ciprofloxacin and levofloxacin (24, 40).
Antimicrobial resistance in Salmonella from swine varies across countries, influenced by production practices, antibiotic use, and regulations. In Brazil, de Quadros et al. (63) found that only 16% of Salmonella strains were fully susceptible, yet enrofloxacin inhibited 100% of them. This is notable, as enrofloxacin was widely used before 2017. However, stricter fluoroquinolone regulations in pork production appear to have reduced resistance, indicating a positive impact of recent policies (41).
However, Kich et al. (64) reported high nalidixic acid resistance (45.9%) in carcasses, while studies on mesenteric lymph nodes found resistance rates of 40.6–65.5% and MDR rates of 70.3–79% (42, 43). In Argentina, Vico et al. (66) observed 52% resistance to fluoroquinolones in mesenteric lymph nodes and 86% MDR. Resistance in S. Typhimurium and S. Derby, a key serotype in swine and pork products worldwide, is particularly concerning (44–46). The role of S. Derby, which is one of the most frequently reported serotypes in swine and pork products, both in Brazil and other regions such as the European Union and China (46, 47). In Colombia, Vidal et al. (67) reported 44% MDR in Salmonella from swine feces, with 55% ciprofloxacin resistance, emphasizing the impact of antibiotic overuse in pig farming.
Recent studies reveal distinct resistance patterns between poultry and swine. Pigs show higher resistance to azithromycin, ampicillin, and chloramphenicol, while poultry exhibit greater resistance to quinolones and sulfonamides, reflecting differences in antibiotic use between both industries (48).
Antimicrobial resistance data for cattle is limited. In Uruguay, high non-susceptibility to FQ has been reported, with 77.3% of isolates non-susceptible to ciprofloxacin and 6.6% to enrofloxacin (41). Additionally, 56% of isolates exhibited MDR particularly in serotypes like S. Typhimurium, S. Newport, and S. Anatum are notably prevalent in intensive cattle production, causing both enteric and invasive diseases such as septicemia (49).
Genotypic studies identify ST19 (S. Typhimurium) and its monophasic variant as key serovars in swine, commonly detected in Brazil and the European Union (50–54). S. Typhimurium is the second most common cause of salmonellosis outbreaks in the EU, with pork as the main source in 2014 (55).
The Ser83Phe mutation is frequently reported in pigs strains with reduced susceptibility to CIP, which has been detected in clinical cases in Peru (56). PMQR genes of the qnr alleles, such as the qnrB19 gene, are associated with the spread of Salmonella strains resistant to CIP in the United States (57). The Thr57Ser mutation has been described as relevant for reducing susceptibility to ciprofloxacin (58). The frequent detection of gyrA (position 83) and parC (position 57) mutations highlights their critical role in fluoroquinolone resistance. These findings underscore the need for targeted surveillance and intervention strategies to mitigate the spread of resistant strains.
4.3 One Health implications and need for coordinated surveillance
The antimicrobial resistance observed to quinolones and fluoroquinolones reflects the selective pressure generated by the use of antibiotics in intensive animal production systems. This resistance can vary considerably between different matrices, such as meat, feces and carcass swabs, highlighting the importance of sampling multiple sources within the production system. Although antibiotics remain a crucial tool for treating bacterial diseases, the increase in antimicrobial resistance has reduced their effectiveness (59). Resistance to fluoroquinolones is particularly concerning, as they are commonly used to treat severe human infections, such as systemic salmonellosis in immunocompromised individuals (60).
These findings underscore the urgency of adopting a coordinated One Health approach that integrates surveillance, policy, and intervention strategies across human, animal, and environmental sectors to effectively mitigate the spread of antimicrobial resistance in South America.
4.4 Limitations
This study has several limitations that should be considered when interpreting the results. First, not all South American countries published research on quinolone- and fluoroquinolone-resistant Salmonella between 2020 and 2024, which limits the representativeness of the data for the entire region. In addition, there was considerable variability in the types of samples or matrices used in the included studies—ranging from feces, tissues, food, and in some cases pooled samples from different animal species—making it difficult to isolate Salmonella-specific information.
Although the study initially sought to include data on fluoroquinolone-resistant Salmonella from rodents, no eligible studies were found from South America during the selected period. However, in veterinary medicine, rodents are recognized as important reservoirs and amplifiers of zoonotic infections, including Salmonella. Their absence in the published literature highlights a relevant knowledge gap that should be addressed in future research.
Finally, although a predefined protocol was used to guide the review process, it was not formally registered in a database such as PROSPERO. This omission is acknowledged as a limitation in the transparency of the study methodology.
Future studies should aim to include a broader range of countries and matrices, and explore the role of rodents in the dissemination of antimicrobial-resistant Salmonella.
5 Conclusion
The antimicrobial resistance to quinolones and fluoroquinolones in Salmonella from poultry, swine, and cattle in South America shows a concerning trend, with high rates of resistance and multidrug resistance across several countries. The indiscriminate use of antibiotics in animal production systems appears to be a key factor in the accelerating pressure of the problem, which could undermine the effectiveness of treatments in both humans and animals. Furthermore, resistance varies significantly across different matrices, highlighting the need for more comprehensive surveillance and the use of more standardized diagnostic techniques. The findings of this study highlight the urgent need for coordinated regional efforts to monitor and control antimicrobial resistance in Salmonella. Policymakers, researchers, and industry stakeholders must collaborate to implement effective strategies that safeguard public health and ensure food safety.
Author contributions
SB-V: Investigation, Data curation, Writing – original draft, Formal analysis. MG-Á: Writing – review & editing, Investigation, Data curation. JV: Writing – review & editing. LG-O: Writing – review & editing. SL-O: Writing – review & editing. JC-G: Wring – review & editing, Funding acquisition.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This project was funded by Group CIBAV, Strategy of sustainability 2023–2025, Faculty of Agrarian Sciences at the University of Antioquia, Medellin, Colombia.
Acknowledgments
We would like to thank the reviewers and collaborators who provided valuable feedback and support throughout this study.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The authors declare that Gen AI was used in the creation of this manuscript. The authors acknowledge the use of ChatGPT, an artificial intelligence (AI) model developed by OpenAI, as a tool to support translation and enhance the clarity and coherence of the manuscript’s writing. However, the authors declare that this AI model was not used for the creation of the manuscript’s content, the formulation of ideas, or the generation, analysis, or interpretation of the results presented.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
1. Oludairo, OO, Kwaga, JKP, Kabir, J, Abdu, PA, Gitanjali, A, Perrets, A, et al. A review on salmonella characteristics, taxonomy, nomenclature with special reference to non-Typhoidal and Typhoidal salmonellosis. Zagazig Vet J. (2022) 50:160–71. doi: 10.21608/zvjz.2022.137946.1179
2. Moura, EAGO, DGD, S, Turco, CH, Sanches, TVC, Storino, GY, Almeida, HMS, et al. Salmonella bacterin vaccination decreases shedding and colonization of Salmonella Typhimurium in pigs. Microorganisms. (2021) 9:1163. doi: 10.3390/microorganisms9061163
3. Chen, HM, Wang, Y, Su, LH, and Chiu, CH. Nontyphoid salmonella infection: microbiology, clinical features, and antimicrobial therapy. Pediatr Neonatol. (2013) 54:147–52. doi: 10.1016/j.pedneo.2013.01.010
4. European Centre for Disease Prevention and Control (ECDC). The European Union one health 2022 zoonoses report. EFSA J. (2023) 21:e8442. doi: 10.2903/j.efsa.2023.8442
5. Márquez, RJA. Sanidad animal y salud pública: el paradigma de la salmonella. New York: Amazing Books (2023).
6. Dudhane, RA, Bankar, NJ, Shelke, YP, and Badge, AK. The rise of non-typhoidal salmonella infections in India: causes, symptoms, and prevention. Cureus. (2023) 15:e46699. doi: 10.7759/cureus.46699
7. Ayala-Romero, C, Ballen-Parada, C, Rico-Gaitán, M, Chamorro-Tobar, I, Zambrano-Moreno, D, Poutou-Piñales, R, et al. Prevalencia de Salmonella spp., en ganglios mesentéricos de porcinos en plantas de beneficio Colombianas. Rev MVZ Córdoba. (2018) 23:6474–86. doi: 10.21897/rmvz.1242
8. Gouvêa, R, Santos, F, Aquino, M, and Pereira, VL. Fluoroquinolones in industrial poultry production, bacterial resistance and food residues: a review. Braz. J Poult Sci. (2015) 17:1–10. doi: 10.1590/1516-635x17011-10
9. Eng, SK, Pusparajah, P, Ab Mutalib, NS, Ser, HL, Chan, KG, and Lee, LH. Salmonella: A review on pathogenesis, epidemiology and antibiotic resistance. Front Life Sci. (2015) 8:284–93. doi: 10.1080/21553769.2015.1051243
10. Mulvey, MR, Boyd, DA, Olson, AB, Doublet, B, and Cloeckaert, A. The genetics of salmonella genomic island 1. Microbes Infect. (2006) 8:1915–22. doi: 10.1016/j.micinf.2005.12.028
11. Martinez, M, McDermott, P, and Walker, R. Pharmacology of the fluoroquinolones: a perspective for the use in domestic animals. Vet J. (2006) 172:10–28. doi: 10.1016/j.tvjl.2005.07.010
12. World Health Organization. Critically important antimicrobials for human medicine. Geneva: World Health Organization (2019).
13. Gu, Y, Huang, L, Wu, C, Huang, J, Hao, H, Yuan, Z, et al. The evolution of fluoroquinolone resistance in salmonella under exposure to sub-inhibitory concentration of enrofloxacin. Int J Mol Sci. (2021) 22:12218. doi: 10.3390/ijms222212218
14. Cuypers, WL, Jacobs, J, Wong, V, Klemm, EJ, Deborggraeve, S, and Van Puyvelde, S. Fluoroquinolone resistance in salmonella: insights by whole-genome sequencing. Microb Genom. (2018) 4:e000195. doi: 10.1099/mgen.0.000195
15. Correia, S, Poeta, P, Hébraud, M, Capelo, JL, and Igrejas, G. Mechanisms of quinolone action and resistance: where do we stand? J Med Microbiol. (2017) 66:551–9. doi: 10.1099/jmm.0.000475
16. Page, MJ, Moher, D, Bossuyt, PM, Boutron, I, Hoffmann, TC, Mulrow, CD, et al. PRISMA 2020 explanation and elaboration: updated guidance and exemplars for reporting systematic reviews. BMJ. (2021) 372:n160. doi: 10.1136/bmj.n160
17. Biemer, JJ. Antimicrobial susceptibility testing by the Kirby-Bauer disc diffusion method. Ann Clin Lab Sci. (1973) 3:135–40.
18. Wiegand, I, Hilpert, K, and Hancock, REW. Agar and broth dilution methods to determine the minimal inhibitory concentration (MIC) of antimicrobial substances. Nat Protoc. (2008) 3:163–75. doi: 10.1038/nprot.2007.521
19. Mullis, KB, and Faloona, FA. Specific synthesis of DNA in vitro via a polymerase-catalyzed chain reaction. Methods Enzymol. (1987) 155:335–50. doi: 10.1016/0076-6879(87)55023-6
20. Huggett, J, Dheda, K, Bustin, S, and Zumla, A. Real-time RT-PCR normalisation; strategies and considerations. Genes Immun. (2005) 6:279–84. doi: 10.1038/sj.gene.6364190
21. Robicsek, A, Strahilevitz, J, Sahm, DF, Jacoby, GA, and Hooper, DC. qnr prevalence in Ceftazidime-resistant Enterobacteriaceae isolates from the United States. Antimicrob Agents Chemother. (2006) 50:2872–4. doi: 10.1128/AAC.01647-05
22. Quainoo, S, Coolen, JPM, van Hijum, SAFT, Huynen, MA, Melchers, WJG, van Schaik, W, et al. Whole-genome sequencing of bacterial pathogens: the future of nosocomial outbreak analysis. Clin Microbiol Rev. (2017) 30:1015–63. doi: 10.1128/CMR.00016-17
23. Köser, CU, Bryant, JM, Becq, J, Török, ME, Ellington, MJ, Marti-Renom, MA, et al. Whole-genome sequencing for rapid susceptibility testing of M. tuberculosis. N Engl J Med. (2013) 369:290–2. doi: 10.1056/NEJMc1215305
24. Herrera-Sánchez, MP, Castro-Vargas, RE, Fandiño-de-Rubio, LC, Rodríguez-Hernández, R, and Rondón-Barragán, IS. Molecular identification of fluoroquinolone resistance in salmonella spp. isolated from broiler farms and human samples obtained from two regions in Colombia. Vet World. (2021) 14:1767–73. doi: 10.14202/vetworld.2021.1767-1773
25. Moreira, JPFF, do Monte, DFM, Lima, C d A, de Oliveira, CJB, da Silva Martins, NR, Berchieri Junior, A, et al. Molecular genotyping reveals inter-regional relatedness among antimicrobial resistant salmonella Minnesota strains isolated from poultry farm and humans, Brazil. Braz J Microbiol. (2022) 53:503–8. doi: 10.1007/s42770-021-00666-1
26. Souza, AIS, Saraiva, MMS, Casas, MRT, Oliveira, GM, Cardozo, MV, Benevides, VP, et al. High occurrence of β-lactamase-producing salmonella Heidelberg from poultry origin. PLoS One. (2020) 15:e0230676. doi: 10.1371/journal.pone.0230676
27. Lapierre, L, Cornejo, J, Zavala, S, Galarce, N, Sánchez, F, Benavides, MB, et al. Phenotypic and genotypic characterization of virulence factors and susceptibility to antibiotics in salmonella Infantis strains isolated from chicken meat: first findings in Chile. Animals. (2020) 10:1049. doi: 10.3390/ani10061049
28. Perin, AP, Martins, BTF, Barreiros, MAB, Yamatogi, RS, Nero, LA, and dos Santos Bersot, L. Occurrence, quantification, pulse types, and antimicrobial susceptibility of salmonella sp. isolated from chicken meat in the state of Paraná, Brazil. Braz J Microbiol. (2020) 51:335–45. doi: 10.1007/s42770-019-00188-x
29. Vilela, FP, dos Prazeres Rodrigues, D, Allard, MW, and Falcão, JP. Genomic analyses of drug-resistant Salmonella enterica serovar Heidelberg strains isolated from meat and related sources between 2013 and 2017 in the south region of Brazil. Curr Genet. (2023) 69:141–52. doi: 10.1007/s00294-023-01264-4
30. Lucca, V, Borges, KA, Furian, TQ, Chitolina, GZ, Streck, AF, da Rocha, DT, et al. Phenotypic and molecular characterisation of salmonella spp. isolates in healthy poultry. Br Poult Sci. (2024) 65:415–23. doi: 10.1080/00071668.2024.2337180
31. Grossi, JL, Yamatogi, RS, Call, DR, and Nero, LA. High prevalence of intermediate resistance to ciprofloxacin in Salmonella enterica isolated from a Brazilian poultry production chain, located in Minas Gerais state. Int J Food Microbiol. (2023) 394:110180. doi: 10.1016/j.ijfoodmicro.2023.110180
32. Monte, DFM, Andrigheto, C, Ribeiro, VB, Landgraf, M, and Destro, MT. Highly clonal relationship among Salmonella Enteritidis isolates in a commercial chicken production chain, Brazil. Braz J Microbiol. (2020) 51:2049–56. doi: 10.1007/s42770-020-00372-4
33. Donado-Godoy, P, Clavijo, V, León, M, Tafur, MA, Gonzales, S, Hume, M, et al. Prevalencia de Salmonella en canales de carne de pollo de engorde al por menor en Colombia. J Food Prot. (2012) 75:1134–8. doi: 10.4315/0362-028X.JFP-11-513
34. de Melo, RT, dos Reis, CT, Peres, PABM, Braz, RF, Monteiro, GP, and Rossi, DA. Salmonella enterica Serovar Minnesota biofilms, susceptibility to biocides, and molecular characterization. Pathogens. (2021) 10:581. doi: 10.3390/pathogens10050581
35. Monte, DF, Lincopan, N, Berman, H, Cerdeira, L, Keelara, S, Thakur, S, et al. Genomic features of high-priority Salmonella enterica Serovars circulating in the food production chain, Brazil, 2000–2016. Sci Rep. (2019) 9:11058. doi: 10.1038/s41598-019-45838-0
36. Koutsoumanis, K, Allende, A, Alvarez-Ordóñez, A, Bolton, D, Bover-Cid, S, Chemaly, M, et al. Salmonella control in poultry flocks and its public health impact. EFSA J. (2019) 17:e05596. doi: 10.2903/j.efsa.2019.5596
37. Saidenberg, ABS, Franco, LS, Reple, JN, Hounmanou, YMG, Casas, MRT, Cardoso, B, et al. Salmonella Heidelberg and salmonella Minnesota in Brazilian broilers: genomic characterization of third-generation cephalosporin and fluoroquinolone-resistant strains. Environ Microbiol Rep. (2023) 15:119–28. doi: 10.1111/1758-2229.13132
38. Parada, J, Galas, M, Faccone, D, Tamiozzo, P, and Carranza, A. Antibiotic resistance and associated resistance determinants in different Salmonella enterica serovars isolated from pigs in Argentina. Vet World. (2022) 15:1215–20. doi: 10.14202/vetworld.2022.1215-1220
39. van den Berg, RR, Dissel, S, Rapallini, MLBA, van der Weijden, CC, Wit, B, and Heymans, R. Characterization and whole genome sequencing of closely related multidrug-resistant Salmonella enterica serovar Heidelberg isolates from imported poultry meat in the Netherlands. PLoS One. (2019) 14:e0219795. doi: 10.1371/journal.pone.0219795
40. Vetting, MW, Park, CH, Hegde, SS, Jacoby, GA, Hooper, DC, and Blanchard, JS. Mechanistic and structural analysis of aminoglycoside N-acetyltransferase AAC (6′)-Ib and its bifunctional, fluoroquinolone-active AAC (6′)-Ib-cr variant. Biochemistry. (2008) 47:9825–35. doi: 10.1021/bi800664x
41. Casaux, ML, D’Alessandro, B, Vignoli, R, and Fraga, M. Phenotypic and genotypic survey of antibiotic resistance in Salmonella enterica isolates from dairy farms in Uruguay. Front Vet Sci. (2023) 10:1055432. doi: 10.3389/fvets.2023.1055432
42. de Azevedo, EC, Martins, BTF, Tiba Casas, MR, Possebon, FS, Araújo Junior, JP, Nero, LA, et al. Multidrug resistance and virulence profiles of salmonella isolated from swine lymph nodes. Microb Drug Resist. (2021) 27:562–70. doi: 10.1089/mdr.2020.0120
43. Possebon, FS, Tiba Casas, MR, Nero, LA, Yamatogi, RS, Araújo, JP, and Pinto, AN. Prevalence, antibiotic resistance, PFGE and MLST characterization of salmonella in swine mesenteric lymph nodes. Prev Vet Med. (2020) 179:105024. doi: 10.1016/j.prevetmed.2020.105024
44. Hsueh, PR, Teng, LJ, Tseng, SP, Chang, CF, Wan, JH, Yan, JJ, et al. Ciprofloxacin-resistant Salmonella enterica Typhimurium and Choleraesuis from pigs to humans, Taiwan. Emerg Infect Dis. (2004) 10:60–8. doi: 10.3201/eid1001.030171
45. Pissetti, C, de Freitas Costa, E, Zenato, KS, and de Itapema Cardoso, MR. Critically important antimicrobial resistance trends in salmonella Derby and Salmonella Typhimurium isolated from the pork production chain in Brazil: a 16-year period. Pathogens. (2022) 11:905. doi: 10.3390/pathogens11080905
46. Simoni, C, de Campos Ausani, T, Laviniki, V, Lopes, GV, and de Itapema Cardoso, MR. Salmonella derby from pig production chain over a 10-year period: antimicrobial resistance, biofilm formation, and genetic relatedness. Braz J Microbiol. (2022) 53:2185–94. doi: 10.1007/s42770-022-00846-7
47. Cai, Y, Tao, J, Jiao, Y, Fei, X, Zhou, L, Wang, Y, et al. Phenotypic characteristics and genotypic correlation between salmonella isolates from a slaughterhouse and retail markets in Yangzhou, China. Int J Food Microbiol. (2016) 222:56–64. doi: 10.1016/j.ijfoodmicro.2016.01.020
48. Gomes, VTM, Moreno, LZ, Silva, APS, Thakur, S, La Ragione, RM, Mather, AE, et al. Characterization of Salmonella enterica contamination in pork and poultry meat from São Paulo/Brazil: serotypes, genotypes and antimicrobial resistance profiles. Pathogens. (2022) 11:358. doi: 10.3390/pathogens11030358
49. Costa, LF, Paixão, TA, Tsolis, RM, Bäumler, AJ, and Santos, RL. Salmonellosis in cattle: advantages of being an experimental model. Res Vet Sci. (2012) 93:1–6. doi: 10.1016/j.rvsc.2012.03.002
50. Biasino, W, De Zutter, L, Mattheus, W, Bertrand, S, Uyttendaele, M, and Van Damme, I. Correlation between slaughter practices and the distribution of salmonella and hygiene indicator bacteria on pig carcasses during slaughter. Food Microbiol. (2018) 70:192–9. doi: 10.1016/j.fm.2017.10.003
51. Bonardi, S. Salmonella in the pork production chain and its impact on human health in the European Union. Epidemiol Infect. (2017) 145:1513–26. doi: 10.1017/S095026881700036X
52. Cabral, CC, Panzenhagen, PHN, Delgado, KF, Silva, GRA, Rodrigues, DDP, Franco, RM, et al. Contamination of carcasses and utensils in small swine slaughterhouses by salmonella in the northwestern region of the state of Rio de Janeiro, Brazil. J Food Prot. (2017) 80:1128–32. doi: 10.4315/0362-028X.JFP-16-387
53. dos Santos Bersot, L, Quintana Cavicchioli, V, Viana, C, Konrad Burin, RC, Camargo, AC, de Almeida Nogueira Pinto, JP, et al. Prevalence, antimicrobial resistance, and diversity of salmonella along the pig production chain in southern Brazil. Pathogens. (2019) 8:204. doi: 10.3390/pathogens8040204
54. Viana, C, Sereno, MJ, Pegoraro, K, Yamatogi, RS, Call, DR, dos Santos Bersot, L, et al. Distribution, diversity, virulence genotypes and antibiotic resistance for salmonella isolated from a Brazilian pork production chain. Int J Food Microbiol. (2019) 310:108310. doi: 10.1016/j.ijfoodmicro.2019.108310
55. EFSA. The European Union summary report on trends and sources of zoonoses, zoonotic agents and food-borne outbreaks in 2016. EFSA J. (2017) 15:e05077.
56. García, C, Lejon, V, Horna, G, Astocondor, L, Vanhoof, R, Bertrand, S, et al. Intermediate Susceptibility to Ciprofloxacin among Salmonella enterica Serovar Typhi Isolates in Lima, Peru. J Clin Microbiol. (2014) 52:968–70. doi: 10.1128/JCM.02663-13
57. Elnekave, E, Hong, SL, Lim, S, Hayer, SS, Boxrud, D, Taylor, AJ, et al. Circulation of plasmids harboring resistance genes to quinolones and/or extended-spectrum cephalosporins in multiple Salmonella enterica serotypes from swine in the United States. Antimicrob Agents Chemother. (2019) 63:e02602. doi: 10.1128/AAC.02602-18
58. Ling, JM, Chan, EW, Lam, AW, and Cheng, AF. Mutations in topoisomerase genes of fluoroquinolone-resistant salmonellae in Hong Kong. Antimicrob Agents Chemother. (2003) 47:3567–73. doi: 10.1128/AAC.47.11.3567-3573.2003
60. Medalla, F, Gu, W, Mahon, BE, Judd, M, Folster, J, Griffin, PM, et al. Estimated incidence of antimicrobial drug–resistant nontyphoidal salmonella infections, United States, 2004–2012. Emerg Infect Dis. (2017) 23:29–37. doi: 10.3201/eid2301.160771
61. Benevides, VP, Saraiva, MMS, Nascimento, CF, Delgado-Suárez, EJ, Oliveira, CJB, Silva, SR, et al. Genomic features and phylogenetic analysis of antimicrobial-resistant salmonella Mbandaka ST413 strains. Microorganisms. (2024) 12:312. doi: 10.3390/microorganisms12020312
62. Burnett, E., Ishida, M., de Janon, S., Naushad, S., Duceppe, M.-O., Gao, R., et al. (2021). Whole-Genome Sequencing Reveals the Presence of the blaCTX-M-65 Gene in Extended-Spectrum β-Lactamase-Producing and Multi-Drug-Resistant Clones of Salmonella Serovar Infantis Isolated from Broiler Chicken Environments in the Galapagos Islands. Antibiotics, 10:3. doi: 10.3390/antibiotics10030267
63. de Quadros, C. L., Manto, L., Mistura, E., Webber, B., Ritterbusch, G. A., Borges, K. A., et al. (2020). Antimicrobial and Disinfectant Susceptibility of Salmonella Serotypes Isolated from Swine Slaughterhouses. Current Microbiology, 77, 1035–1042. doi: 10.1007/s00284-020-01904-9
64. Kich, J. D., Souza, A. I. A., Montes, J., Meneguzzi, M., Costa, E. F., Coldebella, A., et al. (2020). Investigation of Listeria monocytogenes, Salmonella enterica and Yersinia enterocolitica in pig carcasses in Southern Brazil. Pesquisa Veterinária Brasileira, 40, 781–790. doi: 10.1590/1678-5150-PVB-6628
65. Ortiz, F., Weiler, N., Álvarez, M., Orrego, V., Martínez, J., Melgarejo, N., et al. (2023). Mecanismos plasmídicos de resistencia a quinolonas, betalactámicos y colistina en Salmonella enterica. Paraguay 2020-2021. Memorias del Instituto de Investigaciones en Ciencias de la Salud, 21, 13. doi: 10.18004/mem.iics/1812-9528/2023.e21122313
66. Vico, J. P., Lorenzutti, A., Zogbi, A. P., Aleu, G., Sánchez, I. C., Caffer, M. I., et al. (2020). Prevalence, associated risk factors, and antimicrobial resistance profiles of non-typhoidal Salmonella in large scale swine production in Córdoba, Argentina. Research in Veterinary Science, 130. doi: 10.1016/j.rvsc.2020.03.003
67. Vidal, J. L., Clavijo, V., Castellanos, L. R., Kathiresan, J., Kumar, A. M. V., Mehta, K., et al. (2023). Multidrug-resistant Salmonella spp. In fecal samples of pigs with suspected salmonellosis in Antioquia, Colombia, 2019–2021. Revista Panamericana de Salud Pública, 47, 1. doi: 10.26633/RPSP.2023.46
68. Vilela, F. P., Rodrigues, D. d., Ferreira, J. C., Darini, A. L. d. C., Allard, M. W., Falcão, J. P., et al. (2020). Genomic characterization of Salmonella enterica serovar Choleraesuis from Brazil reveals a swine gallbladder isolate harboring colistin resistance gene mcr-1.1. Braz. J. Microbiol, 53, 1799–1806. doi: 10.1007/s42770-022-00812-3
Keywords: quinolones, fluoroquinolones, antimicrobial resistance, Salmonella, South America
Citation: Barrientos-Villegas S, García-Álvarez MI, Vidal JL, Gómez-Osorio LM, López-Osorio S and Chaparro-Gutiérrez JJ (2025) Systematic review: genotypic and phenotypic resistance of fluoroquinolone-resistant Salmonella in livestock in South America (2020–2024). Front. Vet. Sci. 12:1614486. doi: 10.3389/fvets.2025.1614486
Edited by:
Windell L. Rivera, University of the Philippines Diliman, PhilippinesReviewed by:
Chelea Matchawe, Institute of Medical Research and Studies of Medicinal Plants (IMPM), CameroonElizabeth M. Parker, The Ohio State University, United States
Copyright © 2025 Barrientos-Villegas, García-Álvarez, Vidal, Gómez-Osorio, López-Osorio and Chaparro-Gutiérrez. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Jenny J. Chaparro-Gutiérrez, amVubnkuY2hhcGFycm9AdWRlYS5lZHUuY28=