 Gabriel Miranda Macambira1
Gabriel Miranda Macambira1 Carlos Bôa-Viagem Rabello1
Carlos Bôa-Viagem Rabello1 Hélia Sharlane de Holanda Oliveira1†
Hélia Sharlane de Holanda Oliveira1† Marcos José Batista dos Santos1
Marcos José Batista dos Santos1 Apolônio Gomes Ribeiro2Oziel Saturnino Lins Júnior1†Lucas Delano Nascimento de Sousa1Igor Luiz Carvalho Máximo1†
Apolônio Gomes Ribeiro2Oziel Saturnino Lins Júnior1†Lucas Delano Nascimento de Sousa1Igor Luiz Carvalho Máximo1† Lucas Rannier Ribeiro Antonino Carvalho3*†
Lucas Rannier Ribeiro Antonino Carvalho3*† Odrey Mesa Fleitas4
Odrey Mesa Fleitas4- 1Department of Animal Science, Universidade Federal Rural de Pernambuco, Recife, Brazil
- 2Department of Animal Science, Universidade Federal da Paraíba, Areia, Brazil
- 3Department of Physiology and Pharmacology, Stockholm Sweden Biomedicum, Karolinska Institutet, Stockholm, Sweden
- 4Institute of Animal Science (ICA), Havana, Cuba
Moringa oleifera leaf meal (MOL) can be an alternative food in poultry diets. Still, their use is limited due to the presence of non-starch polysaccharides (NSP) and phytates. The supply of leaves associated with exogenous enzymes can influence the weight, size, and morphology of the intestine, accessory glands, and reproductive system of laying hens. The aim was, therefore, to study the influence of MOL in diets supplemented with and without the enzymes xylanase and phytase, whether associated or not, and their effects on the size and weight of the organs of the gastrointestinal tract, accessory glands, and reproductive system, as well as on the morphological characteristics of the small intestine of laying hens intended for egg production during the peak laying period. A total of 288 laying hens of the Dekalb White strain aged 32 weeks were used, distributed in a completely randomized design in a 2 × 4 factorial arrangement (presence and absence of MOL x 4 forms of enzyme supplementation - with or without) for a total of eight treatments with six replicates of six birds per experimental unit. The treatments consisted of a control diet based on corn and soybean meal and a diet with 5% MOL supplementation followed by three forms of enzyme supplementation (xylanase, phytase, and a mix of the two enzymes). Xylanase, alone or in combination with phytase, played an essential role in reducing the relative size of the small intestine and caeca, as well as improving intestinal morphology by increasing villus height, crypt depth, villus height/crypt depth ratio, mucosal length, and villus width, thus characterizing improvements in the processes of digestion and absorption of nutrients. Phytase has not influenced the variables studied in this study. Dietary inclusion of Moringa oleifera leaf (MOL) and exogenous enzymes, particularly xylanase, significantly influenced organ weights and intestinal morphology in laying hens. Xylanase, especially when combined with MOL, enhanced villus height, crypt depth, V:C ratio, and mucosal length across all intestinal segments, while phytase had more variable effects depending on diet context. It can be concluded that the NSP present in MOL have various effects on the intestine, such as increasing the size of the intestine and attached glands and influencing intestinal morphometric characteristics. With the degradation of these nutrients, the action of xylanase re-establishes the birds’ intestinal health, which could lead to better performance from the layers.
1 Introduction
Plants are the most widely used food sources in the formulation of poultry diets, with corn and soybean meal being these animals’ primary feedstuffs. Plant-based ingredients contain varying levels of non-starch polysaccharides (NSPs), which are structural components of the plant cell wall and primarily serve a protective function (1). These fiber compounds are composed of glycosidic bonds (β-1,4 and β-1,6) that cannot be broken down by the animal’s endogenous enzymes. However, they can be degraded by microbial enzymes produced by the gut microbiota (2, 3).
The effect of these compounds on the gastrointestinal tract (GIT) of poultry depends on the solubility of each molecule. Soluble NSP has a high-water retention capacity. They can increase the viscosity and volume of the digesta, decrease intestinal transit, as well as compromise the association of enzymes and substrates, impairing the digestion of proteins, fats, and carbohydrates (4–8). Insoluble NSP, on the other hand, speed up intestinal transit, reducing the time the digested remains in contact with endogenous digestive enzymes, as well as encapsulating nutrients within the plant cell, making them unavailable for use by poultry (2, 6, 9–11).
Herbaceous plants have received considerable attention for improving the performance and health status of commercial layers (12). In this context, Moringa oleifera leaves (MOL) show great potential for use in poultry nutrition due to their rich nutritional profile, including an average protein content of around 25%, as well as high levels of calcium, phosphorus, flavonoids, ascorbic acid, alpha-tocopherol, polyphenols, glycosides, and phenolic compounds. MOL, commonly referred to as the “miracle tree,” is recognized for its exceptional nutritional composition. In poultry nutrition, it has been explored as a natural feed additive due to its multiple beneficial effects. When included at optimal levels in broiler diets, MOL has been shown to improve body weight gain and feed conversion ratio. Additionally, its bioactive compounds help control pathogenic gut bacteria, contributing to improved intestinal health and a reduced need for antibiotic use. The inclusion of MOL also enhances nutrient digestibility. In laying hens, it has been associated with improvements in egg production and egg quality parameters, such as yolk pigmentation and shell thickness, likely due to its content of carotenoids and essential minerals (13–21).
However, thanks to the high NSP content present in this food, its use in poultry nutrition is still limited. Macambira et al. (22) found that most of the fibrous compounds in Moringa oleifera leaves belong to the soluble fraction. Formed by the hemicellulose fractions (xyloglucans, xylans, arabinoxylans, β glucans, among others), gums and pectins, these components have a high capacity for absorbing water, resulting in an increase in the viscosity and volume of the digestate, compromising the association of enzymes and substrates, reduced intestinal transit, changes in the secretion of pancreatic juice and other secretory mechanisms of the GIT, an increase in the size of the liver, pancreas, and intestines, thanks to the more significant presence of undigested substrate in the intestinal lumen (4, 6, 8, 23–25). In addition, they can compromise the intestinal morphological characteristics with reduced villus width and size, reduced crypt depth, and villus height/crypt depth ratio (23, 26–29). MOL also contains significant amounts of phytate, which complexes and make unavailable minerals, such as phosphorus, calcium, magnesium, iron, and zinc, and bind to proteins, fibers, and other nutrients.
Exogenous enzymes, mainly carbohydrates, and phytases, represent promising alternatives that allow greater use of fibrous foods in poultry diets (30–34). Although there are studies that have verified the influence of Moringa oleifera leaves on intestinal morphology in broiler chickens (18, 35, 36), no studies on this topic were found with commercial layers. On the other hand, we are unaware of any studies that have examined the effects of the association of carbohydrates in diets containing this vegetable on the morphological characteristics of the intestine in poultry. Therefore, the results of this study represent a new approach to using this promising plant species in poultry feeding for egg production.
The hypothesis was that dietary inclusion of Moringa oleifera leaves (MOL) at a 5% inclusion level, in combination with exogenous enzymes, would positively influence the morphological characteristics of the small intestine, the size of gastrointestinal organs and associated glands, as well as the reproductive system of laying hens during their peak laying period. The aim was, therefore, to study the influence of MOL in diets supplemented with and without the enzymes xylanase and phytase, whether or not associated, and their effects on the size and weight of the GIT organs, reproductive system, and attached glands, as well as on the morphological characteristics of the small intestine of laying hens destined for egg production during the peak laying period.
2 Materials and methods
The research was approved by the Ethics Committee on the Use of Animals (CEUA), of the Federal Rural University of Pernambuco, in accordance with license number 21/2018.
2.1 Production of Moringa oleifera leaf meal and bromatological analysis
Moringa oleifera leaves, and petioles collected 45 days apart were used to obtain the material which combine green matter production and the nutritional value of the leaves. The plants were cut at a height of approximately 60 cm from the ground. After harvesting, the plants were first dried in a shed until their weight stabilized and then chopped into fodder. The material was then ground in a vertical mill to obtain leaf meal.
Samples of MOL and experimental feed were collected and sent to the Animal Nutrition Laboratory (ANL) of the Animal Science Department of the Federal Rural University of Pernambuco (UFRPE) for determination of the dry matter (DM), crude protein (CP), ether extract (EE) and mineral matter (MM) contents, according to the methodologies proposed by Detmann et al. (37). Neutral detergent fiber (NDF) and acid detergent fiber (ADF) were determined using the method proposed by Van Soest (38). Gross energy (BE) was determined using a calorimetric pump (IKA, model C-200). The determined composition of the MOL is shown in Table 1.
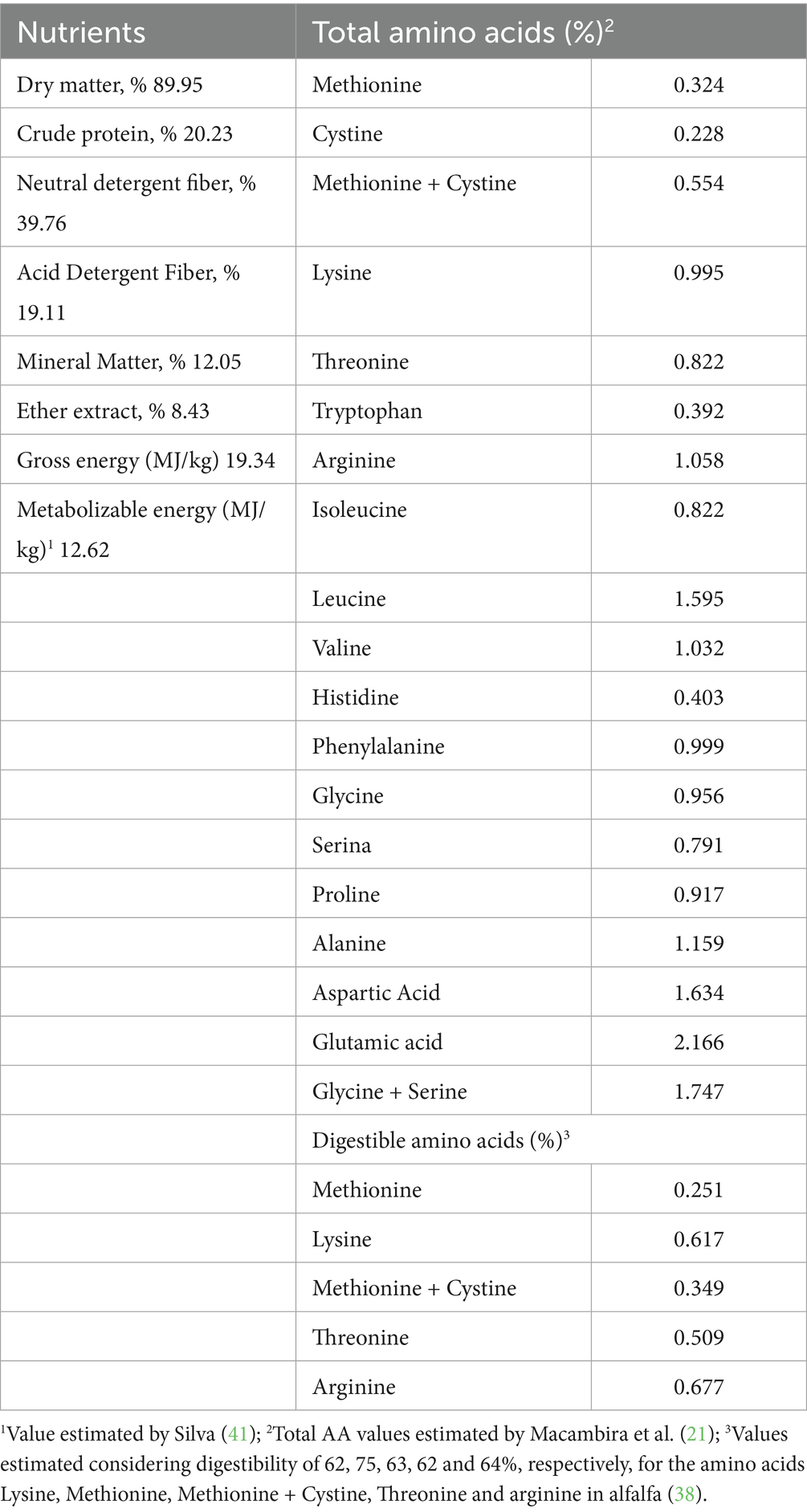
Table 1. Chemical composition of Moringa oleifera leaf meal (in natural matter).
2.2 Birds and facilities
The study used 288 laying hens of the Dekalb White strain aged 32 weeks with an average initial weight of 1,520 kg. They were housed in cages measuring 1.00 × 0.40 × 0.45 m, equipped with a trough for collecting eggs, a trough-type feeder, and an automatic drinker with an attached cup. The birds were weighed at the start of the experimental period to ensure uniformity between the experimental plots. The animals then had their egg production monitored, per experimental unit, for 14 days. Once the weight and egg production were uniform, the treatments were randomly distributed among the experimental units and reared for 18 weeks or 140 days.
The light program was adopted for 17 h, consisting of 12 h of natural light + 5 h of artificial light. The environmental parameters, temperature, and relative humidity were measured daily using a data logger (HOBO, model U12-001), as well as a thermohydrometer (Incoterm Digital, model 7666.02.0.00) installed in the middle of the house at the height of the birds’ backs, throughout the experiment. The average temperature and relative humidity during the experimental period were 25.79°C and 69.92%, respectively. The variations in temperature and relative humidity throughout the period are illustrated in Figure 1.
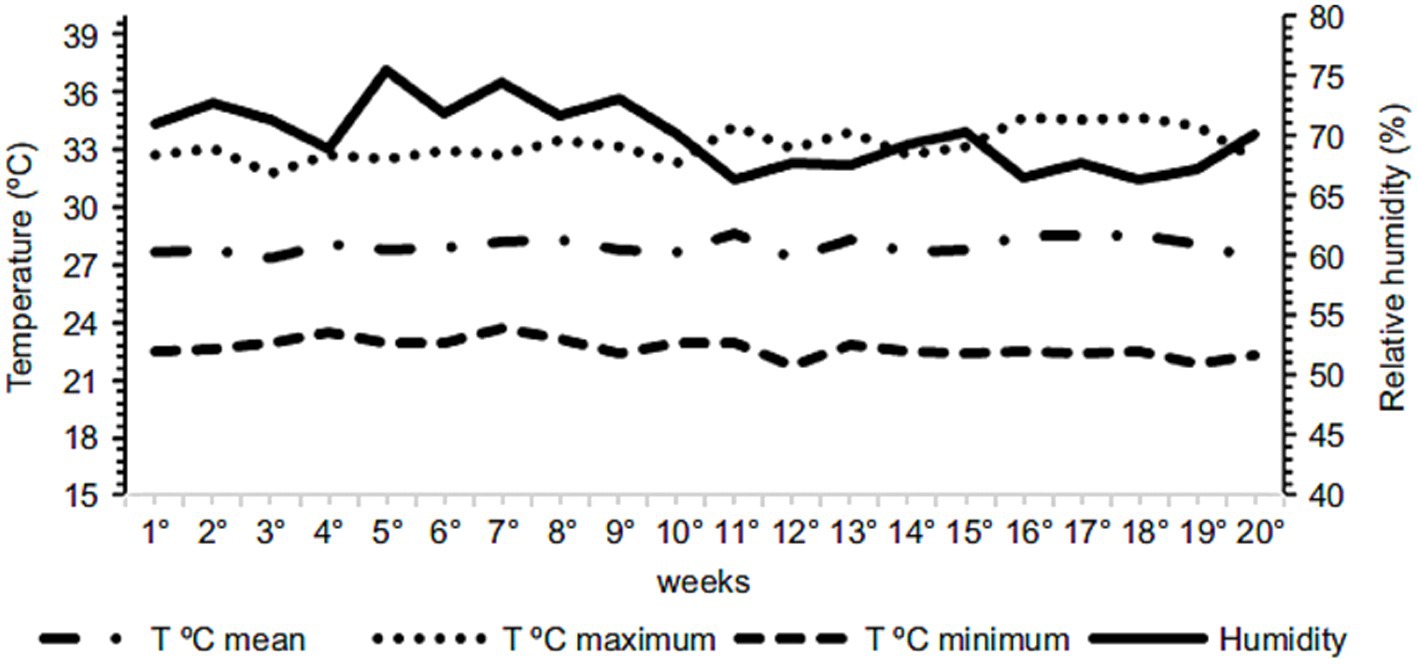
Figure 1. Variation in temperature and relative humidity during the experimental period.
2.3 Design and experimental diets
The birds were distributed in a completely randomized design in a 2 × 4 factorial arrangement (supplementation or not of 5% Moringa oleifera leaf meal x four forms of enzyme supplementation – with or without) for eight treatments. The xylanase used was Econase XT 25P (AB Vista, Florida), a bacterial enzyme expressed in Trichoderma sp. with an activity of 160,000 BXU of endo 1,4-b-xylanase per gram. The supplemented phytase was Quantum-Blue 5 G (AB Vista, Florida), an enzyme isolated from Escherichia coli with an activity of 300FTU. A Beechwood Xylanase Unit (BXU) is the enzyme capable of releasing 1 nmol of birch xylan, measured in xylose equivalents, under assay conditions (AB Enzymes, Germany). The FTU, or active phytase unit, is the amount of enzyme required to release 1 μmol of inorganic phosphorus per minute from a substrate of 0.0051 mol/L of sodium phytate at pH 5.5 and 37°C (39).
The experimental treatments were as follows: a control diet without enzyme supplementation (C); a control diet supplemented with 75 g/tonne of xylanase (CX); a control diet supplemented with 60 g/tonne of phytase (CF); a control diet supplemented with both 75 g/tonne of xylanase and 60 g/tonne of phytase (CMIX); a diet containing 5% Moringa leaf meal without enzyme supplementation (M); a Moringa-based diet supplemented with 75 g/tonne of xylanase (MX); a Moringa-based diet supplemented with 60 g/tonne of phytase (MF); and a Moringa-based diet supplemented with both 75 g/tonne of xylanase and 60 g/tonne of phytase (MMIX). The lowest enzyme dosages recommended by the manufacturers were selected in order to evaluate the minimum effective inclusion level capable of promoting improvements in performance and nutrient utilization. This approach allows for a cost-effective assessment of enzyme efficacy, which is particularly relevant for practical applications in commercial poultry production. Additionally, using the lowest recommended dose helps minimize potential interactions or over-supplementation effects when combined with other dietary components such as Moringa oleifera leaf meal.
Table 2 shows the centesimal composition of the ingredients and the calculated and determined nutritional composition of the experimental rations. The rations were formulated according to the feed composition in the Brazilian Tables for Poultry and Pigs developed by Rostagno et al. (40), except for the MOL, which had its nutritional profile analyzed at the ANL. The apparent metabolizable energy content of 3,014 kcal/kg for MOL was determined by Silva (41) in a metabolism trial with layers. Reference AA data determined by Macambira et al. (22) and estimates of digestible AA considering a digestibility of 62, 75, 63, 62, and 64% for Lysine, Methionine, Methionine + Cystine, Threonine, and Arginine from alfalfa (42) were used to formulate the diets (Table 1). The manual for the breed used met all the birds’ nutritional requirements. The nutritional profile was evaluated for the diets containing the enzymes xylanase and phytase at 100 kcal and 0.15% of available phosphorus, respectively. To better observe their effect on the feed components, all the ingredients were kept stable, with only the amounts of soybean oil and calcium phosphate added varyingpna.
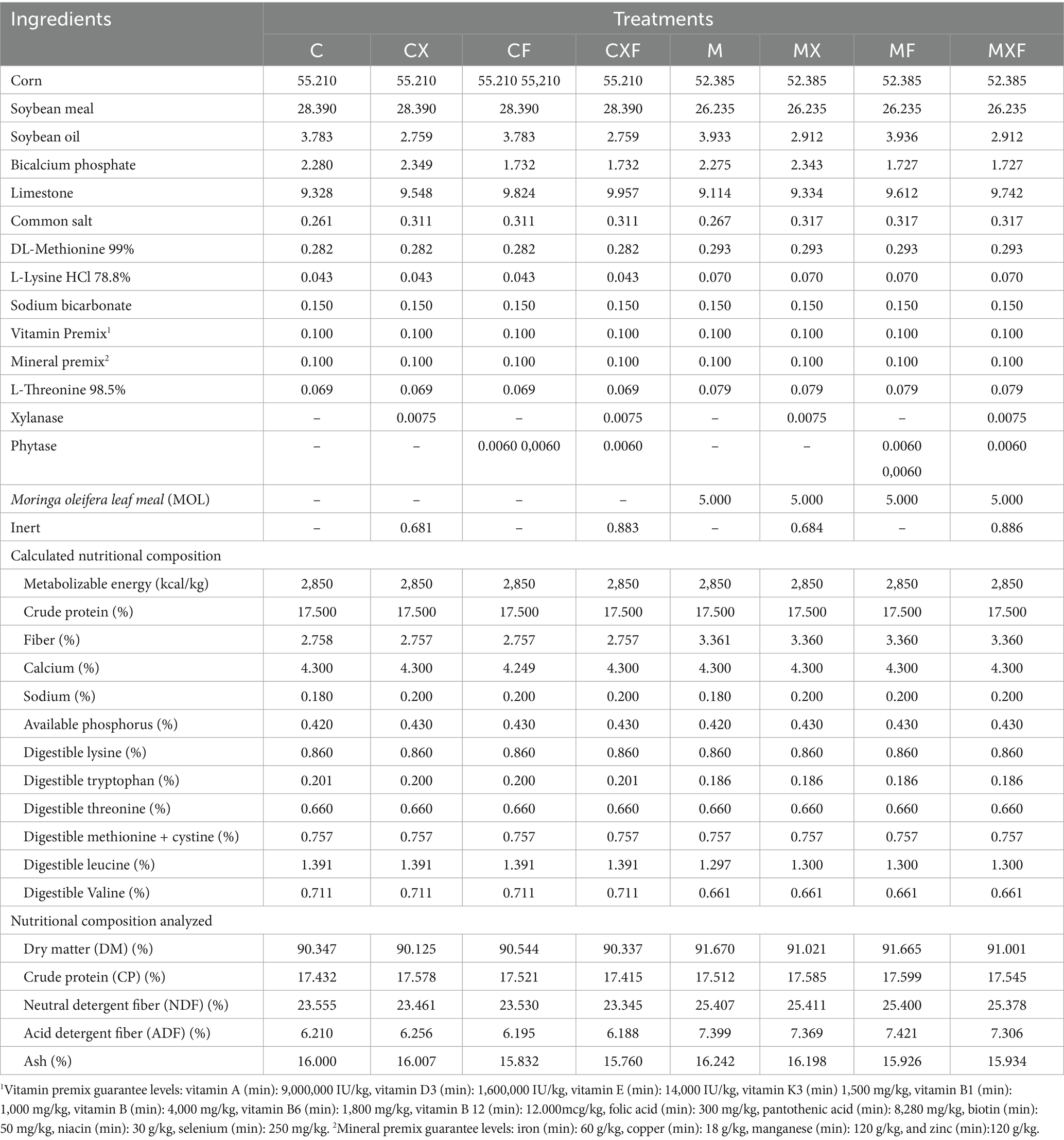
Table 2. Chemical composition and nutritional values of experimental diets.
2.4 Organs of the gastrointestinal tract and productive system
At the end of the experimental period, twelve birds per treatment (two per experimental unit, with an average weight within the same unit) were euthanized by cervical dislocation and sent to the Meat Laboratory of the Animal Science Department at UFRPE to measure the weight of the organs of the gastrointestinal tract (liver, gizzard, small intestine, large intestine, cecum, and pancreas), reproductive system (ovary and oviduct) and spleen, as well as the length of the intestines (small, large and caeca). A 0.01 g precision scale was used to obtain the weight data, while a tape measure was used for the length measurements. Weight results were expressed in absolute weight (g) and length measurements (cm).
2.5 Intestinal histology
At the end of the experimental period, two birds per experimental unit (twelve per treatment), different from those used to assess weight and organ length, were euthanized by cervical detachment. Samples of 4 cm from sections of the small intestine (duodenum, jejunum, and ileum), liver, pancreas, and spleen were collected, washed with saline solution, and immersed in 10% buffered formalin. After these, the tissues were dehydrated in a series of alcohols with increasing concentrations (10, 80, 90, and 100%), immersed in xylene, and embedded in paraffin.
The height of the villi was measured from their apex to their base, while the depth of the crypt was measured from the crypt’s base to the villus’s base. The villus-crypt ratio was calculated as the ratio between villi and crypts. For each segment and variable analyzed, 20 measurements were taken, totaling 40 per experimental unit.
2.6 Statistical analyses
Each cage, housing six birds, was considered an experimental unit. For each treatment, six cages were used, resulting in a total of 48 experimental units across all treatments. The data was analyzed for homoscedasticity and homogeneity of variances. One-way ANOVA was conducted using the GLM procedure in SAS 9.4 (SAS Institute Inc., 2012) (43). The influence of the individual factors and interactions on the variables was analyzed. In the presence of significant differences, the means were compared using the Tukey test at 5% probability. The equation below shows the statistical model.
in which:
Yijk is the observed value, μ is the population average, αi is the effect of the MOL (1–2), βj is the enzymatic supplementation effect (1–4), (αβ)ij is the interaction effect between MOL and enzymatic supplementation, and εijk is the residual error.
3 Results
3.1 Organs of the gastrointestinal tract, accessory glands and reproductive system
According to Table 3, a significant interaction was observed between the experimental factors for liver and pancreas weights, as well as for the lengths of the small intestine and caeca. For gizzard weight, only a main effect of MOL inclusion was detected, with birds fed diets containing 5% MOL exhibiting significantly heavier gizzards. No significant effects (p > 0.05), either individual or interactive, were found for the weights of the small intestine, large intestine, caeca, and spleen, nor for the length of the large intestine.
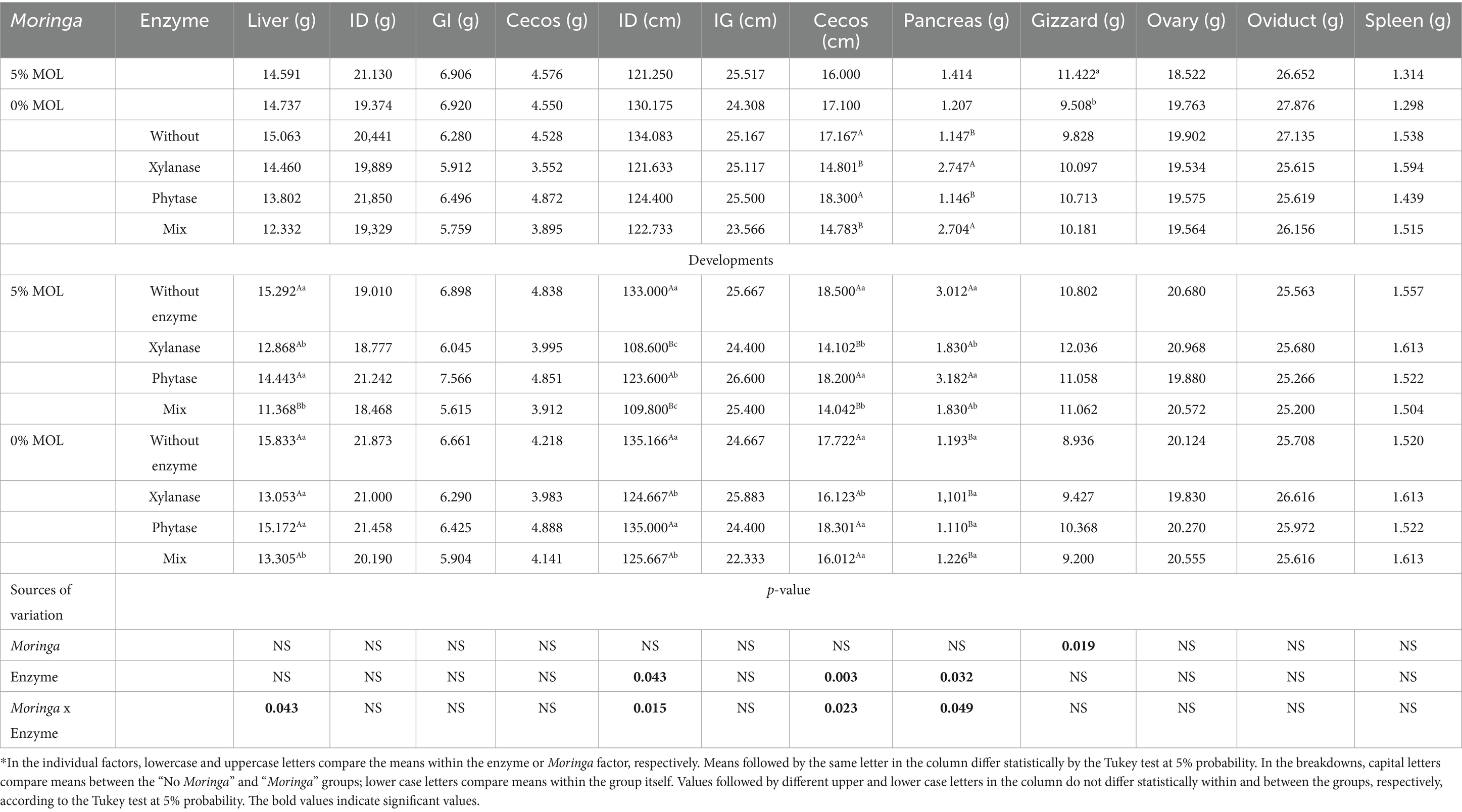
Table 3. Effects of individual factors and unfolding for the variables weight and length of the organs of the digestive and reproductive systems of laying hens fed diets containing Moringa and supplemented with enzymes.
As shown in Figure 2, dietary supplementation with xylanase, either alone or in combination with phytase, significantly reduced liver weight (p < 0.05) in the presence of MOL. In the absence of MOL, individual enzyme supplementation had no significant effect on liver weight; however, the combined use of xylanase and phytase tended to reduce liver weight (p < 0.05).
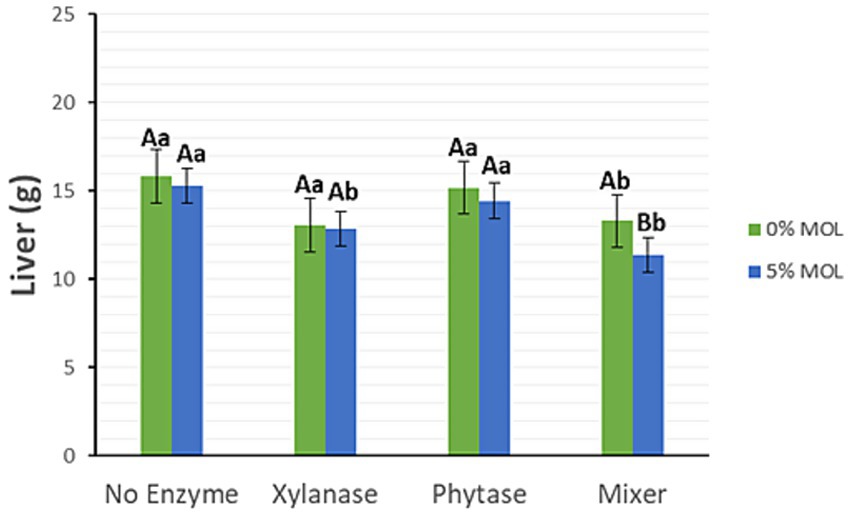
Figure 2. MOL x enzyme interaction for liver weight. Upper-case letters compare means between the “0% MOL” and “5% MOL” groups, lower case letters compare means within itself.
Xylanase supplementation in the presence of MOL, whether administered individually or in combination with phytase, significantly reduced the length of the small intestine (p < 0.05), as illustrated in Figure 3. Birds fed diets without enzyme supplementation, regardless of MOL inclusion, exhibited longer intestines. Diets containing phytase alone also resulted in longer intestines. A similar pattern was observed for caecal length (p < 0.05) (Figure 4).
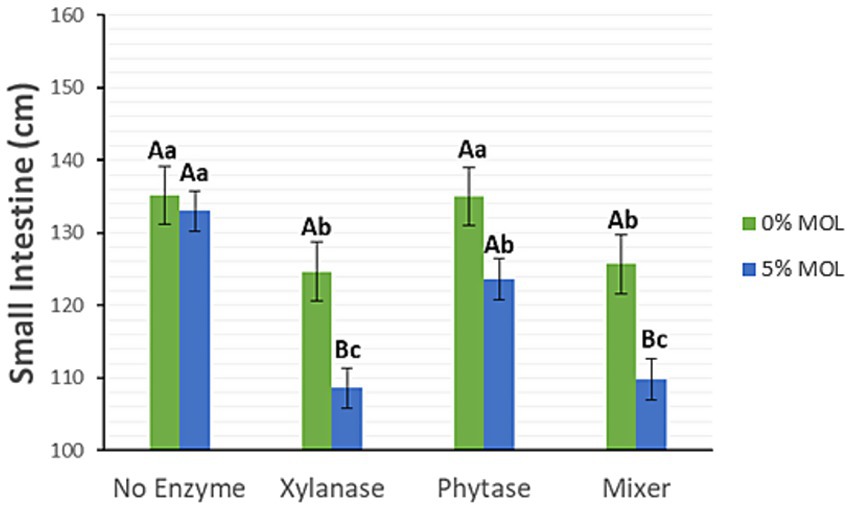
Figure 3. MOL x enzyme interaction for small intestine length. Upper-case letters compare means between the “0% MOL” and “5% MOL” groups, and lowercase letters compare means within the group itself.
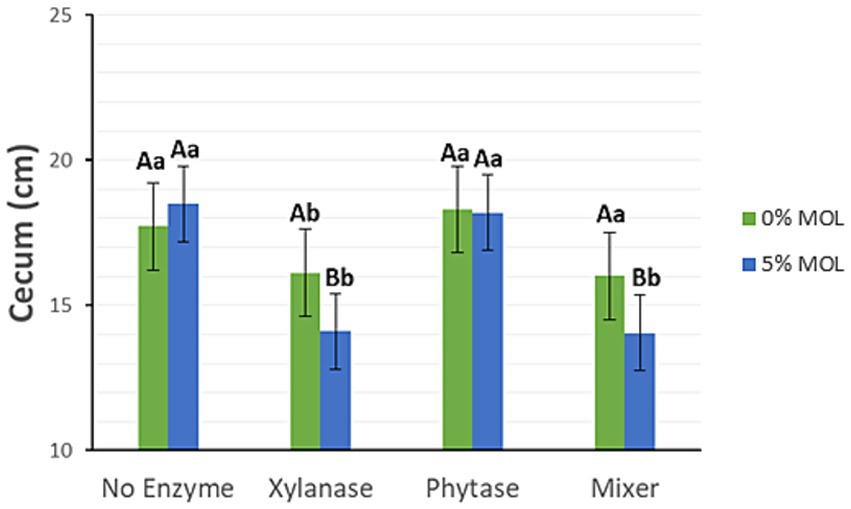
Figure 4. MOL x enzyme interaction for cecum length. Upper-case letters compare means between the “0% MOL” and “5% MOL” groups; lower case letters compare means within the group.
Figure 5 shows that the inclusion of MOL significantly increased pancreas weight. However, laying hens receiving MOL-supplemented diets along with xylanase—either alone or in combination with phytase—had significantly lower pancreas weights compared to those fed diets without enzymes or with phytase alone (p < 0.05).
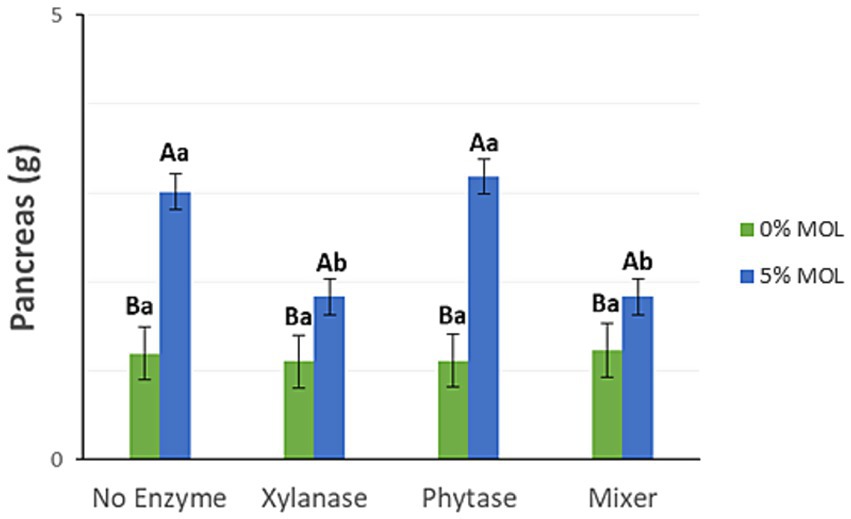
Figure 5. MOL x enzyme interaction for pancreas weight. Upper-case letters compare means between the “0% MOL” and “5% MOL” groups, lower case letters compare means within the group itself.
3.2 Intestinal morphology
Table 4 shows the intestinal morphology results of laying hens fed diets containing MOL supplemented with exogenous enzymes. The three intestinal segments analyzed had an interaction effect for all the variables studied.
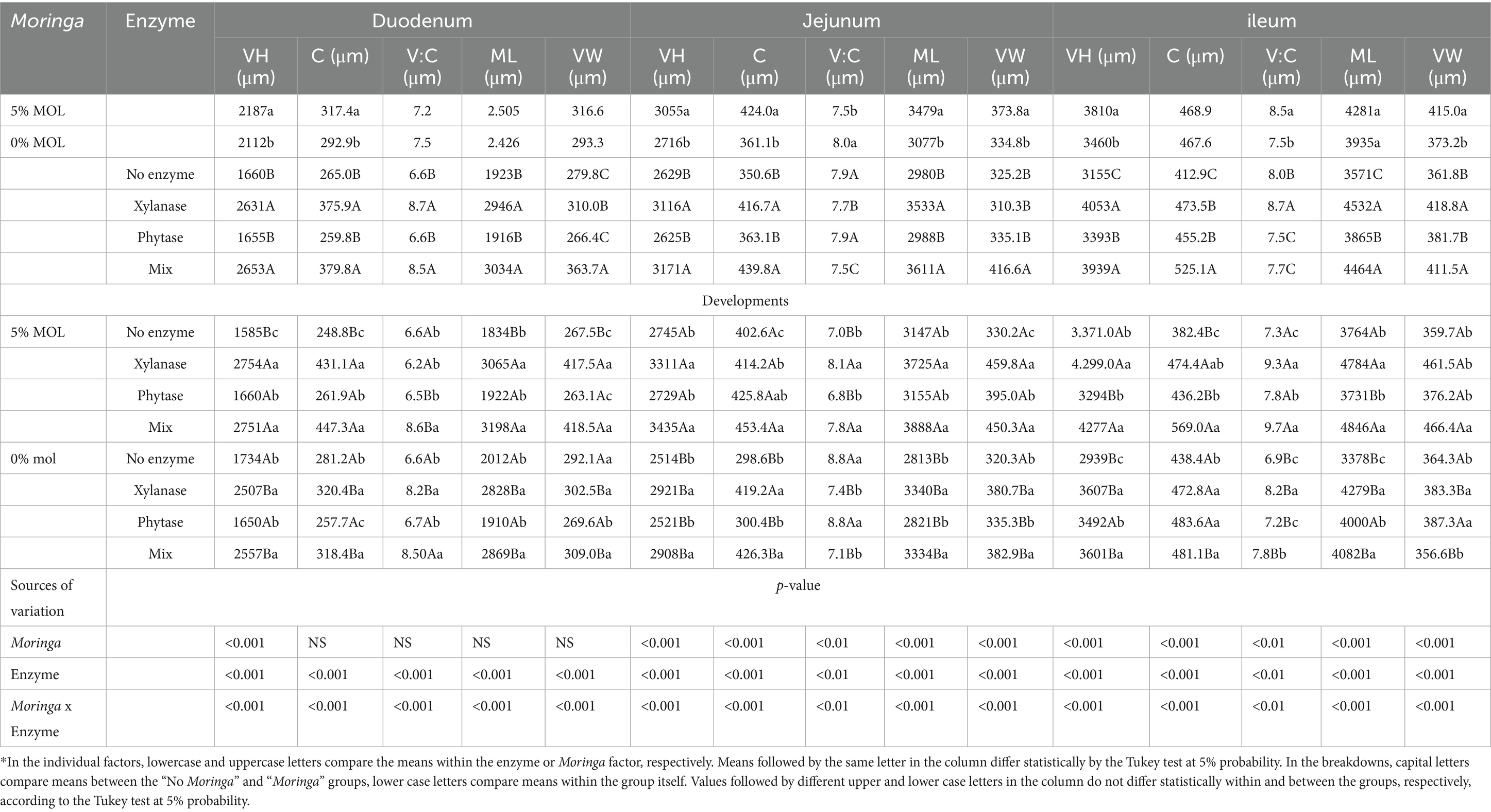
Table 4. Effects of individual factors and unfolding for the variables weight and length of the organs of the digestive and reproductive systems of laying hens fed diets containing Moringa and supplemented with enzymes.
3.3 Duodenum
Figure 6 presents the interaction effects between Moringa oleifera leaf meal (MOL) inclusion and exogenous enzyme supplementation on villus height (VH) and crypt depth (C) in the duodenum of laying hens. The addition of xylanase to MOL-containing diets significantly increased duodenal villus height, both when xylanase was used alone and when combined with phytase (p < 0.05). In contrast, phytase alone reduced VH in MOL-based diets (p < 0.05). Interestingly, the opposite was observed in the control (non-MOL) diets: phytase supplementation in this case increased VH (p < 0.05). A similar interaction pattern was found for crypt depth. Xylanase, whether used alone or in combination with phytase, significantly increased crypt depth (p < 0.05). In diets without MOL, xylanase also increased VH and C, though the effect was less pronounced than in the MOL-based diets (p < 0.05).
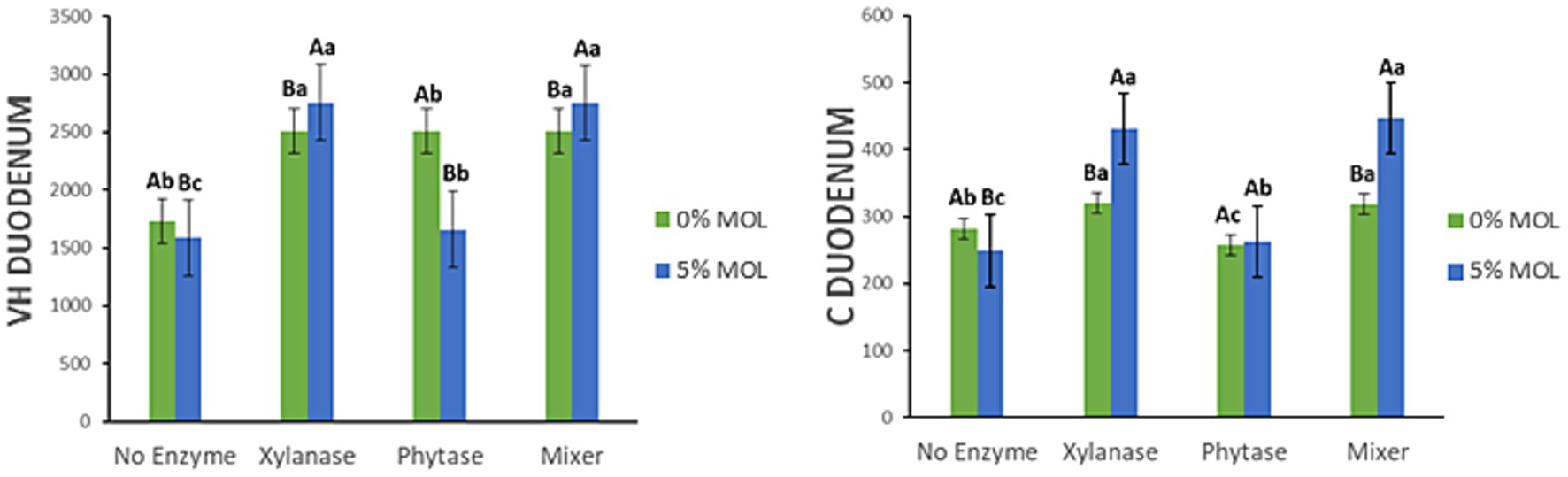
Figure 6. MOL x enzyme interaction for VH and C of the duodenum. Upper-case letters compare means between the “0% MOL” and “5% MOL” groups; lower case letters compare means within the group.
Figure 7 displays the interaction effects on the villus height-to-crypt depth ratio (V:C ratio) and mucosal length (ML) in the duodenum. The V:C ratio was significantly higher (p < 0.05) in hens fed MOL diets supplemented with the enzyme combination (xylanase + phytase), compared to those receiving only one of the enzymes. However, this increase was not significantly different from that observed in the control diet also supplemented with the enzyme mix (p > 0.05). Control diets supplemented with either xylanase or phytase individually also showed a significant increase in the V:C ratio (p < 0.05). Additionally, mucosal length was significantly greater in hens receiving MOL-based diets supplemented with xylanase, either alone or with phytase (p < 0.05). A similar but less marked increase in ML was observed in the control diets with xylanase (p < 0.05), while phytase alone had no significant effect on ML in either dietary context (p > 0.05).
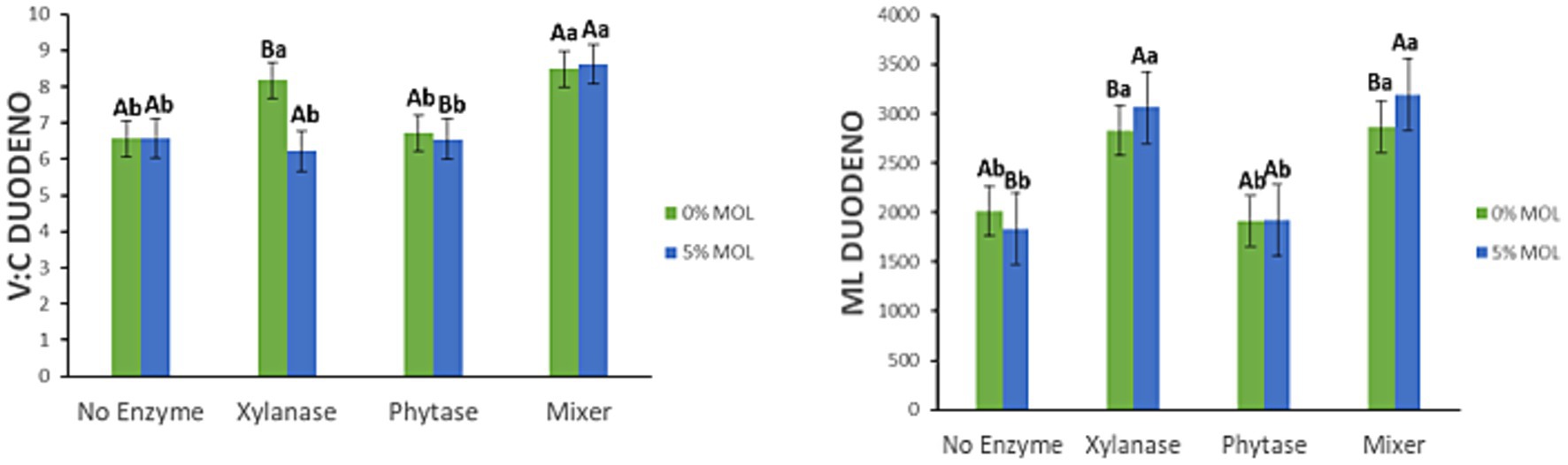
Figure 7. MOL x enzyme interaction for V:C and ML of the duodenum. Upper-case letters compare means between the “0% MOL” and “5% MOL” groups; lower case letters compare means within the group.
Figure 8 highlights the interaction effects on villus width (VW) in the duodenum. Birds fed MOL diets supplemented with xylanase—alone or in combination with phytase—had significantly wider villi (p < 0.05). Phytase on its own had no significant effect on VW, and birds fed MOL without any enzyme supplementation exhibited narrower villi.
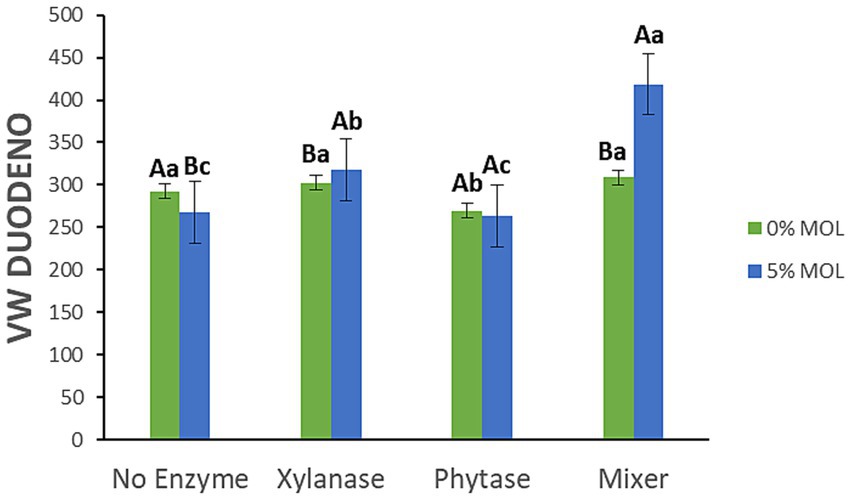
Figure 8. MOL x enzyme interaction for the VW of the duodenum. Upper-case letters compare means between the “0% MOL” and “5% MOL” groups; lower case letters compare means within the group.
3.4 Jejunum
Figure 9 illustrates the interaction effects between Moringa oleifera leaf meal (MOL) inclusion and exogenous enzyme supplementation on villus height (VH) and crypt depth (C) in the jejunum of laying hens. In MOL-based diets, xylanase supplementation significantly increased VH (p < 0.05), with no notable difference between xylanase alone and its combination with phytase. Phytase alone also improved VH (p < 0.05), although the effect was less pronounced compared to xylanase. Interestingly, MOL inclusion without any enzyme supplementation significantly increased both VH and crypt depth (p < 0.05), indicating its inherent positive effect on intestinal morphology. The greatest crypt depth was observed in birds fed MOL in combination with the enzyme mix (p < 0.05), followed by those receiving phytase alone. However, crypt depth values in the xylanase-supplemented groups (alone or in combination) were not statistically different from the phytase group.
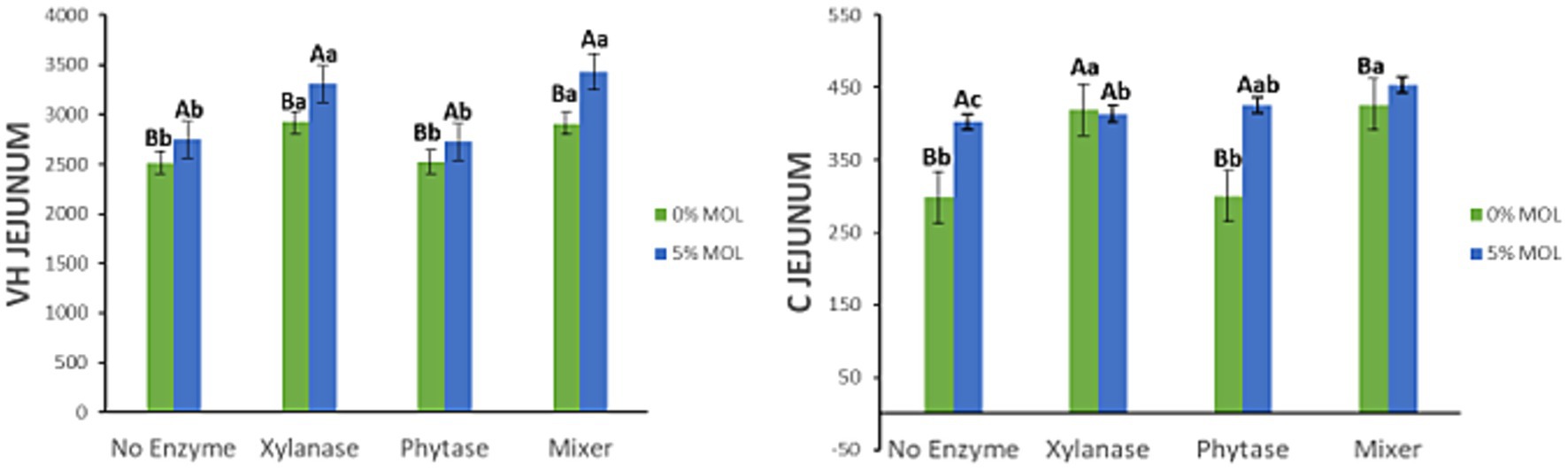
Figure 9. MOL x enzyme interaction for jejunal VH and C. Upper-case compare means between the “0% MOL” and “5% MOL” groups; lower case letters compare means within the group.
Figure 10 presents the interaction effects on the villus height-to-crypt depth ratio (V:C) and mucosal length (ML) in the jejunum. Diets containing MOL supplemented with xylanase, whether alone or in combination with phytase, significantly increased the V:C ratio (p < 0.05). In contrast, lower V:C ratios were observed in birds fed phytase alone or MOL alone (p < 0.05), suggesting that xylanase plays a more prominent role in enhancing intestinal morphology. Similarly, mucosal length was significantly greater in hens fed MOL diets supplemented with xylanase (p < 0.05). Although phytase and MOL alone also contributed to increased ML, the effect was comparatively modest.
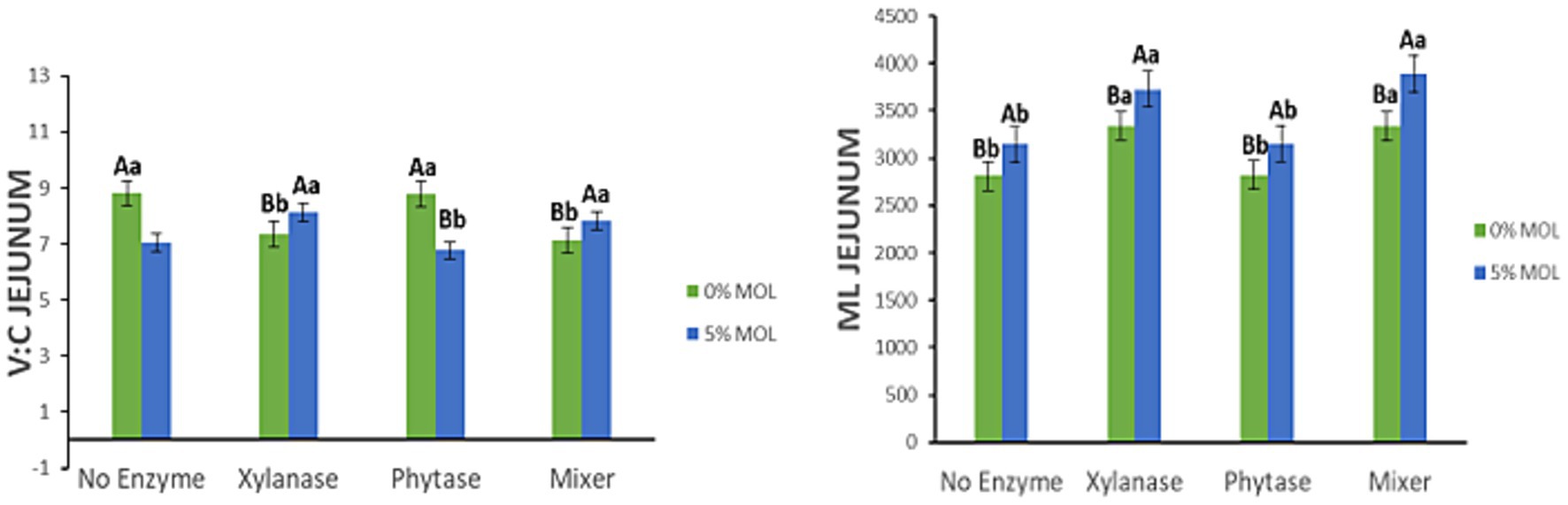
Figure 10. MOL x enzyme interaction for V:C and ML of the jejunum. Upper-case letters compare means between the “0% MOL” and “5% MOL” groups; lower-case letters compare means within the group.
Figure 11 shows the interaction effects on villus width (VW) in the jejunum. Birds fed MOL diets supplemented with xylanase, either alone or in combination with phytase, exhibited significantly wider villi (p < 0.05). No significant differences in VW were observed among birds fed diets without enzyme supplementation, indicating that xylanase was the primary contributor to increased villus width in these treatments.
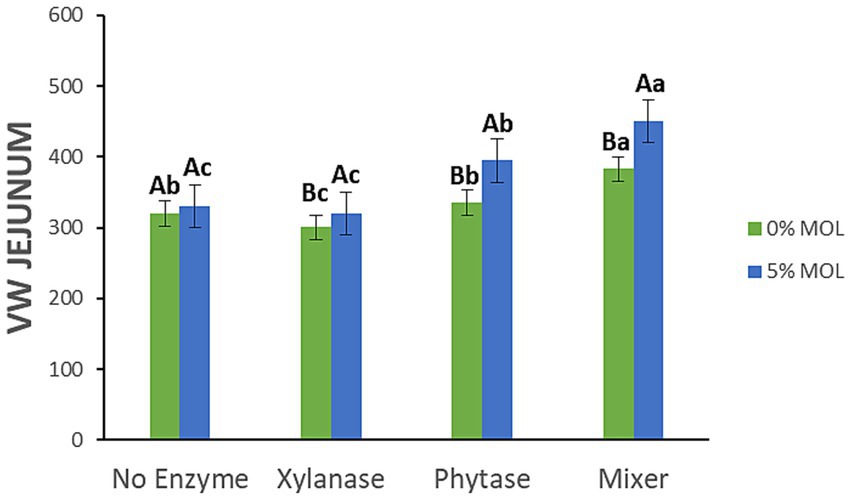
Figure 11. MOL x enzyme interaction for jejunal VW. Upper-case letters compare means between the “0% MOL” and “5% MOL” groups; lower case letters compare means within the group.
3.5 Ileum
Figure 12 shows the interaction effects of Moringa oleifera leaf meal (MOL) and exogenous enzymes on villus height (VH) and crypt depth (C) in the ileum of laying hens. Birds fed diets containing MOL with xylanase—either alone or combined with phytase—exhibited significantly taller ileal villi (p < 0.05). In contrast, phytase supplementation in MOL diets was associated with a significant reduction in villus height (p < 0.05). Notably, MOL inclusion without enzyme supplementation also significantly increased villus height (p < 0.05). Regarding crypt depth, the enzyme combination (MIX) produced the deepest ileal crypts (p < 0.05), while phytase alone and MOL alone were linked to a decrease in crypt depth. Xylanase alone did not significantly affect crypt depth in the ileum (p > 0.05).
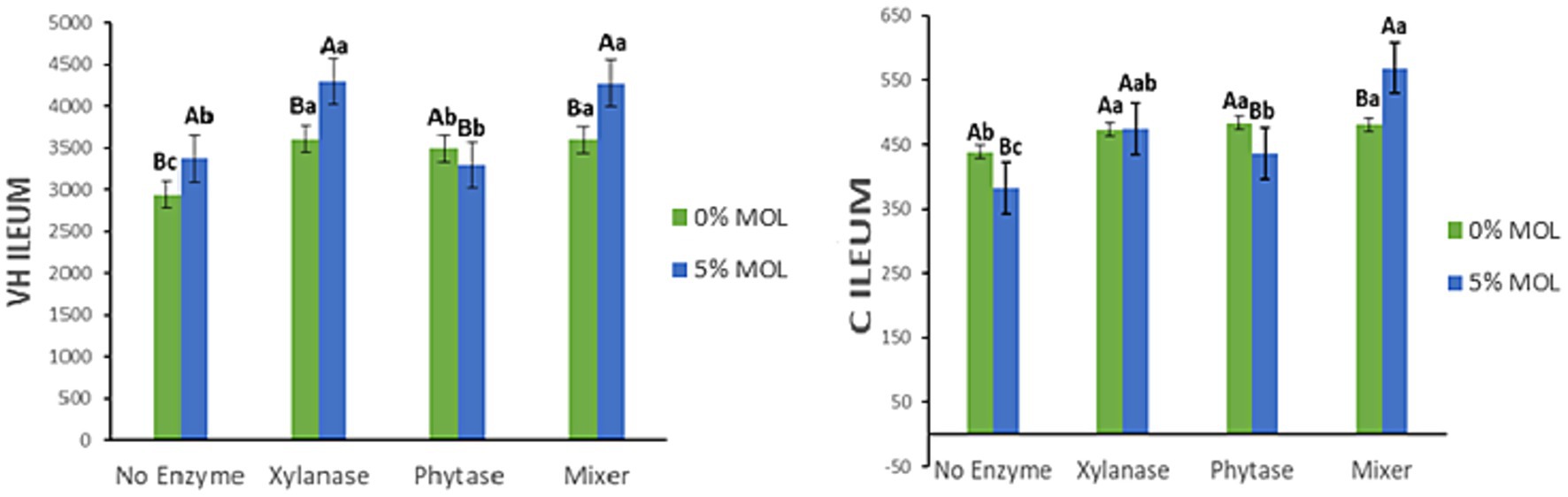
Figure 12. MOL x enzyme interaction for VH and C of the ileum. Upper-case letters compare means between the “0% MOL” and “5% MOL” groups; lower-case letters compare means within the group.
Figure 13 presents the interaction effects on the villus height-to-crypt depth ratio (V:C) and mucosal length (ML) in the ileum. Birds receiving MOL diets supplemented with xylanase, either individually or combined with phytase, had significantly higher V:C ratios (p < 0.05). Phytase and MOL alone also increased the V:C ratio but to a lesser extent (p < 0.05). A similar pattern was observed for mucosal length: xylanase supplementation in MOL diets significantly increased ileal mucosal length (p < 0.05). Conversely, phytase alone reduced mucosal length, while MOL alone increased it when no enzymes were included (p < 0.05).
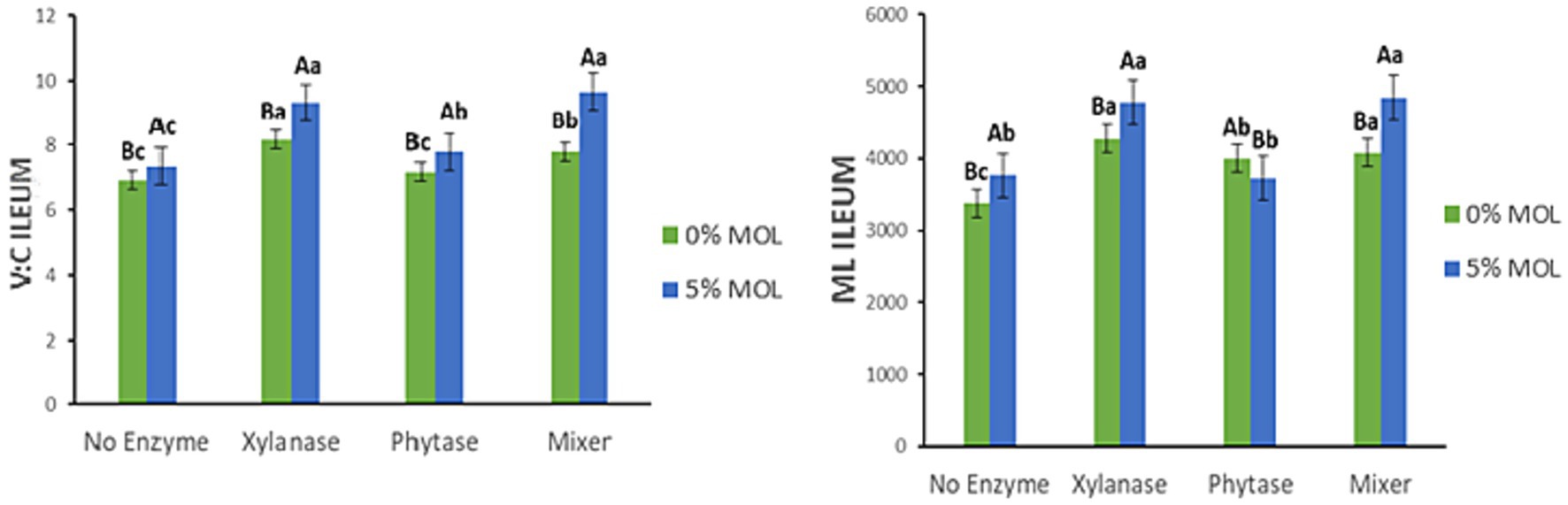
Figure 13. MOL x enzyme interaction for V:C and ML of the ileum. Upper-case letters compare means between the “0% MOL” and “5% MOL” groups; lower-case letters compare means within the group.
Figure 14 illustrates the interaction effects on villus width (VW) in the ileum. Significant increases in villus width were observed in birds fed MOL combined with xylanase, whether supplemented individually or in combination with phytase (p < 0.05). Neither phytase nor MOL alone had a significant effect on villus width in this intestinal segment (p > 0.05).
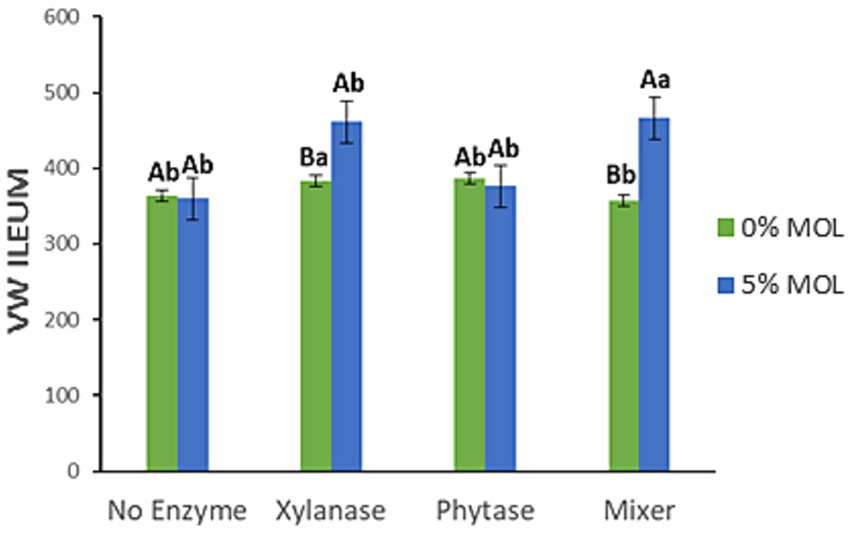
Figure 14. MOL x enzyme interaction for the VW of the ileum. Upper-case letters compare means between the “0% MOL” and “5% MOL” groups, and lower-case letters compare means within the group.
4 Discussion
The liver is the largest internal organ in the body, accounting for around 3% of a chicken’s body weight, and its size is associated with the age and body condition of the animal. In this study, the average body weight of the chickens was 1.725 kg (20). Research has shown that Moringa oleifera leaves possess antioxidant activity due to the presence of compounds such as flavonoids, ascorbic acid, alpha-tocopherol, beta-carotene, polyphenols, thiocarbons, glycosides, and phenolic compounds, which can serve as preventive agents against liver damage (13–15, 19, 20). Although some studies have reported an increase in liver weight in poultry fed Moringa oleifera leaf meal (MOL) (44), most research has found no significant effect of MOL on the relative liver size or function in broilers and laying hens (20, 45–47). These previous findings contrast with the results of the present study, which observed a significant influence of MOL on relative liver weight. The observed reduction in liver size with the combination of xylanase and Moringa oleifera may be explained by improved nutrient digestibility and absorption, leading to decreased metabolic stress on the liver. Furthermore, the antioxidant and anti-inflammatory compounds in MOL could protect liver tissue by preventing inflammation and fat accumulation, which are factors that contribute to liver enlargement. Regarding the pancreas, previous research has shown that Moringa oleifera leaves can increase its size, mainly due to the presence of bioactive compounds (as previously mentioned), which enhance pancreatic activity and inhibit the growth of pathogenic microbes in the intestines of birds, thereby positively affecting chicken metabolism and nutrition (48, 49).
On the other hand, the NSP in corn, soybean meal, and MOL may explain the decreases observed in the size of the liver and pancreas and the length of the small intestine and caeca of the birds when xylanase was supplemented. The basic composition of NSP includes cellulose, hemicellulose, xylans, and arabinoxylans, among others (2, 3). According to Bach Knudsen (50), total NSP in corn and soybean meal are approximately 9.70 and 21.70%, respectively. In contrast, the values reported for MOL by Macambira et al. (22) show that most of the NSP fraction in the leaves belongs to the soluble fiber fraction, as observed in this study, and that a considerable amount of the fiber in MOL is hemicellulose and its constituents. NSP are recognized for their ability to increase the weight and length of the intestine in birds (51, 52). Generally, these animals respond quickly to changes in the fiber content of the feed, showing modifications in intestinal size and passage rate. Amerah et al. (53) observed an increase in the length of the small intestine of broiler chickens fed whole wheat compared to those fed insoluble NSP sources. According to Khan et al. (35), the increase observed in the length of the intestine of broiler chickens fed MOL is due to increased digesta residence time in the gastrointestinal tract induced by the high fiber content of the feed. Although no significant differences were found in the weight of the small and large intestines in this study, the increases in the size of the small intestine and caeca in diets containing MOL may be related to the higher amount of this type of fiber in the diets. NSP, with their adverse effects on digestion and intestinal transit, may have caused this portion of the gastrointestinal tract to enhance its secretory mechanisms, due to the increase in the amount of undigested substrate and, consequently, a greater need for digestive enzymes, which triggered an increase in total intestinal size and the weight of attached glands (23, 25). The increase in viscosity caused by soluble NSP stimulates the secretion of pancreatic juice and increases the spacing of the intestinal mucosal barrier, which hinders the contact of enzymes with substrates, thus impairing the formation of micelles and the digestion of lipids, fats, and carbohydrates (54). According to Sousa et al. (33), soluble NSP increase the metabolic activity of the liver, resulting in enhanced synthesis and secretion of bile acids due to the affinity of this type of fiber for these compounds. This increases their excretion and causes greater liver activity to restore normal levels of bile acids in the gastrointestinal tract, which can lead to increases in liver size.
When xylanase was added to the feed, the NSP were degraded, reducing the relative size of the glands, small intestine, and caeca. Other studies have shown a reduction in the size of the liver and pancreas when xylanase is supplemented in diets containing fibrous poultry feed (23, 55). Hoseini et al. (56) observed shorter small intestine lengths in birds fed wheat-based diets supplemented with carbohydrases. In this study, there were no effects of phytase on decreasing the weight or length of the intestines and glands, indicating that the observed results were mainly due to supplementation with xylanase, either individually or in combination.
The gizzard is an essential organ in feed digestion in poultry; it has strong and thick muscles with the primary function of mechanical digestion for grinding and, consequently, reducing the particle size of the food (20). Moringa oleifera leaves can modify the anatomical structure of the birds’ gastrointestinal tract due to their fibrous content (57, 58). It has already been reported that birds need a minimum amount of fiber to maintain the function of the gizzard and the activity of the gastrointestinal tract (59), which should not exceed levels greater than 5%, according to the recommendations in the manual for the strain used in this study. The inclusion of fiber in the diet improves the development of the gizzard, as the presence of these components in this segment increases its growth, enhances the motility of the digestive tract, and increases the secretion of cholecystokinin (CCK), thus improving the mixing of digestive enzymes with the digesta (10, 22, 60–62). The increase in fiber levels present in the rations containing MOL (Table 2) and the increased activity of the proventriculus to grind these components provide a plausible explanation for the increase in gizzard weight when the leaves were included in the diet. Teteh et al. (57) observed significant increases in the weight of the proventriculus at 56 days of age when they fed laying hens with increasing levels of MOL. However, the authors of that study did not specify the amount of fiber present in the leaves used; the determined levels of crude fiber in the laying rations did not exceed 5%. When xylanase was added to the feed, it degraded NSPs, reducing the relative size of the glands, small intestine, and caeca. Similar reductions in liver and pancreas size with xylanase supplementation in fibrous diets have been reported (23, 55). Hoseini et al. (56) also observed shorter small intestines in birds fed wheat-based diets with carbohydrases. In contrast, phytase had no significant effect on the weight or length of intestines and glands in this study, indicating that the changes were mainly due to xylanase supplementation, alone or combined.
The primary function of the gastrointestinal tract is to promote the digestion and absorption of nutrients for maintenance, growth, and production. Maintaining intestinal health is crucial for profitable and sustainable production systems, as disorders can negatively impact production efficiency, animal welfare, and environmental protection (63). The duodenum, jejunum, and ileum are key sites of digestion and nutrient absorption, with their surface area and epithelial properties influencing absorption capacity (30, 64, 65). Morphological characteristics such as villus height (VH), crypt depth (C), villus height/crypt depth ratio (V:C), mucosal length (ML), and villus width (VW) are commonly used to assess intestinal functional capacity and response to diet.
NSPs adversely affect the enteric mucosa, causing villus shortening and widening, reduced crypt depth, mucosal atrophy, and increased goblet cells, all impairing intestinal function (26, 28). Consistent with this, the present study found that NSPs in Moringa oleifera leaf meal (MOL) negatively influenced the small intestine histomorphology, evidenced by significant reductions in VH, C, V:C, ML, and VW across the duodenum, jejunum, and ileum of laying hens.
In contrast, supplementation with xylanase—either alone or in combination with phytase—mitigated these negative effects and significantly improved all evaluated intestinal parameters. These improvements align with previous findings reporting that xylanase enhances intestinal morphology by hydrolyzing NSPs and reducing their antinutritional effects (56, 66, 67). For example, the increased VH observed in our study suggests a greater number of enterocytes and enteroendocrine cells, along with enhanced expression of brush border enzymes, which contribute to improved digestion and nutrient absorption (68, 69).
Furthermore, our findings support the interpretation by Yason et al. (70), who stated that crypt depth reflects the regenerative activity of the intestinal lining. In our study, the greater crypt depth observed with xylanase supplementation indicates a healthier mucosal renewal process compared to the MOL-only treatment, where crypt depth was significantly reduced. Likewise, the lower VH and crypt depth seen in the MOL treatment are indicative of impaired absorption, while the higher V:C ratio in the xylanase-supplemented groups points to a more developed and functionally efficient intestinal mucosa (71). Similarly, the increases in villus width observed with xylanase supplementation in our study are consistent with literature linking wider villi to enhanced nutrient absorption due to increased surface area (30, 72).
Although the NSP content of the diets was not directly measured in this study, the significant improvements in intestinal morphology following xylanase supplementation strongly suggest the presence of hydrolysable NSP substrates in the MOL-based diets. This further supports findings from previous studies demonstrating the beneficial effects of xylanase in improving intestinal morphometry in poultry fed fibrous diets (23, 30, 65, 72–74). Thus, our results confirm that xylanase plays an important role in attenuating the negative effects of dietary fiber and in promoting intestinal health in laying hens fed MOL-containing diets.
However, it seems that MOL, depending on the supplementation level, maturity stage, and gastrointestinal tract section of the birds, exert a positive influence on the morphological characteristics of the intestines of these animals (35). These same researchers observed an increase in VH and the V:C ratio of the jejunum and ileum of broiler chickens fed diets containing 1.2% MOL, with a decline at higher inclusion levels. It should be noted that laying hens, being older animals and consequently having a more mature gastrointestinal tract, have a greater capacity to digest fiber. The effects observed in this study, where MOL without enzyme supplementation was able to increase VH, C, and ML of the jejunum, as well as ML and the V:C ratio of the ileum, show that the birds, even when fed diets without enzyme supplementation, seem to tolerate higher levels of fiber without significant impairment of the digestive and absorptive processes. In addition, Moringa oleifera leaves contain L-glutamine, an amino acid derivative of glutamate, which plays an important role in maintaining the integrity of the intestinal mucosa (75, 76). According to Rao and Sama (76), L-glutamine increases the rate of protein synthesis in the intestine, reduces proteolysis in enterocytes, and is used as an energy source for the proliferation of intestinal epithelial cells, which promotes greater intestinal integrity and, consequently, an improvement in intestinal morphology. Other studies have found better morphological characteristics in all sections of the intestines of birds fed MOL at inclusion levels ranging from 1.0 to 5.0% (36, 75, 77–80).
The effects of phytase on intestinal morphology observed in this study were limited and primarily restricted to the jejunum, where the enzyme increased villus height (VH), crypt depth (C), and mucosal length (ML) in diets containing Moringa oleifera leaf meal (MOL). Phytate levels in Moringa leaves can reach approximately 2.5% (81), and this compound is known to form insoluble complexes with minerals such as phosphorus, calcium, magnesium, iron, and zinc, as well as with proteins and other nutrients (82–85). Phytase supplementation has been reported to improve intestinal morphology mainly by reducing the anti-nutritional effects of phytate. This occurs through the hydrolysis of phytate, which not only enhances nutrient bioavailability but also limits the amount of substrate available for pathogenic microbial fermentation. Consequently, phytase may help modulate gut microbiota composition, reduce the inflammatory response, and minimize mucosal damage, thereby contributing to improved epithelial structure and function (86–88). In this study, such effects were evident through localized improvements in the jejunal mucosa of laying hens fed MOL-based diets.
5 Conclusion
MOL inclusion in the diet had multiple effects on the GIT of laying hens, including increasing the size of intestinal segments and associated glands, as well as modulating key histomorphological parameters. The supplementation of xylanase, through its ability to degrade NSP, contributes to the restoration of intestinal integrity and function, potentially improving nutrient absorption and overall performance in laying hens. However, further studies are warranted to characterize the specific NSP profile of Moringa oleifera leaves to better understand their interactions with exogenous enzymes and optimize their inclusion in poultry diets.
Data availability statement
The original contributions presented in the study are included in the article/supplementary material, further inquiries can be directed to the corresponding author.
Ethics statement
The animal study was approved by Ethics Committee on the Use of Animals (CEUA), of the Federal Rural University of Pernambuco, in accordance with license number 21/2018. The study was conducted in accordance with the local legislation and institutional requirements.
Author contributions
GM: Resources, Writing – original draft, Investigation, Formal analysis, Visualization, Supervision, Project administration, Conceptualization, Methodology, Writing – review & editing, Data curation. CR: Data curation, Supervision, Investigation, Methodology, Conceptualization, Writing – review & editing, Validation, Resources, Visualization, Formal analysis, Funding acquisition, Project administration, Writing – original draft. HO: Formal analysis, Data curation, Writing – review & editing, Conceptualization. MS: Formal analysis, Conceptualization, Data curation, Software, Writing – review & editing. AR: Formal analysis, Writing – review & editing, Data curation, Conceptualization. OL: Writing – review & editing, Formal analysis, Data curation. LS: Formal analysis, Data curation, Writing – review & editing. IM: Conceptualization, Data curation, Writing – review & editing. LC: Data curation, Conceptualization, Writing – review & editing. OF: Conceptualization, Writing – review & editing, Data curation.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This work was supported by Karolinska Institutet through an institutional publication agreement. The funding covered the open access publication.
Acknowledgments
The authors thank the Coordination for the Improvement of Higher Education Personnel (CAPES), the National Council for Scientific and Technological Development (CNPq).
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The authors declare that no Gen AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
1. Bederska-Łojewska, D, Świątkiewicz, S, Arczewska-Włosek, A, and Schwarz, T. Rye non-starch polysaccharides: their impact on poultry intestinal physiology, nutrients digestibility and performance indices - a review. Ann Anim Sci. (2017) 17:351–69. doi: 10.1515/aoas-2016-0090
2. Smits, CHM, and Annison, G. Non-starch plant polysaccharides in broiler nutrition - towards a physiologically valid approach to their determination. Worlds Poult Sci J. (1996) 52:203–21. doi: 10.1079/WPS19960016
3. Fortes, BDA, Cafe, MB, Stringhini, JH, Brito, JÁG, Rezende, PP, and Silva, RD. Evaluation of nutritional programs using carbohydrases and phytase in broiler feed. Ciência Animal Brasileira. (2012) 13:24–32. doi: 10.5216/cab.v13i1.8705
4. Adeola, O, and Bedford, MR. Exogenous dietary xylanase ameliorates viscosity-induced anti-nutritional effects in wheat-based diets for white Pekin ducks (Anas platyrinchos domesticus). Br J Nutr. (2004) 92:87–94. doi: 10.1079/BJN20041180
5. Tahir, M, Saleh, F, Ohtsuka, A, and Hayashi, K. An effective combination of carbohydrases that enables reduction of dietary protein in broilers: importance of hemicellulase. Poult Sci. (2008) 87:713–8. doi: 10.3382/ps.2007-00340
6. Tavernari, FC, Carvalho, TS, Assis, AP, and Lima, HJD. Soluble non-starch polysaccharides in pig and poultry diets. Revista Etrônica Nutritime. (2008) 5:673–89. Available at: https://nutritime.com.br/artigos/artigo-068-polissacarideo-nao-amilaceo-soluvel-na-dieta-de-suinos-e-aves/
7. Nitrayová, S, Heger, J, Patráš, P, Kluge, H, and Brož, J. Effect of xylanase on apparent ileal and total tract digestibility of nutrients and energy of rye in young pigs. Arch Anim Nutr. (2009) 63:281–91. doi: 10.1080/17450390903020455
8. Jha, R, and Berrocoso, JD. Review: dietary fiber utilization and its effects on physiological functions and gut health of swine. Animal. (2015) 9:1441–52. doi: 10.1017/S1751731115000919
9. Cao, BH, Zhang, XP, Guo, YM, Karasawa, Y, and Kumao, T. Effects of dietary cellulose levels on growth, nitrogen utilization, retention time of diets in digestive tract and caecal microflora of chickens. Asian Australas J Anim Sci. (2003) 16:863–6. doi: 10.5713/ajas.2003.863
10. Mateos, GG, Jiménez-Moreno, E, Serrano, MP, and Lázaro, RP. Poultry response to high levels of dietary fiber sources varying in physical and chemical characteristics. J Appl Poult Res. (2012) 21:156–74. doi: 10.3382/japr.2011-00477
11. Tejeda, OJ, and Kim, WK. Role of dietary fiber in poultry nutrition. Animals. (2021) 11:461. doi: 10.3390/ani11020461
12. Abdel-Wareth, AAA, and Lohakare, J. Moringa oleifera leaves as eco-friendly feed additive in diets of Hy-line brown hens during the late laying period. Animals. (2021) 11:1–10. doi: 10.3390/ani11041116
13. Makkar, HPS, and BECKER, K. Nutritional value and antinutritional components of whole and ethanol extracted Moringa oleifera leaves. Anim Feed Sci Technol. (1996) 63:211–28. doi: 10.1016/S0377-8401(96)01023-1
14. Siddhuraju, P, and Becker, K. Antioxidant properties of various solvent extracts of total phenolic constituents from three different agroclimatic origins of drumstick tree (Moringa oleifera lam.) leaves. J Agric Food Chem. (2003) 51:2144–55. doi: 10.1021/jf020444+
15. Anwar, F, Latif, S, Ashraf, M, and Gilani, AH. Moringa oleifera: a food plant with multiple medicinal uses. Phytopheraphy Research. (2007) 21:17–25. doi: 10.1002/ptr.2023
16. Rajanandh, MG, and Kavitha, J. Quantitative estimation of β-sitosterol, total phenolic and flavonoid compounds in the leaves of Moringa oleifera. Int J PharmTech Res. (2010) 2:1409–14. Available at: https://www.researchgate.net/publication/237844717_Quantitative_estimation_of_b-Sitosterol_total_phenolic_and_flavonoid_compounds_in_the_leaves_of_Moringa_oleifera
17. Moyo, B, Masika, PJ, Hugo, A, and Muchenje, V. Nutritional characterization of Moringa (Moringa oleifera lam.) leaves. Afr J Biotechnol. (2011) 10:12925–33. doi: 10.5897/AJB10.1599
18. Nkukwana, TT, Muchenje, V, Pieterse, E, Masika, PJ, Mabusela, TP, Hoffman, LC, et al. Effect of Moringa oleifera leaf meal on growth performance, apparent digestibility, digestive organ size and carcass yield in broiler chickens. Livest Sci. (2014) 161:139–46. doi: 10.1016/j.livsci.2014.01.001
19. Hassan, HMA, El-Moniary, MM, Hamouda, Y, El-Daly, EF, Youssef, AW, Abd El-Azeem, NA, et al. Effect of different levels of moringa oleifera leaves meal on productive performance, carcass characteristics and some blood parameters of broiler chicks reared under heat stress conditions. Asian J Animal Vet Advan. (2016) 11:60–6. doi: 10.3923/ajava.2016.60.66
20. Mardewi, NK, Suwitari, NKE, Rukmini, NKS, IGADS, R, and NMGR, A. Effect of Moringa (Moringa oleifera) leaf meal supplementation in broiler chicken ration on weight of internal organs, HDL and triglyceride levels. SEAS. (2017) 1:46–51. doi: 10.22225/seas.1.2.397.46-51
21. Han, Z, Li, C, and Liu, G. Recent advances in the extraction, purification and analytical techniques for flavonoids from plants: taking hawthorn as an example. J Food Compos Anal. (2025) 144:107372. doi: 10.1016/j.jfca.2025.107372
22. Macambira, GM, Rabello, CBV, Navarro, MIV, Ludke, MCMM, Silva, JCR, Lopes, EC, et al. Chemical and nutritional characterization of moringa oleifera leaves for broilers. Arquivo Brasileiro de Medicina Veterinaria e Zootecnia. (2018) 70:570–8. doi: 10.1590/1678-4162-9522
23. Wang, ZR, Qiao, SY, Lu, WQ, and Li, DF. Effects of enzyme supplementation on performance, nutrient digestibility, gastrointestinal morphology, and volatile fatty acid profiles in the hindgut of broilers fed wheat-based diets. Poult Sci. (2005) 84:875–81. doi: 10.1093/ps/84.6.875
24. Vahjen, W, Osswald, T, Schäfer, K, and Simon, O. Comparison of a xylanase and a complex of non starch polysaccharide- degrading enzymes with regard to performance and bacterial metabolism in weaned piglets. Arch Anim Nutr. (2007) 61:90–102. doi: 10.1080/17450390701203881
25. Abdollahi, A, Karimi, A, Sadeghi, AA, Bedford, MR, and Ashengroph, M. The effects of the fiber source and xylanase supplementation on production, egg quality, digestibility, and intestinal morphology in the aged laying hen. Poult Sci. (2021) 100:100936. doi: 10.1016/j.psj.2020.12.033
26. Viveros, A, Brenes, A, Pizarro, M, and Castaño, M. Effect of enzyme supplementation of a diet based on barley, and autoclave treatment, on apparent digestibility, growth performance and gut morphology of broilers. Anim Feed Sci Technol. (1994) 48:237–51. doi: 10.1016/0377-8401(94)90175-9
27. Santin, E, Maiorka, A, Macari, M, Grecco, M, Sanchez, JC, Okada, TM, et al. Performance and intestinal mucosa development of broiler chickens fed diets containing Saccharomyces cerevisiae cell wall. J Appl Poult Res. (2001) 10:236–44. doi: 10.1093/japr/10.3.236
28. Hetland, H, Choct, M, and Svihus, B. Role of insoluble non-starch polysaccharides in poultry nutrition. Worlds Poult Sci J. (2004) 60:415–22. doi: 10.1079/WPS200325
29. Baurhoo, B, Phillip, L, and Ruiz-Feria, CA. Effects of purified lignin and mannan oligosaccharides on intestinal integrity and microbial populations in the ceca and litter of broiler chickens. Poult Sci. (2007) 86:1070–8. doi: 10.1093/ps/86.6.1070
30. Apperson, KD, and Cherian, G. Effect of whole flax seed and carbohydrase enzymes on gastrointestinal morphology, muscle fatty acids, and production performance in broiler chickens. Poult Sci. (2017) 96:1228–34. doi: 10.3382/ps/pew371
31. Rojas, IYM, González, EA, Menocal, JA, Santos, TT, Arguello, JR, and Coello, CL. Assessment of a phytase included with lactic acid on productive parameters and on deposition of phosphorus, calcium, and zinc in laying hens fed with sorghum-soybean-meal-based diets. J Appl Anim Res. (2018) 46:314–21. doi: 10.1080/09712119.2017.1299740
32. Taylor, AE, Bedford, MR, Pace, SC, and Miller, HM. The effects of phytase and xylanase supplementation on performance and egg quality in laying hens. Br Poult Sci. (2018) 59:554–61. doi: 10.1080/00071668.2018.1483575
33. Sousa, LS, Carvalho, TSM, Nogueira, FA, Saldanha, MM, Vaz, DP, Bertechini, AG, et al. Fiber source and xylanase on performance, egg quality, and gastrointestinal tract of laying hens. Rev Bras Zootec. (2019) 48:1–10. doi: 10.1590/rbz4820170286
34. Chen, Y, Shi, Y, Li, M, Ming, D, Liu, W, Xu, X, et al. Phase separation-mediated multienzyme assembly in vivo. J Agric Food Chem. (2025) 73:7867–76. doi: 10.1021/acs.jafc.4c09585
35. Khan, I, Zaneb, H, Masood, S, Yousaf, MS, Rehman, HF, and Rehman, H. Effect of Moringa oleifera leaf powder supplementation on growth performance and intestinal morphology in broiler chickens. Animal Physiol Animal Nutrit. (2017) 101:114–21. doi: 10.1111/jpn.12634
36. Moreno-Mendoza, Y, López-Villarreal, KD, Hernández-Martínez, CA, Rodríguez-Tovar, LE, Hernández-Coronado, AC, Soto-Domínguez, A, et al. Effect of moringa leaf powder and agave inulin on performance, intestinal morphology, and meat yield of broiler chickens. Poult Sci. (2021) 100:738–45. doi: 10.1016/j.psj.2020.11.058
37. Detmann, E, Souza, MA, Valadares Filho, SC, Queiroz, AC, Berchielli, TT, Saliba, EOS, et al. Methods for food analysis. Visconde do Rio Branco: Suprema (2012). 214 p.
38. Van Soest, PJ. Development of a comprehensive system of feed analyses and its application to forages. J Anim Sci. (1967) 26:119–28. doi: 10.2527/jas1967.261119x
39. Engelen, AJ, Van Der Heeft, FC, Randsdorp, PH, and Smtt, EL. Simple and rapid determination of phytase activity. J AOAC Int. (1994) 77:760–4. doi: 10.1093/jaoac/77.3.760
40. Rostagno, HS, Albino, LFT, Hannas, MI, Donzele, JL, Sakomura, NK, Perazzo, FG, et al. Tabelas brasileiras para aves e suínos: composição de alimentos e exigências nutricionais. Minas Gerais: Federal University of Viçosa (2017).
41. Silva, JCR (2018) Use of Moringa oleifera in the feeding of broilers and laying hens. Federal Rural University of Pernambuco - UFRPE, PhD thesis, Department of Animal Science (Recife, PE). 86p. Available at: http://www.tede2.ufrpe.br:8080/tede2/bitstream/tede2/8102/2/Jaqueline%20de%20Cassia%20Ramos%20da%20Silva.pdf
42. FEDNA. Spanish Foundation for the Development of Animal Nutrition. (2010). Available online at: http://fundacionfedna.org/ecuaciones/ecuaciones-de-regresion-tablas-fedna-2010-harinas-alfalfa (Accessed May 27, 2024).
43. SAS Institute Inc. SAS/STAT software: Changes and enhancements through release 9.4. Carey, NC, USA: SAS Institute Inc. (2012).
44. Onunkwo, DN, and George, OS. Effects of Moringa oleifera leaf meal on the growth performance and carcass characteristics of broiler birds. IOSR J Agri Vet Sci II. (2015) 8:2319–72. Available at: https://iosrjournals.org/iosr-javs/papers/vol8-issue3/Version-2/M08326366.pdf
45. Ochi, EB, Elbushra, ME, Fatur, M, Abubakr, OI, and Hafiz, A. Effect of moringa (Moringa oleifera lam) seeds on the performance and carcass characteristics of broiler chickens. J Nat Sci Res. (2015) 5:66–73. Available at: https://www.researchgate.net/publication/324543462_Effect_of_Moringa_Moringa_oleifera_Lam_Seeds_on_the_Performance_and_Carcass_Characteristics_of_Broiler_Chickens
46. Lu, W, Wang, J, Zhang, HJ, Wu, SG, and Qi, GH. Evaluation of Moringa oleifera leaf in laying hens: effects on laying performance,egg quality,plasma biochemistry and organ histopathological indices. Ital J Anim Sci. (2016) 15:658–65. doi: 10.1080/1828051X.2016.1249967
47. Ashour, EA, El-Kholy, MS, Alagawany, M, Abd El-Hack, ME, Mohamed, LA, Taha, AE, et al. Effect of dietary supplementation with moringa oleifera leaves and/or seeds powder on production, egg characteristics, hatchability and blood chemistry of laying Japanese quails. Sustainability (Basel). (2020) 12:1–9. doi: 10.3390/su12062463
48. Teteh, A, Lawson, E, Tona, K, Decuypere, E, and Gbeassor, M. Moringa oleifera leave: hydro-alcoholic extract and effects on growth performance of broilers. Int J Poult Sci. (2013) 12:401–5. doi: 10.3923/ijps.2013.401.405
49. Antara, IKJ, Bidura, IGNG, and Siti, NW. Effects of Moringa oleifera leaf and probiotics mixed fermented extract on the egg production and cholesterol contents in egg of laying hens. Int J Fauna Biol Stud. (2019) 6:6–12. Available at: https://www.faunajournal.com/archives/2019/vol6issue5/PartA/6-4-19-999.pdf
50. Bach Knudsen, KE. The nutritional significance of "dietary fiber" analysis. Anim Feed Sci Technol. (2001) 90:3–20. doi: 10.1016/S0377-8401(01)00193-6
51. Jørgensen, H, Zhao, XQ, Knudsen, KE, and Eggum, BO. The influence of dietary fibre source and level on the development of the gastrointestinal tract, digestibility and energy metabolism in broiler chickens. Br J Nutr. (1996) 75:379–95. doi: 10.1079/bjn19960141
52. Smits, CHM, Veldman, A, Verstegen, MWA, and Beynen, AC. Dietary carboxymethylcellulose with high instead of low viscosity reduces macronutrient digestion in broiler chickens. J Nutr. (1997) 127:483–7. doi: 10.1093/jn/127.3.483
53. Amerah, AM, Ravindran, V, and Lentle, RG. Influence of insoluble fibre and whole wheat inclusion on the performance, digestive tract development and ileal microbiota profile of broiler chickens. Br Poult Sci. (2009) 50:366–75. doi: 10.1080/00071660902865901
54. Mirzaie, S, Zaghari, M, Aminzadeh, S, Shivazad, M, and Mateos, GG. Effects of wheat inclusion and xylanase supplementation of the diet on productive performance, nutrient retention, and endogenous intestinal enzyme activity of laying hens. Poult Sci. (2012) 91:413–25. doi: 10.3382/ps.2011-01686
55. Pirgozliev, V, Whiting, IM, Mansbridge, SC, Enchev, S, Rose, SP, Kljak, K, et al. Effect of rearing temperature on physiological measures and antioxidant status of broiler chickens fed stevia (Stevia rebaudiana B.) leaf meal and exogenous xylanase. Curr Res Biotechnol. (2021) 3:173–81. doi: 10.1016/j.crbiot.2021.05.005
56. Hoseini, SM, Akbari, MR, Alzawqari, MH, Mehrban, H, and Moharrery, A. Effect of L-threonine and NSP-degrading enzyme on the performance, intestinal during the starter period. Poult Sci. (2021) 9:7–18. doi: 10.22069/psj.2021.18281.1617
57. Teteh, A, Voemesse, K, Agbonon, A, Gbeassor, M, Decuypere, E, and Tona, K. Effect of Moringa oleifera leaves on feed transit and morphometric parameters of the digestive tract of layer pullets and laying hens. Euro Poultry Sci. (2017) 81:1–11. doi: 10.1399/eps.2017.173
58. Macambira, GM, Rabello, CB-V, Navarro, MIV, Lopes, CC, Lopes, EC, Nascimento, GR, et al. Effects of Moringa oleifera leaf meal on performance and carcass yield of broilers. Rev Bras Zootec. (2022) 51:e20210203. doi: 10.37496/rbz5120210203
59. Jiménez-Moreno, E, González-Alvarado, JM, González-Sánchez, D, Lázaro, R, and Mateos, GG. Effects of type and particle size of dietary fiber on growth performance and digestive traits of broilers from 1 to 21 days of age. Poult Sci. (2010) 89:2197–212. doi: 10.3382/ps.2010-00771
60. Hetland, H, Svihus, B, and Choct, M. Role of insoluble fiber on gizzard activity in layers. J Appl Poult Res. (2005) 14:38–46. doi: 10.1093/japr/14.1.38
61. Svihus, B, Juvik, E, Hetland, H, and Krogdahl, A. Causes for improvement in nutritive value of broiler chicken diets with whole wheat instead of ground wheat. Br Poult Sci. (2004) 45:55–60. doi: 10.1080/00071660410001668860
62. Macambira, GM, Rabello, CBV, Oliveira, HSH, Santos, MJB, Silva, DA, Santos, ACF, et al. Xylanase and phytase supplementation of Moringa oleifera diets: effects on the performance, egg quality and blood profile of laying hens. Anim Prod Sci. (2024) 64:AN24194. doi: 10.1071/AN24194
63. Yan, F, Dibner, JJ, Knight, CD, and Vazquez-Anon, M. Effect of carbohydrase and protease on growth performance and gut health of young broilers fed diets containing rye, wheat, and feather meal. Poult Sci. (2017) 96:817–28. doi: 10.3382/ps/pew300
64. Ferrer, C, Pedragosa, E, Torras-Llort, M, Parcerisa, X, Rafecas, M, Ferrer, R, et al. Dietary lipids modify brush border membrane composition and nutrient transport in chicken small intestine. J Nutr. (2003) 133:1147–53. doi: 10.1093/jn/133.4.1147
65. Westbrook, LA, and Cherian, G. Egg quality, fatty-acid composition and gastrointestinal morphology of layer hens fed whole flaxseed with enzyme supplementation. Br Poult Sci. (2019) 60:146–53. doi: 10.1080/00071668.2018.1556783
66. Kalmendal, R, and Tauson, R. Effects of a xylanase and protease, individually or in combination, and an ionophore coccidiostat on performance, nutrient utilization, and intestinal morphology in broiler chickens fed a wheat-soybean meal-based diet. Poult Sci. (2012) 91:1387–93. doi: 10.3382/ps.2011-02064
67. Gonzalez-Ortiz, G, Sola-Oriol, D, Martinez-Mora, M, Perez, JF, and Bedford, MR. Response of broiler chickens fed wheat-based diets to xylanase supplementation. Poult Sci. (2017) 96:2776–85. doi: 10.3382/ps/pex092
68. Caspary, WF. Physiology and pathophysiology of intestinal absorption. Am J Clin Nutr. (1992) 55:299–308. doi: 10.1093/ajcn/55.1.299s
69. Uni, Z, Zaiger, G, Gal-Garber, O, Pines, M, Rozenboim, I, and Reifen, R. Vitamin a deficiency interferes with proliferation and maturation of cells in the chicken small intestine. Br Poult Sci. (2000) 41:410–5. doi: 10.1080/713654958
70. Yason, CV, Summers, BA, and Schat, KA. Pathogenesis of rotavirus infection in various age groups of chickens and turkeys: pathology. Am J Vet Res. (1987) 48:927–38. doi: 10.2460/ajvr.1987.48.06.927
71. Xu, ZR, Hu, CH, Xia, MS, Zhan, XA, and Wang, MQ. Effects of dietary fructooligosaccharide on digestive enzyme activities, intestinal microflora and morphology of male broilers. Poult Sci. (2003) 82:1030–6. doi: 10.1093/ps/82.6.1030
72. Fallah, R, Saghafi, M, Rezaei, H, and Parvar, R. Effect of bioplus 2B and Protoxin probiotics supplementation on ® growth performance, small intestinal morphology and carcass characteristics of broiler chickens. British J Poultry Sci. (2013) 2:11–5. doi: 10.5829/idosi.bjps.2013.2.2.73137
73. Pan, S, Zou, J, Mao, H, Hu, Z, Sun, S, Wu, W, et al. Available phosphorus levels modulate growth performance, serum indices, metabolome, rumen fermentation, and microorganism in Hu lambs. Anim Feed Sci Technol. (2025) 322:116259. doi: 10.1016/j.anifeedsci.2025.116259
74. Zhang, Y, Zhang, X, Cao, D, Yang, J, Mao, H, Sun, L, et al. Integrated multi-omics reveals the relationship between growth performance, rumen microbes and metabolic status of Hu sheep with different residual feed intakes. Animal Nutrition. (2024) 18:284–95. doi: 10.1016/j.aninu.2024.04.021
75. NkukwanaI, TT, MuchenjeI, V, MasikaII, PJ, and Mushonga, B. Intestinal morphology, digestive organ size and digesta pH of broiler chickens fed diets supplemented with or without Moringa oleifera leaf meal. S Afr J Anim Sci. (2015) 45:362–71. doi: 10.4314/SAJAS.V45I4.2
76. Rao, R, and Samak, G. Role of glutamine in protection of intestinal epithelial tight junctions. J Epithel Biol Pharmacol. (2012) 5:47–54. doi: 10.2174/1875044301205010047
77. Hezaveh, MSS, Ghasemi, HA, Hajkhodadadi, I, and Moradi, MH. Single and combined effects of phytase and citric acid on growth performance, nutrient digestibility, bone characteristics, intestinal morphology, and blood components in meat-type quails fed low-phosphorous diets. Anim Feed Sci Technol. (2020) 269:114677. doi: 10.1016/j.anifeedsci.2020.114677
78. Chang, G, Tian, S, Luo, X, Xiang, Y, Cai, C, Zhu, R, et al. Hypoglycemic effects and mechanisms of polyphenols from Myrica rubra pomace in type 2 diabetes (db/db) mice. Mol Nutr Food Res. (2025) 69:e202400523. doi: 10.1002/mnfr.202400523
79. Wang, Y, Wang, L, Du, Y, Yao, F, Zhao, M, Cai, C, et al. Metabolomics study reveals DON-induced intestinal toxicity in adult zebrafish through disruption of amino acid metabolism and sphingolipid signaling pathway. Aquat Toxicol. (2025) 282:107324. doi: 10.1016/j.aquatox.2025.107324
80. Wang, X, Liu, Y, Xu, Y, Gao, S, Xu, Q, Gong, H, et al. Structural characterization of a pectic polysaccharide from Rubus chingii Hu. Unripe fruits and its efficacy in inhibiting intestinal lipid absorption in vivo. Carbohydr Polym. (2025) 363:123728. doi: 10.1016/j.carbpol.2025.123728
81. Valdivié-Navarro, M, Martínez-Aguilar, Y, Mesa-Fleitas, O, Botello-León, A, Hurtado, CB, and Velázquez-Martí, B. Review of Moringa oleifera as forage meal (leaves plus stems) intended for the feeding of non-ruminant animals. Anim Feed Sci Technol. (2020) 260:1–9. doi: 10.1016/j.anifeedsci.2019.114338
82. Stech, MR, Carneiro, DJ, and Carvalho, MRB. Anti-nutritional factors and protein apparent digestibility coefficients of soybeans sources to pacu (Piaractus mesopotamicus). Acta Scientiarum. (2010) 32:255–62. doi: 10.4025/actascianimsci.v32i3.5819
83. Zhang, GQ, Dong, XF, Wang, ZH, Zhang, Q, Wang, HX, and Tong, JM. Purification, characterization, and cloning of a novel phytase with low pH optimum and strong proteolysis resistance from aspergillus ficuum NTG-23. Bioresour Technol. (2010) 101:4125–31. doi: 10.1016/j.biortech.2010.01.001
84. Falowo, AB, Mukumbo, FE, Idamokoro, EM, Lorenzo, JM, Afolayan, AJ, and Muchenje, V. Multi-functional application of Moringa oleifera lam. In nutrition and animal food products: a review. Food Res Int. (2018) 106:317–34. doi: 10.1016/j.foodres.2017.12.079
85. Chen, F, Wang, Y, Wang, K, Chen, J, Jin, K, Peng, K, et al. Effects of litsea cubeba essential oil on growth performance, blood antioxidation, immune function, apparent digestibility of nutrients, and fecal microflora of pigs. Front Pharmacol. (2023) 14:1166022. doi: 10.3389/fphar.2023.1166022
86. Cook, RH, and Bird, FH. Duodenal villus area and epithelial cell migration in conventional and germ-free chicks. Poult Sci. (1973) 52:2276–80. doi: 10.3382/ps.0522276
87. Aydin, A, Pekel, AY, Issa, G, Demirel, G, and Patterson, PH. Effects of dietary copper, citric acid, and microbial phytase on digesta pH and ileal and carcass microbiota of broiler chickens fed a low available phosphorus diet. J Appl Poult Res. (2010) 19:422–31. doi: 10.3382/japr.2009-00123
Keywords: alternative feed, carbohydrases, intestine, histology, non-starch polysaccharides
Citation: Macambira GM, Rabello CB-V, Oliveira HSdH, Santos MJBd, Ribeiro AG, Lins Júnior OS, Sousa LDNd, Máximo ILC, Carvalho LRRA and Fleitas OM (2025) Effects of xylanase and phytase supplementation in diets containing Moringa oleifera leaf meal on intestinal morphology and the relative size and weight of internal organs of laying hens. Front. Vet. Sci. 12:1615214. doi: 10.3389/fvets.2025.1615214
Edited by:
Arda Yıldırım, Gaziosmanpaşa University, TürkiyeReviewed by:
Baseer Ahmad, Muhammad Nawaz Shareef University of Agriculture, PakistanAndrea Toschi, Vetagro S.p.A., Italy
Mohamed Yusuf, King Salman International University, Egypt
Copyright © 2025 Macambira, Rabello, Oliveira, Santos, Ribeiro, Lins Júnior, Sousa, Máximo, Carvalho and Fleitas. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Lucas Rannier Ribeiro Antonino Carvalho, bHVjYXMuY2FydmFsaG9Aa2kuc2U=
†ORCID: Hélia Sharlane de Holanda Oliveira, orcid.org/0000-0002-4314-4827
Oziel Saturnino Lins Júnior, orcid.org/0000-0002-1203-4318
Igor Luiz Carvalho Máximo, orcid.org/0000-0003-3299-1307
Lucas Rannier Ribeiro Antonino Carvalho, orcid.org/0000-0002-8385-4188