 Sumayya1Hassan A. Rudayni2
Sumayya1Hassan A. Rudayni2 Muhammad Shuaib1
Muhammad Shuaib1 Bichun Li1Hongyan Sun1Obaid Ullah3Abdullah S. Alawam2
Bichun Li1Hongyan Sun1Obaid Ullah3Abdullah S. Alawam2 Ahmed A. Allam2
Ahmed A. Allam2 Ahmed A. Elolimy4*Mohamed E. Abd El-Hack5
Ahmed A. Elolimy4*Mohamed E. Abd El-Hack5- 1Key Laboratory of Animal Breeding, Reproduction and Molecular Design for Jiangsu Province, College of Animal Sciences and Technology, Yangzhou University, Yangzhou, China
- 2Department of Biology, College of Science, Imam Mohammad Ibn Saud Islamic University (IMSIU), Riyadh, Saudi Arabia
- 3Department of Veterinary Clinical Sciences, Faculty of Veterinary and Animal Sciences, University of Poonch, Rawalakot, Pakistan
- 4Department of Integrative Agriculture, College of Agriculture and Veterinary Medicine, United Arab Emirates University, Abu Dhabi, United Arab Emirates
- 5Department of Industrial Pharmacy, College of Pharmaceutical Sciences and Drug Manufacturing, Misr University for Science and Technology (MUST), Giza, Egypt
Research on circular RNA (circRNA) in poultry increasingly explores its role as a potential regulator in various diseases. It has also shown differential expression of circRNAs in infected tissues, signifying their participation in disease pathogenesis and immune response mechanisms. Additionally, it can function as microRNA sponges, blocking the binding of microRNAs to their target mRNAs and modifying the expression of specific genes. circRNAs are being explored as potential biomarkers for early disease detection and monitoring of disease progression. They are highly expressed and stable in various chicken tissues, offering new insights into their role in tumor development. circRNAs are also being researched as a tool for gene editing, where they could potentially be utilized to increase or repress particular genes in therapeutic contexts. The vast quantity of circRNAs with unknown functions in the eukaryotic transcriptome means that, despite recent progress, our knowledge of the crucial role of circRNAs in viral infections and antiviral immune responses is still unclear. circRNA is a viable contender as a biomarker for bacterial infections due to its stability and distinct expression patterns. Numerous viral disorders have been found to exhibit altered circRNA expression profiles, suggesting their potential use in disease diagnostics. This review briefly introduces the biogenesis, characteristics, and functions of circRNAs, focusing on their progress in research on poultry diseases.
1 Introduction
Circular RNA (circRNA) is a non-coding, single-stranded RNA formed by back-splicing of precursor mRNAs (pre-mRNAs), a process where a downstream splice donor site is joined to an upstream splice acceptor site, creating a covalently closed circular structure. Research on circRNA in poultry is increasingly exploring its role as a potential regulator in various avian diseases, particularly in the context of viral infections like Marek’s disease virus (MDV) and avian leukosis virus (ALV), where several studies demonstrate differential expressions of circRNAs in infected tissues, suggesting their involvement in disease pathogenesis and immune response mechanisms. High-throughput RNA sequencing now profiles circRNAs in chicken tissues, identifying novel disease-associated species (1). circRNA structure is more stable and conserved than linear RNA, as circRNA forms a covalently closed loop without a 5′ end cap or a 3′ poly(A) tail (2). circRNAs typically range from 1 to 5 exons, with about 25% retaining introns in their mature form (3). Recent findings suggest the half-life of circRNAs varies between 8 and 50 h, and their peculiar circular shape allows them to be resistant to RNA exonucleases, resulting in stable expression (1). circRNAs are found either in the nucleus, especially multiexon-circles with retained introns, or in the cytoplasm, where most circRNAs primarily reside (4). Due to their covalently closed structure and resistance to exonucleases, circRNAs exhibit tissue-specific expression and are stable, making them promising diagnostic biomarkers. Moreover, circRNAs have been identified as potential treatment goals for various diseases, such as diabetes, heart disease, neurological conditions, chronic inflammation, and cancer. Their enrichment in exosomes (small extracellular vesicles) further supports their potential as diagnostic markers.
circRNAs have been detected in eukaryotes, including fungi, protists, plants, worms, fish, insects, and mammals, exhibiting cell-type- and tissue-specific expression patterns, often conserved across species and developmental stages (5). These molecules act as microRNA sponges, up-regulating target mRNA expression, and are involved in multiple regulatory functions, such as transcriptional and translational control, protein interaction facilitation, and even direct translation into proteins (6). For instance, circFOXO3 interacts with proteins associated with stress and senescence, inhibiting their nuclear translocation and mitigating senescence and stress in cardiac fibroblasts (7).
As biotechnology and molecular medicine have developed, artificial circRNAs have been created as innovative vaccines for both the prevention and therapy of disease (8). Since circRNAs can show proteins correctly in cells and exhibit reduced immunogenicity, synthetic purification circRNAs are appropriate for in vivo applications, like vaccines or therapeutic agents. circRNA vaccines and gene therapies have gained importance, and further research into circRNAs has highlighted their crucial functions in both physiological and pathological mechanisms, particularly in regulating gene expression at multiple levels (9).
In poultry, certain circRNAs, including circRNA_3238 and circRNA_3079, have been implicated in avian leukosis virus (ALV) infection (10). circRNAs are highly expressed and stable in various chicken tissues, offering new insights into their role in tumor development in poultry (11). Qiu et al. (12) explored circRNA expression in both healthy and ALV-J-infected chicken spleen tissues, indicating a role for circRNAs in tumor genesis. Researchers are investigating the specific functions of identified circRNAs by studying their interaction with miRNAs and the downstream effects on gene expression (3).
Given their ubiquitous presence and diversity, circRNAs likely play significant roles in both normal cellular physiology and pathological processes. Further research is required to elucidate the detailed molecular process by which circRNAs control gene expression and contribute to disease pathogenesis. We hypothesize that circRNAs function as key regulatory molecules that modulate disease pathogenesis and immune responses in poultry through multiple mechanisms, including miRNA sponging, protein scaffolding, and transcriptional regulation. This review aims to test this hypothesis by examining the current evidence for circRNA involvement in poultry diseases and their potential as diagnostic biomarkers and therapeutic targets. Therefore, this review briefly introduces the properties, roles, and biogenesis of circRNAs, emphasizing their research progress in poultry diseases. Specifically, Section 2 provides historical context; Sections 3 and 4 describe circRNA biogenesis and characteristics; Section 5 focuses on regulatory mechanisms; Sections 6 and 7 review circRNAs in poultry diseases; and Section 9 concludes with future perspectives.
2 Historical background
Understanding the historical progression of circRNA research provides crucial context for evaluating our hypothesis that circRNAs function as key regulatory molecules in poultry diseases, as the evolution from “splicing artifacts” to recognized regulatory elements mirrors their potential importance in avian pathology. Circular RNAs were first identified in 1976 when Sanger et al. observed RNA circles in viroids using electron microscopy (13). Soon after, Hsu and Coca-Prados detected them in bodily fluids, including the cytoplasm of human HeLa cells, and in 1980 they were also found in the mitochondrial genome of yeast (14). Initially thought to be exclusive to viroids, circRNAs were later detected in the hepatitis delta virus in 1986 (15), and in adenoviruses infecting animal cells. At the time, they were considered rare and biologically insignificant splicing by-products in mammals, although early evidence hinted at their presence in higher organisms (16). In the 1990s and early 2000s, genes producing circRNAs were found across eukaryotes from flies to mammals, but many discoveries were dismissed during the so-called “Artifact Era” due to detection limitations and the challenge of distinguishing them from linear RNAs (17). By the mid-2000s, advances in bioinformatics and high-throughput sequencing allowed reliable identification of circRNAs as distinct RNA species, prompting recognition of their potential regulatory roles in gene expression (18).
In 2012, high-throughput RNA sequencing (RNA-seq) transformed non-coding RNA research and revealed numerous circRNAs in mammalian cells. A landmark study by Salzman et al. (18) showed that circRNAs are abundant, stable, and not splicing errors, but rather a distinct class of transcripts with potential biological functions. This marked a paradigm shift in RNA biology, distinguishing circRNAs from linear mRNAs. Subsequent studies revealed that circRNAs can serve as crucial regulators of cellular processes. Advances in RNA-seq have since confirmed that thousands of circRNA types are expressed across metazoans. Figure 1 summarizes the historical background of circRNAs.
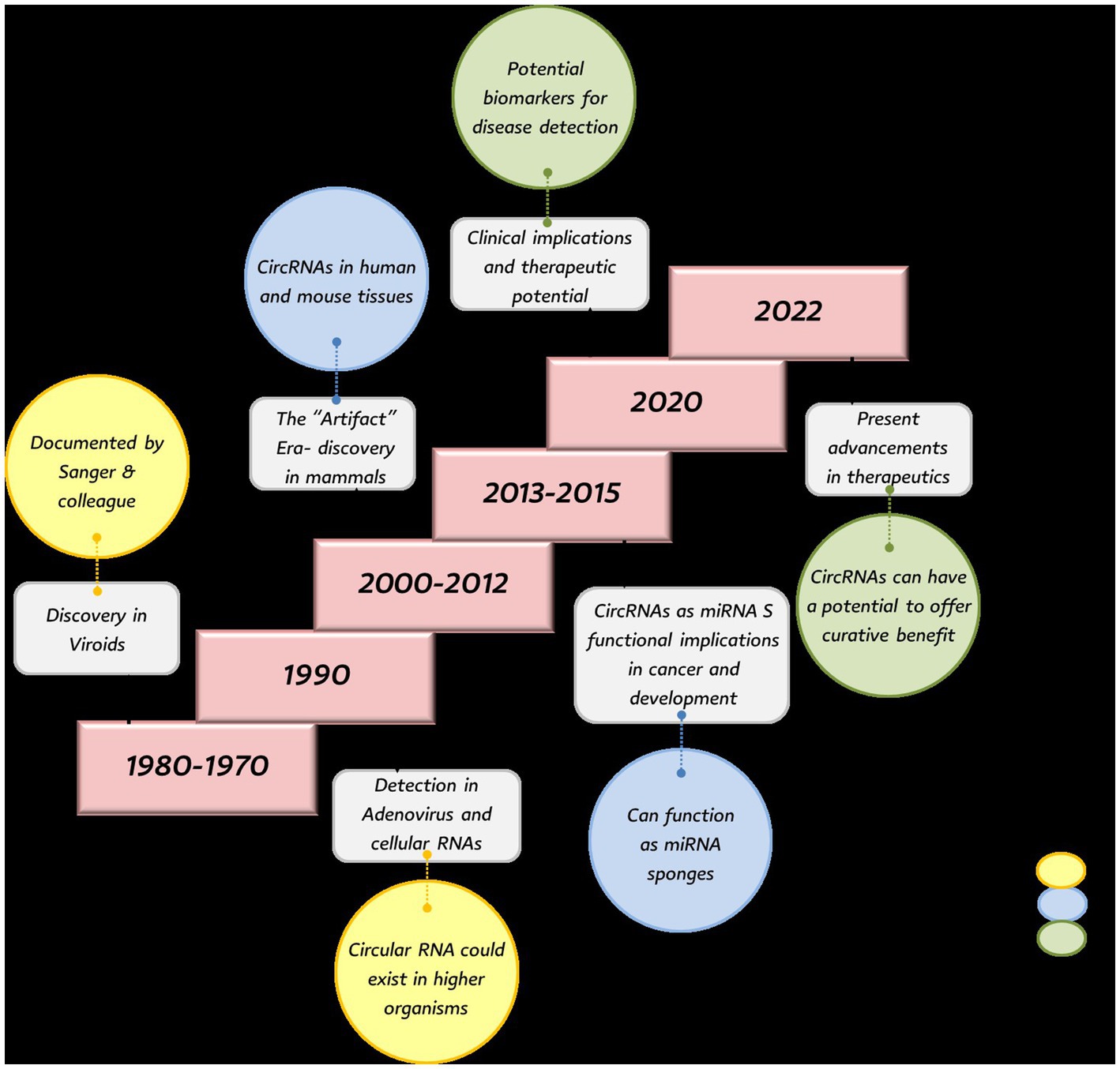
Figure 1. Historical background of circular RNAs.
Following their discovery, circRNAs were recognized for important regulatory functions. Hansen (19) demonstrated that they can act as “miRNA sponges,” sequestering microRNAs (miRNAs) to prevent them from binding target mRNAs, thereby influencing gene expression. Jeck (20) linked circRNAs to cell growth, differentiation, and apoptosis; for instance, CDR1as (ciRS-7) regulates cancer cell proliferation by binding miR-7, a microRNA involved in oncogenic pathways. circRNAs are now associated with cancer, neurological disorders, and cardiovascular diseases, with their stability and detection in blood and saliva making them promising biomarkers (21). In cardiovascular contexts, they modulate key pathways critical for heart function (22). Increasingly, their therapeutic potential is being explored through approaches such as CRISPR-Cas9 and RNA interference to modulate expression for treating cancers, neurological conditions, and other diseases (23). Once considered splicing by-products, circRNAs are now established as multifunctional regulators in diverse cellular processes, including immune responses and pathogen-related pathways. Their stability, tissue-specific patterns, and presence in biofluids position them as strong candidates for diagnostic and therapeutic applications.
Although early circRNA research focused on mammals, poultry studies have expanded rapidly due to the economic impact of infectious diseases and interest in host–pathogen interactions. The first avian-focused work in the late 2010s profiled circRNA expression across tissues, with (24) linking circRNAs to resistance against avian leukosis virus subgroup J (ALV-J)-induced tumors, the first evidence that host circRNAs modulate oncogenic viral outcomes in any species. Subsequent studies implicated circRNAs in responses to Newcastle disease virus (NDV) and Eimeria tenella, connecting them to immune pathways such as NF-κB and B cell receptor signaling (25, 26). Chen et al. (25) uncovered 86 NDV-responsive circRNAs in chicken embryo fibroblasts and showed that over-expression of circ-EZH2 inhibited velogenic NDV replication. Most recently, synthetic circRNAs engineered for low immunogenicity have entered pre-clinical pipelines as next-generation poultry vaccines (8).
In Marek’s disease (MD), circRUNX2.2 promoted lymphoma cell proliferation via RUNX2 activation, indicating an oncogenic role (27), and genome-wide profiling revealed both host- and virus-derived circRNAs. ALV-J studies reported differential expression in resistant birds (12) and a ~30% circRNA loss in infected ones, with circHRH4 showing stability and possible tumorigenic involvement (28, 29). In E. tenella infection, specific circRNAs were linked to adaptive immunity and NK cell activity (26). circRNA changes have also been observed under ammonia exposure and in infectious bursal disease (30).
Beyond disease, circRNAs influence productivity traits. In muscle, circTMTC1, circFGFR2, circHIPK3, and circSVIL regulate myoblast proliferation and differentiation via miRNA interactions (30, 31). In abdominal fat, circRNAs associated with lipid metabolism and mTOR/TGF-β pathways provided the first comprehensive fat-tissue profiles (32, 33). Overall, poultry circRNAs are emerging as key regulators in disease, immunity, and production traits, with diagnostic and therapeutic potential. Collectively, this historical trajectory demonstrates how circRNAs have evolved from overlooked splicing byproducts to recognized regulatory molecules across species, providing strong foundational evidence supporting our hypothesis that they serve as key modulators of disease pathogenesis and immune responses in poultry through multiple regulatory mechanisms.
3 circRNA biogenesis and classification
Elucidating the biogenesis and classification of circRNAs is fundamental to testing our hypothesis, as understanding how these molecules are generated and categorized reveals the mechanistic basis for their regulatory potential in poultry disease contexts. The first phase of circRNA synthesis involves the transcription of eukaryotic precursor mRNAs (pre-mRNAs) by RNA polymerase II, followed by splicing to excise introns and ligate exons, resulting in mature mRNAs (34). circRNA is produced via a unique kind of alternative splicing called head-to-tail splicing or back-splicing, in which an exon’s 3′ end ligates to the 5′ end of either its own or an upstream exon via 3′–5′ phosphodiester bonds. This produces a closed-loop configuration, including a back-splicing junction point. Initially, circRNAs were considered splicing anomalies, including “scrambled exons” (35).
circRNAs are classified into three types: exonic, exon-intron, and circular intronic based on their composition: exonic circRNAs (ecircRNAs) composed only of exons, exon-intron circRNAs (EIciRNAs) including both exons and introns, and circular intronic RNAs (ciRNAs) originating from introns (36). More than 80% of known circRNAs are ecircRNAs, mostly located in the cytoplasm. Conversely, EIciRNAs and ciRNAs are primarily situated in the nucleus, where they are believed to be involved in the control of gene transcription (37). Circular RNAs can modulate gene transcription in the nucleus by interacting directly with the promoter region of their parental genes, recruiting transcription factors or RNA polymerase II, and forming RNA–DNA hybrid structures (R-loops). Depending on the specific circRNA and cellular context, this can either enhance or repress transcription, functioning as a “scaffold” for the transcriptional machinery at the gene locus. The three canonical circularization models (intron-pairing, lariat-driven, RBP-assisted) were established in mammalian cells; orthologous intronic architecture and repeat elements have been reported in chickens (e.g., reverse-complementary matches overlapping CR1 repeats), supporting—but not proving—mechanistic conservation in avian species.
Recent studies have identified three models of biosynthesis of circRNA that reflect the contest between back-splicing and canonical splicing: (i) Intron-pairing-driven circularization, (ii) lariat-driven circularization or exon skipping, and (iii) RNA-binding protein (RBP)-driven circularization (38). In the intron-pairing concept, back-splicing is promoted by base-pairing among different introns, especially involving repetitive sequences such as ALU repeat. In the lariat-driven model, circularization occurs when exons are eliminated for creating linear RNA, allowing splice site joining. In the RBP-driven model, Introns close to splice sites are bound by RBPs, promoting the formation of circular RNAs (39). Translation, transcription or splicing regulation, protein interaction, and RNA interaction represent a few of the possible processes of circRNAs. Circular RNAs can also be released into extracellular space after being enclosed in exosomes, which are produced by vesicle cells.
Circular RNAs arise from spliceosome-mediated back-splicing of pre-mRNAs through three routes: intron pairing (complementary repeats bring flanking splice sites together), lariat-driven back-splicing (exon skipping followed by processing of the intron lariat), and RNA-binding protein–assisted circularization (RBPs bridge flanking introns). The figure also summarizes key functions: regulate transcription/splicing, sponge miRNAs and interact with mRNA/lncRNA (ceRNA), bind proteins as decoys/scaffolds, sort into exosomes for extracellular release, and translate into peptides via cap-independent initiation (IRES/m6A). Together, these biogenesis mechanisms and classification systems illustrate how circRNAs are generated through highly regulated processes that determine their cellular localization and functional capacity, supporting our hypothesis that their diverse origins and structural features enable them to function as versatile regulatory molecules in poultry disease pathogenesis and immune responses.
4 Characteristics of circRNAs
The unique structural and functional characteristics of circRNAs provide the molecular foundation for testing our hypothesis, as their stability, tissue specificity, and conservation across species suggest they are well-positioned to serve as key regulatory molecules in poultry health and disease. Circular RNAs are a unique class of single-stranded, covalently closed RNA molecules found across a wide range of species, including eukaryotes such as animals and plants. They are primarily generated from precursor messenger RNAs (pre-mRNAs) through a non-canonical process called back-splicing, in which a downstream splice donor site is joined to an upstream splice acceptor site. Unlike linear RNAs, circRNAs lack both a 5′ cap and a 3′ poly(A) tail, a structural feature that makes them highly resistant to exonuclease-mediated degradation, such as by RNase R, and results in remarkable stability, often two to five times greater than that of their linear counterparts (40, 41). This closed-loop structure, combined with their widespread occurrence, enhances their potential as biomarkers and therapeutic targets in clinical and veterinary research (42–44). General features such as exon dominance, nuclear versus cytoplasmic partitioning, and RNase R resistance derive primarily from mammalian studies; where chicken-specific distributions or exceptions have been experimentally validated (e.g., tissue-specific expression hotspots, developmental stage specificity in Gushi chickens), they are explicitly noted with avian data references.
In poultry, circRNAs are broadly distributed across all chromosomes, including sex chromosomes, with certain genomic regions acting as hotspots for circularization events. They vary greatly in length, from under 100 nucleotides to nearly 100,000 nucleotides, depending on the tissue and breed. Most identified chicken circRNAs are derived from exonic regions, 84.95% in Gushi chicken adipose tissue and 81.5% in pectoralis muscle, while a smaller proportion originates from introns, intergenic sequences, or untranslated regions (45). The expression of circRNAs is often tissue-specific and developmentally regulated, with some showing distinct patterns such as bimodal expression peaks in Gushi chicken adipose tissue (45, 46). Their expression is generally lower than that of linear mRNAs, yet their stability allows them to accumulate and persist in specific cell types. External factors, such as ammonia exposure, viral infections (including Avian Leukosis Virus subgroup J, Marek’s disease virus, and H5N1 avian influenza virus), bacterial infections (Salmonella enterica serovar Enteritidis), and even insulin administration, can dynamically regulate circRNA expression in chickens (47).
Functionally, circRNAs participate in a diverse range of biological processes. Many act as microRNA (miRNA) sponges, sequestering specific miRNAs and thus modulating post-transcriptional gene regulation (42, 44). Others interact with RNA-binding proteins (RBPs), influence transcription, affect pre-mRNA splicing, and, in some cases, may even serve as templates for protein or peptide translation, particularly when they contain internal ribosome entry site (IRES) elements. While translation has been experimentally confirmed for some mammalian circRNAs, definitive evidence in poultry or plants remains lacking, though the potential is strong. Subcellular localization also correlates with function, exonic circRNAs are predominantly cytoplasmic, whereas intronic and exon–intron circRNAs are often nuclear (48).
Circular RNAs are highly conserved across species, indicating their evolutionary and functional importance. For example, circINSR, derived from the insulin receptor (INSR) gene in chickens, shares approximately 77% sequence similarity with its mammalian counterparts, including humans, mice, and bovines (47). This conservation supports the idea that circRNAs carry out essential, preserved roles in cellular regulation. Their stability, tissue specificity, and involvement in key physiological and pathological processes, including muscle development, fat metabolism, and immune responses, underscore their potential not only as diagnostic and prognostic biomarkers but also as targets for therapeutic interventions (29, 46). Given their multifaceted biological significance, continued in-depth studies are essential to fully understand their functions and translate these findings into practical applications for both human and animal health (43, 49, 50). Collectively, these distinctive characteristics, including exceptional stability, tissue-specific expression, evolutionary conservation, and environmental responsiveness, provide compelling evidence supporting our hypothesis that circRNAs function as key regulatory molecules in poultry, with their unique properties enabling them to effectively modulate disease pathogenesis, immune responses, and serve as reliable diagnostic biomarkers and therapeutic targets.
5 Regulatory mechanism of circRNA
The diverse regulatory mechanisms employed by circRNAs provide direct evidence for testing our central hypothesis, as their ability to function through multiple pathways—including miRNA sponging, protein scaffolding, translation, and transcriptional regulation—demonstrates their capacity to serve as key modulators of disease pathogenesis and immune responses in poultry. Circular RNAs serve multiple functions within the cell. One of their most extensively studied roles is as master regulators of gene expression. They can act as competing endogenous RNAs (ceRNAs), sequestering or “sponging” regulatory molecules, particularly microRNAs (miRNAs) (51). Additionally, some circRNAs have the capacity to be transformed into proteins. Since circRNAs can encode proteins in cells while eliciting minimal immunogenicity, synthetic circRNAs are promising for in vivo applications such as vaccines or therapeutic agents. Mechanistic models summarized below are largely defined in mammalian systems; poultry-specific validations are called out in each subsection. Table 1 summarizes only avian-derived circRNA mechanisms and phenotypes (species/cell type, disease/process, outcomes, targets).
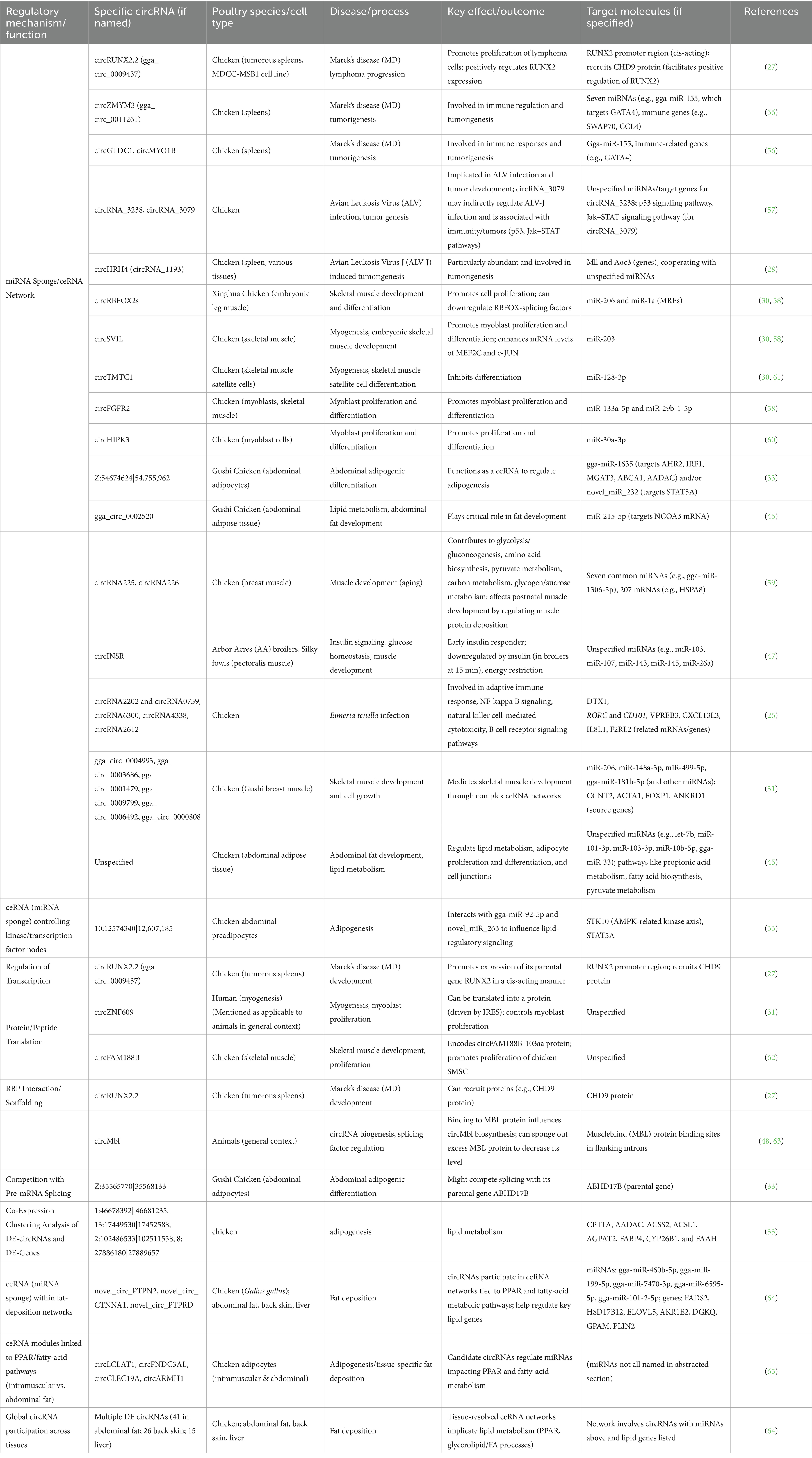
Table 1. Poultry circRNA regulatory mechanisms, targets, and phenotypes (avian experimental evidence).
5.1 circRNAs regulate gene expressions as miRNA sponges
CircRNAs function as “super sponges” that provide quantitative control over miRNA availability, establishing them as master regulators that can amplify or dampen entire gene expression programs through single molecular interactions. This regulatory amplification model is supported by their superior miRNA-binding affinity compared to other endogenous RNAs and their capacity to sequester multiple miRNA copies simultaneously (52). The regulatory power of circRNA sponging is exemplified by CDR1as/CiRS-7, which demonstrates how a single circRNA can reshape entire cellular programs. With 73 binding sites for miR-7, CiRS-7 creates a molecular sink that depletes miR-7 availability throughout the cell, simultaneously derepressing all miR-7 target genes (53). This multiplexed regulation means that CiRS-7 expression changes do not simply adjust individual genes, they coordinately regulate entire miR-7-controlled genetic programs. The regulatory principal scales beyond individual circRNA-miRNA pairs to create hierarchical control networks. The 70 conserved miRNA target sites in circCDR1 illustrate how circRNAs can simultaneously regulate multiple miRNA families, creating regulatory hubs that integrate diverse cellular signals (54). This hub architecture explains why circRNA dysregulation has such profound pathological effects: disrupting a single circRNA node can cascade through multiple miRNA-controlled pathways, amplifying regulatory perturbations across entire cellular networks. In pathological contexts, this amplification transforms circRNAs from simple molecular sponges into disease-driving master regulators. Furthermore, circRNAs have been connected to many pathological processes through their ability to sponge different miRNAs. By competing with mRNAs for miRNA binding, circRNAs can negatively regulate miRNA activity, indirectly influencing mRNA levels. Consequently, circRNAs can alleviate or reduce the inhibition of genes targeted by miRNAs, thereby modulating the expression of those genes. For example, overexpression of circLRP6 has been shown to accelerate the sponging of miR-145 to lead to atherosclerosis (55).
Across poultry systems, circRNAs frequently act as competing endogenous RNAs (ceRNAs) that tune disease and development by sequestering miRNAs and derepressing key targets. In Marek’s disease (MD), circRUNX2.2 (gga_circ_0009437) promotes lymphoma-cell proliferation by enhancing its parental gene in cis and recruiting CHD9 at the RUNX2 promoter, consistent with a ceRNA-coupled transcriptional mechanism (27). Additional MD-linked circRNAs—including circZMYM3, circGTDC1, and circMYO1B—are embedded in immune/tumorigenic ceRNA networks featuring gga-miR-155 and downstream immune genes such as GATA4, SWAP70, and CCL4 (56). In avian leukosis, circRNA_3079 aligns with p53 and JAK/STAT signaling enrichment and likely modulates ALV-J infection via miRNA pathways, while circHRH4 (circRNA_1193) is abundant across tissues during ALV-J tumorigenesis (28, 57). In skeletal muscle, circRBFOX2s, circSVIL, circTMTC1, circFGFR2, and circHIPK3 govern myoblast proliferation/differentiation through ceRNA control of miR-206/miR-1a, miR-203, miR-128-3p, miR-133a-5p/miR-29b-1-5p, and miR-30a-3p, respectively (30, 58–60). Adipogenic ceRNA axes include Z:54674624|54755962 (gga-miR-1635; novel_miR_232) and gga_circ_0002520 (miR-215-5p → NCOA3), with broader networks in breast muscle and abdominal fat engaging miR-206/148a-3p/499-5p/181b-5p and lipid-metabolism pathways (31, 45, 59). Notably, circINSR behaves as an early insulin responder in pectoralis muscle, linking ceRNA regulation to glucose and growth phenotypes (47). In coccidiosis, differentially expressed circRNAs associate with adaptive immunity, B-cell receptor signaling, NK-cell cytotoxicity, and NF-κB modules during Eimeria tenella infection, reinforcing an immune-centric ceRNA layer (26). These poultry-specific examples of circRNA-mediated ceRNA networks clearly demonstrate how circRNAs function as key regulatory molecules in avian systems, supporting our hypothesis through their documented roles in modulating viral pathogenesis, immune responses, muscle development, and metabolic processes via miRNA sequestration mechanisms.
5.2 Circular RNAs as protein decoys
circRNAs can directly engage with circRNA-binding proteins (cRBPs), serving as protein decoys that influence various cellular functions. By binding to specific proteins, circRNAs can control the translocation of these proteins within the cell. For instance, the circRNA circ-Ccnb1 forms a ternary complex with Ccnb1 and Cdk1, which inhibits the activity of Ccnb1, thereby promoting cell proliferation and survival (66). The interaction of circAmotl1 with PDK1 and AKT1 in primary cardiomyocytes results in the phosphorylation and nuclear translocation of AKT1, hence diminishing apoptosis and facilitating cardiac healing. Likewise, circFOXO3 promotes the interaction between MDM2 and p53, leading to reduced p53 protein levels and the consequent initiation of apoptosis (67).
CircACC1 is another example that contributes to metabolic adaptation to serum deprivation by increasing the enzymatic activity of the AMP-activated protein kinase (AMPK) holoenzyme. This transpires via establishing a ternary complex with the AMPK β and γ regulatory subunits (68). However, bioinformatics analyses indicate that circRNAs exhibit decreased RBP binding density compared to linear RNAs, suggesting that few circRNAs may be unable to effectively interact with proteins. Thus, there is a critical requirement for innovative approaches to facilitate more efficient studies on circRNA-protein interactions. Most protein-decoy/scaffold interactions (e.g., circFOXO3–MDM2–p53; circAmotl1–AKT1) are mammalian; direct RBP-binding demonstrations in poultry remain limited and are identified explicitly where available. Evidence for circRNA–RBP architecture in poultry includes circRUNX2.2 recruitment of CHD9 during MD progression (27); more broadly, animal models show that circMbl binds the splicing factor MBL to regulate its own biogenesis and buffer excess MBL—a mechanism likely conserved but still underexplored experimentally in avian tissues (48, 63). While most protein-scaffolding mechanisms derive from mammalian studies, the emerging evidence from poultry systems, particularly circRUNX2.2’s recruitment of CHD9, supports our hypothesis by demonstrating that avian circRNAs can indeed function as key regulatory molecules through direct protein interactions that influence disease progression.
5.3 circRNAs translation into proteins
Most circRNAs are found in the cytosol, suggesting their potential involvement in ribosome assembly for protein translation. Studies have demonstrated that circRNAs containing translation of an internal ribosome entry site (IRES) are available in peptides both in vitro and in vivo (67). While some circRNAs are localized in the nucleus, most are predominantly abundant in the cytoplasm. This cytoplasmic localization allows circRNAs to interact with proteins and microRNAs, thereby regulating gene expression (69). Recent research has shown that the translation of ecircRNAs, manufactured from back-splicing in human cells that have been vectorized, is indeed possible (70).
However, there is currently no evidence to suggest that spliceosome-generated ecircRNAs can function as mRNAs. For instance, circ-ZNF609 contains a 753-nucleotide open reading frame (ORF) that spans from the start codon to an in-frame stop codon. Notably, circ-ZNF609 has the potential to encode two distinct protein isoforms by both splicing-dependent and—independent mechanisms, marking the initial proof that endogenous circRNAs are capable of translation (48). Translationally competent circRNAs are emerging in muscle biology: while circZNF609 (IRES-driven) exemplifies a conserved myogenic paradigm in animals (31), chicken circFAM188B encodes the peptide circFAM188B-103aa that promotes skeletal-muscle satellite-cell proliferation, highlighting an endogenous coding output from avian circRNAs (62). The demonstration that circRNAs can be translated into functional peptides provides additional evidence supporting our hypothesis by showing that circRNAs serve as multifunctional regulatory molecules in poultry through both coding and non-coding mechanisms.
5.4 Transcriptional regulation
Circular RNAs (circRNAs) are highly stable non-coding RNAs produced by back-splicing; beyond acting post-transcriptionally they also participate directly in transcriptional regulation and are themselves subject to complex transcriptional control. Some nuclear circRNAs, such as circular intronic RNAs (ciRNAs) and exon–intron circRNAs (EIciRNAs), interact with RNA polymerase II (Pol II), the enzyme that copies DNA into RNA, to boost the activity of the genes they come from. For example, ciRNAs such as ci-ANKRD52 enhance Pol II transcription at their host loci, while EIciRNAs (e.g., circEIF3J, circPAIP2) bind U1 snRNP and Pol II at promoters to stimulate transcription (35, 71, 72). Nuclear circRNAs can also recruit chromatin remodelers or transcription cofactors to promoters—chicken circRUNX2.2 recruits CHD9 to the RUNX2 promoter and promotes RUNX2 transcription—demonstrating a direct, locus-specific activating mechanism (27, 30). Figure 2 shows the four models of circRNA functions.
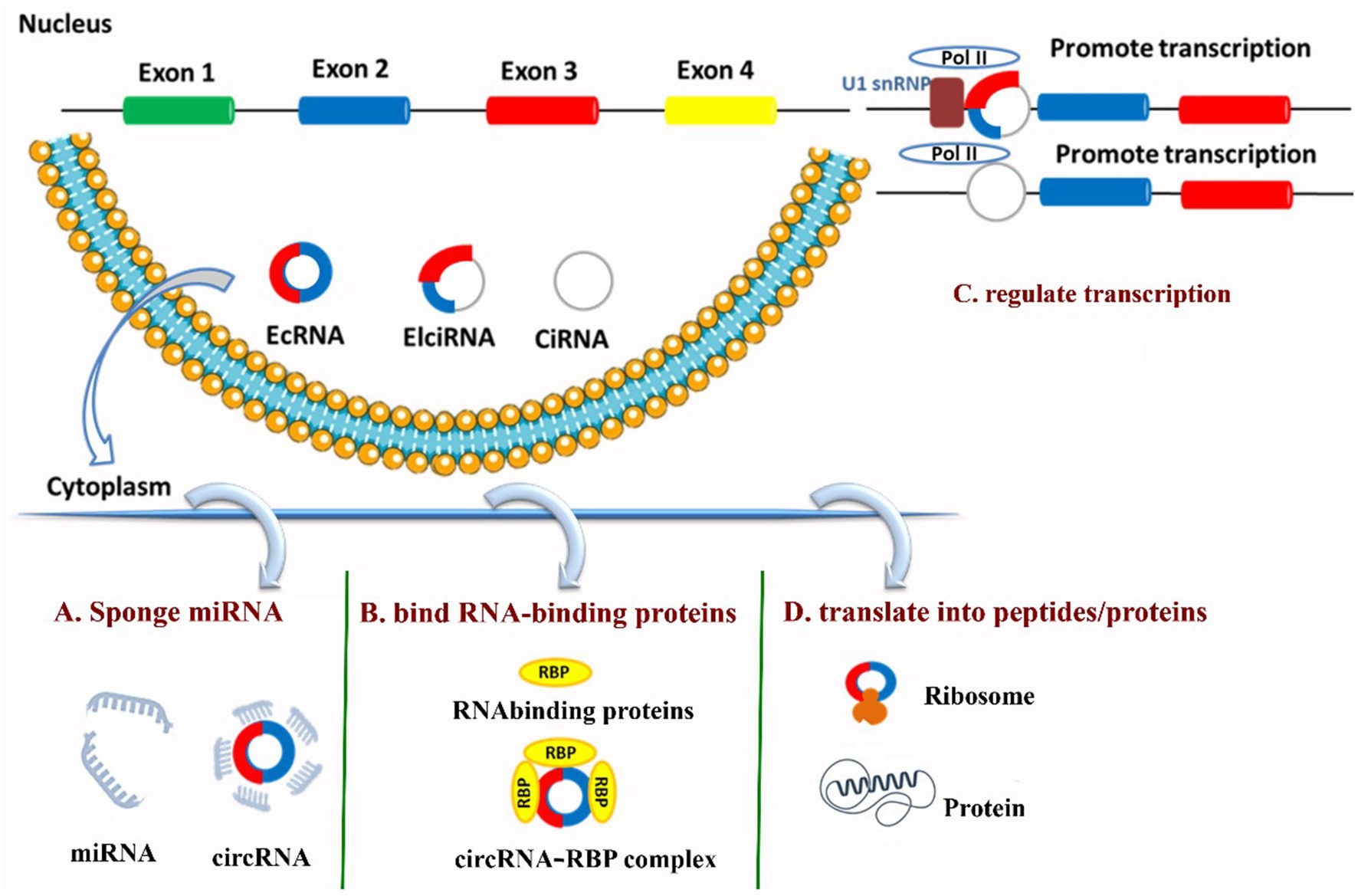
Figure 2. Four canonical circRNA functions [adapted from Zhang et al. (52)]. (A) Sponge miRNA and relieve repression on target mRNAs (ceRNA model). (B) Bind RNA-binding proteins and act as decoy/scaffold to modulate protein localization and activity. (C) Regulate transcription of parental genes, with EIciRNAs/ciRNAs engaging RNA polymerase II and/or U1 snRNP at promoters. (D) Translate into peptides/proteins via cap-independent initiation mediated by IRES elements or m6A.
The circRNAs may also repress transcription through alternative mechanisms: some bind their parent loci to form R-loops (RNA–DNA hybrids) that cause Pol II pausing or termination (e.g., circSMARCA5), or they act as decoys/scaffolds that sequester transcription factors or RNA-binding proteins away from promoters, altering transcription factor binding and chromatin states (71, 73, 74). Epigenetic associations are common: circRNA loci frequently overlap regulatory regions marked by active histone marks (e.g., H3K27ac) and can be integrated into transcription factor networks (75, 76). Importantly, circRNAs can act both in cis (at their host gene) and in trans (on distant genes) by recruiting or sequestering regulatory proteins.
Conversely, circRNA biogenesis (their “transcriptional” control) is tightly regulated. Back-splicing competes with canonical linear splicing and is promoted by intron-pairing (complementary flanking sequences/inverted repeats), RNA-binding proteins (RBPs) that bridge splice sites (e.g., MBL, QKI), and lariat/exon-skipping pathways. In chickens, reverse complementary matches (RCMs), including elements overlapping CR1 repeats, facilitate exon circularization; deleting RCMs reduces circRNA formation (27, 30, 46). Biogenesis can thus reduce linear mRNA output (negative correlation between exonic circRNAs and parental mRNAs in some tissues) or, alternatively, correlate positively (as reported for some intronic circRNAs in chicken adipocytes), depending on context and mechanism. Post-transcriptional modifiers also affect circularization: ADAR1-mediated A → I editing can disrupt intronic base-pairing and inhibit circRNA formation (and has been implicated in AR-linked suppression of circRNAs in HCC), whereas m6A modification can promote production and translation of certain circRNAs with ORFs (48, 62, 77).
In sum, circRNAs both shape and are shaped by transcriptional programs: they modulate Pol II activity, recruit or sequester regulatory proteins, form R-loops, influence chromatin and TF binding, and compete with linear splicing—while their biogenesis is controlled by intronic architecture, RBPs, RNA editing, and epitranscriptomic marks. These two-way interactions make circRNAs integral players in transcriptional regulation and attractive candidates for functional studies and therapeutic targeting in poultry and other species (71, 75, 78, 79).
For poultry studies, beyond post-transcriptional miRNA sponging, avian circRNAs operate across multiple regulatory layers relevant to disease and production traits. A kinase/TF–focused ceRNA circuit is exemplified by 10:12574340|12607185, which interacts with gga-miR-92-5p and novel_miR_263 to influence STK10 (an AMPK-related kinase) and STAT5A during adipogenesis (33). At the transcriptional level, circRUNX2.2 elevates RUNX2 in tumorous spleens by acting in cis at the promoter and recruiting CHD9, illustrating that circRNAs can couple enhancer-like activity with ceRNA functions in Marek’s disease (27). Biogenesis competes with linear splicing, as Z:35565770|35568133 may alter processing of its parental gene ABHD17B in Gushi chicken adipocytes (33). Systems analyses map DE-circRNAs to lipid-metabolism hubs (CPT1A, AADAC, ACSS2, ACSL1, AGPAT2, FABP4, CYP26B1, FAAH), consistent with coordinated control of fatty-acid activation/transport and glycerolipid synthesis (33). Tissue-resolved ceRNA modules, novel_circ_PTPN2, novel_circ_CTNNA1, novel_circ_PTPRD, intersect with PPAR and fatty-acid pathways via specific miRNAs to regulate lipid genes across abdominal fat, back skin, and liver of Gallus gallus (64), while depot-specific candidates (circLCLAT1, circFNDC3AL, circCLEC19A, circARMH1) help partition intramuscular versus abdominal fat programs (65). Consistently, cross-tissue remodeling (41, 26, and 15 DE circRNAs in abdominal fat, back skin, and liver, respectively) converges on PPAR, glycerolipid, and fatty-acid metabolism, aligning circRNA regulation with carcass fatness phenotypes (64).
Collectively, these four regulatory mechanisms, miRNA sponging, protein scaffolding, translation, and transcriptional regulation, provide comprehensive evidence supporting our central hypothesis. The documented examples from poultry systems demonstrate that circRNAs indeed function as multifaceted key regulatory molecules that modulate disease pathogenesis and immune responses through diverse, interconnected mechanisms, establishing them as promising targets for diagnostic and therapeutic applications in avian health.
6 Role of circular RNA in diseases
The advent of high-throughput RNA sequencing and refined bioinformatics tools has revealed their widespread expression across viruses, plants, archaea, and animals, along with their pivotal roles in regulating cellular homeostasis and disease processes. Recent studies have confirmed the presence of circRNAs in both DNA and RNA viruses, where they play crucial roles such as evading the host’s immune response, contributing to disease pathogenesis, enhancing protein translation, serving as miRNA sponges, and regulating viral replication and cellular division. Endometrial fibrosis, endometriosis-related infertility, and pre-eclampsia (PE) are among the most common gynecological disorders worldwide, all of which have been linked to circRNAs (80). Most exemplars derive from mammalian research provides essential context for circRNA biology. These mammalian insights set the stage for understanding disease associations, while poultry-specific data and mechanisms are addressed separately in Section 7.
Recent data suggests that many circRNAs may interact with miRNAs, therefore participating in several physiological and pathological processes. For example, hsa_circ_0009361 modulates the expression of adenomatous polyposis coli 2 by interacting with miR-582, thus impeding the advancement of colorectal cancer (81). Microvascular complications are prevalent in diabetic retinopathy (DR) and may lead to blindness. Recent studies have correlated three RNA molecules—miRNAs, long noncoding RNAs (lncRNAs), and circRNAs—with the occurrence and development of diabetic retinopathy (DR), indicating their potential role as upstream regulators or participants in the functional processes related to this disease (82). Moreover, research indicates that circRNAs produced by viruses or differently expressed host circRNAs may function as potential biomarkers for viral infections (11). Along with their roles as supreme controllers of gene appearance, circRNAs present intriguing potential as novel biomarkers for cancer and other diseases (83).
Beyond these examples, the dysregulation of circRNAs is now recognized as a hallmark in a broad spectrum of human diseases, where they may act as either pathogenic drivers or protective modulators.
• Inflammatory Bowel Disease (IBD): Aberrant circRNA expression has been documented in ulcerative colitis and Crohn’s disease, affecting intestinal epithelial integrity, immune homeostasis, and fibrotic remodeling. For instance, circ_103516 correlates positively with TNF-α and IFN-γ levels while suppressing IL-10, thereby amplifying inflammation through miR-19b-1-5p sequestration. Other circRNAs such as circSMAD4 activate JAK2/STAT3 signaling, whereas circPABPN1 impairs autophagy by disrupting HuR–ATG16L1 interactions. Certain circRNAs also serve as non-invasive biomarkers capable of distinguishing Crohn’s disease from ulcerative colitis with high diagnostic accuracy (84).
• Neurological Disorders: The mammalian brain exhibits high circRNA abundance, with dynamic expression during development, aging, and neurodegeneration (85). For instance, circRNA dysregulation is strongly linked to Alzheimer’s disease (AD), with altered expression profiles correlating with cognitive decline. Key examples include: circAPP, which influences microglial polarization via the miR-1906/CLIC1 axis; CDR1as, which sponges miR-7, leading to α-synuclein buildup and impaired protein degradation; circHOMER1, downregulated in AD and potentially regulating PSEN1/PSEN2; circHDAC9, neuroprotective through miR-142-5p sponging; circLPAR1, promoting Aβ-induced apoptosis and inflammation; circNF1-419, enhancing autophagy via PI3K/Akt/AMPK/mTOR signaling; circAβ-a, generating Aβ polypeptides; circCORO1C, reducing miR-105 and increasing APP; circPSEN1, affecting TGF-β1/Notch pathways via miR-4668-5p and miR-5584-5p; and circRIMS2, whose m6A modification contributes to synaptic and memory deficits (63, 84, 86, 87).
• In Parkinson’s disease, circSLC8A1 is linked to oxidative stress via miR-128/miR-132; circ_0004381 and circ_0017204, early PD biomarkers; circ_0085869, circ_0004381, and circ_0090668, distinguishing disease stages; circEps15, which promotes dopaminergic recovery via miR-24/PINK1-PRKN mitophagy; circSV2b, restoring dopamine synthesis via miR-5107-5p/Foxk1/Akt1; and circPANK1 and circSNCA, which boost α-synuclein by sponging miR-7, driving neurotoxic aggregation. Comparable mechanistic roles are reported in amyotrophic lateral sclerosis, Huntington’s disease, schizophrenia, bipolar disorder, multiple sclerosis, and epilepsy (63, 84, 85).
• Cancer: circRNAs can function as oncogenes or tumor suppressors. In prostate cancer, circSMARCC1 promotes tumor-associated macrophage infiltration and M2 polarization, while in glioblastoma, circMTO1 and circHIPK3 facilitate tumor progression through miRNA sponging and pathway modulation. In colorectal cancer, circCCDC66 and circITGA7 regulate metastatic behavior, whereas in hepatocellular carcinoma, circATP5H enhances tumor growth via TNFAIP3 regulation through miR-138-5p (85, 88, 89).
• Autoimmune Diseases: Elevated CiRS-7 in rheumatoid arthritis disrupts mTOR signaling by sequestering miR-7. Multiple sclerosis patients display distinctive leukocyte circRNA profiles, highlighting potential biomarker applications. Similar immunoregulatory roles are observed in systemic lupus erythematosus and other immune-mediated disorders (63, 85, 86).
• Cardiovascular Diseases: circRNAs influence cardiomyocyte fate by modulating apoptosis, autophagy, pyroptosis, necroptosis, and ferroptosis through pathways such as NF-κB, PI3K/AKT, and TGF-β1. Examples include circANRIL in atherosclerosis and circDICAR in myocardial ischemia/reperfusion injury (84, 86).
• Pathogenic Infections: Both host- and pathogen-derived circRNAs participate in antiviral or proviral mechanisms. In viral infections, hsa_circRNA_001387 and EBV-derived circRPMS1 serve as nasopharyngeal carcinoma markers, while KSHV produces circRNAs from its PAN RNA region and HPV generates circE7, encoding the E7 oncoprotein linked to high-risk cancers. Coronaviruses, including SARS-CoV-2, produce circRNAs such as circ_3205, which promotes infection by sponging miR-298 and upregulating PRKCE and KCNMB4. Due to their stability, long translation times, and low immunogenicity, circRNAs are promising candidates for durable viral and cancer vaccines (63, 84). In bacterial infections, S. aureus alters circRNA profiles in milk-derived extracellular vesicles, and reduced circ_0009128 and circ_0005836 in PBMCs may indicate tuberculosis (63).
• circRNAs hold strong potential as diagnostic and therapeutic tools due to their stability, disease- and tissue-specific expression, and presence in diverse biofluids such as blood, saliva, urine, cerebrospinal fluid, and exosomes. They can aid in diagnosis, prognosis, and treatment monitoring. Therapeutic strategies under investigation include RNA interference targeting back-splice junctions, viral vector-mediated overexpression, nanocarrier or exosome-based delivery, and synthetic circRNAs or aptamers as direct therapeutic agents (63, 85, 88, 89).
This mechanistic framework, immune hijacking and metabolic reprogramming, provides the conceptual foundation for interpreting circRNA functions in poultry diseases, where similar regulatory principles likely operate through species-specific molecular networks.
7 Research progress of circular RNA in poultry
This section presents direct experimental evidence from poultry systems or avian cell/tissue datasets that test our central hypothesis, and developmental processes provide species-specific validation of their function as key regulatory molecules in avian disease pathogenesis and immune responses. Recent investigations have begun to illuminate the specific circRNAs associated with poultry diseases such as infectious bronchitis, avian influenza, and coccidiosis. Analyzing the expression patterns, functional roles, and diagnostic potential of circRNAs in these contexts represents a crucial step toward harnessing their therapeutic and diagnostic capabilities in poultry health. circRNAs have a vital function in various biological processes and the progression of disorders. For instance, the investigation of circRNA_3079 in chickens reveals that it is a stable circular transcript predominantly located in the cytoplasm (90). This circRNA is broadly expressed across different tissues, particularly in the lung, spleen, lymph nodes, and bursa of Fabricius. circRNA_3079 may indirectly regulate the process of avian leukosis virus subgroup J (ALV-J) infection. Bioinformatics analyses have indicated that circRNA_3079 and its predicted target genes are enriched in several mechanisms that are associated with immunity and tumors, such as the p53 signaling pathway and the Jak–STAT signaling pathway (57). Additionally, circRNAs have been shown to act as miRNA sponges that regulate chicken muscle development. For instance, Chen et al. verified that circFGFR2 promotes myoblast differentiation and proliferation by interacting with miR-133a-5p and miR-29b-1-5p (30). Thus, circRNAs significantly impact various tissues and organs in poultry, playing an essential role in health and disease. Figure 3 provides a system-level snapshot of how poultry circRNAs wire ceRNA networks into core signaling pathways to shape outcomes across viral, parasitic, and non-infectious conditions.
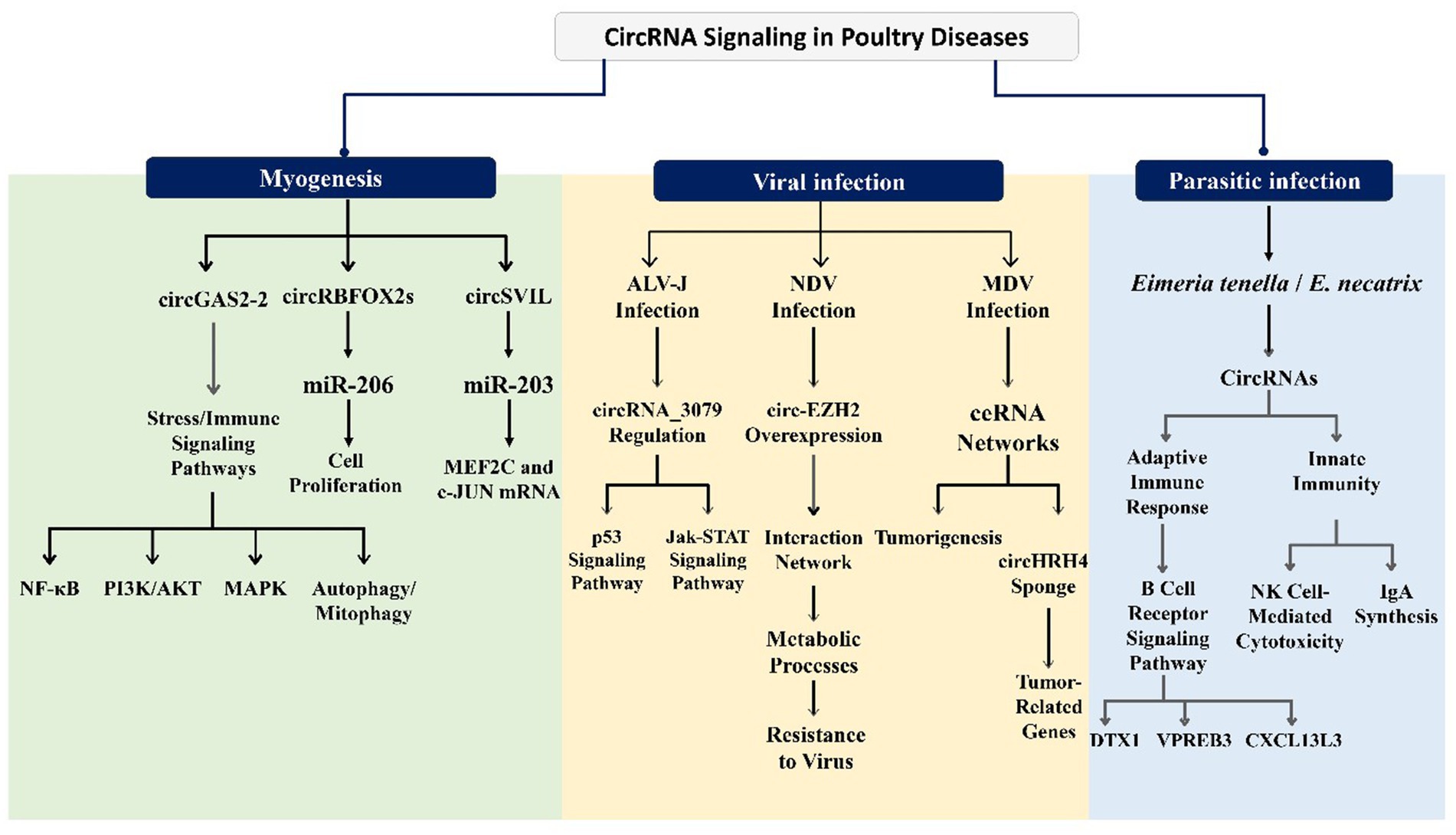
Figure 3. Mechanistic map of circRNA-centered signaling across major poultry diseases.
This figure highlights circRNA functions in myogenesis, viral infections, and parasitic infections. In muscle development, circRNAs such as circGAS2-2, circRBFOX2s, and circSVIL regulate cell proliferation, immune/stress pathways (NF-κB, PI3K/AKT, MAPK, autophagy), and transcription factors (MEF2C, c-JUN). During viral infections, circRNA_3079 modulates the p53 and Jak–STAT pathways in ALV-J, circ-EZH2 enhances resistance to NDV via metabolic regulation, and a circHRH4 ceRNA network drives tumorigenesis in MDV. In parasitic infections (Eimeria tenella and E. necatrix), circRNAs influence adaptive immunity (B cell receptor signaling via DTX1, VPREB3) and innate immunity (NK cell-mediated cytotoxicity, IgA synthesis, CXCL13L3).
7.1 Viral infection
7.1.1 Avian leukosis virus subgroup J (ALV-J)
The oncogenic exogenous retrovirus known as Avian Leukosis Virus Subgroup J (ALV-J) leads to many tumors, slowed growth, high mortality rates, and decreased immunity, and it has been related to poor egg-laying abilities. ALV-J infection fundamentally rewires the host circRNA landscape, creating a molecular environment that promotes tumorigenesis through coordinated disruption of immune surveillance mechanisms. The evidence for this regulatory hijacking comes from multiple converging studies. Zhang et al. used high-throughput transcriptome sequencing of ALV-J–infected chicken macrophages and fibroblasts to identify 7,684 differentially expressed circRNAs. RT-qPCR validation confirmed their association with immune regulation and oncogenic signaling, suggesting roles in ALV-J pathogenesis (28). Critically, these changes were not random but showed systematic enrichment in immune regulatory and oncogenic signaling pathways, suggesting that circRNA perturbation is a central mechanism by which ALV-J establishes and maintains tumorigenic conditions.
The magnitude of this circRNA response indicates that viral oncogenesis depends on comprehensive reprogramming of the host regulatory network rather than simple immune evasion. Supporting this concept, circRNA_3079 and circRNA_3238 emerge as key regulatory nodes during ALV-J infection, with bioinformatics analyses revealing their integration into p53 and JAK/STAT signaling cascades, pathways fundamental to both immune defense and tumor suppression (57). This coordinated targeting of multiple tumor suppressor pathways through circRNA modulation provides a mechanistic explanation for ALV-J’s potent oncogenic effects and suggests that circRNA-mediated regulation represents a vulnerability that could be therapeutically exploited.
As one of the major non-coding RNAs, circRNAs have essential functions in different biological processes, consisting of skeletal muscle development (91). They uniquely regulate both muscle growth in humans and animals and associated physiological and pathological aspects (92). However, research on circRNAs regulating skeletal muscle development in chickens remains limited, particularly during embryonic stages. The 14-day embryo represents a critical period for myoblast proliferation and differentiation into myotubes, which ultimately fuse into muscle fibers by 20 days of age. Wu et al. (91) collected leg muscles from 14 to 20-day-old Bian chicken embryos for RNA-seq, aiming to recognize key circRNAs involved in expansion of skeletal muscle.
These findings from ALV-J studies provide direct evidence supporting our hypothesis, demonstrating that circRNAs function as key regulatory molecules in poultry viral pathogenesis through their differential expression patterns, association with immune and oncogenic pathways, and potential roles in disease resistance mechanisms.
7.1.2 Newcastle disease virus (NDV)
Newcastle disease virus (NDV) is avian paramyxovirus serotype-1 (APMV-1), which was first isolated from pigeons in the Middle East in 1978 (93). NDV infection demonstrates that host circRNAs can function as molecular switches that determine viral replication outcomes through metabolic reprogramming. This regulatory model is supported by compelling functional evidence from Chen et al., who identified 86 differentially expressed circRNAs during NDV infection, with pathway enrichment analyses revealing systematic targeting of host metabolic processes (25). The key insight emerges from circ-EZH2, which exemplifies how individual circRNAs can serve as master regulators of antiviral resistance. Functional validation proves that circ-EZH2 acts as a critical determinant of infection outcome: its overexpression significantly inhibited velogenic NDV replication, while knockdown promoted viral propagation (25). This bidirectional effect establishes circ-EZH2 as a host restriction factor that controls viral replication through downstream metabolic regulation. The mechanistic significance extends beyond individual circRNAs—the coordinated expression changes in 86 circRNAs suggest that NDV resistance depends on comprehensive metabolic reprogramming mediated by multiple circRNA-mRNA-miRNA regulatory networks. This finding reframes NDV pathogenesis from simple viral cytotoxicity to a battle for control over host cellular metabolism, with circRNAs serving as the molecular battlefield where infection outcomes are determined. The NDV infection studies strongly support our hypothesis by showing that circRNAs, particularly circ-EZH2, function as key regulatory molecules in poultry antiviral responses through their ability to modulate metabolic processes and directly inhibit viral replication, demonstrating their therapeutic potential.
7.1.3 Marek’s disease virus (MDV)
The Marek’s disease virus (MDV), which causes Marek’s disease, poses a significant risk to the poultry sector as it is a lymphotropic neoplastic disease. RNA sequencing of tumorous spleens (TS) was completed in order to investigate the expression profiles of host circRNAs and microRNAs (miRNAs). Two spleen types were analyzed: spleens from survivors (SS) without lesions after MDV infection and non-infected chicken spleens (NS). This study identified a total of 2,169 circRNAs, with more than 80% derived from exons. Among these, 113 circRNAs were classified as differentially expressed circRNAs (DECs) (56). There have been more and more circRNAs discovered in recent years in hepatocellular carcinoma (HCC), assisting in the development of tumors and their spread, and showing possible use as biomarkers by their roles as competitive endogenous RNAs (ceRNAs), acting as miRNA sponges, possibly by interactions with proteins that bind RNA (RBPs). Chasseur conducted full-length sequencing and viral circRNA expression analysis in MDV infections and discovered that circRNAs from the same essential virulence genes were highly expressed in vivo (94).
Viral circRNA sequences described from in vitro and in vivo infections showed many noncanonical junctions, which is surprising because the detected back-splice junctions do not follow a unique canonical pattern consistent with the U2-dependent splicing machinery. Qiu conducted a study on when the avian leukosis virus, subgroup J, causes cancer in hens, and competing endogenous RNA networks are shown by circular RNA and mRNA profiling. Transcript structure analysis revealed that chicken circRNAs had similar GC content and comparatively shorter transcripts than mRNAs and lncRNAs. Differential expression analysis identified 152 circRNAs, including 106 up-regulated and 46 down-regulated circRNAs (12). Through the comparison of differentially expressed circRNA host genes and mRNAs, alongside ceRNA network analysis, many tumor or immune-related genes were found, including four genes (Dock4, Fmr1, Zfhx3, Ralb) and two genes (Mll, Aoc3) associated with the ceRNA network. They also identified a circRNA generated from the HRH4 gene inside the ceRNA network, termed circHRH4 (95–98). These MDV investigations provide compelling evidence for our hypothesis, revealing that circRNAs serve as key regulatory molecules in poultry oncogenic viral diseases through multiple mechanisms including transcriptional regulation (circRUNX2.2), ceRNA networks (circHRH4), and both host and viral circRNA interactions that modulate tumorigenesis.
7.2 Parasitic and bacterial infections (coccidiosis; Eimeria tenella/E. necatrix)
circRNAs are long non-coding RNAs that perform vital functions in inflammatory reactions and a number of infectious illnesses. These circRNAs are not merely bystanders but are actively involved in shaping the host’s immune response to the parasite. The E. tenella parasitizes mostly resulting in bloody stools and bleeding of the cecum epithelium in chickens (99). High-throughput sequencing was used to detect circRNAs in the cecal tissues of chickens from the susceptible (JS), resistant (JR), and control (JC) groups on day 4.5 post-infection. This research found 104 differentially expressed circRNAs associated with pathways relevant to Eimeria tenella infection (26). In poultry, Chen et al. (100) found that chicken ammonia poisoning elevated the expression of five differentially expressed circRNAs. Liu et al. (9) discovered that 27 differently expressed circRNAs contributed to the immune response to chronic bursal infection in chickens. Fan et al. (101) performed transcriptome sequencing on the small intestine tissues of chickens infected with E. necatrix and identified 13 differentially expressed circRNAs, including circRNA2673, circRNA3106, and circRNA1579, which were essential to the E. necatrix infection process. During E. tenella infection, circRNAs function as molecular selectors that shape resistance or susceptibility by directing innate versus adaptive immune pathways. Resistant birds showed circRNA enrichment in NF-κB and NK cell cytotoxicity pathways, while susceptible birds displayed adaptive pathway bias. Key circRNA–gene pairs (e.g., circRNA2202/DTX1, circRNA4338/VPREB3, CXCL13L3) illustrate their role in fine-tuning immune cell fate and determining infection outcomes (26). The functional enrichment of these differentially expressed circRNAs was significantly prevalent in the adaptive immune response and B cell receptor signaling pathways (101).
Adaptive immunity regulates antigen-specific immune responses to prevent the pathogen from colonizing and proliferating inside the host. It is an essential element of anti-coccidial protective immunity (102). The B cells are essential components of adaptive immune responses in avians (103). B cells with identical surface immunoglobulin may persist in division and proliferation to enhance host immunity after coccidia infection, during which they produce antigen-specific surface immunoglobulin molecules that bind to the antigen (104). Differentially expressed circRNAs were considerably enriched in the natural killer cell-mediated cytotoxicity pathway, associated with the innate immunity to coccidian infection and involved in the defense against coccidia’s invasion of the gut mucosa (105).
Therefore, in susceptible chickens, DE circRNAs were associated with the adaptive immune response. In contrast, in resistant birds, the enriched pathways included NF-kappa B signaling and natural killer (NK) cell-mediated cytotoxicity such as IgA synthesis during coccidia infection. Specific circRNAs, such as circRNA2202 and circRNA0759, were found to be associated with the DTX1 gene, while circRNA4338 was linked to VPREB3 and CXCL13L3, all of which are involved in immune regulation (26). This highlights how circRNA-mediated ceRNA networks can differentiate between resistant and susceptible host responses, providing valuable insights into the molecular mechanisms of disease resistance in poultry. These results indicated that these circRNAs have a role in coccidian infection by affecting chickens’ innate and adaptive immune responses. Together, these coccidiosis studies provide strong evidence supporting our hypothesis by demonstrating that circRNAs function as key regulatory molecules in poultry parasitic diseases through their modulation of both adaptive and innate immune responses, with specific circRNAs serving as molecular switches that differentiate between resistant and susceptible host responses.
7.3 Muscle development processes (myogenesis)
Myogenesis in poultry operates through temporally coordinated waves of circRNA expression that synchronize metabolic reprogramming with cell fate transitions, ensuring that muscle fiber formation proceeds in precise developmental sequence. Studies have identified a plethora of circRNAs that are dynamically expressed during chicken embryonic and post-hatching muscle development. These myogenesis-associated circRNAs form intricate ceRNA networks that regulate genes involved in muscle metabolism, including glycolysis/gluconeogenesis, amino acid biosynthesis, and energy production (59). For example, a circRNA-miRNA-mRNA network was constructed involving 68 circRNAs, 361 miRNAs, and 599 mRNAs, all showing a tendency to be upregulated with age, consistent with the rapid muscle growth observed after hatching (59). These circRNAs are not only involved in the intrinsic regulation of muscle development but also exhibit crosstalk with stress and immune signaling pathways. This suggests that the physiological state of the bird, including stress levels, can influence muscle growth through circRNA-mediated mechanisms, integrating immune, metabolic, and developmental processes (106). This integration ensures that muscle growth proceeds only when metabolic resources are sufficient and immune challenges are minimal, optimizing developmental efficiency in response to environmental conditions.
To investigate circRNA expression during duck muscle development, Liu et al. (107) collected pectoral muscle samples from Shan Ma ducks at two critical embryonic time points: day 13 (E13), when myoblasts remain undifferentiated, and day 19 (E19), when myoblasts have differentiated. RNA sequencing identified 16,622 circRNAs, of which nearly 80% were exonic circular RNAs; importantly, 260 exhibited differential expression between E19 and E13. The functional evidence demonstrates that specific circRNAs serve as molecular gatekeepers controlling developmental transitions. CircGAS2-2 exemplifies this gatekeeper function: its overexpression accelerated cell cycle progression and enhanced myoblast proliferative capacity, while knockdown blocked cell cycle advancement and reduced proliferative viability (107). This bidirectional control establishes circGAS2-2 as a master regulator that determines when myoblasts transition from proliferation to differentiation phases. The total quantity of muscle fibers is the critical determinant of muscle mass, controlled throughout embryogenesis or early post-hatch in chickens (108). Likewise, the circRBFOX2s was confirmed to promote cell proliferation via their association with miR-206 (109).
RNA sequencing was conducted on the leg muscles of female Xinghua (XH) chickens at three developmental stages: 11 embryonic days (E11), 16 embryonic days (E16), and 1-day post-hatch (P1) to identify circRNAs involved in chicken embryonic skeletal muscle development. A total of 13,377 circRNAs were discovered, with 3,036 exhibiting high expression levels, mainly originating from coding exons. Similarly, 462 circRNAs exhibited differential expression (fold change > 2; p-value < 0.05), with their parental genes associated with biological activities in muscle tissue. Among them, 946 exonic circRNAs were identified to possess one or more miRNA-binding sites for 150 known miRNAs. Ouyang examined the impact of circSVIL on skeletal muscle development and identified the levels of circSVIL expression in the leg muscle of embryos from E10 to P1 (110). The expression level of circSVIL was elevated during the late embryonic development of skeletal muscle. The dual-luciferase test, RNA immunoprecipitation, and biotin-coupled miRNA pull-down revealed that chicken circSVIL acts as a sponge for miR-203, therefore enhancing the mRNA levels of MEF2C and c-JUN (111). These mechanistic studies collectively demonstrate that circRNAs function as critical regulatory molecules during myogenesis through their ability to modulate miRNA availability and subsequently influence target gene expression. The consistent observation of circRNA-miRNA-mRNA regulatory networks across different developmental stages and muscle types provides strong evidence supporting our hypothesis that circRNAs serve as key mediators of normal physiological processes in poultry, extending their regulatory influence beyond disease contexts to fundamental developmental biology, establishing them as promising diagnostic biomarkers and therapeutic targets.
8 Challenges in circRNA research and therapeutic translation
Circular RNAs (circRNAs) have emerged as an exciting frontier in RNA biology, expanding beyond the earlier discoveries of microRNAs to reveal critical roles in gene regulation and disease processes (6, 112, 113). Their covalently closed-loop structure grants high stability and resistance to exonucleases, while their diverse and often tissue-specific expression patterns make them promising candidates for diagnostic and therapeutic applications. In poultry, circRNAs have been linked to immune defense, metabolism, growth, and disease susceptibility, offering opportunities for novel interventions. However, their translation from bench to field faces multiple interconnected scientific and technical barriers that must be addressed systematically before circRNA-based strategies can be reliably implemented in agricultural or clinical settings.
A central research challenge is accurate identification and quantification of circRNAs in avian species. The closed-loop structure complicates detection using conventional RNA sequencing pipelines, which are optimized for polyadenylated transcripts (17, 114). Even gold-standard enrichment workflows, combining rRNA depletion, RNase R digestion, and divergent primer-based RT-qPCR, introduce biases that can skew abundance estimates. Bioinformatics tools such as CIRI-quant and Sailfish-cir improve detection of back-splice junctions but remain prone to errors in low-abundance or alternatively spliced transcripts, especially in complex poultry tissues (115). The lack of standardized protocols, spike-in controls, and universally accepted nomenclature further hampers reproducibility and biomarker validation.
Beyond detection, functional elucidation of circRNAs remains limited. Although high-throughput sequencing has revealed thousands of circRNAs across tissues such as skeletal muscle, liver, spleen, and granulosa cells (7), few have undergone rigorous functional validation in poultry. For example, CRISPR-mediated knockout of circHIPK3 in broilers reduced abdominal fat but impaired antiviral antibody responses, illustrating the delicate balance between production traits and immune fitness (116, 117). Moreover, environmental stressors such as diet composition and heat stress dynamically alter circRNA expression, effects that are difficult to replicate in vitro due to the scarcity of physiologically relevant avian cell models.
One of the key obstacles in targeted circRNA modulation is achieving tissue- and cell-type specificity. While some circRNAs exhibit highly restricted expression patterns, others are broadly distributed, making them less suitable as selective biomarkers or therapeutic targets (118). Current molecular strategies, such as RNA interference, antisense oligonucleotides (ASOs), and expression plasmids, often lack the precision required to prevent off-target effects, and nanoparticle-based delivery platforms remain in developmental stages (119). Effective delivery remains a central challenge: viral vectors provide stable transduction but are constrained by packaging limits and potential immunogenicity, whereas non-viral systems like lipid nanoparticles face low encapsulation efficiency, rapid clearance, and inefficient endosomal escape (6, 120). In complex organisms such as poultry, delivering therapeutic agents, including small molecules or ASOs, without provoking immune reactions or causing tissue damage is particularly difficult. These limitations hinder treatment efficacy, especially for systemic diseases like viral infections or metabolic disorders. Continued progress in engineering viral vectors and next-generation nanoparticle systems with enhanced precision and tissue selectivity may help overcome these barriers (121).
Another emerging barrier is synthetic circRNA immunogenicity (122). While engineered circRNAs offer potential for therapeutics, vaccines, and protein expression due to their stability, they can still activate innate immune receptors such as RIG-I, MDA5, and PKR if they contain immunostimulatory motifs or contaminants (120). The absence of the m6A alteration was proposed to differentiate foreign circRNAs from endogenous circRNAs (123). Current research is looking for ways to lessen the immunogenicity of synthetic circRNA, such as wrapping them in RBPs and applying chemical changes. Strategies to mitigate immune activation—including chemical modifications and immunologically inert delivery vehicles—are promising but not yet foolproof, especially in large-animal models.
Finally, biogenesis-related complications such as mis-splicing remain a persistent issue. Overexpression systems relying on inverted repeats or intronic complementary sequences can generate unintended linear or aberrantly spliced products, interfering with functional readouts and potentially sequestering regulatory molecules (6, 112). While long-read sequencing can improve structural characterization, it remains labor-intensive and is not yet compatible with high-throughput production.
Collectively, these challenges, spanning detection, functional validation, targeting specificity, delivery efficiency, immunogenicity, and biosynthetic fidelity, underscore the need for a cohesive research roadmap. Addressing these interconnected issues in a staged manner is essential. The most immediate priority should be establishing standardized detection and quantification pipelines for poultry circRNAs, as reliable identification underpins all downstream functional and translational work. Once robust profiling platforms are in place, targeted modulation strategies and delivery systems can be developed and optimized with greater precision, paving the way for circRNA-based diagnostics and therapeutics that can meaningfully enhance poultry health and productivity. Addressing these interconnected challenges through coordinated research efforts will be essential for translating the strong evidence supporting our hypothesis, that circRNAs serve as key regulatory molecules in poultry diseases, into practical diagnostic tools and therapeutic interventions that can meaningfully enhance poultry health and productivity.
9 Conclusions and future perspectives
circRNAs are now recognized as important regulators in cellular physiology and disease, including antiviral immune responses, yet our understanding in poultry remains fragmentary. A persistent challenge is the absence of a unified nomenclature system—current naming conventions based on host genes (e.g., circMTO1) or functions (e.g., ciRS-7, MFACR) hinder cross-study comparisons and database integration. Equally critical are technical barriers in accurate detection, as most standard RNA sequencing pipelines are not optimized for circular transcripts, leading to inconsistent quantification. Functionally, while circRNAs have been linked to modulation of innate immunity and protection against viral infections, the majority of identified circRNAs still lack experimental validation, limiting their translational potential. To address these gaps, we propose four priority actions: (1) develop standardized, poultry-adapted pipelines for circRNA detection and quantification, including reference spike-ins and shared bioinformatic criteria; (2) establish a consensus nomenclature endorsed by the avian research community to facilitate data sharing; (3) invest in targeted functional assays—such as CRISPR/Cas13-mediated knockdowns—in physiologically relevant avian cell models to link circRNAs to immune phenotypes; and (4) accelerate delivery technology development, particularly nanoparticle and exosome systems, for precise circRNA modulation in vivo. By systematically tackling these limitations through coordinated methodological, functional, and translational research, circRNA-based diagnostics and therapeutics could transition from exploratory discovery to practical tools for enhancing poultry health, productivity, and disease resilience.
Author contributions
Sumayya: Conceptualization, Data curation, Formal analysis, Investigation, Methodology, Software, Supervision, Validation, Visualization, Writing – original draft, Writing – review & editing. HR: Conceptualization, Investigation, Validation, Visualization, Writing – original draft, Writing – review & editing. MS: Conceptualization, Investigation, Validation, Visualization, Writing – original draft, Writing – review & editing. BL: Conceptualization, Investigation, Supervision, Validation, Visualization, Writing – original draft, Writing – review & editing. HS: Funding acquisition, Supervision, Validation, Visualization, Writing – original draft, Writing – review & editing. OU: Methodology, Resources, Software, Validation, Visualization, Writing – original draft, Writing – review & editing. ASA: Resources, Software, Supervision, Validation, Writing – original draft, Writing – review & editing. AAA: Investigation, Methodology, Software, Supervision, Writing – original draft, Writing – review & editing. AE: Investigation, Project administration, Software, Supervision, Validation, Visualization, Writing – original draft, Writing – review & editing. MA: Conceptulization, Software, Writing - original draft, Validation & Visualization.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This work was supported and funded by the Deanship of Scientific Research at Imam Mohammad Ibn Saud Islamic University (IMSIU) (grant number IMSIU-DDRSP2501).
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The authors declare that no Gen AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
1. Zhang, Z, Huang, Y, Guo, A, and Yang, L. Research progress of circular RNA molecules in aging and age-related diseases. Ageing Res Rev. (2023) 87:101913. doi: 10.1016/j.arr.2023.101913
2. Zhao, X, Zhong, Y, Wang, X, Shen, J, and An, W. Advances in circular RNA and its applications. Int J Med Sci. (2022) 19:975–85. doi: 10.7150/ijms.71840
3. Bose, R, and Ain, R. Regulation of transcription by circular RNAs In:. Advances in experimental medicine and biology. Singapore: Springer (2018). 81–94.
4. Ebbesen, KK, Hansen, TB, and Kjems, J. Insights into circular RNA biology. RNA Biol. (2017) 14:1035–45. doi: 10.1080/15476286.2016.1271524
5. Huang, A, Zheng, H, Wu, Z, Chen, M, and Huang, Y. Circular RNA-protein interactions: functions, mechanisms, and identification. Theranostics. (2020) 10:3503–17. doi: 10.7150/thno.42174
6. He, AT, Liu, J, Li, F, and Yang, BB. Targeting circular RNAs as a therapeutic approach: current strategies and challenges. Signal Transduct Target Ther. (2021) 6:185. doi: 10.1038/s41392-021-00828-5
7. Misir, S, Wu, N, and Yang, BB. Specific expression and functions of circular RNAs. Cell Death Differ. (2022) 29:481–91. doi: 10.1038/s41418-022-00948-7
8. Niu, D, Wu, Y, and Lian, J. Circular RNA vaccine in disease prevention and treatment. Signal Transduct Target Ther. (2023) 8:123. doi: 10.1038/s41392-023-01561-x
9. Liu, X-d, Song, J, Liu, X, and Shan, H. Research note: circular RNA expressing in different developmental stages of the chicken bursa of Fabricius. Poult Sci. (2020) 99:3846–52. doi: 10.1016/j.psj.2020.04.026
10. Tian, Y, Ma, X, Jiang, Y, Han, J, Zhang, R, Xu, X, et al. Circular RNA circAKIRIN2 participates in the process of stress-induced immunosuppression affecting immune response to infectious bursal disease virus vaccine in chicken. Vet Microbiol. (2023) 281:109746. doi: 10.1016/j.vetmic.2023.109746
11. Xie, H, Sun, H, Mu, R, Li, S, Li, Y, Yang, C, et al. The role of circular RNAs in viral infection and related diseases. Virus Res. (2021) 291:198205. doi: 10.1016/j.virusres.2020.198205
12. Qiu, L, Chang, G, Bi, Y, Liu, X, and Chen, G. Circular RNA and mRNA profiling reveal competing endogenous RNA networks during avian leukosis virus, subgroup J-induced tumorigenesis in chickens. PLoS One. (2018) 13:e0204931. doi: 10.1371/journal.pone.0204931
13. Sanger, HL, Klotz, G, Riesner, D, Gross, HJ, and Kleinschmidt, AK. Viroids are single-stranded covalently closed circular RNA molecules existing as highly base-paired rod-like structures. Proc Natl Acad Sci. (1976) 73:3852–6. doi: 10.1073/pnas.73.11.3852
14. Hensgens, LAM, Arnberg, AC, Roosendaal, E, Van Der Horst, G, Van Der Veen, R, Van Ommen, G-JB, et al. Variation, transcription and circular RNAs of the mitochondrial gene for subunit I of cytochrome c oxidase. J Mol Biol. (1983) 164:35–58. doi: 10.1016/0022-2836(83)90086-4
15. Kos, A, Dijkema, R, Arnberg, AC, van der Meide, PH, and Schellekens, H. The hepatitis delta (δ) virus possesses a circular RNA. Nature. (1986) 323:558–60. doi: 10.1038/323558a0
16. Capel, B, Swain, A, Nicolis, S, Hacker, A, Walter, M, Koopman, P, et al. Circular transcripts of the testis-determining gene Sry in adult mouse testis. Cell. (1993) 73:1019–30. doi: 10.1016/0092-8674(93)90279-Y
17. Jeck, WR, and Sharpless, NE. Detecting and characterizing circular RNAs. Nat Biotechnol. (2014) 32:453–61. doi: 10.1038/nbt.2890
18. Salzman, J, Gawad, C, Wang, PL, Lacayo, N, and Brown, PO. Circular RNAs are the predominant transcript isoform from hundreds of human genes in diverse cell types. PLoS One. (2012) 7:e30733. doi: 10.1371/journal.pone.0030733
19. Hansen, TB, Jensen, TI, Clausen, BH, Bramsen, JB, Finsen, B, Damgaard, CK, et al. Natural RNA circles function as efficient microRNA sponges. Nature. (2013) 495:384–8. doi: 10.1038/nature11993
20. Jeck, WR, Sorrentino, JA, Wang, K, Slevin, MK, Burd, CE, Liu, J, et al. Circular RNAs are abundant, conserved, and associated with ALU repeats. RNA. (2013) 19:141–57. doi: 10.1261/rna.035667.112
21. Kristensen, LS, Andersen, MS, Stagsted, LVW, Ebbesen, KK, Hansen, TB, and Kjems, J. The biogenesis, biology and characterization of circular RNAs. Nat Rev Genet. (2019) 20:675–91. doi: 10.1038/s41576-019-0158-7
22. Zhang, L, Zhang, Y, Wang, Y, Zhao, Y, Ding, H, and Li, P. Circular RNAs: functions and clinical significance in cardiovascular disease. Front Cell Dev Biol. (2020):8. doi: 10.3389/fcell.2020.584051
23. Tang, Q, and Hann, SS. Biological roles and mechanisms of circular RNA in human cancers. Onco Targets Ther. (2020) 13:2067–92. doi: 10.2147/OTT.S233672
24. Zhang, X, Yan, Y, Lei, X, Li, A, Zhang, H, Dai, Z, et al. Circular RNA alterations are involved in resistance to avian leukosis virus subgroup-J-induced tumor formation in chickens. Oncotarget. (2017) 8:34961–70. doi: 10.18632/oncotarget.16442
25. Chen, L, Ruan, J, Chen, Y, Deng, W, Lai, J, Fan, L, et al. RNA sequencing reveals CircRNA expression profiles in chicken embryo fibroblasts infected with velogenic Newcastle disease virus. Front Vet Sci. (2023) 10:1167444. doi: 10.3389/fvets.2023.1167444
26. Yu, H, Mi, C, Wang, Q, Zou, W, Dai, G, Zhang, T, et al. Comprehensive analyses of circRNA expression profiles and function prediction in chicken Cecums after Eimeria tenella infection. Front Cell Infect Microbiol. (2021) 11:628667. doi: 10.3389/fcimb.2021.628667
27. Wang, L, Zheng, G, Yuan, Y, Wang, Z, Wang, Q, Sun, M, et al. CircRUNX2.2, highly expressed in Marek's disease tumor tissues, functions in cis to regulate parental gene RUNX2 expression. Poult Sci. (2024) 103:104045. doi: 10.1016/j.psj.2024.104045
28. Awan, FM, Yang, BB, Naz, A, Hanif, A, Ikram, A, Obaid, A, et al. The emerging role and significance of circular RNAs in viral infections and antiviral immune responses: possible implication as theranostic agents. RNA Biol. (2020) 18:1–15. doi: 10.1080/15476286.2020.1790198
29. Zucko, D, and Boris-Lawrie, K. Circular RNAs are regulators of diverse animal transcriptomes: one health perspective. Front Genet. (2020) 11:999. doi: 10.3389/fgene.2020.00999
30. Huang, Y, Wang, Y, Zhang, C, and Sun, X. Biological functions of circRNAs and their progress in livestock and poultry. Reprod Domest Anim. (2020) 55:1667–77. doi: 10.1111/rda.13816
31. Yuan, P, Zhao, Y, Li, H, Li, S, Fan, S, Zhai, B, et al. CircRNAs related to breast muscle development and their interaction regulatory network in Gushi chicken. Genes (Basel). (2022) 13:1974. doi: 10.3390/genes13111974
32. Jing, Y, Cheng, B, Wang, H, Bai, X, Zhang, Q, Wang, N, et al. The landscape of the long non-coding RNAs and circular RNAs of the abdominal fat tissues in the chicken lines divergently selected for fatness. BMC Genomics. (2022) 23:790. doi: 10.1186/s12864-022-09045-y
33. Tian, W, Zhang, B, Zhong, H, Nie, R, Ling, Y, Zhang, H, et al. Dynamic expression and regulatory network of circular RNA for abdominal preadipocytes differentiation in chicken (Gallus gallus). Front Cell Dev Biol. (2021) 9:761638. doi: 10.3389/fcell.2021.761638
34. Chen, L-L. The expanding regulatory mechanisms and cellular functions of circular RNAs. Nat Rev Mol Cell Biol. (2020) 21:475–90. doi: 10.1038/s41580-020-0243-y
35. Zhou, W-Y, Cai, Z-R, Liu, J, Wang, D-S, Ju, H-Q, and Xu, R-H. Circular RNA: metabolism, functions and interactions with proteins. Mol Cancer. (2020) 19:172. doi: 10.1186/s12943-020-01286-3
36. Kim, E, Kim, YK, and Lee, S-JV. Emerging functions of circular RNA in aging. Trends Genet. (2021) 37:819–29. doi: 10.1016/j.tig.2021.04.014
37. Geng, X, Jia, Y, Zhang, Y, Shi, L, Li, Q, Zang, A, et al. Circular RNA: biogenesis, degradation, functions and potential roles in mediating resistance to anticarcinogens. Epigenomics. (2020) 12:267–83. doi: 10.2217/epi-2019-0295
38. Qu, S, Yang, X, Li, X, Wang, J, Gao, Y, Shang, R, et al. Circular RNA: a new star of noncoding RNAs. Cancer Lett. (2015) 365:141–8. doi: 10.1016/j.canlet.2015.06.003
39. Floris, G, Zhang, L, Follesa, P, and Sun, T. Regulatory role of circular RNAs and neurological disorders. Mol Neurobiol. (2017) 54:5156–65. doi: 10.1007/s12035-016-0055-4
40. Chen, X, and Lu, Y. Circular RNA: biosynthesis in vitro. Front Bioeng Biotechnol. (2021):9. doi: 10.3389/fbioe.2021.787881
41. Enuka, Y, Lauriola, M, Feldman, ME, Sas-Chen, A, Ulitsky, I, and Yarden, Y. Circular RNAs are long-lived and display only minimal early alterations in response to a growth factor. Nucleic Acids Res. (2016) 44:1370–83. doi: 10.1093/nar/gkv1367
42. Nisar, S, Bhat, A, Singh, M, Karedath, T, Rizwan, A, Hashem, S, et al. Insights into the role of CircRNAs: biogenesis, characterization, functional, and clinical impact in human malignancies. Front Cell Dev Biol. (2021) 9. doi: 10.3389/fcell.2021.617281
43. Shang, Q, Yang, Z, Jia, R, and Ge, S. The novel roles of circRNAs in human cancer. Mol Cancer. (2019) 18:6. doi: 10.1186/s12943-018-0934-6
44. Verduci, L, Tarcitano, E, Strano, S, Yarden, Y, and Blandino, G. CircRNAs: role in human diseases and potential use as biomarkers. Cell Death Dis. (2021) 12:468. doi: 10.1038/s41419-021-03743-3
45. Jin, W, Zhao, Y, Zhai, B, Li, Y, Fan, S, Yuan, P, et al. Characteristics and expression profiles of circRNAs during abdominal adipose tissue development in Chinese Gushi chickens. PLoS One. (2021) 16:e0249288. doi: 10.1371/journal.pone.0249288
46. Pan, Z, Yang, C, Zhao, R, Jiang, X, Yu, C, and Li, Z. Characterization of lncRNA/circRNA-miRNA-mRNA network to reveal potential functional ceRNAs in the skeletal muscle of chicken. Front Physiol. (2022) 13:969854. doi: 10.3389/fphys.2022.969854
47. Shao, B, Wang, Z, Luo, P, Du, P, Zhang, X, Zhang, H, et al. Identifying insulin-responsive circRNAs in chicken pectoralis. BMC Genomics. (2025) 26:148. doi: 10.1186/s12864-025-11347-w
48. Wang, M, Yu, F, and Li, P. Circular RNAs: characteristics, function and clinical significance in hepatocellular carcinoma. Cancer. (2018) 10:258. doi: 10.3390/cancers10080258
49. Feng, X-Y, Zhu, S-X, Pu, K-J, Huang, H-J, Chen, Y-Q, and Wang, W. New insight into circRNAs: characterization, strategies, and biomedical applications. Exp Hematol Oncol. (2023) 12:42. doi: 10.1186/s40164-023-00451-w
50. Patop, I, Wüst, S, and Kadener, S. Past, present, and future of circRNAs. EMBO J. (2019) 38:e100836. doi: 10.15252/embj.2018100836
51. Dey, P, Buragohain, T, Das, M, and Banerjee, S. Exploring the role of non-coding RNA mediated regulation of signaling pathways in endometrial cancer. Adv Cancer Biol Metastasis. (2023) 9:100111. doi: 10.1016/j.adcanc.2023.100111
52. Zhang, X, Lu, N, Wang, L, Wang, Y, Li, M, Zhou, Y, et al. Circular RNAs and esophageal cancer. Cancer Cell Int. (2020) 20:242. doi: 10.1186/s12935-020-01670-5
53. Liu, K-S, Pan, F, Mao, X-D, Liu, C, and Chen, Y-J. Biological functions of circular RNAs and their roles in occurrence of reproduction and gynecological diseases. Am J Transl Res. (2019) 11:1.
54. Zhang, Y, Liu, L, Liu, K, Wang, M, Su, X, and Wang, J. Regulatory mechanism of circular RNA involvement in osteoarthritis. Front Surg. (2023) 9:1049513. doi: 10.3389/fsurg.2022.1049513
55. Hall, IF, Climent, M, Quintavalle, M, Farina, FM, Schorn, T, Zani, S, et al. Circ_Lrp6, a circular RNA enriched in vascular smooth muscle cells, acts as a sponge regulating miRNA-145 function. Circ Res. (2019) 124:498–510. doi: 10.1161/CIRCRESAHA.118.314240
56. Wang, L, You, Z, Wang, M, Yuan, Y, Liu, C, Yang, N, et al. Genome-wide analysis of circular RNAs involved in Marek’s disease tumourigenesis in chickens. RNA Biol. (2020) 17:517–27. doi: 10.1080/15476286.2020.1713538
57. Yang, T, Qiu, L, Bai, M, Wang, L, Hu, X, Huang, L, et al. Identification, biogenesis and function prediction of novel circRNA during the chicken ALV-J infection. Anim Biotechnol. (2022) 33:981–91. doi: 10.1080/10495398.2020.1856125
58. Chen, X, Ouyang, H, Wang, Z, Chen, B, and Nie, Q. A novel circular RNA generated by FGFR2 gene promotes myoblast proliferation and differentiation by sponging miR-133a-5p and miR-29b-1-5p. Cells. (2018) 7:199. doi: 10.3390/cells7110199
59. Lei, Q, Hu, X, Han, H, Wang, J, Liu, W, Zhou, Y, et al. Integrative analysis of circRNA, miRNA, and mRNA profiles to reveal ceRNA regulation in chicken muscle development from the embryonic to post-hatching periods. BMC Genomics. (2022) 23:342. doi: 10.1186/s12864-022-08525-5
60. Chen, B, Yu, J, Guo, L, Byers, MS, Wang, Z, Chen, X, et al. Circular RNA circHIPK3 promotes the proliferation and differentiation of chicken myoblast cells by sponging miR-30a-3p. Cells. (2019) 8:177. doi: 10.3390/cells8020177
61. Shen, X, Liu, Z, Cao, X, He, H, Han, S, Chen, Y, et al. Circular RNA profiling identified an abundant circular RNA circTMTC1 that inhibits chicken skeletal muscle satellite cell differentiation by sponging miR-128-3p. Int J Biol Sci. (2019) 15:2265–81. doi: 10.7150/ijbs.36412
62. Yang, Z, He, T, and Chen, Q. The roles of CircRNAs in regulating muscle development of livestock animals. Front Cell Dev Biol. (2021) 9:619329. doi: 10.3389/fcell.2021.619329
63. Gu, A, Jaijyan, DK, Yang, S, Zeng, M, Pei, S, and Zhu, H. Functions of circular RNA in human diseases and illnesses. Noncoding RNA. (2023) 9:38. doi: 10.3390/ncrna9040038
64. Xiao, C, Sun, T, Yang, Z, Zou, L, Deng, J, and Yang, X. Whole-transcriptome RNA sequencing reveals the global molecular responses and circRNA/lncRNA-miRNA-mRNA ceRNA regulatory network in chicken fat deposition. Poult Sci. (2022) 101:102121. doi: 10.1016/j.psj.2022.102121
65. Zhang, M, Han, Y, Zhai, Y, Ma, X, An, X, Zhang, S, et al. Integrative analysis of circRNAs, miRNAs, and mRNAs profiles to reveal ceRNAs networks in chicken intramuscular and abdominal adipogenesis. BMC Genomics. (2020) 21:594. doi: 10.1186/s12864-020-07000-3
66. Zhang, P, Chao, Z, Zhang, R, Ding, R, Wang, Y, Wu, W, et al. Circular RNA regulation of myogenesis. Cells. (2019) 8:885. doi: 10.3390/cells8080885
67. Yu, C-Y, and Kuo, H-C. The emerging roles and functions of circular RNAs and their generation. J Biomed Sci. (2019) 26:29. doi: 10.1186/s12929-019-0523-z
68. Li, Q, Wang, Y, Wu, S, Zhou, Z, Ding, X, Shi, R, et al. CircACC1 regulates assembly and activation of AMPK complex under metabolic stress. Cell Metab. (2019) 30:157–173.e7. doi: 10.1016/j.cmet.2019.05.009
69. Sadhukhan, S, Sinha, T, Dey, S, and Panda, AC. Subcellular localization of circular RNAs: where and why. Biochem Biophys Res Commun. (2024) 715:149937. doi: 10.1016/j.bbrc.2024.149937
70. Chen, B, and Huang, S. Circular RNA: an emerging non-coding RNA as a regulator and biomarker in cancer. Cancer Lett. (2018) 418:41–50. doi: 10.1016/j.canlet.2018.01.011
71. Wei, J, Li, M, Xue, C, Chen, S, Zheng, L, Deng, H, et al. Understanding the roles and regulation patterns of circRNA on its host gene in tumorigenesis and tumor progression. J Exp Clin Cancer Res. (2023) 42:103. doi: 10.1186/s13046-023-02657-6
72. Xu, T, Wang, M, Jiang, L, Li,, Wan, L, Chen, Q-N, et al. Circrnas in anticancer drug resistance: recent advances and future potential. Mol Cancer. (2020) 19:127. doi: 10.1186/s12943-020-01240-3
73. Xu, X, Zhang, J, Tian, Y, Gao, Y, Dong, X, Chen, W, et al. CircRNA inhibits DNA damage repair by interacting with host gene. Mol Cancer. (2020) 19:128. doi: 10.1186/s12943-020-01246-x
74. Zang, J, Lu, D, and Xu, A. The interaction of circRNAs and RNA binding proteins: An important part of circRNA maintenance and function. J Neurosci Res. (2020) 98:87–97. doi: 10.1002/jnr.24356
75. Tang, Z, Li, X, Zhao, J, Qian, F, Feng, C, Li, Y, et al. TRCirc: a resource for transcriptional regulation information of circRNAs. Brief Bioinform. (2018) 20:1566–1573. doi: 10.1093/bib/bby045
76. Eidem, TM, Kugel, JF, and Goodrich, JA. Noncoding RNAs: regulators of the mammalian transcription machinery. J Mol Biol. (2016) 428:2652–9. doi: 10.1016/j.jmb.2016.02.019
77. Shen, H, Liu, B, Xu, J, Zhang, B, Wang, Y, Shi, L, et al. Circular RNAs: characteristics, biogenesis, mechanisms and functions in liver cancer. J Hematol Oncol. (2021) 14:134. doi: 10.1186/s13045-021-01145-8
78. Fischer, J, and Leung, A. Circrnas: a regulator of cellular stress. Crit Rev Biochem Mol Biol. (2017) 52:220–33. doi: 10.1080/10409238.2016.1276882
79. Tao, M, Zheng, M, Xu, Y, Shuo, M, Zhang, W, and Ju, S. CircRNAs and their regulatory roles in cancers. Mol Med. (2021) 27:94. doi: 10.1186/s10020-021-00335-4
80. Chen, X, Zhou, M, Yant, L, and Huang, C. Circular RNA in disease: basic properties and biomedical relevance. WIREs RNA. (2022) 13:e1721. doi: 10.1002/wrna.1723
81. Zhou, Z, Sun, B, Huang, S, and Zhao, L. Roles of circular RNAs in immune regulation and autoimmune diseases. Cell Death Dis. (2019) 10:503. doi: 10.1038/s41419-019-1744-5
82. Chang, X, Zhu, G, Cai, Z, Wang, Y, Lian, R, Tang, X, et al. miRNA, lncRNA and circRNA: targeted molecules full of therapeutic prospects in the development of diabetic retinopathy. Front Endocrinol. (2021) 12:771552. doi: 10.3389/fendo.2021.771552
83. Arnaiz, E, Solé, C, Manterola, L, Iparraguirre, L, Otaegui, D, and Lawrie, C. Circrnas and cancer: biomarkers and master regulators. Semin Cancer Biol. (2019) 58:90–9. doi: 10.1016/j.semcancer.2018.12.002
84. Amelimojarad, M, and Amelimojarad, M. Regulatory mechanism of circular RNAs in brain and neurodegenerative diseases. Front Mol Neurosci. (2025) 18:1507575. doi: 10.3389/fnmol.2025.1507575
85. Galli, V, Vinciguerra, S, Biagioli, M, and Morandell, J. Circular RNAs as disease modifiers of complex neurologic disorders. Front Pharmacol. (2025) 16:1577496. doi: 10.3389/fphar.2025.1577496
86. Pan, R, Koo, C, Su, W, You, Q, Guo, H, and Liu, B. Circular RNAs modulate cell death in cardiovascular diseases. Cell Death Discov. (2025) 11:214. doi: 10.1038/s41420-025-02504-x
87. Wu, DP, Wei, YS, Hou, LX, Du, YX, Yan, QQ, Liu, LL, et al. Circular RNA APP contributes to Alzheimer's disease pathogenesis by modulating microglial polarization via miR-1906/CLIC1 axis. Alzheimer's Res Ther. (2025) 17:44. doi: 10.1186/s13195-025-01698-7
88. Hatzimanolis, O, Sykes, AM, and Cristino, AS. Circular RNAs in neurological conditions - computational identification, functional validation, and potential clinical applications. Mol Psychiatry. (2025) 30:1652–75. doi: 10.1038/s41380-025-02925-1
89. Zheng, X, Song, L, Cao, C, and Sun, S. Multiple roles of circular RNAs in prostate cancer: from the biological basis to potential clinical applications. Eur J Med Res. (2025) 30:140. doi: 10.1186/s40001-025-02382-0
90. Cao, M-X, Xie, X-D, Wang, X-R, Hu, W-Y, Zhao, Y, Chen, Q, et al. Separation, purification, structure analysis, in vitro antioxidant activity and circRNA-miRNA-mRNA regulatory network on PRV-infected RAW264.7 cells of a polysaccharide derived from arthrospira platensis. Antioxidants. (2021) 10:1689. doi: 10.3390/antiox10111689
91. Wu, P, Zhou, K, Zhang, J, Ling, X, Zhang, X, Zhang, L, et al. Identification of crucial circRNAs in skeletal muscle during chicken embryonic development. BMC Genomics. (2022) 23:330. doi: 10.1186/s12864-022-08588-4
92. Das, A, Das, A, Das, D, Abdelmohsen, K, and Panda, AC. Circular RNAs in myogenesis. Biochim Biophys Acta (BBA) Gene Regul Mech. (2020) 1863:194372. doi: 10.1016/j.bbagrm.2019.02.011
93. Miller, PJ, Decanini, EL, and Afonso, CL. Newcastle disease: evolution of genotypes and the related diagnostic challenges. Infect Genet Evol. (2010) 10:26–35. doi: 10.1016/j.meegid.2009.09.012
94. Chasseur, AS, Trozzi, G, Istasse, C, Petit, A, Rasschaert, P, Denesvre, C, et al. Marek’s disease virus virulence genes encode circular RNAs. J Virol. (2022) 96:e0032122. doi: 10.1128/jvi.00321-22
95. Xu, J, Chen, X, Sun, Y, Shi, Y, Teng, F, Lv, M, et al. The regulation network and clinical significance of circular RNAs in breast cancer. Front Oncol. (2021) 11:691317. doi: 10.3389/fonc.2021.691317
96. Nie, MW, Han, YC, Shen, ZJ, and Xie, HZ. Identification of circRNA and mRNA expression profiles and functional networks of vascular tissue in lipopolysaccharide-induced sepsis. J Cell Mol Med. (2020) 24:7915–27. doi: 10.1111/jcmm.15424
97. Yu, X, Xu, H, Xing, Y, Sun, D, Li, D, Shi, J, et al. Identifying essential hub genes and circRNA-regulated ceRNA networks in hepatocellular carcinoma. Int J Mol Sci. (2025) 26:1408. doi: 10.3390/ijms26041408
98. Tang, Z, Li, X, Zheng, Y, Liu, J, Liu, C, and Li, X. The role of competing endogenous RNA network in the development of hepatocellular carcinoma: potential therapeutic targets. Front Cell Dev Biol. (2024) 12:1341999. doi: 10.3389/fcell.2024.1341999
99. Yu, H, Zou, W, Xin, S, Wang, X, Mi, C, Dai, G, et al. Association analysis of single nucleotide polymorphisms in the 5′ regulatory region of the IL-6 Gene with Eimeria tenella resistance in Jinghai yellow chickens. Genes. (2019) 10:890. doi: 10.3390/genes10110890
100. Chen, D, Miao, Z, Peng, M, Xing, H, Zhang, H, and Teng, X. The co-expression of circRNA and mRNA in the thymuses of chickens exposed to ammonia. Ecotoxicol Environ Saf. (2019) 176:146–52. doi: 10.1016/j.ecoenv.2019.03.076
101. Fan, X-C, Liu, T-L, Wang, Y, Wu, X-M, Wang, Y-X, Lai, P, et al. Zhao G-H: genome-wide analysis of differentially expressed profiles of mRNAs, lncRNAs and circRNAs in chickens during Eimeria necatrix infection. Parasites Vectors. (2020) 13:167. doi: 10.1186/s13071-020-04047-9
102. Kim, WH, Chaudhari, AA, and Lillehoj, HS. Involvement of T cell immunity in avian coccidiosis. Front Immunol. (2019) 10:2732. doi: 10.3389/fimmu.2019.02732
103. Girard, F, Fort, G, Yvoré, P, and Quéré, P. Kinetics of specific immunoglobulin a, M and G production in the duodenal and caecal mucosa of chickens infected with Eimeria acervulina or Eimeria tenella. Int J Parasitol. (1997) 27:803–9. doi: 10.1016/S0020-7519(97)00044-1
104. Lillehoj, HS. Role of T lymphocytes and cytokines in coccidiosis. Int J Parasitol. (1998) 28:1071–81. doi: 10.1016/S0020-7519(98)00075-7
105. Min, W, Kim, WH, Lillehoj, EP, and Lillehoj, HS. Recent progress in host immunity to avian coccidiosis: IL-17 family cytokines as sentinels of the intestinal mucosa. Dev Compar Immunol. (2013) 41:418–28. doi: 10.1016/j.dci.2013.04.003
106. Huang, C-J, and Choo, KB. Circular RNA-and microRNA-mediated post-transcriptional regulation of preadipocyte differentiation in adipogenesis: from expression profiling to signaling pathway. Int J Mol Sci. (2023) 24:4549. doi: 10.3390/ijms24054549
107. Liu, J, Liu, S, Zhang, W, Hu, X, Mao, H, Liu, S, et al. Transcriptome RNA sequencing reveals that circular RNAs are abundantly expressed in embryonic breast muscle of duck. Vet Sci. (2023) 10:75. doi: 10.3390/vetsci10020075
108. Halevy, O, Piestun, Y, Allouh, MZ, Rosser, BWC, Rinkevich, Y, Reshef, R, et al. Pattern of Pax7 expression during myogenesis in the posthatch chicken establishes a model for satellite cell differentiation and renewal. Dev Dyn. (2004) 231:489–502. doi: 10.1002/dvdy.20151
109. Ouyang, H, Chen, X, Wang, Z, Yu, J, Jia, X, Li, Z, et al. Circular RNAs are abundant and dynamically expressed during embryonic muscle development in chickens. DNA Res. (2018) 25:71–86. doi: 10.1093/dnares/dsx039
110. Ouyang, H, Chen, X, Li, W, Li, Z, Nie, Q, and Zhang, X. Circular RNA circSVIL promotes myoblast proliferation and differentiation by sponging miR-203 in chicken. Front Genet. (2018) 9:172. doi: 10.3389/fgene.2018.00172
111. Shi, H, He, Y, Li, X, Du, Y, Zhao, J, and Ge, C. Regulation of non-coding RNA in the growth and development of skeletal muscle in domestic chickens. Genes. (2022) 13:1033. doi: 10.3390/genes13061033
112. Kristensen, L, Hansen, T, Venø, M, and Kjems, J. Circular RNAs in cancer: opportunities and challenges in the field. Oncogene. (2017) 37:555–65. doi: 10.1038/s41392-021-00571-0
113. Yang, L, and Chen, L-L. The biogenesis, functions, and challenges of circular RNAs. Mol Cell. (2018) 71:428–42.
114. Szabo, L, and Salzman, J. Detecting circular RNAs: bioinformatic and experimental challenges. Nat Rev Genet. (2016) 17:679–92. doi: 10.1038/nrg.2016.114
115. Chen, L-L, and Yang, L. Regulation of circRNA biogenesis. RNA Biol. (2015) 12:381–8. doi: 10.1080/15476286.2015.1020271
116. Gallala, M. Application of CRISPR/Cas gene editing for infectious disease control in poultry. Open Life Sci. (2025) 20:20251095. doi: 10.1515/biol-2025-1095
117. Alqahtani, S, Alqahtani, T, Venkatesan, K, Sivadasan, D, Ahmed, R, Elfadil, H, et al. Unveiling pharmacogenomics insights into circular RNAs: toward precision medicine in cancer therapy. Biomolecules. (2025) 15:535.
118. Ji, P, Wu, W, Chen, S, Zheng, Y, Zhou, L, Zhang, J, et al. Expanded expression landscape and prioritization of circular RNAs in mammals. Cell Rep. (2019) 26:3444–3460.e5. doi: 10.1016/j.celrep.2019.02.078
119. Tam, Y, Chen, S, and Cullis, P. Advances in lipid nanoparticles for siRNA delivery. Pharmaceutics. (2013) 5:498–507. doi: 10.3390/pharmaceutics5030498
120. Liu, X, Zhang, Y, Zhou, S, Dain, L, Mei, L, and Zhu, G. Circular RNA: An emerging frontier in RNA therapeutic targets, RNA therapeutics, and mRNA vaccines. J Control Rel Off. (2022). doi: 10.3390/biom15040535
121. Zhou, J, Shum, K-T, Burnett, J, and Rossi, J. Nanoparticle-based delivery of RNAi therapeutics: progress and challenges. Pharmaceuticals. (2013) 6:85–107. doi: 10.3390/ph6010085
122. Chen, YG, Kim, MV, Chen, X, Batista, PJ, Aoyama, S, Wilusz, JE, et al. Sensing self and foreign circular RNAs by intron identity. Mol Cell. (2017) 67:228–238.e5. doi: 10.1016/j.molcel.2017.05.022
123. Chen, YG, Chen, R, Ahmad, S, Verma, R, Kasturi, SP, Amaya, L, et al. N6-Methyladenosine modification controls circular RNA immunity. Mol Cell. (2019) 76:96–109.e9. doi: 10.1016/j.molcel.2019.07.016
Keywords: circular RNAs, biogenesis, virus, regulatory mechanism, poultry diseases
Citation: Sumayya, Rudayni HA, Shuaib M, Li B, Sun H, Ullah O, Alawam AS, Allam AA, Elolimy AA and Abd El-Hack ME (2025) Regulatory roles of circular RNAs in poultry diseases: mechanisms and research advances. Front. Vet. Sci. 12:1626464. doi: 10.3389/fvets.2025.1626464
Edited by:
Mariangela Caroprese, University of Foggia, ItalyReviewed by:
Ali Hassan Nawaz, Nanjing Agricultural University, ChinaJunjie Gu, Sichuan University, China
Copyright © 2025 Sumayya, Rudayni, Shuaib, Li, Sun, Ullah, Alawam, Allam, Elolimy and Abd El-Hack. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Ahmed A. Elolimy, ZWxvbGlteUB1YWV1LmFjLmFl