 Ana Šikić1Ema Gagović2
Ana Šikić1Ema Gagović2 Alicia Rojas3Magda Sindičić4Daria Jurković Žilić2Šimun Naletlić5Davor Balić6Adnan Hodžić7
Alicia Rojas3Magda Sindičić4Daria Jurković Žilić2Šimun Naletlić5Davor Balić6Adnan Hodžić7 Relja Beck2*
Relja Beck2*- 1Department for Parasitology and Invasive Diseases, Faculty of Veterinary Medicine, University of Zagreb, Zagreb, Croatia
- 2Laboratory for Parasitology, Department of Microbiology and Parasitology, Croatian Veterinary Institute, Zagreb, Croatia
- 3Laboratory of Helminthology, Faculty of Microbiology, University of Costa Rica, San José, Costa Rica
- 4Department for Hunting and Wild Animals, Faculty of Veterinary Medicine, University of Zagreb, Zagreb, Croatia
- 5Laboratory for Pathology, Department of Pathology, Croatian Veterinary Institute, Zagreb, Croatia
- 6Veterinary Department Vinkovci, Croatian Veterinary Institute, Vinkovci, Croatia
- 7Center for Microbiology and Environmental Systems Science (CMESS), Department of Microbiology and Ecosystem Science, Division of Microbial Ecology (DoME), University of Vienna, Vienna, Austria
This study presents the first molecularly confirmed identification of the cestode Spirometra mansoni in the golden jackals (Canis aureus) in Croatia, and possibly the first such report in Europe. Of 198 jackals examined between 2020 and 2025, adult Spirometra worms were recovered from three individuals. The morphological characteristics of these specimens were consistent with S. mansoni, and their identity was confirmed by PCR and sequencing of the mitochondrial cox1 and nad1 genes. Phylogenetic analysis grouped the obtained sequences within the S. mansoni clade, with strong posterior probability support. This finding expands the known host range and geographic distribution of S. mansoni and underscores the importance of integrating molecular diagnostics in parasitological surveys. Further research is needed to assess the role of golden jackals and other wildlife in the epidemiology of Spirometra spp. in Europe.
Introduction
In recent years, parasites of the genus Spirometra (family Diphyllobothriidae, order Diphyllobothriidea) have increasingly attracted the attention of veterinary and human health professionals in Europe, primarily due to their role as causative agents of the food-borne zoonosis known as sparganosis. The disease was first described in China in 1882 (1, 2), and since then, more than 2,000 cases have been reported globally—predominantly in East and Southeast Asia—with sporadic cases documented in South America, Africa, and Europe (3, 4).
Despite the growing number of reports, many aspects of Spirometra biology remain unresolved, particularly in terms of taxonomy, geographical distribution, host range, zoonotic potential, and disease epidemiology (4–7). Adult Spirometra worms are morphologically similar to those within order Diphyllobothriidea, exhibiting considerable morphological uniformity as well as host-related intraspecific variability. Consequently, reliable identification often requires molecular methods, although there is still a lack of consistent morphological data linked to genetically confirmed specimens (7). Additionally, available molecular data for some Spirometra species have been inconsistent, with many nucleotide sequences available in the GenBank® database being misidentified (7, 8). Therefore, identification within the genus should be supported by molecular, morphological, ecological, or pathological evidence, as previously suggested (9).
Molecular studies suggest the existence of at least seven genetically distinct lineages or species within the genus, as summarized in the comprehensive taxonomic reviews by Kuchta et al. (4, 7). These tapeworms have a complex life cycle involving two intermediate hosts. The first larval stage (coracidium) hatches from eggs in water and is ingested by copepods (Cyclops spp.), in which the procercoid develops. The second intermediate host—typically amphibians, reptiles, or mammals—ingests the copepod, after which the plerocercoid larva develops and migrates to the subcutaneous or muscular tissues. Carnivorous mammals serve as definitive hosts, becoming infected upon consuming the second intermediate host (10, 11).
Spirometra species have been reported from all continents except Antarctica. While S. mansoni (Cobbold, 1882) is considered cosmopolitan, other species appear to have more restricted distributions. However, due to the lack of molecular confirmation in most reports, current distribution maps remain unreliable and subject to revision (4, 7). In Europe, only two species, S. erinaceieuropaei and S. mansoni have been confirmed through molecular analysis (4). Plerocercoid stages have been identified morphologically in 17 European countries, but records of definitive hosts remain sparse. Spirometra erinaceieuropaei is potentially endemic to Central and Eastern Europe and exhibits low genetic diversity (12). Other historical species (Spirometra ranarum, Spirometra raillieti) are considered taxonomically doubtful due to insufficient descriptions and a lack of genetic data (7).
Although European wildlife harbors a wide array of potential definitive and intermediate hosts, there is a significant knowledge gap regarding their role in Spirometra transmission. The genus has been molecularly confirmed in only three definitive host species, namely wolves (Canis lupus), Eurasian lynxes (Lynx lynx), and domestic cats (Felis catus). Moreover, morphological records exist for the European wildcat (Felis silvestris), red fox (Vulpes vulpes), and raccoon dog (Nyctereutes procyonoides) (4, 7, 12, 13). Among all native European carnivores, the golden jackal (Canis aureus) is currently undergoing the most rapid and dramatic range expansion (14). This medium-sized, omnivorous and opportunistic canid is distributed across southern Asia, the Middle East, and outheastern to central Europe, where it inhabits a wide range of habitats, including agricultural landscapes and semi-urban areas (15, 16). Golden jackals are highly adaptable to human-modified habitats, where they readily exploit accessible food sources (17). Due to this ecological flexibility, they have emerged as one of the most relevant species for investigating the role of potential definitive hosts of Spirometra in Europe. This study presents the first molecular identification of Spirometra mansoni in golden jackals in Croatia, contributing to a broader understanding of the parasite's host range and geographic distribution.
Materials and methods
Study area
This study was conducted in Central Croatia, a region situated in the continental part of the country that features lowland and hilly landscapes interspersed with river valleys and forested zones. The sampling locations were located within a mosaic of mixed deciduous forests, agricultural land, shrub vegetation, and rural settlements, typical of the transitional zone between the Pannonian Basin and the Dinaric Mountains. The climate is temperate continental, characterized by hot summers, cold winters, and significant seasonal variations in both temperature and precipitation (18). The region is rich in freshwater resources, with numerous rivers, streams, and wetlands distributed throughout the area. These water bodies, along with extensive forest coverage, provide diverse habitats for a variety of amphibians, reptiles, birds, and mammals (19).
The ecological heterogeneity and the presence of permanent surface water make this landscape particularly favorable for supporting complex food webs and parasite transmission cycles. Specifically, the proximity to natural water sources, forest edges, and agricultural fields creates a dynamic interface suitable for the coexistence of both intermediate and definitive hosts of various parasitic species. The golden jackal is well adapted to such environments and frequently occupies ecotones between natural and anthropogenic habitats.
Parasite collection and morphological identification
A total of 198 golden jackal carcasses were submitted to the Croatian Veterinary Institute between 2020 and 2025. The carcasses were collected either as part of the national rabies surveillance program or through roadkill monitoring. The approximate geographic origin was recorded for each animal. For better visibility of the obtained results, positive samples were mapped using QGIS software version 3.30.0 RC. During the necropsy, strict biosafety measures were implemented to prevent potential infection with Echinococcus spp. The intestines were opened longitudinally, and their contents visually inspected. Macroscopically visible tapeworms, were isolated, washed in water, and preserved in 70% ethanol. Prior to preservation specimens were morphologically identified using available morphological keys (7) under a Zeiss Stereo Discovery V20 and Imager M.2 microscope, equipped with Axiovision and ZEN2 Pro software.
Molecular characterization and phylogenetic analysis
Genomic DNA was extracted from individual worms from each animal using the DNeasy Blood and Tissue Kit (Qiagen, Germany) on the QIAcube robotic workstation (Qiagen, Germany). PCR amplification of the mitochondrial cytochrome c oxidase subunit 1 (cox1) gene was performed using primers Diphyllo-Cox1-F (5′-TAGACTAAGTGTTTTCAAAACACTA−3′) and Diphyllo-Cox1-R (5′-ATAGCATGATGCAAAAGG−3′) (20), while the NADH dehydrogenase subunit 1 (nad1) gene was amplified using primers JB11 and JB12 (21). PCR amplification was conducted with GoTaq® G2 Hot Start Colorless Master Mix (Promega, USA) under the following cycling conditions: 40 cycles of 30 s at 94°C, 30 s at 56°C (cox1) or 50°C (nad1), and 60 s at 72°C.
Amplicons were verified by capillary electrophoresis (QIAxcel, Qiagen, Germany), purified using ExoSAP-IT® PCR Clean-Up Reagent (Thermo Fischer Scientific, USA), and sequenced bidirectionally by Macrogen Europe (macrogen-europe.com). Obtained nucleotide sequences were then aligned and analyzed with SeqMan and EditSeq (Lasergene, DNASTAR) and compared with GenBank® reference sequences using BLASTn (blast.ncbi.nlm.nih.gov). The representative sequences were deposited in GenBank®.
Chromatograms were visually inspected for unresolved nucleotides. Sequences of 367 bp were then aligned and trimmed from primers using MEGA7 and the MUSCLE algorithm. Schistocephalus solidus (GenBank: OP586600) was used as an outgroup. The best-fit substitution model (HKY+G) was selected using JModelTest2, and a Bayesian Inference phylogenetic tree was constructed with the BEAST v2.7.7 package. Chain convergence and tree prior values were evaluated with Tracer v1.7.2. The consensus tree was estimated using TreeAnnotator and visualized in FigTree v1.4.4.
Results
Adult cestodes were identified in the small intestines of three golden jackals (1.5%) originating from three different locations in the central part of Continental Croatia (Figure 1). The Spirometra-positive animals were collected in Jastrebarsko (GPS: 45.662651, 15.679982) on September 5, 2022, in Gredani (GPS: 45.204596, 17.157740) on January 31, 2025, and in Sisak (GPS: 45.506042, 16.405342) on December 6, 2023. The morphological characteristics of the cestodes included an unarmed scolex with well-developed bothria and a narrow neck separating the scolex from the immature proglottids (Figure 2a). Under stereomicroscopy, both immature and mature proglottids were observed (Figures 2b, c). The proglottids contained multiple testes and vitelline glands in the lateral fields, with genital and uterine pores located medially. The genital pore was anterior to the uterine pore, and the uterus showed several convolutions, features consistent with S. mansoni description (7). The presence of a single cestode with distinct morphological characteristics was confirmed in each animal.
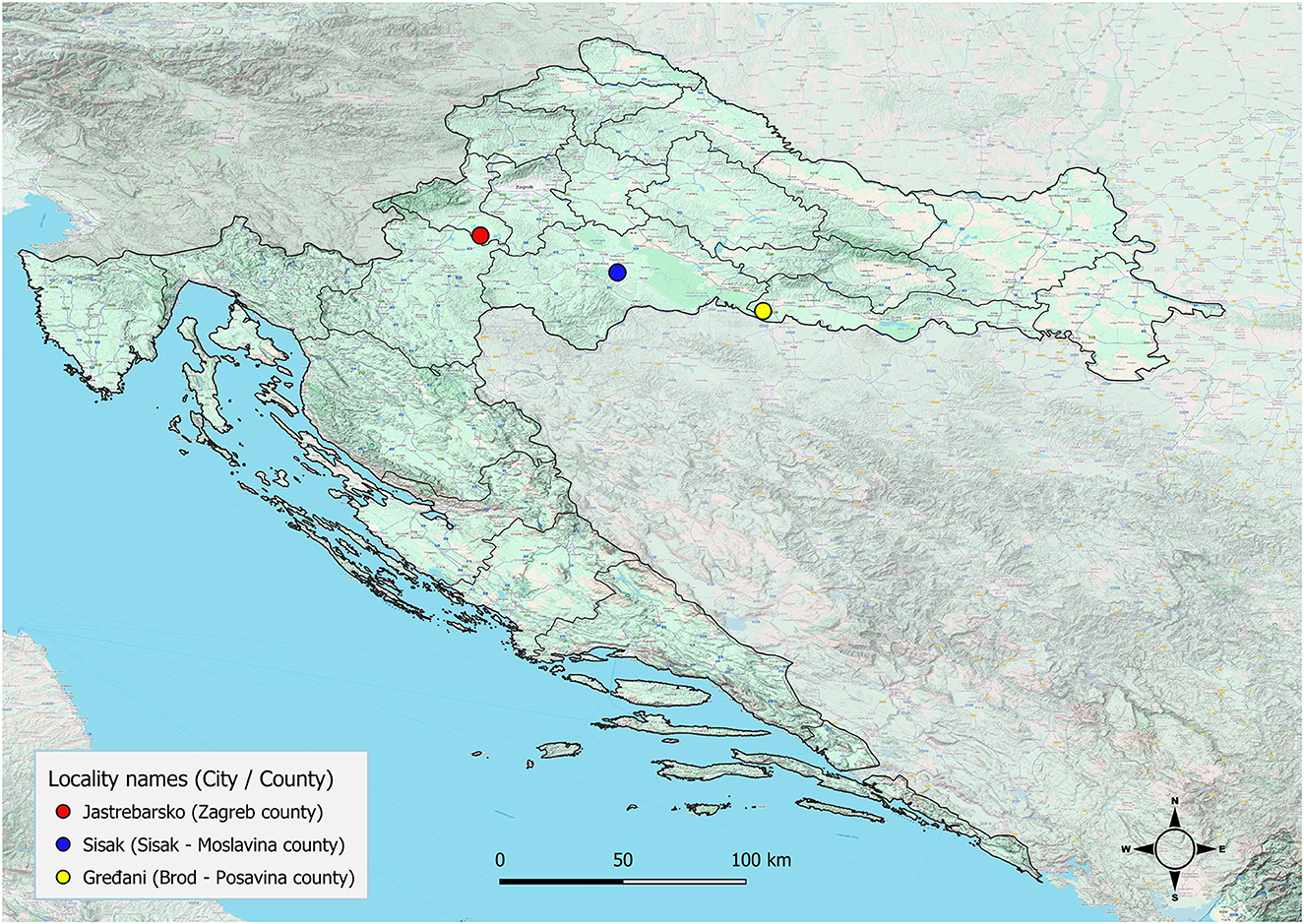
Figure 1. Geographic locations in Croatia where Spirometra mansoni was detected in. Figure was created with QGIS.
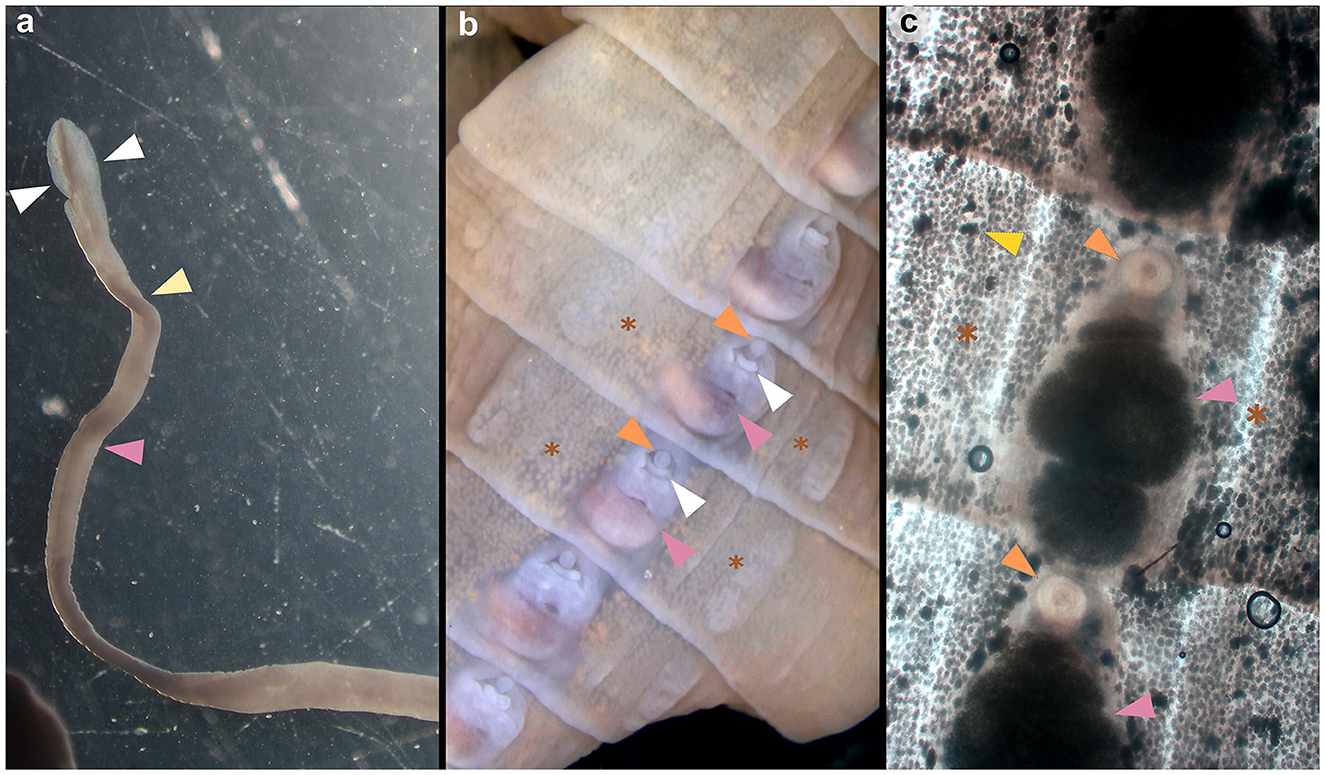
Figure 2. Morphological assessment of the scolex and proglottids of Spirometra mansoni collected from the golden jackal (Canis aureus) in Croatia. (a) Elongated scolex of an adult showing two bothria (marked with white triangles), the neck (yellow triangle) and the beginning of the strobila (pink triangle). (b) Mature proglottid with evident genital pore (marked with an orange triangle), uterine pore (white triangle), coiled uterus (pink triangle) and vitelline glands (brown asterisk). (c) Mature proglottid showing the genital pore (orange triangle), coiled uterus filled with eggs (pink triangle), testis (yellow triangle) and vitelline glands (brown asterisk).
Molecular analysis of the mitochondrial genes confirmed the identification of S. mansoni, with all three tested specimens exhibiting identical nucleotide sequences. The cox1 sequences showed 98.53% similarity with S. mansoni from Pelophylax esculentus (GenBank accession numbers MT131822 and MT131823), with 100% query coverage. Molecular analysis of the nad1 gene revealed sequence similarities ranging from 95.09% to 96.32% with S. erinaceieuropaei (GenBank accession numbers OM935779 and OM935776), 95.30% with S. ranarum (MH298844), and 95.71% with S. decipiens (MN121695), all with 100% query coverage. Phylogenetic reconstruction of the cox1 using Bayesian inference placed the Croatian sequences within the S. mansoni clade with a posterior probability value of 1.0, indicating strong statistical support (Figure 3). This clade included sequences from diverse geographical regions including Asia and Europe (Romania), confirming the wide distribution and relatively low inter-population variability of this species.
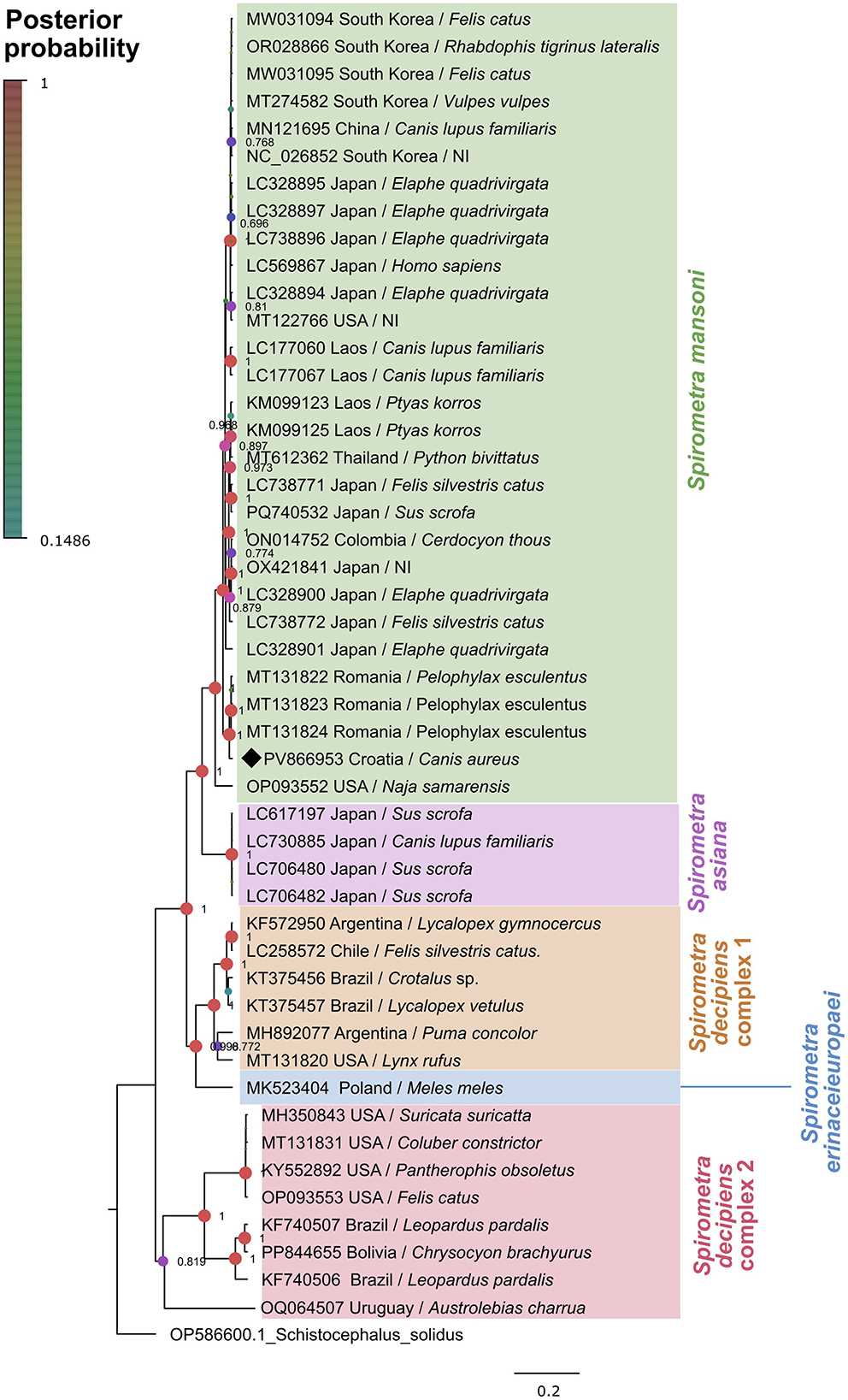
Figure 3. Phylogenetic analysis of cox1 sequences of Spirometra spp. (1,567 bp) from different hosts and geographical regions. Bayesian inference phylogenetic tree showing Spirometra spp. in different color blocks. The Spirometra mansoni sequence obtained this study is marked with a black diamond.
Discussion
This study presents the first confirmed molecular identification of S. mansoni in golden jackals in Croatia and, to our knowledge, in Europe. Gherman and Mihalca (22) cite a case report from 1889 describing the necropsy finding of S. mansoni (syn. Bothriocephalus mansoni) in a golden jackal in Italy (23). However, a golden jackal originated from Egypt and was imported into Italy. Furthermore, the identification was based solely on subcutaneous larvae and lacks morphological or molecular confirmation. The case is not included in the more recent and comprehensive reviews by Kuchta et al. (4, 7), and therefore, we consider the 1889 report to be unreliable. Although sparganosis and its etiological agents have been well documented in Asia and Africa, reports from Europe remain scarce and are largely based on morphological identification alone (4, 7). To date, S. mansoni has been molecularly confirmed in Europe only in water frogs from Romania (4). Our findings support recent recommendations advocating the integration of molecular and morphological methods to accurately delineate species boundaries and improve knowledge of Spirometra distribution (4, 7). Previous evidence of Spirometra spp. in Croatia was based solely on morphological re-examination of parasite eggs found in gray wolf feces (4, 24), without molecular confirmation. Since Spirometra spp. eggs are indistinguishable among species and morphology of mature proglottids alone is insufficient for reliable species confirmation, molecular typing remains essential for accurate identification at specie level (4, 7). Thus, this study represents the first genetically verified record of S. mansoni in a wild canid in the country.
The identification of S. mansoni in a free-ranging wild carnivore in Europe is unexpected, as S. erinaceieuropaei has previously been considered the only endemic species on the continent (4). The sequences clustered within the S. mansoni clade with strong posterior support and showed haplotype identity with strains from Asia and Romania, raising the question about possible introduction pathways, host mobility, and underestimated zoonotic potential in the region. The lack of nucleotide variation among the three Croatian isolates indicates haplotype homogeneity within the local population. This may reflect either a highly conserved lineage or a recent introduction of S. mansoni into the region; however, the limited sample size precludes definitive conclusions. Therefore, more extensive sampling including other carnivore species and populations from neighboring countries is required to better assess local genetic diversity.
The golden jackal is a highly adaptable and increasingly widespread carnivore in Europe (16, 25). Its opportunistic feeding habits, including possible predation or scavenging on amphibians and reptiles (26), which are known second intermediate hosts of Spirometra, make it a suitable definitive host. Central Croatia, a transitional ecological zone between the Pannonian lowlands and the Dinaric Alps (where the jackals were sampled), is rich in natural water sources, and high biodiversity, creating optimal conditions for complex parasitic life cycles. The role of golden jackal as both a reservoir and a bridge host for emerging parasites in Europe was already emphasized (22, 27). The expansion of jackals from Croatia eastward (toward Bulgaria) and westward (toward Italy) has also been genetically confirmed (28). These factors may facilitate the local maintenance and potential spread of S. mansoni to other suitable wild reservoirs. Like other mesocarnivores, golden jackals successfully inhabit human-dominated habitats, where they rely on human-subsidized food sources, including scavenging and occasionally preying on domestic animals (29, 30). This behavioral flexibility facilitates frequent contact with both humans and domestic animals. In Costa Rica, the possible circulation of S. mansoni between wild reservoirs (such as Canis latrans) and domestic animals (cats and dogs) has been described (8). Although the transmission pathways of S. mansoni are not yet fully understood, the presence of infected carnivores in close proximity to human settlements may carry potential public health implications that warrant further investigation.
Although S. mansoni has been associated with human infections in other regions, its zoonotic potential in Europe remains unclear. The presence of this parasite in an expanding and synanthropic carnivore such as the jackal warrants heightened awareness and additional surveillance. Our study highlights the importance of wildlife in maintaining the life cycle of zoonotic cestodes and the utility of molecular tools for reliable species identification. In addition to the complete cox1 gene sequence, we generated and deposited a new nad1 sequence of Spirometra mansoni in GenBank®. While previous sequences of nad1 associated with this species exist (although labeled under different names such as S. erinaceieuropaei, S. ranarum, or S. decipiens), our sequence contributes to the growing molecular reference dataset of S. mansoni, which is crucial for accurate phylogenetic comparisons and lineage resolution (4, 31). Future studies should include a broader range of potential definitive and intermediate hosts within the region, such as amphibians (e.g., frogs), reptiles (e.g., snakes), and domestic carnivores (e.g., cats), to elucidate the full transmission network of Spirometra spp. and assess possible spillover risks to humans.
Conclusions
This study provides the first molecular confirmation of S. mansoni in the golden jackal in Croatia and possibly Europe. The detection of this zoonotic cestode in a free-ranging wild canid expands current knowledge of Spirometra host range and distribution on the continent. These findings support the need for increased public and veterinary awareness, as well as targeted surveillance and education of professionals regarding the zoonotic potential of Spirometra spp. in Europe.
Data availability statement
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found in the article/supplementary material.
Ethics statement
The animal study was approved by Ethic committee of Croatian Veterinary Institute. The study was conducted in accordance with the local legislation and institutional requirements.
Author contributions
AŠ: Methodology, Writing – original draft, Investigation. EG: Writing – original draft, Methodology, Writing – review & editing, Investigation. AR: Investigation, Methodology, Writing – original draft, Writing – review & editing. MS: Writing – original draft, Writing – review & editing, Data curation. DŽ: Investigation, Methodology, Writing – review & editing, Writing – original draft. ŠN: Writing – review & editing, Investigation. DB: Investigation, Writing – review & editing. AH: Writing – review & editing, Methodology. RB: Methodology, Writing – review & editing, Conceptualization, Funding acquisition, Resources, Supervision, Writing – original draft.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This study was funded by the European Union (NextGenerationEU) and supported by the Ministry of Science and Education of the Republic of Croatia through project NPOO 5 at the Croatian Veterinary Institute (Molecular identification of nematodes, tapeworms, flukes and obligate myasis of domestic and wild animals, PARAKOD).
Acknowledgments
The authors express their sincere gratitude to technicians Kristina Skrbin, Ada Vilić, Petra Cikoš, Martina Pavić and Mario Maršić, which significantly contributed to this study.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The authors declare that no Gen AI was used in the creation of this manuscript.
Publisher's note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
1. Li MW, Song HQ, Li C, Lin HY, Xie WT, Lin RQ, et al. Sparganosis in mainland China. Int J Infect Dis. (2011) 15:154–6. doi: 10.1016/j.ijid.2010.10.001
2. Kuchta R, Scholz T, Brabec J, Narduzzi-Wicht B. Diphyllobothrium, diplogonoporus and Spirometra. In:Xiao L, Ryan U, Feng Y, , editors. Biology of Foodborne Parasites (1st edition). Boca Raton, FL: CRC Press (2015). p. 299-326.
3. Liu Q, Li MW, Wang ZD, Zhao GH, Zhu XQ. Human sparganosis, a neglected food borne zoonosis. Lancet Infect Dis. (2015) 15:1226–35. doi: 10.1016/S1473-3099(15)00133-4
4. Kuchta R, Kołodziej-Sobocińska M, Brabec J, Młocicki D, Sałamatin R, Scholz T. Sparganosis (Spirometra) in Europe in the molecular era. Clin Infect Dis. (2021) 72:882–90. doi: 10.1093/cid/ciaa1036
5. Kuchta R, Scholz T. Diphyllobothriidea. In:Caira, JN, Jensen K, , editors. Planetary Biodiversity Inventory (2008–2017): Tapeworms from Vertebrate Bowels of the Earth. University of Kansas, Lawrence, KS, USA (2017). p. 167–89. Natural History Museum, Special Publication No. 25. Available online at: https://hdl.handle.net/1808/24421
6. Scholz T, Kuchta R, Brabec J. Broad tapeworms (Diphyllobothriidae), parasites of wildlife and humans: recent progress and future challenges. Int J Parasitol Parasites Wildl. (2019) 9:359–69. doi: 10.1016/j.ijppaw.2019.02.001
7. Kuchta R, Phillips AJ, Scholz T. Diversity and biology of Spirometra tapeworms (cestoda: diphyllobothriidea), zoonotic parasites of wildlife: a review. Int J Parasitol Parasites Wildl. (2024) 23:100947. doi: 10.1016/j.ijppaw.2024.100947
8. Alvarado-Hidalgo I, Campos-Camacho J, Arguedas-Morales Y, Romero-Vega LM, Alfaro-Alarcón A, Anchia-Ureña G, et al. Molecular, morphological and histopathological evidence of Spirometra mansoni in wild and domestic animals from Costa Rica. Vet Parasitol Reg Stud Reports. (2024) 51:101030. doi: 10.1016/j.vprsr.2024.101030
9. Rojas A, Bass LG, Campos-Camacho J, Dittel-Meza FA, Fonseca C, Huang-Qiu YY, et al. Integrative taxonomy in helminth analysis: protocols and limitations in the twenty-first century. Parasit Vectors. (2025) 18:1–20. doi: 10.1186/s13071-025-06682-6
10. Bowman DD. Georgis Parasitology for Veterinarians. Philadelphia: W.B. Saunders (1999). p. 148–9.
11. Vettorazzi R, Norbis W, Martorelli SR, García G, Ríos N. First report of Spirometra (eucestoda; diphyllobothriidae) naturally occurring in a fish host. Folia Parasitol. (2023) 70:8. doi: 10.14411/fp.2023.008
12. Bazsalovicsová EC, Radačovská A, Lavikainen A, Kuchta R, Králová-Hromadová I. Genetic interrelationships of Spirometra erinaceieuropaei (cestoda: diphyllobothriidea), the causative agent of sparganosis in Europe. Parasite. (2022) 29:8. doi: 10.1051/parasite/2022009
13. Bagrade G, Králová-Hromadová I, Bazsalovicsová E, Radačovská A, Kołodziej-Sobocińska M. The first records of Spirometra erinaceieuropaei (cestoda: diphyllobothriidae), a causative agent of human sparganosis, in Latvian wildlife. Parasitol Res. (2021) 120:365–71. doi: 10.1007/s00436-020-06957-0
14. Trouwborst A, Krofel M, Linnell JDC. Legal implications of range expansions in a terrestrial carnivore: the case of the golden jackal (Canis aureus) in Europe. Biodivers Conserv. (2015) 24:2593–610. doi: 10.1007/s10531-015-0948-y
15. Krofel M, Giannatos G, Cirović D, Stoyanov S, Newsome TM. Golden jackal expansion in Europe: a case of mesopredator release triggered by continent-wide wolf persecution? Hystrix. (2017) 31:26–34. doi: 10.4404/hystrix-28.1-11819
16. Gaál DK, Heltai M, Sándor G, Schally G, Csányi E. Ecotones in the spotlight—habitat selection of the Golden Jackal (Canis aureus Linnaeus, 1758) in the agricultural landscapes of central Europe. Animals. (2025) 15:760. doi: 10.3390/ani15050760
17. Cirović D, Penezić A, Krofel M. Jackals as cleaners: ecosystem services provided by a mesocarnivore in human-dominated landscapes. Biol Conserv. (2016) 199:51–5. doi: 10.1016/j.biocon.2016.04.027
18. Hruševar D, Bakrač K, Miko S, Ilijanić N, Hasan O, Mamić M, et al. Environmental history in central croatia for the last two millennia – vegetation, fire and hydrological changes under climate and human impact. Pril Inst arheol Zagrebu. (2020) 37:117–64. doi: 10.33254/piaz.37.5
19. Brundić D, Curlin S, Deur M, Jardas I, Kerovec M, Lupert-Obradović S, et al. Biodiversity of Croatia. In:Kutle A, , editor. Pregled stanja biološke i krajobrazne raznolikosti Hrvatske sa strategijom i akcijskim planovima zaštite. Zagreb: Zavod za zaštitu okoliša i prirode (1999). p. 151.
20. Yanagida T, Matsuoka H, Kanai T, Nakao M, Ito A. Anomalous segmentation of Diphyllobothrium nihonkaiense. Parasitol Int. (2010) 59:268–70. doi: 10.1016/j.parint.2009.12.006
21. Trachsel D, Deplazes P, Mathis A. Identification of taeniid eggs in the faeces from carnivores based on multiplex PCR using targets in mitochondrial DNA. Parasitol. (2007) 134:911–20. doi: 10.1017/S0031182007002235
22. Gherman CM, Mihalca AD. A synoptic overview of golden jackal parasites reveals high diversity of species. Parasit Vectors. (2017) 10:1–40. doi: 10.1186/s13071-017-2329-8
23. Sonsino P. Studi e notizie elmintologiche. Atti della Societa Toscana di Scienze Naturali di Pisa. (1890) 7:99–114.
24. Hermosilla C, Kleinertz S, Silva LM, Hirzmann J, Huber D, Kusak J, et al. Protozoan and helminth parasite fauna of free-living croatian wild wolves (Canis lupus) analyzed by scat collection. Vet Parasitol. (2017) 233:14–9. doi: 10.1016/j.vetpar.2016.11.011
25. Hatlauf J, Bayer K, Trouwborst A, Hackländer K. New rules or old concepts? The golden jackal (Canis aureus) and its legal status in Central Europe. Eur J Wildl Res. (2021) 67:25. doi: 10.1007/s10344-020-01454-2
26. Lange PN, Lelieveld G, De Knegt HJ. Diet composition of the golden jackal Canis aureus in south-east Europe–a review. Mamm Rev. (2021) 51:207–13. doi: 10.1111/mam.12235
27. Orioles M, Fabbri D, Miani G, Pesaro S, Dorigo L, Bregoli M, et al. Double trouble: co-infection of Angiostrongylus vasorum and Dirofilaria immitis in golden jackal (Canis aureus) in Friuli Venezia Giulia, Italy. Int J Parasitol Parasites Wildl. (2024) 24:100969. doi: 10.1016/j.ijppaw.2024.100969
28. Fabbri E, Caniglia R, Galov A, Arbanasić H, Lapini L, Bošković I, et al. Genetic structure and expansion of golden jackals (Canis aureus) in the north-western distribution range (croatia and eastern Italian Alps). Conserv genet. (2014) 15:187–99. doi: 10.1007/s10592-013-0530-7
29. Yom-Tov Y, Ashkenazi S, Viner O. 1995: cattle predation by the golden jackal Canis aureus in the Golan Heights, Israel. Biol Conserv. (1995) 73:19–22. doi: 10.1016/0006-3207(95)90051-9
30. Cirović D, Penezić A, Milenković M, Paunović M. Winter diet composition of the golden jackal (Canis aureus L., 1758) in Serbia. Mamm Biol. (2014) 79:132–7. doi: 10.1016/j.mambio.2013.11.003
Keywords: Canis aureus, cox1, nad1, Croatia, Spirometra mansoni
Citation: Šikić A, Gagović E, Rojas A, Sindičić M, Žilić DJ, Naletlić, Balić D, Hodžić A and Beck R (2025) First molecular identification of Spirometra mansoni in the golden jackal (Canis aureus) in Croatia. Front. Vet. Sci. 12:1629099. doi: 10.3389/fvets.2025.1629099
Received: 15 May 2025; Accepted: 23 July 2025;
Published: 11 August 2025.
Edited by:
Calin Mircea Gherman, University of Agricultural Sciences and Veterinary Medicine of Cluj-Napoca, RomaniaReviewed by:
Roman Kuchta, Academy of Sciences of the Czech Republic (ASCR), CzechiaJan Brabec, Czech Academy of Sciences, Czechia
Aida Vafae Eslahi, Qazvin University of Medical Sciences, Iran
Copyright © 2025 Šikić, Gagović, Rojas, Sindičić, Žilić, Naletlić, Balić, Hodžić and Beck. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Relja Beck, YmVja0B2ZWluc3QuaHI=