 Morteza Naderi1*†
Morteza Naderi1*† Rupert Palme2†
Rupert Palme2† Kelly Yarnell3†Emrah Çoban4Ayşegül Karaahmetoğlu Çoban5Josip Kusak6,7,8
Kelly Yarnell3†Emrah Çoban4Ayşegül Karaahmetoğlu Çoban5Josip Kusak6,7,8 Çağan H. Şekercioğlu1,4,7†
Çağan H. Şekercioğlu1,4,7†- 1Department of Biology, Faculty of Sciences, Sakarya University, Sakarya, Türkiye
- 2Experimental Endocrinology, Department of Biological Sciences and Pathobiology, University of Veterinary Medicine, Vienna, Austria
- 3School of Animal and Environmental Sciences, Nottingham Trent University, Nottingham, United Kingdom
- 4KuzeyDoga Society, Kars, Türkiye
- 5Division of Clinical Sciences, Department of Wild Animal Diseases, Faculty of Veterinary Medicine, Balıkesir University, Balıkesir, Türkiye
- 6Veterinary Biology Unit, Faculty of Veterinary Medicine, University of Zagreb, Zagreb, Croatia
- 7School of Biological Sciences, University of Utah, Salt Lake City, UT, United States
- 8College of Sciences, Koc University, Istanbul, Türkiye
Brown bears (Ursus arctos) are a keystone species vital for maintaining ecological balance in northeastern Türkiye. However, increasing human activities—such as logging, agriculture, and recreation—pose significant threats to their conservation. It is therefore crucial to assess how these specific anthropogenic pressures influence bears’ physiological stress responses to inform effective conservation strategies. Our hypothesis that increased human activity would correlate with elevated stress markers in bears was tested by collecting blood serum samples from 50 free-ranging bears during live capture. Blood cortisol levels and fecal cortisol metabolites were measured to assess stress responses. We also employed camera trap surveys to quantify human activity levels across different habitat patches, calculating a Relative Abundance Index (RAI). Statistical analyses, including correlation and regression models, were used to assess relationships between cortisol measures, habitat features, and human presence. The study revealed an inverse correlation between bear mass and blood cortisol levels and a significant relationship between fecal cortisol metabolites and human presence, as quantified through camera trap data. These findings highlight the significant impact of human disturbances on bear stress physiology, the urgent need for effective conservation measures to minimize human-wildlife conflicts and support the long-term viability of bear populations in Türkiye. These findings highlight that fecal cortisol metabolites serve as reliable, non-invasive indicators of stress in free ranging brown bears, enabling large-scale monitoring to identify habitat disturbance hotspots, assess the effectiveness of protected areas, and inform targeted management actions to minimize human-wildlife conflicts and enhance habitat quality.
1 Introduction
Brown bears (Ursus arctos) are keystone predators and flagship species in many ecosystems, including northeastern Türkiye, where their presence indicates healthy forest and mountainous habitats (1). They play a vital ecological role by regulating prey populations and maintaining habitat diversity (2). Due to their keystone ecological role and cultural significance, brown bears are often a focal species in community-based conservation programs, encouraging local engagement in habitat protection and conflict mitigation. However, their populations face numerous threats in Türkiye, including habitat fragmentation, human-wildlife conflicts (3), and habitat degradation driven by logging, infrastructure development, and increased human presence (3, 4). These ongoing pressures threaten the long-term viability of Brown bears, underscoring the need for effective conservation strategies that incorporate biological monitoring, welfare, and stress assessment. Stressful situations, particularly those caused by human activities, can both directly and indirectly impact the long-term survival of wild species. This highlights the importance of ecological studies that focus on more vulnerable groups. Gaining insight into how these species respond and adapt to environmental shifts is essential for effective conservation efforts (5). Physiological indicators of animal welfare have been investigated among domestic animals (6, 7) animals in captivity (8), and rehabilitation centers, but this can be much more challenging while aiming for free-ranging animals (9). Among these physiological indicators, cortisol is the primary hormone used to assess stress responses in both domestic and wild animals due to its well-established role in the hypothalamic–pituitary–adrenal (HPA) axis.
In the context of welfare in wild, free-ranging animals, stress is not limited to direct threats or adverse events, but can also result from environmental factors such as habitat disturbance, species interactions, and human activity (9). They can include any species interaction, encounters with predators or rivals, and potentially beneficial interactions such as finding a mate, participating in social play, or engaging in vigorous physical activities. Anthropogenic sources, including human activity in wildlife habitats, can provide stressful conditions for wildlife, too (10).
In bears, blood glucocorticoids (serum GCs) have been measured as an indicator of stress [for an extensive review, see Babic et al. (11), Davenport et al. (12), Malcolm et al. (13)]. These hormones have also been utilized to assess stress responses related to capture techniques and to evaluate baseline levels specific to different bear species. Tryland et al. (14) stated that elevated cortisol levels indicated heightened stress in wild bear populations. Despite the broad use of blood glucocorticoids (GCs) to represent short-term stress status, studies also highlight high variability in cortisol levels depending on stimuli such as capture and handling, as well as factors like the time of day [referred to as “point” samples by (15)], therefore, measuring serum cortisol is not practical for assessing both short- and long-term stress in wildlife, making non-invasive techniques such as fecal cortisol metabolite analysis the preferred approach for studying stress in free-ranging animals (12, 16, 17). In conclusion, to evaluate the long-term stress state of free-ranging animals such as brown bears, it is essential to use reliable non-invasive methods that do not require animal handling or subject the animals to additional stressful procedures during capture and tranquilization. Such methodologies are critical for shaping conservation strategies in wildlife and free-ranging animals. These approaches enable frequent and cumulative physiological measurements with minimal, if any, disturbance to animals. Among them, measuring fecal cortisol and corticosterone metabolites (FCMs) has proven especially useful (see Palme (18) for a comprehensive review and evaluation of all previously published papers) (but for example see also: Hein et al. (19); Hunninck et al. (20); Babic et al. (11)).
FCM analyses in wild animals have been utilized to evaluate a range of stressors, including environmental, ecological, and anthropogenic factors. Natural environmental variables such as vegetation cover, prey availability, and weather conditions (e.g., temperature and precipitation) can significantly impact stress hormone levels, as they directly affect foraging success, reproduction, and overall habitat quality. Recent research on brown bears has demonstrated that physiological stress indicators such as blood cortisol and fecal cortisol metabolites (FCMs) reliably reflect responses to environmental and anthropogenic stressors (17). For example, Piñero et al. (17) conducted an ACTH challenge on unacclimated captive brown bears, illustrating the relationship between serum cortisol and FCM concentrations and highlighting the impact of human activity on stress hormone levels in natural bear populations. Hunninck et al. (20) reported that environmental factors like vegetation cover and quality can significantly influence FCM levels in wildlife, reflecting annual variations in food resources and habitat conditions impacting stress responses in species like impala (Aepyceros melampus) (20). Similarly, in red deer (Cervus elaphus) and chamois (Rupicapra rupicapra), precipitation and temperature were found to significantly influence FCM concentrations, indicating that seasonal environmental changes can modulate stress responses (21). FCM studies have also been carried out on free-living mountain hares (Lepus timidus) (22), capercaillies (Tetrao urogallus) (23), red deer (Cervus elaphus) and roe deer (Capreolus capreolus) (24), Alpine chamois (Rupicapra rupicapra) (25), Geoffroy’s spider monkeys (Ateles geoffroyi) (26), and Mexican gray wolves (Canis lupus) (27).
In some cases, human-related factors like habitat fragmentation, noise, and direct human presence have been shown to induce higher stress levels than natural ecological pressures, such as predator presence (24). These studies suggest that the impact of human activities can overshadow or interact with natural stressors, influencing wildlife behavior and physiology in complex ways. However, the length of exposure to human-induced disturbances can be significant in influencing the magnitude and duration of stress responses in wildlife. For example, a study on chipmunks (Tamias striatus) in urban habitats showed that they experience less stressful conditions than their nature-dwelling counterparts (28). Rangel-Negrin et al. (29) found a positive relationship between FCMs and environmental disturbances, with the lowest amounts of FCMs found among individuals in protected areas where the anthropogenic disturbances were the lowest (29). The authors also reported that habitat fragmentation can be a long-term stressor for spider monkeys. Moreover, anthropogenic activities can also exert a context-dependent effect on animals’ stress physiology (22, 30). However, studies on large carnivores’ welfare have focused less on the impact of habitat fragmentation, habitat patch size, seasonality, and human activity, but have focused on captive animals (4, 29–32).
Our study provided an ideal opportunity to explore physiological stress indices in free-ranging brown bears across a human gradient, which can then be compared to samples from rehabilitation centers in future studies, to find verified stress indices and inducing factors. We explored the relationship between serum GCs and FCMs and whether higher human activity, such as logging, would impact animals’ stress levels as measured via FCMs. This study aims to test the hypothesis that increased human activity correlates with elevated stress indicators in free-ranging brown bears, and to evaluate whether fecal cortisol metabolites serve as reliable non-invasive biomarkers of long-term stress in natural habitats. This research should advance our understanding of the spatial and human-induced factors that affect the stress physiology of free-ranging brown bears, and the findings can be applied to enhance their welfare.
2 Methods
2.1 Study area
Sarıkamış forests- Allahuekber Mountains National Park in north-eastern Türkiye is home to Brown bears (Ursus arctos), Gray wolves (Canis lupus), and Caucasian lynxes (Lynx lynx dinniki) (33, 34) with a relatively high degree of large carnivore-related human-wildlife conflict (34). Villagers inhabiting areas around the forests and organizations as forestry management, routinely use these fragmented Scotch pine forest patches with a total area of 338.5 km2 for logging (1) (Figure 1). Livestock herding, recreation (picnicking), and wild herb and mushroom collection create a high level of human activity, except during the harsh winter season (35). The ambient temperature usually does not fall below 20 °C during the warm season (May–October) but reaches levels lower than −10 °C during the cold months (November–April). Mixed beach-fir forest, mostly Fagus sylvatica, Abies alba, Picea abies, Quercus sp., Castanea sativa, and Pinus sp., constitute the forest community type in the study area (36). Utilizing the Relative Abundance Index (RAI) from camera trap surveys, we quantified human activity intensity within each patch, correlating these findings with physiological stress indicators in bears. The Relative Abundance Index (RAI) is a commonly used metric in camera trap studies that estimates the activity or detection rate of animals or humans within a given area, standardized by sampling effort. It is calculated by dividing the number of detection events at a station by the total trap days and multiplying by 100, providing detections per 100 trap days as an indicator of relative activity levels. Villagers around these forests engage in activities like logging, livestock herding, recreation, and wild herb collection, barring the severe winter months. An open garbage dump is located across the bear’s distribution at the study area and affects behavioral patterns in bears, including seasonal migration habits (35). There is considerable human-wildlife conflict at the garbage dumps, as bears are often drawn closer to human settlements due to the readily available food resources, leading to increased interactions, potential conflicts, and sometimes road mortalities. The Sarıkamış area provides a unique case for substantially advancing brown bear welfare and stress indices.
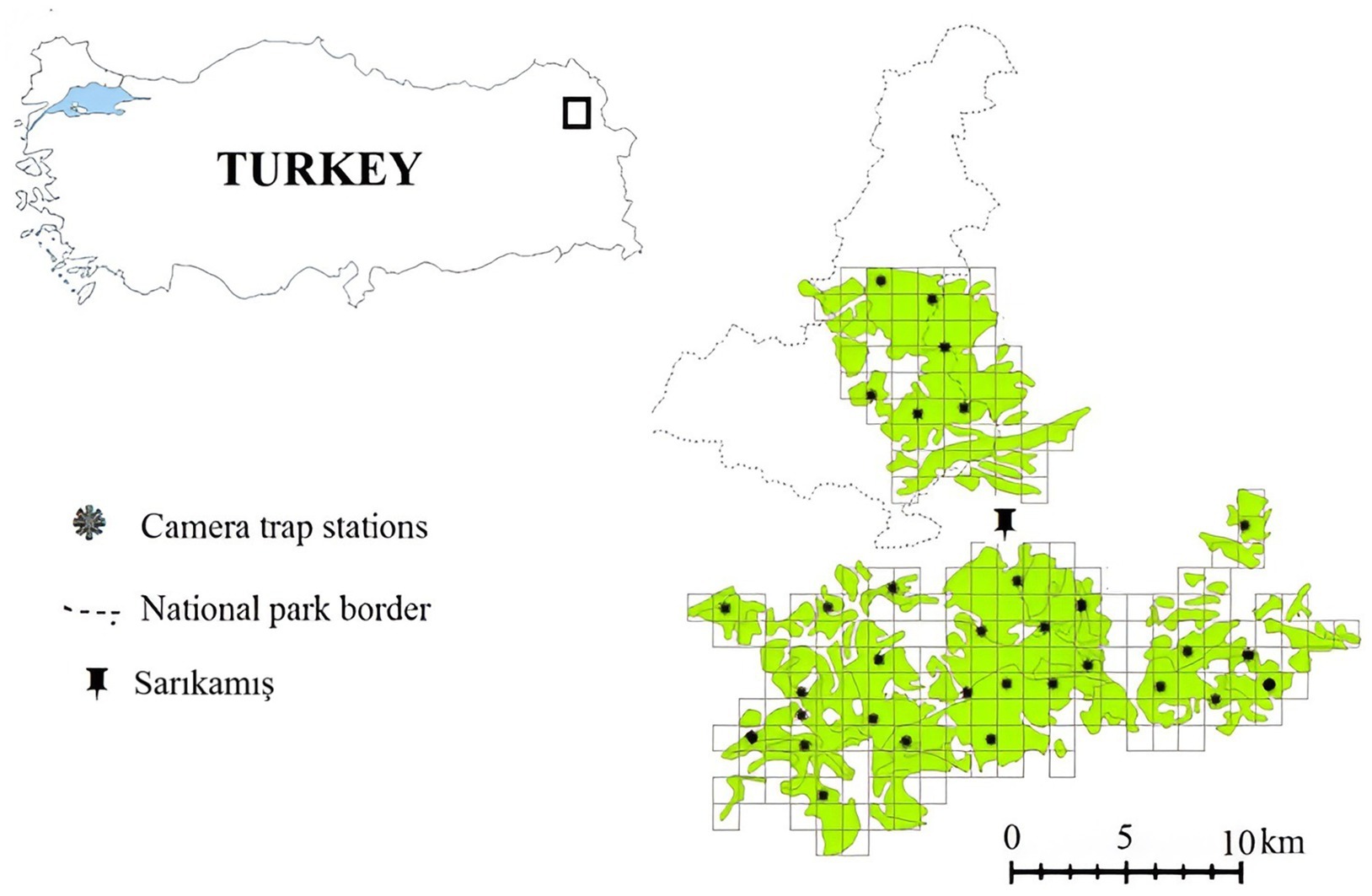
Figure 1. Study area and the location of the camera trapping stations. The black points show the camera traps.
2.2 Camera trapping
A large carnivore study and conservation management project has been performed in the study area during the past 13 years, led by the KuzeyDoğa Society (KD). We used data from camera trapping surveys from 2018 to 2023 to quantify the human activity in the area. During this period, 28–42 cameras (Reconyx Ultrafire XR6 and Reconyx PC900) were deployed systematically throughout the study area, divided into two square km grids, ensuring equal distribution of the cameras in each habitat. A total of 75,000 trap days have been done to collect data during the 6 years. Camera stations were set along roads and intersections of minor forest roads and wildlife trails without using any bait or attractant (37). We installed all the cameras 2 m high in the trees. Cameras were checked regularly every month. Based on Kays et al. (38), all cameras were set to take a series of five photos per trigger without any delay, with medium sensitivity and a 30-s sensor break between series (38). During this study, cameras were routinely shifted based on a uniform random pattern (39) to different study grids (generated in ArcGIS software ver. 10.3.1, ESRI, Redlands, CA) to cover most grid cells (4). To assess the level of human activity within the study area, we utilized the Relative Abundance Index (RAI). This approach allows for comparison across different locations and periods, accounting for variations in sampling effort. In our study, the RAI served as a proxy for human activity intensity because a higher number of detections at a station indicated increased presence or activity of humans in that area. This standardized measure was then used to explore relationships between human disturbance and stress hormone levels in brown bears (4).
2.3 Sample collection
This research forms a segment of an ongoing conservation and population biology initiative focused on the large carnivores of northeastern Türkiye, including brown bears, wolves, and lynxes, spearheaded by the KuzeyDoğa Society since 2006 (40). Bears were captured using Aldrich snares in a “European cubby” (41). A GSM-equipped camera trap was also installed to send instant photos of the possible capture, and the intervention time was then reduced to around 20 min for handling the captured bear. We also used GSM-equipped alarms to send signals on any triggering of traps. In this study, we used a tranquilization protocol involving a combination of Zoletil 100 and Domitor. The dosage was 2.5 mg/kg of Zoletil 100 and 0.05 mg/kg of Domitor. This combination was prepared such that 0.25 mL of the mixed solution was used for each 10 kg of body mass (42). Target animals were then equipped with Vectronic Aerospace GPS-GSM/Iridium collars (Vectronic Aerospace GmbH, Berlin, Germany) with two-axis activity sensors, which continuously recorded the acceleration and stored the values within a range of 0–255 in five-minute intervals. Standard capture forms were filled out for each captured animal, and all necessary data for this study, such as sex, age, and mass, have been recorded. Blood samples were taken from the femoral vein (Vena femoralis) and stored in the fieldwork station’s cool box for less than 1 h and centrifuged at 3,000 rpm for 5 min. Separated plasma samples were stored in a −20 °C freezer for laboratory analysis.
In all cases, captured bears’ fresh scat samples were found around the cubby and immediately transferred to the −20 °C freezer. Collected scats from the mentioned cubbies were very fresh, less than 30 min after defecation. The other scats across the habitats were also selected from the fresh ones that had been defecated during the previous night. In some places, such as the garbage dump, we also collected less than 10 min old scats. As a unique study on free-ranging brown bears across the world, we collect the largest sample size of bears’ blood (n = 50) and 23 scat samples known to belong to the same individuals from which the blood samples were taken. Those scats were categorized based on the animal’s sex since they were collected along with the animal’s live capture while monitored by the real-time cameras installed at the cubbies. For the rest of the scat samples (n = 28) the sex could not be detected as they were collected in different parts of the habitat. We also followed collared bears to exact locations, including dens, daily beds, and resting areas, allowing further fresh scats to be collected. Since the study area has an open garbage dump, some collared bears spend their time foraging in the garbage dump for food. Therefore, by visiting the garbage dump, we could spot the collared bears and collect fecal samples from this area while also determining the sex of the collected scats. This allowed us to explore the impact of anthropogenically driven feeding areas on animal stress status as the feeding area was often shared with dogs that vocalized often. To control for potential seasonal effects, samples were collected throughout only the warm season (May to August), and sampling was evenly distributed across different months of this season; therefore, the seasonal effects have not been considered in this study. All procedures aimed to minimize temporal variability, but inherent seasonal influences on hormone levels were acknowledged as a limitation. Table 1 indicates the number of samples with more details.
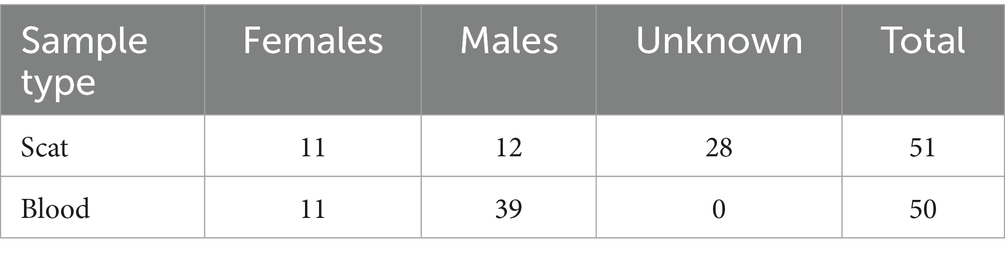
Table 1. Collected samples based on type and sex.
2.4 Steroid analyses
For FCMs, we weighed 0.5 g of each wet scat and then added 5 mL of 80% methanol to the sample. Each falcon tube was vortexed by hand for 2 min, and all samples were centrifuged for 15 min at 2,500 g. Finally, 0.5 mL of supernatant was transferred to new tubes and dried in an oven at 40–50° C for 24 h (43). For serum samples, we took 0.5 mL of serum and added 5 mL of diethyl ether and kept it in the 15 mL falcon tubes, shook it by hand several times, and centrifuged it (15 min at 2,500 g). Falcon tubes were frozen for 2–3 h. Then, gently, we transferred the ether phase to new falcon tubes using micropipettes. Afterwards, the ether phase was dried and redissolved in an assay buffer. FCMs were determined with a cortisol enzyme immunoassay (for details, see Palme and Moestl (44)), which has been successfully validated for brown bears (45). The same cortisol assay was used for the serum extracts.
2.5 Statistical analyses
First, normality of the datasets (serum cortisol, fecal cortisol metabolites) was assessed using the Shapiro–Wilk test for all samples and relevant subgroups (e.g., by sex, mass). For normally distributed variables, parametric tests such as Pearson correlation and t-test were employed; otherwise, non-parametric alternatives like Spearman correlation and Mann–Whitney U test were used. To determine the relationship between FCMs and human activity, the Spearman’s rank correlation coefficient was calculated. The correlation between blood cortisol and fecal cortisol metabolite concentrations within the same individuals was also assessed using Spearman’s correlation. Differences in cortisol levels between sexes were examined using the Mann–Whitney U test, due to small sample sizes and non-normal distribution of subgroup data.
Comparisons of FCM levels among different habitat patches were conducted with the Mann–Whitney U test, as the data did not meet parametric assumptions. All significance levels were set at p < 0.05. Assumptions of each test were checked using residual plots and appropriate diagnostics. Analyses were performed using R, with mentioned packages like ggplot2 and lme4 for statistical modeling and data visualization, and significance levels were set at p < 0.05. Data normality and assumptions for each analysis were checked and addressed accordingly to ensure the reliability of the results. Additionally, we considered the potential influence of covariates, such as age and mass, in the analyses to enhance the accuracy of our interpretations.
2.6 Ethical declarations
All animal procedures followed the Kafkas University local ethical committee for animal experimentation (KAÜ HADYEK) guidelines under KAÜ HADYEK/ 2018-050 and KAÜ-HADYEK/ 2021-083 permission number. Capturing bears was conducted under strict ethical guidelines approved by Turkey’s Department of Nature Conservation and the Ministry of Agriculture and Forestry (Permit No. 72784983-488.04-114100 and E-21264211-288.04-1602322). The procedure was justified by the necessity to obtain biological samples (blood and feces) from individual bears for stress assessment, which cannot be reliably achieved through non-invasive means alone in this context. During immobilization, bears were monitored continuously, and vital parameters were checked regularly. Bears were kept under supervision until full recovery from anesthesia.
3 Results
Blood cortisol levels ranged from 2.3 to 58.5 (median: 17.9) ng/ml, and FCM concentrations from 1.2 to 16.1 (median: 4.5) ng/g. These ranges come from the whole data, including blood and scat samples taken from the same individuals. There was a relationship between levels of blood cortisol and its metabolites in scat samples of the same individuals (r = 0.61, p < 0.05) (Figure 2). To verify the assumption of normality for our dataset, we conducted the Shapiro–Wilk test across variables. The pooled data, not classified by age or sex, demonstrated a p-value of 0.22, indicating a normal distribution. We investigated the correlation between animal mass and blood cortisol levels using Pearson’s correlation coefficient. Our analysis revealed a significant inverse correlation (r = −0.41, p = 0.04, n = 50). This suggests heavier animals tended to have lower cortisol concentrations.
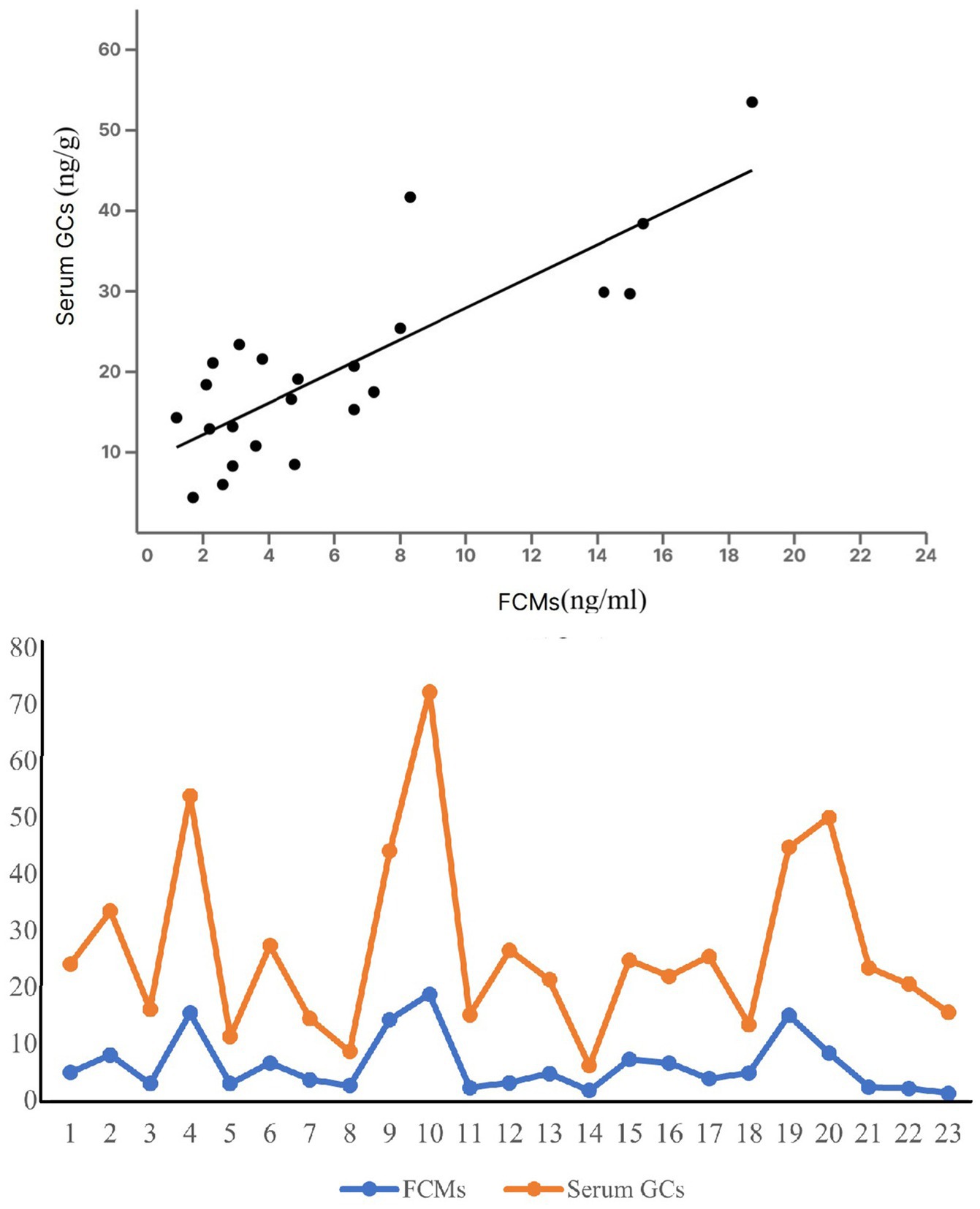
Figure 2. A linear relationship between FCM and blood cortisol concentration in the same individuals.
We conducted a Mann–Whitney U test to evaluate the differences in blood cortisol levels between male and female bears. The analysis did not reveal a significant difference in cortisol levels between male and female bears. Due to the presence of tied values—where multiple observations share the same measurement—in our data, the Mann–Whitney U test was conducted with a correction for ties, which adjusts the test statistic accordingly. The results revealed a U statistic of 159 and a Z value of −0.9674. The analysis failed to reject the null hypothesis, as the p-value was 0.3 (U = 159, Z = −0.9674, p = 0.3). This indicates no statistically significant difference in blood cortisol levels between the sexes. The computed standardized effect size was small (0.14), and the observed common language effect size was 0.4, suggesting a low probability (40%) that a randomly selected cortisol level from a male bear would exceed that from a female bear. In this study, we found a good correlation between measured FCMs and human activity (r = 0.71, p < 0.01; Figure 3), which was estimated from the camera trapping project (calculated as RAI). We found lower mean FCM levels in samples collected from open garbage dumps than in the habitat patches. However, differences were not statistically significant (Z equals 1.34). There was no significant relationship between FCMs and sex/age (p > 0.05).
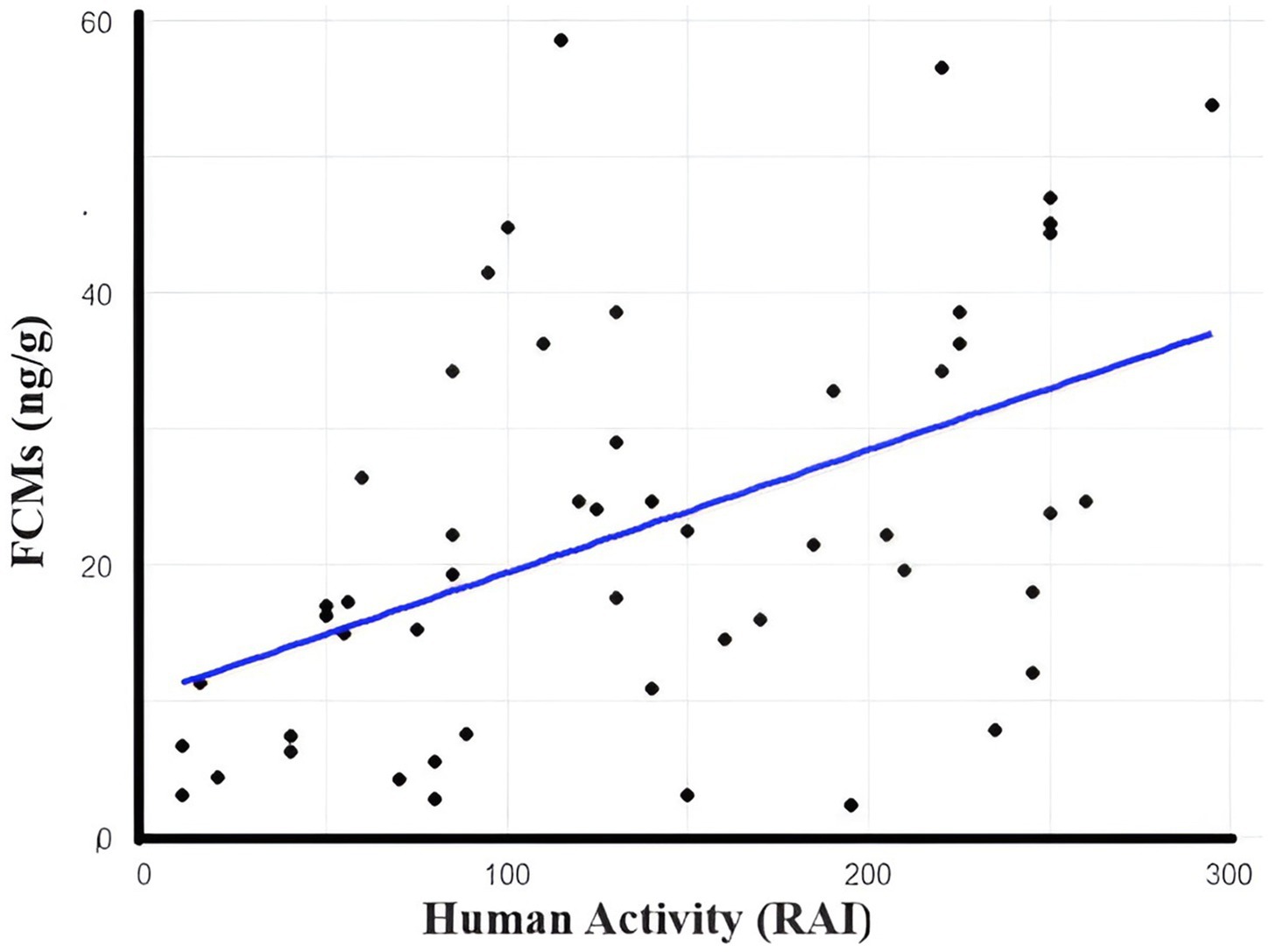
Figure 3. Human activity (RAI) and stress levels of brown bears. This graph indicates that the animals suffer from more stressful conditions in areas with higher human activity.
4 Discussion
Despite recent advances in understanding stress physiology in free-ranging mammals, there remains a significant gap in knowledge regarding the welfare and stress conditions of free-roaming animals such as large carnivores that inhabit large, unbounded areas and encounter a wide range of environmental challenges (9). Specifically, for large carnivores such as brown bears, data on long-term stress indicators are limited, particularly in their natural habitats where multiple anthropogenic and ecological factors intersect. In this study, we explored blood serum cortisol and FCM concentrations utilizing the largest sample size from free-ranging brown bears at their natural habitats, since most similar studies have been carried out on captive brown bears (17). Both measures were correlated, and we found an inverse correlation between bear mass and blood cortisol levels and a significant relationship between FCMs and human presence, as quantified through camera trap data.
We believe that our findings can contribute to advancing research of this type, particularly focusing on large carnivores like brown bears. Our samples were highly valuable and unique because we were able to collect both blood and scat samples from the same individuals in their natural habitat. Therefore, we were able to evaluate the relationship between blood cortisol and FCM concentrations, which has also been done in a previous study (17). In this research, we found that FCMs can be a suitable index for free-ranging bears’ welfare and stress status across their natural habitats, because their levels are not influenced by capture stress. Blood cortisol concentrations can be highly variable about the environmental conditions (15) and needs animal live capture and immobilization, which can be highly stressful and expensive (17).
Previous studies suggest that body condition can influence an animal’s stress response (46, 47). Lower cortisol levels in heavier bears suggest that larger, potentially more dominant or better-conditioned bears might experience lower physiological stress, possibly due to better access to resources or superior territory. Larger bears may also be more successful in securing high-quality habitats, which could buffer them from certain stressors such as human disturbance. Conversely, smaller or subordinate individuals might experience higher stress levels due to increased competition or limited access to resources, which could be reflected in increased cortisol. However, this relationship warrants caution; higher body mass may also be associated with other factors like reproductive status or age, which we did not fully control for. Therefore, while a larger size might suggest better access to resources or higher social standing, it’s important to recognize that the relationship between size and stress is complex and may involve various ecological and behavioral factors. Future studies incorporating detailed habitat use and social hierarchy data could provide deeper insights into how size and territoriality modulate stress responses in brown bears.
The lack of significant relationships between FCM or blood cortisol levels with age or sex highlights the complexity of stress responses in wildlife, which has also been confirmed by previous studies. For example, Pinero et al. (17) also reported that there was no relationship between sex and both blood cortisol and FCM concentrations. However, this does not mean that the situation can be the same among other large mammals. For example, an opposite finding has been reported for Coyotes (Canis latrans) (48). These results suggest that factors other than age and sex, such as environmental conditions and individual health status, may play more crucial roles in determining stress hormone levels in brown bears (49, 50). Human activity emerged as a critical factor influencing bear stress levels. The significant relationship between human presence (as measured by RAI) and higher FCM concentrations indicates that bears in areas with higher human activity experienced increased stress. This finding is consistent with other studies documenting the impact of anthropogenic disturbances on wildlife stress physiology (24, 51). Human activities such as logging, livestock herding, and recreational activities can create disturbances that elevate stress hormones in wildlife (22).
Other studies indicated a relationship between FCMs and long-term stressful conditions, such as anthropogenic activities and stress conditions inferred from FCMs. For instance, Asiatic black bears (Ursus thibetanus) had higher levels of FCMs outside nature reserves with higher human activities (13). Nevertheless, Babic et al. (11) indicated that FCMs may not accurately reflect chronic or long-term stress or the impact of habitat conditions on bear welfare due to various factors such as adaptations, reproduction status, hibernation, etc.
While not statistically significant, comparing samples from garbage dumps and natural habitats provided valuable insights. The lower mean FGM levels in garbage dump samples could be attributed to the availability of accessible food resources, reducing the need for bears to engage in energy-intensive foraging activities. However, the lack of statistical significance suggests that other factors, such as the presence of dogs and humans, may offset the potential stress-relieving effects of accessible food sources (35, 52). Bears exposed to higher levels of human traffic exhibited increased cortisol concentrations, indicating heightened physiological stress (52). Roads and human presence can fragment habitats and create barriers to movement, leading to increased stress and decreased habitat quality for wildlife (3, 4). This study demonstrates that fecal cortisol metabolites are reliable, non-invasive indicators of long-term stress in free-ranging brown bears, closely correlating with serum cortisol levels. Our findings highlight the impact of human activities on bear physiology, emphasizing the importance of minimizing disturbances and habitat fragmentation. Incorporating stress biomarkers into conservation strategies can enhance monitoring efforts, identify stress hotspots, and support long-term population sustainability. Future research should expand spatial and seasonal sampling, include behavioral data, and examine cumulative effects of multiple stressors, aiding the development of adaptive management plans that prioritize both species welfare and ecosystem resilience.
4.1 Study limitations
This approach provided valuable insights into how these stress markers relate under real-world conditions. However, several limitations must be acknowledged. First, the uncontrolled conditions inherent to wild environments—such as variable diets, fluctuating weather patterns, and differences in individual activity levels—may influence hormone levels independently of human disturbance or other stressors. Additionally, physiological measures like cortisol are subject to natural circadian and seasonal variations (18), which can complicate interpretation when sampling occurs at different times or seasons without precise control. Furthermore, factors such as animal age, reproductive status, health condition, and prior exposure to stressors were not fully controlled or standardized in our sampling process. These variables can potentially influence hormone levels and confound the relationship between observed stress responses and environmental factors. Finally, as with all field-based studies, the inability to manipulate external variables limits the extent to which causality can be established. Despite these limitations, we believe our findings provide meaningful contributions toward understanding the stress physiology of large carnivores in their natural habitats and can serve as a foundation for future, more controlled research. We suggest that subsequent studies incorporate repeated measures across seasons and habitats, alongside controlled assessments of diet and activity budgets, to better isolate the effects of specific stressors. Moreover, integrating GPS movement data and behavioral observations could help contextualize hormonal fluctuations with behavioral responses, providing a more comprehensive understanding of animal welfare under varying environmental conditions.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Ethics statement
The animal study was approved by Türkiye’s Department of Nature Conservation and National Parks and the Ministry of Agriculture and Forestry for granting the permit for this research (No. 72784983-488.04-114100 and E-21264211-288.04-1602322). All animal procedures followed the Kafkas University local ethical committee for animal experimentation (KAÜ HADYEK) guidelines under KAÜ HADYEK/ 2018-050 and KAÜ-HADYEK/ 2021-083 permission number. The study was conducted in accordance with the local legislation and institutional requirements.
Author contributions
MN: Conceptualization, Data curation, Formal analysis, Funding acquisition, Investigation, Methodology, Project administration, Resources, Software, Supervision, Validation, Visualization, Writing – original draft, Writing – review & editing. RP: Conceptualization, Formal analysis, Methodology, Resources, Software, Supervision, Validation, Writing – review & editing. KY: Conceptualization, Visualization, Writing – review & editing. EÇ: Data curation, Writing – review & editing. AKÇ: Data curation, Writing – review & editing, Validation. JK: Validation, Writing – review & editing, Conceptualization. ÇŞ: Conceptualization, Writing – review & editing, Supervision.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This project has been funded by the Wild Animal Initiative (WAI) under the project code C-2023-00005.
Acknowledgments
Many thanks to Luke Hecht for his constructive comments on the early draft and Sabine Macho-Maschler for steroid analyses. We are grateful to the Wild Animal Initiative for its financial support.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The authors declare that no Gen AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
1. Şekercioğlu, ÇH, Anderson, S, Akçay, E, and Bilgin, R. Turkey’s rich natural heritage under assault. Science (1979) (2011) 334:1637 LP – 1639. doi: 10.1126/science.334.6063.1637-b
2. Ashrafzadeh, MR, Khosravi, R, Mohammadi, A, Naghipour, AA, Khoshnamvand, H, Haidarian, M, et al. Modeling climate change impacts on the distribution of an endangered brown bear population in its critical habitat in Iran. Sci Total Environ. (2022) 837:155753. doi: 10.1016/j.scitotenv.2022.155753
3. Sıkdokur, E, Naderi, M, Çeltik, E, Kemahlı Aytekin, MÇ, Kusak, J, Sağlam, İK, et al. Human-brown bear conflicts in Türkiye are driven by increased human presence around protected areas. Ecol Inform. (2024) 81:102643. doi: 10.1016/j.ecoinf.2024.102643
4. Naderi, M, Kusak, J, Bojarska, K, Chynoweth, M, Green, A, and Şekercioğlu, HÇ. Hares, humans, and lynx activity rhythms: who avoids whom? Hystrix Ital J Mammal. (2021) 32:147–52. doi: 10.4404/hystrix-00462-2021
5. Hofer, H, and East, ML. Biological conservation and stress. Adv Study Behav. (1998) 27:405–525. doi: 10.1016/S0065-3454(08)60370-8
6. Broom, DM, and Johnson, KG. Stress and animal welfare. Cham: Springer International Publishing (2019).
7. Balcı, ES, and Sabuncuoğlu Çoban, N. Measuring stress in animals by noninvasive methods. Vet Sci Pract. (2024) 19:52–8. doi: 10.17094/vetsci.1471401
8. Serra, M, Wolkers, CP, and Urbinati, E. Physiological indicators of animal welfare. Revista Brasileira de Zoociências. (2018) 19:70–96. doi: 10.34019/2596-3325.2018.v19.24726
9. Reeder, DM, and Kramer, KM. Stress in free-ranging mammals: integrating physiology, ecology, and natural history. J Mammal. (2005) 86:225–35. doi: 10.1644/BHE-003.1
10. Karaer, MC, Čebulj-Kadunc, N, and Snoj, T. Stress in wildlife: comparison of the stress response among domestic, captive, and free-ranging animals. Front Vet Sci. (2023) 10:1167016. doi: 10.3389/fvets.2023.1167016
11. Babic, NL, Johnstone, CP, Reljić, S, Sergiel, A, Huber, Đ, and Reina, RD. Evaluation of physiological stress in free-ranging bears: current knowledge and future directions. Biol Rev. (2023) 98:168–90. doi: 10.1111/brv.12902
12. Davenport, MD, Tiefenbacher, S, Lutz, CK, Novak, MA, and Meyer, JS. Analysis of endogenous cortisol concentrations in the hair of rhesus macaques. Gen Comp Endocrinol. (2006) 147:255–61. doi: 10.1016/j.ygcen.2006.01.005
13. Malcolm, KD, McShea, WJ, Garshelis, DL, Luo, S-J, Van Deelen, TR, Liu, F, et al. Increased stress in Asiatic black bears relates to food limitation, crop raiding, and foraging beyond nature reserve boundaries in China. Glob Ecol Conserv. (2014) 2:267–76. doi: 10.1016/j.gecco.2014.09.010
14. Tryland, M, Brun, E, Derocher, AE, Arnemo, JM, Kierulf, P, Ølberg, R-A, et al. Plasma biochemical values from apparently healthy free-ranging polar bears from Svalbard. J Wildl Dis. (2002) 38:566–75. doi: 10.7589/0090-3558-38.3.566
15. Davies, N, Gillett, A, McAlpine, C, Seabrook, L, Baxter, G, Lunney, D, et al. The effect of ACTH upon faecal glucocorticoid excretion in the koala. J Endocrinol. (2013) 219:1–12. doi: 10.1530/JOE-13-0123
16. Thompson, DP, Crouse, JA, McDonough, TJ, Barboza, PS, and Jaques, S. Acute thermal and stress response in moose to chemical immobilization. J Wildl Manag. (2020) 84:1051–62. doi: 10.1002/jwmg.21871
17. Piñero, JA, Jansen, HT, Robbins, CT, Vincent, EP, and Lafferty, DJR. Blood cortisol and faecal cortisol metabolite concentrations following an ACTH challenge in unanaesthetized brown bears (Ursus arctos). Conserv Physiol. (2025) 13:coae093. doi: 10.1093/conphys/coae093
18. Palme, R. Non-invasive measurement of glucocorticoids: advances and problems. Physiol Behav. (2019) 199:229–43. doi: 10.1016/j.physbeh.2018.11.021
19. Hein, A, Palme, R, Baumgartner, K, von Fersen, L, Woelfing, B, Greenwood, AD, et al. Faecal glucocorticoid metabolites as a measure of adrenocortical activity in polar bears (Ursus maritimus). Conserv Physiol. (2020) 8:coaa012. doi: 10.1093/conphys/coaa012
20. Hunninck, L, May, R, Jackson, CR, Palme, R, Røskaft, E, and Sheriff, MJ. Consequences of climate-induced vegetation changes exceed those of human disturbance for wild impala in the Serengeti ecosystem. Conserv Physiol. (2020) 8:coz117. doi: 10.1093/conphys/coz117
21. Anderwald, P, Campell Andri, S, and Palme, R. Reflections of ecological differences? Stress responses of sympatric alpine chamois and red deer to weather, forage quality, and human disturbance. Ecol Evol. (2021) 11:15740–53. doi: 10.1002/ece3.8235
22. Rehnus, M, Wehrle, M, and Palme, R. Mountain hares Lepus timidus and tourism: stress events and reactions. J Appl Ecol. (2014) 51:6–12. doi: 10.1111/1365-2664.12174
23. Thiel, D, Jenni-Eiermann, S, Braunisch, V, Palme, R, and Jenni, L. Ski tourism affects habitat use and evokes a physiological stress response in capercaillie Tetrao urogallus: a new methodological approach. J Appl Ecol. (2008) 45:845–53. doi: 10.1111/j.1365-2664.2008.01465.x
24. Zbyryt, A, Bubnicki, JW, Kuijper, DPJ, Dehnhard, M, Churski, M, and Schmidt, K. Do wild ungulates experience higher stress with humans than with large carnivores? Behav Ecol. (2018) 29:19–30. doi: 10.1093/beheco/arx142
25. Corlatti, L, Palme, R, Valencak, TG, and Gomez, KM. Season-dependent impact of forage quality on stress in alpine chamois. Ecol Evol. (2023) 13:e10045. doi: 10.1002/ece3.10045
26. Rodrigues, MA, Wittwer, D, and Kitchen, DM. Measuring stress responses in female Geoffroy’s spider monkeys: validation and the influence of reproductive state. Am J Primatol. (2015) 77:925–35. doi: 10.1002/ajp.22421
27. Escobar-Ibarra, I, Mayagoitia-Novales, L, Alcántara-Barrera, A, Cerda-Molina, AL, Mondragón-Ceballos, R, Ramírez-Necoechea, R, et al. Long-term quantification of faecal glucocorticoid metabolite concentrations reveals that Mexican grey wolves may habituate to captivity. Eur Zool J. (2017) 84:311–20. doi: 10.1080/24750263.2017.1332111
28. Lyons, J, Mastromonaco, G, Edwards, DB, and Schulte-Hostedde, AI. Fat and happy in the city: eastern chipmunks in urban environments. Behav Ecol. (2017) 28:1464–71. doi: 10.1093/beheco/arx109
29. Rangel-Negrín, A, Alfaro, JL, Valdez, RA, Romano, MC, and Serio-Silva, JC. Stress in Yucatan spider monkeys: effects of environmental conditions on fecal cortisol levels in wild and captive populations. Anim Conserv. (2009) 12:496–502. doi: 10.1111/j.1469-1795.2009.00280.x
30. Carbillet, J, Hollain, M, Rey, B, Palme, R, Pellerin, M, Regis, C, et al. Age and spatio-temporal variations in food resources modulate stress-immunity relationships in three populations of wild roe deer. Gen Comp Endocrinol. (2023) 330:114141. doi: 10.1016/j.ygcen.2022.114141
31. Farahani, R, and Asgharzadeh, A. Golestan National Park’s ecosystem services, a case with brown bears habitat suitability. Sci Rep Life Sci. (2023) 4:1–7. doi: 10.5281/zenodo.7632690
32. Şekercioğlu, ÇH. Turkey’s first wildlife corridor links bear, wolf and lynx populations to the Caucasus forests – news watch. Natl Geogr Mag (2012) (Accessed August 17, 2024).
33. Chynoweth, MW, Çoban, E, and Şekercioğlu, ÇH. Conservation of a new breeding population of Caucasian lynx (Lynx lynx dinniki) in eastern Turkey. Turk J Zool. (2015) 39:541–3. doi: 10.3906/zoo-1405-10
34. Capitani, C, Chynoweth, MW, and Kusak, J. Wolf diet in an agricultural landscape of North-Eastern Turkey. Mammalia. (2016) 80:329–34. doi: 10.1515/mammalia-2014-0151
35. Cozzi, G, Chynoweth, M, Kusak, J, Çoban, E, Çoban, A, Ozgul, A, et al. Anthropogenic food resources foster the coexistence of distinct life history strategies: year-round sedentary and migratory brown bears. J Zool. (2016) 300:142–50. doi: 10.1111/jzo.12365
36. Atalay, İ. A general survey of the vegetation of north - eastern Anatolia. Ege Coğrafya Derg. (1983) 1:14–39.
37. Tobler, M, and Powell, G. Estimating jaguar densities with camera traps: problems with current designs and recommendations for future studies. Biol Conserv. (2013) 159:109–18. doi: 10.1016/j.biocon.2012.12.009
38. Kays, R, Kranstauber, B, Jansen, P, Carbone, C, Rowcliffe, M, Fountain, T, et al. Camera traps as sensor networks for monitoring animal communities. Local Computer Networks, 2009. LCN 2009. IEEE 34th Conference on (2009), 811–818.
39. Luo, G, Wei, W, Dai, Q, and Ran, J. Density estimation of unmarked populations using camera traps in heterogeneous space. Wildl Soc Bull. (2020) 44:173–81. doi: 10.1002/wsb.1060
40. Akkucuk, U, and Sekercioglu, C. NGOs for environmental sustainability: the case of KuzeyDoga society. Fresenius Environ Bull. (2016) 25:6038–44.
41. Huber, D, Kusak, J, and Radisic, B. Analysis of efficiency in live-capture of European Brown bears. J Wildlife Res. (1996) 1:158–62.
43. Palme, R, Touma, C, Lepschy, M, Arias, N, and Dominchin, F. Steroid extraction: get the best out of faecal samples. Wien Tierarztl Monatsschr. (2013) 100:238–46.
44. Palme, R, and Moestl, E. Measurement of cortisol metabolites in faeces of sheep as a parameter of cortisol concentration in blood. Z Saugetierkd. (1997) 62:192–7. Available online at: https://api.semanticscholar.org/CorpusID:85798546
45. Dalerum, F, Ganswindt, A, Palme, R, Bettega, C, Delgado, M, Dehnhard, M, et al. Methodological considerations for using fecal glucocorticoid metabolite concentrations as an indicator of physiological stress in the brown bear (Ursus arctos). Physiol Biochem Zool. (2020) 93:197–204. doi: 10.1086/708630
46. Harlow, HJ, Lindzey, FG, Sickle, WDVan, and Gern, WA. Stress response of cougars to nonlethal pursuit by hunters. Can J Zool (1992) 70:136–139. doi: 10.1139/z92-020
47. Kitaysky, AS, Wingfield, JC, and Piatt, JF. Dynamics of food availability, body condition and physiological stress response in breeding black-legged kittiwakes. Funct Ecol. (1999) 13:577–84. doi: 10.1046/j.1365-2435.1999.00352.x
48. Stevenson, ET, Gese, EM, Neuman-Lee, LA, and French, SS. Levels of plasma and fecal glucocorticoid metabolites following an ACTH challenge in male and female coyotes (Canis latrans). J Comp Physiol B. (2018) 188:345–58. doi: 10.1007/s00360-017-1125-2
49. Bechshøft, TØ, Sonne, C, Rigét, FF, Letcher, RJ, Novak, MA, Henchey, E, et al. Polar bear stress hormone cortisol fluctuates with the North Atlantic oscillation climate index. Polar Biol. (2013) 36:1525–9. doi: 10.1007/s00300-013-1364-y
50. Sheriff, MJ, Dantzer, B, Delehanty, B, Palme, R, and Boonstra, R. Measuring stress in wildlife: techniques for quantifying glucocorticoids. Oecologia. (2011) 166:869–87. doi: 10.1007/s00442-011-1943-y
51. Creel, S, Fox, JE, Hardy, A, Sands, J, Garrott, B, and Peterson, RO. Snowmobile activity and glucocorticoid stress responses in wolves and elk. Conserv Biol. (2002) 16:809–14. doi: 10.1046/j.1523-1739.2002.00554.x
Keywords: brown bears, stress indicators, glucocorticoid metabolites, wildlife conservation, environmental impact
Citation: Naderi M, Palme R, Yarnell K, Çoban E, Çoban AK, Kusak J and Şekercioğlu ÇH (2025) Stress responses in free ranging brown bears (Ursus arctos) in eastern Türkiye. Front. Vet. Sci. 12:1639623. doi: 10.3389/fvets.2025.1639623
Edited by:
Daniel Mota-Rojas, Metropolitan Autonomous University, MexicoReviewed by:
James Edward Brereton, Sparsholt College, United KingdomAdriana Domínguez-Oliva, Metropolitan Autonomous University, Mexico
Copyright © 2025 Naderi, Palme, Yarnell, Çoban, Çoban, Kusak and Şekercioğlu. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Morteza Naderi, bW9ydGV6YW5hZGVyaUBzYWthcnlhLmVkdS50cg==
†ORCID: Morteza Naderi, orcid.org/0000-0002-7578-4159
Rupert Palme, orcid.org/0000-0001-9466-3662
Kelly Yarnell, orcid.org/0000-0001-7464-8764
Çağan H. Şekercioğlu, orcid.org/0000-0003-3193-0377