 Zacharia Waithaka Ng’ang’a1,2
Zacharia Waithaka Ng’ang’a1,2 Nuria Tous1
Nuria Tous1 Maria Ballester3
Maria Ballester3 Jakob Leskovec1
Jakob Leskovec1 Beatriz Jimenez-Moya1
Beatriz Jimenez-Moya1 Raúl Beltrán-Debón2
Raúl Beltrán-Debón2 David Torrallardona1†
David Torrallardona1† Joan Tarradas1*†
Joan Tarradas1*†- 1IRTA, Animal Nutrition, Mas Bové, Constantí, Catalonia, Spain
- 2MoBioFood Research Group, Department of Biochemistry and Biotechnology, Universitat Rovira i Virgili, Tarragona, Spain
- 3IRTA, Animal Breeding and Genetics, Torre Marimon, Caldes de Montbui, Catalonia, Spain
Zinc oxide (ZnO) has been used at pharmacological levels to promote gut health and growth performance in the critical postweaning (PW) phase of piglets. The pharmacological use of ZnO in piglet diets has been banned in Europe and other countries due to antimicrobial resistance and environmental concerns. Therefore, understanding its mode of action, including its molecular mechanisms, is crucial for developing effective and sustainable alternatives. We investigated the mechanisms by which dietary supplementation with 3,000 mg/kg ZnO supports gut health and improves growth performance of piglets during the first 14 days PW. During the 2 weeks of trial (0–14 d PW), ZnO fed piglets had higher average daily gain (165 vs. 123 g/d; p < 0.01), and tended to have increased average daily feed intake (204 vs. 181 g/d; p < 0.1) and improved gain-to-feed ratio (0.669 vs. 0.774; p < 0.05) compared to control piglets. Feces from piglets in the ZnO group were also more consistent during the 2 weeks of trial (p < 0.01). At day 14 PW, ZnO piglets had lower calprotectin concentrations in serum (p < 0.01). Dietary ZnO downregulated several genes, involved in immune, oxidative and inflammatory responses, in jejunal (GPX2, REG3G, IL-8, IL-6, IL-22, and TGFβ1) and ileal (REG3G, IL-17A, IL-1β, and TLR2) mucosa (p < 0.05). It also downregulated the expression of the zinc transporter SLC39A4, that is associated with zinc homeostasis, in both tissues. Notably, PPARGC1A, which promotes energy production and lipid metabolism through fatty acid oxidation, was upregulated by ZnO in ileum. In conclusion, the current results suggest that high dietary levels of ZnO reduce the expression of inflammatory cytokines, the oxidative enzyme GPX2, pathogen recognition proteins, and zinc transporters while promoting the expression of PPARGC1A gene related with energy metabolism in the intestine. Therefore, ZnO can facilitate a smoother weaning transition to reduce weaning related gut health disturbances, ultimately contributing to gut homeostasis and improved performance.

Graphical Abstract. Zinc oxide (ZnO) supplementation (3 kg/ton for 2 weeks post-weaning) improved piglet performance and gut homeostasis by modulating immune and oxidative stress responses in both jejunum and ileum. ZnO reduced calprotectin concentration in blood, which is normally released by activated neutrophils to bind zinc and stimulate inflammation. In jejunum, ZnO appeared to support Th17/Treg balance to regulate inflammation by reducing IL-6, IL-22 and TGF-β1 as well as CXCL-8 production while tending to decrease NFKB1 expression. ZnO also suppressed Reg3G, an antimicrobial peptide and GPX2, a modulator of oxidative status, in jejunum. ZnO downregulated TLR2, IL1B and IL-17A while limiting Reg3G-induced epithelial responses in ileum, helping preserve gut integrity. Additionally, ZnO upregulated PPARGC1A (PPAR-α) to promote energy production in the ileum. SLC39A4 was decreased in both tissues probably to limit excess zinc absorption. Overall, ZnO appears to support gut health through coordinated, site-specific transcriptional regulation of inflammation and gut barrier function pathways. PW, post-weaning (Figure created using draw.io).
1 Introduction
The supplementation of zinc oxide (ZnO) in the diets of weaned piglets has been a common strategy to mitigate post-weaning (PW) enteric infections, reduce the use of antibiotics, and enhance the growth performance of piglets (1–4). Weaning is a critical and stressful phase that is associated with reduced feed intake and impaired gut health, which is evidenced by gut inflammation, disrupted intestinal integrity, microbial imbalances, immunodepression and increased susceptibility to post-weaning diarrhea (PWD) (5–9). The use of pharmacological concentrations of ZnO (2,000–3,000 mg/kg) in postweaning feeds has been shown to improve growth performance of weanling pigs (1, 4, 10).
Zinc (Zn), commonly provided as ZnO, supports piglet gut health probably through several intertwined mechanisms to prevent PWD. It plays an essential role in numerous physiological processes including the synthesis of over 300 enzymes related to immune function and gut health (11–13). Firstly, ZnO at high doses may enhance intestinal barrier function, nutrient absorption, and intestinal development (14–16). ZnO supports the integrity of the gut barrier by promoting the activity of intestinal digestive enzymes and regenerating epithelial tissue (2, 17), upregulating the expression of tight junctions, such as ZO-1 (18), and improving intestinal architecture, thereby promoting enterocyte proliferation (18, 19), enhancing nutrient absorption, and reducing gut permeability (14, 20, 21). Secondly, ZnO modulates intestinal immunity and redox balance (22), by suppressing pro-inflammatory cytokines and up-regulating anti-inflammatory factors (2, 13, 18). During infections, it reduces the expression of innate immunity related genes (23, 24), and alleviates oxidative stress induced by inflammatory processes (18, 25, 26). Finally, high ZnO doses also appear to modulate gut microbiota composition (27) by increasing alpha-diversity (28), similarly to certain growth-promoting antibiotics (15). In several studies, it has been shown that ZnO shifts the small intestinal microbiota towards non-pathological bacterial populations, contributing to a more stable intestinal environment during the PW period (6, 15, 27–29). A recent dose–response study revealed that ZnO inclusions above 2,400 mg/kg are required to markedly reshape the gut microbiota and modulate blood metabolic profile (30). These combined effects of ZnO on the piglets’ barrier integrity, digestive function, immune regulation, antioxidant defense, and microbial composition can explain the consistent reduction in PWD prevalence and improved performance (2, 11).
Zinc deficiency is known to affect all body organs, induce immune dysfunctions, and increase susceptibility to diseases (13). Conversely, the withdrawal of pharmacological doses of ZnO in pig diets may have detrimental effects on PW growth performance and gut health (31), including villi atrophy, reduced absorptive surface area and digestive enzyme secretion (32). During the immediate PW period, the piglets’ low feed intake may result in Zn deficiency, which could be overcome by the use pharmacological levels of ZnO in diet to help in meeting the animal’s Zn requirements (33). However, after a few days when feed intake is normalized, these high levels may become problematic as they might cause zinc overload (>2 ppm in serum) which has been associated with significant reductions in serum copper and selenium concentrations (34), as well as chronic pancreatitis (35). It has also been shown that pharmacological ZnO can increase the accumulation of Zn in the small intestine, and activate the expression of apoptosis-related genes, causing damage to the intestinal epithelium (36).
In addition, dietary zinc is poorly absorbed, and it accumulates in manure leading to soil contamination and toxicity concerns (11, 37, 38). Moreover, pharmacological doses of ZnO may interact negatively with other nutrients (38), promote the acquisition of antibiotic multi-resistant genes in zoonotic pathogens such as enterotoxigenic Escherichia coli (ETEC) (39), contributing to the emergence of antimicrobial resistances (AMR) and, finally, exacerbating public health concerns (33, 40, 41). Consequently, these concerns over environmental pollution, toxicity and AMR have prompted regulatory authorities to impose limits on permissible dietary zinc levels in pig production (11). For instance, the European Union has banned pharmacological ZnO use since 2022 and set the maximum allowable dietary zinc level for weaned piglets at 150 mg/kg (42). Hansen et al. (33) indicated that, at this concentration, newly weaned piglets with low voluntary feed intake cannot meet their zinc nutritional requirements, impairing gut barrier integrity and this may predispose the piglets to enteric infections (43). This could partially explain why piglets fed diets with 150 mg/kg of Zn are more susceptible to ETEC than those fed pharmacological levels of ZnO (44). In response to the ban, alternative strategies to pharmacological ZnO such as probiotics, prebiotics, organic acids, essential oils, spray dried porcine plasma have been explored (45–48), even though their efficacy in matching ZnO benefits has been inconsistent.
Even with the advancements in ZnO research, the mode of action of ZnO in the different regions of the intestine of piglets during the early post-weaning period is still not completely deciphered and its characterization remains critical for the future development of new alternatives (2, 45). To address this knowledge gap, we hypothesized that ZnO modulates the concentrations of key gut health biomarkers, and the expression of intestinal genes related to barrier function, immune response, oxidative stress, metabolic activity and digestive enzymes, to partially explain its mode of action. Therefore, we studied the effects of pharmacological doses of ZnO on selected gut health markers and expression of specific genes related to different gut functionalities in jejunum, ileum and caecum, aiming to contribute critical insights to the understanding of its mechanisms of action.
2 Materials and methods
This study adhered to the EU principles for the protection of animals used for scientific purposes and complied with Directive 2010/63/EU of 22 September 2010, as well as the Spanish regulations for the care and use of animals in research (Real Decreto 53/2013). The experimental procedures received approval from IRTA’s Ethical Committee for Animal Experimentation and the Commission on Animal Experimentation of Generalitat de Catalunya, Spain (Project number 11766).
2.1 Experimental design
The study was conducted in the postweaning facilities of the experimental farm of Animal Nutrition, IRTA (Centre Mas Bové, Constantí, Spain). A total of one hundred and eighty newly weaned piglets ([Large White × Landrace] × Pietrain; mixed sexes; 7.03 ± 1.12 kg BW) of around 26 days of age, in two separate batches (3 weeks apart) from IRTA’s experimental sow herd, were used. For each batch, the piglets were housed in two weaning rooms with 12 and 6 pens, respectively. The experimental herd had a relatively high-health status without clinical disease outbreaks having been recorded for at least the 12 months preceding the study. Therefore, the facilities were not cleaned or disinfected before the start of the trial with the intention of creating slightly challenging conditions for the animals. The rooms are provided with automatic heating, forced ventilation and completely slatted floors. Feed was distributed ad libitum in one feeding trough per pen with four feeding spaces. Water was also provided ad libitum throughout all the animals and experimental periods.
At the start of each batch, 90 piglets were selected and randomly distributed by initial body weight into nine blocks per batch. Each block consisted of two pens with five piglets each. There were two experimental treatments: Control diet and the same diet containing zinc oxide at 3,000 mg/kg (ZnO). Within each block, the two treatments were randomly distributed between pens according to a randomized complete block design.
At the start of each batch (day 0 PW) piglets were offered diets with pre-starter specifications until day 14. Feeds were formulated according to the minimum nutrient requirements as recommended by FEDNA (49) and the composition of the diets is presented in Table 1. The vitamin-mineral premix used in the basal diet contained 110 mg ZnO per kg of feed. For the ZnO diet, ZnO (0.3%) was added on top of the basal diet directly into the feed mixer. The feeds were presented in pelleted form and offered ad libitum.
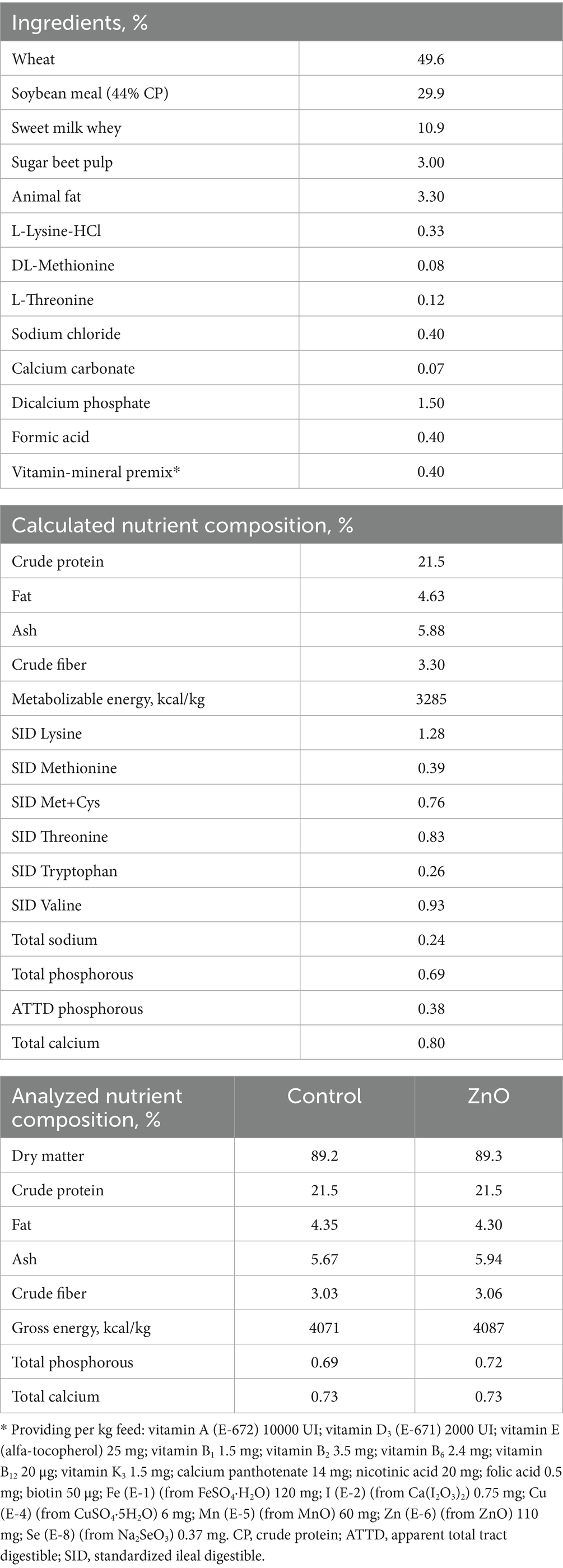
Table 1. Ingredient composition and calculated nutrient contents of the basal diet and analyzed nutrient contents of the experimental diets (as-fed basis).
2.2 Performance and fecal scores
Feed (per pen) and individual piglets were weighed at days 0 and 14 PW. Pen’s average initial and final body weight, daily weight gain, feed intake and gain-to-feed ratio were calculated for the period 0–14 days. Fecal consistency was individually assessed using a 5-category score system (0 = firm and shaped; 1 = soft and shaped; 2 = soft without shape; 3 = loose; 4 = watery) as a slight modification of the scale proposed by Pederson and Toft (50). This was done by observing the feces in the pens; if they were firm and shaped, it was considered that all the piglets in that pen had a score of 0, if not, the piglets in that pen were individually scored by direct inspection. The highest individual scores given during weeks 1 and 2, respectively, were used for the statistical analysis.
2.3 Sample collection
At day 14 post-weaning, one animal per pen (the animal with intermediate initial body weight in each pen) was sampled for blood and euthanized to obtain samples of bile and intestinal mucosa. Blood serum and bile samples were collected into cryotubes and stored at −80 °C until analysis. Tissue fragments of 10 cm from jejunum (400 cm proximal from the ileo-cecal valve), ileum (50 cm proximal from the ileo-cecal valve) and caecum (5 cm distal from the ileo-cecal valve) were obtained and rinsed with saline solution, and the corresponding mucosa was scrapped and collected in 1 mL RNAlater (2 mL cryotube; 2 tubes per sample) before being frozen at −80 °C until analysis.
2.4 Gut barrier and immune indicators—ELISA
Commercial ELISA kits were employed, following the manufacturers’ protocols, for the quantitative measurement of calprotectin and citrulline in blood serum, and secretory immunoglobulin A (sIgA) in bile samples (CSB-EQ013485PI, Cusabio, China; K 6600, Immundiagnostik AG, Bensheim, Germany; and abx258080, Abbexa, Cambridge, United Kingdom, respectively). Absorbance was read at 450 nm using a microplate reader (Biotek Epoch2, Biotek Instruments, Winooski, Vermont, United States).
2.5 RNA extraction and reverse transcription
RNA was isolated from the jejunal, ileal, and cecal mucosa of the individual pigs using the miRNeasy Mini Kit (Qiagen, Hilden, Germany) according to the manufacturer’s protocol. The RNA concentrations and purities were assessed with a NanoDrop 1000 spectrophotometer (Thermo Scientific, Waltham, MA, United States). Extracted RNA samples were stored at −80 °C for future use. One μg of each RNA sample was used to synthesize complementary DNA (cDNA) using the PrimeScript RT Reagent Kit (Takara, RR037A, Dalian, China), following the instructions provided by the manufacturer.
2.6 Pre-amplification
Pooled cDNA samples were utilized to create a standard (STD) curve for quantification during qPCR. The pooled and individual cDNA samples diluted 1:5 in DNA suspension buffer (10 mM Tris–HCl, 1 mM EDTA, pH 8.0) underwent pre-amplification using the Preamp Master Mix (St Standard Biotools, San Francisco, CA, United States) and a custom primer mix containing 48 primer pairs at 500 nM each for subsequent qPCR analysis. The pre-amplification process included an initial denaturation at 95 °C for 10 min, followed by 15 cycles of 95 °C for 15 s and 60 °C for 4 min. Pre-amplification reactions were cleaned using Exonuclease I (Exo I) to remove residual primers. The pre-amplified cDNA -Exo I treated samples were subsequently diluted 1:20 using DNA suspension buffer prior to downstream analysis.
2.7 High-throughput gene expression analysis—qPCR
Quantitative real-time PCR (qPCR) was performed on the BioMark™ X9 system (Standard BioTools™, United States—formerly Fluidigm Corporation) utilizing 96.96 Dynamic Array Integrated Fluidic Circuit (IFC) chips. This setup allowed for 48 primer pairs in duplicates across 96 samples. The primers included in-house designs and modified versions of those previously reported by González-Solé et al. (51), sourced from Condalab (Spain), as detailed in Supplementary Table S1. This includes genes involved in: barrier function (OCLN, CLDN15, ZO1, CLDN1, MUC2, MUC13, and CLDN4), immune response, such as pattern recognition receptors, cytokines, chemokines and stress proteins (pBD3, REG3G, CXCL8—also referred as IL8, IKBKB, IL6, TLR4, pBD2, GBP1, IFNG, IL22, TGFB1, TNFa, IFNGR1, TLR2, NFKBIA, DEFB1, IL10, IL17A, CCL20, NFKB1, and IL1B), coding for enzymes and hormones implicated in the digestion process (GPX2, DAO, PPARGC1A, IGF1R, IDO1, FAXDC2, SOD2, HNMT, CCK, and ALPI), nutrient transport (SLC5A1, SLC7A8, SLC16A1, and SLC39A4), and stress response (HSPA4 and HSD11B1); analyzed relative to reference genes (GAPDH, ACTB, HPRT1 and TBP). Genes with minimal detection across samples were excluded from subsequent analyzes.
The qPCR sample mix was prepared by combining 2.7 μL of pre-amplified and Exo-I treated samples (diluted 1:20 DNA suspension buffer), 3.3 μL of 20 × EvaGreen DNA Binding Dye (Biotium, PN 31000), and 0.5 μL of 20 × DNA Binding Dye Sample Loading Reagent (Fluidigm, PN 100-0388). For each assay, the mix consisted of 27 μL of DNA suspension buffer, 30 μL of 2X Assay Loading Reagent and 3 μL of forward and reverse primers, each at a final concentration of 5 μM.
On the 96.96 Dynamic Array IFC chips, 5 μL of each sample mix and 5 μL of each assay mix were loaded into the corresponding inlets. Non-template controls (NTCs) to monitor potential contamination or nonspecific amplification and STD curve dilutions, prepared from pooled cDNA samples were included to the chip. The pre-amplified STD was diluted 1/5 with DNA suspension buffer and followed by five ¼ dilutions. The chips were processed on the BioMark™ X9 system under the following thermal cycling conditions: initial activation at 95 °C for 60 s, followed by 35 cycles of 96 °C for 5 s and 60 °C for 20 s. Fluorescence data were recorded at each cycle, and melting curve analyzes confirmed specific amplification.
Initial data quality control and threshold settings were performed using the BioMark™ system Real-Time PCR Analysis software (version 1.0.2; Standard BioTools, United States). Further normalization and efficiency corrections were conducted using DAG Expression software1 (52). Four porcine intestinal reference genes—GAPDH, ACTB, HPRT1, and TBP—were evaluated for stability, with TBP and GAPDH identified as the most stable reference genes for normalization.
2.8 Statistical analysis
All the performance parameters measured were compared between treatments by ANOVA using the PROC MIXED procedure of the statistical package SAS (SAS 9.4; SAS Institute Inc., Cary, NC, United States). A randomized complete block design was used with initial weight, batch and pen location as block criteria. For the statistical analysis, the block as a random factor and treatment as a fixed effect were included in the model and the pen was used as the experimental unit. Data in tables are presented as least-square means. Fecal consistency assessed using the maximum score given to each individual piglet each week. The score frequencies were analyzed using a Chi-square test of independence utilizing the PROC FREQ procedure in SAS software. ELISA data were Log transformed and tested for normality using the Shapiro–Wilk test, and then the differences between the Control and ZnO treatment groups were assessed using ANOVA in the GLIMMIX procedure of SAS software. The model included treatment as a fixed effect and block (same criteria as for performance) as a random effect. Differential gene expression values across the jejunum, ileum, and cecum were assessed using the GLIMMIX procedure in SAS. The model incorporated treatment as a fixed effect and block (same criteria as for performance) as a random effect. All the results were considered statistically significant at p < 0.05, while values of 0.05 ≤ p < 0.10 were interpreted as a tendency.
3 Results
3.1 Growth performance
The performance results (0–14 days) are shown in Table 2. Piglets from the ZnO group had higher final BW (p < 0.01) and average daily weight gain (ADG) than those from the control group (p < 0.01) and also tended to increase their average daily feed intake (ADFI) (p < 0.1). Moreover, ZnO improved the gain-to-feed ratio (GFR) compared to control (p < 0.05).
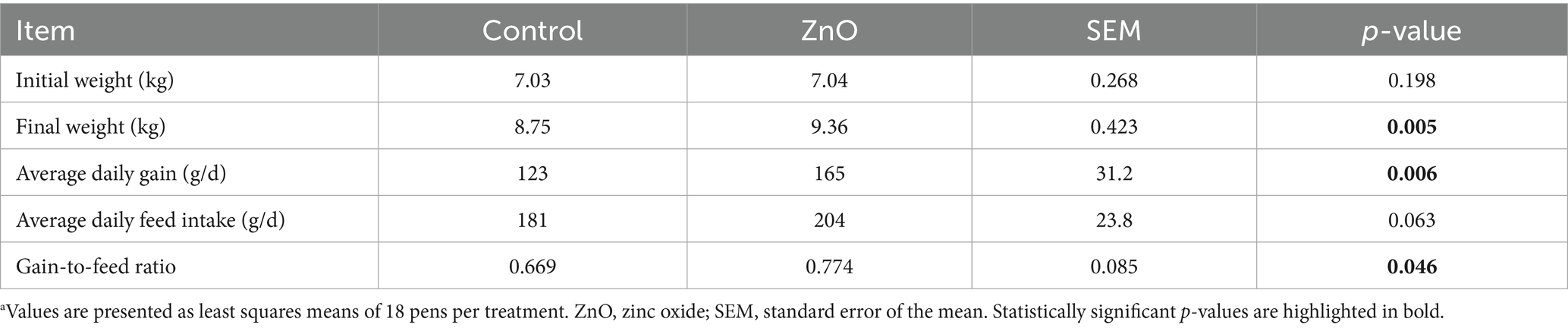
Table 2. Productive parameters of piglets fed control or ZnO diets between 0 and 14 days postweaninga.
3.2 Fecal scores
The weekly fecal consistency results are shown in Figure 1. The analysis of individual fecal score frequencies revealed significant differences between treatments during week 1 (Chi-square = 16.82, p < 0.01) and week 2 (Chi-square = 14.05, p < 0.01). Notably, for the first week, 51% of the Control piglets had a fecal score of 3 (loose), compared to only 30% in the ZnO group. Additionally, 12% of the control pigs had a more consistent score of 1 (soft and shaped), compared to 27% of pigs in the ZnO group. During the second week, there were more piglets with loose (score of 3; Control = 31%, ZnO = 21%) and watery feces (score of 4; Control = 30%, ZnO = 20%) in the control group than in the ZnO group.
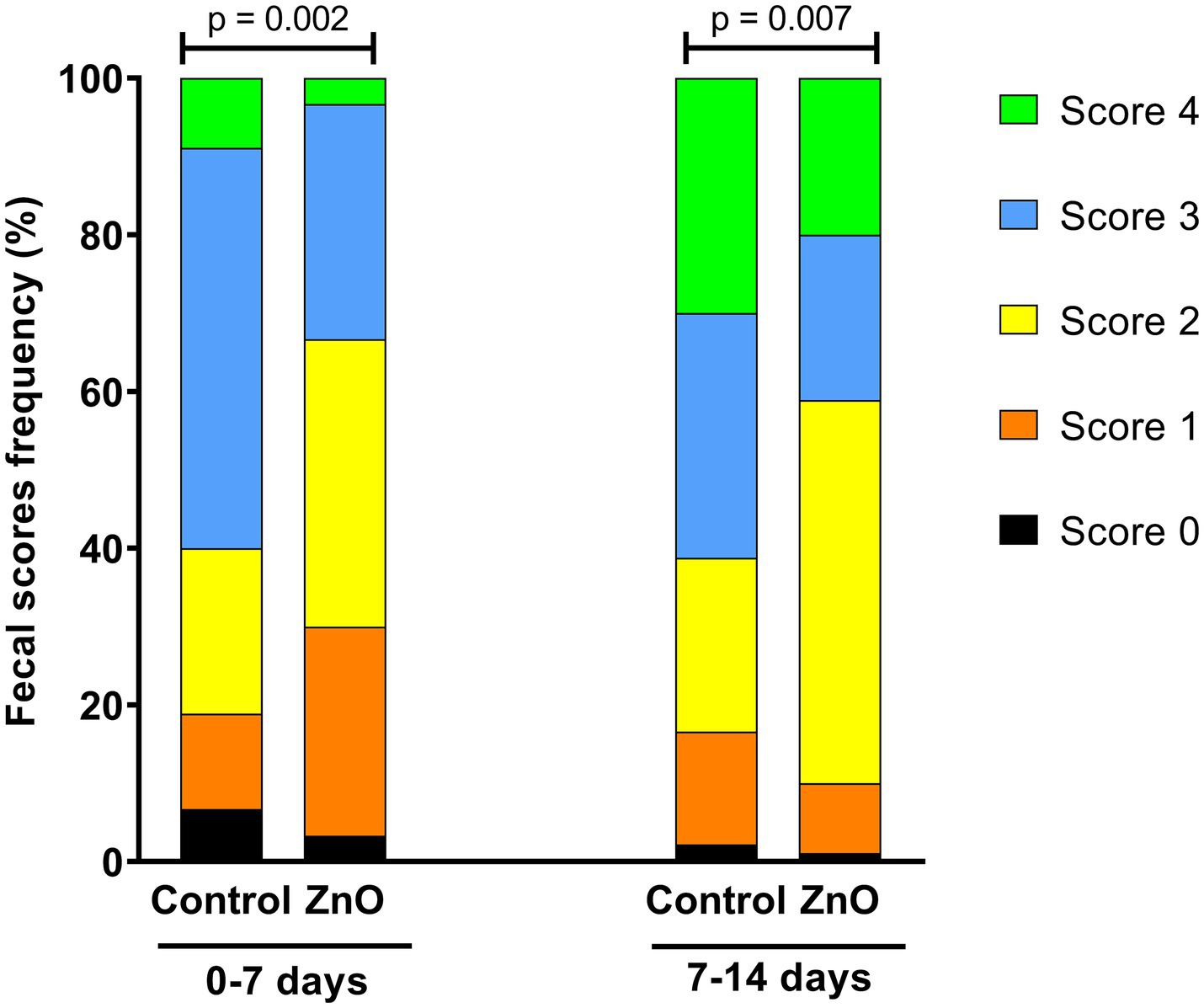
Figure 1. Fecal scores frequency (0 = firm and shaped; 1 = soft and shaped; 2 = soft without shape; 3 = loose; 4 = watery) of piglets between 0–7 days and 7–14 days postweaning.
3.3 Biomarkers of gut health
The concentration levels of the intestinal health biomarkers analyzed in this study are shown in Table 3. Serum calprotectin levels, a marker of intestinal inflammation, were lower in ZnO-treated compared to control piglets (355 vs. 240 ng/mL, p < 0.01). Serum citrulline, as a biomarker of intestinal integrity, and bile sIgA, as an indicator of mucosal innate immunity, were unaffected by ZnO supplementation.
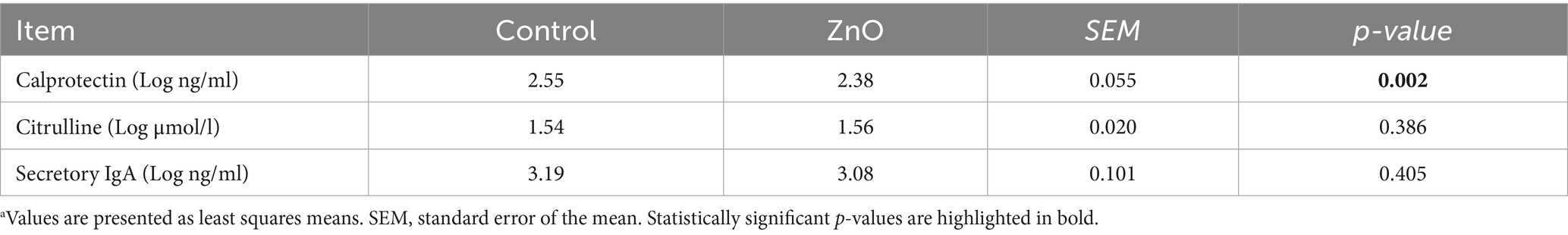
Table 3. Effects of ZnO on the Log-transformed concentrations of calprotectin and citrulline in blood serum and sIgA in bile at day 14 post-weaninga.
3.4 Intestinal gene expression profile
Gene expression analysis using microfluidic qPCR provided insights into the molecular effects of ZnO on gut health. All the measured relative gene expression values in jejunum, ileum and cecum are shown in Table 4.
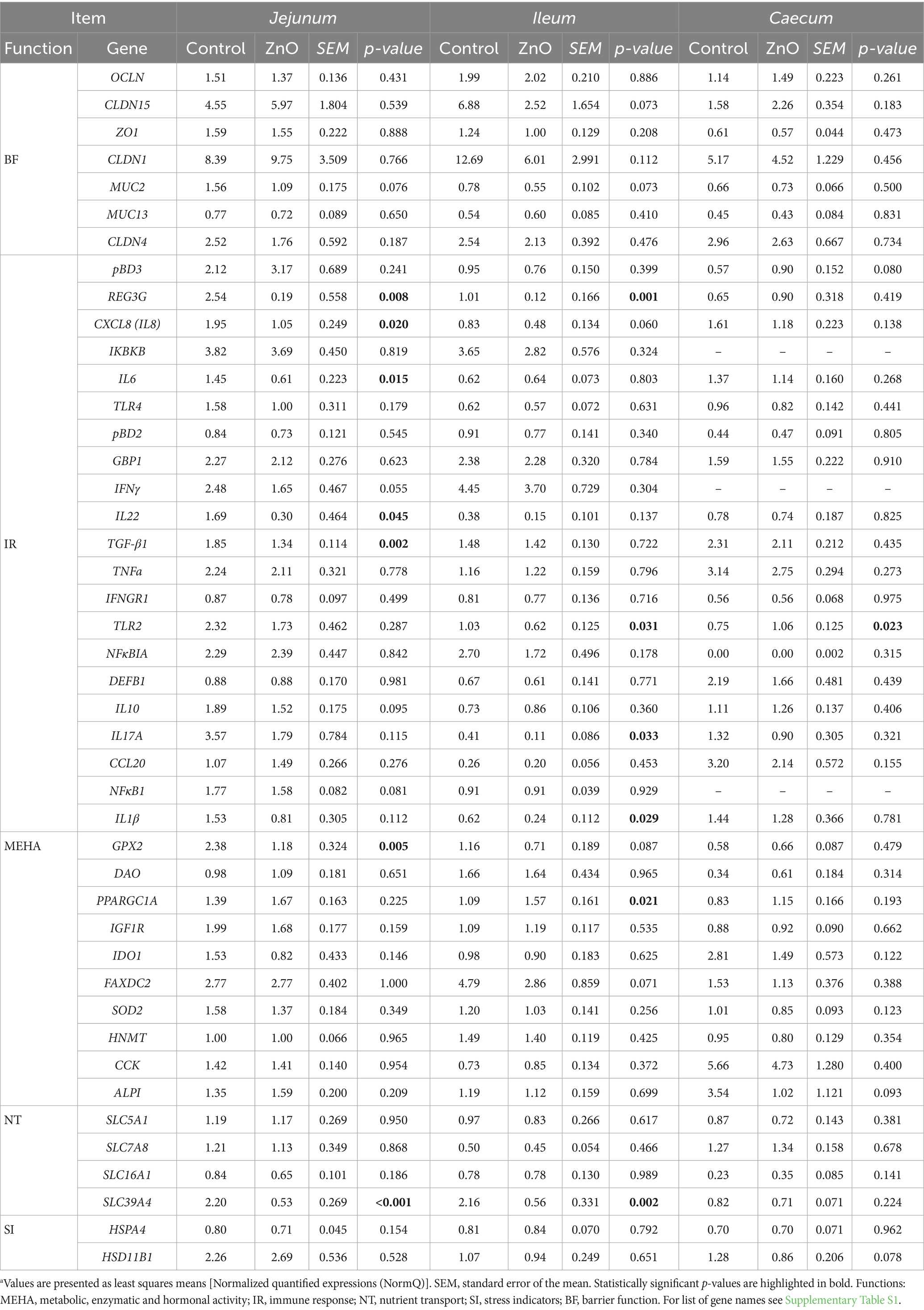
Table 4. Relative gene expression in the piglets’ jejunal, ileal and cecal mucosa of control and ZnO treatment relative to the reference genes at day 14 post-weaninga.
In Jejunum, several genes showed differential expression between the control and ZnO treatments. The expression of the immune-related genes REG3G, CXCL8/IL8, IL-6, IL-22, and TGFB1 was downregulated in the ZnO group compared to the control group (p < 0.05). The expressions of the Zn transporter gene SLC39A4 and the antioxidative-related gene GPX2 were also decreased in the ZnO fed piglets (p < 0.01). Finally, ZnO also tended to downregulate the expressions of IFNγ, MUC2, IL-10, and NFκB1 (p < 0.1).
In the ileum, ZnO downregulated pro-inflammatory and other immune-related genes at d 14 PW, such as REG3G, IL-17A, IL-1B, and TLR2 (p < 0.05). The expression of SLC39A4 was also downregulated in the ileum (p < 0.001) of ZnO fed piglets. The only gene that was upregulated by ZnO in the ileum was PPARGC1A (p < 0.05). Moreover, tendencies for the downregulation of the gene expressions for CLDN15, MUC2, CXCL8/IL-8, GPX2, and FAXDC2 were also observed in the ileum of the ZnO group (p < 0.1).
In the caecum, TLR2 gene was upregulated in ZnO fed piglets compared to the control (p < 0.05).
4 Discussion
Weaning is a critical phase in swine production in which environmental and pathogenic stressors are often linked to impairment of gut health, high incidence of PWD, reduced performance and increased mortality (5, 9). Dietary ZnO at high doses of up to 3,000 mg/kg has been widely used in pre-starter diets for weaned pigs due to its potent antimicrobial and growth performance enhancing capacities (2, 18). However, the regulatory ban on this use of ZnO in Europe, driven by concerns over Zn environmental pollution and antimicrobial resistances emergence among others, has generated the need to develop sustainable effective alternatives to ZnO, for which a deeper understanding of its mechanisms of action are required (11, 42). With the aim to better understand the modes of action of ZnO in weaned pigs, the present study investigated the effects of high levels of ZnO supplementation on growth performance, gut health biomarkers in blood and bile, and the gene expression in different segments of the intestine for key genes related to barrier function, immune response, metabolic and enzymatic activity, stress, and nutrient transport.
In the current trial, we observed that high doses of ZnO increased ADG, ADFI, and feed efficiency, results that are consistent with many previous studies (1, 10, 53). Hollis et al. (54) demonstrated that pigs fed 2,000 mg/kg ZnO gained weight faster and consumed more feed than those fed either 500 mg/kg ZnO or organic Zn sources. Similarly, Hill et al. (10) observed that supplemental ZnO at 1,500 to 2,000 mg Zn/kg improved postweaning pig performance, and in another study the supplementation with 3000 mg/kg ZnO increased BW after 14 d PW (18).
These performance benefits are often attributed to the ability of ZnO to reduce the incidence of PWD (15, 55). In our study, the piglets from the ZnO group had better fecal consistency, indicative of a lower prevalence of diarrhea, underscoring the efficacy of ZnO to manage post-weaning challenges. This is in line with numerous studies that have reported similar results in weaned pigs (18, 20, 56). These improvements are likely mediated by antimicrobial properties of ZnO, which can inhibit the adhesion of pathogenic bacteria to intestinal cells and prevent the subsequent disruption of intestinal integrity (11, 21). Direct effects of ZnO on pathogens have not been completely defined, although electrostatic forces, production of Zn2+ ions and the generation of reactive oxygen species have been described as possible pathways of the bactericidal action of ZnO (57). Recent research proposes that the mechanism of action of ZnO involves the reduction of toxic and inflammatory microbial metabolites resulting from a lower abundance of pathogenic bacteria in the gut (30, 58).
A key finding of our study was the reduction of serum calprotectin levels in the piglets fed ZnO. Calprotectin, an innate immune protein complex with antimicrobial activity, can bind Zn and other metals with high affinity, and starves bacteria of these essential nutrients (59, 60). As a marker of inflammation that is released by activated neutrophils, calprotectin in serum is elevated in conditions of gut inflammation and barrier dysfunction in different species (61, 62). In the current study, high levels of dietary ZnO may have directly prevented neutrophil activation and subsequent calprotectin release triggered by the recognition of bacterial pathogens. Moreover, it has been shown in mouse jejunal organoids that calprotectin increases the expression of gene SLC39A4 which encodes for the Zn transporter ZIP4 (63). This transporter can modulate the intracellular and extracellular concentrations and availability of Zn2+ in response to available Zn2+ levels (63). In our study we observed a clear downregulation of SLC39A4 expression in both jejunum and ileum of the ZnO piglets, which might be explained by the reduction of gut inflammation (less calprotectin release), or because the high circulant Zn2+ available makes unnecessary the overexpression of Zn transporters for Zn2+ cell internalization. ZIP4 is important for Zn homeostasis and absorption from the small intestine, as well as for the regulation of intestinal epithelial function (64, 65); it can therefore be indicative of Zn absorption and utilization (66). Similarly to what we have observed in both the jejunum and the ileum, high doses of Zn lead to the downregulation of SLC39A4 in the jejunum of piglets (67, 68). Interestingly, the expression of ZIP4 is regulated by Zn, being increased in conditions of deficiency and decreased with adequate levels (69, 70). This suggests an adequate Zn status with the high ZnO diets, limiting further uptake and the possibly of mitigating Zn induced cytotoxicity (71). Alternatively, it may also reflect that the dose of Zn administered exceeded the animals’ physiological needs, suggesting that such high levels may not be necessary to meet the nutritional requirements of healthy piglets.
Serum citrulline, a biomarker for intestinal enterocyte mass and function (72), was unaffected by dietary ZnO at day 14 PW. This is in line with longitudinal studies showing that citrullinemia falls only during the first 7 to 10 days after weaning and returns to baseline levels by day 15, thus making it less sensitive to the inflammatory changes relieved by ZnO in the current study (73–75). Secretory IgA provides the first line of innate mucosal defense by neutralizing luminal pathogens without provoking inflammation (76). Although most of the sIgA in the gut lumen is released locally, a proportion is delivered into the upper intestine via the biliary tract (77–80), and biliary sIgA has been used to assess mucosal immune status in pigs (81, 82). We found that pharmacological ZnO had no effect on biliary sIgA levels at 14 d PW. However, Ramis et al. (83) observed reduced levels of fecal sIgA with ZnO supplementation at 8 d PW but that was not observed at later stages (11–15 d PW). Although the lack of differences in the current study might be attributed to the limitations of using bile instead of intestinal contents to measure sIgA, it could also be due to the temporal dynamics of the immune response to ZnO.
During the PW period, piglets experience both prolonged and transient changes of gene expression of inflammatory cytokines like IFN-γ, IL-1β, IL-6, IL-8, IL-10, IL-12α, and TGF-β in jejunum, ileum, and colon (84). Our study revealed that ZnO reduces the expression of inflammatory genes in the small intestine. Notably, ZnO downregulated REG3G, IL-6, IL-8, IL-22, and TGF-β1 in the jejunum and IL-1β, REG3G, TLR2, and IL-17A in the ileum. Contrarily, ZnO increased TLR2 in the cecum. These results align with previous studies where ZnO prevented the induction of inflammatory processes, pointing out to a coordinated effort to maintain gut homeostasis in the PW period (18, 23, 24, 85). Our observation that ZnO downregulated jejunal IL-6, and IL-8, and tended to downregulate that of IFNγ and NFκB1, supports its proposed capacity to modulate pro-inflammatory signaling (86). Consistent with our findings in jejunum, Hu et al. (87) observed decreased mucosal mRNA levels of IL-6 and IFN-γ at day 7 PW using diosmectite ZnO at 500 mg/kg in early weaned pigs. Thus, ZnO may contribute to preserve epithelial integrity by suppressing the expression of proinflammatory cytokines which can lead to increased intestinal permeability (88).
Interestingly, in ZnO piglets at day 14, the gene expression for cytokines TGF-β1 and IL-22 was downregulated in jejunum, and that of IL-17A in ileum; cytokines that are important key regulators for the epithelial barrier function and repair (89, 90). These results contrast with previous findings showing that ZnO increased TGF-β expression in the intestine of weaned piglets at 8 d PW (91) and jejunal mucosa at 28 d PW (18), suggesting a role in repair. Our data suggest that in a context of overall reduced inflammation, the stimuli for TGF-β1 production are also reduced, pointing to a state of quiet homeostasis rather than active repair. This discrepancy may be explained by the pleiotropic nature of TGF-β1, which, together with IL-6, contributes to the regulatory signaling of T helper 17 (Th17) cells and regulatory T-cells (Tregs) (92–94). Zn is a known modulator of this axis, promoting the development of anti-inflammatory Treg cells while suppressing the differentiation of pro-inflammatory Th17 cells (95). Th17 cells can also promote tissue repair and homeostasis, an effect mediated by IL-22 by promoting the regeneration of epithelial tissues (96). Specifically, ZnO has been shown to directly inhibit phosphorylation of STAT3, a key transcription factor required for Th17 differentiation, thereby reducing the production of Th17-associated cytokines like IL-17A and IL-22 (97). Taken together, the downregulation of IL-22 in jejunum and IL-17A in ileum in ZnO-fed piglets is consistent with the lower serum calprotectin concentrations observed, because both cytokines can induce calprotectin expression in epithelial cells (98). This could be a possible mode of action of ZnO to dampen a major pathway of intestinal inflammation in jejunum. However, these observations require other functional assays to validate these potential mechanisms.
The gene REG3G, which encodes for an important C-type lectin that has bactericidal activity against Gram-positive bacteria, is induced by IL-22 via STAT3 pathway activation (99). Notably, we observed decreased expression of REG3G in both jejunum and ileum, despite the expression of IL-22 only being reduced in jejunum. Interestingly, Schokker et al. (28) also observed REG3G downregulation in both tissues from clinically healthy piglets fed with high doses of ZnO. This gene is also a regulator of Th17 cell mediated immunity as has been observed in mice, playing an important role in the physical segregation of microbiota from the host as well as in the immune response induced by pulmonary pathogens (100). All these findings support the hypothesis that ZnO suppresses inflammatory processes and consequently, Th17 responses, which aligns with the mechanistic evidence provided by Kitabayashi et al. (97), who showed that zinc directly inhibits Th17 cell development by attenuating STAT3 activation.
Beyond immunomodulation, ZnO has been shown to enhance the redox balance and prevent apoptosis in the jejunum of piglets at 15 d PW, effectively reducing weaning-related intestinal dysfunction and nutrient malabsorption (25). In our study, the jejunal downregulation of GPX2, a key antioxidant enzyme, suggests that ZnO may mitigate oxidative stress to protect enterocytes from damage, which is consistent with the reported decrease of plasma malondialdehyde levels, an indicator of oxidative stress or lipid peroxidation, in ZnO fed piglets (18).
In the ileum, ZnO downregulated the expression of IL-1β and TLR2 and tended to downregulate that of IL-8 and FAXDC2. The reduced IL-1β expression in our study is consistent with the results of Zhu et al. (18) who observed a downregulation of IL-1β expression in the jejunal mucosa by ZnO signifying its role in dampening inflammation. The observed trend for gene expression downregulation of IL-8 in our study, which is involved in neutrophil recruitment, might be linked to its regulation by the TLR2/MyD88/NF-κB signaling pathway (101). Although the influence of Zn on this pathway is not well established, other studies have shown that ZnO supplementation downregulates TLR2 and TLR4 expression (102) and influences MAPK signaling pathway in weaned piglets (91). We observed a TLR2 downregulation in the ileum of ZnO-fed piglets, which is noteworthy considering that TLR2 mRNA expression is naturally high in the Peyer’s patches of the ileum (103). The suppression of TLR2 expression in ileum by ZnO may consequently attenuate NF-κB activation, thereby reducing the production of pro-inflammatory cytokines (85, 104). As it is known that Zn deficiency activates NF-κB and MAPK signaling (105), our findings reinforce the immunoregulatory role of ZnO in supporting both innate and adaptive immunity, helping to alleviate intestinal inflammation and enhance gut defense mechanisms in weaned piglets.
In the cecum, TLR2 was upregulated in ZnO fed piglets, suggesting an enhanced state of immune readiness in the hindgut. TLR2 triggers inflammatory and immune reactions by recognizing diverse microbial components (106). This expression in the cecum may be driven by the dense and diverse local microbiota, which provides potent TLR2 ligands like peptidoglycan and lipoproteins and determines the region-specific immune status (106–108). In fact, expression patterns of intestinal TLRs differ significantly across gut segments, with microbiota composition and physiological events such as weaning influencing markedly higher expression in the colon and cecum (109–111). This might explain our observed downregulation in the ileum compared to the upregulation in cecum with the use of ZnO. Considering that ZnO is known to alter gut microbiota, the amplified TLR2 signaling in the cecum may therefore reflect the site-specific immunomodulation driven by diet-induced microbial changes, thereby potentially contributing to improved mucosal barrier integrity, gut health and PWD management (102, 112, 113).
The gene PPARGC1A, also known as PGC-1α, was the only upregulated gene in the ileum of ZnO supplemented piglets. PPARGC1A encodes a transcriptional coactivator that interacts with PPARγ, regulating genes involved in lipid metabolism, ATP production, and glucose homeostasis, while also suppressing inflammation through negative crosstalk with the NF-κB signaling pathway (114, 115). Supporting this role, Handschin et al. (116), showed that mice lacking PGC-1α in skeletal muscle exhibited elevated circulating IL-6 and impaired glucose regulation, while Pérez et al. (117), demonstrated that PGC-1α deficiency in the pancreas amplified NF-κB-mediated IL-6 expression and worsened inflammation during pancreatitis. In our study, we observed upregulation of PPARGC1A in the ileum and downregulation of IL-6 expression in the jejunum of ZnO-supplemented piglets, suggesting a broader anti-inflammatory effect of ZnO, potentially mediated through PGC-1α–dependent suppression of NF-κB and IL-6 signaling. This is supported by evidence that PPAR signaling requires Zn to exert its anti-inflammatory effects (115). Previous studies have shown that Zn increased the mRNA expression of PPARγ in weaned piglets (118), while Zn deficiency decreased PPARγ signaling in porcine endothelial cells (119). In a study by Bao et al. (114), Zn increased PPARGC1A expression in the ileum of elderly humans, while weaning in piglets has been shown to reduce the expression of PGC-1α (120). The current study, therefore, offers key evidence that ZnO may support PPARγ signaling, contributing to its anti-inflammatory and antioxidant actions, likely through NF-κB inhibition.
Collectively, these findings support the reported immunomodulatory and antimicrobial role of ZnO by modulating innate immunity, probably by reducing pathogenic bacterial loads and stabilizing microbiota consistently across the small intestine (2, 21, 121). From our findings, it could be speculated that ZnO dampens inflammatory responses across the small intestine by inhibiting the NF-κB signaling pathway and the Th17 response through STAT3 inactivation, as suggested by the reduced expression of REG3G, to achieve gut homeostasis in a multi-faceted way. The inhibition of NF-κB by Zn via TLR, calprotectin and PPARγ signaling pathways is therefore a possible mechanism through which Zn attenuates pro-inflammatory processes and mitigates oxidative stress. The pronounced modulation observed in jejunum, followed by some similar but lower influences in ileum, with minimal changes in the caecum, may be attributed to their critical functions as the primary segments exposed to dietary changes after weaning, making them more susceptible to ZnO. Importantly, our study provides novel intestinal segment-specific molecular insights that can be used when evaluating potential alternatives to ZnO, with the objective of replicating its beneficial effects on gut health. We also note the need to maintain a delicate balance, where host immune cells require an adequate Zn supply to sustain an effective immune response against stressors, while simultaneously restricting Zn availability to pathogens and toxicity risks.
However, we acknowledge that a key limitation of our study is that protein levels and activity were not evaluated to establish causality. While transcriptomic analysis provides valuable insight into potential mechanisms, mRNA levels do not always translate directly to abundance or functional state of proteins due to complex post-transcriptional and post-translational regulation. Therefore, future research is warranted to validate our segment-specific findings at the protein level using targeted proteomic and functional assays.
5 Conclusion
The understanding of the molecular mechanisms by which ZnO preserves gut health is essential for the identification of pharmacological targets and the development of alternatives. Pharmacological levels of ZnO improved performance and fecal consistency in weaned piglets. Additionally, ZnO modulated the expression of genes related to the maintenance of intestinal health, mainly in the ileum and the jejunum. These include the suppression of oxidative stress (GPX2) and the modulation of gut microbiota (REG3G) and innate immunity (TLR2). ZnO reduced the gene expression of pro- and anti-inflammatory cytokines (IL8, IL6, IL22, IL17A, IL1B, and TGFB1), reduced Zn transport and absorption (SLC39A4), enhanced mitochondrial function (PPARGC1A), and attenuated neutrophil-mediated inflammation (serum calprotectin). These findings are indicative of the roles of ZnO to regulate different pathways such as NF-κB, STAT3, Th17/Treg balance, and PPARγ signaling pathway. Collectively, the performance benefits of dietary supplementation with high doses of ZnO in weaning piglets may be explained through the maintenance of intestinal homeostasis, particularly that of the gut immune function, resulting from the anti-inflammatory, antioxidant, and antimicrobial properties of ZnO.
Data availability statement
The datasets used and/or analyzed to support the findings of this study are available from the corresponding author on a reasonable request.
Ethics statement
The animal study was approved by IRTA’s Ethical Committee for Animal Experimentation and the Commission on Animal Experimentation of Generalitat de Catalunya (Project number 11766). The studies were conducted in accordance with the local legislation and institutional requirements.
Author contributions
ZN: Conceptualization, Data curation, Formal analysis, Investigation, Methodology, Software, Writing – original draft. NT: Conceptualization, Formal analysis, Investigation, Writing – review & editing. MB: Formal analysis, Methodology, Software, Validation, Writing – review & editing. JL: Formal analysis, Writing – review & editing. BJ-M: Investigation, Methodology, Writing – review & editing. RB-D: Formal analysis, Investigation, Supervision, Writing – review & editing. DT: Conceptualization, Formal analysis, Funding acquisition, Investigation, Methodology, Project administration, Supervision, Writing – review & editing. JT: Conceptualization, Data curation, Formal analysis, Investigation, Methodology, Supervision, Visualization, Writing – review & editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. ZN received funding from the European Union’s Horizon 2020 research and innovation programme under the Marie Skłodowska-Curie grant agreement No. 945413 and from the Universitat Rovira i Virgili (URV) and the Institute of Agrifood Research and Technology (IRTA).
Acknowledgments
The authors would like to thank the barn staff at IRTA-Facilities, for taking care of the animal management and its support during sampling. The authors also want to thank laboratory staff at IRTA-Animal Nutrition and Animal Breeding and Genetics programmes for laboratory analysis of the samples provided. Special thanks to Núria Paris and Olga González for their technical support especially in the microfluidic assay.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The authors declare that no Gen AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fvets.2025.1645900/full#supplementary-material
Footnotes
References
1. Sales, J. Effects of pharmacological concentrations of dietary zinc oxide on growth of post-weaning pigs: a meta-analysis. Biol Trace Elem Res. (2013) 152:343–9. doi: 10.1007/s12011-013-9638-3
2. Tang, X, Xiong, K, Zeng, Y, and Fang, R. The mechanism of zinc oxide in alleviating diarrhea in piglets after weaning: a review from the perspective of intestinal barrier function. Int J Mol Sci. (2024) 25:10040. doi: 10.3390/ijms251810040
3. Duffy, R, Yin, M, and Redding, LE. A review of the impact of dietary zinc on livestock health. J Trace Elem Miner. (2023) 5:100085. doi: 10.1016/j.jtemin.2023.100085
4. Poulsen, HD. Zinc oxide for weanling piglets. Acta Agric Scand A Anim Sci. (1995) 45:159–67. doi: 10.1080/09064709509415847
5. Campbell, JM, Crenshaw, JD, and Polo, J. The biological stress of early weaned piglets. J Anim Sci Biotechnol. (2013) 4:1–4. doi: 10.1186/2049-1891-4-19
6. Pluske, JR, Hampson, DJ, and Williams, IH. Factors influencing the structure and function of the small intestine in the weaned pig: a review. Livest Prod Sci. (1997) 51:215–36. doi: 10.1016/S0301-6226(97)00057-2
7. Moeser, AJ, Pohl, CS, and Rajput, M. Weaning stress and gastrointestinal barrier development: implications for lifelong gut health in pigs. Animal Nutr. (2017) 3:313–21. doi: 10.1016/j.aninu.2017.06.003
8. Hu, CH, Xiao, K, Luan, ZS, and Song, J. Early weaning increases intestinal permeability, alters expression of cytokine and tight junction proteins, and activates mitogen-activated protein kinases in pigs. J Anim Sci. (2013) 91:1094–101. doi: 10.2527/jas.2012-5796
9. Tang, X, Xiong, K, Fang, R, and Li, M. Weaning stress and intestinal health of piglets: a review. Front Immunol. (2022) 13:1042778. doi: 10.3389/fimmu.2022.1042778
10. Hill, GM, Mahan, DC, Carter, SD, Cromwell, GL, Ewan, RC, Harrold, RL, et al. Effect of pharmacological concentrations of zinc oxide with or without the inclusion of an antibacterial agent on nursery pig performance. J Anim Sci. (2001) 79:934–41. doi: 10.2527/2001.794934x
11. Bonetti, A, Tugnoli, B, Piva, A, and Grilli, E. Towards zero zinc oxide: feeding strategies to manage post-weaning diarrhea in piglets. Animals. (2021) 11:642. doi: 10.3390/ani11030642
12. Aimo, L, Cherr, GN, and Oteiza, PI. Low extracellular zinc increases neuronal oxidant production through nadph oxidase and nitric oxide synthase activation. Free Radic Biol Med. (2010) 48:1577–87. doi: 10.1016/j.freeradbiomed.2010.02.040
13. Gammoh, NZ, and Rink, L. Zinc and the immune system. In: M Mahmoudi and N Rezaei, editors. Nutrition and immunity. Cham: Springer (2019). 127–58.
14. Pearce, SC, Sanz Fernandez, MV, Torrison, J, Wilson, ME, Baumgard, LH, and Gabler, NK. Dietary organic zinc attenuates heat stress–induced changes in pig intestinal integrity and metabolism. J Anim Sci. (2015) 93:4702–13. doi: 10.2527/jas.2015-9018
15. Højberg, O, Canibe, N, Poulsen, HD, Hedemann, MS, and Jensen, BB. Influence of dietary zinc oxide and copper sulfate on the gastrointestinal ecosystem in newly weaned piglets. Appl Environ Microbiol. (2005) 71:2267–77. doi: 10.1128/AEM.71.5.2267-2277.2005
16. Li, BT, Van Kessel, AG, Caine, WR, Huang, SX, and Kirkwood, RN. Small intestinal morphology and bacterial populations in ileal digesta and feces of newly weaned pigs receiving a high dietary level of zinc oxide. Can J Anim Sci. (2001) 81:511–6. doi: 10.4141/A01-043
17. Siddiqi, KS, Ur Rahman, A, and Tajuddin, HA. Properties of zinc oxide nanoparticles and their activity against microbes. Nanoscale Res Lett. (2018) 13:1–13. doi: 10.1186/s11671-018-2532-3
18. Zhu, C, Lv, H, Chen, Z, Wang, L, Wu, X, Chen, Z, et al. Dietary zinc oxide modulates antioxidant capacity, small intestine development, and jejunal gene expression in weaned piglets. Biol Trace Elem Res. (2017) 175:331–8. doi: 10.1007/s12011-016-0767-3
19. Grilli, E, Tugnoli, B, Vitari, F, Domeneghini, C, Morlacchini, M, Piva, A, et al. Low doses of microencapsulated zinc oxide improve performance and modulate the ileum architecture, inflammatory cytokines and tight junctions expression of weaned pigs. Animal. (2015) 9:1760–8. doi: 10.1017/S1751731115001329
20. Zhang, B, and Guo, Y. Supplemental zinc reduced intestinal permeability by enhancing occludin and zonula occludens protein-1 (ZO-1) expression in weaning piglets. Br J Nutr. (2009) 102:687–93. doi: 10.1017/S0007114509289033
21. Roselli, M, Finamore, A, Garaguso, I, Britti, MS, and Mengheri, E. Zinc oxide protects cultured enterocytes from the damage induced by Escherichia coli. J Nutr. (2003) 133:4077–82. doi: 10.1093/jn/133.12.4077
22. Wessels, I, Maywald, M, and Rink, L. Zinc as a gatekeeper of immune function. Nutrients. (2017) 9:1286. doi: 10.3390/nu9121286
23. Sargeant, HR, McDowall, KJ, Miller, HM, and Shaw, MA. Dietary zinc oxide affects the expression of genes associated with inflammation: transcriptome analysis in piglets challenged with ETEC K88. Vet Immunol Immunopathol. (2010) 137:120–9. doi: 10.1016/j.vetimm.2010.05.001
24. Sargeant, HR, Miller, HM, and Shaw, MA. Inflammatory response of porcine epithelial IPEC J2 cells to enterotoxigenic E. coli infection is modulated by zinc supplementation. Mol Immunol. (2011) 48:2113–21. doi: 10.1016/j.molimm.2011.07.002
25. Wang, X, Ou, D, Yin, J, Wu, G, and Wang, J. Proteomic analysis reveals altered expression of proteins related to glutathione metabolism and apoptosis in the small intestine of zinc oxide-supplemented piglets. Amino Acids. (2009) 37:209–18. doi: 10.1007/s00726-009-0242-y
26. Martínez-Montemayor, MM, Hill, GM, Raney, NE, Rilington, VD, Tempelman, RJ, Link, JE, et al. Gene expression profiling in hepatic tissue of newly weaned pigs fed pharmacological zinc and phytase supplemented diets. BMC Genomics. (2008) 9:1–14. doi: 10.1186/1471-2164-9-421
27. Starke, IC, Pieper, R, Neumann, K, Zentek, J, and Vahjen, W. The impact of high dietary zinc oxide on the development of the intestinal microbiota in weaned piglets. FEMS Microbiol Ecol. (2014) 87:416–27. doi: 10.1111/1574-6941.12233
28. Schokker, D, Kar, SK, Willems, E, Bossers, A, Dekker, RA, and Jansman, AJM. Dietary supplementation of zinc oxide modulates intestinal functionality during the post-weaning period in clinically healthy piglets. J Anim Sci Biotechnol. (2023) 14:1–16. doi: 10.1186/s40104-023-00925-1
29. Pieper, R, Vahjen, W, Neumann, K, Van Kessel, AG, and Zentek, J. Dose-dependent effects of dietary zinc oxide on bacterial communities and metabolic profiles in the ileum of weaned pigs. J Anim Physiol Anim Nutr (Berl). (2012) 96:825–33. doi: 10.1111/j.1439-0396.2011.01231.x
30. Assadi, ES, Nielsen, TS, Hedemann, MS, and Curtasu, MV. Integrated faecal microbiota and blood metabolic changes following different dietary zinc oxide levels in weaned piglets. Sci Rep. (2025) 15:12089. doi: 10.1038/s41598-025-03103-7
31. Broom, LJ, Miller, HM, Kerr, KG, and Toplis, P. Removal of both zinc oxide and avilamycin from the post-weaning piglet diet: consequences for performance through to slaughter. Anim Sci. (2003) 77:79–84. doi: 10.1017/S1357729800053674
32. Heo, JM, Opapeju, FO, Pluske, JR, Kim, JC, Hampson, DJ, and Nyachoti, CM. Gastrointestinal health and function in weaned pigs: a review of feeding strategies to control post-weaning diarrhoea without using in-feed antimicrobial compounds. J Anim Physiol Anim Nutr (Berl). (2013) 97:207–37. doi: 10.1111/j.1439-0396.2012.01284.x
33. Hansen, SV, Nørskov, NP, Nørgaard, JV, Woyengo, TA, Poulsen, HD, and Nielsen, TS. Determination of the optimal level of dietary zinc for newly weaned pigs: a dose-response study. Animals. (2022) 12:1552. doi: 10.3390/ani12121552
34. Burrough, ER, De Mille, C, and Gabler, NK. Zinc overload in weaned pigs: tissue accumulation, pathology, and growth impacts. J Vet Diagn Invest. (2019) 31:537–45. doi: 10.1177/1040638719852144
35. Komatsu, T, Sugie, K, Inukai, N, Eguchi, O, Oyamada, T, Sawada, H, et al. Chronic pancreatitis in farmed pigs fed excessive zinc oxide. J Vet Diagn Invest. (2020) 32:689–94. doi: 10.1177/1040638720944368
36. Zhou, X, Li, Y, Li, Z, Cao, Y, Wang, F, and Li, CM. Effect of dietary zinc on morphological characteristics and apoptosis related gene expression in the small intestine of Bama miniature pigs. Acta Histochem. (2017) 119:235–43. doi: 10.1016/j.acthis.2017.01.006
37. Brugger, D, and Windisch, WM. Subclinical zinc deficiency impairs pancreatic digestive enzyme activity and digestive capacity of weaned piglets. Br J Nutr. (2016) 116:425–33. doi: 10.1017/S0007114516002105
38. Jondreville, C, Revy, PS, and Dourmad, JY. Dietary means to better control the environmental impact of copper and zinc by pigs from weaning to slaughter. Livest Prod Sci. (2003) 84:147–56. doi: 10.1016/j.livprodsci.2003.09.011
39. Ciesinski, L, Guenther, S, Pieper, R, Kalisch, M, Bednorz, C, and Wieler, LH. High dietary zinc feeding promotes persistence of multi-resistant E. coli in the swine gut. PLoS One. (2018) 13:e0191660. doi: 10.1371/journal.pone.0191660
40. Vahjen, W, Pietruszyńska, D, Starke, IC, and Zentek, J. High dietary zinc supplementation increases the occurrence of tetracycline and sulfonamide resistance genes in the intestine of weaned pigs. Gut Pathog. (2015) 7:1–5. doi: 10.1186/s13099-015-0071-3
41. Bednorz, C, Oelgeschläger, K, Kinnemann, B, Hartmann, S, Neumann, K, Pieper, R, et al. The broader context of antibiotic resistance: zinc feed supplementation of piglets increases the proportion of multi-resistant Escherichia coli in vivo. Int J Med Microbiol. (2013) 303:396–403. doi: 10.1016/j.ijmm.2013.06.004
42. European Commission. Commission implementing regulation (EU) 2016/1095 of 6 July 2016 concerning the authorisation of zinc acetate dihydrate, zinc chloride anhydrous, zinc oxide, zinc sulphate heptahydrate, zinc sulphate monohydrate, zinc chelate of amino acids hydrate, zinc chelate of protein hydrolysates, zinc chelate of glycine hydrate (solid) and zinc chelate of glycine hydrate (liquid) as feed additives for all animal species and amending regulations (EC) No 1334/2003, (EC) No 479/2006, (EU) No 335/2010 and Implementing Regulations (EU) No 991/2012 and (EU) No 636/2013. Off J Eur Union (2016). L182:7–27. Available online at: https://eur-lex.europa.eu/eli/reg_impl/2016/1095/oj/eng (accessed February 5, 2025).
43. Guevarra, RB, Lee, JH, Lee, SH, Seok, MJ, Kim, DW, Kang, BN, et al. Piglet gut microbial shifts early in life: causes and effects. J Anim Sci Biotechnol. (2019) 10:1–10. doi: 10.1186/s40104-018-0308-3
44. Hansen, SV, Canibe, N, Nielsen, TS, and Woyengo, TA. Zinc status and indicators of intestinal health in enterotoxigenic Escherichia coli F18 challenged newly weaned pigs fed diets with different levels of zinc. J Anim Sci. (2024) 102:skae018. doi: 10.1093/jas/skae018
45. Pejsak, Z, Kaźmierczak, P, Butkiewicz, AF, Wojciechowski, J, and Woźniakowski, G. Alternatives to zinc oxide in pig production. Pol J Vet Sci. (2023) 26:319–30. doi: 10.24425/pjvs.2023.145033
46. Torrallardona, D. Spray dried animal plasma as an alternative to antibiotics in weanling pigs – a review. Asian Australas J Anim Sci. (2010) 23:131–48. doi: 10.5713/ajas.2010.70630
47. Luise, D, Correa, F, Negrini, C, Virdis, S, Mazzoni, M, Dalcanale, S, et al. Blend of natural and natural identical essential oil compounds as a strategy to improve the gut health of weaning pigs. Animal. (2023) 17:101031. doi: 10.1016/j.animal.2023.101031
48. López-Gálvez, G, López-Alonso, M, Pechova, A, Mayo, B, Dierick, N, and Gropp, J. Alternatives to antibiotics and trace elements (copper and zinc) to improve gut health and zootechnical parameters in piglets: a review. Anim Feed Sci Technol. (2021) 271:114727. doi: 10.1016/j.anifeedsci.2020.114727
49. FEDNA. Fundación Española para el Desarrollo de la Nutrición Animal. “Necesidades Nutricionales Para Ganado Porcino Normas.,” In: C. BlasDe, G.G. Mateos, and P García-Rebollar, editors. 2nd ed. Madrid, Spain (2013). Available online at: http://www.ciap.org.ar/Sitio/Archivos/Normas%20PORCINO_2013rev.pdf (accessed October 17, 2024).
50. Pedersen, KS, and Toft, N. Intra- and inter-observer agreement when using a descriptive classification scale for clinical assessment of faecal consistency in growing pigs. Prev Vet Med. (2011) 98:288–91. doi: 10.1016/j.prevetmed.2010.11.016
51. González-Solé, F, Criado-Mesas, L, Villodre, C, García, WC, Farré, M, Borda, E, et al. Porcine digestible peptides (PDP) in weanling diets regulates the expression of genes involved in gut barrier function, immune response and nutrient transport in nursery pigs. Animals. (2020) 10:2368. doi: 10.3390/ani10122368
52. Ballester, M, Cordón, R, and Folch, JM. DAG expression: high-throughput gene expression analysis of real-time PCR data using standard curves for relative quantification. PLoS One. (2013) 8:e80385. doi: 10.1371/journal.pone.0080385
53. Huang, F, Wang, Q, Wang, Z, Lv, L, and Feng, J. Effects of organic zinc on the growth performance of weanling pigs: a Meta-analysis. Biol Trace Elem Res. (2024) 202:5051–60. doi: 10.1007/s12011-024-04070-2
54. Hollis, GR, Carter, SD, Cline, TR, Crenshaw, TD, Cromwell, GL, Hill, GM, et al. Effects of replacing pharmacological levels of dietary zinc oxide with lower dietary levels of various organic zinc sources for weanling pigs. J Anim Sci. (2005) 83:2123–9. doi: 10.2527/2005.8392123x
55. Canibe, N, Højberg, O, Kongsted, H, Vodolazska, D, Lauridsen, C, Nielsen, TS, et al. Review on preventive measures to reduce post-weaning Diarrhoea in piglets. Animals. (2022) 12:2585. doi: 10.3390/ani12192585
56. Kim, S, Jae, KCH, Park, BC, Lee, CY, and Han, JH. Effects of a lipid-encapsulated zinc oxide dietary supplement, on growth parameters and intestinal morphology in weanling pigs artificially infected with enterotoxigenic Escherichia coli. J Anim Sci Technol. (2015) 57:1–5. doi: 10.1186/s40781-014-0038-9
57. Mendes, CR, Dilarri, G, Forsan, CF, De MR, SV, PRM, L, De Moraes, PB, et al. Antibacterial action and target mechanisms of zinc oxide nanoparticles against bacterial pathogens. Sci Rep. (2022) 12:1–10. doi: 10.1038/s41598-022-06657-y
58. Ortiz Sanjuán, JM, Argüello, H, Cabrera-Rubio, R, Crispie, F, Cotter, PD, Garrido, JJ, et al. Effects of removing in-feed antibiotics and zinc oxide on the taxonomy and functionality of the microbiota in post weaning pigs. Anim Microbiome. (2024) 6:32. doi: 10.1186/s42523-024-00306-7
59. Damo, SM, Kehl-Fie, TE, Sugitani, N, Holt, ME, Rathi, S, Murphy, WJ, et al. Molecular basis for manganese sequestration by calprotectin and roles in the innate immune response to invading bacterial pathogens. Proc Natl Acad Sci USA. (2013) 110:3841–6. doi: 10.1073/pnas.1220341110
60. Zackular, JP, Chazin, WJ, and Skaar, EP. Nutritional immunity: S100 proteins at the host-pathogen Interface. J Biol Chem. (2015) 290:18991–8. doi: 10.1074/jbc.R115.645085
61. Yui, S, Nakatani, Y, and Mikami, M. Calprotectin (S100A8/S100A9), an inflammatory protein complex from neutrophils with a broad apoptosis-inducing activity. Biol Pharm Bull. (2003) 26:753–60. doi: 10.1248/bpb.26.753
62. Dal Pont, GC, Belote, BL, Lee, A, Bortoluzzi, C, Eyng, C, Sevastiyanova, M, et al. Novel models for chronic intestinal inflammation in chickens: intestinal inflammation pattern and biomarkers. Front Immunol. (2021) 12:676628. doi: 10.3389/fimmu.2021.676628
63. González, R, Ceacero-Heras, D, Tena-Garitaonaindia, M, Álvarez-Mercado, A, Gámez-Belmonte, R, Chazin, WJ, et al. Intestinal inflammation marker calprotectin regulates epithelial intestinal zinc metabolism and proliferation in mouse jejunal organoids. Biomed Pharmacother. (2024) 174:116555. doi: 10.1016/j.biopha.2024.116555
64. Geiser, J, Venken, KJT, de Lisle, RC, and Andrews, GK. A mouse model of acrodermatitis Enteropathica: loss of intestine zinc transporter ZIP4 (Slc39a4) disrupts the stem cell niche and intestine integrity. PLoS Genet. (2012) 8:e1002766. doi: 10.1371/journal.pgen.1002766
65. Dufner-Beattie, J, Wang, F, Kuo, YM, Gitschier, J, Eide, D, and Andrews, GK. The acrodermatitis Enteropathica gene ZIP4 encodes a tissue-specific, zinc-regulated zinc transporter in mice. J Biol Chem. (2003) 278:33474–81. doi: 10.1074/jbc.M305000200
66. Hennigar, SR, Olson, CI, Kelley, AM, and McClung, JP. Slc39a4 in the small intestine predicts zinc absorption and utilization: a comprehensive analysis of zinc transporter expression in response to diets of varied zinc content in young mice. J Nutr Biochem. (2022) 101:108927. doi: 10.1016/j.jnutbio.2021.108927
67. Martin, L, Lodemann, U, Bondzio, A, Gefeller, EM, Vahjen, W, Aschenbach, JR, et al. A high amount of dietary zinc changes the expression of zinc transporters and metallothionein in jejunal epithelial cells in vitro and in vivo but does not prevent zinc accumulation in jejunal tissue of piglets. J Nutr. (2013) 143:1205–10. doi: 10.3945/jn.113.177881
68. Montoya, D, D’angelo, M, Martín-orúe, SM, Rodríguez-sorrento, A, Saladrigas-garcía, M, Araujo, C, et al. Effectiveness of two plant-based in-feed additives against an Escherichia coli F4 Oral challenge in weaned piglets. Animals. (2021) 11:2024. doi: 10.3390/ani11072024
69. Weaver, BP, Dufner-Beattie, J, Kambe, T, and Andrews, GK. Novel zinc-responsive post-transcriptional mechanisms reciprocally regulate expression of the mouse Slc39a4 and Slc39a5 zinc transporters (Zip4 and Zip5). Biol Chem. (2007) 388:1301–12. doi: 10.1515/BC.2007.149
70. Medida, RL, Sharma, AK, Guo, Y, Johnston, L, Urriola, PE, Gomez, A, et al. Dietary zinc restriction affects the expression of genes related to immunity and stress response in the small intestine of pigs. J Nutr Sci. (2022) 11:e104. doi: 10.1017/jns.2022.105
71. Mao, X, Kim, BE, Wang, F, Eide, DJ, and Petris, MJ. A histidine-rich cluster mediates the ubiquitination and degradation of the human zinc transporter, hZIP4, and protects against zinc cytotoxicity. J Biol Chem. (2007) 282:6992–7000. doi: 10.1074/jbc.M610552200
72. Maric, S, Restin, T, Muff, JL, Camargo, SM, Guglielmetti, LC, Holland-Cunz, SG, et al. Citrulline, biomarker of enterocyte functional mass and dietary supplement. Metabolism, transport, and current evidence for clinical use. Nutrients. (2021) 13:2794. doi: 10.3390/nu13082794
73. Soraci, AL, Decundo, JM, Dieguez, SN, Martínez, G, Pérez Gaudio, DS, Amanto, FA, et al. Citrullinemia is a suitable biomarker for post weaning performance in piglets under intensive farming. J Am Vet Med Assoc. (2023) 261:858–64. doi: 10.2460/javma.22.12.0543
74. Berkeveld, M, Langendijk, P, Verheijden, JHM, Taverne, MAM, Van Nes, A, Van Haard, P, et al. Citrulline and intestinal fatty acid-binding protein: longitudinal markers of postweaning small intestinal function in pigs? J Anim Sci. (2008) 86:3440–9. doi: 10.2527/jas.2008-1167
75. Berkeveld, M, Langendijk, P, Soede, NM, Kemp, B, Taverne, MAM, Verheijden, JHM, et al. Improving adaptation to weaning: effect of intermittent suckling regimens on piglet feed intake, growth, and gut characteristics. J Anim Sci. (2009) 87:3156–66. doi: 10.2527/jas.2008-1764
76. Corthésy, B. Role of secretory IgA in infection and maintenance of homeostasis. Autoimmun Rev. (2013) 12:661–5. doi: 10.1016/j.autrev.2012.10.012
77. Snoeck, V, Peters, IR, and Cox, E. The IgA system: a comparison of structure and function in different species. Vet Res. (2006) 37:455–67. doi: 10.1051/vetres:2006010
78. Orlans, E, Peppard, J, Reynolds, J, and Hall, J. Rapid active transport of immunoglobulin a from blood to bile. J Exp Med. (1978) 147:588–92. doi: 10.1084/jem.147.2.588
79. Vaerman, JP, Langendries, A, Pabst, R, and Rothkötter, HJ. Contribution of serum IgA to intestinal lymph IgA, and vice versa, in minipigs. Vet Immunol Immunopathol. (1997) 58:301–8. doi: 10.1016/S0165-2427(97)00041-X
80. Brown, WR, and Kloppel, TM. The liver and IgA: immunological, cell biological and clinical implications. Hepatology. (1989) 9:763–84. doi: 10.1002/hep.1840090518
81. Badia, R, Lizardo, R, Martinez, P, Badiola, I, and Brufau, J. The influence of dietary locust bean gum and live yeast on some digestive immunological parameters of piglets experimentally challenged with Escherichia coli. J Anim Sci. (2012) 90:260–2. doi: 10.2527/jas.53894
82. Ng’ang’a, ZW, Tous, N, Hussain, M, Ballester, M, Polo, J, Beltrán-Debón, R, et al. Spray-dried porcine plasma improves piglets’ performance and modulates gut immune-related genes in the first week post-weaning. Anim Biosci. (2025). doi: 10.5713/ab.25.0185
83. Ramis, G, Murciano, F, Orengo, J, González-Guijarro, B, Cuellar-Flores, A, Serrano, D, et al. Is Oral vaccination against Escherichia coli influenced by zinc oxide? Animals. (2023) 13:1754. doi: 10.3390/ani13111754
84. De Groot, N, Fariñas, F, Cabrera-Gómez, CG, Pallares, FJ, and Ramis, G. Weaning causes a prolonged but transient change in immune gene expression in the intestine of piglets. J Anim Sci. (2021) 99:1–12. doi: 10.1093/jas/skab065
85. Hu, CH, Song, ZH, Xiao, K, Song, J, Jiao, LF, and Ke, YL. Zinc oxide influences intestinal integrity, the expressions of genes associated with inflammation and TLR4-myeloid differentiation factor 88 signaling pathways in weanling pigs. Innate Immun. (2014) 20:478–86. doi: 10.1177/1753425913499947
86. Peng, P, Deng, D, Chen, S, Li, C, Luo, J, Romeo, A, et al. The effects of dietary porous zinc oxide supplementation on growth performance, inflammatory cytokines and tight junction’s gene expression in early-weaned piglets. J Nutr Sci Vitaminol (Tokyo). (2020) 66:311–8. doi: 10.3177/jnsv.66.311
87. Hu, C, Song, J, Li, Y, Luan, Z, and Zhu, K. Diosmectite–zinc oxide composite improves intestinal barrier function, modulates expression of pro-inflammatory cytokines and tight junction protein in early weaned pigs. Br J Nutr. (2013) 110:681–8. doi: 10.1017/S0007114512005508
88. Al-Sadi, R, Boivin, M, and Ma, T. Mechanism of cytokine modulation of epithelial tight junction barrier. Front Biosci. (2009) 14:2765–78. doi: 10.2741/3413
89. Hering, NA, Andres, S, Fromm, A, van Tol, EA, Amasheh, M, Mankertz, J, et al. Transforming growth factor-β, a whey protein component, strengthens the intestinal barrier by upregulating Claudin-4 in HT-29/B6 Cells1,2. J Nutr. (2011) 141:783–9. doi: 10.3945/jn.110.137588
90. Patnaude, L, Mayo, M, Mario, R, Wu, X, Knight, H, Creamer, K, et al. Mechanisms and regulation of IL-22-mediated intestinal epithelial homeostasis and repair. Life Sci. (2021) 271:119195. doi: 10.1016/j.lfs.2021.119195
91. Song, ZH, Xiao, K, Ke, YL, Jiao, LF, and Hu, CH. Zinc oxide influences mitogen-activated protein kinase and TGF-β1 signaling pathways, and enhances intestinal barrier integrity in weaned pigs. Innate Immun. (2015) 21:341–8. doi: 10.1177/1753425914536450
92. Wang, J, Zhao, X, and Wan, YY. Intricacies of TGF-β signaling in Treg and Th17 cell biology. Cell Mol Immunol. (2023) 20:1002–22. doi: 10.1038/s41423-023-01036-7
93. Kimura, A, and Kishimoto, T. IL-6: regulator of Treg/Th17 balance. Eur J Immunol. (2010) 40:1830–5. doi: 10.1002/eji.201040391
94. Mangan, PR, Harrington, LE, O’Quinn, DB, Helms, WS, Bullard, DC, Elson, CO, et al. Transforming growth factor-β induces development of the TH17 lineage. Nature. (2006) 441:231–4. doi: 10.1038/nature04754
95. Dünkelberg, S, Maywald, M, Schmitt, AK, Schwerdtle, T, Meyer, S, and Rink, L. The interaction of sodium and zinc in the priming of T cell subpopulations regarding Th17 and Treg cells. Mol Nutr Food Res. (2020) 64:1900245. doi: 10.1002/mnfr.201900245
96. Dudakov, JA, Hanash, AM, and Van Den Brink, MRM. Interleukin-22: immunobiology and pathology. Annu Rev Immunol. (2015) 33:747–85. doi: 10.1146/annurev-immunol-032414-112123
97. Kitabayashi, C, Fukada, T, Kanamoto, M, Ohashi, W, Hojyo, S, Atsumi, T, et al. Zinc suppresses Th17 development via inhibition of STAT3 activation. Int Immunol. (2010) 22:375–86. doi: 10.1093/intimm/dxq017
98. Liang, SC, Tan, XY, Luxenberg, DP, Karim, R, Dunussi-Joannopoulos, K, Collins, M, et al. Interleukin (IL)-22 and IL-17 are coexpressed by Th17 cells and cooperatively enhance expression of antimicrobial peptides. J Exp Med. (2006) 203:2271–9. doi: 10.1084/jem.20061308
99. Hendrikx, T, Duan, Y, Wang, Y, Oh, JH, Alexander, LM, Huang, W, et al. Bacteria engineered to produce IL-22 in intestine induce expression of REG3G to reduce ethanol-induced liver disease in mice. Gut. (2019) 68:1504–15. doi: 10.1136/gutjnl-2018-317232
100. McAleer, J, Choi, SM, Chen, K, Binnie, M, Sheppard, D, Qin, S, et al. Regenerating islet-derived 3-gamma regulates pulmonary Th17 immunity by altering the gut microbiome (120.37). J Immunol. (2012) 188:120.37. doi: 10.4049/jimmunol.188.Supp.120.37
101. Qin, Y, Li, H, and Qiao, J. TLR2/MyD88/NF-κB signalling pathway regulates IL-8 production in porcine alveolar macrophages infected with porcine circovirus 2. J Gen Virol. (2016) 97:445–52. doi: 10.1099/jgv.0.000345
102. Liu, P, Pieper, R, Rieger, J, Vahjen, W, Davin, R, Plendl, J, et al. Effect of dietary zinc oxide on morphological characteristics, mucin composition and gene expression in the colon of weaned piglets. PLoS One. (2014) 9:e91091. doi: 10.1371/journal.pone.0091091
103. Tohno, M, Shimosato, T, Moue, M, Aso, H, Watanabe, K, Kawai, Y, et al. Toll-like receptor 2 and 9 are expressed and functional in gut-associated lymphoid tissues of presuckling newborn swine. Vet Res. (2006) 37:791–812. doi: 10.1051/vetres:2006036
104. Magalhaes, JG, Tattoli, I, and Girardin, SE. The intestinal epithelial barrier: how to distinguish between the microbial flora and pathogens. Semin Immunol. (2007) 19:106–15. doi: 10.1016/j.smim.2006.12.006
105. Xu, Y, Li, A, Li, X, Deng, X, and Gao, XJ. Zinc deficiency induces inflammation and apoptosis via oxidative stress in the kidneys of mice. Biol Trace Elem Res. (2023) 201:739–50. doi: 10.1007/s12011-022-03166-x
106. Álvarez, B, Revilla, C, Doménech, N, Pérez, C, Martínez, P, Alonso, F, et al. Expression of toll-like receptor 2 (TLR2) in porcine leukocyte subsets and tissues. Vet Res. (2008) 39:27. doi: 10.1051/vetres:2007051
107. Takeda, K, and Akira, S. Toll-like receptors in innate immunity. Int Immunol. (2005) 17:1–14. doi: 10.1093/intimm/dxh186
108. Maradiaga, N, Aldridge, B, Zeineldin, M, and Lowe, J. Gastrointestinal microbiota and mucosal immune gene expression in neonatal pigs reared in a cross-fostering model. Microb Pathog. (2018) 121:27–39. doi: 10.1016/j.micpath.2018.05.007
109. Tao, X, Xu, Z, and Wan, J. Intestinal microbiota diversity and expression of pattern recognition receptors in newly weaned piglets. Anaerobe. (2015) 32:51–6. doi: 10.1016/j.anaerobe.2014.12.005
110. Price, AE, Shamardani, K, Lugo, KA, Deguine, J, Roberts, AW, Lee, BL, et al. A map of toll-like receptor expression in the intestinal epithelium reveals distinct spatial, cell type-specific, and temporal patterns. Immunity. (2018) 49:560–575.e6. doi: 10.1016/j.immuni.2018.07.016
111. Arnaud, AP, Rome, V, Richard, M, Formal, M, David-Le Gall, S, and Boudry, G. Post-natal co-development of the microbiota and gut barrier function follows different paths in the small and large intestine in piglets. FASEB J. (2020) 34:1430–46. doi: 10.1096/fj.201902514r
112. Xia, P, Wu, Y, Lian, S, Yan, L, Meng, X, Duan, Q, et al. Research progress on toll-like receptor signal transduction and its roles in antimicrobial immune responses. Appl Microbiol Biotechnol. (2021) 105:5341–55. doi: 10.1007/s00253-021-11406-8
113. Cario, E, Gerken, G, and Podolsky, DK. Toll-like receptor 2 controls mucosal inflammation by regulating epithelial barrier function. Gastroenterology. (2007) 132:1359–74. doi: 10.1053/j.gastro.2007.02.056
114. Bao, B, Prasad, AS, Beck, FWJ, Fitzgerald, JT, Snell, D, Bao, GW, et al. Zinc decreases C-reactive protein, lipid peroxidation, and inflammatory cytokines in elderly subjects: a potential implication of zinc as an atheroprotective agent. Am J Clin Nutr. (2010) 91:1634–41. doi: 10.3945/ajcn.2009.28836
115. Reiterer, G, Toborek, M, and Hennig, B. Peroxisome proliferator activated receptors α and γ require zinc for their anti-inflammatory properties in porcine vascular endothelial cells. J Nutr. (2004) 134:1711–5. doi: 10.1093/jn/134.7.1711
116. Handschin, C, Cheol, SC, Chin, S, Kim, S, Kawamori, D, Kurpad, AJ, et al. Abnormal glucose homeostasis in skeletal muscle–specific PGC-1α knockout mice reveals skeletal muscle–pancreatic β cell crosstalk. J Clin Invest. (2007) 117:3463–74. doi: 10.1172/JCI31785
117. Pérez, S, Rius-Pérez, S, Finamor, I, Martí-Andrés, P, Prieto, I, García, R, et al. Obesity causes PGC-1α deficiency in the pancreas leading to marked IL-6 upregulation via NF-κB in acute pancreatitis. J Pathol. (2019) 247:48–59. doi: 10.1002/path.5166
118. Zhang, HB, Wang, MS, Wang, ZS, Zhou, AM, Zhang, XM, Dong, XW, et al. Supplementation dietary zinc levels on growth performance, carcass traits, and intramuscular fat deposition in weaned piglets. Biol Trace Elem Res. (2014) 161:69–77. doi: 10.1007/s12011-014-0078-5
119. Meerarani, P, Reiterer, G, Toborek, M, and Hennig, B. Zinc modulates PPARγ signaling and activation of porcine endothelial cells. J Nutr. (2003) 133:3058–64. doi: 10.1093/jn/133.10.3058
120. Zhu, LH, Zhao, KL, Chen, XL, and Xu, JX. Impact of weaning and an antioxidant blend on intestinal barrier function and antioxidant status in pigs. J Anim Sci. (2012) 90:2581–9. doi: 10.2527/jas.2011-4444
Keywords: zinc oxide, gut health, gene expression, immune response, inflammation
Citation: Ng’ang’a ZW, Tous N, Ballester M, Leskovec J, Jimenez-Moya B, Beltrán-Debón R, Torrallardona D and Tarradas J (2025) Impact of zinc oxide on gut health, immunity, and growth in weaned piglets: exploring potential modes of action. Front. Vet. Sci. 12:1645900. doi: 10.3389/fvets.2025.1645900
Edited by:
Jinsu Hong, University of Minnesota Twin Cities, United StatesReviewed by:
Hyunjun Choi, North Carolina State University, United StatesTingting Ju, Purdue University, United States
Copyright © 2025 Ng’ang’a, Tous, Ballester, Leskovec, Jimenez-Moya, Beltrán-Debón, Torrallardona and Tarradas. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Joan Tarradas, am9hbi50YXJyYWRhc0BpcnRhLmNhdA==
†These authors have contributed equally to this work and share last authorship