 Raimundo Nonato Colares Camargo-Júnior1,2*
Raimundo Nonato Colares Camargo-Júnior1,2* Cláudio Vieira de Araújo3
Cláudio Vieira de Araújo3 Marina de Nadai Bonin Gomes4
Marina de Nadai Bonin Gomes4 José Ribamar Felipe Marques5
José Ribamar Felipe Marques5 Welligton Conceição da Silva2
Welligton Conceição da Silva2 Carlos Eduardo Lima Sousa6Rubens Lima de Andrade6Albiane Sousa de Oliveira4
Carlos Eduardo Lima Sousa6Rubens Lima de Andrade6Albiane Sousa de Oliveira4 Éder Bruno Rebelo da Silva1Jaqueline Rodrigues Ferreira Cara4José de Brito Lourenço-Júnior1Alison Miranda Santos1
Éder Bruno Rebelo da Silva1Jaqueline Rodrigues Ferreira Cara4José de Brito Lourenço-Júnior1Alison Miranda Santos1 André Guimaraes Maciel e Silva1
André Guimaraes Maciel e Silva1- 1Postgraduate Program in Animal Science (PPGCAN), Institute of Veterinary Medicine, Federal University of Para (UFPA), Federal Rural University of the Amazon (UFRA), Brazilian Agricultural Research Corporation (EMBRAPA), Castanhal, Brazil
- 2Federal Institute of Education, Science and Technology of Pará (IFPA), Santarém, Brazil
- 3Department of Agricultural and Environmental Sciences, Federal University of Mato Grosso (UFMT), Sinop, Brazil
- 4Meat Technology and Processing Laboratory of the College of Veterinary Medicine and Animal Science, Universidade Federal do Mato Grosso do Sul, Campo Grande, Brazil
- 5Embrapa Eastern Amazon, Belém, Brazil
- 6Department of Veterinary Medicine, University Center of the Amazon (UNAMA), Santarem, Brazil
The genetic evaluation of Murrah buffaloes can be optimized by associating milk production, genetic value of sires, and age at first calving. Therefore, the aim of this study was to compare the Linear Mixed Model with the Threshold Model and their genetic association with milk production and the genetic evaluation of sires in the estimation of variance components of age at first calving in Murrah buffalo. The dataset comprised information on total milk production and age at first calving of Murrah buffaloes. The mixed linear animal model, designated as Model 1, was employed to estimate variance components. In a subsequent analysis, designated as Model 2, the age at first calving was examined in conjunction with the milk production. The variance components were obtained by Bayesian inference, using the Gibbs sampler to obtain posterior means. The t-test was then applied in order to compare the means of two samples. The additive genetic correlations between milk production and age at first calving were low in both models, with values equal to 0.11 and 0.17 for Models 1 and 2, respectively. The descriptive analysis of the predicted breeding values revealed that, irrespective of the model, the values for milk production exhibited minimal variation. In a separate analysis, Model 2 exhibited a reduced amplitude for age at first calving and enhanced prediction accuracy, particularly for sires with negative breeding values for this trait. Consequently, the Threshold Model strategy for analyzing age at first calving variance components is more efficient than a Linear Mixed Model. It provides more accurate genetic value estimates for sires without affecting milk production predictions.
1 Introduction
Research on Murrah buffaloes is justified by their importance as a dairy breed, entailing physiological and management complexities inherent to dairy buffalo farming. The relentless pursuit of high milk yields in Murrah buffaloes often is associated with specific reproductive challenges, such as prolonged anestrus, silent estrus, and reduced conception rates. These challenges directly affect overall productivity by prolonging calving intervals and decreasing lifetime production. Consequently, it is essential to focus jointly on reproductive disorders, along with the nutritional and management demands specific to the productive and reproductive needs of Murrah buffaloes. This joint approach can significantly increase reproductive efficiency and overall productivity in this specific breed. This, in turn, has the potential to generate substantial benefits for this segment of the buffalo agribusiness.
It is imperative to underscore certain limitations, including the occurrence of calving in buffaloes. As in cattle, challenges often arise, primarily dystocia due to fetal-maternal disproportion and nutritional deficiencies that lead to conditions such as placental retention or puerperal paresis (1, 2). These complications result in increased intervals between calving, compromised subsequent fertility, and high calf mortality (3, 4). Effective peripartum nutrition management and meticulous prepartum assessment are crucial (5, 6). Such difficulties can have a significant impact on herd productivity and economic viability, necessitating proactive clinical management (7).
Strategic supplementation with specific feed additives critically enhances buffalo reproductive health (8). Essential trace minerals (e.g., Se, Cu, Zn, Mn), vital vitamins (e.g., A, E, D), and omega-3 fatty acids optimize ovarian function and conception rates (5, 9). These nutrients bolster immune response, mitigate postpartum disorders, and improve overall reproductive efficiency (6, 10). By correcting nutritional deficiencies, these additives support the high metabolic demands of reproduction, making their precise dietary integration paramount for sustainable breeding success (4, 6).
During late gestation, particularly the final trimester, buffaloes are susceptible to prevalent metabolic diseases, including ketosis, hypocalcemia, and acidosis (4, 11). These conditions arise from metabolic imbalances like energy deficits, calcium dysregulation, or altered ruminal pH (8, 11, 12). Such disorders severely compromise maternal health, impair immune function, and jeopardize fetal viability. Consequently, parturition success, subsequent lactation, and reproductive performance are critically impacted, necessitating proactive nutritional and management interventions to mitigate adverse outcomes (13).
These interventions can and should benefit from promising research results in dairy farming. For example, it has been demonstrated in dairy calf breeds (14) that organic mineral supplementation is an effective strategy for improving mineral bioavailability and supporting the health of these animals during their first years of life. In another analysis, a feed additive mixture of condensed and hydrolysable tannins (15) improved reproductive efficiency in lactating cows, reducing the number of services per conception.
Other favorable results include the use of new methodologies, such as mid-infrared spectroscopy (16), to predict blood metabolic indicators in Holstein cow milk, which has proven to be an alternative for identifying cows at risk of negative energy balance and subclinical ketosis. Similarly, feed additives with immunomodulatory capabilities (17) are also emerging as strategies to improve metabolic and immunological responses to subacute ruminal acidosis in dairy cows.
Beyond these immediate clinical and nutritional interventions, the long-term genetic progress of a herd critically relies on the rigorous genetic evaluation of breeding animals, such as buffalo bulls (18, 19). It allows the selection of the best animals, resulting in superior productivity and quality (20).
By analyzing performance data and, with increasing frequency, genomic information, the genetic value of breeding animals can be estimated, reflecting their capacity to enhance characteristics such as milk production and age at first calving (21–23).
Consequently, selection for increased milk production is predicted to have a positive effect on the age at first calving, as demonstrated in Nili-Ravi (24) and Murrah (25) buffaloes. In other analyses, Tamboli et al. (26), Seno et al. (25), Mathur and RoyChoudhury (27), and Calanni Macchio et al. (28) evaluated data from Murrah buffaloes and Italian buffaloes. These analyses demonstrated that the opposite is also true; that is, selection by age at first calving showed improvement in milk production and other characteristics. This was evidenced by a negative association between age at first calving and standard milk production in the first lactation, productive life, total milk production throughout life, standard milk production throughout life, and reproductive efficiency. The findings of these studies indicate that decreasing the age at first calving would enhance various performance traits throughout the animal’s lifespan, extending beyond merely milk production.
It is imperative to note that this genetic evaluation is predicated on the precise quantification of genetic and environmental influences. Consequently, the estimation of variance components is essential. This approach enables precise measurement of additive genetic variance, as well as other crucial factors such as environmental variance and genotype-Environment interaction, as emphasized by Ranjan et al. (29) and Zhang et al. (30).
In addition to variance components, environmental factors, including calving period and season (31, 32) and age at first calving (32), were identified as significant determinants of performance. Furthermore, advanced evaluations consider direct, maternal, and permanent maternal effects (33) and employ methods such as Principal Component Analysis (33) to assess genetic trends and relationships between genetic values.
Among the widely used methods for estimating these components, the Linear Mixed Model and Threshold Models stand out. These are statistical tools used to analyze dependent data, i.e., data where observations are related or influence one another (34, 35). The aim was to compare the Linear Mixed Model with the Threshold Model and their genetic association with milk production and the genetic evaluation of sires in the estimation of variance components of age at first calving in Murrah buffalo.
2 Materials and methods
The analysis was based on a dataset comprising 2,866 records of total milk production and 661 records of age at first calving for 1,122 Murrah buffaloes participating in the Brazilian Buffalo Improvement Program (PROMEBULL).
Total milk production (TMP) was regressed as a function of lactation length (LL). Subsequently, the TMP was corrected for 305 days of lactation using the expression: MP305 = TMP + (5.42705) (305 − LL). This expression, referred to as “total milk yield,” offers a succinct representation of the relationship between TMP and LL. Consequently, MP305 is regarded as the rectified value for total milk yield, which will be utilized for subsequent analysis. This value is here in after referred to as milk production (MP).
Direct heterozygosity effect (HTZ) was estimated using breed information, defined as the deviation from the Murrah breed, which varies from zero (0) to one (1) (36).
The heterozygosity value (HTZ) was calculated using the equation (37) as follows:
Where:
MS is the degree of racial composition of the Murrah breed from the sire’s contribution.
OM is the degree of racial composition of the other breed from the dam’s contribution.
MM is the degree of racial composition of the Murrah breed from the dam’s contribution.
OS is the degree of racial composition of the other breed from the sire’s contribution.
The calving months were grouped into four calving seasons (CS) to adapt the analysis to regional climatic adversities, considered as follows: CS = 1 the period from January to March; CS = 2 the period from April to June; CS = 3 the period from July to September; and CS = 4 the period from October to December.
Contemporary groups were a combination of the fixed effects of herd, year, and CS. Contemporary group information with at least three observations was included.
The mixed linear animal model, designated as Model 1, was employed to estimate variance components. Subsequently, the method was employed to forecast breeding values, which are denoted as:
Where:
𝑦 is a vector of observations.
𝛽 is a vector of fixed effects (group of contemporaries and linear effect of the heterosis covariate).
𝑎 is the vector of direct additive genetic effect.
𝑒 is the vector of residual effect.
𝑋 is the incidence matrix that associates 𝛽 with 𝑦.
𝑍 is the incidence matrix of the direct genetic effect.
The age at first calving (AFC) was subsequently categorized into two classes. Class 1 encompassed values below the median, defined as less than or equal to 36.32 months of age. Class 2 comprised values greater than or equal to the median. In a subsequent analysis, designated as Model 2, the AFC was examined in conjunction with the MP. The AFC was conceptualized as a dichotomous variable, while the MP was considered a continuous variable within a Threshold Model. It was hypothesized that the scale underlying the AFC exhibited a continuous normal distribution and was represented as:
Where:
is the base scale vector; is the vector of location parameters (the true ‘unobserved’ breeding values); is the incidence matrix; is the identity matrix, is the residual variance.
The (co)variance components were obtained by Bayesian inference, using the Gibbs sampler with the GIBBS1F90 program (38) to obtain posterior means. A chain size of 600,000 cycles was used, with cycles saved every 20 cycles and a discard period of 60,000 initial cycles. The Geweke criterion (39) was employed to assess the quality of the chains, with a 5% probability level set as the cutoff.
In order to compare the posterior heritability means between the models, 30 sub-samples of 2,700 cycles were randomly obtained from the Gibbs chains for each model. The Gibbs chains were used to generate the posterior heritability means. The heritability averages were obtained from these subsamples, resulting in 30 values for each model. The t-test was then applied to compare the means of two samples, with a significance level of 0.05.
Subsequent to acquiring the predicted breeding values for MP and AFC for the sires, the Spearman correlations were calculated in three distinct scenarios and for each model. In the initial scenario, the sample was comprised of all sires with progenies exhibiting production. In the second scenario, the consideration was limited to sires with positive progenies for MP. Finally, the third scenario encompassed sires with progeny that were negative for AFC.
The estimated means and standard deviations of the minimum and maximum values observed for MP and AFC are shown in Table 1.
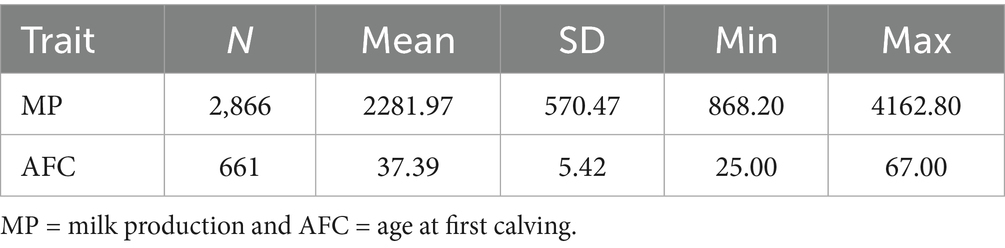
Table 1. Number of observations (N), estimates of means and standard deviation (SD), minimum (Min) and maximum (Max) values for MP and AFC.
3 Results and discussion
The estimation of the additive and residual genetic variance components for milk production (MP) and age at first calving (AFC) was demonstrated to be statistically robust using Bayesian inference with Markov chains via Monte Carlo (MCMC). The convergence of the chains was assessed by the Geweke test, whose values were found to be close to zero and with associated p-values exceeding 0.05 for all parameters. This indicates satisfactory convergence at the 5% significance level (see Table 2 for details).
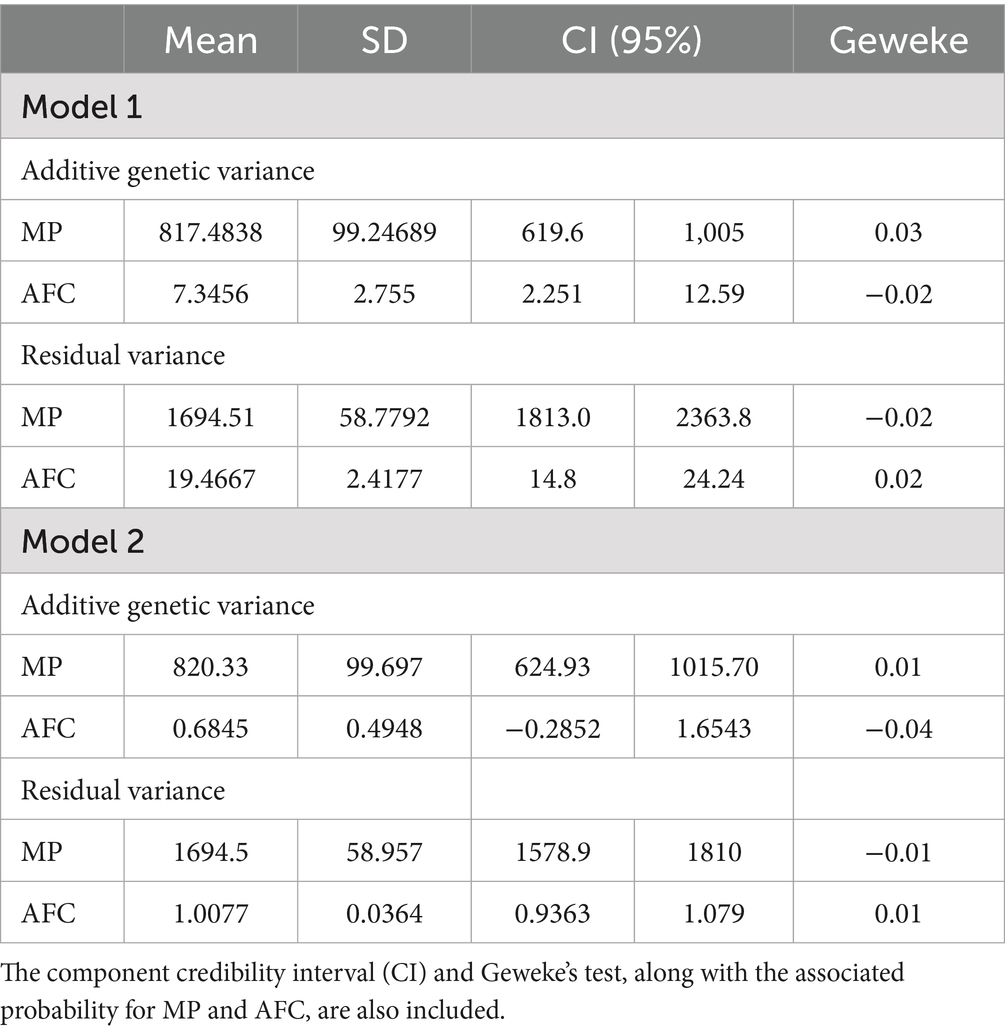
Table 2. The posterior means, along with their respective SD, are presented for the variance components of additive genetic and residual effects.
In Model 1, an additive genetic variance of 817.48 was observed for MP and 7.35 for AFC, with residual variances of 1694.51 and 19.47, respectively. In a separate analysis, Model 2 revealed that MP maintained comparable estimates for genetic (820.33) and residual (1694.5) variance, while AFC exhibited a substantial reduction in the genetic (0.68) and residual (1.01) components.
As demonstrated in Table 3, the posterior means for heritabilities are consistent across models for milk production, with a mean of 0.32 ± 0.03. However, for AFC, the estimates exhibited greater variability. The mean values obtained for Models 1 and 2 were 0.27 ± 0.09 and 0.37 ± 0.13, respectively. A t-test performed with independent subsamples of the Gibbs chain demonstrated a statistically significant discrepancy between the models for AFC (t = 187.53; p < 0.0001).
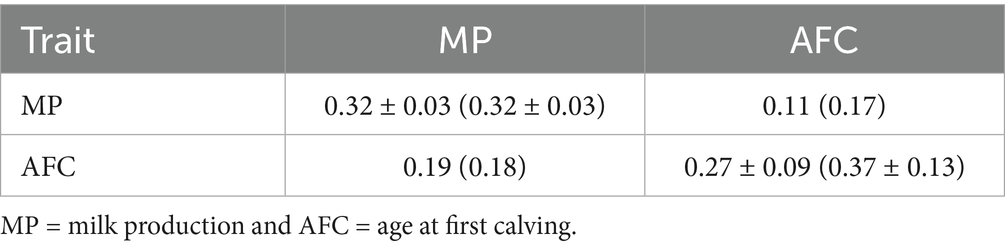
Table 3. The posterior means of heritability, with respective standard deviations, are presented on the diagonal for Model 1 and Model 2 (in parentheses), with the posterior means of additive genetic correlation for the Model 1 (above the diagonal) and Model 2 (below the diagonal) for milk production (MP) and age at first calving (AFC).
The additive genetic correlations between MP and AFC were low in both models, with values equal to 0.11 and 0.17 for Models 1 and 2, respectively. Spearman correlations between the estimated breeding values (see Table 4) provide further evidence for this pattern, with values ranging from 0.24 (Model 1) to 0.38 (Model 2). There is also high consistency between models in the ordering of the sires within each trait (ρ = 0.99 for MP and 0.72 for AFC).
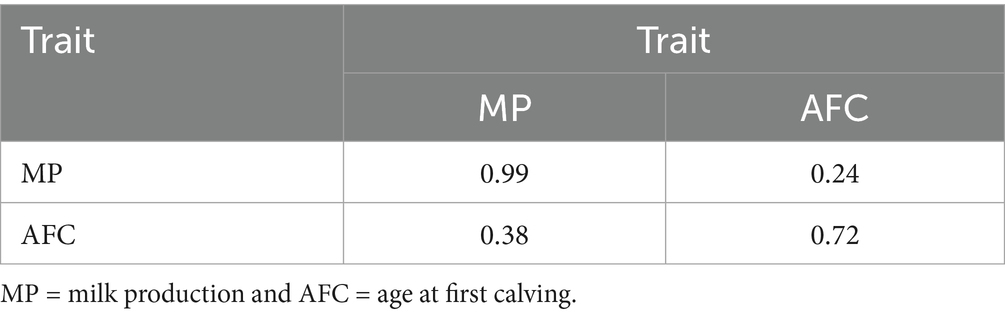
Table 4. Estimates of Spearman’s correlations between the breeding values of sires for MP and AFC are presented for Models 1 and 2, respectively, with the correlations between models illustrated on the diagonal and those between models on the above and below the diagonal.
The descriptive analysis presented in Table 5 revealed the descriptive analysis of the predicted breeding values revealed that, irrespective of the model, the values for MP demonstrated minimal variation. In a separate analysis, Model 2 exhibited a reduced amplitude for AFC and enhanced prediction accuracy, particularly for sires with negative breeding values for this trait. In all situations analyzed, the accuracy for MP remained constant (0.70–0.72), in contrast to the increase observed for AFC, which rose in Model 2 from 0.49 to 0.55.
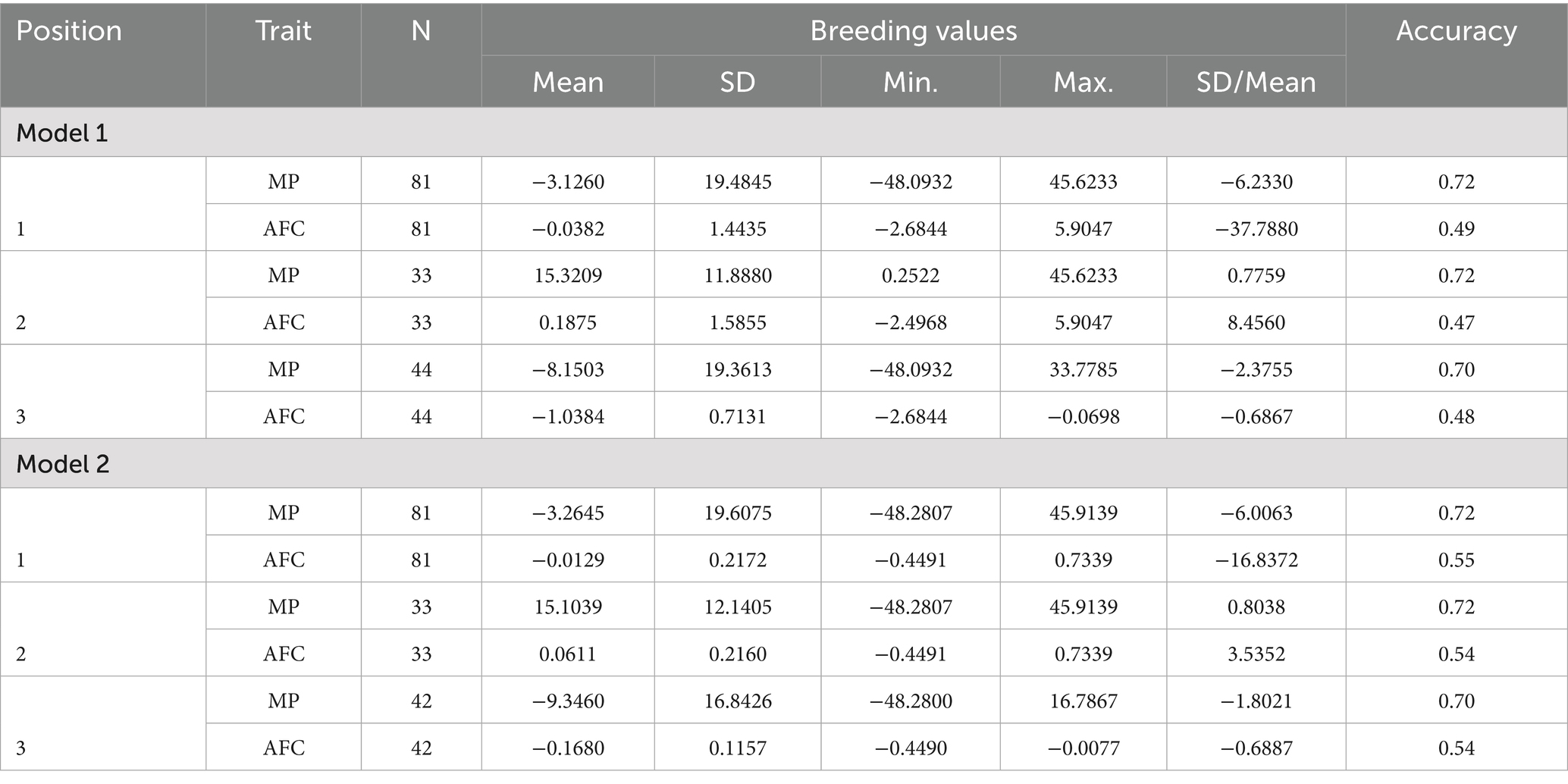
Table 5. Descriptive analysis of the Number of observations (N), minimum (Min) and maximum (Max) predicted breeding values of sires for MP and AFC are presented for Models 1 and 2, respectively.
The results demonstrated the robustness of the Bayesian approach in the inference of genetic components, as observed in other surveys (40, 41), particularly with regard to MP in cattle and buffaloes (42–45). In other words, the direct comparison made in this study between Linear Mixed Model and Threshold Models for the age at first calving in Murrah buffaloes, highlighting the Threshold Model as the best for evaluating the sires, represents a significant methodological contribution.
This assertion aligns with the conclusions of Colonia et al. (43), who previously corroborated the efficacy of Threshold Models in the analysis of non-normally distributed characteristics in dairy cattle, notwithstanding divergent prior specifications. While Camargo Júnior et al. (42), Kumar et al. (44), and Mendes-Malhado et al. (45) also employed Bayesian inference to estimate genetic parameters in dairy cattle (milk and growth traits), their focus differs, as they did not make this specific comparison between Linear Mixed Model and Threshold Models for discrete traits.
The low genetic correlations (0.11–0.17) observed between milk production and age at first calving in this study offer specific insights into Murrah reproduction. These insights are contextualized by the broader utility of Bayesian methodologies highlighted by Sun et al. (40) and van de Schoot et al. (41) for complex genetic analyses. The stability observed in the estimates of additive genetic variance and heritability between the two models evaluated suggests that this trait has a robust and well-defined genetic basis.
This consistency, even with different residual structures and parameterizations, indicates that MP is less susceptible to variations in the statistical model. This finding provides substantial credibility to genetic predictions for milk production (MP) and underscores its viability as a selection criterion in genetic enhancement programs for dairy buffalo farming (30, 46–48).
By contrast, AFC was highly sensitive to model specification. There was a marked reduction in additive genetic variance in Model 2, and the heritability estimates differed significantly. This discrepancy can be attributed to the trait’s lower heritability, its intrinsic biological complexity, and the greater influence of environmental and non-genetic factors (20, 26, 49, 50).
The greater accuracy of the Threshold Model for AFC possibly derives from its inherent ability to more effectively capture the underlying distribution of the characteristic when treated categorically, even though it is a biologically continuous trait. By dichotomizing AFC (below or above the median), the Threshold Model can effectively map crucial biological or management thresholds that a Linear Mixed Model would disregard. This approach makes the Threshold Model less sensitive to noise and nonlinearities often present in complex reproductive traits that are strongly influenced by environmental and non-genetic factors.
According to the results obtained, in the case of AFC, noise could come from several sources, such as small variations in nutritional management between animals, seasonal climatic differences not fully captured, data recording errors, or even small individual biological fluctuations that do not have a significant genetic component. By treating AFC as a categorical variable (e.g., early or late using the median as the threshold), the Threshold Model focuses on the classification that is most robust to random disturbances, filtering out the noise and highlighting the most important signal of precocity. Consequently, the model refines the signal-to-noise ratio, focusing on the qualitative outcome of early versus late calving, which has great practical relevance in buffalo selection.
Despite the similarity in the ratio between the standard deviation and the mean of heritability across models, indicating relative consistency in variability, the magnitude of the differences in means underscores the substantial impact of model alterations on genetic estimates for AFC. The present findings highlight the imperative for judiciousness in statistical modeling of reproductive traits, which characteristically manifest elevated environmental variability and diminished direct genetic control (44, 51, 52).
Additionally, it was evident that Model 2 showed a substantial reduction in genetic and residual variances for AFC. This suggests that the Threshold Model was more efficient in discriminating the true sources of variation, resulting in more accurate estimates of additive genetic effects and, consequently, greater predictive accuracy. This is because the Threshold Model deals with non-linearities by assuming that there is a continuous latent variable (not directly observable) – the predisposition or liability to AFC, which follows a normal distribution but manifests itself in observable categories when a certain threshold is exceeded.
By dichotomizing AFC, the Threshold Model implicitly recognizes that genetic impact can be better understood in terms of crossing a threshold. It can therefore better capture the idea that each additional day in AFC can change significantly once certain critical points (the thresholds) are reached, offering a more realistic representation of the biological complexity of this trait. The threshold model design assumes a continuous underlying predisposition or liability, but observes a categorical outcome, proving more suitable for addressing the biological complexity and low heritability typical of reproductive traits.
The estimates of additive genetic and Spearman rank-order correlations between MP and AFC were low. This finding indicates that there is a weak genetic association between these two traits. In practice, this indicates that ranking sires based on genetic merit for one trait does not serve as a reliable predictor of performance for the other (53, 54). Therefore, it is imperative to adopt selection indices that consider both traits jointly and in balance, to avoid limited or even undesirable gains in non-target traits (55–57).
The almost perfect consistency in the ranking of sires for MP between models (ρ = 0.99) confirms the robustness of genetic inference for this trait. This finding indicates that the selection of superior animals remains consistent, even when the model is modified. In another analysis for AFC, although the Spearman correlation between models was moderately high (ρ = 0.72), greater sensitivity to the adopted specifications was evident, indicating that minor changes to the model can alter the genetic ranking of individuals. This is particularly salient in the context of reproductive traits, where variations in the residual structure or the incorporation of supplementary effects can markedly enhance the precision of estimates (58–60).
Another salient point pertains to the analysis of the accuracy of genetic predictions. While the accuracy of MP remained stable for both models and the different sire subsets evaluated, Model 2 demonstrated significant enhancement in AFC accuracy. This finding indicates that the careful selection of the appropriate residual structure can facilitate the extraction of more valuable information from the database and improve the reliability of estimates for traits that are difficult to measure or have low heritability. Additionally, the diminished range of breeding values observed in Model 2 may suggest a more conservative yet precise modeling approach, a strategy that is particularly advantageous in selection contexts where there is a risk of overestimating genetic merit.
Consequently, the necessity of meticulous statistical modeling tailored to the biological characteristics of the studied traits is evident. While MP is a highly stable trait from a genetic point of view, AFC requires more refined analytical strategies that may include environmental, genomic, or management information to better capture its complexity (19, 61). In this sense, the utilization of Bayesian inference in conjunction with convergence verification and the implementation of statistical tests on Gibbs chain samples has been demonstrated to be a potent and adaptable approach (62–66). This approach has been demonstrated to produce reliable point estimates, thereby facilitating more informed decisions in the context of selection programs.
4 Conclusion
Overall, the Threshold Model strategy demonstrated superior efficiency in extracting variance components for AFC when directly compared to its treatment as a continuous variable within a Linear Mixed Model. The Threshold Model provided more accurate genetic value estimates for sires without affecting MP predictions.
The heritability estimates for MP indicate the possibility of substantial genetic progress through selective breeding. However, sires that demonstrated favorable genetic merit for precocity do not necessarily exhibit the greatest merit for MP.
In the pursuit of furthering the modeling of AFC, prospective investigations could entail the incorporation of more intricate environmental data, including temperature-humidity indices, precipitation, and variations in pasture quality or nutritional management during pivotal phases of animal development. This approach would facilitate the capture of environmental changes that exert a significant influence on AFC.
At the genomic level, the application of high-density SNP arrays or whole-genome sequencing would enable the identification of specific genes or Quantitative Trait Loci (QTLs) associated with precocity in buffaloes. The integration of these genomic data with advanced statistical methodologies, such as single-step genomic best linear unbiased prediction (ssGBLUP), holds considerable promise in enhancing the precision of genetic value predictions for AFC in buffalo. This approach would serve to reinforce and complement the advancements already demonstrated by the threshold model.
Data availability statement
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found in the article/supplementary material.
Ethics statement
Ethical approval was not required for the study involving animals in accordance with the local legislation and institutional requirements because this is a study generated from public data, and CEUA approval is not required.
Author contributions
RC-J: Software, Methodology, Writing – review & editing, Supervision, Investigation, Writing – original draft, Resources, Conceptualization, Formal analysis, Funding acquisition, Data curation, Visualization, Validation, Project administration. CA: Writing – review & editing, Investigation, Methodology, Validation, Writing – original draft, Formal analysis. MG: Writing – review & editing, Investigation, Methodology, Writing – original draft. JM: Formal analysis, Methodology, Writing – original draft, Writing – review & editing, Investigation. WS: Writing – review & editing, Methodology, Formal analysis, Investigation, Writing – original draft, Validation. CS: Writing – review & editing, Investigation, Methodology, Writing – original draft. RA: Investigation, Writing – review & editing, Methodology, Writing – original draft. AO: Writing – review & editing, Methodology, Writing – original draft, Investigation. ÉS: Methodology, Writing – original draft, Investigation, Writing – review & editing. JC: Methodology, Writing – original draft, Writing – review & editing, Investigation. JL-J: Investigation, Writing – original draft, Writing – review & editing, Validation, Methodology. AMS: Writing – original draft, Writing – review & editing, Investigation, Methodology. AGMS: Writing – original draft, Writing – review & editing, Methodology, Investigation.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This study was financed in part by the Universidade Federal de Mato Grosso do Sul -Brasil (UFMS) -Finance Code 001. This study was financed in part by the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior -Brasil (CAPES) -Finance Code 001.
Acknowledgments
We would like to express our sincere gratitude to the Coordination for the Improvement of Higher Education Personnel – Brazil (CAPES). We would like to express our sincere gratitude to the following institutions: Federal University of Pará (Brazil), Federal University of Mato Grosso do Sul (Brazil), and Federal University of Mato Grosso (Brazil). We thank EMBRAPA.
Conflict of interest
RJ, ÉS, JL-J, AMS, and AGMS were employed by Brazilian Agricultural Research Corporation (EMBRAPA).
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The authors declare that no Gen AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
1. Hiew, WHM. Prediction of parturition and dystocia in Holstein-friesian cattle, and cesarean section in dystocic beef cattle. West Lafayette: Purdue University (2014).
2. Murikipudi, NS, Injarapu, VSK, Kamineni, S, and Kumar, R. Parturient paresis (Milk fever) In: T Rana, editor. Periparturient diseases of cattle. New York: Wiley (2024). 67–76.
4. Mohd Azmi, AF, Ahmad, H, Mohd Nor, N, Goh, Y-M, Zamri-Saad, M, Abu Bakar, MZ, et al. The impact of feed supplementations on Asian buffaloes: a review. Animals. (2021) 11:2033. doi: 10.3390/ani11072033
5. Meghsham, MP. Evaluation of metabolic profile of purnathadi buffaloes during transition period. India: Master of Veterinary Science Master of Veterinary Science, Maharashtra Animal And Fishery Sciences University (2024).
6. Moghimi-Kandelousi, M, Alamouti, AA, Imani, M, and Zebeli, Q. A meta-analysis and meta-regression of the effects of vitamin E supplementation on serum enrichment, udder health, milk yield, and reproductive performance of transition cows. J Dairy Sci. (2020) 103:6157–66. doi: 10.3168/jds.2019-17556
7. Kumar, R, Sakhahari, DS, Ningwal, D, and Kumar, P. Injuries to the birth canal In: T Rana, editor. Periparturient diseases of cattle. Hoboken, New Jersey: Wiley Blackwell (2024). 153–64.
8. Chaudhary, SK, Dutta, N, Jadhav, SE, Singh, G, Singh, SK, and Tewari, D. Effect of strategic supplementation of a novel feed supplement on zootechnical attributes, metabolic profile, production and reproduction performance of buffaloes. Buf Bull. (2024) 43:209–25. doi: 10.56825/bufbu.2024.4325485
9. Thakur, D, Jain, RK, and Aich, R. Effect of strategic nutrient supplementation on health, reproductive and productive status of buffaloes in the Malwa region of Madhya Pradesh. Buf Bull. (2016) 35:225–36.
10. Anwar, F, Akhtar, M, Lal, C, Lodhi, L, Ayaz, M, Ahmad, I, et al. Effect of prepartum administration of selenium and vitamin E on subsequent postpartum performance in first calf Nili-Ravi buffalo. J Anim Plant Sci. (2014) 24:23–4.
11. Purohit, G, Gaur, M, Saraswat, C, and Bihani, D. Metabolic disorders in the parturient buffalo In: G Purohit, editor. Bubaline theriogenology. United States: International Veterinary Information Service (2013)
12. Purohit, GN. Bubaline Theriogenology. United States: International Veterinary Information Service (2019).
13. Ramadan, MH, Yousef, NaM, Mahmoud, A-EZE-A, Zeidan, AEB, Mickdam, EME, Ahmed, AE, et al. Effects of ketosis and hypocalcemia on the biochemical parameters and subsequent postpartum reproductive performance in buffaloes. SVU-Int J Vet Sci. (2023) 6:1–17. doi: 10.21608/svu.2023.157726.1224
14. Mortazavi, MS, Hajmohammadi, M, Buonaiuto, G, Colleluori, R, Lamanna, M, Cavallini, D, et al. The effect of a pre-mix of essential organic minerals on growth, antioxidant indices, and the diarrhea incidence in dairy calves breed in arid climates. Ruminants. (2025) 5:22. doi: 10.3390/ruminants5020022
15. Castillo, AR, Rienzo, JaD, and Cavallini, D. Effect of a mix of condense and hydrolysable tannins feed additive on lactating dairy cows’ services per conception and days open. Vet Anim Sci. (2025) 27:100434. doi: 10.1016/j.vas.2025.100434
16. Magro, S, Costa, A, Cavallini, D, Chiarin, E, and De Marchi, M. Phenotypic variation of dairy cows’ hematic metabolites and feasibility of non-invasive monitoring of the metabolic status in the transition period. Front Vet Sci. (2024) 11:1437352. doi: 10.3389/fvets.2024.1437352
17. Cavallini, D, Mammi, LME, Palmonari, A, García-González, R, Chapman, JD, Mclean, DJ, et al. Effect of an immunomodulatory feed additive in mitigating the stress responses in lactating dairy cows to a high concentrate diet challenge. Animals. (2022) 12:2129. doi: 10.3390/ani12162129
18. Gowane, GR, and Vohra, V. Advances in Buffalo breeding: a journey from classical breeding to genomic selection In: MS Chauhan and N Selokar, editors. Biotechnological applications in buffalo research. Singapore: Springer (2022). 57–77.
19. Rehman, S, Hassan, F, Luo, X, Li, Z, and Liu, Q. Whole-genome sequencing and characterization of buffalo genetic resources: recent advances and future challenges. Animals. (2021) 11:904. doi: 10.3390/ani11030904
20. Shao, B, Sun, H, Ahmad, MJ, Ghanem, N, Abdel-Shafy, H, Du, C, et al. Genetic features of reproductive traits in bovine and Buffalo: lessons from bovine to Buffalo. Front Genet. (2021) 12:617128. doi: 10.3389/fgene.2021.617128
21. Chafai, N, Hayah, I, Houaga, I, and Badaoui, B. A review of machine learning models applied to genomic prediction in animal breeding. Front Genet. (2023) 14:1150596. doi: 10.3389/fgene.2023.1150596
22. Scott, BA, Haile-Mariam, M, Cocks, BG, and Pryce, JE. How genomic selection has increased rates of genetic gain and inbreeding in the Australian national herd, genomic information nucleus, and bulls. J Dairy Sci. (2021) 104:11832–49. doi: 10.3168/jds.2021-20326
23. Yousuf, M, Yusuf, A, and Mohammed, I. Review on current animal breeding and genetic technologies to increase production and productivity of cattle. Glob J Anim Sci Res. (2024) 12:19–36.
24. Bhatti, S, Sarwar, M, Khan, M, and Hussain, S. Reducing the age at first calving through nutritional manipulations in dairy buffaloes and cows: a review. Pak Vet J. (2007) 27:42–7.
25. Seno, L, Cardoso, V, El Faro, L, Sesana, R, Aspilcueta-Borquis, R, De Camargo, G, et al. Genetic parameters for milk yield, age at first calving and interval between first and second calving in milk Murrah buffaloes. Livest Res Rural Dev. (2010) 22:38. doi: 10.4081/ijas.2007.s2.397
26. Tamboli, P, Bharadwaj, A, Chaurasiya, A, Bangar, YC, and Jerome, A. Association between age at first calving, first lactation traits and lifetime productivity in Murrah buffaloes. Anim Biosci. (2022) 35:1151–61. doi: 10.5713/ab.21.0182
27. Mathur, BS, and Roychoudhury, PN. Effect of age at first calving and first lactation yield on life time production in Italian buffaloes. Zentralbl Veterinarmed A. (1971) 18:499–506. doi: 10.1111/j.1439-0442.1971.tb00605.x
28. Calanni Macchio, A, Santinello, M, Bifulco, G, Matera, R, Biffani, S, Gomez-Carpio, M, et al. The role of age at first calving in shaping production and reproductive outcomes in Italian buffaloes. J Dairy Sci. (2025) 108:7235–47. doi: 10.3168/jds.2025-26369
29. Ranjan, A, Jain, A, Verma, A, Sinha, R, Joshi, P, Gowane, GR, et al. Optimization of test day for milk yield recording and sire evaluation in Murrah buffaloes. J Anim Breed Genet. (2023) 140:400–12. doi: 10.1111/jbg.12767
30. Zhang, XX, An, ZG, Niu, KF, Chen, C, Ye, TZ, Shaukat, A, et al. Evaluation of type traits in relation to production, and their importance in early selection for milk performance in dairy buffaloes. Animal. (2022) 16:100653. doi: 10.1016/j.animal.2022.100653
31. Kumar, A, Vohra, V, Verma, U, and Singh, U. Estimates of genetic parameters for production and reproduction traits in Murrah buffaloes (riverine buffalo) in India. Buf Bull. (2022) 41:135–41. doi: 10.56825/bufbu.2022.4113396
32. Sathwara, RN, Gupta, JP, Chaudhari, JD, Prajapati, BM, Srivastava, AK, Chauhan, HD, et al. Sire evaluation models for estimating breeding values of Mehsana buffaloes. Indian J Anim Sci. (2019) 89:448–52. doi: 10.56093/ijans.v89i4.89148
33. Salem, MMI, Amin, AMS, Ashour, AF, Ibrahim, MME, and Abo-Ismail, MK. Genetic parameters and principal components analysis of breeding value for birth and weaning weight in Egyptian buffalo. Anim Biosci. (2021) 34:12–9. doi: 10.5713/ajas.19.0651
34. Amin, AMS, Abo-Ismail, MK, and Salem, MMI. Genetic parameters and genetic trends for reproductive traits in Egyptian buffalo. Anim Reprod Sci. (2021) 231:106800. doi: 10.1016/j.anireprosci.2021.106800
35. Dahiyta, S, Kumar, M, Ratwan, P, and Dhillod, S. Genetic analysis of biometric traits in Murrah buffaloes using Bayesian approach. Turkish Journal of Veterinary and Animal Sciences. (2022) 46:285–92. doi: 10.55730/1300-0128.4176
36. Diaz, JR. Aspectos genéticos da produção de leite e seus constituíntes em búfalas mestiças. Brazil: Universidade Estadual Paulista Julio de Mesquita Filho (2010).
37. Wolf, J, Zavadilová, L, and Němcová, E. Non-additive effects on milk production in Czech dairy cows. J Anim Breed Genet. (2005) 122:332–9. doi: 10.1111/j.1439-0388.2005.00537.x
38. Misztal, I, Tsuruta, S, Strabel, T, Auvray, B, Druet, T, and Lee, D. BLUPF90 and related programs (BGF90). In Proceedings of the 7th world congress on genetics applied to livestock production, Montpellier, France Genetics Applied to Livestock Production. (2002).
39. Geweke, JF. (1991). Evaluating the accuracy of sampling-based approaches to the calculation of posterior moments. Minneapolis: Federal Reserve Bank of Minneapolis. 169–193.
40. Sun, N, Wang, Y, Chu, J, Han, Q, and Shen, Y. Bayesian approaches in exploring gene-environment and gene-gene interactions: a comprehensive review. Cancer Genomics Proteomics. (2023) 20:669–78. doi: 10.21873/cgp.20414
41. Schoot, RVD, Depaoli, S, King, R, Kramer, B, Märtens, K, Tadesse, MG, et al. Bayesian statistics and modelling. Nat Rev Methods Primers. (2021) 1:2. doi: 10.1038/s43586-020-00001-2
42. Camargo Júnior, RNC, Fernandes, LDS, Panetto, JCDC, Silva, MVGBD, Araújo, CVD, Silva, AGME, et al. Heterogeneity of variance and genetic parameters for milk production in cattle, using Bayesian inference. PLoS One. (2023) 18:e0288257. doi: 10.1371/journal.pone.0288257
43. Colonia, SRR, Oliveira, ADC, and Pilonetto, F. Genetic parameters for milk yield, casein percentage, subclinical mastitis incidence and sexual precocity using Bayesian linear and threshold models. Anim Prod Sci. (2022) 62:792–801. doi: 10.1071/AN20313
44. Kumar, M, Vohra, V, Ratwan, P, and Lathwal, SS. Genetic analysis of milk and milk composition traits in Murrah buffaloes using Bayesian inference. Anim Biotechnol. (2023) 34:3280–6. doi: 10.1080/10495398.2022.2130797
45. Mendes-Malhado, CH, Mendes-Malhado, AC, Amorim-Ramos, A, Carneiro, PLS, Siewerdt, F, and Pala, A. Genetic parameters by Bayesian inference for dual purpose Jaffarabadi buffaloes. Arch Anim Breed. (2012) 55:567–76. doi: 10.5194/aab-55-567-2012
46. Carvalho, ISD, Lázaro, SF, Stefani, G, Silva, AA, Silveira, KRD, Scalez, DCB, et al. Genetic parameters and association between longevity and milk production in buffaloes using the ssGBLUP method. Rev Bras Zootec. (2023) 52:e20220143. doi: 10.37496/rbz5220220143
47. Kumar, M, Vohra, V, Ratwan, P, Gowane, GR, and Malhotra, R. Sustainable multi-trait selection index based on production, reproduction, and health traits for genetic improvement of Murrah buffaloes. Anim Biotechnol. (2023) 34:2505–13. doi: 10.1080/10495398.2022.2101117
48. Muner, RD, Moaeen-ud-Din, M, Saleem, AH, Ullah, Q, Abbas, G, Maqbool, B, et al. Genomic selection, gene editing, and reproductive biotechnology: a triad for the improvement of native buffalo breeds in a developing country perspective like Pakistan. Trop Anim Health Prod. (2025) 57:102. doi: 10.1007/s11250-025-04352-4
49. Amin, AMS, Salem, MMI, Ibrahim, MME, Darweish, SA, and El-Nagar, AG. Factors affecting calving ease in Egyptian buffalo. Reprod Domest Anim. (2021) 56:370–3. doi: 10.1111/rda.13834
50. Verma, R, Kennady, V, Meenakshi, V, Sanjeev, K, and Verma, A. Seasonal influence of age at first calving on genetic variation and subsequent reproductive performances in Murrah buffaloes. Biol Rhythm Res. (2021) 52:622–35. doi: 10.1080/09291016.2019.1627655
51. George, L, Rani, A, Gopal, G, Vikas, V, Pooja, J, Ravi, K, et al. Weighted single step GWAS reveals genomic regions associated with economic traits in Murrah buffaloes. Anim Biotechnol. (2024) 35:2319622. doi: 10.1080/10495398.2024.2319622
52. Yadav, N, Mukherjee, S, and Mukherjee, A. Comparative genetic analysis of frequentist and Bayesian approach for reproduction, production and life time traits showing favourable association of age at first calving in Tharparkar cattle. Anim Biosci. (2023) 36:1806–20. doi: 10.5713/ab.23.0025
53. Schmid, M, Imort-Just, A, Emmerling, R, Fuerst, C, Hamann, H, and Bennewitz, J. Genotype-by-environment interactions at the trait level and total merit index level for milk production and functional traits in Brown Swiss cattle. Animal. (2021) 15:100052. doi: 10.1016/j.animal.2020.100052
54. Talokar, AJ, Kumar, H, Mehrotra, A, Kaisa, K, Saravanan, KA, Panigrahi, M, et al. Recent advances in sire evaluation methods: a review. Indian J Anim Res. (2023) 57:395–401. doi: 10.18805/IJAR.B-4280
55. Cole, JB, Dürr, JW, and Nicolazzi, EL. Invited review: the future of selection decisions and breeding programs: what are we breeding for, and who decides? J Dairy Sci. (2021) 104:5111–24. doi: 10.3168/jds.2020-19777
56. Taylor, EN, Channa, K, Hanks, J, and Taylor, NM. Milk recording data indicates the importance of fertility, including age at first calving, on the progression of first lactation cows to second lactation. PLoS One. (2024) 19:e0297657. doi: 10.1371/journal.pone.0297657
57. Vargas-Leitón, B, Romero-Zúñiga, JJ, Castillo-Badilla, G, and Saborío-Montero, A. Optimal age at first calving in pasture-based dairy systems. Dairy. (2023) 4:581–93. doi: 10.3390/dairy4040040
58. Chen, SY, Freitas, PHF, and Oliveira, HR. Genotype-by-environment interactions for reproduction, body composition, and growth traits in maternal-line pigs based on single-step genomic reaction norms. Genet Sel Evol. (2021) 53:51. doi: 10.1186/s12711-021-00645-y
59. Gutierrez-Reinoso, MA, Aponte, PM, and Garcia-Herreros, M. Genomic analysis, Progress and future perspectives in dairy cattle selection: a review. Animals. (2021) 11:599. doi: 10.3390/ani11030599
60. Wang, X, Shi, S, Wang, G, Luo, W, Wei, X, Qiu, A, et al. Using machine learning to improve the accuracy of genomic prediction of reproduction traits in pigs. J Anim Sci Biotechnol. (2022) 13:60. doi: 10.1186/s40104-022-00708-0
61. Gómez-Carpio, M, Cesarani, A, Zullo, G, Cimmino, R, Neglia, G, Campanile, G, et al. Genetic parameters for reproductive traits in the Italian Mediterranean buffalo using milk yield as a correlated trait. J Dairy Sci. (2023) 106:9016–25. doi: 10.3168/jds.2023-23257
62. Albert, JH, and Chib, S. Bayes inference via Gibbs sampling of autoregressive time series subject to Markov mean and variance shifts. J Bus Econ Stat. (1993) 11:1–15. doi: 10.1080/07350015.1993.10509929
63. Fox, JP, and Glas, CAW. Bayesian estimation of a multilevel IRT model using Gibbs sampling. Psychometrika. (2001) 66:271–88. doi: 10.1007/BF02294839
64. Gelfand, AE. Gibbs sampling. J Am Stat Assoc. (2000) 95:1300–4. doi: 10.1080/01621459.2000.10474335
65. Gelfand, AE, Hills, SE, Amy, RP, and Smith, AFM. Illustration of Bayesian inference in normal data models using Gibbs sampling. J Am Stat Assoc. (1990) 85:972–85. doi: 10.1080/01621459.1990.10474968
Keywords: Bayesian inference, Geweke test, Markov chains via Monte Carlo, Murrah breed, Gibbs sampler
Citation: Camargo-Júnior RNC, de Araújo CV, Gomes MdNB, Marques JRF, da Silva WC, Sousa CEL, de Andrade RL, de Oliveira AS, da Silva ÉBR, Cara JRF, Lourenço-Júnior JB, Santos AM and Silva AGMe (2025) Comparison between linear mixed model and threshold model in the estimation of variance components in age at first calving and milk production in buffaloes. Front. Vet. Sci. 12:1649690. doi: 10.3389/fvets.2025.1649690
Edited by:
Daniel Mota-Rojas, Metropolitan Autonomous University, MexicoReviewed by:
Damiano Cavallini, University of Bologna, ItalyAdriana Domínguez-Oliva, Metropolitan Autonomous University, Mexico
Copyright © 2025 Camargo-Júnior, de Araújo, Gomes, Marques, da Silva, Sousa, de Andrade, de Oliveira, da Silva, Cara, Lourenço-Júnior, Santos and Silva. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Raimundo Nonato Colares Camargo-Júnior, Y2FtYXJnb2p1bmlvckBnbWFpbC5jb20=