 Manuel Louro1
Manuel Louro1 Andreia Valença
Andreia Valença Margarida Alves
Margarida Alves- 1Faculty of Veterinary Medicine, Lusófona University-Lisbon University Centre, Lisbon, Portugal
- 2Research in Veterinary Medicine (I-MVET), Faculty of Veterinary Medicine, Lusófona University-Lisbon University Centre, Lisbon, Portugal
- 3All Pets – Clínica Veterinária de Tires, São Domingos de Rana, Portugal
- 4Superior School of Health, Protection and Animal Welfare, Polytechnic Institute of Lusophony, Lisbon, Portugal
- 5Animal and Veterinary Research Center (CECAV), Faculty of Veterinary Medicine, Lusófona University - Lisbon University Centre, Lisbon, Portugal
- 6Global Health and Tropical Medicine (GHTM), Associate Laboratory in Translation and Innovation Towards Global Health (LA-REAL), Instituto de Higiene e Medicina Tropical (IHMT), Universidade NOVA De Lisboa (UNL), Lisbon, Portugal
Introduction: Mycoplasma agassizii is a well-recognized etiologic agent of upper respiratory tract disease in tortoises. Although frequently reported in both captive and wild populations across Europe, its occurrence in Portugal had not been previously documented. This study aimed to investigate the presence of M. agassizii in apparently healthy captive tortoises in mainland Portugal and to evaluate potential host- and management-related factors associated with infection.
Methods: Oral swabs were collected from 84 tortoises of 13 species across 3 geographic regions. DNA extraction success and sample integrity were confirmed by partial amplification of the tortoise mitochondrial 12S rRNA gene in 92.9% of cases (78/84), which were then screened for M. agassizii using a species-specific PCR targeting the 16S rRNA gene.
Results and discussion: The pathogen DNA was detected in 66.7% (52/78) of individuals. Phylogenetic analysis confirmed species identification, with all sequences forming a strongly supported monophyletic cluster together with M. agassizii reference sequences. A significant association was observed between tortoise genus and M. agassizii infection (p = 0.021), with Chelonoidis exhibiting a significantly lower infection frequency than Testudo (p = 0.029). No statistically significant associations were observed regarding geographic region, housing origin, or group size. These results reveal a high frequency of M. agassizii infection in apparently healthy captive tortoises in Portugal, emphasizing its potential for silent transmission in group or mixed-species settings. Our findings support the inclusion of this pathogen in the differential diagnosis of respiratory disease in tortoises, even when clinical signs are absent and underscore the need for routine molecular surveillance and strengthened biosecurity practices to mitigate transmission risks and foster chelonian conservation efforts.
1 Introduction
Some Mycoplasma species are considered commensals in chelonians, while others can cause severe upper respiratory tract disease (URTD), leading to significant morbidity and mortality in tortoises (1–4). Mycoplasma agassizii is the most commonly reported etiologic agent of URTD in both free-ranging and captive tortoises in the United States and Europe (3–9).
Mycoplasma agassizii infections may persist in chronic or subclinical forms, with intermittent recurrence of clinical signs such as rhinitis (ranging from mild to severe), nasal and ocular discharges, periocular edema, and conjunctivitis (7–10). Lesions are primarily localized to the nasal cavity, impairing their sense of smell and consequently their ability to forage (8, 11).
Although high pathogen loads are often required to induce disease, subclinically infected tortoises can serve as long-term carriers (8, 12–15). This persistence is likely associated with innate immune responses that reduce but do not eliminate infection, allowing recrudescence under stress conditions (13, 16–18). Persistent infection, combined with pathogen transmission among individuals, contributes to the maintenance and spread of M. agassizii within populations (12, 13).
Transmission occurs mainly through direct contact, especially during courtship, mating, or aggressive interactions. Although individuals exhibiting clinical signs are more likely to transmit the pathogen, asymptomatic carriers also play a role in its spread (8, 19, 20). Infection and clinical signs are more frequently reported in captive tortoises, such as those in zoos, rescue centers, breeding facilities, and private collections, likely reflecting increased surveillance and more frequent health assessments. Nevertheless, the close and prolonged contact characteristic of captive settings may also play a significant role in transmission. This has implications for both animal welfare and disease management, with considerable treatment costs (12, 21–25).
Importantly, M. agassizii poses a threat to the conservation of wild tortoise populations, particularly those already impacted by anthropogenic pressures (7, 8, 23, 26). Tortoises living near human settlements show higher prevalence compared to those in remote areas with minimal human contact (22, 27), suggesting that the escape or intentional release of captive tortoises may introduce M. agassizii and other pathogens, such as herpesvirus, into naïve wild populations (8, 21, 22, 28, 29). Habitat degradation due to agriculture, forestry, mining, urban development, and pollution may further disrupt natural behavior and cause physiological stress, increasing the risk of URTD outbreaks (8, 22, 27, 30). Other stressors include handling, translocations, interactions or injuries involving domestic animals or vehicles, and exposure to waste materials (16, 30–32).
Pathogen surveillance in wildlife is particularly challenging in long-lived hosts and for persistent infections (13). M. agassizii is not host-specific and has been detected in several species of free-ranging and captive tortoises (4, 7, 8, 25, 26, 33), raising concerns about interspecies transmission (4, 33). Effective diagnosis and monitoring are therefore crucial, especially in captive populations, to prevent further spread. This is particularly important in the pet trade and conservation programs involving reintroductions, where undetected infections could compromise wild populations. Surveillance of captive tortoises can yield valuable data on the epidemiology of M. agassizii and help clarify disease mechanisms, ultimately supporting the prevention and control of URTD in both captive and wild animals (7, 16, 34, 35).
While M. agassizii has been reported in both wild and captive tortoises across Europe, no data are currently available on its occurrence in Portugal. This study aimed to investigate the presence of M. agassizii in apparently healthy captive tortoises in mainland Portugal and to evaluate potential host- and management-related factors associated with infection.
2 Materials and methods
2.1 Sample collection
A cross-sectional survey was conducted between March 2022 and June 2023 in private households (not for commercial or breeding purposes), breeding facilities, and animal parks located in the North, Lisbon and Tagus Valley, and South regions of mainland Portugal (Figure 1). Sterile flocked oral swabs were collected from 84 apparently healthy captive adult tortoises (i.e., no clinical signs compatible with respiratory or other infections were observed during clinical evaluation) of 13 different species, including Aldabrachelys gigantea (n = 3), Astrochelys radiata (n = 4), Centrochelys sulcata (n = 14), Chelonoidis carbonarius (n = 8), Geochelone elegans (n = 1), Indotestudo elongata (n = 2), Kinixys belliana (n = 1), Malacochersus tornieri (n = 3), Stigmochelys pardalis (n = 7), Testudo graeca (n = 26), Testudo hermanni (n = 4), Testudo horsfieldii (n = 3), and Testudo marginata (n = 2). These animals were distributed across 23 enclosures belonging to 10 different households or facilities. Animals were selected based on availability and accessibility, following a convenience sampling strategy. Immediately after collection, each swab was placed dry (i.e., without transport medium) in a sterile 2 ml microtube. Samples were kept refrigerated (4°C) for a maximum of 24 h after collection and then stored at −80 °C until DNA extraction.
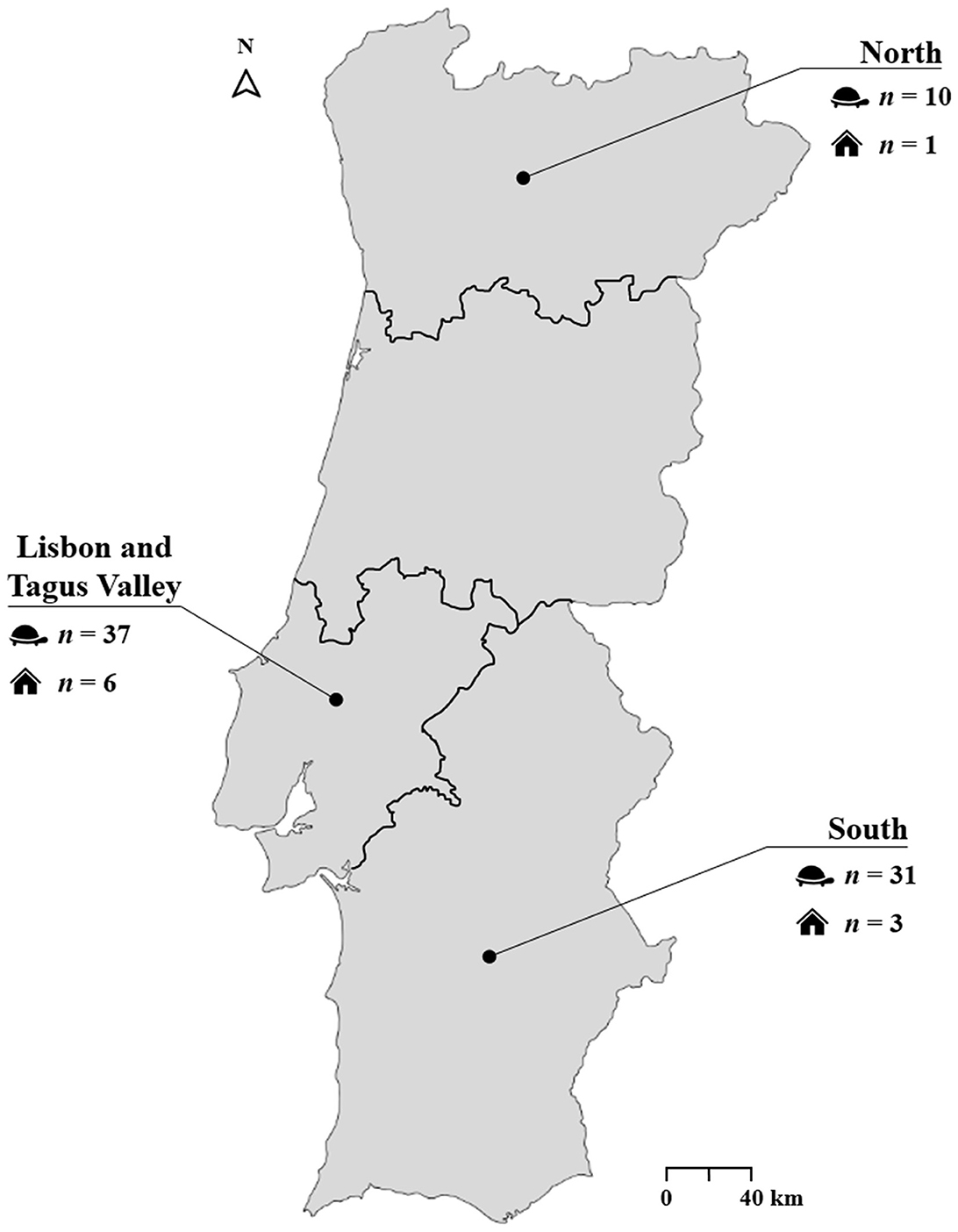
Figure 1. Geographic regions of sample collection.
Additionally, each animal caretaker completed a questionnaire designed to gather information on the general health status of the animals, and relevant housing and husbandry practices.
The study was approved by the Ethics and Animal Welfare Committee (CEBEA) of the Faculty of Veterinary Medicine, Lusófona University—Lisbon University Centre (approval number 5/2022). Written informed consent was obtained from all owners or legal detainers prior to sampling.
2.2 DNA extraction and chelonian housekeeping gene PCR
DNA was extracted from oral swabs using the Invisorb® Spin Universal Kit (Invitek Molecular, Berlin, Germany), according to the manufacturer's instructions.
Tortoise nucleic acid was detected using primers targeting a conserved region of the mitochondrial 12S rRNA gene (Table 1), as previously described (36). PCR reactions were performed in a total volume of 25 μl, containing 12.5 μl of NZYTaq II 2x Green Master Mix (NZYTech®, Lisbon, Portugal), 15 pmol of each primer, 5 μl of total DNA, and sterile ultrapure water to complete the final volume. Non-template controls (NTC) containing water instead of DNA were included in each run. Amplification was carried out on a T100™ Thermal Cycler (Bio-Rad, Hercules, CA, USA) with the following conditions: initial denaturation at 94°C for 5 min; 43 cycles of 94°C for 30 s, 52°C for 45 s, and 72°C for 1 min; followed by a final extension at 72°C for 6 min. PCR products were visualized by electrophoresis on 1.5% (w/v) agarose gels under UV illumination.

Table 1. Primers used to amplify tortoise and Mycoplasma agassizii DNA.
2.3 Molecular detection of M. agassizii
Only DNA samples in which amplification of the 12S rRNA gene fragment was successfully achieved were screened for M. agassizii by PCR targeting the 16S rRNA gene, using species-specific primers previously described by Brown et al. (5) (Table 1).
Reactions were performed in a final volume of 25 μl, containing 12.5 μl of NZYTaq II 2x Green Master Mix (NZYTech®), 10 pmol of each primer, 5 μl of DNA, and sterile ultrapure water to complete the final volume. NTC and positive controls (DNA from a confirmed M. agassizii sample) were included in each run. Amplification was carried out on a T100™ Thermal Cycler (Bio-Rad) using the following conditions: initial denaturation at 94°C for 5 min; 50 cycles at 94°C for 45 s, 55°C for 1 min, and 72°C for 45 s; followed by a final extension at 72°C for 10 min. Amplicons were visualized by electrophoresis on 1.5% (w/v) agarose gels under UV illumination.
All PCR products with amplicons of the expected size were purified using the Jetquick PCR Product Purification Spin Kit (Genomed GmbH, Bad Oeynhausen, Germany), following the manufacturer's instructions. Purified products were submitted for Sanger sequencing at STABVida® (Caparica, Portugal). Sequence data were analyzed using the FinchTV software (Geospiza®), and nucleotide sequence similarity searches were performed with the BLASTn tool (http://blast.ncbi.nlm.nih.gov/Blast.cgi) (37). The nucleotide sequences obtained during this study were deposited in the DDBJ/ENA/GenBank under the accession numbers LC878613-LC878662.
2.4 Phylogenetic inference
Multiple sequence alignments were generated using the G-INS-i iterative refinement method implemented in MAFFT v7 (38). The resulting alignments were treated with Gblocks (39), via SeaView v5.0.5, using default parameters. Phylogenetic reconstruction was performed using the maximum likelihood method implemented in IQ-TREE v1.6.12, with the best-fit substitution model selected according to the Bayesian Information Criterion. Branch support was assessed using the bootstrap method with 1,000 replicates, with values ≥75% considered indicative of strong topological support. Trees were visualized and edited for display using iTOL v6 (40). A list of all Mycoplasma spp. sequences included in the phylogenetic analysis, along with associated metadata (i.e., strain name or molecular ID, host, country of origin, GenBank accession number, and type strain status), is provided in Supplementary Table 1.
2.5 Statistical analysis
Data were compiled in Microsoft Excel v365 and analyzed using IBM SPSS Statistics v25. Exploratory and descriptive analyses were conducted to characterize the dataset. Relative frequencies of M. agassizii-positive cases were calculated, and 95% confidence intervals (CI) were obtained using Wilson's method via the OpenEpi online tool (41).
Associations between categorical variables (i.e., tortoise species, geographic region, origin, and housing type) and M. agassizii infection status were explored using the Chi-square (χ2) test, Fisher's exact test (for 2 × 2 tables), or the Fisher–Freeman–Halton exact test (for larger contingency tables), as appropriate. Adjusted standardized residuals (ASR) were determined to identify cells with significant contributions to the overall test result, with ASR values exceeding ±1.96 considered statistically significant (α = 0.05). Pairwise comparisons were conducted between categories of variables that showed significant overall differences.
The Mann–Whitney U test was used to evaluate differences in the number of animals per enclosure between groups defined by M. agassizii infection status. A p-value < 0.05 was considered statistically significant for all analyses.
3 Results
Partial amplification of the mitochondrial 12S rRNA gene, employed as a host-specific internal control, was successful in 92.9% (78/84) of the sampled tortoises. These 78 samples were considered suitable for further molecular screening of M. agassizii.
Mycoplasma agassizii 16S rRNA gene fragment was detected in 66.7% (52/78) of the screened individuals (Table 2). BLASTn analysis of the obtained sequences showed 100% identity and 99% query cover with M. agassizii reference sequences available in public databases (e.g., GenBank accession no. KY212532).
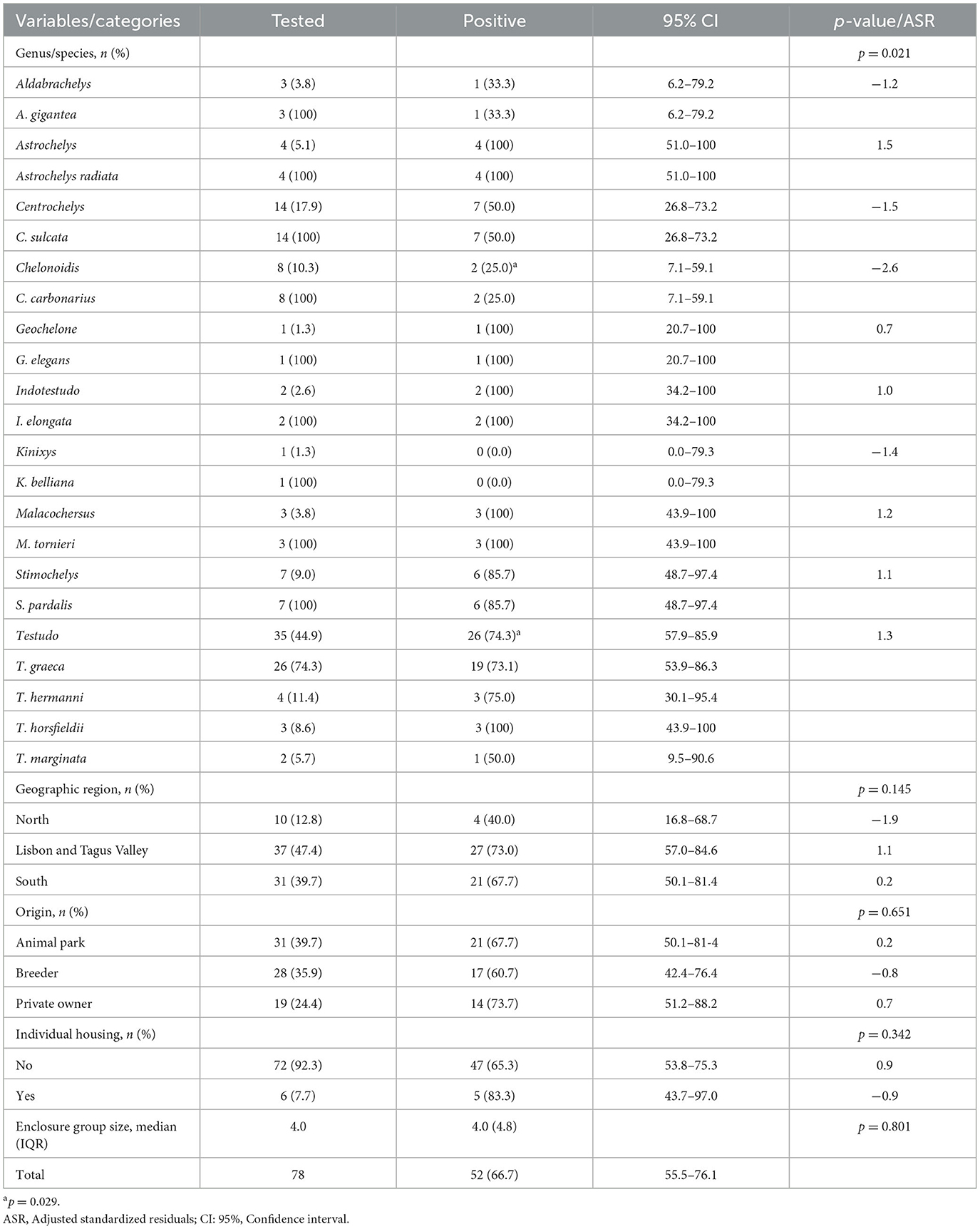
Table 2. Molecular frequency of Mycoplasma agassizii infection in captive chelonians by species, origin, region, and housing conditions.
Phylogenetic analysis revealed that all 16S rRNA sequences obtained in this study segregated into a well-supported monophyletic cluster, exclusively composed of M. agassizii reference sequences (Figure 2).
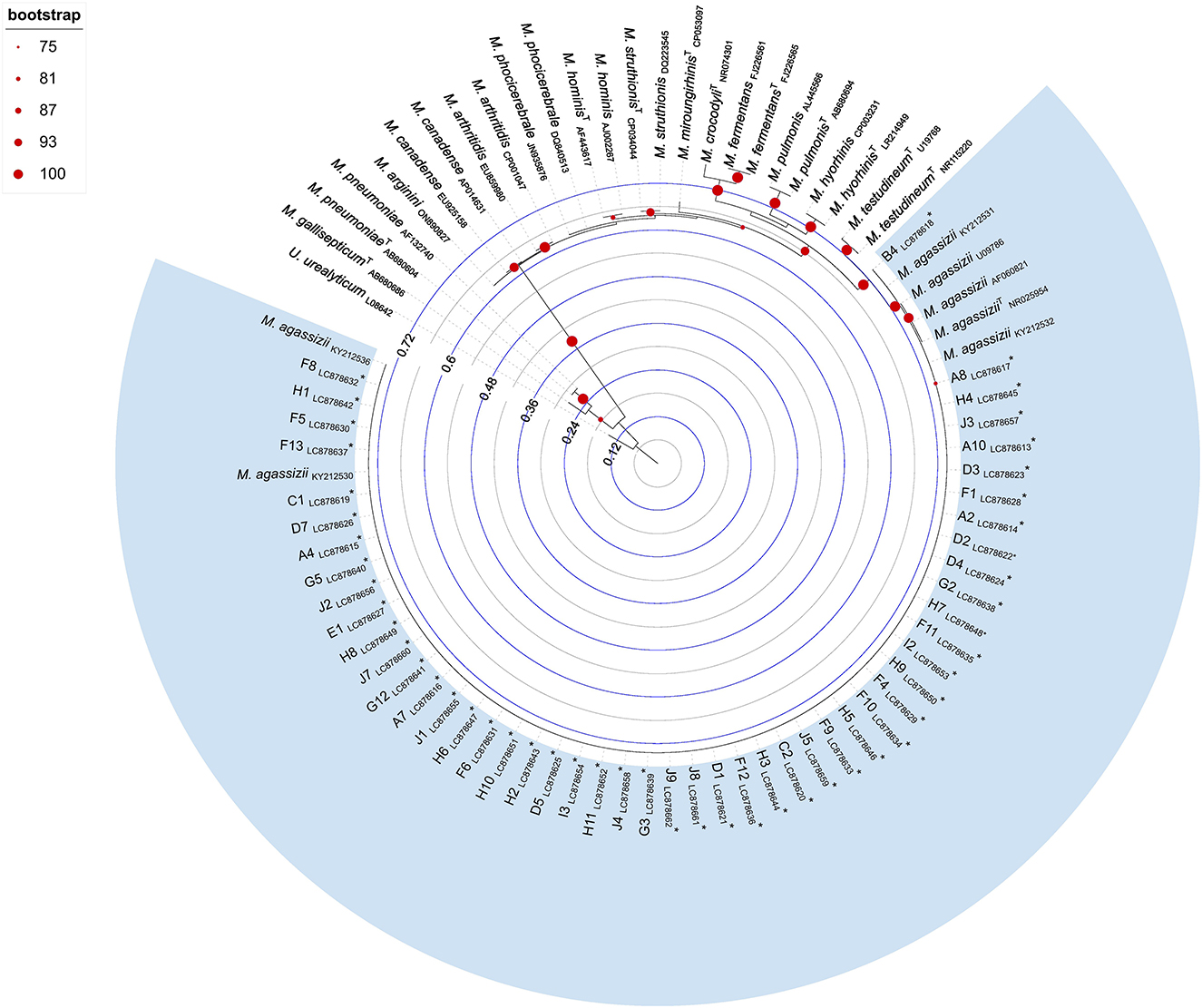
Figure 2. Maximum likelihood phylogenetic tree inferred from Mycoplasma spp. 16S rRNA gene sequences (509 bp). Tree reconstruction was performed in IQ-TREE using the TVMe+I+G4 substitution model, selected as the best-fitting model based on the Bayesian Information Criterion. Node support was assessed using 1,000 bootstrap replicates, and values ≥75% are shown at the corresponding nodes. The tree was rooted using a Ureaplasma urealyticum sequence (GenBank accession number: L08642). Sequences retrieved from public databases are labeled with species name and GenBank accession number; type strains are indicated with a superscript “T” following the species name. Sequences obtained in this study are marked with an asterisk and include the tortoise identifier and GenBank accession numbers LC878613–LC878662. Branch lengths are proportional to the number of nucleotide substitutions per site, as indicated by the internal scale bar.
Infection frequencies across species ranged from 0.0% in K. belliana (n = 1) to 100% in several species, including A. radiata (n = 4), I. elongata (n = 2), M. tornieri (n = 3), G. elegans (n = 1), and T. horsfieldii (n = 3). Although these species had small sample sizes, T. graeca had the highest number of individuals testing positive for M. agassizii DNA (19/26).
Among genera with at least three sampled individuals, Testudo showed the highest detection frequency (74.3%; 26/35). A statistically significant association was observed between tortoise genus and the presence of M. agassizii DNA (p = 0.021; Table 2). Pairwise comparison showed that Chelonoidis had a significantly (p = 0.029) lower frequency (25.0%, 2/8; ASR = −2.6,) of M. agassizii infection compared to other genera, particularly in contrast to Testudo (74.3%, 26/35).
Positive cases were detected across all sampled regions in mainland Portugal. The Lisbon and Tagus Valley region recorded the highest number of infected tortoises (73.0%, 27/37), followed by the South (67.7%, 21/31) and the North (40.0%, 4/10). However, these regional differences were not statistically significant (p = 0.145).
Mycoplasma agassizii DNA was detected in tortoises from all housing origins: 73.7% (14/19) of private households, 67.7% (21/31) of animal parks, and 60.7% (17/28) of breeders. No statistically significant association was found between housing origin and infection status (p = 0.651).
Regarding housing conditions, no association was found between M. agassizii DNA detection and enclosure group size or individual housing. The median number of animals per enclosure did not differ significantly between infected and uninfected tortoises (p = 0.801), nor was any association found with being housed individually (p = 0.342).
4 Discussion
While M. agassizii has been previously reported in both wild and captive tortoises across Europe, this is the first comprehensive study to assess its presence in captive tortoises kept under diverse housing origins and conditions in mainland Portugal. The high detection frequency (66.7%; 52/78) aligns with M. agassizii transmission dynamics, which rely on prolonged direct contact (typically >24–48 h) for successful spread (12). Such conditions are frequently met in captive settings due to co-housing and group management practices (12, 21–25). However, this percentage is higher than those reported in both free-ranging and captive tortoises in other European countries, where prevalence ranged from 0 to 42% (4, 7, 25, 42, 43). Notably, unlike previous studies, none of the animals tested in this study exhibited clinical signs of URTD, suggesting subclinical or chronic infections. By acting as reservoirs, these individuals can contribute to pathogen persistence and spread, complicating sanitary control, especially given the lower pathogen loads often observed in subclinical cases (8, 12–15). This highlights the importance of molecular screening, even in the absence of clinical signs, particularly during quarantine, relocation, or introduction into new enclosures.
Although quarantine periods of 12–18 months are recommended (8, 44), they are often impractical or omitted altogether, especially in illicit trade contexts. Furthermore, subclinical infections may remain undetected throughout quarantine. Therefore, rapid molecular screening at intake, using oral swabs (a non-invasive method compatible with the epithelial tropism of mycoplasmas) offers a practical approach for M. agassizii routine testing.
Most animals in this study were not housed individually, increasing the risk of horizontal transmission. This raises the possibility that some individuals may have become infected only after being introduced in mixed enclosures, thereby underlining the risk of within-collection transmission. Longitudinal follow-up of the six enclosures where both positive and negative individuals were detected could provide further insight into transmission dynamics and infection timelines.
Interestingly, in this study, tortoises of the genus Testudo exhibited a significantly higher infection frequency than those of the genus Chelonoidis. Although tortoises are not autochthonous to Portugal, Testudo species are native to neighboring Mediterranean countries, such as Spain, France, Italy, and Greece (45). The decline of wild Testudo populations is well documented, with IUCN conservation statuses ranging from Near Threatened to Critically Endangered (3, 43). These populations face numerous threats including habitat loss and fragmentation, urban development, predation, delayed maturity, low fecundity, illegal pet trade and movement of exotic animals, competition with exotic tortoise species, and infectious diseases such as URTD (7, 8, 23, 46, 47).
The international pet trade exacerbates conservation challenges by enabling long-distance movement of infected individuals with limited or no sanitary oversight (35, 48). These flows of individuals result in open routes for the expansion of emerging infectious pathogens, such as M. agassizii. Given its high prevalence in captive animals, the introduction of infected individuals into the wild, whether intentional or accidental, could have devastating consequences for naïve wild populations. Studies have shown higher M. agassizii prevalence in tortoises associated with proximity to human settlements, likely reflecting contacts with captive or released individuals (22, 27). Ballouard et al. (42) reported high M. agassizii infection rates in captive and vagrant exotic tortoises co-occurring with native T. h. hermanni in urban or peri-urban areas in southeastern France. These findings raise concerns about the uncontrolled introduction of exotic pet tortoises and the potential transmission from captive to free-ranging tortoises. Conversely, the illegally harvesting of wild tortoises can also introduce M. agassizii into captive settings, thus perpetuating the transmission cycle.
Cross-species transmission of M. agassizii has been documented (4, 33), and many positive animals in this study were housed in mixed-species enclosures, supporting the likelihood of interspecies transmission. The fact that M. agassizii was detected in 12 different tortoise species, including C. sulcata, C. carbonarius, I. elongata, and M. tornieri, confirms that this pathogen is not species-specific. This contrasts with recent findings from Galosi et al. (7), who did not detect the pathogen in these species in Italy. Further research on both mixed-species and species-specific enclosures may provide new insights into cross-species transmission dynamics.
Several studies have reported URTD-like syndromes in mycoplasma-positive turtles of the family Emydidae (2, 25, 49–52). Although M. agassizii primarily affects tortoises, its full host range and transmission dynamics remain incompletely understood. There is some indication of Mycoplasma spp. being detected in freshwater turtles, including unpublished data suggesting PCR detection of M. agassizii in Trachemys scripta elegans (8). Further studies are needed to assess the susceptibility of other chelonian species and to evaluate potential risks associated with interspecific contact in shared environments. Given the ecological overlap between tortoises and freshwater turtle populations, investigating the potential for cross-species transmission remains an important area for future research.
Despite the absence of clinical signs in the tortoises sampled for this study, M. agassizii is recognized as the primary etiological agent of URTD in tortoises (3–9). This infection, even when subclinical, has the potential to progress to overt disease under certain stressors or immunosuppressive conditions (7–10, 13, 16–18). The high frequency observed here reinforces the need to include M. agassizii in differential diagnoses of respiratory conditions in tortoises, particularly in captive settings, new acquisitions, or individuals with recent chelonian contact. This study provides valuable epidemiological evidence to inform clinical decision-making and improve biosecurity protocols in both private and institutional collections.
5 Conclusion
This study provides the first evidence of widespread M. agassizii infection among captive tortoises in mainland Portugal. The pathogen was detected in multiple tortoise species, including species native to the Mediterranean region, yet none exhibited clinical signs at the time of sampling. This highlights the potential role of subclinical carriers in maintaining and spreading the infection, especially in settings with group housing or mixed-species enclosures. The international reptile trade, coupled with insufficient sanitary oversight, remains a driver of pathogen dissemination, posing significant risks to both native and non-native chelonian populations.
Given M. agassizii's established role in URTD and its potential to cause clinical illness under stress or immunosuppression, the results presented here support its inclusion in the differential diagnosis of respiratory disease in tortoises, even in the absence of clinical signs. Furthermore, our results underscore the importance of incorporating molecular diagnostics as a standard practice in routine veterinary care and tortoise collection management. This study offers valuable insights for guiding effective strategies to prevent pathogen transmission and protect vulnerable tortoise populations.
Data availability statement
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found below: https://www.ncbi.nlm.nih.gov/genbank/, LC878613-LC878662.
Ethics statement
The animal studies were approved by Ethics and Animal Welfare Committee (CEBEA) of the Faculty of Veterinary Medicine, Lusófona University - Lisbon University Centre. The studies were conducted in accordance with the local legislation and institutional requirements. Written informed consent was obtained from the owners for the participation of their animals in this study.
Author contributions
ML: Data curation, Investigation, Resources, Visualization, Writing – review & editing. RP: Investigation, Resources, Supervision, Writing – review & editing, Visualization. AP: Resources, Writing – review & editing, Data curation, Formal analysis, Writing – original draft. AV: Conceptualization, Data curation, Formal analysis, Investigation, Methodology, Resources, Supervision, Writing – original draft, Writing – review & editing. MA: Data curation, Resources, Writing – original draft, Writing – review & editing, Conceptualization, Funding acquisition, Investigation, Methodology, Project administration, Supervision.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. Funding was received from FMV-Lusófona University – Scholarship grant FMV (2021–2022).
Acknowledgments
The authors would like to thank FMV-Lusófona University for the financial support provided for this work. We are also grateful to the tortoise pet owners, breeders, and animal facilities that generously agreed to participate and contribute to this research.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Gen AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher's note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fvets.2025.1652362/full#supplementary-material
References
1. Di Ianni F, Dodi PL, Cabassi CS, Pelizzone I, Sala A, Cavirani S, et al. Conjunctival flora of clinically normal and diseased turtles and tortoises. BMC Vet Res. (2015) 11:91. doi: 10.1186/s12917-015-0405-x
2. Farkas SL, Gál J. Adenovirus and mycoplasma infection in an ornate box turtle (Terrapene ornata ornata) in Hungary. Vet Microbiol. (2009) 138:169–73. doi: 10.1016/j.vetmic.2009.03.016
3. Lecis R, Paglietti B, Rubino S, Are BM, Muzzeddu M, Berlinguer F, et al. Detection and characterization of Mycoplasma spp. and Salmonella spp. in free-living European tortoises (Testudo hermanni, Testudo graeca, and Testudo marginata). J Wildl Dis. (2011) 47:717–24. doi: 10.7589/0090-3558-47.3.717
4. Soares JF, Chalker VJ, Erles K, Holtby S, Waters M, McArthur S. Prevalence of Mycoplasma agassizii and chelonian herpesvirus in captive tortoises (Testudo sp) in the United Kingdom. J Zoo Wildl Med. (2004) 35:25–33. doi: 10.1638/02-092
5. Brown MB, McLaughlin GS, Klein PA, Crenshaw BC, Schumacher IM, Brown DR, et al. Upper respiratory tract disease in the gopher tortoise is caused by Mycoplasma agassizii. J Clin Microbiol. (1999) 37:2262–69. doi: 10.1128/JCM.37.7.2262-2269.1999
6. Brown MB, Schumacher IM, Klein PA, Harris D, Corell T, Jacobson ER. Mycoplasma agassizii causes upper respiratory tract disease in the desert tortoise. Infect Immun. (1994) 62:4580–86. doi: 10.1128/iai.62.10.4580-4586.1994
7. Galosi L, Ridolfi N, Fellini C, Pelizzone I, Cusaro S, Marchetti G, et al. Detection and identification of Mycoplasmopsis agassizii in captive tortoises with different clinical signs in Italy. Animals. (2023) 13:588. doi: 10.3390/ani13040588
8. Jacobson ER, Brown MB, Wendland LD, Brown DR, Klein PA, Christopher MM, et al. Mycoplasmosis and upper respiratory tract disease of tortoises: a review and update. Vet J. (2014) 201:257–64. doi: 10.1016/j.tvjl.2014.05.039
9. Luzuriaga-Neira A, Sandmeier FC, Weitzman CL, Tracy CR, Bauschlicher SN, Tillett RL, et al. Mycoplasma agassizii, an opportunistic pathogen of tortoises, shows very little genetic variation across the Mojave and Sonoran Deserts. PLoS ONE. (2021) 16:e0245895. doi: 10.1371/journal.pone.0245895
10. Origgi FC, Jacobson ER. Diseases of the respiratory tract of chelonians. Vet Clin North Am Exot Anim Pract. (2000) 3:537–49. doi: 10.1016/S1094-9194(17)30088-9
11. Germano J, van Zerr VE, Esque TC, Nussear KE, Lamberski N. Impacts of upper respiratory tract disease on olfactory behavior of the Mojave desert tortoise. J Wildl Dis. (2014) 50:354–58. doi: 10.7589/2013-06-130
12. Aiello CM, Nussear KE, Esque TC, Emblidge PG, Sah P, Bansal S, et al. Host contact and shedding patterns clarify variation in pathogen exposure and transmission in threatened tortoise Gopherus agassizii: implications for disease modelling and management. J Anim Ecol. (2016) 85:829–42. doi: 10.1111/1365-2656.12511
13. Sandmeier FC, Maloney KN, Tracy CR, Hyde D, Mohammadpour H, Marlow R, et al. Chronic disease in the Mojave desert tortoise: Host physiology and recrudescence obscure patterns of pathogen transmission. Ecol Evol. (2017) 7:10616–29. doi: 10.1002/ece3.3480
14. Sandmeier FC, Weitzman CL, Tracy CR. An ecoimmunological approach to disease in tortoises reveals the importance of lymphocytes. Ecosphere. (2018) 9:e02427. doi: 10.1002/ecs2.2427
15. Weitzman CL, Sandmeier FC, Tracy CR. Prevalence and diversity of the upper respiratory pathogen Mycoplasma agassizii in Mojave desert tortoises (Gopherus agassizii). Herpetologica. (2017) 73:113–20. doi: 10.1655/Herpetologica-D-16-00079.1
16. Moeller CA, Perales S, Rodriguez W, Martin AM, Eversole CB, Rideout-Hanzak S, et al. Surveillance of Mycoplasma agassizii in Texas tortoises (Gopherus berlandieri) for translocation with emphasis on treatment and recovery. Front Vet Sci. (2025) 11:1525179. doi: 10.3389/fvets.2024.1525179
17. Simecka JW, Davis JK, Davidson MK, Ross SE, Stadtländer CTKH, Cassel GH. “Mycoplasma diseases of animals”. In:Maniloff J, McElhaney RN, Finch LR, Baseman JB, , editors. Mycoplasmas: Molecular Biology and Pathogenesis. Washington, DC: American Society for Microbiology Press (1992). p. 391–445.
18. Zimmerman LM, Vogel LA, Bowden RM. Understanding the vertebrate immune system: insights from the reptilian perspective. J Exp Biol. (2020) 213:661–71. doi: 10.1242/jeb.038315
19. Jacobson ER, Brown MB, Schumacher IM, Collins BR, Harris RK, Klein PA. Mycoplasmosis and the desert tortoise (Gopherus agassizii) in Las Vegas Valley, Nevada. Chelonian Conserv Biol. (1995) 1:279–84.
20. Jacobson ER, Berry KH. Mycoplasma testudineum in free-ranging desert tortoises, Gopherus agassizii. J Wildl Dis. (2012) 48:1063–68. doi: 10.7589/2011-09-256
21. Berry KH, Brown MB, Vaughn M, Gowan TA, Hasskamp MA, Torres MCM. Mycoplasma agassizii in Morafka's desert tortoise (Gopherus morafkai) in Mexico. J Wildl Dis. (2015) 51:89–100. doi: 10.7589/2014-04-083
22. Berry KH, Coble AA, Yee JL, Mack JS, Perry WM, Anderson KM, et al. Distance to human populations influences epidemiology of respiratory disease in desert tortoises. J Wildl Manage. (2015) 79:122–36. doi: 10.1002/jwmg.816
23. Dickinson VM, Schumacher IM, Jarchow JL, Duck T, Schwalbe CR. Mycoplasmosis in free-ranging desert tortoises in Utah and Arizona. J Wildl Dis. (2005) 41:839–42. doi: 10.7589/0090-3558-41.4.839
24. Jones CA. (2008) Mycoplasma agassizii in the Sonoran population of the desert tortoise in Arizona (master's thesis). University of Arizona, Tucson, AZ.
25. Kolesnik E, Obiegala A. Marschang RE. Detection of Mycoplasma spp, herpesviruses, topiviruses, and ferlaviruses in samples from chelonians in Europe. J Vet Diagn Invest. (2017) 29:820–32. doi: 10.1177/1040638717722387
26. Apakupakul K, Nieto-Claudin A, Rakotonanahary TF, Catenacci LS, Rasambainarivo F, Randrianarisoa S, et al. Molecular identification of Mycoplasma agassizii in confiscated. J Wildl Dis. (2025) 61:258–61. doi: 10.7589/JWD-D-24-00040
27. Berry KH, Bailey TY, Anderson KM. Attributes of desert tortoise populations at the National Training Center, Central Mojave Desert, California, USA. J Arid Environ. (2006) 67:165–91. doi: 10.1016/j.jaridenv.2006.09.026
28. Jacobson ER, Berry KH, Wellehan JFX Jr, Origgi F, Childress AL, Braun J, et al. Serologic and molecular evidence for Testudinid herpesvirus 2 infection in wild Agassiz's desert tortoises, Gopherus agassizii. J Wildl Dis. (2012) 48:747–57. doi: 10.7589/0090-3558-48.3.747
29. Johnson AJ, Morafka DJ, Jacobson ER. Seroprevalence of Mycoplasma agassizii and tortoise herpesvirus in captive desert tortoises (Gopherus agassizii) from the Greater Barstow Area, Mojave Desert, California. J Arid Environ. (2006) 67:192–201. doi: 10.1016/j.jaridenv.2006.09.025
30. Berry KH, Spangenberg EK, Homer BL, Jacobson ER. Deaths of desert tortoises following periods of drought and research manipulation. Chelonian Conserv Biol. (2002) 4:436–48.
31. Donoghue S. “Nutrition”. In:Mader DR, , editor. Reptile medicine and surgery. 2nd ed. St. Louis, MO: Saunders Elsevier (2006). p. 251–98. doi: 10.1016/B0-72-169327-X/50022-5
32. Esque TC, Nussear KE, Drake KK, Walde AD, Berry KH, Averill-Murray RC, et al. Effects of subsidized predators, resource variability, and human population density on desert tortoise populations in the Mojave Desert, USA. Endangered Species Res. (2010) 12:167–77. doi: 10.3354/esr00298
33. Salinas M, Francino O, Sánchez A, Altet L. Mycoplasma and herpesvirus PCR detection in tortoises with rhinitis-stomatitis complex in Spain. J Wildl Dis. (2011) 47:195–200. doi: 10.7589/0090-3558-47.1.195
34. Aiello CM, Esque TC, Nussear KE, Emblidge PG, Hudson PJ. The slow dynamics of mycoplasma infections in a tortoise host reveal heterogeneity pertinent to pathogen transmission and monitoring. Epidemiol Infect. (2018) 147:e12. doi: 10.1017/S0950268818002613
35. Auliya M, Altherr S, Ariano-Sanchez D, Baard EH, Brown C, Brown RM, et al. Trade in live reptiles, its impact on wild populations, and the role of the European market. Biol Conserv. (2016) 204:103–19. doi: 10.1016/j.biocon.2016.05.017
36. Kocher TD, Thomas WK, Meyer A, Edwards SV, Pääbo S, Villablanca FX, et al. Dynamics of mitochondrial DNA evolution in animals: amplification and sequencing with conserved primers. Proc Natl Acad Sci USA. (1989) 86:6196–200. doi: 10.1073/pnas.86.16.6196
37. Boratyn GM, Camacho C, Cooper PS, Coulouris G, Fong A, Ma N, et al. BLAST: a more efficient report with usability improvements. Nucleic Acids Res. (2013) 41:W29–33. doi: 10.1093/nar/gkt282
38. Katoh K, Rozewicki J, Yamada KD. MAFFT online service: multiple sequence alignment, interactive sequence choice and visualization. Brief Bioinform. (2019) 20:1160–66. doi: 10.1093/bib/bbx108
39. Castresana J. Selection of conserved blocks from multiple alignments for their use in phylogenetic analysis. Mol Biol Evol. (2000) 17:540–52. doi: 10.1093/oxfordjournals.molbev.a026334
40. Letunic I, Bork P. Interactive Tree of Life (iTOL) v6: recent updates to the phylogenetic tree display and annotation tool. Nucleic Acids Res. (2024) 52:W78–82. doi: 10.1093/nar/gkae268
41. Dean AG, Sullivan KM, Soe MM. OpenEpi: Open Source Epidemiologic Statistics for Public Health, Version 3.01. (2013). Available online at: https://www.openepi.com (accessed June 17, 2025).
42. Ballouard JM, Bonnet X, Jourdan J, Martinez-Silvestre A, Gagno S, Fertard B, et al. First detection of herpesvirus and prevalence of mycoplasma infection in free-ranging Hermann's tortoises (Testudo hermanni), and in potential pet vectors. Peer Community J. (2022) 2:e5. doi: 10.24072/pcjournal.81
43. Marenzoni ML, Stefanetti V, Del Rossi E, Zicavo A, Scuota S, Origgi FC, et al. Detection of Testudinid alphaherpesvirus, Chlamydia spp, Mycoplasma spp, and Salmonella spp in free ranging and rescued Italian Testudo hermanni hermanni. Vet Italiana. (2022) 58:25–34. doi: 10.12834/VetIt.1915.13833.1
44. Highfield AC. Disease Prevention in Tortoise Collections (2014). Tortoise Trust. Available online at: https://www.tortoisetrust.org/articles/disease.html (accessed June 4, 2025).
45. Gracià E, Rodríguez-Caro RC, Ferrández M, Martínez-Silvestre A, Pérez-Ibarra I, Amahjour R, et al. From troubles to solutions: conservation of Mediterranean tortoises under global change. Basic Appl Herpetol. (2020) 34:5–6. doi: 10.11160/bah.196
46. Bertolero A, Cheylan M, Hailey A, Livoreil B, Willemsen RE. “Testudo hermanni (Gmelin 1789) Hermann's Tortoise”. In:Rhodin AGJ, Pritchard PCH, van Dijk PP, Saumure RA, Buhlmann KA, Iverson JB, Mittermeier RA, , eds. Conservation biology of freshwater turtles and tortoises: a compilation project of the IUCN/SSC tortoise and freshwater turtle specialist group. Chelonian Res Monogr. (2011) 5:059.1–20. doi: 10.3854/crm.5.059.hermanni.v1.2011
47. Livoreil B. Distribution of the endangered Hermann's tortoise Testudo hermanni hermanni in Var, France, and recommendations for its conservation. Oryx. (2009) 43:299–305. doi: 10.1017/S0030605307000841
48. Stanford CB, Iverson JB, Rhodin AG, van Dijk PP, Mittermeier RA, Kuchling G, et al. Turtles and tortoises are in trouble. Curr Biol. (2020) 30:721–35. doi: 10.1016/j.cub.2020.04.088
49. Feldman SH, Wimsatt J, Marchang RE, Johnson AJ, Brown W, Mitchell JC, et al. A novel mycoplasma detected in association with upper respiratory disease syndrome in free-ranging eastern box turtles (Terrapene carolina carolina) in Virginia. J Wildl Dis. (2006) 42:279–89. doi: 10.7589/0090-3558-42.2.279
50. Ossiboff RJ, Raphael BL, Ammazzalorso AD, Seimon TA, Niederriter H, Zarate B, et al. Mycoplasma species of Emydidae turtles in the northeastern USA. J Wildl Dis. (2015) 51:466–70. doi: 10.7589/2014-04-086
51. Silbernagel C, Clifford DL, Bettaso J, Worth S, Foley J. Prevalence of selected pathogens in western pond turtles and sympatric introduced red-eared sliders in California, USA. Dis Aquat Organ. (2013) 107:37–47. doi: 10.3354/dao02663
Keywords: captive tortoises, chelonian health, molecular epidemiology, Mycoplasma agassizii, polymerase chain reaction, subclinical infection
Citation: Louro M, Patrício R, Pereira A, Valença A and Alves M (2025) First molecular detection of Mycoplasma agassizii in captive tortoises in Portugal. Front. Vet. Sci. 12:1652362. doi: 10.3389/fvets.2025.1652362
Received: 23 June 2025; Accepted: 04 August 2025;
Published: 25 August 2025.
Edited by:
Scott Henke, Texas A& M University Kingsville, United StatesReviewed by:
Alexandra M. Burne, University of Florida, United StatesJoachim Spergser, University of Veterinary Medicine Vienna, Austria
Copyright © 2025 Louro, Patrício, Pereira, Valença and Alves. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Margarida Alves, bWFyZ2FyaWRhLmFsdmVzQHVsdXNvZm9uYS5wdA==; Andreia Valença, YW5kcmVpYS52YWxlbmNhQHVsdXNvZm9uYS5wdA==
†These authors have contributed equally to this work and share last authorship