 Nahid Akhtar1*
Nahid Akhtar1* Ragini Mishra2Shivakant Tripathi1Santiago Redon-Marin3,4
Ragini Mishra2Shivakant Tripathi1Santiago Redon-Marin3,4 Manik Prabhu Narsing Rao5Andrés Felipe Cuspoca Orduz6Jorge Samuel Leon Magdaleno7
Manik Prabhu Narsing Rao5Andrés Felipe Cuspoca Orduz6Jorge Samuel Leon Magdaleno7 Abdul Rajjak Shaikh1
Abdul Rajjak Shaikh1 Julian Ruiz-Saenz3*Luigi Cavallo7
Julian Ruiz-Saenz3*Luigi Cavallo7 Mohit Chawla7*
Mohit Chawla7*- 1Department of Research and Innovation, STEMskills Research and Education Lab Private Limited, Faridabad, Haryana, India
- 2School of Bioengineering and Biosciences, Lovely Professional University, Phagwara, Punjab, India
- 3Grupo de Investigación en Ciencias Animales - GRICA, Facultad de Medicina Veterinaria y Zootecnia, Universidad Cooperativa de Colombia, Bucaramanga, Colombia
- 4Grupo de Investigaciones Biomédicas Uniremington, Programa de Medicina, Facultad de Ciencias de la Salud, Corporación Universitaria Remington, Medellín, Colombia
- 5Facultad de Ingeniería, Instituto de Ciencias Aplicadas, Universidad Autónoma de Chile, Centro de Investigación e Innovación, Huechuraba, Chile
- 6Grupo de Investigación en Epidemiología Clínica de Colombia (GRECO), Universidad Pedagógica y Tecnológica de Colombia, Tunja, Colombia
- 7Physical Sciences and Engineering Division, KAUST Catalysis Center, King Abdullah University of Science and Technology (KAUST), Thuwal, Saudi Arabia
Feline immunodeficiency virus (FIV) is a retrovirus that infects both domestic and wild cats worldwide, causing a progressive decline in the immune function. FIV infection is a major concern for cat owners, particularly those with outdoor cats or multi-cat households, as it can lead to chronic illness and a reduced lifespan. The development of effective prevention and treatment strategies for FIV is therefore essential to improve the health and welfare of cats. This review article provides an overview of current knowledge on FIV, covering its epidemiology, prevalence, pathogenesis, risk factors, transmission, and management. It also discusses the various FIV subtypes, their geographical distribution, and their associations with different clinical outcomes. In addition, the review examines the clinical and pathophysiological features associated with FIV, including oral and respiratory infections, neurological disorders, renal diseases, and cancer. The review also discusses management strategies for FIV-infected cats, with a focus on advances in the development of antiretroviral drugs and immunomodulators. This review highlights the challenges of developing an effective FIV vaccine and provides a comprehensive summary of the latest advancements in FIV vaccine research. Additionally, it offers an overview of adjuvants used so far in FIV vaccine candidates and explores the potential application of adjuvants currently licensed for other vaccines. Overall, this review paper provides a comprehensive and up-to-date summary of current knowledge on FIV, highlighting key areas that require further research to improve treatment and prevention of this important feline viral infection.
1 Introduction
Feline immunodeficiency virus (FIV) is a lentivirus of the Retroviridae family, first reported in 1986 in California, USA, in domestic cats exhibiting immunodeficiency syndromes (1, 2). Since then, cases of immunosuppressive FIV infections have been reported in domestic cats worldwide (3–8). Beyond domestic cats, FIV has also been reported in bobcats, Pallas' cats, guignas, leopards, pumas, Tsushima leopard cats, and lions (9–15). FIV infection is established through the integration of a provirus, a DNA copy of the viral RNA, into the host genome, leading to lifelong infection (16). The course of infection is characterized by three main phases (17). The first, or the primary infection stage, occurs 3–6 weeks after infection, during which viremia develops and cats exhibit signs of anorexia, pyrexia, lymphopenia, neutropenia, peripheral lymphadenopathy, and malaise, lasting from weeks to months (18, 19). The second phase, the asymptomatic phase, is the longest and can persist for several years. During this period, viral replication occurs at minimal levels, and the cat remains clinically healthy. Notably, some cats may remain in this stage for their entire lifetime (7.5–12.5 years) (16, 20). The final stage is terminal infection, characterized by increased viral replication and the onset of clinical symptoms due to CD4+ lymphocytopenia (16, 18). Many cats infected with FIV can live as long as uninfected cats if provided with appropriate management and high-quality care in household settings. However, they remain predisposed to opportunistic infections and atypical diseases (21, 22). The infecting FIV subtype may also affect clinical outcome. One study reported that, although clinical symptoms and FIV subtypes were not significantly correlated, cats infected with subtype A viruses developed life-threatening conditions, including encephalitis and AIDS-like diseases.
In contrast, cats infected with subtype B viruses displayed either no symptoms or relatively mild manifestations, such as gingivitis and stomatitis (23). Similarly, subtype C has been reported to be more virulent than subtype A, with infected cats exhibiting a greater likelihood of lymphopenia and neutropenia in the first 10–12 weeks of infection. Cats infected with FIV-C also exhibited mean viral RNA levels up to 100-fold higher during the initial weeks of infection compared to those infected with subtype A. Furthermore, FIV-C-infected cats showed significantly elevated levels of proviral DNA in peripheral blood mononuclear cells, and proviral DNA levels in tissues, such as the popliteal lymph nodes, were approximately 10 fold higher at 20 weeks post-infection. These findings were accompanied by more severe histopathological lesions (24).
Since the discovery of FIV, numerous studies on both experimentally and naturally infected domestic cats have provided extensive knowledge regarding the virus, its prevalence, and the pathogenesis of the disease. Furthermore, many studies have been performed to develop vaccine candidates and antiviral drugs for the treatment of FIV. This review summarizes up-to-date information regarding FIV epidemiology, prevalence, and pathogenesis. Additionally, the review delineates the advances made in the search for therapies for the prevention and treatment of FIV in cats.
To ensure a comprehensive coverage of the literature, a systematic search strategy was employed. Relevant publications were retrieved from multiple databases, including PubMed, Web of Science, and Scopus, up to July 2025. The search was performed using the following keywords: “Feline immunodeficiency virus,” “FIV,” “feline immunodeficiency virus” OR “FIV.” Furthermore, the keywords “feline immunodeficiency virus,” or “FIV,” were used in combination with other keywords such as “pathogenesis,” “immune response,” “molecular mechanisms,” “diagnosis,” “treatment,” “vaccine,” and “management.” Some of the combined search terms are (“Feline immunodeficiency virus” AND “Vaccine”), (“Feline immunodeficiency virus” AND “Diagnosis”), and (“Feline immunodeficiency virus” AND “Treatment”). The inclusion criteria comprised (i) peer-reviewed research articles, reviews, and conference proceedings relevant to FIV biology, diagnostics, pathogenesis, and therapeutic approaches published in English and (ii) studies focusing on FIV in domestic cats or closely related felids. The exclusion criteria included studies not directly related to FIV (e.g., studies exclusively on HIV unless comparative) and non-English publications. Additional references were identified by manually screening the bibliographies of key papers.
2 Genome and molecular aspects of FIV
FIV is a positive-stranded RNA virus with a genome of approximately 9,400 nucleotides (25, 26). It contains three genes—gag, pol, and env—which encode the Gag protein, the pol polyprotein, and the envelope polyprotein, respectively (25, 27). The Gag protein is the precursor of structural proteins, including the matrix, capsid, and nucleocapsid proteins. It localizes and captures the viral genomic RNA for packaging within the host cell at the cytoplasmic face of the nuclear envelope (27, 28). In FIV, the packaging signal appears to consist of two parts: the first part spans the initial 250 nucleotides of the 5′ untranslated region, and the second part encompasses the start of the gag gene (27, 29, 30). The Y176/L177 motif in the C-terminal domain of the FIV capsid protein is important for viral infectivity, Gag assembly, and capsid oligomerization (31). Both the C-terminal and N-terminal regions of the capsid protein also contribute to Gag assembly (32). The deletion of the C-terminal p2 peptide of the Gag protein disrupts Gag assembly by eliminating the PSAP budding motif (32). The proximal zinc finger motif of the FIV nucleocapsid protein plays a more significant role in genomic RNA binding and virion production than the distal motif. This conclusion is supported by evidence showing that substituting serine for the first cysteine residue in the proximal zinc finger significantly impaired both genomic RNA binding and virion assembly.
In contrast, mutating the first cysteine residue in the distal zinc finger maintained significant RNA-binding activity in the mutant nucleocapsid protein and had no impact on virion production (33). The Pol polyprotein is cleaved into protease, reverse transcriptase, integrase, and deoxyuridine triphosphatase (26, 34, 35). FIV protease, which is an aspartyl proteinase, cleaves the gag-pol polyproteins during maturation into their respective functional structural proteins and enzymes, along with two other peptides, namely spacer peptide p1 and C-terminal p2 peptide (26).
FIV reverse transcriptase initiates the conversion of the single-stranded RNA genome into double-stranded DNA, a process believed to be regulated by interactions between the extreme 5′ nucleotides of the tRNA primer and a conserved stem-loop in the U5 inverted repeat region (27, 36, 37). Integrase is essential for the insertion of FIV proviral DNA into the host cell genome, with the N-terminal (residues 1–52) and C-subterminal domains (residues 189–235) necessary for 3′-end processing and strand-joining reactions (27, 38). Deoxyuridine triphosphatase functions to lower the concentration of dUTP by converting it to dUMP, which can then be used to synthesize dTTP. This prevents the incorrect insertion of uracil during reverse transcription and thereby reduces the likelihood of mutations in the viral genomic DNA (26, 27). The envelope polyprotein gives rise to surface and transmembrane glycoproteins that mediate FIV attachment and entry into host cells by binding to the CD134 and CXCR4 receptors (39, 40). The envelope glycoprotein gp36 facilitates fusion between FIV and host cells (41). In addition, the envelope protein contains a 175-amino-acid signal peptide at the N-terminus that enables evasion of tetherin, a host restriction factor that inhibits the release of FIV from infected cells (42–44).
Furthermore, the V5 loop of the envelope polyprotein plays an important role in determining whether FIV will be neutralized by virus-neutralizing antibodies in cats (27, 45). Additionally, it has three ORFs: Vif (viral infectivity factor), ORF 2, and Rev. ORF2, which is also known as OrfA, has a role in virion dissemination, transcriptional activation, cell cycle arrest of infected cells, and splicing control (27, 46, 47). Vif is essential for FIV replication (48) and counteracts the activity of apolipoprotein B mRNA-editing catalytic polypeptide 3 (APOBEC3), a feline restriction factor that inhibits FIV viral replication, through a ubiquitin/proteasome-dependent pathway (26, 49). The degradation of APOBEC3 is mediated by the interaction of FIV Vif with elongin B, elongin C, and cullin, which together form an E3 ubiquitination complex (50). In addition, the FIV protease could antagonize APOBEC3 by cleaving it within nascent virions (51). The Rev protein facilitates the export of unspliced and partially spliced FIV RNAs to the cytoplasm (26). Within Rev, amino acids 84–99 contain the nuclear localization signal, whereas amino acids 82–95 form the nucleolar localization signal (52).
3 Genomics and evolution of FIV
Based on the diversity of the V3–V5 region of the env gene, FIV can be classified into six clades (A to F) (23, 53–55). A seventh subtype named U-NZenv has been reported to be regionally distributed only in New Zealand (56, 57). Recently, a molecular study in Egypt proposed a novel FIV subtype, FIV-X-EGY, after phylogenetic analysis of env and gag sequences from infected cats showed that Egyptian strains form a distinct clade, genetically divergent from all known subtypes but with low internal variability and no evidence of recombination (58). Apart from the env gene, the FIV gag gene can also be used for the differentiation into different clades (59, 60). The nested PCR-restriction fragment length polymorphism analysis of a 329-base pair fragment within the FIV gag gene enabled the differentiation of FIV isolates belonging to subtypes A, B, and D, previously classified based on the V3–V5 region of the env gene sequence (60). The phylogenetic analysis of the full genome and the env gene allows the identification of at least six of the seven FIV subtypes (Figure 1), highlighting the high viral diversity among the sequences reported in GenBank. It is important to note that, similar to other retroviruses, FIV evolution is strongly driven by recombination and mutational events (61). A recent study analyzed 60 whole genome sequences (WGS) from the NCBI GenBank and discovered that the majority of recombination events (75%) occurred between wild-type host sequences within similar genomic regions, primarily located at the ends of the pol, ORF1, ORF2, and env genes [60]. In addition, both intra- and inter-subtype recombination events have been observed between the most prevalent FIV subtypes A and B (62, 63), and intra-host viral quasispecies have been reported, collectively contributing to the characteristics of the viral population and increasing viral diversity (64).
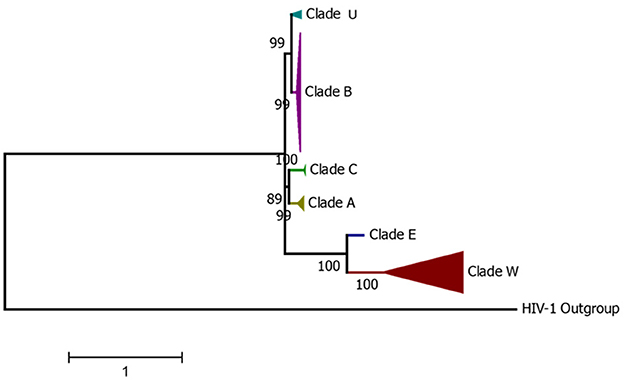
Figure 1. Condensed phylogenetic tree constructed from the alignment of complete FIV genomes from GenBank representing all current FIV-described Clades.
Among these seven subtypes, subtypes A and B are the most widely distributed, while the others are regionally spread (65). Recombinant FIV, such as subtype A/B recombinant and subtype B/F recombinant, has also been reported in North America and South America (66–68). Studies conducted in Brazil and Argentina have shown that subtypes B and E of FIV are present in South America, as revealed through the examination of either partial or complete genome sequences (66, 69). Additionally, studies conducted in Brazil and Colombia have identified subtype A of FIV (66, 70). Subtype D has been reported in Japan and Vietnam (71, 72). Figure 2 shows the subtypes of FIV reported in different countries.
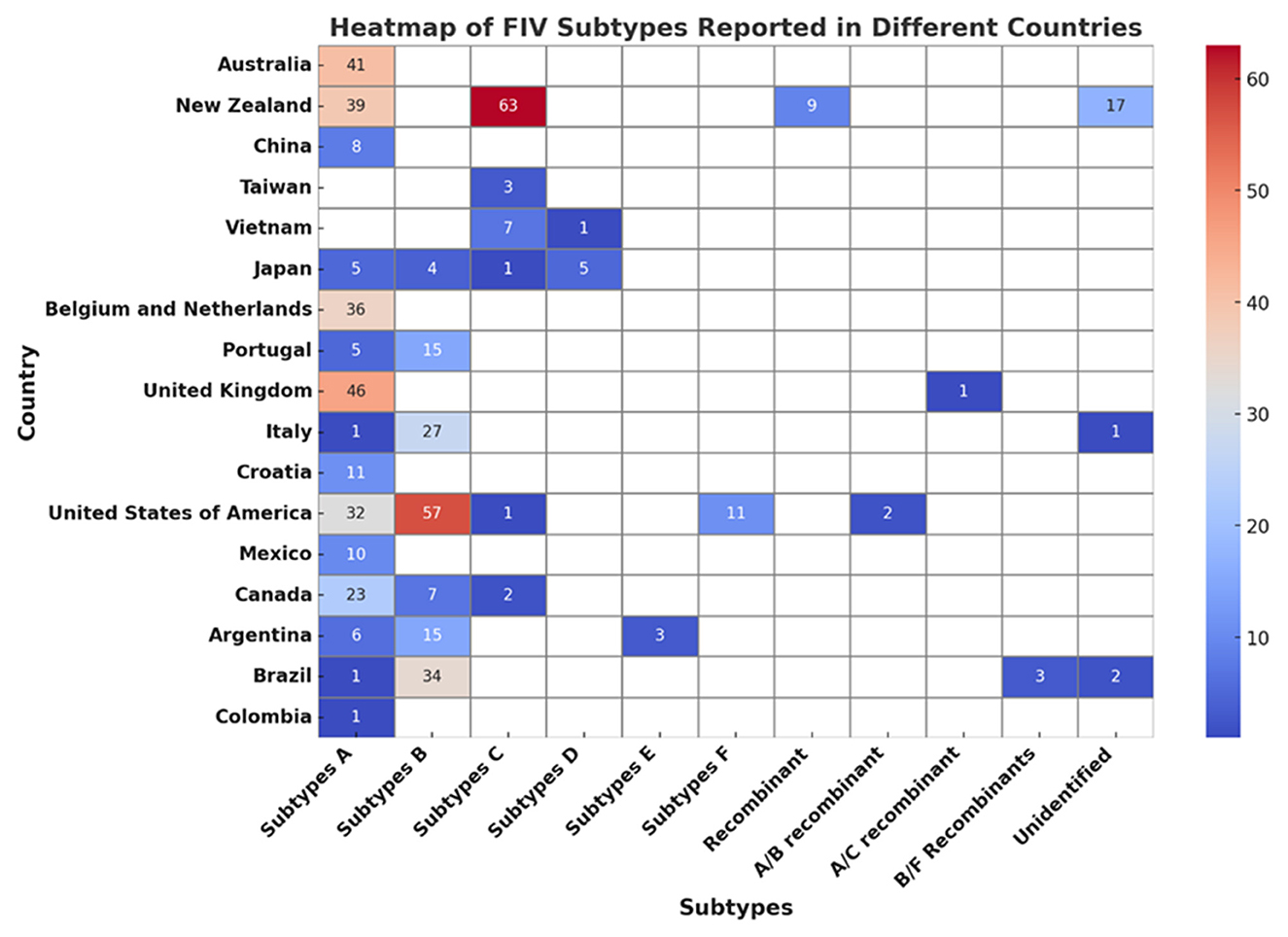
Figure 2. Heatmap analysis of FIV subtypes across different countries.
4 Prevalence of FIV
FIV is found worldwide and exhibits higher seroprevalence in certain regions (7). A recent study found that FIV prevalence typically ranges between 5% and 8%, with a global average of 4.7% (73). The study further reported that the prevalence of FIV ranged from 2.19% to 23% in North America, 2.2–8.8% in Central America, 3.1–27% in Europe, and 3–22% in South America (73). A meta-analysis further revealed seropositivity rates of 5.93% in North America, 8.98% in Europe, 9% in Africa, 9.43% in South America, 10.9% in Central America, 11.9% in Oceania, and 14.34% in Asia (7). The rate of FIV infections in cats has been reported to be 15–16% for adult pet cats with outdoor access in Australia, where subtype A is more prevalent (4, 74). In European countries such as Germany and Ireland, FIV prevalence rates of 3.2% and 10.4%, respectively, have been reported (5, 75). The prevalence of FIV in different countries throughout the world is listed in Table 1. Based on the data from serological tests in Colombia, the prevalence of FIV has been determined to range from 6.7% to 13% (70). Whether the cats were healthy or unhealthy could also affect the prevalence rate of FIV. In a seroprevalence study of FIV in Canada and the USA, the overall seroprevalence was 3.6% but in cats infected with oral disease, respiratory disease, or bite wounds or abscesses, the seroprevalence was 9.7%, 6.4%, and 12.5%, respectively (76). A study indicated that FIV disease is more widespread among cats displaying aggressive behavior. Additionally, the aggressive FIV-infected cats were found to be more prone to having an unhealthy status when compared to their non-aggressive FIV-infected counterparts (77). Moreover, it has been reported that stray cats have a higher FIV seroprevalence (17.8%) than domestic cats that were relinquished by their owners (7.5%) (78). Additionally, FIV is more prevalent in older male cats (73).
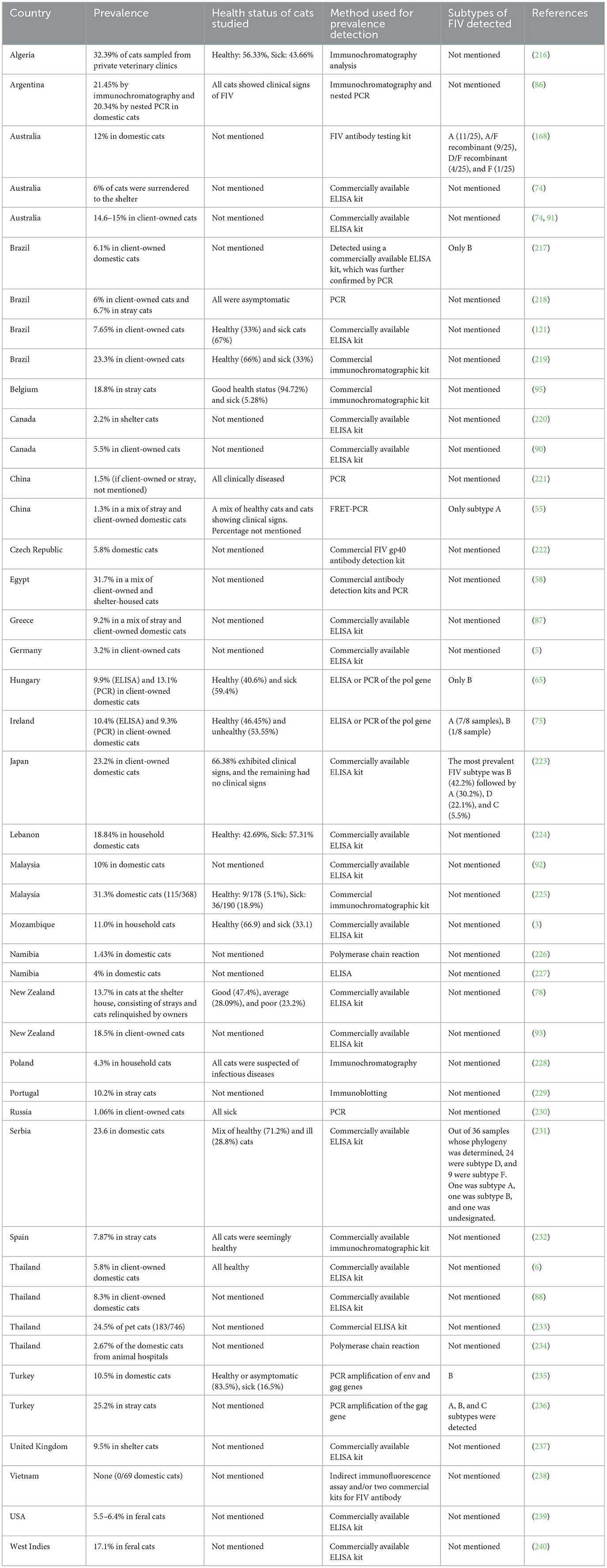
Table 1. Prevalence of FIV in some countries.
5 Infection and transmission of FIV
The transmission of FIV occurs primarily through blood during bites (17). In households with socially well-adjusted cats, the likelihood of transmission is minimal. However, transmission can occur from an infected mother to her kittens, particularly if the mother is experiencing a severe infection. Transplacental transmission can occur in FIV-infected queens, but not all kittens in a litter may be infected. The overall rate of this type of transmission in the first year of infection is approximately 70% (79). The risk of mother-to-kitten transmission is higher in queens with a CD4+ count of less than 200 cells/μL, those showing signs of immunodeficiency, and those who contracted the virus within the last 15 months (80, 81). FIV-infected queen cats could also disseminate the virus to offspring via milk (82). Cats with FIV remain persistently infected, even though they can produce both antibody and cell-mediated immune responses (22). There is also a possibility of sexual transmission of FIV, as cell-free FIV has been detected in semen of naturally and experimentally FIV-infected cats (83). There is a possibility of transmission of FIV from naturally infected cats to uninfected cats in mixed households, but there are conflicting reports and a lack of evidence to support it. A study reported that, despite cohabiting the same household for years, there was no evidence of FIV transmission from infected cats to uninfected cats (84). Experimental infection of FIV in cats has shown that other methods of transmission, such as oral-nasal and rectal/vaginal mucosal transfer, could also be effective for transmitting the virus (85).
6 Risk factors of FIV infection in cats
Adult male cats with outdoor access are at a higher risk of FIV infection (86). Older cats are more frequently infected with FIV, likely due to its prolonged incubation period. During this time, cats can remain in an asymptomatic phase for several years, with minimal effects on morbidity and mortality rates (87). A study reported that male cats are four times more likely to test positive for FIV compared to female cats (87). Other studies have also reported that male cats are more susceptible to FIV infection (3, 76, 88). From a behavioral standpoint, male cats exhibit higher levels of aggression compared to female cats, which increases their risk of sustaining bite wounds and subsequently enhances the likelihood of FIV transmission (87). Another study reported that male urban feral cats exhibiting bold behavior have a higher probability of FIV infection (89). Cats with aggressive behavior are more likely to be infected with FIV (77). Bite wounds, oral diseases, and lethargy are significantly associated with FIV infection (90). Furthermore, mixed-breed and domestic-breed cats are also at higher risk of FIV infection than purebreds (91–93). The low prevalence of FIV among purebred cats may be attributed to their tendency to be kept indoors in smaller groups and their higher likelihood of being vaccinated (92).
Another study found that the American Wirehair and Persian breeds were less susceptible to FIV infection than domestic shorthair cats (88). It has been reported that neutered cats are less likely to be infected with FIV (74, 88). However, other studies have reported that neutered cats are more likely to be FIV-infected (94, 95). Feline leukemia virus co-infection could increase the risk of FIV as well (5). Cats living in multi-cat households are at higher risk of FIV infection than those in single-cat households (88). However, a previous study did not show any evidence to corroborate FIV transmission from naturally FIV-infected cats to non-infected cats in a mixed household (84). FIV transmission among cats sharing the habitat is less common unless they fight (84, 96). Symptoms such as weight loss, skin lesions, and/or pruritus, hyperglobulinemia, and gingivostomatitis are also associated with FIV seropositivity (87).
High testosterone levels have also been reported to be significantly related to FIV infections in cats (97). Aggressiveness in cats is known to be mediated by testosterone, which aligns with the findings showing a higher rate of FIV infection in males with elevated testosterone levels (97). Reduced levels of red blood cells and an albumin-to-globulin ratio below 0.6 are also associated with FIV infection (98). Cats in low socioeconomic status areas are at a higher risk of FIV infection (94). Cats infected with FIV are 1.6–2.3 times more likely to reside in areas of low socioeconomic status. This increased prevalence may be attributed to limited awareness of pet healthcare and a lower willingness or ability to invest in preventive measures, such as vaccinations, among residents in these areas (94).
7 Clinical and pathophysiological features of FIV
Typically, cats infected with the virus do not display any noticeable clinical symptoms for several years, and the development of the disease may be influenced by the strain of the virus that has caused the infection. In some cases, cats may not exhibit any symptoms at all (80). The most common clinical signs of FIV infection are secondary infections and immunodeficiency, resulting from a decrease in CD4+ cells, immunological anergy, and cytokine dysregulation (22, 80, 99). Secondary infections with Toxoplasma gondii, Cladosporium carrion, and Leishmania infantum have been reported in FIV-positive cats (100–102). Similarly, FIV-associated immunodeficiency has also been reported to facilitate parasitic infections such as Eucoleus aerophilus and Cytauxzoon sp. in cats (103, 104). Common symptoms of FIV include stomatitis, weight loss, lethargy, peripheral lymphadenopathy, mild fever, and chronic rhinitis (20, 22, 99). Depending upon the duration of infection, FIV can cause different hematological changes, such as anemia, leukopenia, eosinopenia, lymphopenia, pancytopenia, thrombocytopenia, hypochromia, hyperglobulinemia, and neutropenia in cats (4, 105, 106). FIV-infected cats have higher gamma-globulin concentrations due to increased antibody production by B cells (known as B cell expansion), triggered by the direct and indirect effects of the virus, including altered cytokine production and activation of specific T cell populations (4, 107).
FIV infection can lead to neurological abnormalities, including impaired motor function and cognitive deficits, in both naturally and experimentally infected cats (108, 109). These effects are associated with neuronal loss in the parietal cortex and hippocampus, as well as reduced glutamate receptor expression, which correlates with viral load and neuroinflammation (109). FIV infection may also act synergistically with age-related cognitive impairment, further exacerbating neurocognitive dysfunction in older infected cats (110). Reported behavioral neurological abnormalities in FIV-infected cats include facial and tongue twitching, delayed pupillary reflexes, psychotic behavior, sleep disturbance, and ataxia (99, 111, 112). A recent study has shown that the FIV glycoprotein gp95 increases Alzheimer's disease-related cellular tau pathology through cGMP-dependent kinase II (113). FIV-infected cats are five times more likely to develop tumors than non-infected cats (99). Among these, malignant lymphoma is the most common, although the underlying mechanism remains unclear (114). FIV is believed to contribute to lymphomagenesis primarily through indirect mechanisms, such as defective cell-mediated immunity or chronic lymphocyte activation. Direct involvement has been reported only once, in a case of clonal integration of the FIV genome (99, 114–116). Other tumor types observed in FIV-infected cats include fibrosarcoma, mast cell tumor, leukemia, and squamous cell carcinoma (80, 99, 117, 118). In addition, FIV can cause hyperglobulinemia and, in rare cases, bone marrow suppression (99, 119). Renal damage has also been associated with FIV infection, including glomerulonephritis, glomerulosclerosis, mesangial widening, and both interstitial and glomerular amyloidosis (120).
A study reported a case of plasma cell pododermatitis in a cat co-infected with FIV and feline leukemia virus, presenting clinical symptoms such as erythematous swelling of the paw pads, skin peeling, and alopecia (121). FIV has been linked to myocarditis, inflammatory myopathy, and hypertrophic cardiomyopathy in cats, with evidence of FIV infection detected in inflammatory cells within the myocardium (122, 123). FIV-infected cats have been reported to have significantly lower serum 25-hydroxyvitamin D concentrations than healthy control cats, similar to the lower serum levels of vitamin D in HIV-positive patients (124). FIV-infected cats have been reported to have a higher urinary protein-to-creatinine ratio and serum creatinine than healthy cats (125). Recently, FIV co-infection with T. gondii and Mycoplasma hemominutum has been reported to be associated with hemophagocytic syndrome in cats (126). A statistical association between FIV infection and infection with Mycoplasma hemofelis or Mycoplasma hemominutum has also been described, but it is not clear if FIV infection is a true risk factor for hemoplasmosis in cats (127, 128).
Considering the biology and clinical signs of FIV, FIV infection can lead to a progressive disruption of immunological functions in cats, similar to the pathogenesis of HIV, involving several key mechanisms and clinical manifestations (80). Hematological abnormalities have been reported since cats infected with FIV frequently exhibit lower levels of red blood cells (RBCs), hemoglobin, hematocrit, lymphocytes, and platelets in comparison to uninfected cats (98). Moreover, neutropenia is frequently observed in cats infected with FIV, as well as anemia and thrombocytopenia, compared to cats that are not infected (107). On the other hand, FIV infection results in changes to cytokine profiles, notably with elevated levels of interleukin-10 (IL-10) and interleukin-12 (IL-12) in treated cats, indicating modulation of the immune response. In contrast, untreated cats show a significantly lower IL-10/IL-12 ratio, which suggests a shift toward a more inflammatory immune response (129).
As expected, relevant clinical findings in infected cats have been proposed since FIV leads to a gradual reduction in CD4+ T lymphocytes, resulting in immunodeficiency and heightened vulnerability to secondary infections and neoplasia. Additionally, proteinuria is more frequently observed in FIV-infected cats, affecting 25% of infected individuals compared to 10.3% in non-infected cats (130). Furthermore, neurological symptoms and neoplasia, especially lymphoma, are frequently observed in FIV-infected cats (4). While FIV-infected cats have a higher likelihood of developing lymphoid malignancies, the association is less pronounced compared to feline leukemia virus infection (131).
Distinguishing FIV-related pathology from the comorbidities of aging in cats remains a major clinical challenge, since many conditions can occur independently of viral infection. Differentiating FIV-related pathology from age-associated comorbidities requires a multifaceted diagnostic approach. In naturally infected cats, a progressive inversion of the CD4+/CD8+ ratio (33) and elevated proviral load (34) are strongly associated with immune decline and clinical disease, whereas normal aging may not show these immunologic shifts. Quantitative molecular markers—viral RNA or integrated proviral DNA—have substantial potential for confirming active infection rather than attributing symptoms to age alone, which can be tested using PCR-based assays (35).
Neurologic and cognitive deficits, which may manifest subtly, are often overlooked in older cats unless specifically assessed. Opportunistic infections or neoplasms should be interpreted in the context of immunological indicators (CD4+/CD8+ T-cell ratios, lymphopenia, and hyperglobulinemia) and risk factors (outdoor access, fighting behavior, known FIV exposure), rather than attributed to aging alone. Monitoring immune and viral markers over time can therefore help clinicians distinguish FIV-driven pathology from coincidental age-related disorders.
8 Cellular and tissue reservoirs of FIV
FIV infects various cell types. For entry and infection, FIV targets CD4+ cells by binding its major surface glycoprotein to the CD134 receptor present on CD4+ cells, causing depletion of CD4+ cells (40, 80, 132). FIV can also infect CD8+ T cells by binding to CXCR4 receptors (40). In the later phase of FIV infection, it can also affect B cells, with a study reporting that the FIV provirus was most abundant in B cells in cats infected for more than 5 years (40, 133). FIV has been reported to infect and activate CD4+CD25+ regulatory cells throughout the acute phase of infection (134). CD4+CD25+ regulatory cells negatively regulate the immune response by inhibiting the proliferation and causing apoptosis of activated CD4+ and CD8+ cells (80, 135). FIV is also capable of infecting other leukocytes, including monocytes and dendritic cells (136, 137). In monocytes, adherence induces the expression of viral antigens (136). Additionally, FIV interacts with dendritic cells during the early stages of infection, enabling these cells to transfer the virus to susceptible T cells, thereby initiating a significant burst of viral replication (137). In addition to leukocytes, Feline Immunodeficiency Virus (FIV) has been shown to infect various cells in the central nervous system, including astrocytes and microglial cells. Infection in astrocytes impairs their ability to scavenge extracellular glutamate, while in microglial cells, FIV infection occurs during the subclinical phase and facilitates viral dissemination within brain tissue (138, 139).
Furthermore, megakaryocytes, stromal fibroblasts, and mononuclear cells in the bone marrow have also been reported to be infected by FIV in cats, where these cells could act as targets and reservoirs of the infection (140, 141). Since the primary mode of FIV transmission is biting, the salivary gland of cats could act as a reservoir of FIV during the early stages of infection. A study has reported the infection of epithelial cells of the interlobular duct of the salivary gland in cats (142). Besides cellular reservoirs, various tissues can also act as a reservoir of FIV in cats. Lymph nodes, spleen, thymus, gastrointestinal tract, reproductive tract, liver, brain, and bone marrow can all harbor FIV during the late, asymptomatic phase of infection (17, 143–146).
9 Anti-FIV drugs
Cats infected with retroviruses require specialized care and management, which, when provided, can enable them to live healthy lives for many years. Most retrovirus-infected cats are effectively managed through symptomatic therapy, while antiviral chemotherapy is recommended only in exceptional cases of FIV infections due to the limited proven efficacy and potential toxicity of antiviral drugs (147). The antiviral drugs commonly used in cats have been authorized for use in humans and are specifically designed to treat the infection caused by the human immunodeficiency virus (HIV) (147). Combination antiretroviral therapy (cART) has been reported to alleviate FIV-associated oral disease by maintaining the integrity of the oral mucosal microbiota in FIV-infected cats (148). However, many of these antiviral drugs have shown ineffectiveness in treating FIV in cats or cause adverse side effects (149). FIV-infected cats treated with zidovudine, a nucleoside reverse transcriptase inhibitor, which is used for the treatment of HIV, have been reported to be resistant to the long-term antiretroviral therapy (150). Additionally, fozivudine tidoxil, which is a lipid-zidovudine conjugate, has been reported to decrease plasma and cell-associated viremia during the first 2 weeks of infection but was ineffective in protecting cats from FIV infection, as all cats were infected by 6 weeks (151). Didanosine, another medication used for the treatment of HIV, has been reported to have antiviral activity against FIV in vitro and in animal studies but caused toxic neuropathy in cats (147, 152).
Other than the HIV antiviral drugs, various other strategies have been explored for the treatment of FIV. Various peptides have shown the ability to inhibit the replication of FIV in different feline cell lines (Table 2) (153, 154). Peptides 5–7, spanning amino acids E225 to P264 in a conserved region of the surface protein of the Petaluma isolate of FIV, effectively inhibited FIV-induced syncytium formation and suppressed viral replication in a time-dependent manner (153). Peptide 59, a 20-mer synthetic peptide derived from the membrane-proximal ectodomain of the FIV transmembrane glycoprotein, demonstrated the ability to inhibit the growth of tissue culture-adapted FIV in feline fibroblastoid CrFK cells (154). Similarly, RNA interference technology (lentiviral vector expressing a short hairpin RNA targeting the gag gene of FIV) has also shown the ability to inhibit FIV replication in cell lines that were chronically infected with FIV (155). Seetaha et al. reported the potential of crude extracts of different medicinal mushrooms to inhibit FIV reverse transcriptase in vitro, where ethanol extract from dried fruiting bodies of Inonotus obliquus and hexane extract from dried mycelium of I. obliquus showed the strongest inhibition with IC50 values of 0.80 ± 0.16 μg/mL and 1.22 ± 0.20 μg/mL, respectively (156). Derivatives of different compounds, such as 8-Difluoromethoxy-4-Quinolone, 1,2,3-dithiazole, and T140 derivatives (Table 2) (157, 158).
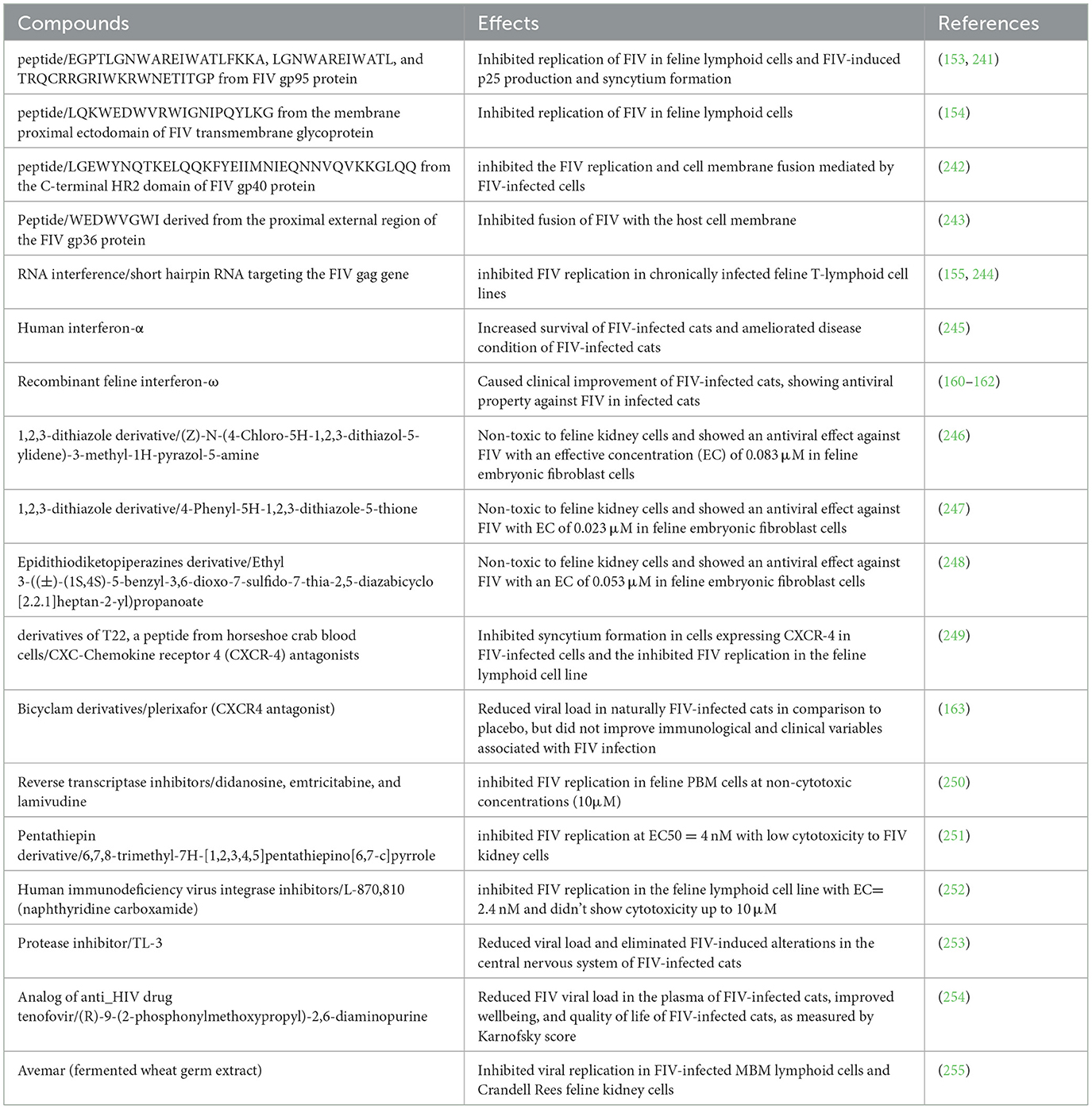
Table 2. FIV inhibition activity of different compounds.
Other therapies, such as the treatment of FIV-infected cats with recombinant human interferon-α, have also been explored. The recombinant human interferon-α therapy did not affect the function of the kidney or liver and ameliorated the clinical signs of FIV infection in cats naturally infected with FIV during the course of the treatment (159). Although the cats remained clinically healthy once the therapy was stopped, signs such as cytopenia and a reduction in CD4+/CD8+ worsened after the therapy discontinuation (159). Recombinant feline interferon omega was the first interferon approved for veterinary use in cats (160). Studies have demonstrated its potential to enhance innate immunity, leading to a reduction in clinical symptoms and co-infections in cats naturally infected with FIV (161, 162). While immunomodulators such as recombinant interferons (e.g., recombinant human IFN-α, recombinant feline IFN-ω) can alleviate clinical signs in FIV-infected cats (36, 37), their long-term use warrants careful monitoring to avoid potential immune dysregulation. The use of interferon can increase the risk of neutropenia, decreased blood counts, kidney dysfunction, infections, and concomitant diseases (36, 38, 39). To balance symptomatic relief with safety, a comprehensive monitoring protocol is essential, including periodic measurements of complete blood counts, thrombocytopenia, leukopenia, creatinine, creatine kinase, and liver function (39). In addition, regular clinical evaluations should assess the emergence of opportunistic infections or organ dysfunction. Employing such multi-parametric surveillance in long-term studies will help ensure that treated cats benefit from improved clinical management without unintended immune side effects.
Although some drugs reduce viral load in naturally FIV-infected cats, other drugs have been unsuccessful in providing protection to cats from acute FIV infection and in ameliorating the clinical and immunological signs of FIV infection in these cats (151, 163). Similarly, treatment with bee venom melittin has improved the general health status of FIV-infected cats but did not influence the immunological parameters such as the CD4/CD8 ratio and lymphocyte subpopulations (164).
Given the limited efficacy and tolerability of HIV-derived drugs in cats, future antiviral discovery for FIV should follow a feline-specific prioritization framework. The most promising candidates will combine a strong safety profile in cats with a high genetic barrier to resistance and broad activity across diverse FIV clades. Agents should demonstrate activity in primary feline lymphocytes and macrophages and show predictable pharmacokinetics suitable for long-term use. Equally important are resistance surveillance, cost-effective formulation for veterinary practice, and translational value from cross-lentivirus research, while ensuring host compatibility in cats.
10 FIV vaccines
According to the Vaccination Guidelines Group (VGG) of the World Small Animal Veterinary Association (WSAVA), the FIV vaccine is considered one of the non-core vaccines for pet cats (165). Significant efforts have been made to create a preventive vaccine for FIV, resulting in Fel-O-Vax FIV. This vaccine contains two strains of FIV—FIV Petaluma (subtype A) and FIV Shizuoka (subtype D)—and was approved for use in cats over 8 weeks old in Australia, Canada, and the USA (166, 167). A case-control field study showed that Fel-O-Vax, the only FDA-approved vaccine against FIV, has only a 56% protective rate among client-owned cats in Australia, raising doubt about the efficacy of the vaccine under field conditions (168). Similarly, another study has reported that the cats vaccinated with Fel-O-Vax did not generate broadly neutralizing antibodies, suggesting that Fel-O-Vax protection may not be effective against certain highly infectious recombinant strains of FIV that are currently circulating in Australia (169). Due to this, the Fel-O-Vax vaccine was removed from the market in the USA and Canada in 2017, although it remains available in Japan, Australia, and New Zealand. According to the WSAVA, it has never been licensed in Europe (165).
Researchers worldwide have been actively engaged in the development of vaccines against FIV (Table 3). However, FIV vaccines developed to date have shown limited effectiveness (168, 170, 171).
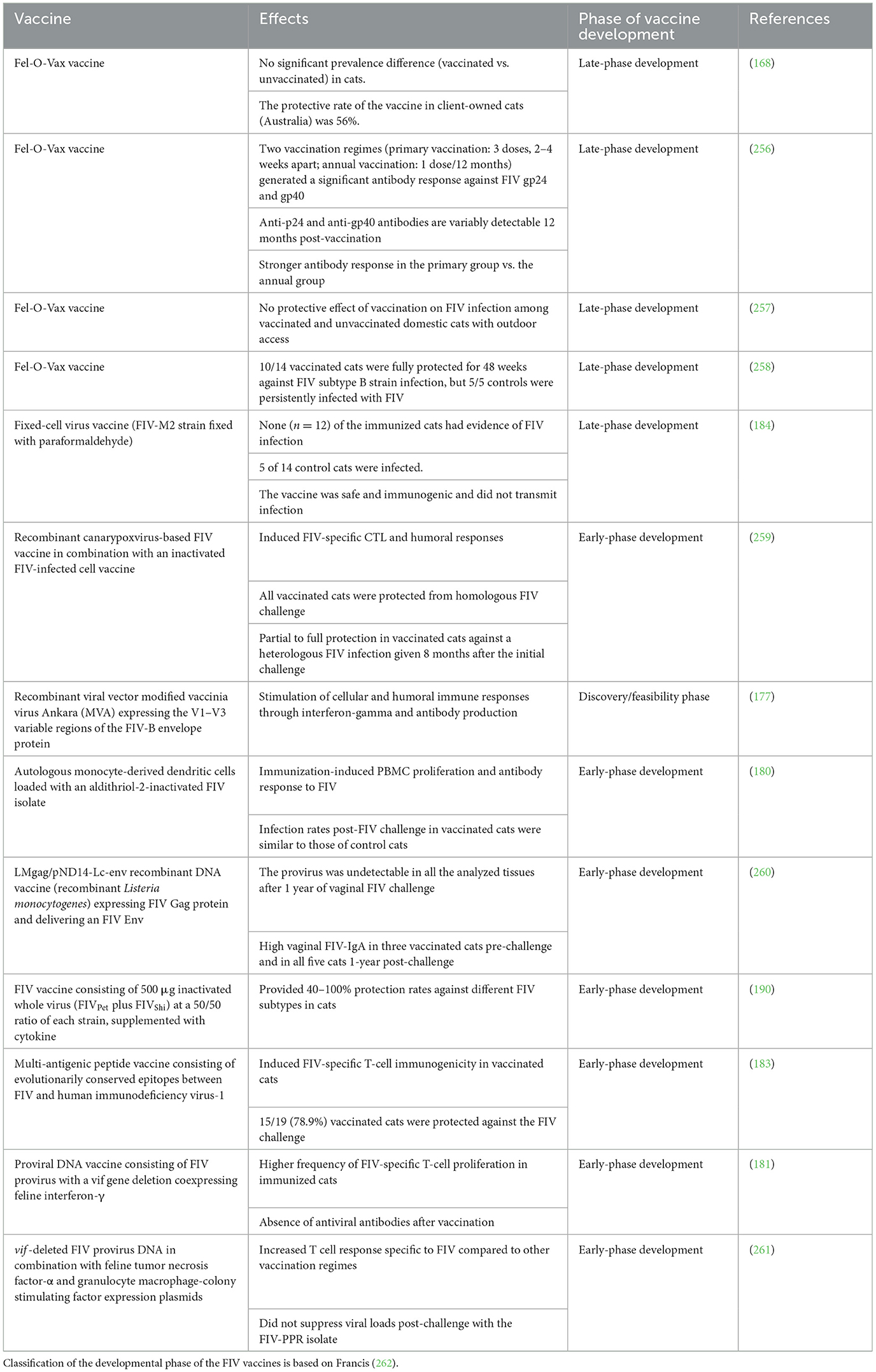
Table 3. FIV vaccines and their status.
Inactivated or attenuated FIV preparations, as well as FIV proteins administered with or without adjuvants, have demonstrated only limited efficacy in protecting against FIV infection in cats (171–174). Vector-based approaches have also been largely unsuccessful: Semliki Forest virus and modified vaccinia virus Ankara (MVA) vectors expressing FIV Rev and OrfA did not elicit sufficient cell-mediated and humoral immunity and were unsuccessful in protecting cats against FIV (175). Moreover, one study reported increased viremia in cats vaccinated with recombinant vaccinia virus expressing the FIV envelope protein incorporated into an immune-stimulating complex (176). Recently, Andrade et al. developed an MVA-based vaccine expressing the variable V1–V3 region of the FIV-B envelope protein, which successfully induced both cellular and humoral immune responses in mice. However, the efficacy of this vaccine candidate has not yet been evaluated in cats (177).
A vaccine based on purified feline immunodeficiency virus (FIV), incorporated into immune-stimulating complexes (ISCOMs), and recombinant FIV p24 ISCOMs successfully elicited antibodies against the core protein p24. However, it failed to produce virus-neutralizing antibodies and did not protect cats from FIV infection when challenged intraperitoneally with 20 infectious units of FIV. Notably, vaccinated cats became viremic earlier than their unvaccinated counterparts (171). Moreover, the ISCOM adjuvanted vaccine candidate containing FIV OrfA and Rev proteins also showed immunogenicity but could not protect cats from the FIV challenge (178). Similarly, other vaccine candidates have also shown ineffectiveness in providing protection against FIV (173, 174).
The attenuated FIV strain lacking ORF-A was tested as a vaccine candidate in nine cats challenged with wild-type FIV (179). Of the nine cats, only three were free from the challenge virus, and in the remaining vaccinated cats, CD4 lymphocyte counts and viral loads were either unaffected or changed only slightly and transiently (179). Similarly, dendritic cells loaded with inactivated FIV generated FIV-specific antibodies, but the frequency at which the challenge virus infected the vaccinated cats was similar to that in the control animals (180). Additionally, another vaccine candidate, an attenuated FIV in which the vif gene has been deleted and co-expressed interferon-γ, was also unable to provide protection to immunized cats from challenge with a biological FIV isolate (181).
Another vaccine candidate consisting of FIV CD134 or surface glycoprotein alone or in complex generated neutralizing antibodies against CD134 and surface glycoprotein, but it could not provide protection to vaccinated cats from FIV challenge (174). A DNA vaccine consisting of replication-defective FIV due to deletion of FIV integrase (ΔIN) or reverse transcriptase (ΔRT) genes provided protection to only 5/18 (ΔIN) and 2/12 (ΔRT) vaccinates challenged with a low virulence strain of FIV, i.e., FIV-Petaluma. However, rechallenging the cats protected from the FIV-Petaluma strain with the relatively more virulent FIV Glasgow-8 strain did not provide sterilizing immunity against the more virulent FIV strain (182). The vaccine candidates studied so far have not shown the desired effectiveness; hence, new avenues should be continuously explored for the development of novel vaccine candidates against FIV. One approach could be to develop multi-antigenic peptides consisting of immunogenic T cell and B cell epitopes as vaccine candidates. Sahay et al. designed a multi-epitope vaccine containing conserved T-cell epitopes from reverse transcriptase and p24 FIV proteins, which protected 78.9% of vaccinated cats from FIV challenge (183).
11 Adjuvants used in FIV vaccines
Several experimental FIV vaccine candidates have been developed and tested in cats, some of which have included adjuvants to improve their effectiveness. In one study, dendritic cells were used as live adjuvants to improve the immune response of a whole-inactivated FIV vaccine (180). Animals that received the vaccine showed obvious signs of increased peripheral blood mononuclear cell proliferation and antibody titers specific to FIV following immunization. Nevertheless, the challenge of vaccinated cats with the homologous virus was unsuccessful in providing protective immunity against the infection and further reduced CD4+T lymphocyte numbers in the vaccinated cats post-challenge (180). Another study has used alum, an adjuvant that has been allowed for use in cats, but the formulated vaccine did not protect cats from FIV infection (174). Similarly, recombinant protein vaccines with ISCOM adjuvant were also unsuccessful in protecting cats from the FIV challenge (178). The use of incomplete Freund's adjuvant in a fixed-cell FIV vaccine and adenyl-muramyl dipeptide adjuvant in a fixed-cell dual-subtype FIV vaccine has also been reported in FIV vaccine candidates (184, 185). In some vaccine candidates against FIV, adjuvants have not been used (175, 177, 186, 187). One experimental FIV vaccine, which is approved for use in cats, known as the Fel-O-Vax FIV vaccine, contains an oil emulsion adjuvant called Fort Dodge-1 adjuvant (188–191). A multi-antigenic peptide vaccine candidate against FIV has also been used with Fort Dodge-1 adjuvant (183).
Different DNA vaccines against FIV have used molecular adjuvants, including IL-18 DNA, IL-12 plus IL-18 DNA, and IFN-γ (181, 182, 188, 192). However, these vaccines were also not very effective in providing protection from the FIV challenge in cats (188). The inclusion of IFN-γ DNA adjuvant did not enhance the efficacy of immunization with a DNA vaccine (FIV-pPPΔvif DNA) (181). In another study, DNA vaccination with an IFN-γ DNA adjuvant did not protect cats from post-challenge FIV infection (193). Cats immunized with a DNA vaccine (FIVGL8ΔRT), along with IL-12 plus IL-18 DNA or IL-12 alone, did not produce antiviral antibodies and showed a reduction in virus-specific CTL activity. However, immunization with a viral DNA (FIVGL8ΔIN) with an IL-18 DNA adjuvant induced virus-specific CTL activity. Nevertheless, both DNA vaccines, FIVGL8ΔRT and FIVGL8ΔIN, with cytokine DNA adjuvants, were not able to provide sterilizing immunity against the virulent FIV strain challenge post-vaccination in cats (182). Vaccination with an FIV DNA vaccine containing FIV gp140 DNA along with feline IL-12 DNA showed that the addition of IL-12 DNA significantly enhanced the response against FIV, and this vaccine protected three out of four cats from challenge infection (192). Similarly, cats immunized with an FIV gp140 DNA vaccine along with feline IL-16 or feline cytosine phosphoguanosine (CpG) had less proviral DNA in PBMCs and became less viremic after FIV challenge infection compared to cats vaccinated only with FIV gp140 DNA, suggesting the potential of IL-16 and CpG as possible adjuvants in FIV vaccines (194).
Overall, there is only one licensed FIV vaccine available that uses an adjuvant. The development and testing of experimental FIV vaccines with adjuvants suggest that adjuvants may be an important component in the development of an effective FIV vaccine. Further research is needed to determine the most effective adjuvant strategies for FIV vaccines. In future adjuvant system families, such as AS01, AS03, AS04, MF59, and CpG, which are used in other licensed or nearly licensed virus vaccine candidates, can be tested as adjuvants for FIV vaccines (195, 196). Furthermore, the adjuvant potential of a new adjuvant called Matrix-M, containing fraction-A and fraction-C of Quillaja saponaria Molina extract and used in a COVID-19 vaccine authorized by the European Medicines Agency, can be explored for developing FIV vaccines (197). AS01 and AS04 contain MPL (3-deacylated monophosphoryl lipid), a toll-like receptor-4 (TLR-4) agonist.
Similarly, other adjuvants that can activate other TLR molecules, such as TLR-7 and TLR-8, should also be analyzed for FIV vaccines. In this regard, Alhydroxiquim-II, an adjuvant used in the COVID-19 vaccine COVAXIN, can be used.
12 Immunoinformatics on FIV research
Immunoinformatics has recently been employed in the development of viral agent vaccines, involving the application of computational methods and resources to study immune system functions. Immunoinformatics enables the precise storage and analysis of immune-related data, facilitating a deeper understanding of immune system mechanisms and functions, which in turn helps in the development of vaccines (198). Epitope-based vaccines assisted by computational tools, derived from viral immunodominant antigens, have been used to develop vaccine alternatives since their activation of helper CD4+ T cells, CD8+ cytotoxic T cells, and B cell activation through helper T cells, allowing them to differentiate into plasma cells that produce antibodies, which are essential for the complete clearance of viruses from the host (199). Although there is no significant research based on the immunoinformatics approach for FIV, different studies have employed this approach to advance new vaccines for veterinary viral agents, such as canine distemper virus (200), canine circovirus (201), canine parvovirus (202), and FIPV (203), among others. Moreover, immunoinformatics has been utilized in the context of HIV to explore T- and B-cell epitopes based on genomic information and antigenic proteins, such as gp120 (204–206). Thus, further studies must be conducted to develop a new generation vaccine based on computationally predicted multiple epitopes for FIV.
13 Future perspectives
High-throughput genomics, transcriptomics, and proteomics in cats could provide valuable insights into the signaling pathways and molecular mechanisms underlying FIV infection. These approaches may also reveal immune-modulatory pathways and novel biomarkers, thereby facilitating the development of targeted therapies and the discovery of new drug targets.
Recent advances in gene-editing technologies, particularly CRISPR/Cas systems, offer exciting opportunities for the control of FIV. CRISPR-Cas systems hold promise for reducing the proviral load of FIV by suppressing viral transcription and limiting the production of infectious virions and potentially achieving a functional cure (207). Such strategies have been investigated in HIV research with promising outcomes (208), and adapting similar approaches for FIV could open novel therapeutic avenues. Beyond proviral excision, CRISPR-based tools can also be used to edit host factors and co-receptors critical for viral entry to generate resistance in susceptible feline cells (209). In addition to therapeutic applications, gene-editing techniques hold potential for vaccine development. CRISPR/Cas can accelerate the design of attenuated or replication-deficient viral strains that serve as safe and immunogenic vaccine candidates. These strategies may overcome the limitations of conventional vaccine platforms, which have shown inconsistent efficacy in cats.
The development of antiviral drugs for FIV remains an underexplored area compared to HIV research, despite the structural and pathological similarities between the two viruses. Future efforts should prioritize drug discovery targeting conserved viral proteins, which are essential for FIV replication and represent viable therapeutic targets. Structure-based drug design, aided by advances in crystallography, molecular docking, and molecular dynamics simulations, can accelerate the identification of small-molecule/natural compound inhibitors with high specificity and low toxicity (210).
mRNA-based vaccine approaches also represent a promising avenue for FIV prevention. Unlike traditional inactivated or recombinant protein vaccines, mRNA vaccines can be rapidly designed to encode multiple conserved FIV antigens and delivered using lipid nanoparticles, eliciting both strong humoral and cellular immune responses. The success of mRNA platforms against emerging human viral pathogens, such as SARS-CoV-2 (211, 212), demonstrates their flexibility and scalability; similar strategies could be adapted for veterinary use. For FIV, mRNA vaccines encoding Env and p24 epitopes, combined with potent adjuvant systems, may overcome limitations of past vaccines by inducing broader, durable, and cross-clade immunity in cats. Additionally, viral vector–mediated mRNA vaccines can be optimized to enhance immunity in cats against FIV. Future research should focus on selecting optimal viral vectors (e.g., adenoviral, modified vaccinia Ankara, or lentiviral platforms) (213), assessing safety and long-term immunogenicity in cats, and evaluating efficacy against diverse FIV subtypes in both experimental and natural challenge models. If successful, viral vector–delivered mRNA vaccines may represent a new generation of FIV vaccines with the potential to overcome the limitations of earlier approaches.
Multi-epitope vaccines represent a promising strategy for overcoming the limitations of conventional FIV vaccines, which often provide incomplete or strain-specific protection. By combining conserved B-cell and T-cell epitopes from multiple viral proteins, such as Env, Gag, and p24, these vaccines can elicit broader and more durable immune responses across diverse FIV subtypes. Advances in bioinformatics and immunoinformatics now make it possible to predict and design epitope combinations with high immunogenic potential while minimizing off-target effects. Incorporating these epitopes into delivery systems, such as nanoparticles, viral vectors, or DNA/mRNA platforms, may further enhance immunogenicity and the longevity of protection. In addition, multi-epitope vaccines offer the flexibility to target both humoral and cellular arms of the immune system. By incorporating epitopes from multiple viral proteins, such vaccines can elicit broad and robust immune responses while addressing major challenges in FIV vaccine design, including antigenic shifts, antigenic drifts, and genetic variability among viral strains (214). This strategy enhances the likelihood of cross-protection against diverse FIV subtypes and reduces the risk of immune escape, which has historically hindered the success of conventional vaccines. Future work should focus on the experimental validation of in silico–designed epitope constructs, optimization of adjuvant formulations, and evaluation of cross-protection against circulating FIV strains in natural populations. If successful, multi-epitope vaccines may provide a next-generation solution with greater global applicability and efficacy in FIV prevention.
Additionally, comprehensive molecular epidemiology studies are required to characterize regional FIV variants, evaluate the cross-protection of vaccines, and assess the role of viral recombination in vaccine escape. Such studies will ensure that future vaccines and therapeutics are effective across a broad range of viral subtypes. A critical future goal in FIV vaccine research is the development of formulations capable of conferring protection against the predominant circulating subtypes, as well as the circulating recombinant forms (190). Given the high genetic diversity and recombination potential of FIV, next-generation vaccine strategies must focus on inducing broad-spectrum and durable immunity that remains effective across both subtype-specific and recombinant viral strains. To overcome this, future vaccine strategies should prioritize the inclusion of conserved epitopes from across circulating subtypes, identified through immunoinformatics and comparative genomics, to maximize cross-protection. In addition, multivalent formulations may help broaden immune responses against diverse strains. Multivalent vaccines offer an advantage over monovalent approaches by inducing polyclonal antibody responses against multiple FIV variants in a single formulation, thereby providing broader protection against recombinant and emerging subtypes (215).
Much of the current understanding of FIV comes from experimental infections in controlled laboratory settings. While these studies have provided valuable insights into viral pathogenesis and immune dysregulation, they may not fully capture the variability observed in naturally infected cats. In real-world conditions, prevalence and disease progression are influenced by multiple factors, including co-infections, environmental stressors, nutritional status, and host genetic background. As a result, naturally infected cats often display a broader range of clinical outcomes, including milder or atypical manifestations compared to laboratory models. For example, some FIV-positive cats remain asymptomatic for life, whereas others develop severe opportunistic infections or neoplasia.
Additionally, evidence from naturally infected cats also underscores the importance of considering real-world conditions when interpreting laboratory data. For example, outdoor FIV-positive cats have been reported to show more pronounced hypergammaglobulinemia and elevated total protein levels compared to indoor cats, likely reflecting greater antigenic exposure and co-infections in outdoor environments (31). Such findings illustrate how environmental and lifestyle factors shape immune responses and clinical manifestations in naturally infected cats, in contrast to the more uniform outcomes observed in controlled laboratory infections. Moreover, it remains difficult to directly compare treatment outcomes between experimentally infected cats maintained under laboratory conditions and pet cats naturally infected with diverse FIV field strains. This discrepancy underscores the importance of real-world evidence. While laboratory models provide controlled insight into mechanisms and drug activity, only studies in naturally infected cats can capture the influence of co-infections, environmental stressors, and viral diversity. Therefore, future progress will depend on well-designed, double-blinded, placebo-controlled clinical trials in naturally FIV-infected cats to rigorously determine the efficacy, safety, and long-term tolerability of novel antiviral compounds (32). These complexities underscore the need to interpret experimental data with caution and highlight the importance of integrating findings from naturally infected populations to obtain a more accurate picture of FIV epidemiology and clinical impact. Future research should therefore prioritize longitudinal studies in naturally infected cats, which will be critical for validating laboratory findings, refining vaccine efficacy, and guiding therapeutic interventions under real-world conditions.
Future research should also aim to overcome the geographical bias evident in current FIV studies, which are predominantly concentrated in developed countries such as Australia, the USA, and Japan. Expanding epidemiological surveys and vaccine evaluations in Africa, Southeast Asia, and Eastern Europe will be essential to generate more representative data on prevalence and vaccine efficacy. Addressing this gap will require investment in diagnostic infrastructure, capacity-building initiatives, and targeted funding to support local research. International collaborations and global data-sharing platforms can further strengthen surveillance and ensure that findings are globally applicable, ultimately improving strategies for FIV prevention and control.
14 Conclusion
FIV is prevalent among both stray and household domestic cats throughout the world. FIV infects a wide variety of cells and causes mild to severe clinical signs, whereas some cats may not show any signs at all. FIV infection has been associated with neoplastic diseases, neurological dysfunctions, and renal diseases. Environmental factors, host characteristics, and genetic variations may influence the clinical signs and pathogenesis of FIV. The variation in genomic sequences and the presence of different subtypes of FIV pose challenges in the development of an effective vaccine. Most vaccine candidates have shown poor efficacy, and although most challenge studies with a commercially available dual-subtype FIV vaccine have shown satisfactory efficacy against FIV infection in controlled settings, its effectiveness in real-world situations still needs to be established.
Similar to the development of FIV vaccine candidates, several factors have also hindered the development of antiviral drugs against this disease. FIV has shown resistance to some antiretroviral drugs used for the treatment of HIV, while other HIV antiviral drugs could be toxic and ineffective in cats. In vitro studies have shown the potential of derivatives of different compounds, peptides, and interferon as antivirals against FIV, but their efficacy has not been determined in FIV-infected cats in properly designed trials. Furthermore, it is uncertain if the findings of laboratory experiments on infected cats regarding FIV antiviral drugs and vaccines can be applied to pet cats infected with naturally occurring strains of the virus. Due to this uncertainty, it is crucial to conduct more carefully planned trials that are double-blind and placebo-controlled in the future. These trials should involve naturally infected cats with retroviruses, and different antiviral compounds should be studied to determine their effectiveness and any potential adverse effects. Moreover, in the future, the potential of multi-epitope vaccines in protecting cats from FIV infection can be explored. Furthermore, future research is required to identify the best adjuvants that can be used in FIV vaccines, including the immunoinformatic approach.
Additionally, bridging molecular insights with clinical management is essential for improving the care of FIV-positive cats. From a veterinary perspective, clinical decision-making should emphasize early detection, ongoing monitoring, and preventive management in multi-cat environments. Routine monitoring—including physical examinations, complete blood counts, serum biochemistry, and analysis of the CD4+/CD8+ T-cell ratio—can help track disease progression and guide timely interventions. Molecular markers such as proviral load and viral RNA quantification, although currently more common in research, may eventually serve as adjunct diagnostic tools in practice for risk stratification. In multi-cat households, segregating FIV-positive cats from aggressive or uninfected cats, neutering to reduce fighting behavior, and testing all cats before group housing remain best practices for minimizing transmission. By integrating molecular knowledge with structured monitoring and practical management guidelines, veterinarians can make informed decisions that balance long-term health outcomes with the quality of life in FIV-infected cats.
Author contributions
NA: Writing – original draft, Writing – review & editing. RM: Writing – review & editing. ST: Writing – review & editing. SR-M: Writing – review & editing. MN: Writing – review & editing. AC: Writing – review & editing. JL: Writing – review & editing. AS: Writing – review & editing. JR-S: Writing – original draft, Writing – review & editing. LC: Writing – original draft, Writing – review & editing. MC: Writing – original draft, Writing – review & editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This research was supported by the KAUST and Universidad Cooperativa de Colombia, Bucaramanga.
Acknowledgments
The authors thank KAUST, Universidad Cooperativa de Colombia in Bucaramanga, and STEMskills Research and Education Lab, a private limited company, for their support.
Conflict of interest
NA, ST, and AS were employed by STEMskills Research and Education Lab Private Limited.
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Gen AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher's note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Abbreviations
FIV, Feline Immunodeficiency Virus; AIDS, Acquired Immunodeficiency Syndrome; PCR, Polymerase Chain Reaction; ELISA, Enzyme-Linked Immunosorbent Assay; VIF, Viral Infectivity Factor; RNA, Ribonucleic Acid; DNA, Deoxyribonucleic Acid; MHC-I, Major Histocompatibility Complex Class I; MHC-II, Major Histocompatibility Complex Class II; ORF, Open Reading Frame.
References
1. Pedersen NC, Ho EW, Brown ML, Yamamoto JK. Isolation of a T-lymphotropic virus from domestic cats with an immunodeficiency-like syndrome. Science. (1987) 235:790–3. doi: 10.1126/science.3643650
2. McDonnel SJ, Sparger EE, Murphy BG. Feline immunodeficiency virus latency. Retrovirology. (2013) 10:69. doi: 10.1186/1742-4690-10-69
3. Tchamo CC, De Rugeriis M, Noormahomed EV. Occurrence of feline immunodeficiency virus and feline leukaemia virus in Maputo city and province, Mozambique: a pilot study. JFMS Open Rep. (2019) 5:2055116919870877. doi: 10.1177/2055116919870877
4. Carlton C, Norris JM, Hall E, Ward MP, Blank S, Gilmore S, et al. Clinicopathological and epidemiological findings in pet cats naturally infected with feline immunodeficiency virus (FIV) in Australia. Viruses. (2022) 14:2177. doi: 10.3390/v14102177
5. Gleich SE, Krieger S, Hartmann K. Prevalence of feline immunodeficiency virus and feline leukaemia virus among client-owned cats and risk factors for infection in Germany. J Feline Med Surg. (2009) 11:985–92. doi: 10.1016/j.jfms.2009.05.019
6. Sprißler F, Jongwattanapisan P, Luengyosluechakul S, Pusoonthornthum R, Reese S, Bergmann M, et al. Prevalence and risk factors of feline immunodeficiency virus and feline leukemia virus infection in healthy cats in Thailand. Front Vet Sci. (2022) 8:764217. doi: 10.3389/fvets.2021.764217
7. Bezerra JAB, Limeira CH, Maranhão ACP de M, Antunes JMA de P, de Azevedo SS. Global seroprevalence and factors associated with seropositivity for feline immunodeficiency virus (FIV) in cats: a systematic review and meta-analysis. Prev Vet Med. (2024) 231:106315. doi: 10.1016/j.prevetmed.2024.106315
8. Chhetri BK, Berke O, Pearl DL, Bienzle D. Comparison of the geographical distribution of feline immunodeficiency virus and feline leukemia virus infections in the United States of America (2000–2011). BMC Vet Res. (2013) 9:2. doi: 10.1186/1746-6148-9-2
9. Sacristán I, Acuña F, Aguilar E, García S, José López M, Cabello J, et al. Cross-species transmission of retroviruses among domestic and wild felids in human-occupied landscapes in Chile. Evol Appl. (2021) 14:1070–82. doi: 10.1111/eva.13181
10. Antunes A, Troyer JL, Roelke ME, Pecon-Slattery J, Packer C, Winterbach C, et al. The evolutionary dynamics of the lion panthera leo revealed by host and viral population genomics. PLoS Genet. (2008) 4:e1000251. doi: 10.1371/journal.pgen.1000251
11. Troyer JL, Pecon-Slattery J, Roelke ME, Johnson W, VandeWoude S, Vazquez-Salat N, et al. Seroprevalence and genomic divergence of circulating strains of feline immunodeficiency virus among felidae and hyaenidae species. J Virol. (2005) 79:8282–94. doi: 10.1128/JVI.79.13.8282-8294.2005
12. Brown MA, Munkhtsog B, Troyer JL, Ross S, Sellers R, Fine AE, et al. Feline immunodeficiency virus (FIV) in wild pallas' cats. Vet Immunol Immunopathol. (2010) 134:90. doi: 10.1016/j.vetimm.2009.10.014
13. Lagana DM, Lee JS, Lewis JS, Bevins SN, Carver S, Sweanor LL, et al. Characterization of regionally associated feline immunodeficiency virus (FIV) in bobcats (Lynx rufus). J Wildl Dis. (2013) 49:718–22. doi: 10.7589/2012-10-243
14. Hayama S, Yamamoto H, Nakanishi S, Hiyama T, Murayama A, Mori H, et al. Risk analysis of feline immunodeficiency virus infection in Tsushima leopard cats (Prionailurus bengalensis euptilurus) and domestic cats using a geographic information system. J Vet Med Sci. (2010) 72:1113–8. doi: 10.1292/jvms.09-0502
15. Barr MC, Calle PP, Roelke ME, Scott FW. Feline immunodeficiency virus infection in nondomestic felids. J Zoo Wildl Med. (1989) 20:265–72.
16. Westman ME, Malik R, Norris JM. Diagnosing feline immunodeficiency virus (FIV) and feline leukaemia virus (FeLV) infection: an update for clinicians. Aust Vet J. (2019) 97:47–55. doi: 10.1111/avj.12781
17. Eckstrand CD, Sparger EE, Murphy BG. Central and peripheral reservoirs of feline immunodeficiency virus in cats: a review. J Gen Virol. (2017) 98:1985–96. doi: 10.1099/jgv.0.000866
18. Taniwaki SA, Figueiredo AS, Jr JPA. Virus–host interaction in feline immunodeficiency virus (FIV) infection. Comp Immunol Microbiol Infect Dis. (2013) 36:549. doi: 10.1016/j.cimid.2013.07.001
19. Callanan JJ, Thompson H, Toth SR, O'Neil B, Lawrence CE, Willett B, et al. Clinical and pathological findings in feline immunodeficiency virus experimental infection. Vet Immunol Immunopathol. (1992) 35:3–13. doi: 10.1016/0165-2427(92)90116-8
20. Murphy BG, Castillo D, Cook S, Eckstrand C, Evans S, Sparger E, et al. The late asymptomatic and terminal immunodeficiency phases in experimentally FIV-infected cats—a long-term study. Viruses. (2023) 15:1775. doi: 10.3390/v15081775
21. de Mello LS, Ribeiro PR, de Almeida BA, Bandinelli MB, Sonne L, Driemeier D, et al. Diseases associated with feline leukemia virus and feline immunodeficiency virus infection: a retrospective study of 1470 necropsied cats (2010–2020). Comp Immunol Microbiol Infect Dis. (2023) 95:101963. doi: 10.1016/j.cimid.2023.101963
22. Hosie MJ, Addie D, Belák S, Boucraut-Baralon C, Egberink H, Frymus T, et al. Feline immunodeficiency. ABCD guidelines on prevention and management. J. Feline Med Surg. (2009) 11:575–84. doi: 10.1016/j.jfms.2009.05.006
23. Nishimura Y, Goto Y, Pang H, Endo Y, Mizuno T, Momoi Y, et al. Genetic heterogeneity of env gene of feline immunodeficiency virus obtained from multiple districts in Japan. Virus Res. (1998) 57:101–12. doi: 10.1016/S0168-1702(98)00085-9
24. Pedersen NC, Leutenegger CM, Woo J, Higgins J. Virulence differences between two field isolates of feline immunodeficiency virus (FIV-APetaluma and FIV-CPGammar) in young adult specific pathogen free cats. Vet Immunol Immunopathol. (2001) 79:53–67. doi: 10.1016/S0165-2427(01)00252-5
25. Olmsted RA, Hirsch VM, Purcell RH, Johnson PR. Nucleotide sequence analysis of feline immunodeficiency virus: genome organization and relationship to other lentiviruses. Proc Natl Acad Sci USA. (1989) 86:8088–92. doi: 10.1073/pnas.86.20.8088
26. González SA, Affranchino JL. Properties and functions of feline immunodeficiency virus gag domains in virion assembly and budding. Viruses. (2018) 10:261. doi: 10.3390/v10050261
27. Kenyon JC, Lever AML. The molecular biology of feline immunodeficiency virus (FIV). Viruses. (2011) 3:2192–213. doi: 10.3390/v3112192
28. Kemler I, Meehan A, Poeschla EM. Live-cell coimaging of the genomic RNAs and gag proteins of two lentiviruses. J Virol. (2010) 84:6352–66. doi: 10.1128/JVI.00363-10
29. Browning MT, Mustafa F, Schmidt RD, Lew KA, Rizvi TA. Delineation of sequences important for efficient packaging of feline immunodeficiency virus RNA. J Gen Virol. (2003) 84:621–7. doi: 10.1099/vir.0.18886-0
30. Kemler I, Azmi I, Poeschla EM. The critical role of proximal gag sequences in feline immunodeficiency virus genome encapsidation. Virology. (2004) 327:111–20. doi: 10.1016/j.virol.2004.06.014
31. Ovejero CA, González SA, Affranchino JL. The conserved Tyr176/Leu177 motif in the α-Helix 9 of the feline immunodeficiency virus capsid protein is critical for gag particle assembly. Viruses. (2019) 11:816. doi: 10.3390/v11090816
32. Abdusetir Cerfoglio JC, González SA, Affranchino JL. Structural elements in the Gag polyprotein of feline immunodeficiency virus involved in Gag self-association and assembly. J Gen Virol. (2014) 95:2050–9. doi: 10.1099/vir.0.065151-0
33. Manrique ML, Rauddi ML, González SA, Affranchino JL. Functional domains in the feline immunodeficiency virus nucleocapsid protein. Virology. (2004) 327:83–92. doi: 10.1016/j.virol.2004.06.019
34. Elder JH, Lerner DL, Hasselkus-Light CS, Fontenot DJ, Hunter E, Luciw PA, et al. Distinct subsets of retroviruses encode dUTPase. J Virol. (1992) 66:1791. doi: 10.1128/jvi.66.3.1791-1794.1992
35. North TW, Cronn RC, Remington KM, Tandberg RT, Judd RC. Characterization of reverse transcriptase from feline immunodeficiency virus. J Biol Chem. (1990) 265:5121–8. doi: 10.1016/S0021-9258(19)34093-1
36. Miller JT, Ehresmann B, Hübscher U, Le Grice SF, A. novel interaction of tRNA(Lys,3) with the feline immunodeficiency virus RNA genome governs initiation of minus strand DNA synthesis. J Biol Chem. (2001) 276:27721–30. doi: 10.1074/jbc.M100513200
37. González SA, Affranchino JL. The life cycle of feline immunodeficiency virus. Virology. (2025) 601:110304. doi: 10.1016/j.virol.2024.110304
38. Shibagaki Y, Holmes ML, Appa RS, Chow SA. Characterization of feline immunodeficiency virus integrase and analysis of functional domains. Virology. (1997) 230:1–10. doi: 10.1006/viro.1997.8466
39. Poeschla EM, Looney DJ. CXCR4 is required by a nonprimate lentivirus: heterologous expression of feline immunodeficiency virus in human, rodent, and feline cells. J Virol. (1998) 72:6858–66. doi: 10.1128/JVI.72.8.6858-6866.1998
40. de Parseval A, Chatterji U, Sun P, Elder JH. Feline immunodeficiency virus targets activated CD4+ T cells by using CD134 as a binding receptor. Proc Natl Acad Sci USA. (2004) 101:13044–9. doi: 10.1073/pnas.0404006101
41. Di Marino D, Bruno A, Grimaldi M, Scrima M, Stillitano I, Amodio G, et al. Binding of the anti-FIV peptide C8 to differently charged membrane models: from first docking to membrane tubulation. Front Chem. (2020) 8:493. doi: 10.3389/fchem.2020.00493
42. Morrison JH, Guevara RB, Marcano AC, Saenz DT, Fadel HJ, Rogstad DK, et al. Feline immunodeficiency virus envelope glycoproteins antagonize tetherin through a distinctive mechanism that requires virion incorporation. J Virol. (2014) 88:3255–72. doi: 10.1128/JVI.03814-13
43. Dietrich I, McMonagle EL, Petit SJ, Vijayakrishnan S, Logan N, Chan CN, et al. Feline tetherin efficiently restricts release of feline immunodeficiency virus but not spreading of infection. J Virol. (2011) 85:5840–52. doi: 10.1128/JVI.00071-11
44. Morrison JH, Poeschla EM. The feline immunodeficiency virus envelope signal peptide is a tetherin antagonizing protein. mBio 14:e00161-23. doi: 10.1128/mbio.00161-23
45. Hosie MJ, Pajek D, Samman A, Willett BJ. Feline immunodeficiency virus (FIV) neutralization: a review. Viruses. (2011) 3:1870–90. doi: 10.3390/v3101870
46. Gemeniano MC, Sawai ET, Sparger EE. Feline immunodeficiency virus Orf-A localizes to the nucleus and induces cell cycle arrest. Virology. (2004) 325:167–74. doi: 10.1016/j.virol.2004.05.007
47. Gemeniano MC, Sawai ET, Leutenegger CM, Sparger EE. feline immunodeficiency virus Orf-A is required for virus particle formation and virus infectivity. J Virol. (2003) 77:8819–30. doi: 10.1128/JVI.77.16.8819-8830.2003
48. Shen X, Leutenegger CM, Stefano Cole K, Pedersen NC, Sparger EE, A. feline immunodeficiency virus vif-deletion mutant remains attenuated upon infection of newborn kittens. J Gen Virol. (2007) 88:2793–9. doi: 10.1099/vir.0.83268-0
49. Zhang Z, Gu Q, Marino D, Lee K-L, Kong I-K, Häussinger D, et al. Feline APOBEC3s, barriers to cross-species transmission of FIV? Viruses. (2018) 10:186. doi: 10.3390/v10040186
50. Gu Q, Zhang Z, Gertzen CGW, Häussinger D, Gohlke H, Münk C. Identification of a conserved interface of human immunodeficiency virus type 1 and feline immunodeficiency virus Vifs with Cullin 5. J Virol. (2018) 92:e01697–17. doi: 10.1128/JVI.01697-17
51. Yoshikawa R, Takeuchi JS, Yamada E, Nakano Y, Misawa N, Kimura Y, et al. Feline immunodeficiency virus evolutionarily acquires two proteins, Vif and protease, capable of antagonizing feline APOBEC3. J Virol. (2017) 91:e00250–17. doi: 10.1128/JVI.00250-17
52. Marchand C, Lemay G, Archambault D. Identification of the nuclear and nucleolar localization signals of the Feline immunodeficiency virus Rev protein. Virus Res. (2020) 290:198153. doi: 10.1016/j.virusres.2020.198153
53. Weaver EA, A. Detailed Phylogenetic Analysis of FIV in the United States. PLoS ONE. (2010) 5:e12004. doi: 10.1371/journal.pone.0012004
54. Sodora DL, Shpaer EG, Kitchell BE, Dow SW, Hoover EA, Mullins JI. Identification of three feline immunodeficiency virus (FIV) env gene subtypes and comparison of the FIV and human immunodeficiency virus type 1 evolutionary patterns. J Virol. (1994) 68:2230–8. doi: 10.1128/jvi.68.4.2230-2238.1994
55. Zhang J, Wang L, Li J, Kelly P, Price S, Wang C. First Molecular characterization of feline immunodeficiency virus in domestic cats from Mainland China. PLoS ONE. (2017) 12:e0169739. doi: 10.1371/journal.pone.0169739
56. Hayward JJ, Taylor J, Rodrigo AG. Phylogenetic analysis of feline immunodeficiency virus in feral and companion domestic cats of New Zealand. J Virol. (2007) 81:2999–3004. doi: 10.1128/JVI.02090-06
57. Hayward JJ, Rodrigo AG. Molecular epidemiology of feline immunodeficiency virus in the domestic cat (Felis catus). Vet Immunol Immunopathol. (2010) 134:68–74. doi: 10.1016/j.vetimm.2009.10.011
58. Safwat MS, Bahr AD, Bakry NM, Amer HM, Yousif AA, Shehata AA, et al. Ancient and dominant: a novel feline immunodeficiency virus subtype “X-EGY” identified in Egyptian cats associated with high prevalence. BMC Vet Res. (2025) 21:497. doi: 10.1186/s12917-025-04943-1
59. Steinrigl A, Klein D. Phylogenetic analysis of feline immunodeficiency virus in Central Europe: a prerequisite for vaccination and molecular diagnostics. J Gen Virol. (2003) 84:1301–7. doi: 10.1099/vir.0.18736-0
60. Hohdatsu T, Motokawa K, Usami M, Amioka M, Okada S, Koyama H. Genetic subtyping and epidemiological study of feline immunodeficiency virus by nested polymerase chain reaction-restriction fragment length polymorphism analysis of the gag gene. J Virol Methods. (1998) 70:107–11. doi: 10.1016/S0166-0934(97)00167-5
61. Hayward JJ, Rodrigo AG. Recombination in feline immunodeficiency virus from feral and companion domestic cats. Virol J. (2008) 5:76. doi: 10.1186/1743-422X-5-76
62. Biondo D, Kipper D, Maciel JG, Santana W de O, Streck AF, Lunge VR. Phylogenetic classification of feline immunodeficiency virus. Acta Sci Vet. (2023) 51:129530. doi: 10.22456/1679-9216.129530
63. Pecon-Slattery J, Troyer JL, Johnson WE, O'Brien SJ. Evolution of feline immunodeficiency virus in Felidae: implications for human health and wildlife ecology. Vet Immunol Immunopathol. (2008) 123:32–44. doi: 10.1016/j.vetimm.2008.01.010
64. Beczkowski PM, Beatty JA. Feline immunodeficiency virus: current knowledge and future directions. Adv Small Anim Care. (2022) 3:145–59. doi: 10.1016/j.yasa.2022.05.007
65. Szilasi A, Dénes L, Krikó E, Heenemann K, Ertl R, Mándoki M, et al. Prevalence of feline immunodeficiency virus and feline leukaemia virus in domestic cats in Hungary. JFMS Open Rep. (2019) 5:2055116919892094. doi: 10.1177/2055116919892094
66. Cano-Ortiz L, Junqueira DM, Comerlato J, Costa CS, Zani A, Duda NB, et al. Phylodynamics of the Brazilian feline immunodeficiency virus. Infect Genet Evol. (2017) 55:166–71. doi: 10.1016/j.meegid.2017.09.011
67. Beczkowski PM, Hughes J, Biek R, Litster A, Willett BJ, Hosie MJ. Feline immunodeficiency virus (FIV) env recombinants are common in natural infections. Retrovirology. (2014) 11:80. doi: 10.1186/s12977-014-0080-1
68. Reggeti F, Bienzle D. Feline immunodeficiency virus subtypes A, B and C and intersubtype recombinants in Ontario, Canada. J Gen Virol. (2004) 85:1843–52. doi: 10.1099/vir.0.19743-0
69. Huguet M, Novo SG, Bratanich A. Detection of feline immunodeficiency virus subtypes A and B circulating in the city of Buenos Aires. Arch Virol. (2019) 164:2769–74. doi: 10.1007/s00705-019-04363-1
70. Taniwaki SA, Jiménez-Villegas T, Santana-Clavijo NF, Cruz TF, Silva SOS, Valencia-Bacca JD, et al. Near-complete genome sequence of feline immunodeficiency virus from Colombia. Microbiol Resour Announc. (2020) 9:e00754–20. doi: 10.1128/MRA.00754-20
71. Nakamura K, Suzuki Y, Ikeo K, Ikeda Y, Sato E, Nguyen NTP, et al. Phylogenetic analysis of Vietnamese isolates of feline immunodeficiency virus: genetic diversity of subtype C. Arch Virol. (2003) 148:783–91. doi: 10.1007/s00705-002-0954-8
72. Kakinuma S, Motokawa K, Hohdatsu T, Yamamoto JK, Koyama H, Hashimoto H. Nucleotide sequence of feline immunodeficiency virus: classification of Japanese isolates into two subtypes which are distinct from non-Japanese subtypes. J Virol. (1995) 69:3639–46. doi: 10.1128/jvi.69.6.3639-3646.1995
73. Nehring M, Dickmann EM, Billington K, VandeWoude S. Study of feline immunodeficiency virus prevalence and expert opinions on standards of care. J Feline Med Surg. (2024) 26:1098612X241245046. doi: 10.1177/1098612X241245046
74. Westman ME, Paul A, Malik R, McDonagh P, Ward MP, Hall E, et al. Seroprevalence of feline immunodeficiency virus and feline leukaemia virus in Australia: risk factors for infection and geographical influences (2011–2013). JFMS Open Rep. (2016) 2:2055116916646388. doi: 10.1177/2055116916646388
75. Szilasi A, Dénes L, Krikó E, Murray C, Mándoki M, Balka G. Prevalence of feline leukaemia virus and feline immunodeficiency virus in domestic cats in Ireland. Acta Vet Hung. (2021) 68:413–20. doi: 10.1556/004.2020.00056
76. Burling AN, Levy JK, Scott HM, Crandall MM, Tucker SJ, Wood EG, et al. Seroprevalences of feline leukemia virus and feline immunodeficiency virus infection in cats in the United States and Canada and risk factors for seropositivity. J Am Vet Med Assoc. (2017) 251:187–94. doi: 10.2460/javma.251.2.187
77. Azadian A, Hanifeh M, Firouzamandi M. The incidence of aggressive behavior in cats naturally infected with Feline Immunodeficiency Virus (FIV) and its interaction with FIV disease progression. Vet Ital. (2020) 56:169–76. doi: 10.12834/VetIt.1795.9466.3
78. Gates MC, Vigeant S, Dale A. Prevalence and risk factors for cats testing positive for feline immunodeficiency virus and feline leukaemia virus infection in cats entering an animal shelter in New Zealand. N Z Vet J. (2017) 65:285–91. doi: 10.1080/00480169.2017.1348266
79. O'Neil LL, Burkhard MJ, Diehl LJ, Hoover EA. Vertical transmission of feline immunodeficiency virus. Semin Vet Med Surg Small Anim. (1995) 10:266–78.
80. Sykes JE. Feline immunodeficiency virus infection. Canine Feline Infect Dis. (2014) 209–223. doi: 10.1016/B978-1-4377-0795-3.00021-1
81. O'Neil LL, Burkhard MJ, Hoover EA. Frequent perinatal transmission of feline immunodeficiency virus by chronically infected cats. J Virol. (1996) 70:2894–901. doi: 10.1128/jvi.70.5.2894-2901.1996
82. Allison RW, Hoover EA. Feline immunodeficiency virus is concentrated in milk early in lactation. AIDS Res Hum Retroviruses. (2003) 19:245–53. doi: 10.1089/088922203763315759
83. Jordan HL, Howard J, Barr MC, Kennedy-Stoskopf S, Levy JK, Tompkins WA. Feline immunodeficiency virus is shed in semen from experimentally and naturally infected cats. AIDS Res Hum Retroviruses. (1998) 14:1087–92. doi: 10.1089/aid.1998.14.1087
84. Litster AL. Transmission of feline immunodeficiency virus (FIV) among cohabiting cats in two cat rescue shelters. Vet J. (2014) 201:184–8. doi: 10.1016/j.tvjl.2014.02.030
85. Obert LA, Hoover EA. Feline immunodeficiency virus clade C mucosal transmission and disease courses. AIDS Res Hum Retroviruses. (2000) 16:677–88. doi: 10.1089/088922200308909
86. Galdo Novo S, Bucafusco D, Diaz LM, Bratanich AC. Viral diagnostic criteria for Feline immunodeficiency virus and Feline leukemia virus infections in domestic cats from Buenos Aires, Argentina. Rev Argentina Microbiol. (2016) 48:293–7. doi: 10.1016/j.ram.2016.07.003
87. Kokkinaki KG, Saridomichelakis MN, Leontides L, Mylonakis ME, Konstantinidis AO, Steiner JM, et al. prospective epidemiological, clinical, and clinicopathologic study of feline leukemia virus and feline immunodeficiency virus infection in 435 cats from Greece. Comp Immunol Microbiol Infect Dis. (2021) 78:101687. doi: 10.1016/j.cimid.2021.101687
88. Rungsuriyawiboon O, Jarudecha T, Hannongbua S, Choowongkomon K, Boonkaewwan C, Rattanasrisomporn J. Risk factors and clinical and laboratory findings associated with feline immunodeficiency virus and feline leukemia virus infections in Bangkok, Thailand. Vet World. (2022) 15:1601–9. doi: 10.14202/vetworld.2022.1601-1609
89. Natoli E, Say L, Cafazzo S, Bonanni R, Schmid M, Pontier D. Bold attitude makes male urban feral domestic cats more vulnerable to Feline Immunodeficiency Virus. Neurosci Biobehav Rev. (2005) 29:151–7. doi: 10.1016/j.neubiorev.2004.06.011
90. Ravi M, Wobeser GA, Taylor SM, Jackson ML. Naturally acquired feline immunodeficiency virus (FIV) infection in cats from western Canada: prevalence, disease associations, and survival analysis. Can Vet J. (2010) 51:271–6.
91. Liem BP, Dhand NK, Pepper AE, Barrs VR, Beatty JA. Clinical findings and survival in cats naturally infected with feline immunodeficiency virus. J Vet Intern Med. (2013) 27:798–805. doi: 10.1111/jvim.12120
92. Sivagurunathan A, Atwa AM, Lobetti R. Prevalence of feline immunodeficiency virus and feline leukaemia virus infection in Malaysia: a retrospective study. JFMS Open Rep. (2018) 4:2055116917752587. doi: 10.1177/2055116917752587
93. Luckman C, Gates MC. Epidemiology and clinical outcomes of feline immunodeficiency virus and feline leukaemia virus in client-owned cats in New Zealand. JFMS Open Rep. (2017) 3:2055116917729311. doi: 10.1177/2055116917729311
94. Tran V, Kelman M, Ward M, Westman M. Risk of feline immunodeficiency virus (FIV) infection in pet cats in Australia is higher in areas of lower socioeconomic status. Animals (Basel). (2019) 9:592. doi: 10.3390/ani9090592
95. Garigliany M, Jolly S, Dive M, Bayrou C, Berthemin S, Robin P, et al. Risk factors and effect of selective removal on retroviral infections prevalence in Belgian stray cats. Vet Rec. (2016) 178:45. doi: 10.1136/vr.103314
96. Dezubiria P, Amirian ES, Spera K, Crawford PC, Levy JK. Animal shelter management of feline leukemia virus and feline immunodeficiency virus infections in cats. Front Vet Sci. (2023) 9:1003388. doi: 10.3389/fvets.2022.1003388
97. Hellard E, Fouchet D, Rey B, Mouchet A, Poulet H, Pontier D. Differential association between circulating testosterone and infection risk by several viruses in natural cat populations: a behavioural-mediated effect? Parasitology. (2013) 140:521–9. doi: 10.1017/S0031182012001862
98. Bezerra JAB, Landim CP, Ribeiro YSR, Tertulino MD, Santos Junior R de F, Miranda Maranhão ACP de, et al. Epidemiological and clinicopathological findings of feline immunodeficiency virus and feline leukemia virus infections in domestic cats from the Brazilian semiarid region. Prev Vet Med. (2024) 226:106167. doi: 10.1016/j.prevetmed.2024.106167
99. Hartmann K. Clinical aspects of feline retroviruses: a review. Viruses. (2012) 4:2684–710. doi: 10.3390/v4112684
100. Moore A, Burrows AK, Malik R, Ghubash RM, Last RD, Remaj B. Fatal disseminated toxoplasmosis in a feline immunodeficiency virus-positive cat receiving oclacitinib for feline atopic skin syndrome. Vet Dermatol. (2022) 33:435–9. doi: 10.1111/vde.13097
101. Zambelli AB, Griffiths CA. South African report of first case of chromoblastomycosis caused by Cladosporium (syn Cladophialophora) carrionii infection in a cat with feline immunodeficiency virus and lymphosarcoma. J Feline Med Surg. (2015) 17:375–80. doi: 10.1177/1098612X14559954
102. Priolo V, Masucci M, Donato G, Solano-Gallego L, Martínez-Orellana P, Persichetti MF, et al. Association between feline immunodeficiency virus and Leishmania infantum infections in cats: a retrospective matched case-control study. Parasit Vect. (2022) 15:107. doi: 10.1186/s13071-022-05230-w
103. Elhamiani Khatat S, Rosenberg D, Benchekroun G, Polack B. Lungworm Eucoleus aerophilus (Capillaria aerophila) infection in a feline immunodeficiency virus-positive cat in France. JFMS Open Rep. (2016) 2:2055116916651649. doi: 10.1177/2055116916651649
104. Panait LC, Stock G, Globokar M, Balzer J, Groth B, Mihalca AD, et al. First report of Cytauxzoon sp. infection in Germany: organism description and molecular confirmation in a domestic cat. Parasitol Res. (2020) 119:3005–11. doi: 10.1007/s00436-020-06811-3
105. Shelton GH, Linenberger ML, Grant CK, Abkowitz JL. Hematologic manifestations of feline immunodeficiency virus infection. Blood. (1990) 76:1104–9. doi: 10.1182/blood.V76.6.1104.1104
106. Costa FVA da, Valle S de F, Machado G, Corbellini LG, Coelho EM, Rosa RB, et al. Hematological findings and factors associated with feline leukemia virus (FeLV) and feline immunodeficiency virus (FIV) positivity in cats from southern Brazil. Pesq Vet Bras. (2017) 37:1531–6. doi: 10.1590/s0100-736x2017001200028
107. Gleich S, Hartmann K. Hematology and serum biochemistry of feline immunodeficiency virus-infected and feline leukemia virus-infected cats. J Vet Intern Med. (2009) 23:552–8. doi: 10.1111/j.1939-1676.2009.0303.x
108. Power C. Neurologic disease in feline immunodeficiency virus infection: disease mechanisms and therapeutic interventions for NeuroAIDS. J Neurovirol. (2018) 24:220–8. doi: 10.1007/s13365-017-0593-1
109. Maingat F, Vivithanaporn P, Zhu Y, Taylor A, Baker G, Pearson K, et al. Neurobehavioral performance in feline immunodeficiency virus infection: integrated analysis of viral burden, neuroinflammation, and neuronal injury in cortex. J Neurosci. (2009) 29:8429–37. doi: 10.1523/JNEUROSCI.5818-08.2009
110. Azadian A, Gunn-Moore DA. Age-related cognitive impairments in domestic cats naturally infected with feline immunodeficiency virus. Vet Record. (2022) 191:e1683. doi: 10.1002/vetr.1683
111. Prospéro-García O, Herold N, Phillips TR, Elder JH, Bloom FE, Henriksen SJ. Sleep patterns are disturbed in cats infected with feline immunodeficiency virus. Proc Natl Acad Sci USA. (1994) 91:12947–51. doi: 10.1073/pnas.91.26.12947
112. Phillips TR, Prospero-Garcia O, Wheeler DW, Wagaman PC, Lerner DL, Fox HS, et al. Neurologic dysfunctions caused by a molecular clone of feline immunodeficiency virus, FIV-PPR. J Neurovirol. (1996) 2:388–96. doi: 10.3109/13550289609146904
113. Sathler MF, Doolittle MJ, Cockrell JA, Nadalin IR, Hofmann F, VandeWoude S, et al. HIV and FIV glycoproteins increase cellular tau pathology via cGMP-dependent kinase II activation. J Cell Sci. (2022) 135:jcs259764. doi: 10.1242/jcs.259764
114. Kaye S, Wang W, Miller C, McLuckie A, Beatty JA, Grant CK, et al. Role of feline immunodeficiency virus in lymphomagenesis–going alone or colluding? ILAR J. (2016) 57:24–33. doi: 10.1093/ilar/ilv047
115. Beatty JA, Lawrence CE, Callanan JJ, Grant CK, Gault EA, Neil JC, et al. Feline immunodeficiency virus (FIV)-associated lymphoma: a potential role for immune dysfunction in tumourigenesis. Vet Immunol Immunopathol. (1998) 65:309–22. doi: 10.1016/S0165-2427(98)00164-0
116. Callanan JJ, Jones BA, Irvine J, Willett BJ, McCandlish IA, Jarrett O. Histologic classification and immunophenotype of lymphosarcomas in cats with naturally and experimentally acquired feline immunodeficiency virus infections. Vet Pathol. (1996) 33:264–72. doi: 10.1177/030098589603300302
117. Hutson CA, Rideout BA, Pedersen NC. Neoplasia associated with feline immunodeficiency virus infection in cats of southern California. J Am Vet Med Assoc. (1991) 199:1357–62. doi: 10.2460/javma.1991.199.10.1357
118. Terry A, Callanan JJ, Fulton R, Jarrett O, Neil JC. Molecular analysis of tumours from feline immunodeficiency virus (FIV)-infected cats: an indirect role for FIV? Int J Cancer. (1995) 61:227–32. doi: 10.1002/ijc.2910610215
119. Shelton GH, Linenberger ML. Hematologic abnormalities associated with retroviral infections in the cat. Semin Vet Med Surg Small Anim. (1995) 10:220–33.
120. Poli A, Tozon N, Guidi G, Pistello M. Renal Alterations in feline immunodeficiency virus (FIV)-infected cats: a natural model of lentivirus-induced renal disease changes. Viruses. (2012) 4:1372–89. doi: 10.3390/v4091372
121. Biezus G, de Cristo TG, da Silva Schade MF, Ferian PE, Carniel F, Miletti LC, et al. Plasma cell pododermatitis associated with feline leukemia virus (felv) and concomitant feline immunodeficiency virus (FIV) infection in a cat. Top Companion Anim Med. (2020) 41:100475. doi: 10.1016/j.tcam.2020.100475
122. Rolim VM, Casagrande RA, Wouters ATB, Driemeier D, Pavarini SP. Myocarditis caused by feline immunodeficiency virus in five cats with hypertrophic cardiomyopathy. J Comp Pathol. (2016) 154:3–8. doi: 10.1016/j.jcpa.2015.10.180
123. Prisco F, Vaccaro E, Cardillo L, Fusco G, Papparella S, Santoro P, et al. Inflammatory myopathy and myocarditis are relevant complications of natural feline immunodeficiency virus infection. Vet Pathol. (2025) 2025:3009858251338849. doi: 10.1177/03009858251338849
124. Titmarsh HF, Lalor SM, Tasker S, Barker EN, Berry J, Gunn-More D, et al. Vitamin D status in cats with feline immunodeficiency virus. Vet Med Sci. (2015) 1:72–8. doi: 10.1002/vms3.11
125. Ghys LFE, Paepe D, Taffin ERL, Vandermeulen E, Duchateau L, Smets PMY, et al. Serum and urinary cystatin C in cats with feline immunodeficiency virus infection and cats with hyperthyroidism. J Feline Med Surg. (2016) 18:658–65. doi: 10.1177/1098612X15592343
126. Fonseca J, Silveira J, Faísca P, de Almeida PM. Presumptive hemophagocytic syndrome associated with co-infections with FIV, Toxoplasma gondii, and Candidatus mycoplasma haemominutum in an adult cat. Vet Clin Pathol. (2023) 52:324–33. doi: 10.1111/vcp.13205
127. Tasker S, Hofmann-Lehmann R, Belák S, Frymus T, Addie DD, Pennisi MG, et al. Haemoplasmosis in cats: European guidelines from the ABCD on prevention and management. J Feline Med Surg. (2018) 20:256–61. doi: 10.1177/1098612X18758594
128. Macieira DB, de Menezes R de CAA, Damico CB, Almosny NRP, McLane HL, Daggy JK, et al. Prevalence and risk factors for hemoplasmas in domestic cats naturally infected with feline immunodeficiency virus and/or feline leukemia virus in Rio de Janeiro — Brazil. J Feline Med Surg. (2008) 10:120–9. doi: 10.1016/j.jfms.2007.08.002
129. Gómez NV, Castillo VA, Gisbert MA, Pisano P, Mira G, Fontanals A, et al. Immune-endocrine interactions in treated and untreated cats naturally infected with FIV. Vet Immunol Immunopathol. (2011) 143:332–7. doi: 10.1016/j.vetimm.2011.06.012
130. Baxter K, Levy J, Edinboro C, Vaden S, Tompkins M. Renal disease in cats infected with feline immunodeficiency virus. J Vet Intern Med. (2012) 26:238–43. doi: 10.1111/j.1939-1676.2011.00871.x
131. Shelton GH, Grant CK, Cotter SM, Gardner MB, Hardy WD, DiGiacomo RF. Feline immunodeficiency virus and feline leukemia virus infections and their relationships to lymphoid malignancies in cats: a retrospective study (1968-1988). J Acquir Immune Defic Syndr.) (1990) 3:623–30.
132. Shimojima M, Miyazawa T, Ikeda Y, McMonagle EL, Haining H, Akashi H, et al. Use of CD134 as a primary receptor by the feline immunodeficiency virus. Science. (2004) 303:1192–5. doi: 10.1126/science.1092124
133. Dean GA, Reubel GH, Moore PF, Pedersen NC. Proviral burden and infection kinetics of feline immunodeficiency virus in lymphocyte subsets of blood and lymph node. J Virol. (1996) 70:5165–9. doi: 10.1128/jvi.70.8.5165-5169.1996
134. Mexas AM, Fogle JE, Tompkins WA, Tompkins MB. CD4+CD25+ regulatory t cells are infected and activated during acute fiv infection. Vet Immunol Immunopathol. (2008) 126:263–72. doi: 10.1016/j.vetimm.2008.08.003
135. Wang F-J, Cui D, Qian W-D. Therapeutic effect of CD4+CD25+ regulatory T cells amplified in vitro on experimental autoimmune neuritis in rats. CPB. (2018) 47:390–402. doi: 10.1159/000489919
136. Dow SW, Mathiason CK, Hoover EA. In vivo monocyte tropism of pathogenic feline immunodeficiency viruses. J Virol. (1999) 73:6852–61. doi: 10.1128/JVI.73.8.6852-6861.1999
137. Sprague WS, Robbiani M, Avery PR, O'Halloran KP, Hoover EA. Feline immunodeficiency virus dendritic cell infection and transfer. J Gen Virol. (2008) 89:709–15. doi: 10.1099/vir.0.83068-0
138. Yu N, Billaud JN, Phillips TR. Effects of feline immunodeficiency virus on astrocyte glutamate uptake: implications for lentivirus-induced central nervous system diseases. Proc Natl Acad Sci USA. (1998) 95:2624–9. doi: 10.1073/pnas.95.5.2624
139. Hein A, Martin JP, Koehren F, Bingen A, Dörries R. In vivo infection of ramified microglia from adult cat central nervous system by feline immunodeficiency virus. Virology. (2000) 268:420–9. doi: 10.1006/viro.1999.0152
140. Beebe AM, Gluckstern TG, George J, Pedersen NC, Dandekar S. Detection of feline immunodeficiency virus infection in bone marrow of cats. Vet Immunol Immunopathol. (1992) 35:37–49. doi: 10.1016/0165-2427(92)90119-B
141. Tanabe T, Yamamoto JK. Phenotypic and functional characteristics of FIV infection in the bone marrow stroma. Virology. (2001) 282:113–22. doi: 10.1006/viro.2000.0822
142. Park HS, Kyaw-Tanner M, Thomas J, Robinson WF. Feline immunodeficiency virus replicates in salivary gland ductular epithelium during the initial phase of infection. Vet Microbiol. (1995) 46:257–67. doi: 10.1016/0378-1135(95)00090-W
143. Eckstrand CD, Sparger EE, Pitt KA, Murphy BG. Peripheral and central immune cell reservoirs in tissues from asymptomatic cats chronically infected with feline immunodeficiency virus. PLoS ONE. (2017) 12:e0175327. doi: 10.1371/journal.pone.0175327
144. Eckstrand CD, Hillman C, Smith AL, Sparger EE, Murphy BG. Viral reservoirs in lymph nodes of FIV-infected progressor and long-term non-progressor cats during the asymptomatic phase. PLoS ONE. (2016) 11:e0146285. doi: 10.1371/journal.pone.0146285
145. Bach JM, Hurtrel M, Chakrabarti L, Ganiere JP, Montagnier L, Hurtrel B. Early stages of feline immunodeficiency virus infection in lymph nodes and spleen. AIDS Res Hum Retroviruses. (1994) 10:1731–8. doi: 10.1089/aid.1994.10.1731
146. Woo JC, Dean GA, Pedersen NC, Moore PF. Immunopathologic changes in the thymus during the acute stage of experimentally induced feline immunodeficiency virus infection in juvenile cats. J Virol. (1997) 71:8632–41. doi: 10.1128/jvi.71.11.8632-8641.1997
147. Hartmann K. Efficacy of antiviral chemotherapy for retrovirus-infected cats: what does the current literature tell us? J Feline Med Surg. (2015) 17:925–39. doi: 10.1177/1098612X15610676
148. Bashor L, Rawlinson JE, Kozakiewicz CP, Behzadi E, Miller C, Kim J, et al. Impacts of antiretroviral therapy on the oral microbiome and periodontal health of feline immunodeficiency virus-positive cats. Viruses. (2025) 17:257. doi: 10.3390/v17020257
149. Hartmann K, Wooding A, Bergmann M. Efficacy of antiviral drugs against feline immunodeficiency virus. Vet Sci. (2015) 2:456–76. doi: 10.3390/vetsci2040456
150. Medeiros S de O, Abreu CM, Delvecchio R, Ribeiro AP, Vasconcelos Z, Brindeiro R de M, et al. Follow-up on long-term antiretroviral therapy for cats infected with feline immunodeficiency virus. J Feline Med Surg. (2016) 18:264–72. doi: 10.1177/1098612X15580144
151. Fogle JE, Tompkins WA, Campbell B, Sumner D, Tompkins MB. Fozivudine tidoxil as single-agent therapy decreases plasma and cell-associated viremia during acute feline immunodeficiency virus infection. J Vet Intern Med. (2011) 25:413–8. doi: 10.1111/j.1939-1676.2011.0699.x
152. Zhu Y, Antony JM, Martinez JA, Glerum DM, Brussee V, Hoke A, et al. Didanosine causes sensory neuropathy in an HIV/AIDS animal model: impaired mitochondrial and neurotrophic factor gene expression. Brain. (2007) 130:2011–23. doi: 10.1093/brain/awm148
153. Lombardi S, Massi C, Indino E, La Rosa C, Mazzetti P, Falcone ML, et al. Inhibition of feline immunodeficiency virus infection in vitro by envelope glycoprotein synthetic peptides. Virology. (1996) 220:274–84. doi: 10.1006/viro.1996.0315
154. Giannecchini S, Di Fenza A, D'Ursi AM, Matteucci D, Rovero P, Bendinelli M. Antiviral activity and conformational features of an octapeptide derived from the membrane-proximal ectodomain of the feline immunodeficiency virus transmembrane glycoprotein. J Virol. (2003) 77:3724–33. doi: 10.1128/JVI.77.6.3724-3733.2003
155. Baba K, Goto-Koshino Y, Mizukoshi F, Setoguchi-Mukai A, Fujino Y, Ohno K, et al. Inhibition of the replication of feline immunodeficiency virus by lentiviral vector-mediated RNA interference in feline cell lines. J Vet Med Sci. (2008) 70:777–83. doi: 10.1292/jvms.70.777
156. Seetaha S, Ratanabunyong S, Tabtimmai L, Choowongkomon K, Rattanasrisomporn J, Choengpanya K. Anti-feline immunodeficiency virus reverse transcriptase properties of some medicinal and edible mushrooms. Vet World. (2020) 13:1798–806. doi: 10.14202/vetworld.2020.1798-1806
157. Kashiwase H, Katsube T, Kimura T, Nishigaki T, Yamashita M. 8-Difluoromethoxy-4-quinolone derivatives as anti-feline immunodeficiency virus (FIV) agents: important structural features for inhibitory activity of FIV replication. J Vet Med Sci. (2000) 62:499–504. doi: 10.1292/jvms.62.499
158. Asquith CRM, Meili T, Laitinen T, Baranovsky IV, Konstantinova LS, Poso A, et al. Synthesis and comparison of substituted 1,2,3-dithiazole and 1,2,3-thiaselenazole as inhibitors of the feline immunodeficiency virus (FIV) nucleocapsid protein as a model for HIV infection. Bioorg Med Chem Lett. (2019) 29:1765–8. doi: 10.1016/j.bmcl.2019.05.016
159. Gomez-Lucia E, Collado VM, Miró G, Martín S, Benítez L, Doménech A. Clinical and hematological follow-up of long-term oral therapy with type-I interferon in cats naturally infected with feline leukemia virus or feline immunodeficiency virus. Animals (Basel). (2020) 10:1464. doi: 10.3390/ani10091464
160. Leal RO, Gil S. The use of recombinant feline interferon omega therapy as an immune-modulator in cats naturally infected with feline immunodeficiency virus: new perspectives. Vet Sci. (2016) 3:32. doi: 10.3390/vetsci3040032
161. Gil S, Leal RO, McGahie D, Sepúlveda N, Duarte A, Niza MMRE, et al. Oral Recombinant Feline Interferon-Omega as an alternative immune modulation therapy in FIV positive cats: clinical and laboratory evaluation. Res Vet Sci. (2014) 96:79–85. doi: 10.1016/j.rvsc.2013.11.007
162. Gil S, Leal RO, Duarte A, McGahie D, Sepúlveda N, Siborro I, et al. Relevance of feline interferon omega for clinical improvement and reduction of concurrent viral excretion in retrovirus infected cats from a rescue shelter. Res Vet Sci. (2013) 94:753–63. doi: 10.1016/j.rvsc.2012.09.025
163. Hartmann K, Stengel C, Klein D, Egberink H, Balzarini J. Efficacy and adverse effects of the antiviral compound plerixafor in feline immunodeficiency virus-infected cats. J Vet Intern Med. (2012) 26:483–90. doi: 10.1111/j.1939-1676.2012.00904.x
164. Hartmann AD, Wilhelm N, Erfle V, Hartmann K. Clinical efficacy of melittin in the treatment of cats infected with the feline immunodeficiency virus. Tierarztl Prax Ausg K Kleintiere Heimtiere. (2016) 44:417–23. doi: 10.15654/TPK-150890
165. Squires RA, Crawford C, Marcondes M, Whitley N. 2024 guidelines for the vaccination of dogs and cats – compiled by the Vaccination Guidelines Group (VGG) of the World Small Animal Veterinary Association (WSAVA). J Small Animl Pract. (2024) 65:277–316. doi: 10.1111/jsap.13718
166. Dunham SP, Bruce J, MacKay S, Golder M, Jarrett O, Neil JC. Limited efficacy of an inactivated feline immunodeficiency virus vaccine. Vet Rec. (2006) 158:561–2. doi: 10.1136/vr.158.16.561
167. Pu R, Coleman J, Coisman J, Sato E, Tanabe T, Arai M, et al. Dual-subtype FIV vaccine (Fel-O-Vax® FIV) protection against a heterologous subtype B FIV isolate. J Feline Med Surg. (2005) 7:65–70. doi: 10.1016/j.jfms.2004.08.005
168. Westman ME, Malik R, Hall E, Harris M, Norris JM. The protective rate of the feline immunodeficiency virus vaccine: an Australian field study. Vaccine. (2016) 34:4752–8. doi: 10.1016/j.vaccine.2016.06.060
169. Beczkowski PM, Harris M, Techakriengkrai N, Beatty JA, Willett BJ, Hosie MJ. Neutralising antibody response in domestic cats immunised with a commercial feline immunodeficiency virus (FIV) vaccine. Vaccine. (2015) 33:977–84. doi: 10.1016/j.vaccine.2015.01.028
170. Richardson J, Moraillon A, Baud S, Cuisinier AM, Sonigo P, Pancino G. Enhancement of feline immunodeficiency virus (FIV) infection after DNA vaccination with the FIV envelope. J Virol. (1997) 71:9640–9. doi: 10.1128/jvi.71.12.9640-9649.1997
171. Hosie MJ, Osborne R, Reid G, Neil JC, Jarrett O. Enhancement after feline immunodeficiency virus vaccination. Vet Immunol Immunopathol. (1992) 35:191–7. doi: 10.1016/0165-2427(92)90131-9
172. Coleman JK, Pu R, Martin M, Sato E, Yamamoto JK. HIV-1 p24 vaccine protects cats against feline immunodeficiency virus infection. AIDS. (2005) 19:1457–66. doi: 10.1097/01.aids.0000183627.81922.be
173. Cuisinier AM, Mallet V, Meyer A, Caldora C, Aubert A, DNA. vaccination using expression vectors carrying FIV structural genes induces immune response against feline immunodeficiency virus. Vaccine. (1997) 15:1085–94. doi: 10.1016/S0264-410X(97)00004-2
174. Miller C, Emanuelli M, Fink E, Musselman E, Mackie R, Troyer R, et al. vaccine with receptor epitopes results in neutralizing antibodies but does not confer resistance to challenge. NPJ Vaccines. (2018) 3:16. doi: 10.1038/s41541-018-0051-y
175. Huisman W, Schrauwen EJA, Tijhaar E, Süzer Y, Pas SD, van Amerongen G, et al. Evaluation of vaccination strategies against infection with feline immunodeficiency virus (FIV) based on recombinant viral vectors expressing FIV Rev and OrfA. Vet Immunol Immunopathol. (2008) 126:332–8. doi: 10.1016/j.vetimm.2008.09.005
176. Osterhaus AD, Tijhaar E, Huisman RC, Huisman W, Darby IH, Francis MJ, et al. Accelerated viremia in cats vaccinated with recombinant vaccinia virus expressing envelope glycoprotein of feline immunodeficiency virus. AIDS Res Hum Retroviruses. (1996) 12:437–41. doi: 10.1089/aid.1996.12.437
177. Andrade LAF, Versiani AF, Barbosa-Stancioli EF, Dos Reis JKP, Dos Reis JGAC, da Fonseca FG. Developing a feline immunodeficiency virus subtype B Vaccine prototype using a recombinant MVA vector. Vaccines (Basel). (2022) 10:1717. doi: 10.3390/vaccines10101717
178. Huisman W, Schrauwen EJA, Pas SD, van Amerongen G, Rimmelzwaan GF, Osterhaus ADME. Evaluation of ISCOM-adjuvanted subunit vaccines containing recombinant feline immunodeficiency virus Rev, OrfA and envelope protein in cats. Vaccine. (2008) 26:2553–61. doi: 10.1016/j.vaccine.2008.03.023
179. Pistello M, Bonci F, Isola P, Mazzetti P, Merico A, Zaccaro L, et al. Evaluation of feline immunodeficiency virus ORF-A mutants as candidate attenuated vaccine. Virology. (2005) 332:676–90. doi: 10.1016/j.virol.2004.12.004
180. Freer G, Matteucci D, Mazzetti P, Tarabella F, Catalucci V, Ricci E, et al. Evaluation of feline monocyte-derived dendritic cells loaded with internally inactivated virus as a vaccine against feline immunodeficiency virus. Clin Vacc Immunol. (2008) 15:452–9. doi: 10.1128/CVI.00421-07
181. Gupta S, Leutenegger CM, Dean GA, Steckbeck JD, Cole KS, Sparger EE. Vaccination of cats with attenuated feline immunodeficiency virus proviral DNA vaccine expressing gamma interferon. J Virol. (2007) 81:465–73. doi: 10.1128/JVI.00815-06
182. Dunham SP, Flynn JN, Rigby MA, Macdonald J, Bruce J, Cannon C, et al. Protection against feline immunodeficiency virus using replication defective proviral DNA vaccines with feline interleukin-12 and−18. Vaccine. (2002) 20:1483–96. doi: 10.1016/S0264-410X(01)00507-2
183. Sahay B, Aranyos AM, Mishra M, McAvoy AC, Martin MM, Pu R, et al. Immunogenicity and efficacy of a novel multi-antigenic peptide vaccine based on cross-reactivity between feline and human immunodeficiency viruses. Viruses. (2019) 11:136. doi: 10.3390/v11020136
184. Matteucci D, Poli A, Mazzetti P, Sozzi S, Bonci F, Isola P, et al. Immunogenicity of an anti-clade B feline immunodeficiency fixed-cell virus vaccine in field cats. J Virol. (2000) 74:10911–9. doi: 10.1128/JVI.74.23.10911-10919.2000
185. Hohdatsu T, Okada S, Motokawa K, Aizawa C, Yamamoto JK, Koyama H. Effect of dual-subtype vaccine against feline immunodeficiency virus infection. Vet Microbiol. (1997) 58:155–65. doi: 10.1016/S0378-1135(97)00164-8
186. Kohmoto M, Miyazawa T, Sato E, Uetsuka K, Nishimura Y, Ikeda Y, et al. Cats are protected against feline immunodeficiency virus infection following vaccination with a homologous AP-1 binding site-deleted mutant. Arch Virol. (1998) 143:1839–45. doi: 10.1007/s007050050422
187. Broche-Pierre S, Richardson J, Moraillon A, Sonigo P. Evaluation of live feline immunodeficiency virus vaccines with modified antigenic properties. J Gen Virol. (2005) 86:2495–506. doi: 10.1099/vir.0.80469-0
188. Uhl EW, Heaton-Jones TG, Pu R, Yamamoto JK, FIV. vaccine development and its importance to veterinary and human medicine: a review: FIV vaccine 2002 update and review. Vet Immunol Immunopathol. (2002) 90:113–32. doi: 10.1016/S0165-2427(02)00227-1
189. Pu R, Coleman J, Omori M, Arai M, Hohdatsu T, Huang C, et al. Dual-subtype FIV vaccine protects cats against in vivo swarms of both homologous and heterologous subtype FIV isolates. AIDS. (2001) 15:1225. doi: 10.1097/00002030-200107060-00004
190. Coleman JK, Pu R, Martin MM, Noon-Song EN, Zwijnenberg R, Yamamoto JK. Feline immunodeficiency virus (FIV) vaccine efficacy and FIV neutralizing antibodies. Vaccine. (2014) 32:746–54. doi: 10.1016/j.vaccine.2013.05.024
191. Omori M, Pu R, Tanabe T, Hou W, Coleman JK, Arai M, et al. Cellular immune responses to feline immunodeficiency virus (FIV) induced by dual-subtype FIV vaccine. Vaccine. (2004) 23:386–98. doi: 10.1016/j.vaccine.2004.05.032
192. Boretti FS, Leutenegger CM, Mislin C, Hofmann-Lehmann R, König S, Schroff M, et al. Protection against FIV challenge infection by genetic vaccination using minimalistic DNA constructs for FIV env gene and feline IL-12 expression. AIDS. (2000) 14:1749. doi: 10.1097/00002030-200008180-00009
193. Hosie MJ, Dunsford T, Klein D, Willett BJ, Cannon C, Osborne R, et al. Vaccination with inactivated virus but not viral DNA reduces virus load following challenge with a heterologous and virulent isolate of feline immunodeficiency virus. J Virol. (2000) 74:9403–11. doi: 10.1128/JVI.74.20.9403-9411.2000
194. Leutenegger CM, Boretti FS, Mislin CN, Flynn JN, Schroff M, Habel A, et al. Immunization of cats against feline immunodeficiency virus (FIV) infection by using minimalistic immunogenic defined gene expression vector vaccines expressing FIV gp140 alone or with feline interleukin-12 (IL-12), IL-16, or a CpG motif. J Virol. (2000) 74:10447–57. doi: 10.1128/JVI.74.22.10447-10457.2000
195. Garçon Garçon N, Di Pasquale A. From discovery to licensure, the Adjuvant System story. Hum Vaccin Immunother. (2016) 13:19–33. doi: 10.1080/21645515.2016.1225635
196. Pulendran B. S. Arunachalam P, O'Hagan DT. Emerging concepts in the science of vaccine adjuvants. Nat Rev Drug Discov. (2021) 20:454–75. doi: 10.1038/s41573-021-00163-y
197. Facciolà A, Visalli G, Laganà A, Di Pietro A. An overview of vaccine adjuvants: current evidence and future perspectives. Vaccines (Basel). (2022) 10:819. doi: 10.3390/vaccines10050819
198. Oli AN, Obialor WO, Ifeanyichukwu MO, Odimegwu DC, Okoyeh JN, Emechebe GO, et al. Immunoinformatics and vaccine development: an overview. Immunotargets Ther. (2020) 9:13–30. doi: 10.2147/ITT.S241064
199. Rakib A, Sami SA, Islam MA, Ahmed S, Faiz FB, Khanam BH, et al. Epitope-based immunoinformatics approach on nucleocapsid protein of severe acute respiratory syndrome-coronavirus-2. Molecules. (2020) 25:5088. doi: 10.3390/molecules25215088
200. Rendon-Marin S, Ruíz-Saenz J. Universal peptide-based potential vaccine design against canine distemper virus (CDV) using a vaccinomic approach. Sci Rep. (2024) 14:16605. doi: 10.1038/s41598-024-67781-5
201. Jain P, Joshi A, Akhtar N, Krishnan S, Kaushik V. An immunoinformatics study: designing multivalent T-cell epitope vaccine against canine circovirus. J Genet Eng Biotechnol. (2021) 19:121. doi: 10.1186/s43141-021-00220-4
202. Paul B, Alam J, Hossain MMK, Hoque SF, Bappy MNI, Akter H, et al. Immunoinformatics for novel multi-epitope vaccine development in canine parvovirus infections. Biomedicines. (2023) 11:2180. doi: 10.3390/biomedicines11082180
203. Chawla M, Cuspoca AF, Akthar N, Magdaleno JSL, Rattanabunyong S, Suwattanasophon C, et al. Immunoinformatics-aided rational design of a multi-epitope vaccine targeting feline infectious peritonitis virus. Front Vet Sci. (2023) 10:1280273. doi: 10.3389/fvets.2023.1280273
204. Abdulla F, Adhikari UK, Uddin MK. Exploring T & B-cell epitopes and designing multi-epitope subunit vaccine targeting integration step of HIV-1 lifecycle using immunoinformatics approach. Microb Pathog. (2019) 137:103791. doi: 10.1016/j.micpath.2019.103791
205. Hashempour A, Khodadad N, Akbarinia S, Ghasabi F, Ghasemi Y, Nazar MMKA, et al. Reverse vaccinology approaches to design a potent multiepitope vaccine against the HIV whole genome: immunoinformatic, bioinformatics, and molecular dynamics approaches. BMC Infect Dis. (2024) 24:873. doi: 10.1186/s12879-024-09775-2
206. Habib A, Liang Y, Xu X, Zhu N, Xie J. Immunoinformatic identification of multiple epitopes of gp120 protein of HIV-1 to enhance the immune response against HIV-1 infection. Int J Mol Sci. (2024) 25:2432. doi: 10.3390/ijms25042432
207. Murphy BG, Wolf T, Vogel H, Castillo D, Woolard K. An RNA-directed gene editing strategy for attenuating the infectious potential of feline immunodeficiency virus-infected cells: a proof of concept. Viruses. (2020) 12:511. doi: 10.3390/v12050511
208. Hussein M, Molina MA, Berkhout B, Herrera-Carrillo E, A. CRISPR-Cas cure for HIV/AIDS. Int J Mol Sci. (2023) 24:1563. doi: 10.3390/ijms24021563
209. Xiao Q, Guo D, Chen S. Application of CRISPR/Cas9-based gene editing in HIV-1/AIDS therapy. Front Cell Infect Microbiol. (2019) 9:69. doi: 10.3389/fcimb.2019.00069
210. Uddin MB, Praseetha PK, Ahmed R, Shaheen MM, Kaouche FC, Bairagi RD, et al. Identification of natural compounds as potential antiviral drug candidates against Hepatitis E virus through molecular docking and dynamics simulations. J Indian Chem Soc. (2024) 101:101446. doi: 10.1016/j.jics.2024.101446
211. Polack FP, Thomas SJ, Kitchin N, Absalon J, Gurtman A, Lockhart S, et al. Safety and efficacy of the BNT162b2 mRNA COVID-19 vaccine. N Engl J Med. (2020) 383:2603–15. doi: 10.1056/NEJMoa2034577
212. Leong KY, Tham SK, Poh CL. Revolutionizing immunization: a comprehensive review of mRNA vaccine technology and applications. Virol J. (2025) 22:71. doi: 10.1186/s12985-025-02645-6
213. Tang J, Amin MA, Campian JL. Past, present, and future of viral vector vaccine platforms: a comprehensive review. Vaccines (Basel). (2025) 13:524. doi: 10.3390/vaccines13050524
214. Oyarzún P, Kobe B. Recombinant and epitope-based vaccines on the road to the market and implications for vaccine design and production. Hum Vaccin Immunother. (2015) 12:763–7. doi: 10.1080/21645515.2015.1094595
215. Kumari M, Liang K-H, Su S-C, Lin H-T, Lu Y-F, Wu M-J, et al. Multivalent mRNA vaccine elicits broad protection against SARS-CoV-2 variants of concern. Vaccines. (2024) 12:714. doi: 10.3390/vaccines12070714
216. Yahiaoui F, Kardjadj M, Ben-Mahdi MH. First seroprevalence study of feline leukemia and feline immunodeficiency infections among cats in Algiers (Algeria) and associated risk factors. Vet Sci. (2024) 11:546. doi: 10.3390/vetsci11110546
217. Teixeira BM, Taniwaki SA, Menezes PMM, Rodrigues AKPP, Mouta AN, Arcebispo TLM, et al. Feline immunodeficiency virus in Northern Ceará, Brazil. JFMS Open Rep. (2019) 5:2055116919859112. doi: 10.1177/2055116919859112
218. Lacerda LC, Silva AN, Freitas JS, Cruz RDS, Said RA, Munhoz AD. Feline immunodeficiency virus and feline leukemia virus: frequency and associated factors in cats in northeastern Brazil. Genet Mol Res. (2017) 16:gmr16029633. doi: 10.4238/gmr16029633
219. Feitosa TF, Costa FTR, Ferreira LC, Silva SS, Santos A, Silva WI, et al. High rate of feline immunodeficiency virus infection in cats in the Brazilian semiarid region: occurrence, associated factors and coinfection with Toxoplasma gondii and feline leukemia virus. Comp Immunol Microbiol Infect Dis. (2021) 79:101718. doi: 10.1016/j.cimid.2021.101718
220. Munro HJ, Berghuis L, Lang AS, Rogers L, Whitney H. Seroprevalence of feline immunodeficiency virus (FIV) and feline leukemia virus (FeLV) in shelter cats on the island of Newfoundland, Canada. Can J Vet Res. (2014) 78:140–4.
221. Liu C, Liu Y, Qian P, Cao Y, Wang J, Sun C, et al. Molecular and serological investigation of cat viral infectious diseases in China from 2016 to 2019. Transbound Emerg Dis. (2020) 67:2329–35. doi: 10.1111/tbed.13667
222. Knotek Z, Hájková P, Svoboda M, Toman M, Raska V. Epidemiology of feline leukaemia and feline immunodeficiency virus infections in the Czech Republic. Zentralbl Veterinarmed B. (1999) 46:665–71. doi: 10.1046/j.1439-0450.1999.00302.x
223. Nakamura Y, Nakamura Y, Ura A, Hirata M, Sakuma M, Sakata Y, et al. An updated nation-wide epidemiological survey of feline immunodeficiency virus (FIV) infection in Japan. J Vet Med Sci. (2010) 72:1051–6. doi: 10.1292/jvms.09-0574
224. Khalife S, Kassaa IA. Occurrence and risk factors of feline immunodeficiency virus (FIV) and feline leukaemia virus (FeLV) in cats of Lebanon. Comp Immunol Microbiol Infect Dis. (2023) 93:101931. doi: 10.1016/j.cimid.2022.101931
225. Bande F, Arshad SS, Hassan L, Zakaria Z, Sapian NA, Rahman NA, et al. Prevalence and risk factors of feline leukaemia virus and feline immunodeficiency virus in peninsular Malaysia. BMC Vet Res. (2012) 8:33. doi: 10.1186/1746-6148-8-33
226. Franzo G, de Villiers L, Coetzee LM, Villiers M de, Molini U. Molecular survey of feline immunodeficiency virus (FIV) infection in Namibian cats. Acta Trop. (2024) 253:107184. doi: 10.1016/j.actatropica.2024.107184
227. de Villiers L, Penzhorn BL, Schoeman JP, Molini U, de Villiers M, Byaruhanga C, et al. A multi-modal investigation into the prevalence and diagnostic evaluation of vector-borne pathogens and retroviruses in domestic cats throughout Namibia. Acta Trop. (2025) 268:107738. doi: 10.1016/j.actatropica.2025.107738
228. Rypuła K, Płoneczka-Janeczko K, Bierowiec K, Kumala A, Sapikowski G. Prevalence of viral infections in cats in southwestern Poland in the years 2006 to 2010. Berl Munch Tierarztl Wochenschr. (2014) 127:163–5.
229. Duarte A, Castro I, Pereira da Fonseca IM, Almeida V, Madeira de Carvalho LM, Meireles J, et al. Survey of infectious and parasitic diseases in stray cats at the Lisbon Metropolitan Area, Portugal. J Feline Med Surg. (2010) 12:441–6. doi: 10.1016/j.jfms.2009.11.003
230. Demkin VV, Kazakov AA. Prevalence of hemotropic mycoplasmas and coinfection with feline leukemia virus and feline immunodeficiency virus in cats in the Moscow region, Russia. Prev Vet Med. (2021) 190:105339. doi: 10.1016/j.prevetmed.2021.105339
231. Sarvani E, Tasker S, Kovac?ević Filipović M, Francuski Andrić J, Andrić N, Aquino L, et al. Prevalence and risk factor analysis for feline haemoplasmas in cats from Northern Serbia, with molecular subtyping of feline immunodeficiency virus. JFMS Open Rep. (2018) 4:2055116918770037. doi: 10.1177/2055116918770037
232. Villanueva-Saz S, Martínez M, Giner J, González A, Tobajas AP, Pérez MD, et al. A cross-sectional serosurvey of SARS-CoV-2 and co-infections in stray cats from the second wave to the sixth wave of COVID-19 outbreaks in Spain. Vet Res Commun. (2023) 47:615–29. doi: 10.1007/s11259-022-10016-7
233. Sukhumavasi W, Bellosa ML, Lucio-Forster A, Liotta JL, Lee ACY, Pornmingmas P, et al. Serological survey of Toxoplasma gondii, Dirofilaria immitis, feline immunodeficiency virus (FIV) and feline leukemia virus (FeLV) infections in pet cats in Bangkok and vicinities, Thailand. Vet Parasitol. (2012) 188:25–30. doi: 10.1016/j.vetpar.2012.02.021
234. So-In C, Watayotha L, Sonsupee T, Khankhum S, Sunthamala N. Molecular detection of vector-borne pathogens and their association with feline immunodeficiency virus and feline leukemia virus in cats from Northeastern Thailand. Animals (Basel). (2025) 15:2065. doi: 10.3390/ani15142065
235. Koç BT, Oguzoglu TÇ. A phylogenetic study of feline immunodeficiency virus (FIV) among domestic cats in Turkey. Comp Immunol Microbiol Infect Dis. (2020) 73:101544. doi: 10.1016/j.cimid.2020.101544
236. Muz D, Can H, Karakavuk M, Döşkaya M, Özdemir HG, Degirmenci Döşkaya A, et al. The molecular and serological investigation of Feline immunodeficiency virus and Feline leukemia virus in stray cats of Western Turkey. Comp Immunol Microbiol Infect Dis. (2021) 78:101688. doi: 10.1016/j.cimid.2021.101688
237. Stavisky J, Dean RS, Molloy MH. Prevalence of and risk factors for FIV and FeLV infection in two shelters in the United Kingdom (2011-2012). Vet Rec. (2017) 181:451. doi: 10.1136/vr.103857
238. Miyazawa T, Ikeda Y, Maeda K, Horimoto T, Tohya Y, Mochizuki M, et al. Seroepidemiological survey of feline retrovirus infections in domestic and leopard cats in northern Vietnam in 1997. J Vet Med Sci. (1998) 60:1273–5. doi: 10.1292/jvms.60.1273
239. Edinboro CH. Feral cat populations and feline retrovirus prevalence in san mateo county, california in three time periods between 2001 and 2016. Animals (Basel). (2022) 12:3477. doi: 10.3390/ani12243477
240. Chi X, Fang K, Koster L, Christie J, Yao C. Prevalence of feline immunodeficiency virus and Toxoplasma gondii in feral cats on St. Kitts, West Indies. Vet Sci. (2021) 8:16. doi: 10.3390/vetsci8020016
241. Massi C, Indino E, Lami C, Fissi A, Pieroni O, La Rosa C, et al. The antiviral activity of a synthetic peptide derived from the envelope SU glycoprotein of feline immunodeficiency virus maps in correspondence of an amphipathic helical segment. Biochem Biophys Res Commun. (1998) 246:160–5. doi: 10.1006/bbrc.1998.8580
242. Mizukoshi F, Baba K, Goto Y, Setoguchi A, Fujino Y, Ohno K, et al. Antiviral activity of membrane fusion inhibitors that target gp40 of the feline immunodeficiency virus envelope protein. Vet Microbiol. (2009) 136:155–9. doi: 10.1016/j.vetmic.2008.10.009
243. Grimaldi M, Stillitano I, Amodio G, Santoro A, Buonocore M, Moltedo O, et al. Structural basis of antiviral activity of peptides from MPER of FIV gp36. PLoS One. (2018) 13:e0204042. doi: 10.1371/journal.pone.0204042
244. Baba K, Mizukoshi F, Goto-Koshino Y, Setoguchi-Mukai A, Fujino Y, Ohno K, et al. Application of RNA interference for inhibiting the replication of feline immunodeficiency virus in chronically infected cell lines. Vet Microbiol. (2007) 120:207–16. doi: 10.1016/j.vetmic.2006.10.033
245. Pedretti E, Passeri B, Amadori M, Isola P, Di Pede P, Telera A, et al. Low-dose interferon-alpha treatment for feline immunodeficiency virus infection. Vet Immunol Immunopathol. (2006) 109:245–54. doi: 10.1016/j.vetimm.2005.08.020
246. Laitinen T, Meili T, Koyioni M, Koutentis PA, Poso A, Hofmann-Lehmann R, et al. Synthesis and evaluation of 1,2,3-dithiazole inhibitors of the nucleocapsid protein of feline immunodeficiency virus (FIV) as a model for HIV infection. Bioorg Med Chem. (2022) 68:116834. doi: 10.1016/j.bmc.2022.116834
247. Asquith CRM, Konstantinova LS, Laitinen T, Meli ML, Poso A, Rakitin OA, et al. Evaluation of substituted 1,2,3-dithiazoles as inhibitors of the feline immunodeficiency virus (FIV) nucleocapsid protein via a proposed zinc ejection mechanism. ChemMedChem. (2016) 11:2119–26. doi: 10.1002/cmdc.201600260
248. Asquith CRM, Sil BC, Laitinen T, Tizzard GJ, Coles SJ, Poso A, et al. Novel epidithiodiketopiperazines as anti-viral zinc ejectors of the Feline Immunodeficiency Virus (FIV) nucleocapsid protein as a model for HIV infection. Bioorg Med Chem. (2019) 27:4174–84. doi: 10.1016/j.bmc.2019.07.047
249. Mizukoshi F, Baba K, Goto-Koshino Y, Setoguchi-Mukai A, Fujino Y, Ohno K, et al. Inhibitory effect of newly developed CXC-chemokine receptor 4 antagonists on the infection with feline immunodeficiency virus. J Vet Med Sci. (2009) 71:121–4. doi: 10.1292/jvms.71.121
250. Schwartz AM, McCrackin MA, Schinazi RF, Hill PB, Vahlenkamp TW, Tompkins MB, et al. Antiviral efficacy of nine nucleoside reverse transcriptase inhibitors against feline immunodeficiency virus in feline peripheral blood mononuclear cells. Am J Vet Res. (2014) 75:273–81. doi: 10.2460/ajvr.75.3.273
251. Asquith CRM, Laitinen T, Konstantinova LS, Tizzard G, Poso A, Rakitin OA, et al. Investigation of the pentathiepin functionality as an inhibitor of feline immunodeficiency virus (FIV) via a potential zinc ejection mechanism, as a model for HIV infection. ChemMedChem. (2019) 14:454–61. doi: 10.1002/cmdc.201800718
252. Savarino A, Pistello M, D'Ostilio D, Zabogli E, Taglia F, Mancini F, et al. Human immunodeficiency virus integrase inhibitors efficiently suppress feline immunodeficiency virus replication in vitro and provide a rationale to redesign antiretroviral treatment for feline AIDS. Retrovirology. (2007) 4:79. doi: 10.1186/1742-4690-4-79
253. Huitron-Resendiz S, De Rozières S, Sanchez-Alavez M, Bühler B, Lin Y-C, Lerner DL, et al. Resolution and prevention of feline immunodeficiency virus-induced neurological deficits by treatment with the protease inhibitor TL-3. J Virol. (2004) 78:4525–32. doi: 10.1128/JVI.78.9.4525-4532.2004
254. Taffin E, Paepe D, Goris N, Auwerx J, Debille M, Neyts J, et al. Antiviral treatment of feline immunodeficiency virus-infected cats with (R)-9-(2-phosphonylmethoxypropyl)-2,6-diaminopurine. J Feline Med Surg. (2015) 17:79–86. doi: 10.1177/1098612X14532089
255. Tarcsai KR, Hidvégi M, Corolciuc O, Nagy K, Abbas AA, Ablashi DV, et al. The effects of Avemar treatment on feline immunodeficiency virus infected cell cultures. Vet Med Sci. (2023) 9:1446–55. doi: 10.1002/vms3.1141
256. Westman M, Yang D, Green J, Norris J, Malik R, Parr YA, et al. Antibody responses in cats following primary and annual vaccination against feline immunodeficiency virus (FIV) with an inactivated whole-virus vaccine (Fel-O-Vax® FIV). Viruses. (2021) 13:470. doi: 10.3390/v13030470
257. Stickney A, Ghosh S, Cave NJ, Dunowska M. Lack of protection against feline immunodeficiency virus infection among domestic cats in New Zealand vaccinated with the Fel-O-Vax® FIV vaccine. Vet Microbiol. (2020) 250:108865. doi: 10.1016/j.vetmic.2020.108865
258. Huang C, Conlee D, Gill M, Chu H-JS. Dual-subtype feline immunodeficiency virus vaccine provides 12 months of protective immunity against heterologous challenge. J Feline Med Surg. (2010) 12:451–7. doi: 10.1016/j.jfms.2009.12.016
259. Tellier MC, Pu R, Pollock D, Vitsky A, Tartaglia J, Paoletti E, et al. Efficacy evaluation of prime-boost protocol: canarypoxvirus-based feline immunodeficiency virus (FIV) vaccine and inactivated FIV-infected cell vaccine against heterologous FIV challenge in cats. AIDS. (1998) 12:11–8. doi: 10.1097/00002030-199801000-00002
260. Stevens R, Howard KE, Nordone S, Burkhard M, Dean GA. Oral immunization with recombinant listeria monocytogenes controls virus load after vaginal challenge with feline immunodeficiency virus. J Virol. (2004) 78:8210–8. doi: 10.1128/JVI.78.15.8210-8218.2004
261. Maksaereekul S, Dubie RA, Shen X, Kieu H, Dean GA, Sparger EE. Vaccination with vif-deleted feline immunodeficiency virus provirus, GM-CSF, and TNF-alpha plasmids preserves global CD4 T lymphocyte function after challenge with FIV. Vaccine. (2009) 27:3754–65. doi: 10.1016/j.vaccine.2009.03.081
Keywords: feline immunodeficiency virus, vaccine, prevalence, antiviral, retrovirus, symptoms
Citation: Akhtar N, Mishra R, Tripathi S, Redon-Marin S, Narsing Rao MP, Cuspoca Orduz AF, Leon Magdaleno JS, Shaikh AR, Ruiz-Saenz J, Cavallo L and Chawla M (2025) Feline immunodeficiency virus: current insights into pathogenesis, clinical impact, and advances in treatment and vaccine development. Front. Vet. Sci. 12:1665999. doi: 10.3389/fvets.2025.1665999
Received: 14 July 2025; Accepted: 05 September 2025;
Published: 25 September 2025.
Edited by:
Francisco José Pallarés, University of Cordoba, SpainReviewed by:
Aoxing Tang, Chinese Academy of Agricultural Sciences (CAAS), ChinaSelim Ahmed, Patuakhali Science and Technology University, Bangladesh
Copyright © 2025 Akhtar, Mishra, Tripathi, Redon-Marin, Narsing Rao, Cuspoca Orduz, Leon Magdaleno, Shaikh, Ruiz-Saenz, Cavallo and Chawla. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Nahid Akhtar, bmFoeWQuYWtodGFyMjRAZ21haWwuY29t; Mohit Chawla, bW9oaXQuY2hhd2xhQGthdXN0LmVkdS5zYQ==; bW9oaXRjaGF3bGEuYnRAZ21haWwuY29t; Julian Ruiz-Saenz, anVsaWFuLnJ1aXpzQGNhbXB1c3VjYy5lZHUuY28=