 Michal Benovics1,2
Michal Benovics1,2 Lívia Švantnerová1Ondřej Mikulka3,4
Lívia Švantnerová1Ondřej Mikulka3,4 Lucie Škorpíková5
Lucie Škorpíková5 Vlastimil Skoták3,4
Vlastimil Skoták3,4 Jan Cukor4,6*
Jan Cukor4,6*- 1Department of Zoology, Faculty of Natural Sciences, Comenius University in Bratislava, Bratislava, Slovakia
- 2Unit for Environmental Sciences and Management, North-West University, Potchefstroom, South Africa
- 3Faculty of Forestry and Wood Technology, Mendel University in Brno, Brno, Czechia
- 4Forestry and Game Management Research Institute, Jíloviště, Czechia
- 5Department of Botany and Zoology, Faculty of Science, Masaryk University, Brno, Czechia
- 6Faculty of Forestry and Wood Sciences, Czech University of Life Sciences Prague, Prague, Czechia
Baylisascaris procyonis, a zoonotic nematode originated in North America, is a significant cause of larva migrans in humans and wildlife. Here we report the first confirmed record of B. procyonis in free-ranging invasive Northern raccoons (Procyon lotor) in the Czech Republic, based on combined morphological and molecular evidence. 10 raccoons were examined from the Doupov Highlands subpopulation between 2023 and 2025, B. procyonis were recovered from three individuals. Morphological and molecular analyses of ITS and COI regions confirmed the species identity, showing 99.3%−100% similarity to known conspecific sequences. This finding extends the recognized range of this zoonotic parasite in Central Europe, underscoring the potential public health risks posed by environmental contamination with infective eggs. Our results highlight the need for ongoing surveillance of invasive raccoon populations to monitor the spread of this pathogenic zoonotic parasites.
Introduction
Biological invasions pose a global challenge for biodiversity conservation, causing a wide range of negative impacts on native ecosystems and their components (1). The introduction of invasive species is typically driven by multiple factors, including propagule pressure associated with international trade and the ecological condition of the new environment (2). One of the most thoroughly researched and consistently confirmed impacts of biological invasions is the disturbance of the environment for native, particularly protected species, resulting in the destabilization of specific habitats. In addition to ecological effects, invasive species may pose direct risks to human health.
This is best exemplified by the Northern raccoon (Procyon lotor), whose native range is in North America, but whose population has been rapidly increasing in Central Europe in recent decades. The European raccoon population is currently considered to be beyond effective control, with Germany as the core distribution area. At the same time the species rapidly expands into other countries such as Poland, Austria, France, and the Czech Republic (3). Along with raccoons, parasites associated with this species—and for which it serves as a suitable host—could also be introduced into Europe. One such common parasite in the native distribution range is Baylisascaris procyonis, whose occurrence has been relatively well documented in North America (4–6) and its presence and potential spread in Europe is apparent from the recent reports from Germany (7–9), Poland (10), Denmark (11), Norway (12), Austria (13), and Italy (14).
Baylisascaris procyonis is an intestinal nematode of raccoons, causing severe or fatal larva migrans syndrome. Over 150 species of birds and mammals (4) are known to be susceptible to infection, including humans (4, 5). Humans serve as paratenic hosts and infection occurs through ingestion of embryonated eggs from contaminated environments, often in urban or peri-urban areas (15, 16). The larvae are neurotropic and can migrate to the central nervous system, leading to eosinophilic meningoencephalitis, especially in young children or immunocompromised individuals (17, 18). It is also recognized as a cause of ocular disease in humans. In contrast to neural larva migrans (NLM), which is primarily restricted to infants and young children, isolated ocular larva migrans (OLM) usually occurs in otherwise healthy adults (19, 20). Due to its environmental persistence and high pathogenicity, B. procyonis is considered an important emerging zoonosis. To date, more than 50 human cases of infection have been confirmed worldwide (21), most of them from the USA, where at least 35 cases have been reported (22). Data from Europe are very scarce. Only two non-fatal clinical cases have been described in the literature (23). In addition to these clinical cases, four individuals with serologically confirmed exposure were reported in Germany (24). Our research therefore aimed to investigate the parasite fauna of raccoons in Czechia, with a special focus on a zoonotic B. procyonis.
Materials and methods
Material collection and identification
Ten raccoons were collected between 2023 and 2025 from the Doupov Highlands (Doupovské Hory, Figure 1) subpopulation, an area characterized by mixed forests, semi-natural broadleaved stands, and open habitats within a military training area. The animals were collected through hunting, a standard wildlife management practice. According to Decree No. 454/2021 Coll. on the designation of invasive species requiring regulation, raccoons are hunted by any hunter holding a hunting license issued by the manager of the respective hunting ground. They were placed in plastic bags, then labeled with information on the sampling location, sex, and body weight. The samples were transported to the laboratory, where the animals were dissected and the intestines and remaining body parts were frozen separately. Before parasitological examination, the intestines were gradually thawed and longitudinally incised. Macroscopic observation revealed the presence of white-yellowish ascarid nematodes in the small intestine of three animals. These were removed, washed and stored in 70% ethanol for subsequent morphological and molecular evaluation. The intestines were examined under a stereomicroscope, and the contents were cleaned using the sieve system [following (25)]. Any additional helminths identified were also preserved in either 70 or 96% ethanol for future studies focusing on the parasite diversity of raccoons in Czechia. The identification of the collected ascarid nematodes followed the guidelines provided by (17, 26, 27). The specimens were then cut into three parts, with the middle section designated for subsequent molecular analyses.
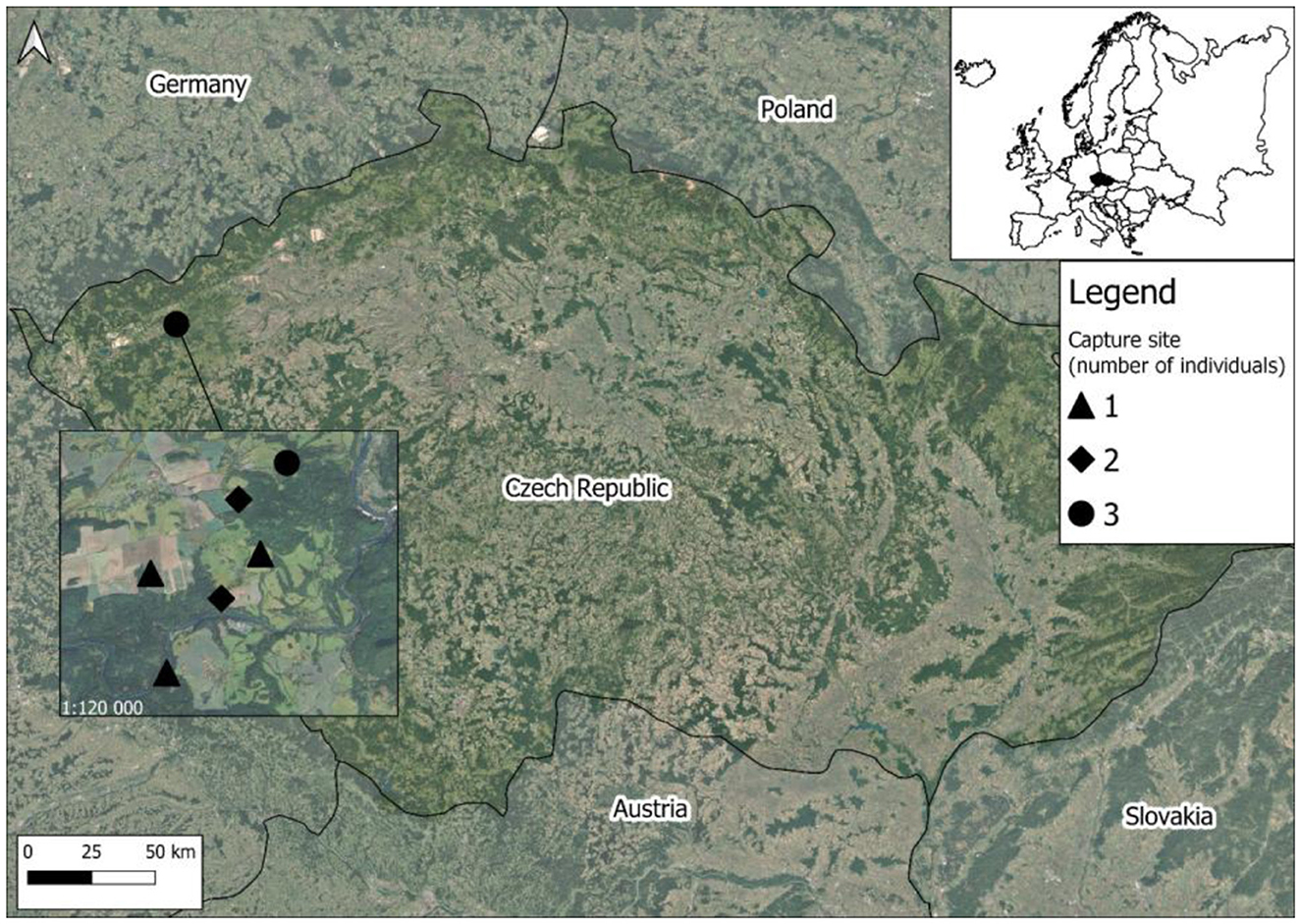
Figure 1. Location of the Doupov Highlands and respective sites where infected raccoons were collected.
Molecular and phylogenetic analyses
Genomic DNA was extracted from cut sections of six nematodes using the DNeasy Blood & Tissue Kit (Qiagen, Hilden, Germany) following the manufacturer's protocol. Two genomic regions were amplified: the ITS1-5.8S-ITS2 ribosomal regions using primers 18SNemF and 26SNemR (28) and the partial mitochondrial COI gene with primers CO1-F and CO1-R (29). The PCR reactions were performed in a 20 μl reaction mixture containing 14 μl nuclease-free water, 4 μl FIREPol Master Mix Ready to Load (Solis BioDyne, Tartu, Estonia), 0.5 μM of each primer, and 1 μl of DNA template. PCR products were detected by electrophoresis in 1% agarose gels stained with GoodView (SBS Genetech, Bratislava, Slovakia). The thermocycler conditions for amplification were retrieved from (30), and resulting amplicons were subjected to Sanger sequencing in Macrogen Europe (Amsterdam, Netherlands) using the PCR primers.
To confirm the species of the collected ascarid nematodes and their phylogenetic relationships to other Ascarididae, additional orthologous COI sequences from congeners or other phylogenetically close species were retrieved from GenBank (accession numbers are included within phylogenetic trees). Three Toxocara spp. ortholog sequences were used as an outgroup for rooting the phylogenetic tree according to (30). The sequences were aligned with the fast Fourier transform algorithm, implemented in MAFFT (31), using the G-INS-i refinement method. The data were treated as codon partitioned, and a general time-reversible model [GTR (32)] was selected independently for each position within the codon, including both a gamma distribution and the proportion of invariable sites. Phylogenetic trees were constructed utilizing Bayesian inference (BI) and maximum likelihood (ML) approaches in MrBayes 3.2. (33) and RAxML 8.1.12 (34, 35), respectively. The BI analysis was run for 2 × 106 generations, sampling trees every 100 generations. The initial 30% of all saved trees were discarded as “burn-in” after checking that the standard deviation split frequency fell below 0.01. The convergence of the runs and the parameters of individual runs were checked using Tracer v. 1.7.1 (36). Posterior probabilities for each tree node were calculated as the frequency of samples recovering a given clade. The clade bootstrap support for ML trees was assessed by simulating 103 pseudoreplicates.
Results
A total of one, two, and eight ascarid nematodes were collected from each infected raccoon. Morphological examinations of all 11 ascarids (Figure 2) revealed characteristics consistent with B. procyonis as reviewed by (26). The majority of the collected specimens were females (F:M ratio 8:3). The length of the collected specimens ranged from 4.2 to 10.1 cm, and the width from 1 to 3 mm. All specimens carried identical ITS1-5.8S-ITS2 and COI alleles, respectively. The newly obtained ITS1-5.8S-ITS2 sequence (1,045 bp long, GenBank accession number PX417439) showed 99.3–100% similarity to conspecific sequences in GenBank (MZ092850-MZ092855) obtained from raccoons in China. The final COI alignment for building phylogenetic trees included 41 taxa and spanned 383 unambiguously aligned nucleotide positions. Both BI and ML analyses generated trees with identical topologies. The BI tree with posterior probabilities and bootstrap values (corresponding to the ML tree) along respective nodes is presented in Figure 3. The resulting trees clearly showed that the newly obtained COI sequences from Czechia (PX417440) are identical to the representative conspecific sequence retrieved from GenBank (MW385530, originating from a USA specimen). Congruently with previous studies, the Baylisascaris species showed a paraphyletic grouping with Parascaris and Ophidascaris species.

Figure 2. Baylisascaris procyonis photo comparison of female and male (A); and microphotographs of morphological details: anterior end with typical lips (B1); male posterior end with tip of spicule (B2); and female posterior end (B3).
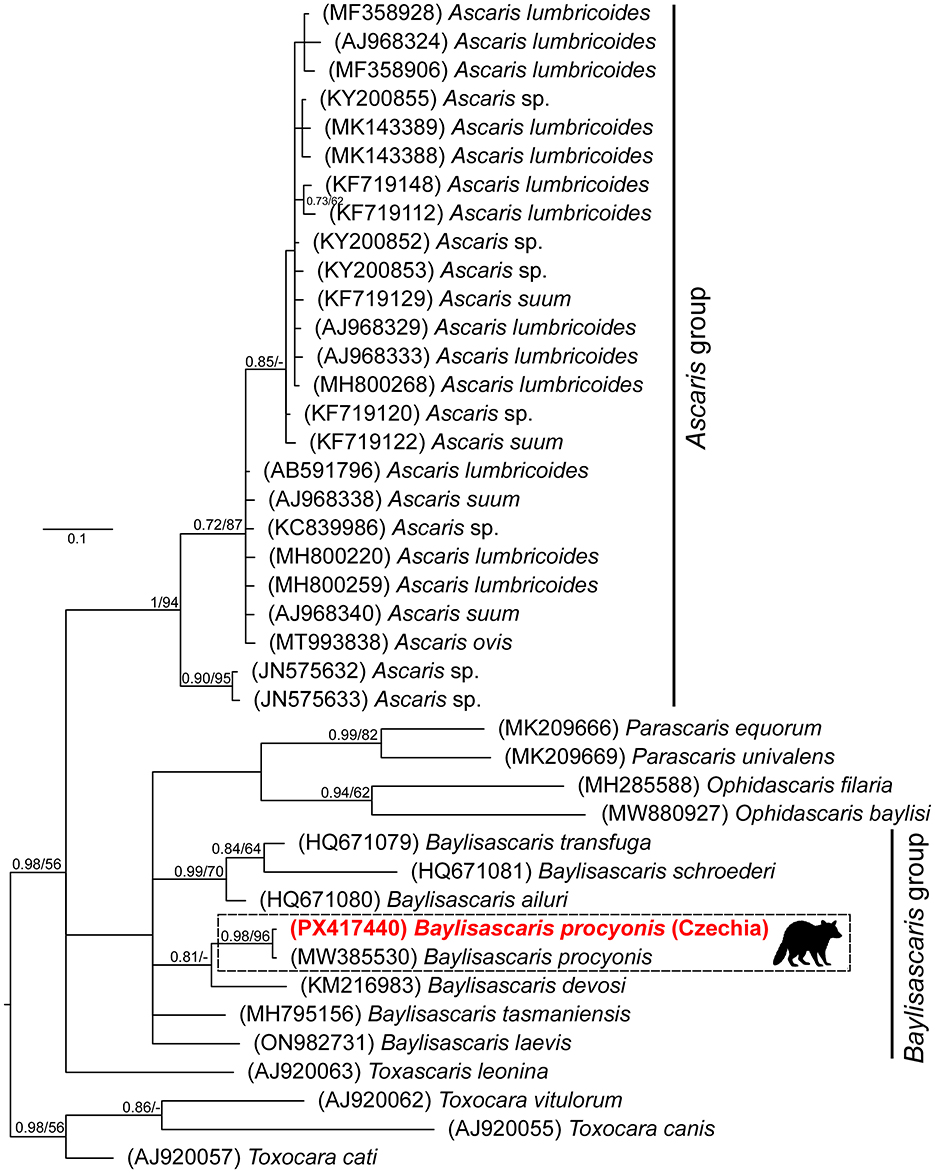
Figure 3. The phylogenetic tree of the Ascarididae species was built from partial COI sequences using Bayesian inference. The tree is rooted using Toxocara spp. as an outgroup. Values at the nodes indicate posterior probabilities from BI and bootstrap values from ML analyses. Dashes indicate nodal support values below 0.70 and 50, respectively. The newly obtained sequence from Czechia is in red.
Discussion
This study represents the first morphologically and molecularly confirmed detection of Baylisascaris procyonis in wild raccoons within the Czech Republic, extending the known range of this zoonotic nematode in Central Europe. Although Tenora and Staněk (37) previously reported this species from the vicinity of Brno based on larvae morphology, the origin of the host raccoons was uncertain, and the identification of nematodes was not confirmed by molecular data. Our findings thus provide the first definitive evidence of B. procyonis established in this region. Although only 3 out of 10 examined raccoons in Doupov Highlands were infected with B. procyonis, this finding is potentially epidemiologically significant given the parasite's capacity to cause severe disease in humans and its increasing prevalence in raccoon populations throughout Europe. In two of the three parasitized raccoons, the B. procyonis specimens were congruent in size with previously recorded conspecifics; i.e., males measuring 6.3 and 6.8 cm, and a single female measuring 10.1 cm [see reviewed meristic data in (14) and (38)]. In contrast, the third raccoon harbored eight roundworms that were comparatively smaller than previously reported, all females measuring 4.2–5.0 cm. The absence of eggs and the presence of only a weakly developed uterus suggest that these specimens were in the subadult stage explaining smaller size.
In neighboring countries, this parasite was recorded with a remarkably high prevalence in Germany, where out of 234 examined raccoons, 95% were carrying B. procyonis (39). In Austria, near the German and Czech borders, one raccoon found dead was parasitized by B. procyonis (13). As the molecular analyses showed the origin of this animal in Germany, this finding further supports the eastward spread of raccoons harboring allochthonous zoonotic diseases. A similar hypothesis was also postulated by Polish authors (10) as the parasite species were coprologically confirmed in raccoons near the Poland-Germany border.
The high prevalence of B. procyonis in some parts of Europe may be attributed to reduced parasite diversity in introduced European raccoon populations and high local host densities [such as in Germany (40)], which facilitate the transmission of diseases. Considering the direct life cycle of the parasite and its environmental contamination with infective eggs via feces, these parasites can easily be transmitted to humans, especially in areas where raccoons are expanding into human settlements (15, 41). Moreover, the higher prevalence of B. procyonis could also be related to Northern raccoon population density, considering the population of raccoons in Germany is approximately eight times bigger than in Czechia, as can be estimated from the harvest numbers obtained in the same hunting season (42, 43).
The detection of B. procyonis in the Czech Republic represents a significant finding in the context of invasive parasite monitoring and public health risk. This report from the western part of the country confirms the continued expansion of B. procyonis beyond its original introduced range in Germany and into neighboring countries. Given the parasite's zoonotic potential and its capacity to cause severe neurological disease in humans, its presence in the Czech Republic raises concerns about environmental contamination and the potential risk to both wildlife and human populations. The finding underscores the importance of continued surveillance of invasive raccoon populations, particularly synanthropic populations, as they often accommodate multiple zoonotic parasite species (39). Therefore, the necessity of eradicating the northern raccoon, an invasive species with potentially substantial impacts on native ecosystems, is further underscored from a public health perspective given the risk of parasite transmission. However, the management strategies aimed at controlling raccoon population size and dispersal potential have failed so far (44) highlighting the need for further research into population control measures.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation. Newly generated sequence data are available in GenBank under accession numbers PX417439 and PX417440.
Ethics statement
Ethical approval was not required for the study involving animals in accordance with the local legislation and institutional requirements because data were obtained from legally hunted individuals according to hunting legislation.
Author contributions
MB: Conceptualization, Data curation, Formal analysis, Investigation, Methodology, Supervision, Validation, Writing – original draft, Writing – review & editing. LŠv: Formal analysis, Methodology, Writing – original draft, Writing – review & editing. OM: Formal analysis, Methodology, Writing – original draft, Writing – review & editing. LŠk: Formal analysis, Writing – original draft, Writing – review & editing. VS: Data curation, Formal analysis, Writing – original draft, Writing – review & editing. JC: Data curation, Funding acquisition, Resources, Supervision, Writing – original draft, Writing – review & editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. The research was funded by the Technology Agency of the Czech Republic (project no. SS07020021), the Ministry of Agriculture of the Czech Republic (project no. QL24020333), the Scientific Grant Agency of the Slovak Republic (project no. VEGA 1/0583/22), and by Czech University of Life Sciences Prague, Faculty of Forestry and Wood Sciences (Excellent Team 2025).
Acknowledgments
We are grateful to Jitka Šišáková (an expert in the field) and Richard Lee Manore (a native speaker) for checking the English of this manuscript.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Gen AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher's note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
1. Simberloff D, Martin JL, Genovesi P, Maris V, Wardle DA, Aronson J, et al. Impacts of biological invasions: what's what and the way forward. Trends Ecol Evol. (2013) 28:58–66. doi: 10.1016/j.tree.2012.07.013
2. Dalmazzone S, Giaccaria S. Economic drivers of biological invasions: a worldwide, bio-geographic analysis. Ecol Econ. (2014) 105:154–65. doi: 10.1016/j.ecolecon.2014.05.008
3. Salgado I. Is the raccoon (Procyon lotor) out of control in Europe? Biodivers Conserv. (2018) 27:2243–56. doi: 10.1007/s10531-018-1535-9
4. French SK, Pearl DL, Peregrine AS, Jardine CM. Baylisascaris procyonis infection in Raccoons: a review of demographic and environmental factors influencing parasite carriage. Vet Parasitol Reg Stud Reports. (2019) 16:100275. doi: 10.1016/j.vprsr.2019.100275
5. Hernandez SM, Galbreath B, Riddle DF, Moore AP, Palamar MB, Levy MG, et al. Baylisascaris procyonis in raccoons (Procyon lotor) from North Carolina and current status of the parasite in the USA. Parasitol Res. (2013) 112:693–8. doi: 10.1007/s00436-012-3186-1
6. Kresta AE, Henke SE, Pence DB. Baylisascaris procyonis in raccoons in Texas and its relationship to habitat characteristics. J Wildl Dis. (2010) 46:843–53. doi: 10.7589/0090-3558-46.3.843
7. Rentería-Solís Z, Birka S, Schmäschke R, Król N, Obiegala A. First detection of Baylisascaris procyonis in wild raccoons (Procyon lotor) from Leipzig, Saxony, Eastern Germany. Parasitol Res. (2018) 117:3289–92. doi: 10.1007/s00436-018-5988-2
8. Heddergott M, Lippert S, Schliephake A, Gaede W, Schleimer A, Frantz AC, et al. Spread of the zoonotic nematode Baylisascaris procyonis into a naive Raccoon population. Ecohealth. (2023) 20:263–72. doi: 10.1007/s10393-023-01655-6
9. Reinhardt NP, Wassermann M, Härle J, Romig T, Kurzrock L, Arnold J, et al. Helminths in invasive raccoons (Procyon lotor) from Southwest Germany. Pathogens. (2023) 12:919. doi: 10.3390/pathogens12070919
10. Popiołek M, Szczesna-Staśkiewicz J, Bartoszewicz M, Okarma H, Smalec, B, Zalewski A. Helminth parasites of an introduced invasive carnivore species, the raccoon (Procyon lotor L), from the Warta Mouth National Park (Poland). J Parasitol. (2011) 97:357–60. doi: 10.1645/GE-2525.1
11. Al-Sabi MNS, Chriél M, Hansen MS, Enemark HL. Baylisascaris procyonis in wild raccoons (Procyon lotor) in Denmark. Vet Parasitol Reg Stud Rep. (2015) 1:55–8. doi: 10.1016/j.vprsr.2016.03.001
12. Davidson RK, Øines Ø, Hamnes IS, Schulze JE. Illegal wildlife imports more than just animals—Baylisascaris procyonis in raccoons (Procyon lotor) in Norway. J Wildl Dis. (2013) 49:986–90. doi: 10.7589/2012-06-154
13. Duscher GG, Frantz AC, Kuebber-Heiss A, Fuehrer HP, Heddergott M. A potential zoonotic threat: first detection of Baylisascaris procyonis in a wild raccoon from Austria. Transbound Emerg Dis. (2020) 68:3034–7. doi: 10.1111/tbed.13963
14. Lombardo A, Brocherel G, Donnini C, Fichi G, Mariacher A, Diaconu EL, et al. First report of the zoonotic nematode Baylisascaris procyonis in non-native raccoons (Procyon lotor) from Italy. Parasit Vectors. (2022) 15:24. doi: 10.1186/s13071-021-05116-3
15. Sorvillo F, Ash LR, Berlin OGW, Yatabe J, Degiorgio C, Morse SA. Baylisascaris procyonis: an emerging helminthic zoonosis. Emerg Infect Dis. (2002) 8:355–9. doi: 10.3201/eid0804.010273
16. Kazacos KR, Jelicks LA, Tanowitz HB. Baylisascaris larva migrans. In:Garcia HH, Tanowitz HB, Del Brutto OH, , editors. Handbook of Clinical Neurology. Amsterdam: Elsevier BV (2013). p. 251–62. doi: 10.1016/B978-0-444-53490-3.00020-0
17. Gavin PJ, Kazacos KR, Shulman ST. Baylisascariasis. Clin Microbiol Rev. (2005) 18:703–18. doi: 10.1128/CMR.18.4.703-718.2005
18. Kazacos KR. Baylisascaris procyonis and related species. In:Samuel WM, Pybus MJ, Kocan AA, , editors. Parasitic Diseases of Wild Mammals. 2nd ed. Ames: Iowa State University Press (2001). p. 301–41.
19. Goldberg MA, Kazacos KR, Boyce WM, Ai E, Katz B. Diffuse unilateral subacute neuritis: morphometric, serologic and epidemiologic support for Baylisascaris as a causative agent. Ophthalmology. (1993) 100:1695–701. doi: 10.1016/S0161-6420(93)31415-6
20. Kazacos KR, Raymond LA, Kazacos EA, Vestre WA. The Raccoon ascarid: a probable cause of human ocular larva migrans. Ophthalmology. (1985) 92:1735–43. doi: 10.1016/S0161-6420(85)34100-3
21. Jones H, Castro PDJ, Burton W, Yabsley MJ. Distribution and prevalence of Baylisascaris in domestic dogs in the United States and Canada, 2017–2023. One Health. (2025) 101095. doi: 10.1016/j.onehlt.2025.101095
22. Vaughan AM. Baylisascariasis (Raccoon roundworm infection) in two unrelated children—Los Angeles County, California, 2024. MMWR Morb Mortal Wkly Rep. (2025) 74. doi: 10.15585/mmwr.mm7428a1
23. Frantz AC, Lippert S, Gaasch J, Heddergott M, Kox L. First records of the raccoon roundworm (Baylisascaris procyonis) in Luxembourg: evidence for natural spread and a novel introduction. Eur J Wildlife Res. (2024) 70:91. doi: 10.1007/s10344-024-01844-w
24. Conraths FJ. Arbeitsplatzbedingte Infektionen des Menschen mit dem Waschbarspulwurm Baylisascaris procyonis. Arbeitsmed Sozialmed Umweltmed. (1996) 31:13–17. German.
25. Benovics M, Nosková E, Klimešová A, Škorpíková L, Jaššová E, Drimaj J, et al. Helminth diversity of nutria (Myocastor coypus) across the Morava basin in the Czech Republic. Parasitology. (2025) 152:61–71. doi: 10.1017/S0031182024001628
26. Kazacos KR. Baylisascaris procyonis and related species. In:Samuel WM, Pybus MJ, Kocan AA, , editors. Parasitic Diseases of Wild Mammals. 2nd ed. Ames: Iowa State University Press (2008). p. 301–41.
27. Sprent JFA. Notes on Ascaris and Toxascaris, with a definition of Baylisascaris gen. nov. Parasitology. (1968) 58:185–98. doi: 10.1017/S0031182000073534
28. Vrain TC, Wakarchuk DA, Levesque AC, Hamilton RI. Intraspecific rDNA restriction fragment length polymorphism in the Xiphinema americanum group. Fundam Appl Nematol. (1992) 15:563–73.
29. Nakano T, Okamoto M, Ikeda Y, Hasegawa H. Mitochondrial cytochrome c oxidase subunit 1 gene and nuclear rDNA regions of Enterobius vermicularis parasitic in captive chimpanzees with special reference to its relationship with pinworms in humans. Parasitol Res. (2006) 100:51–7. doi: 10.1007/s00436-006-0238-4
30. Civánová KríŽová K, Seifertová M, Baruš V, Hodová I, Mašová Š, Nurcahyo W, et al. First study of Ascaris lumbricoides from the semi-wild population of the Sumatran orangutan Pongo abelii in the context of morphological description and molecular phylogeny. Life. (2023) 13:1016. doi: 10.3390/life13041016
31. Katoh K, Rozewicki J, Yamada KD. MAFFT online service: multiple sequence alignment, interactive sequence choice and visualization. Brief Bioinform. (2019) 20:1160–6. doi: 10.1093/bib/bbx108
32. Lanave C, Preparata G, Saccone C, Serio G. A new method for calculating evolutionary substitution rates. J Mol Evol. (1984) 20:86–93. doi: 10.1007/BF02101990
33. Ronquist F, Teslenko M, van der Mark P, Ayres DL, Darling A, Höhna, et al. MrBayes 3.2: efficient Bayesian phylogenetic inference and model choice across a large model space. Syst Biol. (2012) 61:539–42. doi: 10.1093/sysbio/sys029
34. Stamatakis A. RAxML-VI-HPC: maximum likelihood-based phylogenetic analyses with thousands of taxa and mixed models. Bioinformatics. (2006) 22:2688–90. doi: 10.1093/bioinformatics/btl446
35. Stamatakis A. RAxML version 8: a tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics. (2014) 30:1312–3. doi: 10.1093/bioinformatics/btu033
36. Rambaut A, Drummond AJ, Xie D, Baele G, Suchard MA. Posterior summarization in Bayesian phylogenetics using Tracer 1.7. Syst Biol. (2018) 67:901–4. doi: 10.1093/sysbio/syy032
37. Tenora F, Staněk M. Scanning electron microscopy of Toxocara procyonis Sefanski et Zarnowski, 1951 (Nematoda). Helminthologia. (1990) 27:73–7.
38. Gu X-H, Chen H-X, Hu J-J, Li L. Morphology and ASAP analysis of the important zoonotic nematode parasite Baylisascaris procyonis (Stefahski and Zarnowski, 1951), with molecular phylogenetic relationships of Baylisascaris species (Nematoda: Ascaridida). Parasitology. (2024) 151:200–12. doi: 10.1017/S0031182023001312
39. Peter N, Dörge DD, Cunze S, Schantz AV, Skaljic A, Rueckert S, et al. Raccoons contraband—the metazoan parasite fauna of free-ranging raccoons in central Europe. Int J Parasitol Parasites Wildl. (2023) 20:79–88. doi: 10.1016/j.ijppaw.2023.01.003
40. Fischer ML, Sullivan MJP, Greiser G, Guerrero-Casado J, Heddergott M, Hohmann U, et al. Assessing and predicting the spread of non-native raccoons in Germany using hunting bag data and dispersal-weighted models. Biol Invasions. (2016) 18:57–71. doi: 10.1007/s10530-015-0989-x
41. Graef-Teixeira C, Morassutti AL, Kazacos KR. Update on Baylisascariasis, a highly pathogenic zoonotic infection. Clin Microbiol Rev. (2016) 29:375–99. doi: 10.1128/CMR.00044-15
42. Czech Statistical Office. Usmrcení dalších Živočichu. Czechia: Czech Statistical Office (2025). Available online at: https://vdb.czso.cz/vdbvo2/faces/cs/index.jsf?page=vystup-objekt&skupId=2342&z=T&f=TABULKA&katalog=30841&pvo=LES035&pvo=LES035 (Accessed Aug 12, 2025). Czech.
43. Jagdverband D. Jahresjagdstrecke Bundesrepublik Deutschland (Jagdjahr 2023/24). Berlin: Deutscher Jagdverband (2025). Available online at: https://www.jagdverband.de/sites/default/files/2025-02/2025-02_Infografik_Jahresjagdstrecke_Bundesrepublik_Deutschland_2023_2024.jpg (Accessed August 12, 2025). German.
{kind=link}
Keywords: introductions, zoonoses, Nematoda, neurotropic diseases, Doupov Highlands, molecular identification
Citation: Benovics M, Švantnerová L, Mikulka O, Škorpíková L, Skoták V and Cukor J (2025) First record of Baylisascaris procyonis in the wild invasive Northern raccoon (Procyon lotor) in the Czech Republic. Front. Vet. Sci. 12:1686564. doi: 10.3389/fvets.2025.1686564
Received: 15 August 2025; Accepted: 24 September 2025;
Published: 14 October 2025.
Edited by:
Miljenko Bujanić, University of Zagreb, CroatiaReviewed by:
Roman Kuchta, Academy of Sciences of the Czech Republic (ASCR), CzechiaCalin Mircea Gherman, University of Agricultural Sciences and Veterinary Medicine of Cluj-Napoca, Romania
Copyright © 2025 Benovics, Švantnerová, Mikulka, Škorpíková, Skoták and Cukor. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Jan Cukor, Y3Vrb3JAZmxkLmN6dS5jeg==