 Andrea Šimková
Andrea Šimková Martina Dávidová1
Martina Dávidová1 Pavel Hyršl
Pavel Hyršl Markéta Ondračková
Markéta Ondračková- 1Faculty of Science, Department of Botany and Zoology, Masaryk University, Brno, Czechia
- 2Faculty of Fisheries and Protection of Waters, South Bohemian Research Center of Aquaculture and Biodiversity of Hydrocenoses, University of South Bohemia in České Budějovice, Vodňany, Czechia
- 3Faculty of Science, Department of Experimental Biology, Masaryk University, Brno, Czechia
- 4Institute of Vertebrate Biology of the Czech Academy of Sciences, Brno, Czechia
Introduction: Artificial induction of polyploidy in fishes is a widely used method in commercial aquaculture due to its economic potential and its association with changes in cell morphology and physiology that can significantly affect individual fitness. Using tench (Tinca tinca, Cyprinidae), a fish species extensively farmed in European aquaculture, we investigated differences in parasite susceptibility between triploid and diploid fish and analyzed the potential effect of metazoan parasite infection on fish condition, physiology, and health status.
Methods: Amphimictic diploid and induced triploid specimens collected from a breeding pond were examined during four sampling events over the course of one year, focusing on the presence of metazoan parasites and selected fish condition, physiological, and immune parameters.
Results: Diploids generally exhibited a higher overall parasite load than triploids throughout the year; however, this difference was statistically significant only in summer, coinciding with the extremely high abundance of the tench-specific Asymphylodora tincae. Host ploidy appeared to influence susceptibility or resistance to particular parasite species. While amphimictic diploid tench showed higher susceptibility to adult trematode A. tincae, triploid tench were more parasitized by the larval cestode Valipora campylancristrota. No difference in abundance of monogenean Gyrodactylus tincae was observed between amphimictic diploid and induced triploid specimens. Other parasites were relatively rare in both groups. Although no clear association between parasite infection and the measured physiological or immune parameters was found, significant negative correlations were more frequently observed in diploid fish than in triploids.
Discussion: The differences in parasite infection between triploid and diploid tench and the associations between parasite load and condition- and health-related traits may be explained by (1) presumed higher heterozygosity in triploids, (2) physiological differences related to cell size and number of cells in key organs and tissues, (3) variation in feeding performance, and (4) host–parasite coevolutionary interactions.
1 Introduction
Polyploidization, the multiplication of the whole chromosome complement, is a widespread phenomenon in plants and occurs sporadically in animals. Polyploidy represents a special type of mutation and can arise through several mechanisms, such as genomic doubling, gametic nonreduction, or polyspermy (1). It is consistently associated with changes in cell morphology and physiology, which can significantly affect individual fitness (2, 3). In fish, spontaneous polyploidy is known to occur both within species (autopolyploidy) and across species (allopolyploidy) [e.g., Leggatt and Iwama (4), Cunha et al. (5), Havelka et al. (6), and Schreier et al. (7)]. Polyploidy may confer advantages in certain environments, such as increased stress tolerance or altered ecological interactions, resulting in novel traits and enhanced adaptation to new conditions.
In fish selective breeding biotechnological methods, including chromosomal manipulations such as induction of polyploidy, gynogenesis, androgenesis, or sex reversal, have been widely applied (8, 9). In particular, the artificial induction of polyploidy in fishes, most commonly aimed at producing triploid forms, has become an economically attractive and extensively used method in commercial aquaculture. The primary motivation for inducing triploidy in aquaculture is the expectation of beneficial traits, especially enhanced growth performance. This is largely attributed to the sterility of triploids, which prevents the growth depression typically associated with sexual maturation of diploids. Additional advantages include reduced sexual and territorial behavior, leading to lower stress levels and decreased energy expenditure, as well as increased survival in cases where reproduction is associated with elevated mortality of diploids (10, 11). Previous studies have confirmed that diploid and triploid fish differ in growth, survival, slaughter value, and flesh composition [e.g., Piferrer et al. (10), Flajšhans et al. (11), Buchtová et al. (12–14), Tiwary et al. (15), and Tolarová et al. (16)], as well as in haematological or biochemical profiles (2, 10, 15–20), immune performance (2, 10, 16, 21), and behavior (15, 22).
Host–parasite interactions are often characterized by narrow genetic specificity (23), which may explain why parasites can reflect changes in host ploidy status. Higher allelic diversity at immune-related genes in polyploid hosts may allow recognition of a broader range of parasite species (24). Moreover, if additional genome copies are expressed, polyploids may produce greater quantities of immune-related gene products (3), making it more difficult for parasites to evade host recognition mechanisms (24, 25). However, polyploidy may not necessarily generate functional diversity unless increased allelic variation is reflected at the protein level.
The impacts of polyploidy on immune defense in fish have been documented [e.g., Šimková et al. (21), Hakoyama et al. (26), Langston et al. (27), and Fuad et al. (28)]. For example, triploid gynogenetic females of goldfish (Carassius auratus L., 1758) exhibited significantly higher parasite load than diploid sexual females, which was explained by reduced phagocytic activity in triploid form (26). In the study by Šimková et al. (21), the most common triploid MHC (major histocompatibility complex) genotype of gibel carp (Carassius gibelio Bloch, 1782) was more heavily parasitized by gill monogeneans compared to diploids and less frequent triploid MHC variants from the same population. Fuad et al. (28) reported high infection levels of eye trematodes, and underexpression of numerous immunity-related genes in gynogenetic triploid gibel carp compared to sexual diploids, supporting the idea that certain loci, or even whole genomes, may be up- or down-regulated (or even silenced) as ploidy increases, a phenomenon documented in allopolyploids (3). However, polyploidy may produce non-uniform effects on parasites, as demonstrated in plants and phytophagous moths (29), showing that polyploid plants less infected by one phytophagous insect species were significantly more infected by two others.
The present study aimed to investigate the effect of host ploidy status on metazoan parasite infection in tench (Tinca tinca L., 1758, Cyprinidae), a commercially important fish species widely bred in European pond aquaculture. The Czech Republic is one of the leading producers of marketable tench in the Europe. Its production relies either on natural spawning of selected broodstock under controlled conditions or completely on artificial reproduction (30). Artificial production of triploid tench has been shown to result in higher marketable weight and improved product quality (11). However, a high incidence of spontaneous triploidy has also been observed in farmed tench, attributed to natural predisposition for egg overripening and the presence of a recessive allele responsible for the failure of the second polar body extrusion when homozygous (31).
Growth, physiological traits, and immune response of diploid and triploid tench have been extensively studied (11, 16, 17), and the health status of farmed tench has been previously investigated by Svobodová and Kolářová (32) and Piačková and Flajšhans (33). In this integrative study of tench health, we focused on year-long temporal variation in metazoan parasite load, as well as condition, haematological, biochemical, and immunological parameters, to assess (1) differences in parasite susceptibility between induced triploid and amphimictic diploid tench and (2) the potential effect of metazoan parasite infection on fish condition, physiology, and health status.
2 Materials and methods
2.1 Host and parasite collection
Farmed amphimictic diploid and induced triploid specimens of tench were sampled from a breeding pond located at the South Bohemian Research Centre of Aquaculture and Biodiversity of Hydrocenoses in Vodňany, Czech Republic. Triploid tench was produced by cold shock as described by Flajšhans et al. (34). Fish sampling was conducted using seine netting during four seasonal periods (water temperature was measured at the time of fish sample collection): September (16.9 °C), December (2.1 °C), March (7.4 °C), and June (16.4 °C). A total of 160 tench aged five to six years were examined, including 81 diploid and 79 triploid specimens.
Immediately after capture, skin mucus was gently stripped and placed into sterile tube. Blood samples were drawn from the caudal vein following Pravda and Svobodová (35) and mixed with heparin (50 IU.ml−1 of blood, Zentiva). Fish were subsequently transported to the laboratory and individually euthanised by spinal transection prior to dissection. Standard and total body length (in mm), total body weight, and eviscerated (without internal organs) body weight (to the nearest 0.1 g) were recorded. Wet weights of gonads, liver and spleen were measured (to the nearest 0.001 g) and used for calculation of physiological indices: hepatosomatic index (HSI), splenosomatic index (SSI), and gonadosomatic index (GSI) [see Rohlenová et al. (36) for more details]. The number of fish examined per group (ploidy/sampling period), total length, and all physiological parameters are presented in Table 1. Ploidy level of each specimen was verified from the blood sample by means of flow cytometry as relative DNA content in peripheral blood cells. Samples were processed according to Vindelov and Christensen (37).
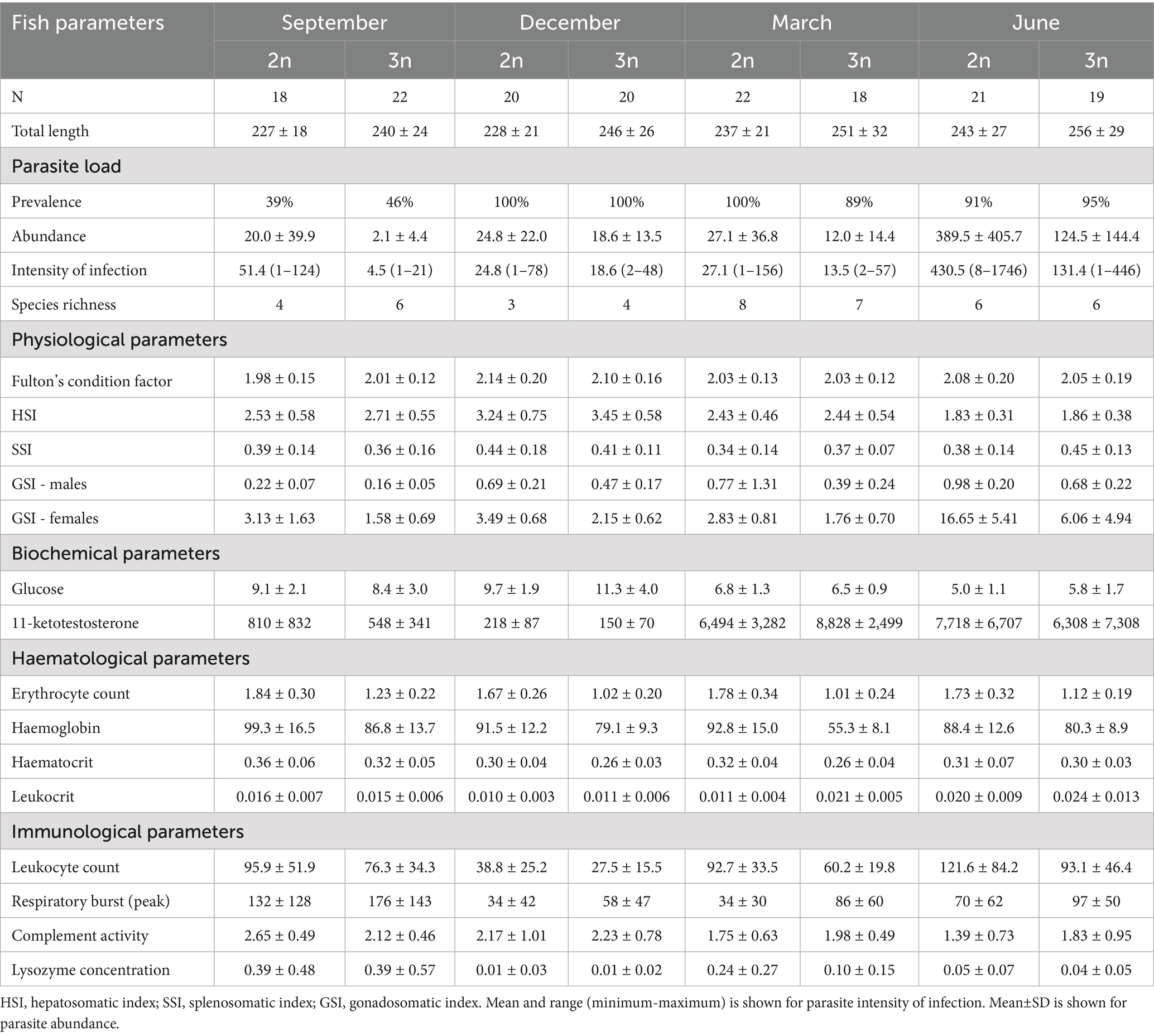
Table 1. Sample size (N), total fish length (in mm, mean±SD), parasite load, physiological, biochemical and immunological parameters (mean±SD is shown for each parameter) for amphimictic diploid (2n) and induced triploid (3n) tench.
Tench specimens were screened for metazoan parasites using binocular stereomicroscopes. All parasites were removed from examined organs and preserved according to standard parasitological procedures (38). Monogenea and Myxosporea were preserved as semi-permanent slides using a mixture of ammonium picrate and glycerine (39). Digenea, Cestoda, Mollusca, Crustacea, and Acarina were preserved in 4% formaldehyde or 70% ethanol (38), with digeneans and cestodes subsequently stained with ferric acetocarmine and mounted in Canada balsam (40). Parasites were identified using a light microscope (Olympus BX 50) equipped with phase-contrast, differential interference contrast (DIC according to Nomarski), and measured using Olympus Digital Image Analysis.
Characteristics of parasite infection, including prevalence, abundance, and intensity of infection, were calculated according to Bush et al. (41). Prevalence was defined as the percentage of fish infected by a given parasite species in a sample; mean abundance as the average number of parasites per host (including infected and uninfected individuals); and mean intensity of infection as the average number of parasites per infected host. Intensity range indicates the minimum and maximum number of parasites per infected host. Parasite species richness, defined as the total number of parasite species, was calculated for each fish group and per individual host. Parasite species strictly associated with tench were classified as specialists [e.g., Gusev (42) and Našincová and Scholz (43)], while those with a broad host range [based on published studies, e.g., Moravec (44)] were considered generalists (Table 2).
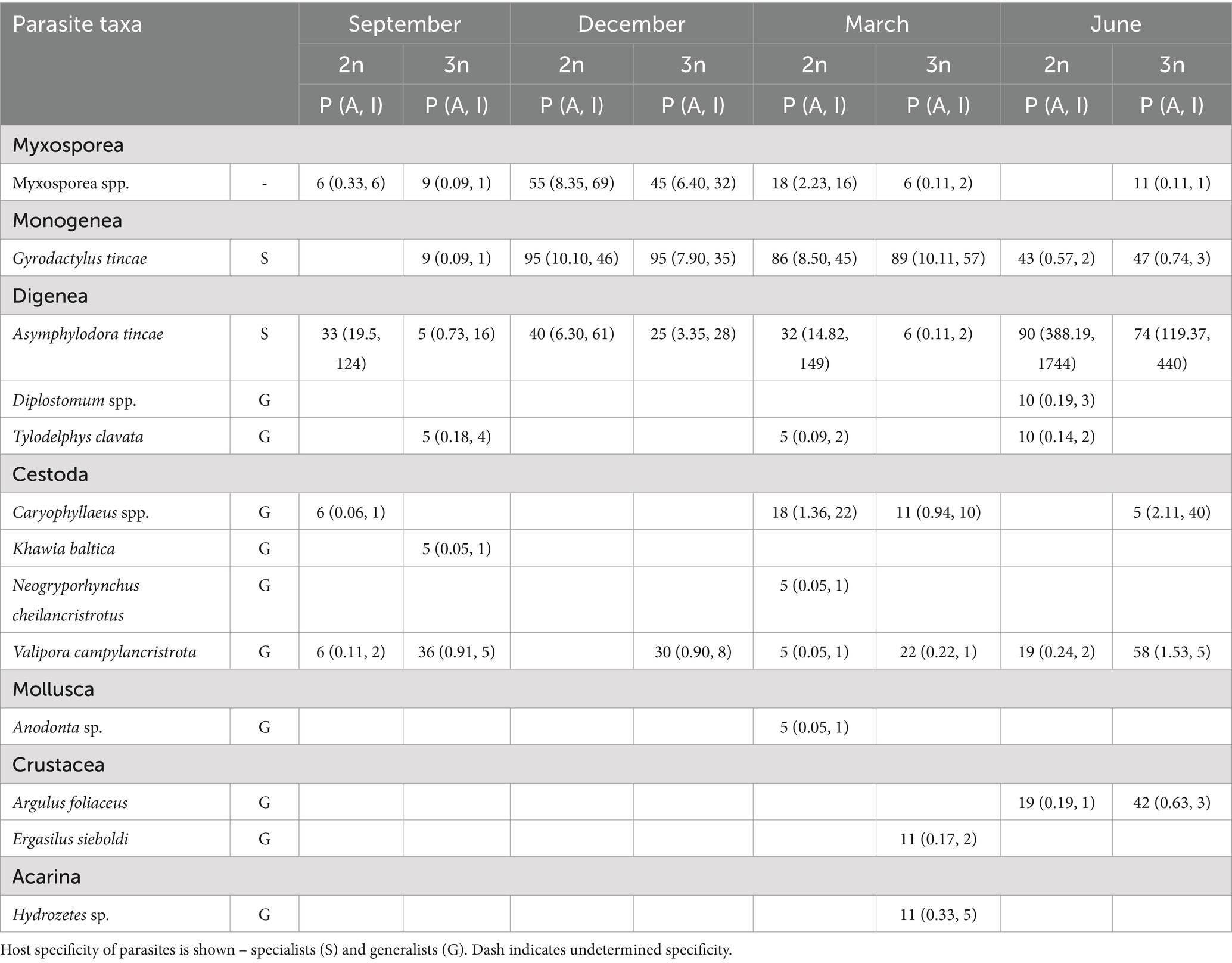
Table 2. Prevalence (P in %) and mean abundance and maximum intensity of infection (A, I) of metazoan parasites infecting farmed amphimictic diploid (2n) and induced triploid (3n) tench.
2.2 Haematological, biochemical, and immunological parameters
Total erythrocyte count (expressed in T.l−1) and leukocyte count (in G.l−1) were determined using a Bürker hemocytometer after dilution of heparinised blood in Natt-Herrick solution at 1:200 ratio (45, 46). Haemoglobin content (in g.l −1) was assessed photometrically at 540 nm (Helios Unicam, USA), using Kampen-Zijlster transformation medium (33). Haematocrit and leukocrit values (in l.l−1) were measured using 75 mm long heparinized microcapillaries with an inner volume of 60 μm. Centrifugation was performed at 12,000 g for 3 min using a haematocrit centrifuge (45).
Blood samples for respiratory burst activity were prepared for each fish according to Buchtíková et al. (47) and analyzed within two hours of collection. The kinetics of luminol-enhanced chemiluminescence were measured at 22 °C for one hour at room temperature using LM01-T luminometer (Immunotech, Czech Republic). The peak curve, expressed in relative light units (RLU) represented the maximal intensity of respiratory burst. Complement activity was assessed following Buchtíková et al. (47) and calculated as the inverse of the time (in h−1) required to kill 50% of Escherichia coli, based on the difference between the maximum measurement time (4 h) and the observed time to 50% lysis.
Lysozyme concentration in skin mucus (in mg.ml−1) was determined by radial diffusion in agarose containing Micrococcus luteus (CCM 169) according to Poisot et al. (48). The concentration of 11-ketotestosterone (in pg.ml−1) in male plasma was measured using a commercial competitive enzyme immunoassay kit (Cayman Chemical, Estonia). Plasma glucose levels (in mmol.l−1) were analyzed using the commercial Biolatest Glukosa Liquid 500 kit (PLIVA-Lachema Diagnostika s.r.o., Brno, Czech Republic). All measured haematological, biochemical, and immunological parameters are presented in Table 1.
2.3 Data analysis
The effects of season, ploidy, and sex on response variables were tested using either linear models (LM) or generalized linear models (GLM). GLMs were applied to all parasitological characteristics and leukocyte count (all assumed to follow a negative binomial distribution, except parasite species richness, which was modelled using a quasi-Poisson distribution). LMs were used for all other condition, physiological, and immunological parameters.
The null model for each response variable included season, ploidy, and sex as predictors. Interactions were excluded due to limited number of replications. For GSI (analyzed separately for males and females) and 11-ketotestosterone (measured only in males), the null model contained only season and ploidy. A backward stepwise approach was used to simplify models by removing predictors that did not significantly affect the fit of the model, as determined by likelihood ratio tests (49). Tukey’s HSD test was applied to control for Type II error in multiple post-hoc pairwise comparisons [using the glht and mcp functions from multcomp package; Hothorn et al. (50)]. Where necessary, i.e., when visual inspection of residuals revealed patterns indicating biased models, such as trends in residuals and their variance, the response variables were log-transformed using log (x + 1), and the model was re-evaluated. This adjustment was applied to male and female GSI, 11-ketotestosterone, leukocrit, lysozyme concentration, respiratory burst peak, and glucose level.
Spearman correlation tests were used to assess associations between total parasite abundance or abundance of the four most abundant parasite species and fish condition, physiological, haematological, and immunological parameters, calculated separately for diploids and triploids. Bonferroni correction was applied to adjust significance levels and reduce the probability of committing a Type I error. All statistical analyses were performed in R version 4.1.2 (51), using the MASS (52), multcomp (50), and emmeans (53) packages.
3 Results
3.1 Parasite load
A total of thirteen taxa of parasitic metazoans were identified on farmed tench, with ten taxa present in diploid and ten in triploid fish (Table 2). Parasite species richness did not differ significantly between ploidy levels (GLM, p > 0.05; Figure 1B). However, the overall parasite abundance was significantly lower in triploids compared to diploids (GLM, p < 0.001; Figure 1A). Triploids exhibited significantly reduced abundance of specialist parasites (GLM, p < 0.001), whereas the abundance of generalists was significantly lower in diploids than in triploids (GLM, p < 0.001) (Table 3; Figures 1C,D).
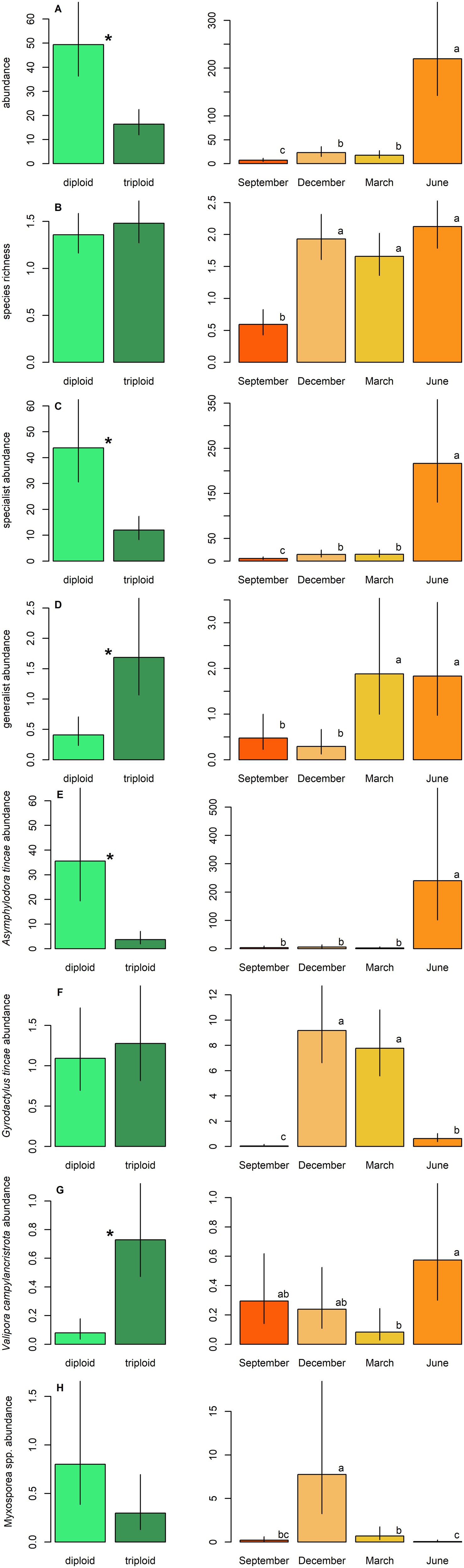
Figure 1. Parasite abundance and species richness in tench, differences between diploids and triploids (green bars) and among sampled seasons (orange bars) are shown. (A) Total abundance, (B) species richness, (C) abundance of specialists, (D) abundance of generalists, (E) abundance of Asymphylodora tincae, (F) abundance of Gyrodactylus tincae, (G) abundance of Valipora campylancristrota, (H) abundance of Myxosporea spp. Ploidy bars with *differ significantly from each other. Season bars with the different letters differ significantly from each other.
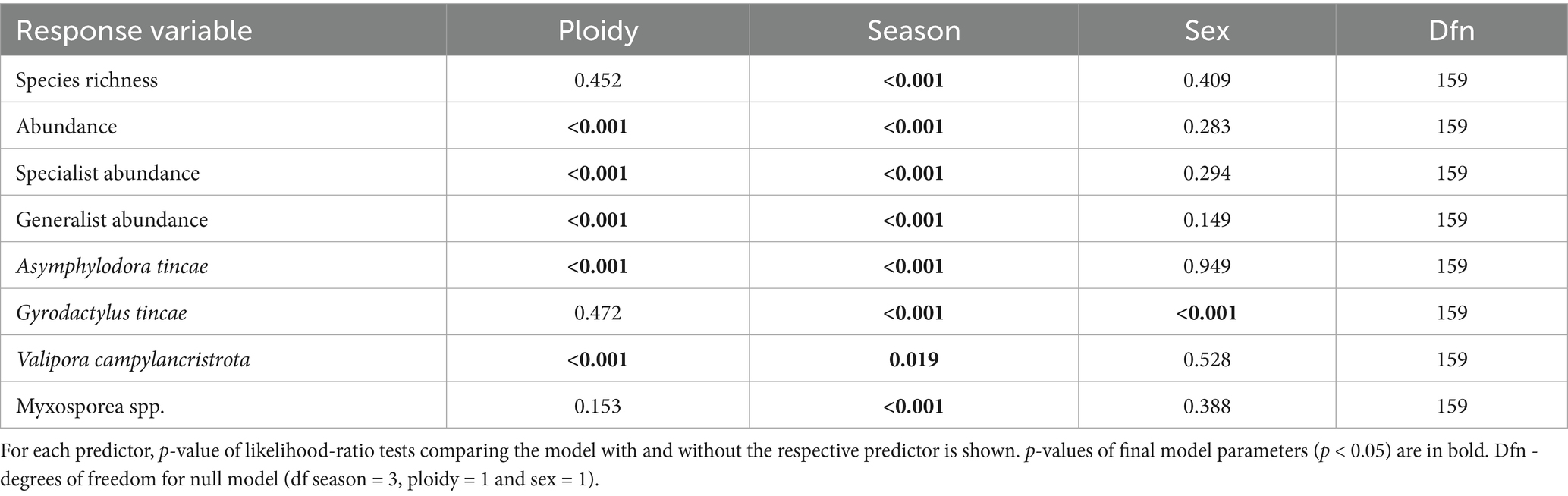
Table 3. Contribution of three predictors (season, ploidy and sex) in models explaining tench parasite assemblage characteristics.
Four taxa were consistently present throughout the year: the monogenean Gyrodactylus tincae Malmberg, 1957, the adult trematode Asymphylodora tincae (Modeer, 1790), the larval cestode Valipora campylancristrota (Wedl, 1855), and an unspecified group of myxosporeans (Myxosporea spp.) (Table 2). Asymphylodora tincae was the most abundant and one of two the most prevalent parasite species, reaching also the highest maximum intensity of infection, followed by G. tincae as the second most abundant. Higher abundance of A. tincae along with lower abundance of V. campylancristrota was observed in diploid tench (GLM, both p < 0.001) (Table 3; Figures 1E,G). No significant effect of ploidy was observed on the abundance of Myxosporea spp. or G. tincae (GLM, both p > 0.05) (Table 3; Figures 1F,H).
Sex had no significant effect on most parasitological parameters, with the exception of abundance of G. tincae, which was significantly lower in males than in females (GLM; p < 0.001; Table 3). Seasonal variation significantly affected all measured characteristics of parasite infection (GLM; P all < 0.05; Table 3; Figure 1). Overall parasite abundance, as well as abundance of A. tincae, V. campylancristrota, and specialist parasites (see Table 2), peaked in June (Figures 1A,C,E,G). Generalist abundance was highest in March and June (Figure 1D), while the Myxosporea spp. and G. tincae reached their highest abundance in December or December and March, respectively (Table 3; Figures 1F,H). Overall, parasite infection levels were lowest in September.
3.2 Physiological parameters
GSI was significantly higher in diploids compared to triploids for both sexes (LM, p < 0.001; Table 4; Figures 2D,E). Other condition-related parameters showed no significant dependence on ploidy (LM, p > 0.05, Table 4). Seasonal variation significantly affected all condition parameters (Table 4; Figure 2). GSI peaked in June for both sexes, while HSI reached its highest in December and lowest in June. Fulton’s condition coefficient remained relatively stable across seasons, with December values significantly exceeding those recorded in September (Figure 2A). SSI and HSI were higher in females than in males, whereas the opposite trend was observed in Fulton’s condition coefficient (Table 4).
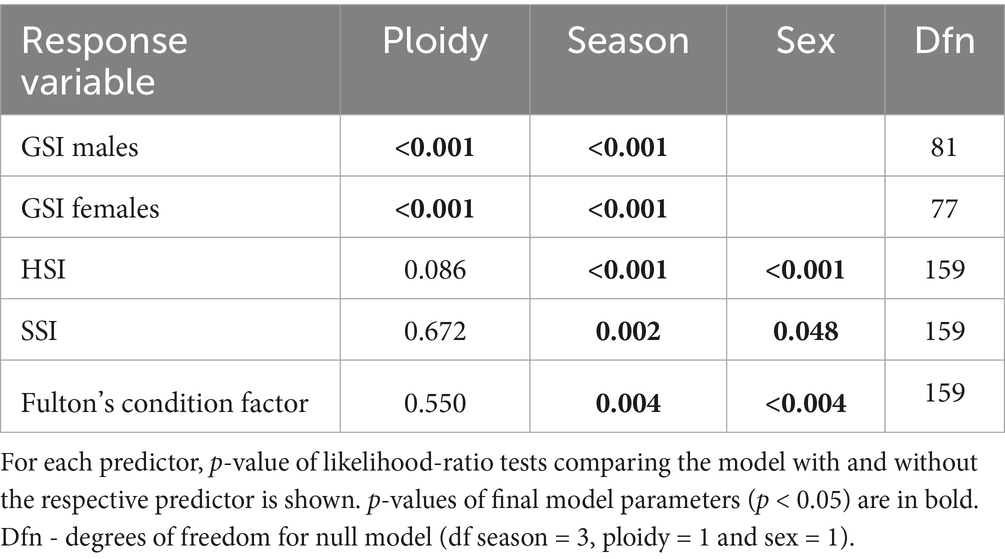
Table 4. Contribution of three predictors (season, ploidy and sex) in models explaining tench physiological characteristics.
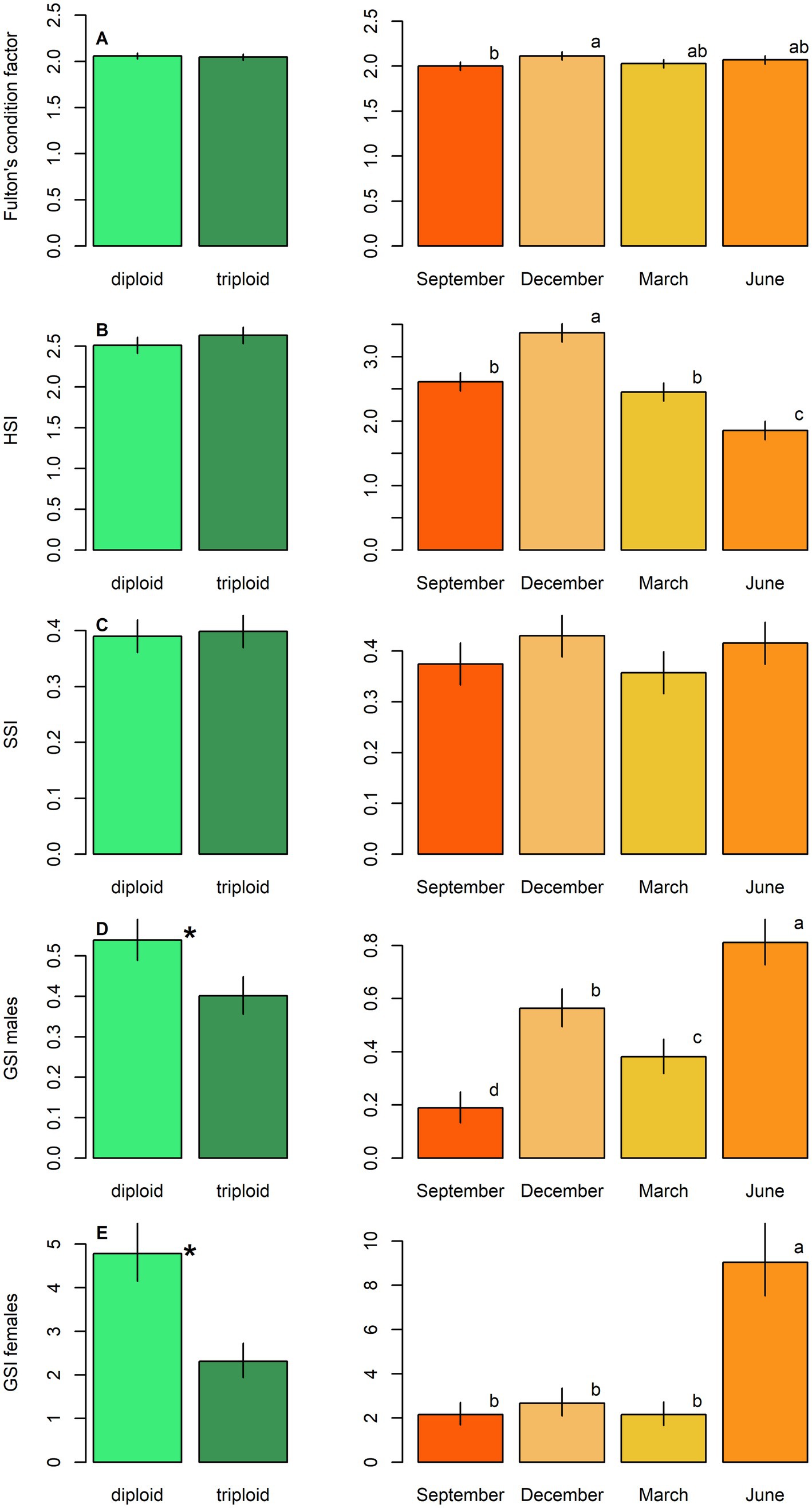
Figure 2. Physiological parameters, including: (A) Fulton’s condition factor, (B) hepatosomatic index (HSI), (C) splenosomatic index (SSI), and (D) gonadosomatic index in males (GSI males) and (E) females (GSI females) in diploid and triploid tench (green bars), and their seasonal fluctuation (orange bars). Ploidy bars with *differ significantly from each other. Season bars with the different letters differ significantly from each other.
3.3 Haematological, biochemical and immunological parameters
Diploids exhibited higher erythrocyte counts, haematocrit, and haemoglobin content compared to triploids, while the reverse pattern was observed for leukocrit (Table 5; Figure 3). No significant effect of ploidy was found for glucose levels and 11-ketotestosterone concentrations (Table 5; Figure 4). Leukocyte counts were significantly higher in diploids (Figure 5A), whereas triploids showed elevated respiratory burst peak (Figure 5B). No ploidy-related differences were detected in complement activity or lysozyme concentration (Table 5; Figures 5C,D). Haematocrit, leukocrit, and leukocyte counts were significantly higher in females than in males (Table 5).
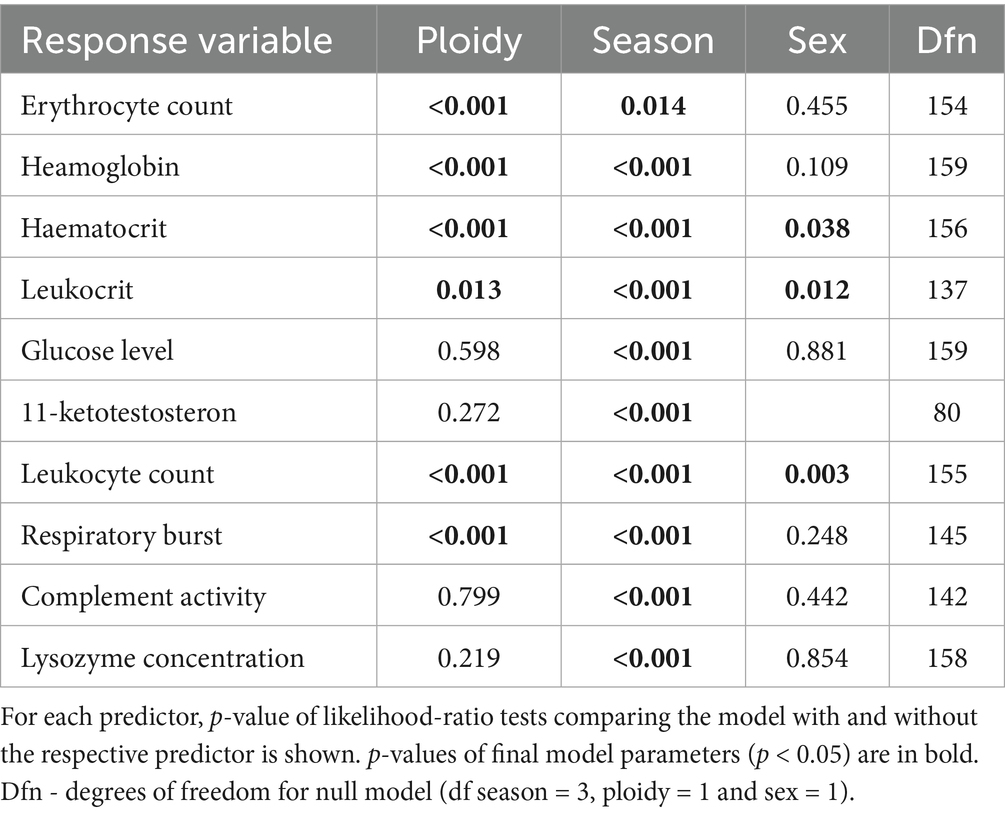
Table 5. Contribution of three predictors (season, ploidy and sex) in models explaining tench haematological, biochemical, and immunological parameters.
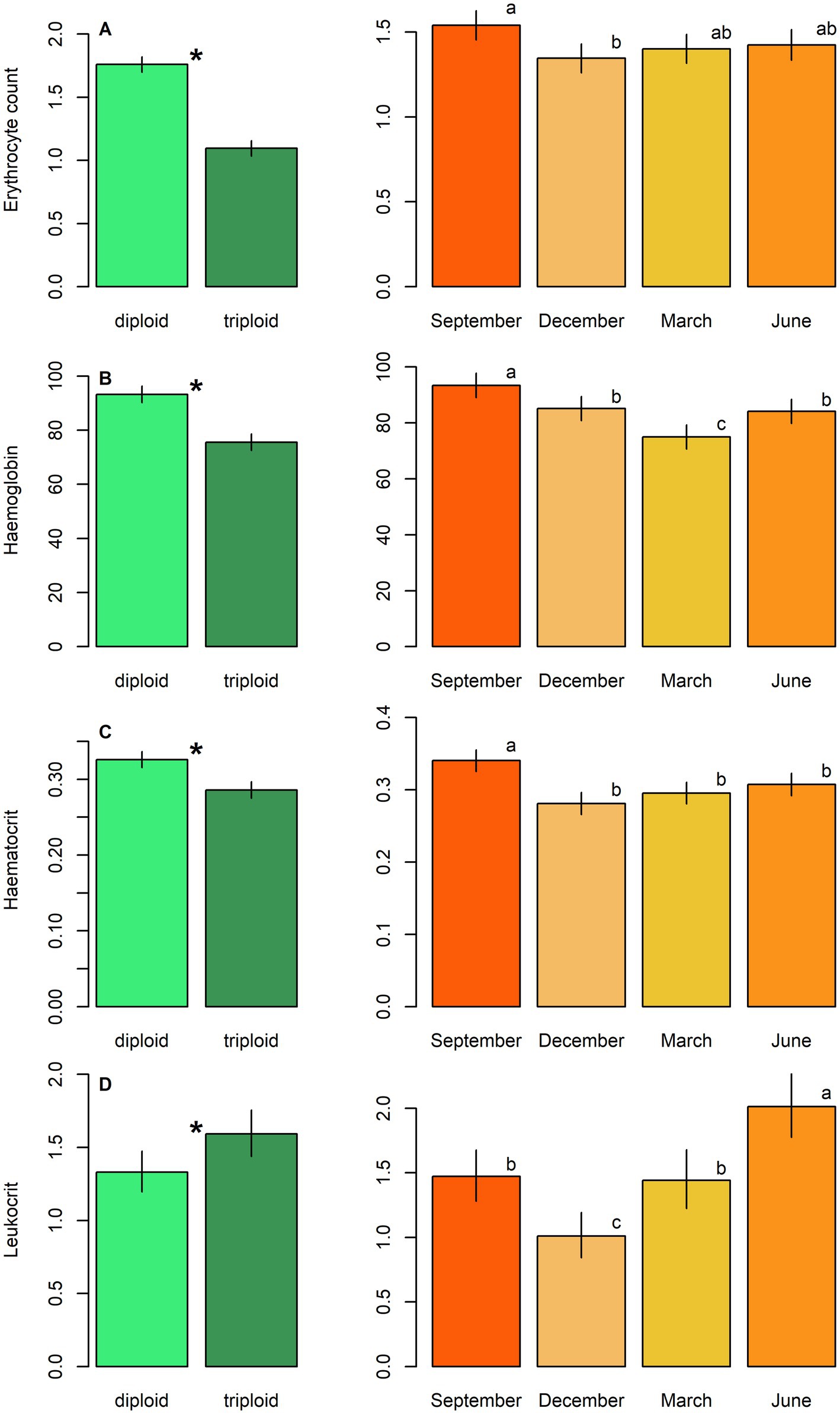
Figure 3. Haematological parameters, including: (A) Erythrocyte count, (B) haemoglobin concentration, (C) haematocrit and (D) leukocrit in plasma samples of diploid and triploid tench (green bars), and their seasonal fluctuation (orange bars). Ploidy bars with * differ significantly from each other. Season bars with the different letters differ significantly from each other.
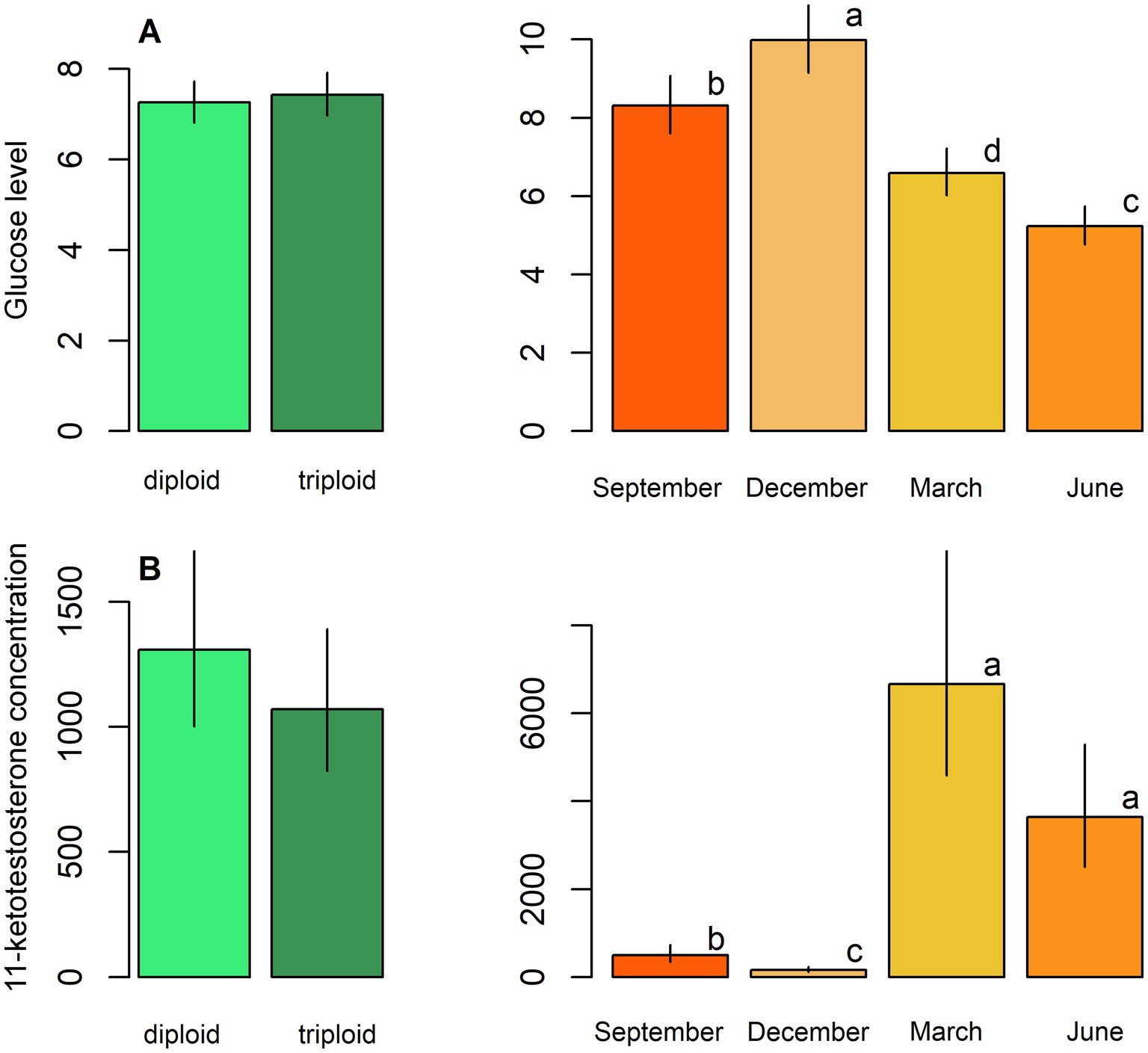
Figure 4. Biochemical parameters, including: (A) Glucose level and (B) 11-ketotestosterone concentration in males in plasma samples of diploid and triploid tench (green bars), and their seasonal fluctuation (orange bars). Season bars with the different letters differ significantly from each other.
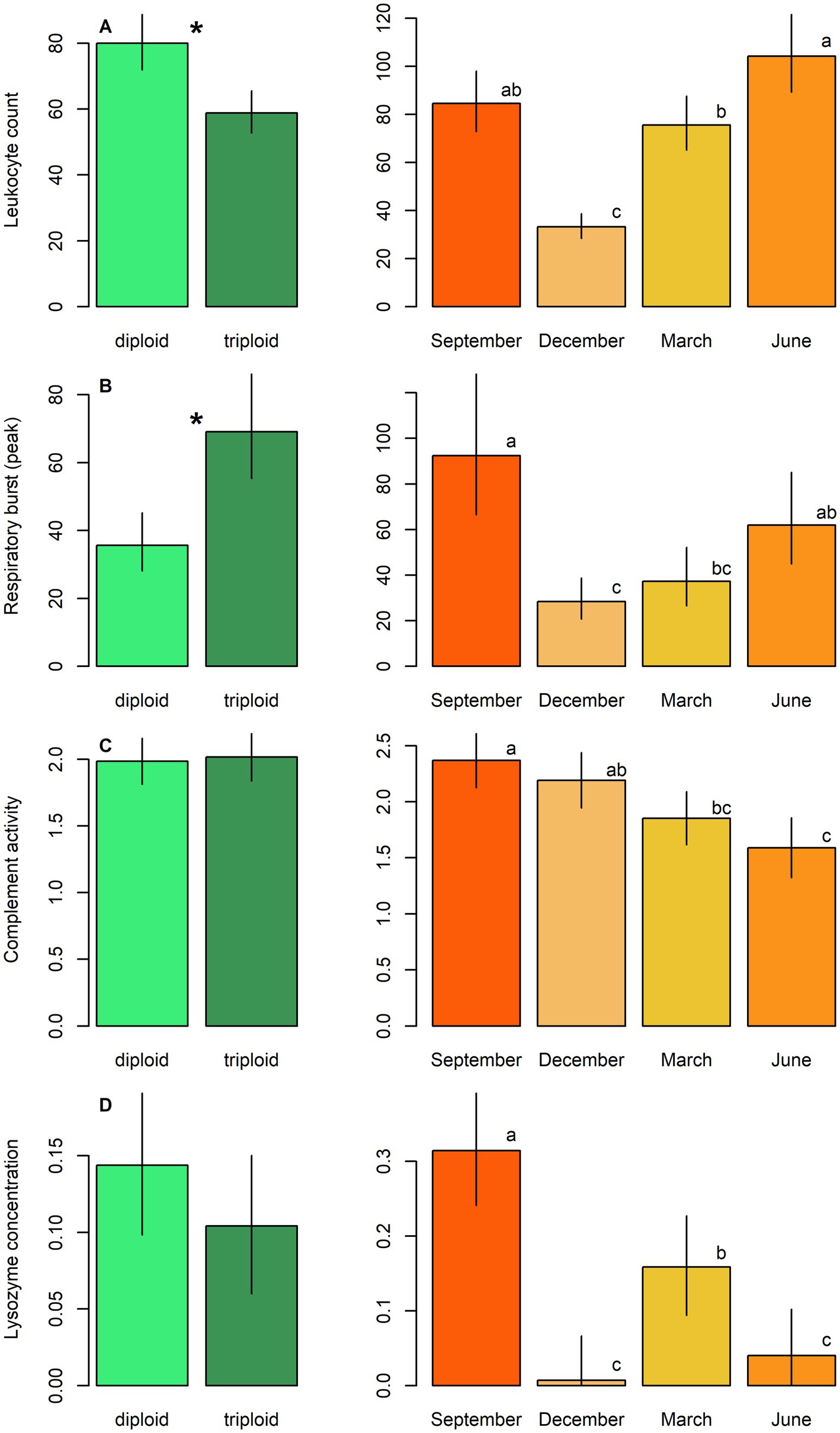
Figure 5. Immunological parameters, including: (A) Leukocyte count, (B) respiratory burst peak, (C) complement activity, and (D) lysozyme concentration in diploid and triploid tench (green bars), and their seasonal fluctuation (orange bars). Ploidy bars with *differ significantly from each other. Season bars with the different letters differ significantly from each other.
Seasonal variation was evident across all measured parameters. Erythrocyte count, haematocrit, and haemoglobin peaked in September, while leukocrit was highest in June and lowest in December (Figure 3). Glucose levels peaked in December, and 11-ketotestosterone concentrations were highest in March and June (Table 5; Figure 4). Respiratory burst peak, lysozyme concentration, and complement activity reached their maximum in September (Figures 5B–D). The lowest values of respiratory burst peak and leukocyte count were observed in December, while leukocyte counts peaked in June (Table 5; Figures 5A,B).
3.4 The associations between parasite load and host parameters
Total parasite abundance was positively associated with GSI in both sexes, regardless of ploidy. However, this association became insignificant in triploid females following Bonferroni correction (Table 6). In diploids, parasite abundance was negatively associated with glucose levels and lysozyme concentration (r = −0.437 and −0.380, respectively; both p < 0.001). Other physiological, biochemical, haematological, and immunological parameters showed no consistent associations with parasite abundance, or their significance diminished after correction for multiple analyses - specifically HSI, haemoglobin, and complement activity in diploids, and lysozyme concentration in triploids (Table 6).
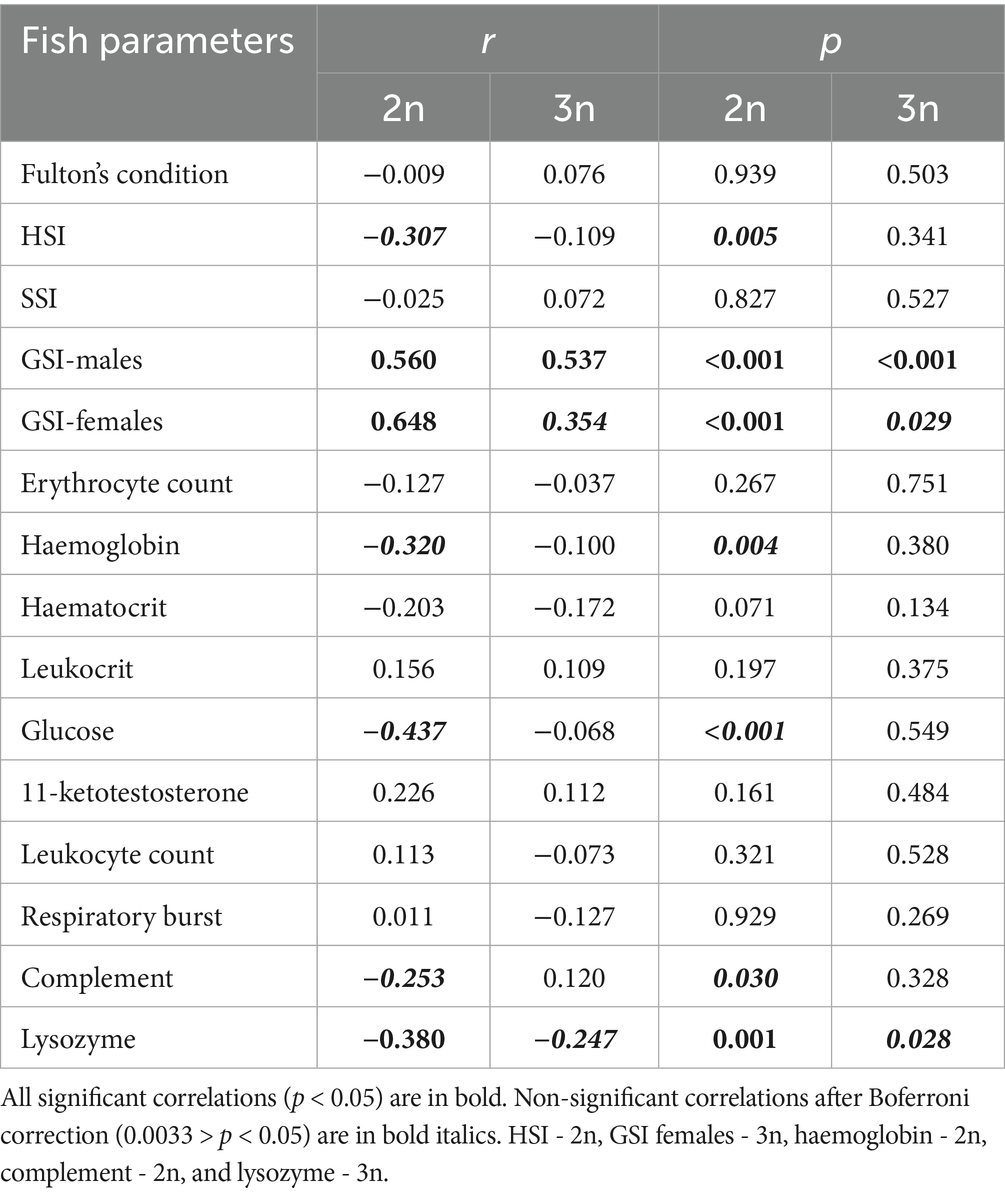
Table 6. Spearman correlation coefficients (r) and their respective p-values for correlations between total parasite abundance and fish physiological (Fulton’s condition factor, hepatosomatic index (HSI), splenosomatic index (SSI), gonadosomatic index (GSI) in males and females); haematological (erythrocyte count, haemoglobin content, haematocrit, leukocrit); biochemical (glucose level in plasma, 11-ketotestosterone concentration in males); and immunological (leukocyte count, respiratory burst peak, complement activity, lysozyme concentration) parameters, calculated separately for diploids (2n) and triploids (3n).
The abundance of A. tincae tended to increase with GSI in both sexes and decrease with HSI and glucose levels in diploids. Abundance of G. tincae tended to increase with HSI in both sexes, while negative associations were found with haematocrit and haemoglobin in triploids, and with leukocrit and respiratory burst peak in diploids, after Bonferroni corrections. No significant associations were found between abundance of V. campylancristrota and host parameters following correction. In contrast, abundance of Myxosporea spp. (present on the gills) was negatively correlated with leukocyte counts in both diploid a triploid tench (Supplementary Table S1).
4 Discussion
Investigating parasite load in diploid and triploid individuals of commercially important fish is a crucial aspect of assessing fish health in aquaculture. Artificially induced polyploid fish are of particular interest due to their enhanced growth performance, improved organoleptic qualities, and greater economic viability across diverse farming environments (10). Polyploidization is associated with changes in cell morphology and physiology, which can affect fish fitness and resistance to pathogens (2, 3, 54). Although polyploids are expected to exhibit advantages in coping with environmental stressors, including pathogens, empirical evidence remains inconclusive [e.g., Osnas and Lively (25), Hakoyama et al. (26), Nuismer and Thompson (29), Ozerov et al. (55), and Van de Peer et al. (56)].
In this study, we examined the relationship between ploidy level and metazoan parasite infection in tench over a one-year period, encompassing four seasonal sampling points: autumn, winter, early spring, and summer. Our findings indicate that ploidy status plays an important role in parasite infection in tench aquaculture. Induced triploids tended to have lower parasitic load than amphimictic diploids over the course of the year, primarily due to higher infection rates of diploids by the most abundant host-specific digenean parasite A. tincae, which peaked in summer. Adult A. tincae are found exclusively in digestive tract of tench; previous reports of its occurrence in other fish species likely stem from misidentification or confusion with other species of the genus Parasymphylodora that infect different fish hosts (43, 57). Tench serve as definitive hosts, becoming infected through ingestion of pulmonate snails carrying rediae with mature cercariae (43).
Conversely, triploid tench reached higher prevalence of generalist endoparasite V. campylancristrota across all seasons, with a summer peak. However, the abundance remained low throughout the year. This cestode parasitizes a broad range of freshwater fish, which act as second intermediate hosts. Infection occurs via ingestion of cyclopoid copepods, with the plerocercus stage residing in the fish gall bladder until consumed by the definitive bird host (58).
The contrasting infection patterns of A. tincae and V. campylancristrota between diploid and triploid tench suggest that polyploidy may exert non-uniform effects on host–parasite interactions. A similar phenomenon was observed by Nuismer and Thompson (29) in Heuchera grossulariifolia Rydb., where polyploid plants were less attacked by one phytophagous insect species but more susceptible to others.
Several mechanisms may explain the observed difference in infection by the two most common parasite species. Increased ploidy may not uniformly enhance resistance; it can either strengthen or weaken host defenses depending on allele interactions between hosts and parasites. Triploid fish may keep three distinct alleles per single locus, providing them with higher levels of heterozygosity what can potentially accrue associated fitness benefits over diploid fish (1). Greater heterozygosity may impede parasite evasion of host recognition, potentially conferring increased resistance (59). Thus, we can hypothesize that the reduced susceptibility of triploid tench to A. tincae infection may be linked to their higher allelic diversity and expression of immune-related genes, although these aspects were not directly assessed in this study. To the best of our knowledge, no any study yet dealt directly with the assessment of heterozygosity in triploid tench but several studies on farmed crustacean, shellfish and fish species revealed higher heterozygosity levels for induced triploids if compared to their diploid counterparts, with the magnitude and/or detectability of triploidy-induced changes in heterozygosity depending greatly on the genetic background of the population studied, as recently reviewed, e.g., by Flajšhans et al. (60). Furthermore, host–parasite interactions often involve tight genetic specificity, which may favour polyploid advantages against coevolving parasites (25), such as host-specific species like A. tincae. Beyond genetic factors, host specificity of parasites is also shaped by the ecological, behavioral, physiological, and biochemical traits of hosts (23).
Physiological and feeding behavioral differences between diploid and triploid tench may further contribute to variation in parasite abundance. Triploid organisms are generally characterized by a reduced number of cells that are proportionally larger in size (61), a phenomenon well documented also in fish species (62). Reduction in cell numbers in central nervous system and reduced hormone levels of hormones in triploid fish could result in diminished responsiveness to environmental stimuli leading to expression of altered behavior (2, 15). Triploids have also been reported to be less aggressive than diploids, potentially affecting competitive feeding success (22, 63). In our study, both ploidy groups were reared in the same pond under conditions of potential food competition.
The other two most abundant parasite taxa were the ectoparasitic host-specific monogenean G. tincae and undetermined Myxosporea spp. Gyrodactylus parasites, due to their hyperviviparity and rapid reproduction (a single worm can produce thousands of progeny), may pose a significant threat in aquaculture (64–66). In polyploid fish, triploid Atlantic salmon were found to be more susceptible to Gyrodactylus salaris Malmberg, 1957, probably due to impaired complement-dependent immune responses (55). In our study, unlike A. tincae and V. campylancristrota, the prevalence and abundance of G. tincae and Myxosporea spp. peaked in winter and early spring (or winter only), with no significant differences between diploid and triploid tench. Notably, the maximum intensity of infection of the host-specific A. tincae was 30 times higher than that of G. tincae. Piačková and Flajšhans (33) reported a higher prevalence of undetermined Gyrodactylus spp. in meiotic gynogenic tench, with no apparent difference between induced triploids and amphimictic diploids, consistent with our findings. However, their conclusions were based solely on visual estimates of parasite prevalence as their study did not include data on intensity of infection. In contrast, our study supports this observation by providing original quantitative data of metazoan parasite load.
Previous studies on polyploid fish [e.g., Benfey (2), Ballarin et al. (18), Peruzzi et al. (19), Levy-Pereira et al. (67), and Rożyński et al. (68)], including those focussing on tench (11, 16, 17, 69), have highlighted that ploidy can alter fish condition, physiology, haematological profile, and immune function. Although induced triploid tench generally exhibit faster growth and improved weight gain [as reviewed by Flajšhans et al. (11)], our study did not reveal differences in body condition factor, energy reserves (measured by HSI), or immunocompetence (expressed by SSI). However, reproductive investment (measured by GSI) followed the trend described in previous tench studies, with diploid females and males showing higher GSI values compared to their triploid counterparts (12, 70). Regarding reproductive potential, triploid females are sterile, while triploid males may produce aneuploid spermatozoa with variable DNA content capable of initiating embryo development (11). Histological studies have revealed underdeveloped or retarded ovaries in females and testes in males (13, 70).
Haematological parameters of 6 + year-old induced triploid and amphimictic diploid tench were previously analyzed by Tolarová et al. (16), who reported lower erythrocyte counts (compensated by larger erythrocyte size), haemoglobin, and haematocrit in triploids compared to diploids, suggesting reduced oxygenation capacity and potentially less effective immune response (11). Similar findings have been reported in other studies on farmed tench (17, 33). A comparable pattern was observed for leukocyte counts in our study and has also been previously documented in various triploid fish (2, 19, 69, 71).
However, we found an opposite trend for leukocrit, with significantly higher values in triploids than in diploids. Since leukocrit reflects the volume fraction of white blood cells (leukocytes) in whole blood, this may indicate that larger leukocytes in triploids contribute more to leukocrit values than the higher leukocyte numbers observed in diploids. This distinction should be considered when interpreting leukocrit as an indicator or immune status, particularly in response to pathogen infection.
In addition to the leukocyte count and leukocrit, we analyzed parameters of innate immunity, including respiratory burst, complement activity, and lysozyme concentration. As reported by Tolarová et al. (16), only respiratory burst, a rapid increase in oxygen consumption by immune cells such as neutrophils and macrophages important for combating pathogens, varied with ploidy status in tench. A study of turbot (Psetta maxima L., 1758) by Budiño et al. (72) found higher respiratory burst and phagocytic activity in triploid fish, although the number of neutrophils was higher in diploids, resulting in comparable respiratory burst activity and the phagocytosis per microliter of blood. Similarly, Chalmers et al. (73) showed that triploid Atlantic salmon exhibited higher respiratory burst activity but lower blood cell counts following furunculosis vaccination, suggesting that reduced cell numbers may be compensated by increased cellular activity. In contrast, a study of Astyanax altiparanae Garutti and Britski, 2000 showed that triploid induction led to reduced phagocytic capacity (67).
Our findings align with those of Budiño et al. (72) and Chalmers et al. (73), indicating that many components of the innate immune system function similarly in diploid and triploid tench, and that lower leukocyte counts in triploids may be offset by higher cellular activity. Differences in physiology, haematology, and immunity between triploid and diploid fish, including those reported in our study, may thus be explained by (1) increased heterozygosity of triploids, (2) a reduced number of larger cells in key organs (brain, retina, kidney, liver, testes, and ovaries) and tissues (blood, cartilage, muscle, epithelium), and (3) impaired gametogenesis as suggested by Benfey (2) and Maxime (74).
Although we anticipated associations between parasite infection and fish condition, haematological profile, physiology, and immunity in diploid and triploid tench, the observed relationships were generally weak. This may be due to overall low parasite abundance and species richness in tench. Moreover, significant seasonal and ploidy effects on parasite infection, along with variable responses among the four most common parasite species, may obscure correlations between overall parasite load and parameters linked to condition, physiology or immunity. Nonetheless, trends in associations between parasite infection and health-related traits were more pronounced in diploid tench than in triploids. This may be attributable to presumed higher heterozygosity of triploids, physiological differences related to cell size, and host–parasite coevolutionary interactions between diploid fish and their associated parasites. Reproductive investment, expressed by GSI, in both diploid and triploid females and males significantly correlated with total parasite abundance and with abundance of A. tincae. These correlations were stronger for diploids, suggesting a greater impact of this trematode parasite (likely weakened immunity) when increasing reproductive investment. This finding is consistent with previous studies on gonad development, which reported delayed or reduced gonadal growth in triploid tench (13, 30, 70).
Correlations between haematological and immune parameters and total parasite load or abundance of the four most frequent parasite species in tench were generally weak and often disappeared after correction for multiple testing. Therefore, these correlations should be interpreted with caution. Published studies on various fish species under different experimental conditions have yielded mixed results regarding immune function in diploid versus triploid fish. In some cases, triploid induction has been associated with immune impairments and reduced disease resistance as indicated by lower expression of immunity-related genes (IL-1 and TGF-β) in induced triploid A. altiparanae compared to diploids (67). A recent study by Jablonska et al. (75) found differences in the TLR gene expression between diploid and triploid sterlet (Acipenser ruthenus L., 1758), with TLRs being key components of the innate immune system responsible for recognizing pathogen-associated molecular patterns.
In contrast, other studies suggest that fish ploidy does not affect the pathogen susceptibility or innate serum immune responses. For example, no differences were found in the manifestation or severity of Neoparamoeba perurans Young, Crosbie, Adams, Nowak and Morrison, 2007 infection (causative agent of amoebic gill disease) in diploid and triploid Atlantic salmon (76). Similarly, a study on natural triploid (gynogenetic females) and diploid specimens of gibel carp (Carassius gibelio) showed that triploids were unaffected by parasite assemblage composition in terms of innate immunity and physiology (77). However, another study using C. gibelio under experimental infection with Diplostomum pseudospathaceum Niewiadomska, 1984 revealed a limited ability of triploid gynogenetic fish to cope with higher parasite infection, likely due to impaired immune gene activation (28). We propose that further studies on tench immunity should focus on differential gene expression and quantification of key immune genes, ideally under controlled experimental infections with a single parasite species, to clarify the role of ploidy in parasite susceptibility and resistance.
5 Conclusion
In conclusion, our study demonstrated that the ploidy status in tench affected metazoan parasite infection, reproductive investment (measured by GSI), haematological parameters, and immune function (specifically, leukocyte count and phagocyte activity expressed by respiratory burst). We revealed that triploids tended to have lower parasite load than diploids throughout the year. Nevertheless, our findings indicate that polyploidy exerts non-uniform effects on infection by different parasite species. Triploids exhibited lower abundance of the host-specific adult trematode A. tincae, yet significantly higher abundance of the larval cestode V. campylancristrota. Highly prevalent and moderately abundant monogenean specialist G. tincae was found in triploids and diploids at the similar intensity of infection. The other parasite species exhibited generally low infection parameters. We also identified an association between parasite infection and gonadosomatic condition, indicating that increased reproductive investment may elevate infection risk, particularly in diploid tench. Our results suggest potential effects of the three most abundant parasite taxa (Gyrodactylus tincae, Asymphylodora tincae and Myxosporea spp.) on tench immunity. The observed differences in parasite infection between triploid and diploid tench and the associations between parasite load and condition- and health-related traits may be explained by several factors: (1) presumed higher heterozygosity in triploids, (2) physiological differences related to cell size and number of cells in key organs and tissues, (3) variation in feeding performance, and (4) host–parasite coevolutionary interactions between diploid tench and their associated parasites.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Ethics statement
The animal study was approved by the Animal Care and Use Committee of the Faculty of Science, Masaryk University, Brno (Permit No. 031/2011). The method of fish sampling and killing complied with the legal requirements of the Czech Republic (§ 7 Law No. 114/1992 on the Protection of Nature and Landscape and § 6, 7, 9 and 10 Regulation No. 419/2012 on the Care, Breeding and Use of Experimental Animals). The study was conducted in accordance with the local legislation and institutional requirements.
Author contributions
AŠ: Conceptualization, Funding acquisition, Investigation, Project administration, Writing – original draft, Writing – review & editing. MD: Investigation, Methodology, Validation, Writing – original draft. PH: Methodology, Validation, Writing – review & editing. MJ: Formal analysis, Validation, Writing – review & editing. MF: Investigation, Resources, Writing – review & editing. MO: Investigation, Visualization, Writing – original draft, Writing – review & editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. The study was funded by the Czech Science Foundation, Projects No. G524/07/0188 and no. P505/12/G112, and supported by the Ministry of Education, Youth and Sports, Operational Programme Johannes Amos Comenius, Project No. CZ.02.01.01/00/23_021/0012616 and “Biodiversity” no. CZ.02.1.01/0.0/0.0/16_025/0007370.
Acknowledgments
The authors would like to thank Radim Blažek, Kateřina Francová, Monika Mendlová, Petra Navrátilová, Naďa Musilová, Eva Řehulková and Mária Seifertová (Masaryk University, Brno, Czechia) for their assistance with parasite collection. For the help with cestode determination we thank Roman Kuchta (Biological Centre, Czech Academy of Sciences, Czechia) and Mikuláš Oros (Parasitological Institute SAS, Košice, Slovakia).
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The authors declare that no Gen AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fvets.2025.1686708/full#supplementary-material
References
1. Otto, SP, and Whitton, J. Polyploid incidence and evolution. Annu Rev Genet. (2000) 34:401–37. doi: 10.1146/annurev.genet.34.1.401
2. Benfey, TJ. The physiology and behaviour of triploid fishes. Rev Fish Sci. (1999) 7:39–67. doi: 10.1080/10641269991319162
3. King, KC, Seppälä, O, and Neiman, M. Is more better? Polyploidy and parasite resistance. Biol Lett. (2012) 8:598–600. doi: 10.1098/rsbl.2011.1152
4. Leggatt, RA, and Iwama, GK. Occurrence of polyploidy in the fishes. Rev Fish Biol Fish. (2003) 13:237–46. doi: 10.1023/B:RFBF.0000033049.00668.fe
5. Cunha, C, Doadrio, I, Abrantes, J, and Coelho, MM. The evolutionary history of the allopolyploid Squalius alburnoides (Cyprinidae) complex in the northern Iberian Peninsula. Heredity. (2011) 106:100–12. doi: 10.1038/hdy.2010.70
6. Havelka, M, Hulák, M, Ráb, P, Rábová, M, Lieckfeldt, D, Ludwig, A, et al. Fertility of a spontaneous hexaploid male Siberian sturgeon, Acipenser baerii. BMC Genet. (2014) 15:5. doi: 10.1186/1471-2156-15-5
7. Schreier, AD, May, B, and Gille, DA. Incidence of spontaneous autopolyploidy in cultured populations of white sturgeon, Acipenser transmontanus. Aquaculture. (2013) 416:141–5. doi: 10.1016/j.aquaculture.2013.09.012
9. Manan, H, Noor Hidayati, AB, Lyana, NA, Amin-Safwan, A, Ma, H, Kasan, NA, et al. A review of gynogenesis manipulation in aquatic animals. Aquac Fish. (2022) 7:1–6. doi: 10.1016/j.aaf.2020.11.006
10. Piferrer, F, Beaumont, A, Falguière, J-C, Flajšhans, M, Haffray, P, and Colombo, L. Polyploid fish and shellfish: production, biology and applications to aquaculture for performance improvement and genetic containment. Aquaculture. (2009) 293:125–56. doi: 10.1016/j.aquaculture.2009.04.036
11. Flajšhans, M, Gela, D, Kocour, M, Buchtová, H, Rodina, M, Pšenička, M, et al. A review on the potential of triploid tench for aquaculture. Rev Fish Biol Fish. (2010) 20:317–29. doi: 10.1007/s11160-009-9144-z
12. Buchtová, H, Svobodová, Z, Flajšhans, M, and Vorlová, L. Analysis of growth, weight and relevant indices of diploid and triploid population of tench Tinca tinca (Linnaeus 1758). Aquac Res. (2003) 34:719–26. doi: 10.1046/j.1365-2109.2003.00874.x
13. Buchtová, H, Smutná, M, Vorlová, L, Svobodová, Z, and Flajšhans, M. Fatty acid composition of diploid and triploid populations of tench (Tinca tinca L.). Acta Vet Brno. (2004) 73:235–45. doi: 10.2754/avb200473020235
14. Buchtová, H, Smutná, M, Vorlová, L, Svobodová, Z, and Flajšhans, M. Amino acid composition of muscle proteins of diploid and triploid tench (Tinca tinca, Linnaeus 1758). Acta Vet Brno. (2005) 74:329–37. doi: 10.2754/avb200574030329
15. Tiwary, BK, Kirubagaran, R, and Ray, AK. The biology of triploid fish. Rev Fish Biol Fish. (2004) 14:391–402. doi: 10.1007/s11160-004-8361-8
16. Tolarová, S, Dávidová, M, Šimková, A, Flajšhans, M, and Hyršl, P. The seasonal changes of innate immunity of tench, Tinca tinca (L.) with different ploidy level. Aquaculture. (2014) 432:46–52. doi: 10.1016/j.aquaculture.2014.04.006
17. Svobodová, Z, Kolářová, J, and Flajšhans, M. The first findings of the differences in complete blood count between diploid and triploid tench, Tinca tinca L. Acta Vet Brno. (1998) 67:243–8. doi: 10.2754/avb199867040243
18. Ballarin, L, Dall’Oro, M, Bertotto, D, Libertini, A, Francescon, A, and Barbaro, A. Haematological parameters in Umbrina cirrosa (Teleostei, Sciaenidae): a comparison between diploid and triploid specimens. Comp Biochem Physiol A. (2004) 138:45–51. doi: 10.1016/j.cbpb.2004.02.019
19. Peruzzi, S, Varsamos, S, Chatain, B, Fauvel, C, Menu, B, Falguière, JC, et al. Haematological and physiological characteristics of diploid and triploid sea bass, Dicentrarchus labrax L. Aquaculture. (2005) 244:359–67. doi: 10.1016/j.aquaculture.2004.11.028
20. Vetešník, L, Halačka, K, and Šimková, A. The effect of ploidy and temporal changes in the biochemical profile of gibel carp (Carassius gibelio): a cyprinid fish species with dual reproductive strategies. Fish Physiol Biochem. (2013) 39:171–80. doi: 10.1007/s10695-012-9688-z
21. Šimková, A, Košař, M, Vetešník, L, and Vyskočilová, M. MHC genes and parasitism in Carassius gibelio, a diploid-triploid fish species with dual reproduction strategies. BMC Evol Biol. (2013) 13:122. doi: 10.1186/1471-2148-13-122
22. O’Keffe, RA, and Benfey, TJ. The feeding response of diploid and triploid Atlantic salmon and brook trout. J Fish Biol. (1997) 51:989–97. doi: 10.1111/j.1095-8649.1997.tb01537.x
23. Poulin, R. Evolutionary ecology of parasites: From individuals to communities. London: Chapman & Hall (1998).
24. Spurgin, LG, and Richardson, DS. How pathogens drive genetic diversity: MHC, mechanisms and misunderstandings. Proc R Soc Lond B. (2010) 277:979–88. doi: 10.1098/rspb.2009.2084
25. Osnas, EE, and Lively, CM. Host ploidy, parasitism and immune defence in a coevolutionary snail-trematode system. J Evol Biol. (2006) 19:42–8. doi: 10.1111/j.1420-9101.2005.00994.x
26. Hakoyama, H, Nishimura, T, Matsubara, N, and Iguchi, K. Difference in parasite load and nonspecific immune reaction between sexual and gynogenetic forms of Carassius auratus. Biol J Linn Soc. (2001) 72:401–7. doi: 10.1006/bijl.2000.0507
27. Langston, AL, Johnstone, R, and Ellis, AE. The kinetics of the hypoferraemic response and changes in levels of alternative complement activity in diploid and triploid Atlantic salmon, following injection of lipopolyssacharide. Fish Shellfish Immunol. (2001) 11:333–45. doi: 10.1006/fsim.2000.0319
28. Fuad, MMH, Tichopád, T, Ondračková, M, Civánová Křížová, K, Seifertová, M, Voříšková, K, et al. Trematode Diplostomum pseudospathaceum inducing differential immune gene expression in sexual and gynogenetic gibel carp (Carassius gibelio): parasites facilitating the coexistence of two reproductive forms of the invasive species. Front Immunol. (2024) 15:1392569. doi: 10.3389/fimmu.2024.1392569
29. Nuismer, SL, and Thompson, JN. Plant polyploidy and non-uniform effects on insect herbivores. Proc R Soc Lond B. (2001) 268:1937–40. doi: 10.1098/rspb.2001.1760
30. Flajšhans, M. Reproduction sterility caused by spontaneous triploidy in tench (Tinca tinca). Pol Arch Hydrobiol. (1997) 44:39–45.
31. Flajšhans, M, Kvasnička, P, and Ráb, P. Genetic studies in tench (Tinca tinca L.): high incidence of spontaneous triploidy. Aquaculture. (1993) 110:243–8. doi: 10.1016/0044-8486(93)90372-6
32. Svobodová, Z, and Kolářová, J. A review of the diseases and contaminant related mortalities of tench (Tinca tinca L.). Vet Med. (2004) 49:19–34. doi: 10.17221/5671-VETMED
33. Piačková, V, and Flajšhans, M. Long-term examination of health conditions in monoculture of communally tested amphimictic diploid, diploid gynogenic and triploid tench, Tinca tinca (L.). Aquac Int. (2006) 14:43–59. doi: 10.1007/S10499-005-9013-2
34. Flajšhans, M, Linhart, O, and Kvasnička, P. Genetic studies of tench (Tinca tinca L.): induced triploidy and tetraploidy and first performance data. Aquaculture. (1993) 113:301–12. doi: 10.1016/0044-8486(93)90401-J
35. Pravda, D, and Svobodová, Z. Haematology of fishes In: J Doubek, J Bouda, M Doubek, M Fürll, Z Knotková, and S Pejřilová, et al., editors. Veterinary Haematology. Brno: Noviko (2003). 381–97.
36. Rohlenová, K, Morand, S, Hyršl, P, Tolarová, S, Flajšhans, M, and Šimková, A. Are fish immune systems really affected by parasites? An immunoecological study of common carp (Cyprinus carpio). Parasit Vectors. (2011) 4:120. doi: 10.1186/1756-3305-4-120
37. Vindelov, LL, and Christensen, IJ. Review of techniques and results obtained in one laboratory by an integrated system of methods designed for routine clinical flow cytometric DNA analysis. Cytometry. (1990) 11:753–70. doi: 10.1002/cyto.990110702
39. Malmberg, G. The excretory systems and the marginal hooks as a basis for the systematics of Gyrodactylus (Trematoda, Monogenea). Ark Zool. (1970) 23:1–235.
40. Georgiev, B, Biserkov, V, and Genov, T. In toto staining method for cestodes with iron acetocarmine. Helminthologia. (1986) 23:279–81.
41. Bush, AO, Lafferty, KD, Lotz, JM, and Shostak, AW. Parasitology meets ecology onits own terms: Margolis et al. revisited. J Parasitol. (1997) 83:575–83. doi: 10.2307/3284227
42. Gusev, AV. Monogenea In: ON Bauer, editor. Key to the parasites of the freshwater fish fauna of the USSR, vol. 2. Leningrad: Nauka (1985)
43. Našincová, V, and Scholz, T. The life cycle of Asymphylodora tincae (Modeer 1790) (Trematoda, Monorchiidae) – a unique development in monorchiid trematodes. Parasitol Res. (1994) 80:192–7. doi: 10.1007/BF00932673
44. Moravec, F. Checklist of the metazoan parasites of fishes of the Czech Republic and the Slovak Republic. Prague: Academie (2001).
45. Svobodová, Z, Pravda, D, and Paláčková, J. Universal methods of haematological investigations in fish. Edice Metodik: VÚRH Vodňany, Czech Republic (1986).
46. Hrubec, TC, and Smith, SA. Hematology of fish In: BV Feldman, JG Zinkl, and NC Jain, editors. Schalm's veterinary hematology. 5th ed. Ames, Iowa: Blackwell Publishing (2000). 1120–5.
47. Buchtíková, S, Šimková, A, Rohlenová, K, Flajšhans, M, Lojek, A, Lilius, E-M, et al. The seasonal changes in innate immunity of the common carp (Cyprinus carpio). Aquaculture. (2011) 318:169–75. doi: 10.1016/j.aquaculture.2011.05.013
48. Poisot, T, Šimková, A, Hyršl, P, and Morand, S. Interactions between immunocompetence, somatic condition and parasitism in the chub Leuciscus cephalus in early spring. J Fish Biol. (2009) 75:1667–82. doi: 10.1111/j.1095-8649.2009.02400.x
49. Zuur, A, Ieno, EN, Walker, N, Saveliev, AA, and Smith, GM. Mixed effects models and extensions in ecology with R. New York: Springer (2009).
50. Hothorn, T, Bretz, F, and Westfall, P. Simultaneous inference in general parametric models. Biom J. (2008) 50:346–63. doi: 10.1002/bimj.200810425
51. R Core Team. R: A language and environment for statistical computing. R foundation for statistical computing. Vienna, Austria (2021).
52. Venables, WN, and Ripley, BD. Modern applied statistics with S. Fourth edition. New York: Springer (2002).
53. Lenth, RV. Emmeans: Estimated Marginal Means, aka Least-Squares Means. R package version 1.7.2. (2022). Available online at: https://CRAN.R-project.org/package=emmeans.
54. Oswald, BP, and Nuismer, SL. Neopolyploidy and pathogen resistance. Proc R Soc B. (2007) 274:2393–7. doi: 10.1098/rspb.2007.0692
55. Ozerov, MY, Lumme, J, Päkk, P, Rintamäki, P, Ziętara, MS, Barskaya, Y, et al. High Gyrodactylus salaris infection rate in triploid Atlantic salmon Salmo salar. Dis Aquat Org. (2010) 91:129–36. doi: 10.3354/dao02242
56. Van de Peer, Y, Ashman, T-L, Soltis, PS, and Soltis, DE. Polyploidy: an evolutionary and ecological force in stressful times. Plant Cell. (2021) 33:11–26. doi: 10.1093/plcell/koaa015
57. Petkevičiūtė, R, Stunžėnas, V, and Stanevičiūtė, G. Diversity of European lissorchiid trematodes from fish and snail hosts with comments on the validity of the genus Parasymphylodora Szidat, 1943. J Helminthol. (2022) 96:e67. doi: 10.1017/S0022149X22000542
58. Scholz, T, Bray, RA, Kuchta, R, and Řepová, R. Larvae of gryporhynchid cestodes (Cyclophyllidea) from fish: a review. Folia Parasitol. (2004) 51:131–52. doi: 10.14411/fp.2004.018
59. Nuismer, SL, and Otto, SP. Host–parasite interactions and the evolution of ploidy. Proc Natl Acad Sci USA. (2004) 101:11036–9. doi: 10.1073/pnas.0403151101
60. Flajšhans, M, Kašpar, V, Prchal, M, Musatova, I, Prášková, E, Kašpar, J, et al. The assessment of the effects of genomic heterozygosity and sterility on the performance of triploid brook trout Salvelinus fontinalis. Front Anim Sci. (2025) 6:1481117. doi: 10.3389/fanim.2025.1481117
61. Leung, K, van de Zande, L, and Beukeboom, LW. Effects of polyploidization and their evolutionary implications are revealed by heritable polyploidy in the haplodiploid wasp Nasonia vitripennis. PLoS One. (2023) 18:e0288278. doi: 10.1371/journal.pone.0288278
62. Goo, IB, Im, JH, Gil, HW, Lim, SG, and Park, IS. Comparison of cell and nuclear size difference between diploid and induced triploid in marine medaka, Oryzias dancena. Dev Reprod. (2015) 19:127–34. doi: 10.12717/DR.2015.19.3.127
63. Carter, CG, McCarthy, ID, Houlihan, DF, Johnstone, R, Walsingham, MV, and Mitchell, AI. Food consumption, feeding behaviour, and growth of triploid and diploid Atlantic salmon, Salmo salar L., parr. Can J Zool. (1994) 72:609–17. doi: 10.1139/z94-083
64. Johnsen, BO, and Jensen, AJ. The Gyrodactylus story in Norway. Aquaculture. (1991) 98:289–302. doi: 10.1016/0044-8486(91)90393-L
65. Bakke, TA, Cable, J, and Harris, PD. The biology of gyrodactylid monogeneans: the "Russian-doll killers". Adv Parasitol. (2007) 64:161. doi: 10.1016/S0065-308X(06)64003-7
66. Buchmann, K. “Gyrodactylus salaris and Gyrodactylus derjavinoides” in Fish parasites: Pathobiology and protection, eds. P.T.K. Woo and K. Buchmann (CABI: Digital Library), (2012) 193–208.
67. Levy-Pereira, N, Carriero, MM, Yasui, GS, Meira, CM, de Moro Sousa, RL, Maia, AAM, et al. Effects of triploid induction on innate immunity and hematology in Astyanax altiparanae. Fish Shellfish Immunol. (2021) 116:12–8. doi: 10.1016/j.fsi.2021.05.004
68. Rożyński, R, Dobosz, S, Rożyński, M, and Ocalewicz, K. The effect of triploidy on gonadal development, hematology and biochemistry in the European Grayling (Thymallus thymallus). Animals. (2025) 15:481. doi: 10.3390/ani15040481
69. Svobodová, Z, Flajšhans, M, Kolářová, J, Modrá, H, Svoboda, M, and Vajcová, V. Leukocyte profile of diploid and triploid tench, Tinca tinca L. Aquaculture. (2001) 198:159–68. doi: 10.1016/S0044-8486(00)00587-1
70. Linhart, O, Rodina, M, Flajšhans, M, Mavrodiev, N, Nebesářová, J, Gela, D, et al. Studies on sperm of diploid and triploid tench (Tinca tinca (L.)). Aquac Int. (2006) 14:9–25. doi: 10.1007/s10499-005-9010-5
71. Benfey, TJ, and Biron, M. Acute stress response in triploid rainbow trout (Oncorhynchus mykiss) and brook trout (Salvelinus fontinalis). Aquaculture. (2000) 184:167–76. doi: 10.1016/S0044-8486(99)00314-2
72. Budiño, B, Cal, RM, Piazzon, CM, and Jesús, L. The activity of several components of the innate immune system in diploid and triploid turbot. Comp Biochem Physiol A Mol Integr Physiol. (2006) 145:108–13. doi: 10.1016/j.cbpa.2006.05.007
73. Chalmers, L, Thompson, KD, Taylor, JF, Black, S, Migaud, H, North, B, et al. A comparison of the response of diploid and triploid Atlantic salmon (Salmo salar) siblings to a commercial furunculosis vaccine and subsequent experimental infection with Aeromonas salmonicida. Fish Shellfish Immunol. (2016) 57:301–8. doi: 10.1016/j.fsi.2016.08.049
74. Maxime, V. The physiology of triploid fish: current knowledge and comparison with tidploid fish. Fish Fisher. (2008) 9:67–78. doi: 10.1111/j.1467-2979.2007.00269.x
75. Jablonska, O, Duda, S, Gajowniczek, S, Nitkiewicz, A, and Fopp-Bayat, D. Toll-like receptor type 2 and 13 gene expression and immune cell profiles in diploid and triploid sterlets (Acipenser ruthenus): insights into immune competence in polyploid fish. Int J Mol Sci. (2025) 26:3986. doi: 10.3390/ijms26093986
76. Chalmers, L, Taylor, JF, Roy, W, Preston, AC, Migaud, H, and Adams, A. A comparison of disease susceptibility and innate immune response between diploid and triploid Atlantic salmon (Salmo salar) siblings following experimental infection with Neoparamoeba perurans, causative agent of amoebic gill disease. Parasitology. (2017) 144:1229–42. doi: 10.1017/S0031182017000622
Keywords: tench, ploidy, metazoan parasites, fish condition, physiology, immunity
Citation: Šimková A, Dávidová M, Hyršl P, Janáč M, Flajšhans M and Ondračková M (2025) Susceptibility to metazoan parasite infection in amphimictic diploid and induced triploid tench (Tinca tinca L., 1758): the role of parasites in fish aquaculture. Front. Vet. Sci. 12:1686708. doi: 10.3389/fvets.2025.1686708
Edited by:
Gustavo Viozzi, National University of Comahue, ArgentinaReviewed by:
Patricio Torres, Austral University of Chile, ChileMohamed Bessat, Alexandria University, Egypt
Copyright © 2025 Šimková, Dávidová, Hyršl, Janáč, Flajšhans and Ondračková. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Andrea Šimková, c2lta292YUBzY2kubXVuaS5jeg==