 Refaat Ras1,2*
Refaat Ras1,2* Amanallah El-Bahrawy3
Amanallah El-Bahrawy3 Adel Abdelkhalek4
Adel Abdelkhalek4 Florica Morariu5
Florica Morariu5 Ayman N. Elsayed6
Ayman N. Elsayed6 Doaa S. Nouh7
Doaa S. Nouh7 Anamaria Plesko8
Anamaria Plesko8 Marius Stelian Ilie8*
Marius Stelian Ilie8* Manar AbdelMageed9,10
Manar AbdelMageed9,10- 1Department of Microbiology and Parasitology, Faculty of Veterinary Medicine, Badr University in Cairo (BUC), Badr City, Cairo, Egypt
- 2Department of Parasitology, Faculty of Veterinary Medicine, Zagazig University, Zagazig, Egypt
- 3Department of Pathology, Faculty of Veterinary Medicine, University of Sadat City, Sadat City, Minoufyia, Egypt
- 4Department of Food Hygiene, Safety and Technology, Faculty of Veterinary Medicine, Badr University in Cairo (BUC), Badr City, Cairo, Egypt
- 5Department of Biotechnologies, University of Life Sciences “King Mihai I” from Timișoara, Timişoara, Romania
- 6Department of Zoology and Entomology, Faculty of Science (Boys), Al-Azhar University, Nasr City, Cairo, Egypt
- 7Department of Anatomy and Embryology, Faculty of Veterinary Medicine, Zagazig University, Zagazig, Egypt
- 8Department of Parasitology and Parasitic Disease, Faculty of Veterinary Medicine, University of Life Sciences “King Mihai I” from Timișoara, Timișoara, Romania
- 9Department of Pathology and Clinical Pathology, Faculty of Veterinary Medicine, Badr University in Cairo (BUC), Badr City, Cairo, Egypt
- 10Department of Pathology, Faculty of Veterinary Medicine, Zagazig University, Zagazig, Egypt
Introduction: Taenia pisiformis (T. pisiformis), the etiological agent of cysticercosis in rabbits, poses a considerable health risk to domestic lagomorphs and contributes to economic losses in rabbit farming. This study aimed to ascertain the prevalence, risk factors, and molecular characteristics of T. pisiformis in rabbits from three Egyptian regions: Badr City (Cairo Province), Sadat City (Monufia Province), and Assiut City (Assiut Province).
Methods: A total of 150 samples were collected from both home-raised (n = 77) and farm-raised (n = 73) rabbits from January 2024 to December 2024. T. pisiformis cysts were identified morphologically and histologically, with tissue samples processed using hematoxylin and eosin (H&E) staining. Molecular confirmation was performed using polymerase chain reaction (PCR) targeting the cytochrome oxidase subunit 1 (cox1) and NADH dehydrogenase subunit 1 (nad1) mitochondrial genes, followed by sequencing and phylogenetic analysis. Statistical associations between infection and risk factors (age, location, season, and management system) were evaluated using the chi-square test and Fisher’s exact test.
Results: The overall infection prevalence rate was 21.3% (32/150), which was significantly higher in home-raised rabbits (31/77, 40.2%) than in farm-raised rabbits (1/73, 1.3%; χ2 = 31.5, p < 0.001). Infection rates were also strongly linked to season, with fall and winter showing higher prevalence. Cysts were mostly found in the mesentery and varied in number (1–5 per rabbit). Morphologically, the cysts contained a scolex with distinctive features, including suckers and rostellar hooks. Histology showed a thick cyst wall and characteristic tissue structures. Molecular analysis confirmed the parasite as T. pisiformis, with sequence similarities ranging from 97.64 to 100%, indicating a close relationship to global sequences.
Discussion: These findings underscore the influence of management practices and seasonal factors on infection dynamics and highlight the importance of molecular tools in parasite surveillance.
1 Introduction
Cysticercus pisiformis (C. pisiformis), the larval form of Taenia pisiformis (T. pisiformis), is the most frequent cestode and is widely distributed worldwide, primarily affecting rabbits and hares (1). The lifecycle involves canids, specifically dogs and foxes, as definitive hosts. Rabbits acquire infection through the infection of feed contaminated with eggs shed in the feces of these definitive hosts (2). Following ingestion, the eggs hatch, and the hexacanth embryos subsequently penetrate the intestinal wall. Then, they migrate through the portal vein and differentiate into the larval stage or metacestode, specifically C. pisiformis, which appears as a fluid-filled cyst characterized by an armed scolex (3).
In Egypt, the majority of rabbit production (approximately 57%) is carried out by rural families who typically raise small flocks (4). Severe larval infection can result in hepatic damage in intermediate hosts, potentially leading to hepatitis, cirrhosis, gastrointestinal disturbances, and impaired immune responses. Consequently, this can lead to secondary bacterial infection and economic losses in the rabbit breeding industry due to their deleterious effects on rabbit health and productivity (5–7). Moreover, T. pisiformis infestation affects the behavioral and productive characteristics of rabbits, and obesity exacerbates the consequences of infection (8). In addition, infection reduces fecundity in rabbits, with infected does showing decreased embryo implantation and smaller embryo vesicle sizes compared to uninfected does (9). However, no zoonotic cases of C. pisiformis were recorded (10), but potential risks should be considered for future studies.
Several studies have reported cysticercosis in rabbits. In southern Spain, a study of 2,923 wild rabbits reported an overall prevalence of 2.8% (6). In contrast, a study in Poland reported a significantly higher prevalence of gastrointestinal parasites of 79.56% in slaughtered rabbits, with T. pisiformis being specifically found in 4.74% of the examined animals (11). Furthermore, in Mexico, the first formal report of infection with T. pisiformis metacestodes revealed a prevalence of approximately 70% (12). Although prevalence studies offer valuable insights into the geographical distribution and burden of T. pisiformis, a more comprehensive understanding of the parasite’s biology and epidemiology requires molecular approaches, particularly genetic characterization (13). Mitochondrial DNA is a highly powerful and reliable molecular marker for analyzing population structure and evolutionary history (14). The mitochondrial genes, such as cytochrome oxidase subunit 1 (cox1) and NADH dehydrogenase subunit 1 (nad1), have been widely used to investigate the genetics of taeniid cestodes, including their genetic origins, population boundaries, and intraspecific and interspecific variation (15–17).
Hence, in Egypt, raising rabbits in small backyard colonies has traditionally served as a means of supplementing household income. However, this practice has recently evolved into a distinct source of meat production. Rabbits are considered optimal animals for meat production because of their quick feed-to-meat conversion, short lifespan, and short pregnancy period (18, 19). With the increasing importance of rabbit farming as a commercial meat source, the impact of parasitic diseases such as T. pisiformis on productivity and animal welfare is becoming a growing concern. Although T. pisiformis is one of the most prevalent parasites that severely affect rabbit breeding in Egypt, there are no formal reports on the infection of rabbit populations with T. pisiformis metacestodes. There is only one study that was conducted in the Qena Governorate, Egypt, which identified T. pisiformis in domestic rabbits (20).
In addition to this isolated case, data on the prevalence and distribution of T. pisiformis in Egypt remain limited and geographically restricted, offering no nationwide perspective on its impact. Furthermore, the genetic variability of circulating T. pisiformis strains in Egypt has never been characterized using molecular tools, despite the value of such information in understanding transmission routes and informing targeted control measures. The lack of effective vaccines and antiparasitic medications has resulted in the poor control of parasitic infections at present. Consequently, integrating molecular diagnostics into routine surveillance could significantly enhance disease management in rabbit production systems.
However, to date, no studies have utilized mitochondrial gene markers to characterize T. pisiformis strains in Egypt, representing a significant gap in the molecular epidemiology of this parasite in the region.
Therefore, this study aimed to investigate the prevalence, determine the morphological and histopathological characteristics, and conduct a phylogenetic analysis of T. pisiformis in domestic rabbits in Egypt, using both traditional and molecular tools.
2 Materials and methods
2.1 Ethical approval
All procedures conducted in this study were approved by the Research Ethics Committee at the Faculty of Pharmacy, Badr University in Cairo, Cairo, Egypt (Protocol No. BUC-IACUC/VET/161/A/2024).
2.2 Sample collection
In this study, a total of 150 domestic rabbits were collected from three different locations: Badr City (Cairo Province), Sadat City (Monufia Province), and Assiut (Assiut Province). The examined animals included rabbits that had died from illness and were submitted for diagnostic evaluation, as well as apparently healthy rabbits reared within the same localities. All rabbits were consecutively received at the Department of Pathology and Clinical Pathology, School of Veterinary Medicine, Badr University in Cairo, Egypt, between January and December 2024. To ensure the reliability of parasitological and pathological examination, carcasses presented more than 12 h postmortem were excluded. Apparently healthy rabbits, which were submitted by veterinarians or owners due to suspected herd health concerns and as part of diagnostic investigations within the affected localities, were humanely euthanized to allow a thorough examination of internal parasites and associated lesions. Euthanasia was conducted in full accordance with international and institutional ethical guidelines using intravenous administration of an overdose of sodium pentobarbital (100 mg/kg). Death was confirmed by the absence of vital signs prior to decapitation.
During necropsy, a thorough examination of all organs was performed, and cysts indicative of T. pisiformis were carefully extracted and subjected to both macroscopic and microscopic morphological evaluation. Data on management systems were recorded, classifying rabbits as either home-raised (small-scale backyard rearing) or farm-raised (small-scale commercial farming). Age was also documented and considered a potential risk factor in epidemiological analysis.
2.3 Histological examination
Tissue specimens of suspected T. pisiformis cysts, liver, and other organ tissues were collected from necropsied rabbits and immediately fixed in 10% neutral-buffered formalin after which they were sequentially processed for the preparation of paraffin blocks. Subsequently, the blocks were cut into 4–5 μm thin sections and stained with hematoxylin and eosin (H&E) for histopathological assessment (21). Photomicrographs were obtained using a light microscope equipped with an AmScope MU1803-HS microscope digital camera.
2.4 Molecular identification of identified Cysticercus pisiformis
2.4.1 DNA extraction
DNA was extracted from seven cyst-positive isolates from tissues using the DNeasy Blood & Tissue Mini-Kit (Qiagen, Hilden, Germany) following the manufacturer’s recommendations (22, 23). Briefly, 25 mg of tissue was sectioned into small fragments and incubated with 20 μL of Proteinase K at 56 °C until complete tissue lysis occurred. Subsequently, 200 μL of AL buffer and absolute ethyl alcohol were added sequentially and homogenized via vortexing. The resulting homogeneous solution was then transferred to a silica column and centrifuged. Subsequently, the samples were purified and centrifuged according to the manufacturer’s protocol. The eluted DNA was stored at −20 °C for further analysis.
2.4.2 DNA amplification by PCR
Extracted DNA was amplified by conventional PCR using two specific primer sets targeting the cox1 and nad1 mitochondrial genes. For cox1, amplification was performed using the forward primer JB3 (5′-TTTTTTGGGCATCCTGAGGTTTAT −3′) and the reverse primer JB4.5 (5′-TAAAGAAAGAACATAATGAAAATG-3′) (24). The second set of primers targeting the nad1 gene were forward primer JP11 (5′-AGATTCGTAAGGGGCCTAATA-3′) and reverse primer JP12 (5′-ACCACTAACTAATTCACTTTC-3′) (25). Each PCR reaction was carried out in a total volume of 50 μL of mixture containing 25 μL of GeneDireX OnePCR™ Master Mix (Cat# MB203-0050), 1 μL of DNA template, 1 μL of each 10 μM the forward primer and the reverse primer, and 22 μL of nuclease-free water. The cycling conditions of PCR were the same for both genes and were set up as follows: an initial denaturation for 5 min at 94 °C, followed by 35 cycles of denaturation at 94 °C for 40 s, annealing at 50 °C for 1 min, extension at 72 °C for 2 min, and a final extension step at 72 °C for 5 min. The reactions were performed on a gradient thermal cycler (Benchmark, Thermal Cycler, United States). The amplicons, as well as a 100 bp ladder (Cat# DM003-R500), were separated on 1.5% agarose gel containing 0.4 μg/mL of ethidium bromide in 1x TAE buffer and visualized under a UV transilluminator. To ensure data reliability, Cysticercus tenuicollis DNA (previously confirmed in our earlier studies) was used as a positive control, and nuclease-free water was used as a negative control in all PCR runs. All amplification steps were performed in separate pre- and post-PCR areas to minimize contamination.
2.4.3 Sample selection and sequencing
Although multiple samples yielded successful amplification for both mitochondrial markers, four representative isolates per gene were selected for sequencing based on the intensity and clarity of their PCR bands, as well as to ensure geographic representation from the three collection sites (Badr, Menoufia, and Assiut). The purified PCR products were sequenced in the forward direction using the same forward primers and the Sanger dideoxy method (Macrogen Inc., Seoul, South Korea; https://dna.macrogen.com).
Chromatograms were visually inspected to confirm base-calling accuracy. All sequences showed clear, high-quality peaks with no ambiguous positions, and no “N” symbols were required in the final sequences. Low-quality ends (Phred <20) were trimmed prior to alignment. The resulting sequences had no unresolved bases and were directly used for BLAST comparison on the NCBI database and phylogenetic analysis. The sequences of the current study were deposited in the GenBank database with available data.
2.4.4 Sequencing and phylogenetic analysis
Multiple sequence alignment of the cox1 and nad1 sequences with related Taenia species retrieved from the GenBank database was performed using Clustal W and Clustal X v2.0 (26). Alignments were visualized and manually adjusted using Jalview v2.11.0 (27). Evolutionary relationships were inferred using both the neighbor-joining (NJ) and maximum likelihood (ML) methods implemented in MEGA 11 (28). For NJ analysis, evolutionary distances were computed using the maximum composite likelihood method (29) with 1,000 bootstrap replicates to assess branch support (30). For ML analysis, the best-fit substitution models were determined using the model-test option in MEGA 11: HKY + G + I for cox1 and HKY + G for nad1 (31). ML trees were reconstructed with 1,000 bootstrap replicates, and the resulting bootstrap consensus trees were used for interpretation (30).
Diphyllobothrium sp. was selected as the outgroup to root all phylogenetic trees. Bootstrap values were displayed next to the corresponding nodes.
2.4.5 Quality control and data validation
All molecular analyses were performed under strict quality control conditions. Separate workstations were used for DNA extraction, PCR setup, and post-PCR analysis to avoid cross-contamination.
Each sequence chromatogram was reviewed to confirm peak clarity and exclude sequencing artifacts. Trimming, alignment, and phylogenetic procedures were performed consistently across all samples to ensure reproducibility and comparability between the cox1 and nad1 datasets.
2.5 Statistical analysis
Statistical analyses were conducted using R 4.4.1 software (R Foundation for Statistical Computing, Vienna, Austria). Descriptive statistics were computed for infection status as the outcome and risk factors, respectively. Independent variables of interest were the age at sample collection (2, 3, or 4 months), management systems (home or farm), sites of collection Badr City (Cairo), Sadat City (Monufia), or Assiut City (Assiut), and seasons of collection (fall, winter, or spring). To investigate the association between these risk factors and the outcome, a chi-square test was used, and statistical significance was set at a p-value of < 0.05. When the assumptions of the chi-square test were not met, Fisher’s exact test was used instead. To determine which specific groups differed significantly when the overall test indicated a significant result, standardized residuals were calculated to measure the difference between the observed and expected frequencies for each cell (32). There is a significant deviation whenever residuals are greater than 1.96.
3 Results
3.1 Descriptive statistics
A total of 150 samples were collected from either home-raised or farm-raised rabbits in Badr City (Cairo), Sadat City (Monufia), and Assiut City (Assiut Province). Of those samples, 32 (21.3%) tested positive for the presence of T. pisiformis. One cyst was isolated from 17 (53%) of the positive samples, two cysts were isolated from 8 (25%) of the samples, three cysts were recovered from 5 (16%) of the samples, and four cysts were isolated from one sample, and five cysts from another one sample, which together accounted for 3% of the samples.
Of the 150 samples, 77 (51.3%) were collected from home-raised rabbits, while the rest of the samples (73, 48.7%) were collected from farm-raised rabbits. Interestingly, only one sample tested positive from farm-raised rabbits (1/73, 1.3%), while 31 samples tested positive from home-raised (40.2%). More information on the distribution of positive and negative samples by age, sampling sites, management systems, and season is provided in Figures 1A–D and Table 1. However, data on seasonal variation were available for only 105 samples.
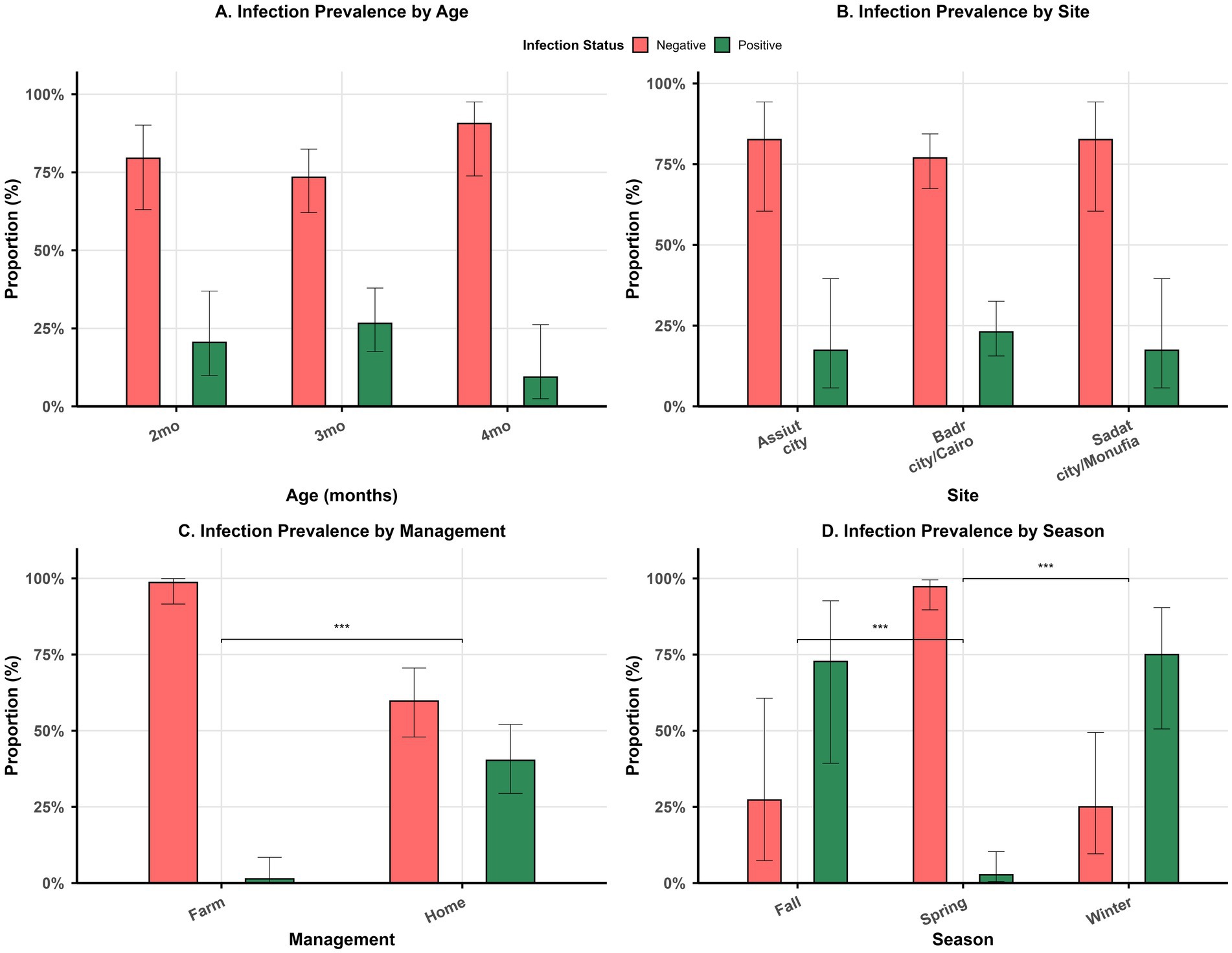
Figure 1. Bar plots show the distribution of positive versus negative samples by age (A), sites of sample collection (B), management systems (C), and season (D). Asterisk *** indicates significant difference in (C,D).
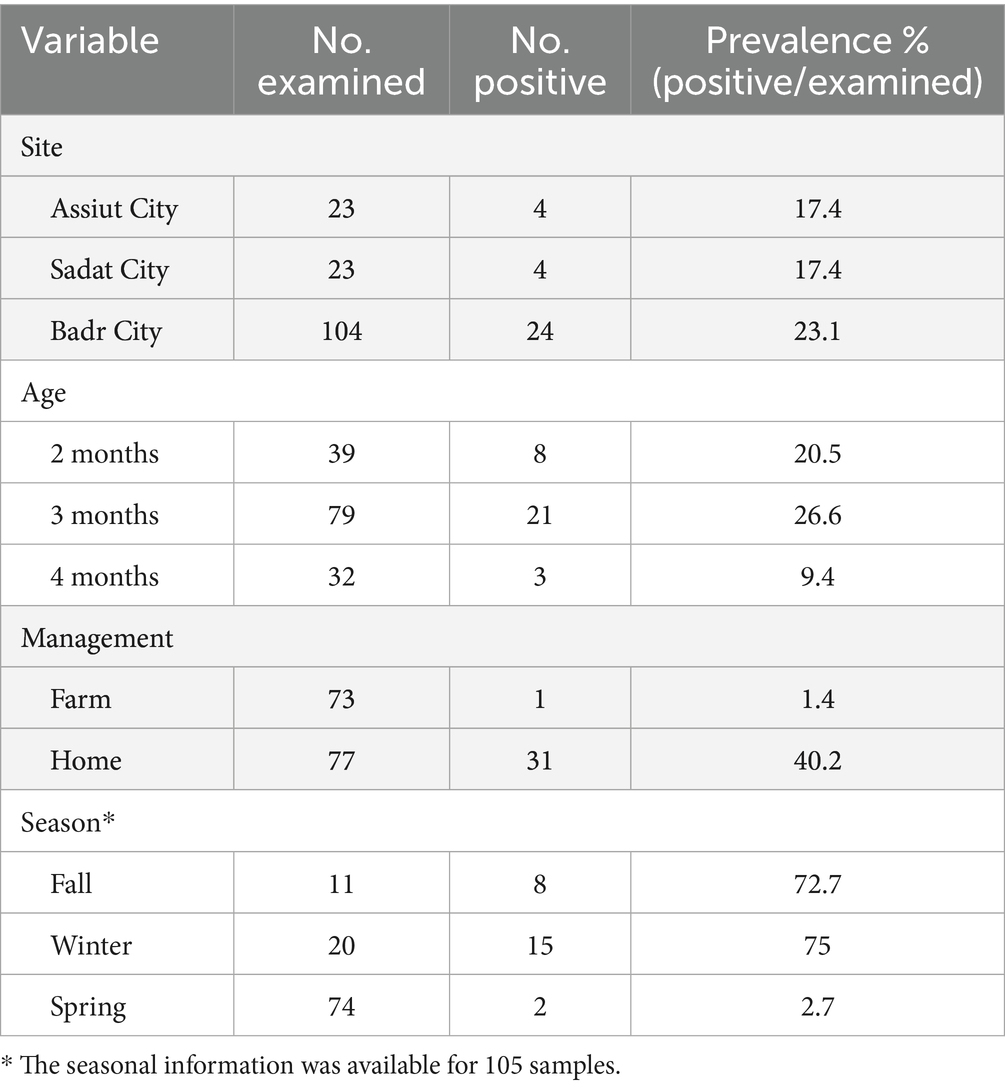
Table 1. Prevalence of T. pisiformis in relation to various risk factors.
3.2 Association between infection status and risk factors
Management was significantly associated with the infection status (X2 = 31.5, p < 0.001). Standardized residuals of 5.8 indicated that farm-raised rabbits were more likely to have negative samples, whereas the corresponding residuals for home-raised rabbits indicated its association with positive samples for T. pisiformis. Furthermore, the season at sample collection was significantly associated with T. pisiformis infection (p < 0.001), where residuals of −3.7 indicated that samples collected in spring were more likely to be negative in contrast to residuals of 3.3 and 4.6 for fall and winter, respectively, indicating their strong association with positive samples for T. pisiformis. On the other hand, neither the age of the animals nor the sites of sample collection were significantly associated with infection status (X2 = 4.03, p = 0.13 for age; p = 0.86 for sites).
3.3 Morphological characteristics
The cystic stage of T. pisiformis appeared as a small, bladder-like structure measuring diameter of 5–8 mm, distinguished by a single invaginated scolex. The mature scolices displayed a substantial size, each bearing four prominent lateral suckers. The rostellum exhibited two distinct rows of hooks characteristic of Taeniidae morphology, consisting of a blade, handle, and guard structure. The total number of hooks was 36, organized into two rows with 18 large hooks and 18 small hooks. The large hooks measured between 112 and 201 μm in length, whereas the smaller hooks ranged from 65 to 121 μm. The suckers were spherical with thick muscular walls and displayed diameters between 96 and 188 μm (Figures 2, 3).
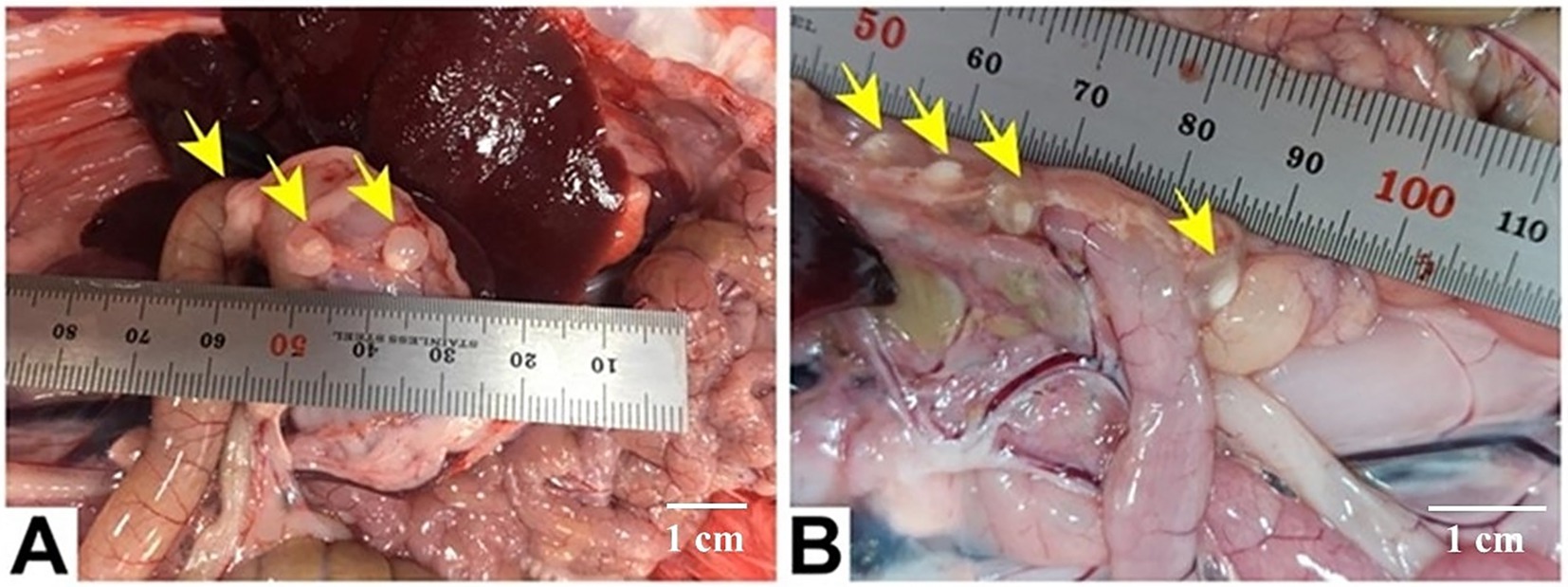
Figure 2. Gross appearance of C. pisiformis cysts (yellow arrows) represented by several small bladder cysts, each containing a single whitish scolex and clear fluid, attached to viscera in the peritoneal cavity near the stomach (A) and the intestine (B). Scale bar = 1 cm in (A,B).

Figure 3. Micrographs of C. pisiformis scolex: the scolex has four suckers, and an armed rostellum has double rows of hooks. Suckers (star) and hooks (arrow). Scale bar = 200 um (not stained 4x (left)). Scale bar = 100 um (not stained 10x (right)).
3.4 Histopathological findings
Larval cysts of C. pisiformis in the peritoneal cavity exhibited a solitary invaginated scolex surrounded by a thick fibrous cyst wall. The invaginated scolex featured the typical suckers and hooks of cysticerci. The larval parenchyma was surrounded by the serrated cuticle and contained numerous basophilic calcareous corpuscles. The liver section showed bile ductular reaction and portal fibrous connective tissue proliferation. The lung section showed vascular medial hypertrophy and mild emphysema (Figure 4).
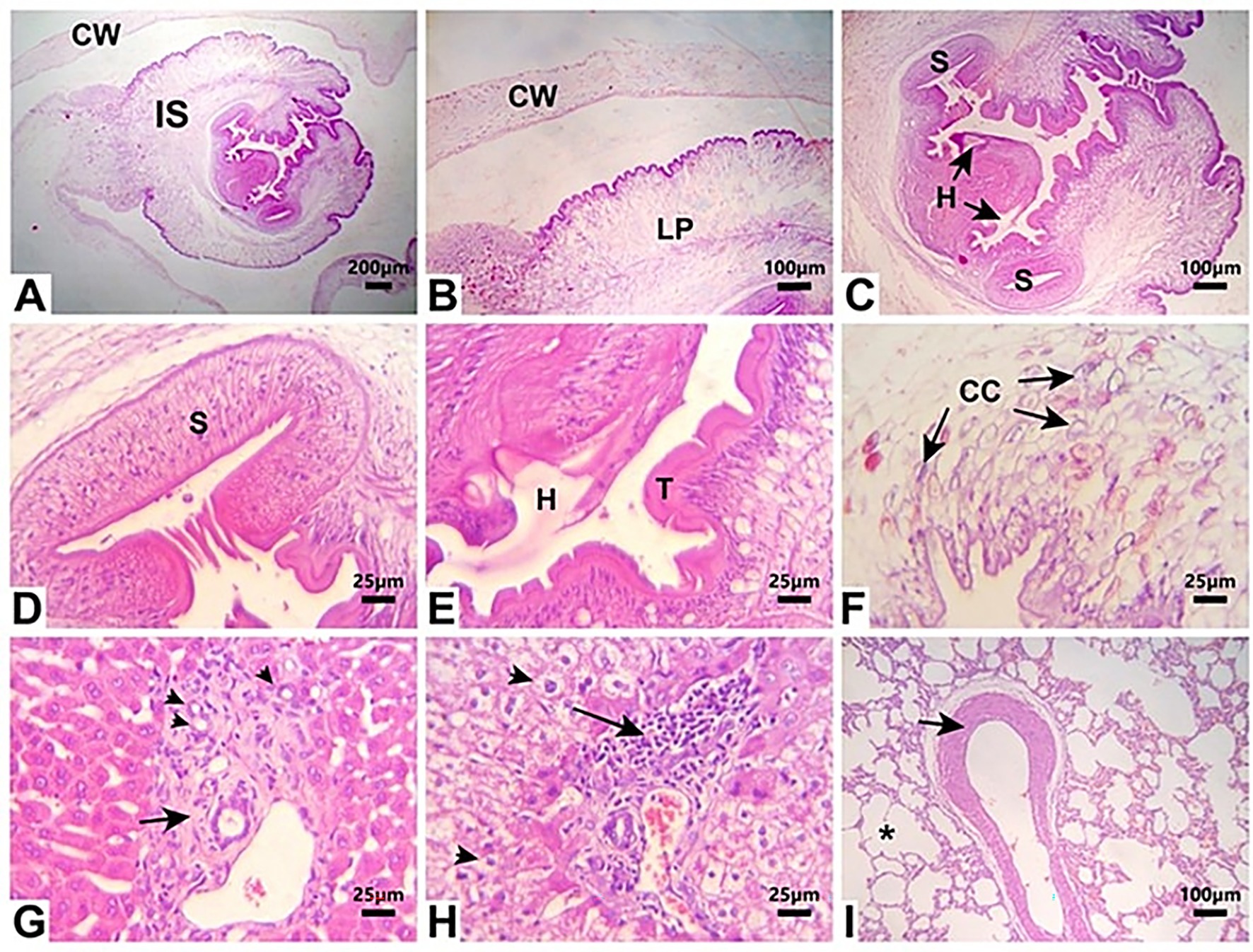
Figure 4. Photomicrographs of H&E-stained tissue sections from necropsied rabbits (A–F) C. pisiformis larval cyst shows a single invaginated scolex (IS) surrounded by a thick fibrous cyst wall (CW). The invaginated scolex displays suckers (S) and hooks (H). The larval parenchyma (LP) is surrounded by a serrated cuticle (T) and contains numerous basophilic calcareous corpuscles (CC). (G) Liver tissue section with portal area shows proliferation of reactive bile ducts (arrowheads) and fibrous connective tissue proliferation (arrow). (H) Liver tissue section shows the vacuolation of hepatocytes (arrowheads) and moderate periportal lymphoplasmacytic infiltration. (I) Lung tissue section shows medial hypertrophy of a blood vessel (arrow) and mild emphysema (asterisk). Scale bar = 200 um in (A). Scale bar = 100 um in (B,C,I). Scale bar = 25 um in (D–H).
3.5 Molecular and phylogenetic analyses
3.5.1 Molecular characterization
Several T. pisiformis cyst samples were successfully amplified using PCR, yielding distinct mitochondrial fragments of approximately 450 bp for the cox1 gene and 500 bp for the nad1 gene. From these, four representative isolates per gene, exhibiting strong and well-defined amplification profiles and originating from the three primary collection sites (Badr, Menoufia, and Assiut), were selected for sequencing.
For the cox1 gene, four sequences were obtained and deposited in the GenBank database under accession numbers PQ135830, PQ144537, PQ144538, and PQ144539. BLAST analysis revealed 99.21–100% identity with T. pisiformis sequences from the GenBank database (query cover 100%), confirming accurate molecular identification. Multiple sequence alignment of the cox1 sequences from Egyptian rabbit isolates, compared with global isolates from China and Poland, showed a high level of nucleotide conservation interspersed with variable positions. Several single-nucleotide polymorphisms (SNPs) were detected at positions 35, 68, 209, and 308, indicating low but measurable intraspecific variation among the isolates. These variations were visualized using Jalview v2.11.0, as shown in Figure 5.

Figure 5. Multiple sequence alignment of the cox1 gene of T. pisiformis. Sequences obtained in this study are indicated by asterisks (*). The alignment compares these sequences with homologous T. pisiformis sequences retrieved from the GenBank database.
Similarly, four nad1 sequences were successfully obtained and registered in the GenBank database under accession numbers PQ156139–PQ156142. BLAST results demonstrated 97.64–100% identity with T. pisiformis isolates from rabbit and dog hosts (query cover 100%). The alignment of the nad1 sequences from Egypt, China, and Poland revealed clear conservation across large regions, with variable sites detected at positions 37, 93, 102, 135, 136, 158, 216, 257, 303, 319, 330, 331, 345, 373, 429, and 458.
These polymorphisms suggest the presence of minor genetic variation potentially related to host or geographic origin, as shown by Jalview v2.11.0 in Figure 6.

Figure 6. Multiple sequence alignment of the nad1 gene of T. pisiformis. Sequences obtained in this study are indicated by asterisks (*). The alignment compares these sequences with homologous T. pisiformis sequences retrieved from the GenBank database.
3.5.2 Phylogenetic analysis (neighbor-joining method)
Phylogenetic analyses were first performed using the neighbor-joining (NJ) method based on 19 cox1 and 17 nad1 nucleotide sequences. Ambiguous positions were removed from each sequence pair, resulting in final alignments of 318 and 327 positions, respectively. In both trees, the Egyptian isolates clustered within the T. pisiformis clade and showed minimal intraspecific variation.
The cox1-based NJ tree grouped the Egyptian sequences closely with isolates from Poland (MZ287426) and China (NC013844, MW350140, JN870101, and JN870104), confirming their genetic identity and close evolutionary relationship (Figure 7). Additionally, sequences from wolves and coyotes (PQ328516, OQ281684, and PP555939) formed closely related but distinct subclusters, suggesting potential host-specific adaptation or early divergence. Other Taenia species, including Taenia saginata (OR143747, AB533173), Taenia multiceps (OP782685, JX535568), and Taenia cakuepengi (MT882036 and PP140883), formed distinct and separate clades, whereas Diphyllobothrium sprakeri (MW596662) served as the outgroup and was positioned on the most distant branch, confirming clear phylogenetic separation.
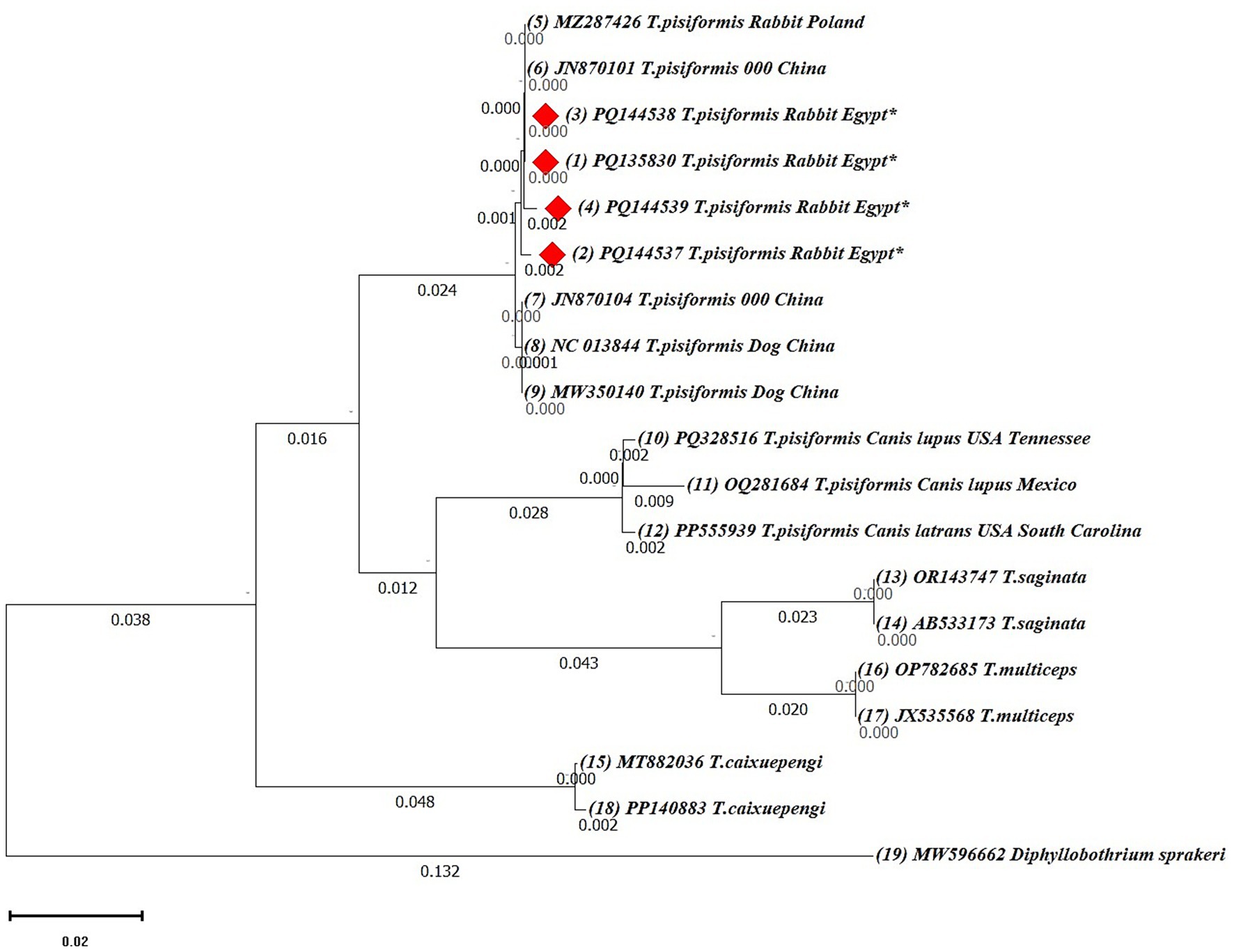
Figure 7. Neighbor-joining (NJ) phylogenetic tree of cox1 gene sequences of Taenia species. The tree was constructed based on evolutionary distances computed using the maximum composite likelihood method. Branch lengths are drawn to scale, representing the number of substitutions per site. The tree is rooted with Diphyllobothrium sprakeri as the outgroup. Sequences from this study are marked with red diamonds (♦).
The nad1-based NJ tree also revealed a consistent topology, with the Egyptian T. pisiformis isolates clustering strongly with global sequences from rabbit and dog hosts, as well as a fox-derived isolate (AJ239109), which grouped within the clade but exhibited a slightly higher genetic distance, suggesting potential host-specific adaptation or early divergence (Figure 8). Other Taenia species, including T. multiceps (KR604806), Taenia ovis (NC021138, KU995333), Taenia serialis (AM503337, DQ410137), and Taenia crocutae (NC024591), formed distinct and separate clades, whereas Diphyllobothrium schistochilos (MW602528) served as the outgroup and was positioned on the most distant branch, confirming clear phylogenetic separation.

Figure 8. The neighbor-joining (NJ) phylogenetic tree of nad1 gene sequences of Taenia species. The tree was constructed based on evolutionary distances computed using the maximum composite likelihood method. Branch lengths are drawn to scale, representing the number of substitutions per site. The tree is rooted with Diphyllobothrium schistochilos as the outgroup. Sequences from this study are marked with red diamonds (♦).
3.5.3 Phylogenetic analysis (maximum likelihood method)
To validate the NJ-based results, additional phylogenetic trees were constructed using the maximum likelihood (ML) method for both cox1 and nad1 genes (Figures 9, 10).
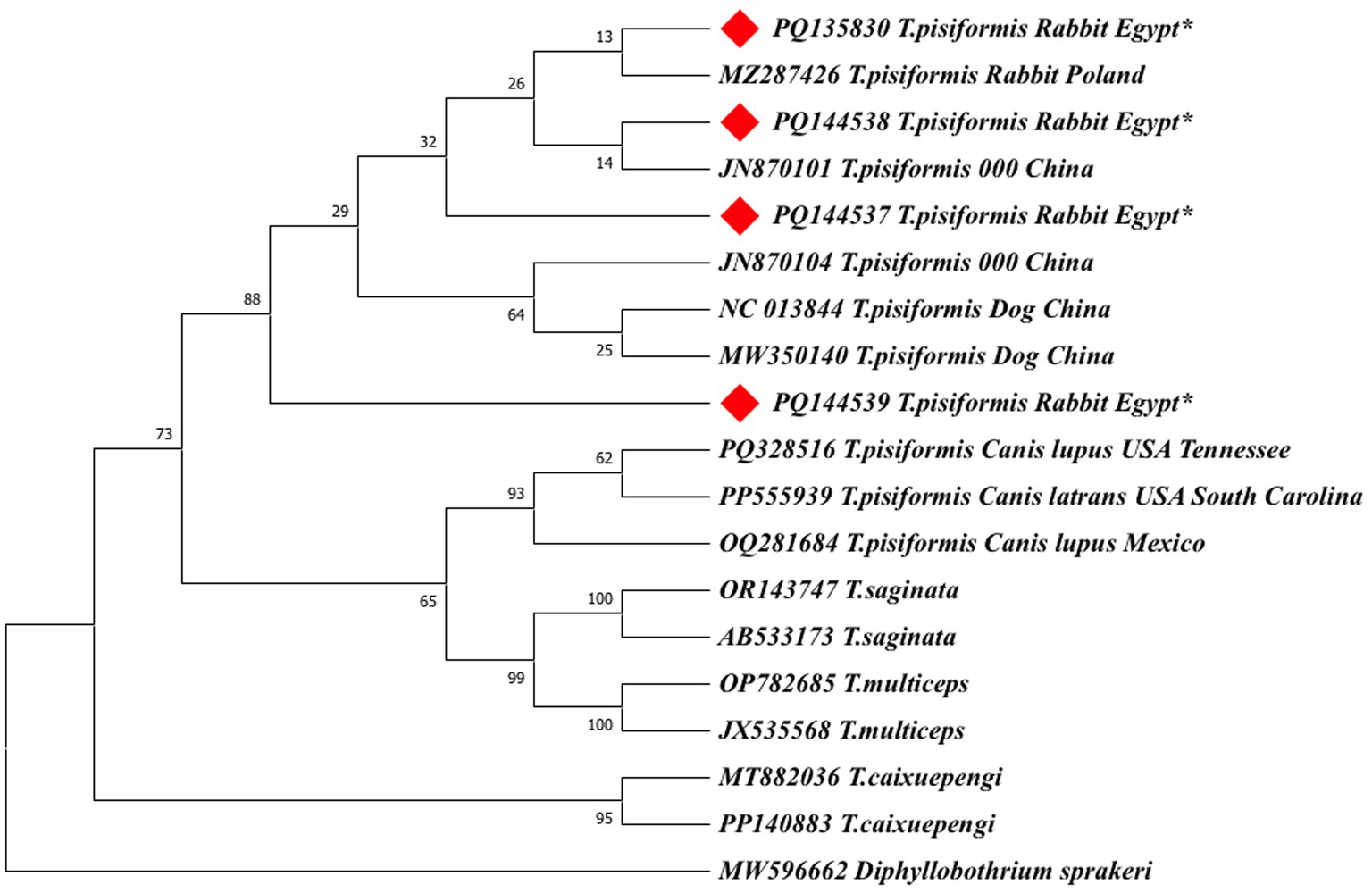
Figure 9. The maximum likelihood (ML) phylogenetic tree based on cox1 gene sequences of Taenia species, inferred under the Hasegawa–Kishino–Yano (HKY) + G + I model. Bootstrap values (1,000 replicates) are shown at the nodes. Branch lengths are proportional to the number of substitutions per site. The tree was rooted with Diphyllobothrium sprakeri. Sequences from this study are marked with red diamonds (♦).
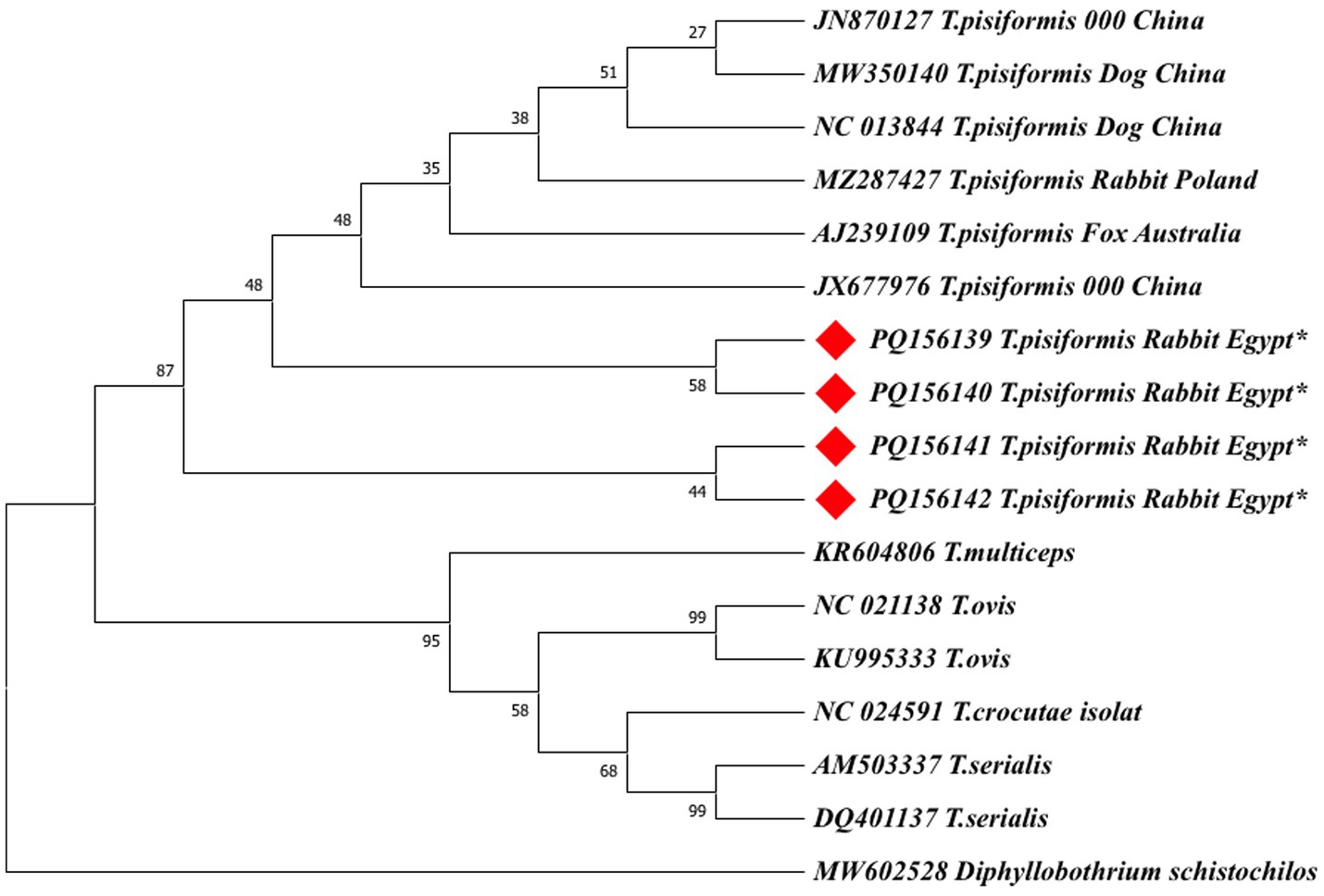
Figure 10. The maximum likelihood (ML) phylogenetic tree based on nad1 gene sequences of Taenia species, inferred under the HKY + G model. Bootstrap values (1,000 replicates) are shown at the nodes. Branch lengths are proportional to the number of substitutions per site. The tree was rooted with Diphyllobothrium schistochilos. Sequences from this study are marked with red diamonds (♦).
Both ML trees showed consistent topologies with the NJ trees, supporting the robustness of the phylogenetic inference.
In the cox1-based ML tree (Figure 9), the Egyptian isolates formed a well-supported clade (bootstrap values >70%) with other global T. pisiformis sequences from rabbits and dogs, confirming their genetic identity within the species. Sequences from wild canids again formed distinct subgroups, supporting the potential host-specific patterns observed in the NJ tree. Other Taenia species formed clearly separated clades, with Diphyllobothrium sprakeri as the outgroup.
In the nad1-based ML tree (Figure 10), the Egyptian isolates form a monophyletic cluster with strong support (bootstrap = 95%) within a larger clade containing global T. pisiformis sequences from rabbits, dogs, and a fox. The fox-derived isolate (AJ239109) grouped within the main T. pisiformis clade but showed a slightly separate position with weak nodal support (bootstrap = 35%), reinforcing the possibility of host-associated genetic variation. Other Taenia species formed distinct and well-separated clades with varying bootstrap support: T. multiceps (95%), T. ovis (58%), T. serialis (99%), and T. crocutae (68%). The clear separation of these congeneric species was further confirmed by the maximum support (bootstrap = 99%) for the deeper nodes defining their distinct lineages. Diphyllobothrium schistochilos served as the outgroup, occupying the most distant branch and confirming robust phylogenetic structure. These findings, despite the minor SNPs detected in the alignments, collectively affirm that T. pisiformis isolates from Egyptian rabbits are genetically homogeneous and closely related to T. pisiformis isolates from other geographic regions.
3.5.4 Genetic distance estimation
The evolutionary divergence among cox1 and nad1 sequences of Taenia species and outgroup taxa was estimated using the maximum composite likelihood model in the MEGA11 software.
The average evolutionary divergence values were 0.0846 for cox1 (with D. sprakeri as the outgroup) and 0.1512 for nad1 (with D. schistochilos as the outgroup). These data, summarized in Supplementary Tables 1, 2, indicate moderate interspecific genetic distances between T. pisiformis and other Taenia species.
4 Discussion
T. pisiformis, one of the most common intestinal parasites in canines, significantly affects rabbit health and causes notable economic losses in the rabbit breeding industry (33). In Egypt, research on T. pisiformis has been limited, with only one study reported to date (20). In the current study, an overall infection rate of 21.3% was observed, with a significantly higher prevalence in home-raised rabbits (40.2%) than in farm-raised rabbits (1.3%). This difference may be attributed to variations in management systems and feeding practices, with home-raised rabbits reared in less controlled environmental and hygienic conditions, reared in closer contact with dogs (the definitive host), and that did not undergo regular deworming. In contrast, farm-raised rabbits are generally maintained under better sanitary conditions and limited exposure to dogs, resulting in lower infection rates.
Our results align with recent findings reporting a 30% prevalence in domestic rabbits in Egypt and a 23.33% infection rate in local breed rabbits in Iraq, respectively (20, 34). This finding indicates the persistent circulation of this parasite in small-scale or household rabbit-rearing systems. Such findings further emphasize the need to improve hygiene and management practices to reduce transmission risk and control infection.
In contrast, much lower prevalence rates have been recorded in wild rabbits in Europe. In southern Spain, only 2.8% (n = 2,923) of wild rabbits were infected (6); in Scotland, a prevalence of 3% (n = 786) was reported (35); and in northern Spain, 3.7% (n = 267) was detected among lagomorphs (36). Similarly, a Polish study identified a prevalence of 4.74% (n = 274) in slaughter rabbits from both small-scale and commercial farms, with no major differences linked to production system or season (11). However, other European studies reported higher prevalence rates in wild rabbits, ranging from 16.1% (n = 153) in southern Spain to 33.3% (n = 62) in the northeastern region (37). In the Canary Islands, prevalence rates in wild rabbits were found to range between 12% (n = 292) and 15% (n = 104) (38, 39). A higher average prevalence of 28% (n = 95) was also recorded in wild rabbits from England (40).
The lower prevalence in wild populations has been linked to better hygiene practices, such as mandatory antiparasitic treatments for dogs and public awareness campaigns about proper carcass disposal (6). Backyard home-raised rabbits, with more exposure to outdoor environments, appear more vulnerable due to possible contact with feces of definitive hosts such as dogs and foxes (41). Discrepancies in prevalence across studies may be due to differences in numbers of tested animals, populations sampled, sampling design, rabbit type (domestic vs. wild), and the distribution of stray dogs (6, 34). A convenience sampling approach was used in this study based on the ease of accessibility to available samples. This may have introduced potential bias in the prevalence estimates, as such a sample might not accurately reflect the true prevalence of T. pisiformis in Egyptian rabbits. Therefore, this represents a limitation of the study. Future research should employ random sampling of both home- and farm-raised rabbits from different Egyptian governorates to allow for more representative and reliable estimates of parasite prevalence. As emphasized in previous research, future studies should focus on identifying other potential definitive or intermediate hosts and understanding the molecular epidemiology of the parasite (6). Another limitation of the study was the inability to analyze the infection status with T. pisiformis as a dichotomous outcome in relation to age, management, and season using a logistic regression model. This was due to high collinearity and the small number of available variables, which together prevented us from proceeding with the multivariable analysis.
In this study, positive samples for T. pisiformis were more likely to occur in fall and winter compared to those collected in spring. This finding aligns with a previous study that also reported positive cases during the hunting season, spanning from August to March (6). However, in the latter study, sampling was limited to this specific period (i.e., hunting season), making direct seasonal comparisons difficult. A more reliable comparison would require regular sampling throughout the entire year.
It is worth noting that the histopathological evaluation in this study was descriptive rather than semi-quantitative. Although this approach allowed detailed characterization of lesion patterns, the absence of a standardized scoring system represents a limitation that future studies could address to enable more objective comparison of lesion severity.
The present study utilized the mitochondrial cox1 and nad1 genes to genetically characterize T. pisiformis cysts from Egyptian rabbits and elucidate their phylogenetic relationship with global isolates. These markers were selected for their well-established utility in providing robust species-level resolution and detecting intraspecific variation within cestodes due to their high evolutionary rates and conserved flanking regions (42–44).
Phylogenetic reconstructions using both the neighbor-joining (NJ) and maximum likelihood (ML) methods yielded congruent topologies, consistently placing the Egyptian isolates within the major T. pisiformis clade. The strong clustering with sequences from China and Poland highlights a shared genetic ancestry and suggests limited phylogeographic structuring for this parasite on a global scale (43).
The modest sample size (n = 4), which is the primary limitation of this study, is important to consider. However, the consistent clustering of all Egyptian isolates from different governorates with global sequences strongly indicates a genetically homogeneous population of T. pisiformis in the region. The identified SNPs are therefore more parsimoniously interpreted as natural microvariation within a conserved genetic background rather than as indicators of distinct sub-lineages, a phenomenon previously documented in globally distributed T. pisiformis populations (3).
An intriguing finding was the formation of distinct subclusters by sequences derived from wild canids. The fox-derived sequence (AJ239109), while nested within the T. pisiformis clade, exhibited a longer branch length and weaker nodal support, suggesting a degree of genetic distance and providing tentative evidence for potential host-associated divergence. It is plausible that the ecological separation and different evolutionary pressures in wild canid hosts may drive subtle genetic structuring within the species, a hypothesis supported by earlier studies (45).
The extensive gene flow facilitated by the wide host range of T. pisiformis (45–47) likely maintains the overall genetic homogeneity but may not entirely preclude localized adaptation. As expected, both ML and NJ trees provided clear interspecific differentiation, with other Taenia species forming well-supported, distinct clades that corroborate their established taxonomic status.
The present findings underscore a major gap in understanding the economic and public health implications of T. pisiformis in Egypt. While local data are scarce, infections elsewhere have been linked to reduced growth performance and carcass value in rabbits. Given the parasite’s life cycle involving dogs, a minor zoonotic risk cannot be excluded. Also, this study confirms the genetic congruence of Egyptian T. pisiformis isolates with a globally conserved lineage. While mitochondrial markers (cox1 and nad1) effectively resolved species identity, subtle indications of host-associated variation highlight the need for further investigation with larger sample sizes and advanced genomic tools, such as nuclear markers or full mitogenomes, to better elucidate potential population-level diversity and host adaptation dynamics. In addition, future studies should quantify economic losses and evaluate preventive measures such as regular deworming of farm dogs, improved sanitation, and enhanced health monitoring in rabbit production systems.
5 Conclusion
This study highlights the metacestode stage of T. pisiformis as a significant parasitic threat to domestic rabbits in Egypt, especially those raised at home. A high prevalence rate, especially during fall and winter in these environments, highlights the consequences of poor management and environmental exposure. Infection rates are further affected by seasonal changes, particularly in the fall and winter, whereas factors such as the age of the rabbits and their geographic location seem to be less influential. Molecular analysis using cox1 and nad1 genes confirmed the parasite’s identity and showed genetic diversity among local strains, which closely resembled global isolates. These findings emphasize the urgent need for improved husbandry for comprehensive control measures, including better husbandry practices and regular molecular monitoring, and the integration of molecular tools of C. pisiformis in rabbit farming. Furthermore, educating breeders on appropriate carcass disposal methods is essential for public health. Future surveillance initiatives targeting dogs, with the implementation of deworming protocols, are also recommended to disrupt the parasite’s life cycle.
Data availability statement
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found in the article/Supplementary material.
Ethics statement
The animal study was approved by the Research Ethics Committee at Faculty of Pharmacy, Badr University in Cairo, Cairo, Egypt. The study was conducted in accordance with the local legislation and institutional requirements.
Author contributions
RR: Conceptualization, Data curation, Formal analysis, Investigation, Methodology, Resources, Software, Supervision, Validation, Visualization, Writing – original draft, Writing – review & editing. AE-B: Writing – original draft, Writing – review & editing, Conceptualization, Data curation, Formal analysis, Investigation, Methodology, Resources, Validation, Visualization. AA: Conceptualization, Data curation, Formal analysis, Investigation, Methodology, Resources, Software, Supervision, Validation, Visualization, Writing – original draft, Writing – review & editing. FM: Data curation, Formal analysis, Funding acquisition, Investigation, Methodology, Resources, Validation, Visualization, Writing – original draft, Writing – review & editing. AE: Conceptualization, Data curation, Formal analysis, Investigation, Methodology, Resources, Software, Validation, Visualization, Writing – original draft, Writing – review & editing. DN: Data curation, Formal analysis, Investigation, Methodology, Resources, Software, Supervision, Validation, Visualization, Writing – original draft, Writing – review & editing. AP: Data curation, Formal analysis, Funding acquisition, Investigation, Methodology, Resources, Validation, Visualization, Writing – original draft, Writing – review & editing. MI: Data curation, Formal analysis, Funding acquisition, Investigation, Methodology, Resources, Validation, Visualization, Writing – original draft, Writing – review & editing. MA: Conceptualization, Data curation, Formal analysis, Investigation, Methodology, Resources, Software, Supervision, Validation, Visualization, Writing – original draft, Writing – review & editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. The article processing charges (APC) was funded by the University of Life Science “King Mihai I” from Timișoara.
Acknowledgments
The authors wish to express their sincere gratitude to Ahmed Abdel-Hamied, Ahmed El Nemr, and Mai Saad, Faculty of Veterinary Medicine, Badr University in Cairo (BUC), Badr City, Cairo, Egypt, for their technical assistance.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The authors declare that no Gen AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fvets.2025.1701083/full#supplementary-material
References
1. Yang, D, Ren, Y, Fu, Y, Xie, Y, Nie, H, Nong, X, et al. Genetic variation of Taenia pisiformis collected from Sichuan, China, based on the mitochondrial cytochrome B gene. Korean J Parasitol. (2013) 51:449–452. doi: 10.3347/kjp.2013.51.4.449,
2. Yang, Q, Li, J, Zhang, L, Zhao, N, Sun, X, and Wang, Z. Type I cystatin derived from Cysticercus pisiformis—Stefins, suppresses LPS-mediated inflammatory response in RAW264. 7 cells. Microorganisms. (2024) 12:850. doi: 10.3390/microorganisms12050850,
3. Samorek-Pieróg, M, Karamon, J, Brzana, A, Bilska-Zając, E, Zdybel, J, and Cencek, T. Molecular confirmation of massive Taenia pisiformis cysticercosis in one rabbit in Poland. Pathogens. (2021) 10:1029. doi: 10.3390/pathogens10081029,
4. Ashour, G, and El-Bahrawy, KA. Genetic resources and optimal utilization of rabbits in Egypt. Egypt J Rabbit Sci. (2025) 35:1–32. doi: 10.21608/ejrs.2025.399452
5. Chen, G, Pu, G, Wang, L, Li, Y, Liu, T, Li, H, et al. Cysticercus pisiformis-derived novel-miR1 targets TLR2 to inhibit the immune response in rabbits. Front Immunol. (2023) 14:1201455. doi: 10.3389/fimmu.2023.1201455,
6. Remesar, S, Castro-Scholten, S, Jiménez-Martín, D, Camacho-Sillero, L, Morrondo, P, Rouco, C, et al. Spatiotemporal monitoring of Cysticercus pisiformis in European wild rabbit (Oryctolagus cuniculus) in Mediterranean ecosystems in southern Spain. Prev Vet Med. (2021) 197:105508. doi: 10.1016/j.prevetmed.2021.105508,
7. Zhang, S. Comparative transcriptomic analysis of the larval and adult stages of Taenia pisiformis. Genes. (2019) 10:507. doi: 10.3390/genes10070507,
8. Arias-Hernández, D, Flores-Pérez, FI, Domínguez-Roldan, R, Báez-Saldaña, A, Carreon, RA, García-Jiménez, S, et al. Influence of the interaction between cysticercosis and obesity on rabbit behavior and productive parameters. Vet Parasitol. (2019) 276:108964. doi: 10.1016/j.vetpar.2019.108964,
9. Rosa, D-R, David, A-H, Emmanuel, D-G, Edda, S, Virginio, A-F, Ivan, F-P, et al. Decreased embryo implantation in rabbits infected with Taenia pisiformis. Parasitol Res. (2022) 121:3689–3692. doi: 10.1007/s00436-022-07694-2,
10. Stancampiano, L, Ravagnan, S, Capelli, G, and Militerno, G. Cysticercosis by Taenia pisiformis in Brown hare (Lepus europaeus) in northern Italy: epidemiologic and pathologic features. Int J Parasitol Parasites Wildl. (2019) 9:139–143. doi: 10.1016/j.ijppaw.2019.04.004,
11. Szkucik, K, Pyz-Łukasik, R, Szczepaniak, KO, and Paszkiewicz, W. Occurrence of gastrointestinal parasites in slaughter rabbits. Parasitol Res. (2014) 113:59–64. doi: 10.1007/s00436-013-3625-7,
12. Domínguez-Roldan, R, Pérez-Martínez, M, Rosetti, M, Arias-Hernández, D, Bernal-Fernández, G, Flores-Pérez, F, et al. High frequency of Taenia pisiformis metacestodes and high sex-associated susceptibility to cysticercosis in naturally infected wild rabbits. Parasitol Res. (2018) 117:2201–2206. doi: 10.1007/s00436-018-5907-6
13. Nakao, M, Lavikainen, A, Iwaki, T, Haukisalmi, V, Konyaev, S, Oku, Y, et al. Molecular phylogeny of the genus Taenia (Cestoda: Taeniidae): proposals for the resurrection of Hydatigera Lamarck, 1816 and the creation of a new genus Versteria. Int J Parasitol. (2013) 43:427–437. doi: 10.1016/j.ijpara.2012.11.014
14. Zink, RM, and Barrowclough, GF. Mitochondrial DNA under siege in avian phylogeography. Mol Ecol. (2008) 17:2107–2121. doi: 10.1111/j.1365-294X.2008.03737.x,
15. Jeon, H-K, Kim, K-H, and Eom, KS. Complete sequence of the mitochondrial genome of Taenia saginata: comparison with T. Solium and T. asiatica. Parasitol Int. (2007) 56:243–246. doi: 10.1016/j.parint.2007.04.001,
16. Nakao, M, Xiao, N, Okamoto, M, Yanagida, T, Sako, Y, and Ito, A. Geographic pattern of genetic variation in the fox tapeworm Echinococcus multilocularis. Parasitol Int. (2009) 58:384–9. doi: 10.1016/j.parint.2009.07.010,
17. Tsubota, K, Nakatsuji, S, Matsumoto, M, Fujihira, S, Yoshizawa, K, Okazaki, Y, et al. Abdominal cysticercosis in a cynomolgus monkey. Vet Parasitol. (2009) 161:339–41. doi: 10.1016/j.vetpar.2009.01.024,
18. Abdullatif, AF, Mahmoud, AF, Hafez, AESE, Abdelkhalek, A, and Ras, R. Rabbit meat consumption: a mini review on the health benefits, potential hazards and mitigation. J Adv Vet Res 2023;13:681–684. Available online at: https://advetresearch.com/index.php/AVR/article/view/1300.
19. Morshdy, AEM, Alsayeqh, AF, Aljasir, MF, Mohieldeen, H, El-Abody, SG, Mohamed, ME, et al. Rabbit meat as a potential source of Staphylococcus aureus and Salmonella spp. Slov Vet Res. (2023) 60:439–445. doi: 10.26873/svr-1674-2023
20. Rabie, SA, Abuelwafa, WA, Eldin, MMM, and Hussein, NM. Infection of Egyptian domestic rabbits, Oryctolagus cuniculus, with Cysticercus pisiformis (Cestoda: Taeniidae): morphological, molecular, and histopathological diagnostic tools. J Parasit Dis. (2024) 48:810–822. doi: 10.1007/s12639-024-01699-7
21. Suvarna, SK, Layton, C, and Bancroft, JD. Bancroft’s theory and practice of histological techniques (8th ed) Churchill Livingstone. Amsterdam, The Netherlands: Elsevier (2019).
22. Ayana, M, Cools, P, Mekonnen, Z, Biruksew, A, Dana, D, Rashwan, N, et al. Comparison of four DNA extraction and three preservation protocols for the molecular detection and quantification of soil-transmitted helminths in stool. PLoS Negl Trop Dis. (2019) 13:e0007778. doi: 10.1371/journal.pntd.0007778,
23. Rubiola, S, Chiesa, F, Zanet, S, and Civera, T. Molecular identification of Sarcocystis spp. in cattle: partial sequencing of cytochrome C oxidase subunit 1 (COI). Ital J Food Saf. (2019) 7:7725. doi: 10.4081/ijfs.2018.7725,
24. Bowles, J, Blair, D, and McManus, DP. Genetic variants within the genus Echinococcus identified by mitochondrial DNA sequencing. Mol Biochem Parasitol. (1992) 54:165–173. doi: 10.1016/0166-6851(92)90109-W,
25. Qingqiu, Z, Xiaohui, S, Xu, W, Xiaodong, W, Xiaoming, W, Youzhong, D, et al. Taeniid cestodes in Tibetan foxes (Vulpes Ferrilata) detected by copro-PCR: applications and challenges. Int J Parasitol Parasites Wildl. (2020) 12:242–249. doi: 10.1016/j.ijppaw.2020.06.008,
26. Larkin, MA, Blackshields, G, Brown, NP, Chenna, R, McGettigan, PA, McWilliam, H, et al. Clustal w and clustal x version 2.0. Bioinformatics. (2007) 23:2947–2948. doi: 10.1093/bioinformatics/btm404,
27. Waterhouse, AM, Procter, JB, Martin, DM, Clamp, M, and Barton, GJ. Jalview version 2—a multiple sequence alignment editor and analysis workbench. Bioinformatics. (2009) 25:1189–1191. doi: 10.1093/bioinformatics/btp033,
28. Tamura, K, Stecher, G, and Kumar, S. MEGA11: molecular evolutionary genetics analysis version 11. Mol Biol Evol. (2021) 38:3022–3027. doi: 10.1093/molbev/msab120,
29. Tamura, K, Nei, M, and Kumar, S. Prospects for inferring very large phylogenies by using the neighbor-joining method. Proc Natl Acad Sci USA. (2004) 101:11030–11035. doi: 10.1073/pnas.0404206101,
30. Felsenstein, J. Confidence limits on phylogenies: an approach using the bootstrap. Evolution. (1985) 39:783–791. doi: 10.1111/j.1558-5646.1985.tb00420.x,
31. Hasegawa, M, Kishino, H, and Yano, T. Dating of the human-ape splitting by a molecular clock of mitochondrial DNA. J Mol Evol. (1985) 22:160–174. doi: 10.1007/bf02101694,
32. García-Pérez, MA, Núñez-Antón, V, and Alcalá-Quintana, R. Analysis of residuals in contingency tables: another nail in the coffin of conditional approaches to significance testing. Behav Res Methods. (2015) 47:147–161. doi: 10.3758/s13428-014-0472-0,
33. Wang, L-Q, Liu, T-L, Liang, P-H, Zhang, S-H, Li, T-S, Li, Y-P, et al. Characterization of exosome-like vesicles derived from Taenia pisiformis cysticercus and their immunoregulatory role on macrophages. Parasit Vectors. (2020) 13:318. doi: 10.1186/s13071-020-04186-z,
34. Al-Khayat, KKK, and Al-Azawi, AKA. Sequence and morphology of cysticercus pisiformis in local breed rabbits. Adv Anim Vet Sci. (2024) 12:392–398. doi: 10.17582/journal.aavs/2024/12.3.392.398
35. Boag, B. The incidence of helminth parasites from the wild rabbit Oryctolagus cuniculus (L.) in eastern Scotland. J Helminthol. (1985) 59:61–69. doi: 10.1017/S0022149X00034507,
36. Espinosa, J, Ferreras, MC, Benavides, J, Cuesta, N, Pérez, C, García Iglesias, MJ, et al. Causes of mortality and disease in rabbits and hares: a retrospective study. Animals (Basel). (2020) 10:158. doi: 10.3390/ani10010158,
37. Blasco, S, Torres, J, Feliu, C, Casanova, J, Miquel, J, and Moreno, S. The helminthfauna of Oryctolagus cuniculus (Linnaeus, 1758) in the Iberian Peninsula. Faunistic and ecological considerations. Parasite. (1996) 3:327–333. doi: 10.1051/parasite/1996034327
38. Foronda, P, Del Castillo, A, Abreu, N, Figueruelo, E, Piñero, J, and Casanova, J. Parasitic helminths of the wild rabbit, Oryctolagus cuniculus, in different bioclimatic zones in Tenerife, Canary Islands. J Helminthol. (2003) 77:305–309. doi: 10.1079/JOH2003182,
39. Fernández-Álvarez, Á, Feliu, C, Miquel, J, Torres, J, and Foronda, P. Helminth fauna of wild rabbit Oryctolagus cuniculus in the Canary Islands, Spain. Helminthologia. (2013) 50:155–160. doi: 10.2478/s11687-013-0125-3
40. Allan, J, Craig, P, Sherington, J, Rogan, M, Storey, D, Heath, S, et al. Helminth parasites of the wild rabbit Oryctolagus cuniculus near Malham Tarn, Yorkshire, UK. J Helminthol. (1999) 73:289–294. doi: 10.1017/S0022149X99000487,
41. Jansen, F, Dorny, P, Gabriël, S, Dermauw, V, Johansen, MV, and Trevisan, C. The survival and dispersal of Taenia eggs in the environment: what are the implications for transmission? A systematic review. Parasit Vectors. (2021) 14:88. doi: 10.1186/s13071-021-04589-6
42. Jia, W-Z, Yan, H-B, Guo, A-J, Zhu, X-Q, Wang, Y-C, Shi, W-G, et al. Complete mitochondrial genomes of Taenia multiceps, T. Hydatigena and T. Pisiformis: additional molecular markers for a tapeworm genus of human and animal health significance. BMC Genomics. (2010) 11:447. doi: 10.1186/1471-2164-11-447,
43. Yang, D, Ren, Y, Fu, Y, Xie, Y, Nong, X, Gu, X, et al. Genetic characteristics of Chinese isolates of the tapeworm Taenia pisiformis based on two mitochondrial genes. J Helminthol. (2015) 89:502–505. doi: 10.1017/S0022149X14000236,
44. Gasser, RB, Zhu, X, and McManus, DP. NADH dehydrogenase subunit 1 and cytochrome c oxidase subunit I sequences compared for members of the genus Taenia (Cestoda). Int J Parasitol. (1999) 29:1965–1970. doi: 10.1016/S0020-7519(99)00153-8,
45. Camacho-Giles, V, Hortelano-Moncada, Y, Torres-Carrera, G, Gil-Alarcón, G, Oceguera-Figueroa, A, García-Prieto, L, et al. Helminths of free-ranging dogs and cats in an urban natural reserve in Mexico City and their potential risk as zoonotic agents. PLoS One. (2024) 19:e0310302. doi: 10.1371/journal.pone.0310302,
46. Baker, E, Dennis, M, Miller, D, Su, C, Rosypal von Dohlen, A, Abouelkhair, MA, et al. Pathology and parasitology of free-ranging coyotes from Tennessee and South Carolina. PLoS One. (2025) 20:e0318645. doi: 10.1371/journal.pone.0318645,
Keywords: cysticercosis, pisiformis , rabbits, prevalence, molecular
Citation: Ras R, El-Bahrawy A, Abdelkhalek A, Morariu F, Elsayed AN, Nouh DS, Plesko A, Ilie MS and AbdelMageed M (2025) A survey of morphological, molecular, and histopathological characteristics of Taenia pisiformis metacestode in Egyptian rabbits (Oryctolagus cuniculus). Front. Vet. Sci. 12:1701083. doi: 10.3389/fvets.2025.1701083
Edited by:
Mutee Murshed, King Saud University, Saudi ArabiaReviewed by:
Mohammed M. Qaid, King Saud University, Saudi ArabiaMohammed Mares, King Saud University, Saudi Arabia
Copyright © 2025 Ras, El-Bahrawy, Abdelkhalek, Morariu, Elsayed, Nouh, Plesko, Ilie and AbdelMageed. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Refaat Ras, UmVmYWF0LmF0ZWZAYnVjLmVkdS5lZw==; Marius Stelian Ilie, bWFyaXVzaWxpZUB1c3Z0LnJv