 María C. Albareda1
María C. Albareda1 María A. Natale1
María A. Natale1 Ana M. De Rissio1
Ana M. De Rissio1 Marisa Fernandez1Alicia Serjan2María G. Alvarez3Gretchen Cooley4Huifeng Shen4Rodolfo Viotti3Jacqueline Bua1
Marisa Fernandez1Alicia Serjan2María G. Alvarez3Gretchen Cooley4Huifeng Shen4Rodolfo Viotti3Jacqueline Bua1 Melisa D. Castro Eiro1Myriam Nuñez5Laura E. Fichera1Bruno Lococo3Karenina Scollo1Rick L. Tarleton4
Melisa D. Castro Eiro1Myriam Nuñez5Laura E. Fichera1Bruno Lococo3Karenina Scollo1Rick L. Tarleton4 Susana A. Laucella1,3*
Susana A. Laucella1,3*- 1Instituto Nacional de Parasitología Dr. M. Fatala Chaben, Buenos Aires, Argentina
- 2Hospital Fernandez, Buenos Aires, Argentina
- 3Hospital Interzonal General de Agudos Eva Perón, Buenos Aires, Argentina
- 4Center for Tropical and Emerging Global Diseases, Athens, GA, United States
- 5Department of Mathematical Physics, Facultad Farmacia y Bioquímica, Universidad de Buenos Aires, Buenos Aires, Argentina
Background: In contrast to adults, Trypanosoma cruzi-infected children have more broadly functional Trypanosoma cruzi-specific T cells, and the total T-cell compartment exhibits fewer signs of immune exhaustion. However, not much is known about the link between immunocompetence and the treatment efficacy for human Chagas disease.
Methods: Using cytokine enzyme-linked immunosorbent spot (ELISPOT) polychromatic flow cytometry, cytometric bead assay, multiplex serological assays and quantitative PCR, we evaluated T. cruzi-specific T-cell and antibody immune responses, T-cell phenotypes and parasitemia in children in the early chronic phase of Chagas disease undergoing anti-Trypanosoma cruzi treatment.
Results: Treatment with benznidazole or nifurtimox induced a decline in T. cruzi-specific IFN-γ- and IL-2-producing cells and proinflammatory cytokines and chemokines. T-cell responses became detectable after therapy in children bearing T-cell responses under background levels prior to treatment. The total frequencies of effector, activated and antigen-experienced T cells also decreased following anti-T. cruzi therapy, along with an increase in T cells expressing the receptor of the homeostatic cytokine IL-7. Posttreatment changes in several of these markers distinguished children with a declining serologic response suggestive of successful treatment from those with sustained serological responses in a 5-year follow-up study. A multivariate analysis demonstrated that lower frequency of CD4+CD45RA−CCR7−CD62L− T cells prior to drug therapy was an independent indicator of successful treatment.
Conclusions: These findings further validate the usefulness of alternative metrics to monitor treatment outcomes. Distinct qualitative and quantitative characteristics of T cells prior to drug therapy may be linked to treatment efficacy.
Introduction
Therapy with benznidazole or nifurtimox is recommended during both acute and early chronic phases of Trypanosoma cruzi infection, and the efficacy of treatment is typically assessed by tracking a decline in serological responses over time posttreatment. The rate of conversion to negative serology by conventional tests is high when treatment is provided during the first year of life (1), and the treatment of 5- to 14-year-old children resulted in a slower decay of T. cruzi-specific antibodies over time (2, 3). However, treatment in adult chronic patients is not widely used mainly because of the lack of early metrics of treatment efficacy. Direct parasite detection is difficult prior to treatment, and thus, failure to detect parasites after treatment is not a failproof way to determine efficacy. We have shown that the use of a multiplex-based serologic assay and the monitoring of T-cell responses were more sensitive tools to evaluate treatment efficacy in adult patients (4, 5).
A detailed characterization of the T-cell compartment in children in relatively early phases of the chronic infection showed that in contrast to adults with longer-term infections, T. cruzi-infected children have more broadly functional T. cruzi-specific T cells and exhibit fewer indicators of immune exhaustion (6, 7). These findings prompted the question of whether the surrogate parameters of treatment efficacy previously assessed in T. cruzi-infected adult subjects might change more rapidly in children. In the present study, we serially monitored cellular and humoral immune responses and parasitemia after the etiological treatment of children in the early chronic phase of T. cruzi infection. We found that posttreatment changes in a set of immune parameters distinguished children with declining serologic responses, indicative of successful treatment, from those with sustained serological responses after therapy.
Materials and Methods
Study Subjects
Five- to Sixteen-year-old children were enrolled at the Instituto Nacional de Parasitología Dr. Mario Fatala Chaben (Buenos Aires, Argentina) and at the Hospital Eva Perón (Buenos Aires, Argentina). T. cruzi infection was determined by indirect immunofluorescence (IIF), hemagglutination (IHA) and ELISA assays (8). Age- and sex-matched children with negative serological findings were recruited as uninfected controls (Table 1). Etiological treatment consisted of 5 mg/kg per day of benznidazole for 60 days or 10 mg/kg per day of nifurtimox for 60 days. Clinical, serological and immunological analyses were conducted prior to treatment, at 6 and 12 months following treatment, and at yearly intervals thereafter. Children with any impaired health condition were excluded from the present study. This protocol (No. 14-0004) was approved by the Institutional Review Boards of the Hospital Eva Perón, and Centro Nacional de Genética, Buenos Aires, Argentina. Informed written consent was obtained from the parents of all children included in the study, and written assent was also obtained from children older than seven years of age.

Table 1. Baseline characteristics of the study population.
Collection of Peripheral Blood Mononuclear Cells (PBMCs) and Sera
Approximately 10 mL of blood was drawn by venipuncture into heparinized tubes (Vacutainer; BD Biosciences). PBMCs were isolated by density gradient centrifugation on Ficoll-Hypaque (Amersham) and stored in liquid nitrogen at a density of 1.5 × 107 PBMCs/mL in newborn bovine serum containing 10% DMSO. On the day of the assay, PBMCs were thawed and washed in RPMI media containing 10% newborn bovine serum, 100 units/mL penicillin, 0.1 mg/mL streptomycin, 2 mM L-glutamine and 10 mM Hepes buffer. The cell viability was assessed by trypan blue staining with a viability range of 80–90%. An additional two milliliters of blood was collected, allowed to coagulate at 37°C and centrifuged at 1,000 g for 15 min for sera separation. Due to sample availability, assays were not run for all samples.
Antigens
Protein lysate from T. cruzi amastigotes was obtained by freeze/thaw cycles, followed by sonication as previously reported (9). Thirteen peptides 9–10 amino acids in length derived from trans-sialidase proteins and bearing high binding affinities for the six most common class I HLA supertypes were synthesized by Pepscan Systems (Lelystad, the Netherlands). Lyophilized peptides were dissolved at 10–20 mg/mL in DMSO, aliquoted and stored at −20°C.
IFN-γ and IL-2 Enzyme-Linked Immunosorbent Spot (ELISPOT) Assays
The number of T. cruzi antigen-responsive IFN-γ- and IL-2-secreting T cells was determined by ex vivo ELISPOT using commercial kits (BD Biosciences), as previously described (4, 9, 10).
Briefly, cryopreserved PBMCs were seeded at a concentration of 4 × 105 cells/well in triplicate wells and stimulated with T. cruzi lysate (10 μg/mL), a peptide pool from the trans-sialidase protein family (5 μg/mL/peptide; 10), or with media alone for 16–20 h at 37°C and 5% CO2. The stimulation of PBMCs with 20 ng/mL of phorbol 12-myristate 13-acetate (PMA, Sigma) plus 500 ng/mL of ionomycin (Sigma) in media was used as a positive control for cytokine secretion. Spot-forming cells (SFCs) were automatically enumerated using the ImmunoSpot analyzer (CTL). The mean number of spots in triplicate wells was calculated for each condition, and the number of specific IFN-γ and IL-2-secreting T cells was calculated by subtracting the value of the wells containing media alone from the antigen-stimulated spot count. Responses were considered statistically significant if a minimum of 10 spots/4 × 105 PBMCs was present per well; additionally, this number was at least twice the value of that in the wells with media alone (5, 11).
Whole Blood Surface Staining Assays
For phenotypic analysis of total CD4+ and CD8+ T cells, 50 μL of whole blood was incubated with different combinations of antihuman-CD4 peridinin chlorophyll (PerCP, clone Leu-3a), anti-CD8 (PerCP, clone Leu-3a), anti-CD45RA fluorescein isothiocyanate (FITC, clone HI100), anti-CD45RA allophycocyanin (APC, clone HI100), anti-CD27 phycoerythrin (PE, clone M-T271), anti-CD28 (PE, clone CD28.2), anti-CD127 (PE, clone Hil-7R-M21), anti-HLA-DR (FITC, clone TU36) (BD Biosciences) and anti-KLRG1 (APC, clone 2F1) (e-Bioscience), followed by red cell lysis and white cell fixation in FACS Lysing Solution (Pharmingen). Typically, 500,000 lymphocytes were acquired on the FACSCalibur (Becton Dickinson Immunocytometry Systems) and analyzed by using FlowJo software (TreeStar, Inc.).
Multiplex Serodiagnostic Assay
Serum specimens were screened for antibodies reactive to a panel of nine recombinant T. cruzi proteins in a Luminex-based format, as previously described (4, 12). The serologic responses to each individual T. cruzi protein were decreased during the study period if the MFI declined by 50% relative to that of a pretherapy sample assessed concurrently.
Cytometric Bead Array (CBA)
CBA assays were conducted according to the manufacturer's instructions (BD Biosciences) using cell supernatants derived from T. cruzi-antigen-stimulated PBMCs or PBMCs cultured with media alone. Stimulation with PMA plus ionomycin served as a positive control to ensure that lymphocyte stimulation conditions were appropriate. The samples were acquired on a FACScalibur flow cytometer and analyzed by using BD CBA Software, version 1.4.
Quantitative T. cruzi DNA Amplification
Two and a half milliliters of peripheral blood collected from subjects after treatment with benznidazole was mixed with the same volume of 6 M guanidine hydrochloride and 0.2 M EDTA, pH 8 (GEB), and maintained at room temperature for 1 week and then at 4°C until further use. Sample DNA isolation and parasite quantification, amplifying a T. cruzi satellite sequence, were performed as previously described (13).
Statistics
The normality of the variable distribution was assessed by using the Kolmogorov-Smirnov criterion. The results are presented as medians or medians and interquartile ranges.
Differences between groups were evaluated by the Mann-Whitney U-test or Student's t-test. Univariate analysis was conducted by Fisher's exact test, Wilcoxon rank sum test or two-sample t-test, as appropriate. The serologic treatment response was defined as a significant decrease in two out of three conventional serologic assays (i.e., 30% reduction in ELISA titers, and a 2-fold dilution by IHA or IIF) and a 50% reduction in the reactivity of at least two proteins in the multiplex assay. For multivariate analysis, a principal component (PC) analysis was conducted to reduce the dimensionality of a large number of interrelated variables. The eigenvalue-one criterion (Kaiser criterion) was used for extracting relevant PCs (eigenvalue >0.7). The candidate markers in the final logistic regression analysis were transformed into log scale and odds ratios with 95% confidence intervals. Associations between continuous variables were assessed using the nonparametric Spearman's correlation coefficient test. To evaluate changes in immune parameters over time posttreatment compared to those at baseline, a linear mixed model with compound symmetry, time as a fixed effect and the serologic treatment response as an interaction term was used. Since a linear mixed model could not be applied to evaluate the changes of T. cruzi-specific antibodies, an ANOVA for repeated measures was applied for the available data. Two-tailed P < 0.05 were considered statistically significant. Statistical analysis was conducted using IBM SPSS Statistics v23.0 (IBM Corp) and the Analytical Software Statistix 8.0.
Results
Clinical Characteristics and Tolerance to Drug Treatment in Children in the Early Stages of Chagas Disease
The T. cruzi-infected group included 18 children born in areas endemic for T. cruzi infection and 34 children born in Buenos Aires, Argentina, where T. cruzi infection is not endemic. Prior to treatment, electrocardiography revealed eight seropositive and two seronegative children who showed abnormal findings not related to Chagas disease (Table 1).
Forty-five children were treated with benznidazole, and because benznidazole was not available in the country, seven additional children were treated with nifurtimox. Mild adverse drug reactions were observed in eight out of the 40 (20%) subjects under treatment with benznidazole, while five subjects (12.5%) showed severe side effects that resulted in treatment suspension. The five children who received incomplete benznidazole dosing were then treated with nifurtimox. Cutaneous rash and dermatitis were the main adverse drug reactions with benznidazole treatment, whereas nifurtimox was well tolerated. No electrocardiographic changes related to Chagas disease were observed during the follow-up period.
Monitoring of IFN-γ and IL-2 Secreting T Cells Responsive to T. cruzi Antigens Following Treatment With Benznidazole or Nifurtimox
In this study, we measured IFN-γ- and IL-2-producing T cells in response to T. cruzi antigens in 41 children treated with benznidazole or nifurtimox, with a median follow-up of 36 months (range 12–60 months). For the analysis, children were grouped according to pretreatment (i.e., baseline) IFN-γ ELISPOT responses to an amastigote lysate into those with positive IFN-γ ELISPOT responses, as defined in the Materials and Methods, and those with IFN-γ ELISPOT responses under background levels. Four different kinetics of changes in ELISPOT responses were observed following treatment. Relative to pretreatment, levels of T. cruzi-specific T cells producing IFN-γ were decreased below background levels in 12 children (Table 2, Group 1; Figure 1A), while in 4 children, IFN-γ T-cell levels decreased at least 3-fold following treatment (Table 2, Group 2; Figure 1B). Eight of the 12 children in whom IFN-γ-producing T cells posttreatment had become undetectable within 12 months after treatment showed a subsequent rebound in T-cell responses between 24 and 48 months posttreatment (Figure 1A). An increase in T-cell responses was also observed in subjects who tested negative for T. cruzi-induced IFN-γ-producing cells prior to treatment (Table 2, Group 3; Figure 1C). In contrast, a smaller proportion of children showed no alterations in T-cell responses following drug therapy (Table 2, Group 4; Figure 1D). In addition, IL-2-producing T cells changed in concert with IFN-γ+ T-cell responses (Figures 1A–D).
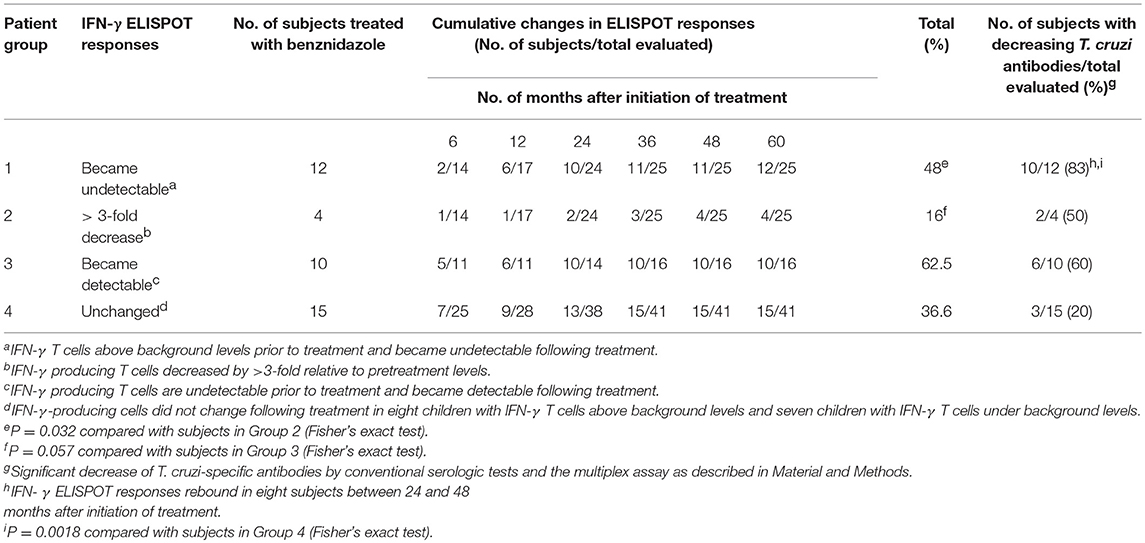
Table 2. Changes in IFN-γ ELISPOT T-cell responses specific for Trypanosoma cruzi antigens in children treated with benznidazole or nifurtimox.
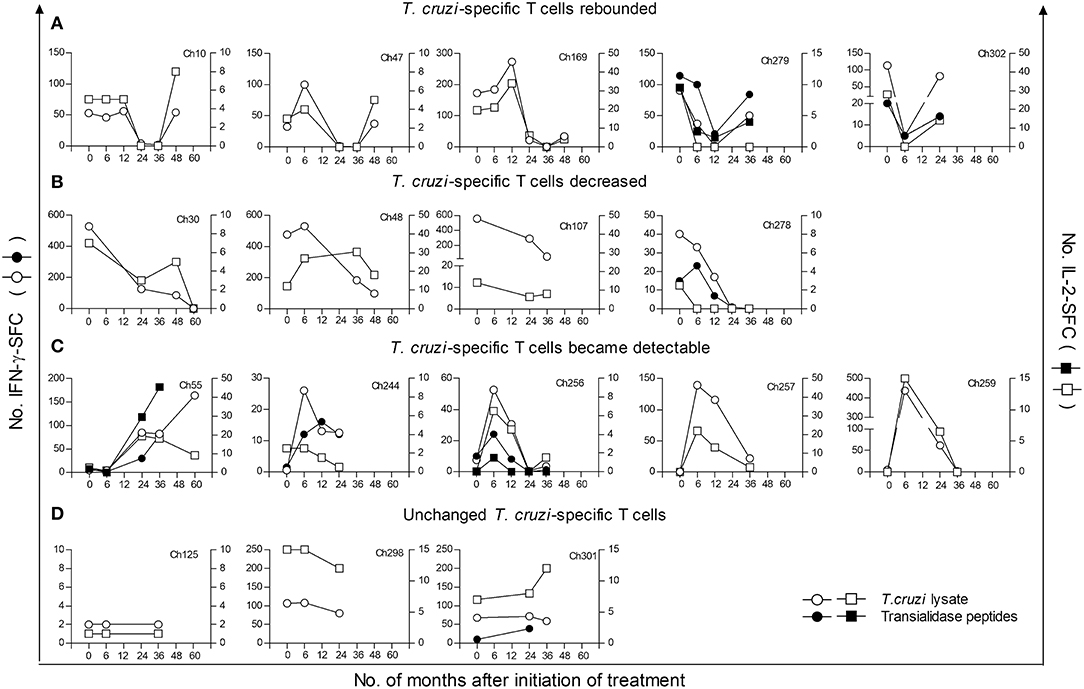
Figure 1. Monitoring of IFN–γ and IL 2-secreting cells in response to T. cruzi antigens in children at early stages of chronic Chagas disease following anti-T. cruzi therapy. IFN-γ- (circles) or IL-2- (squares) producing T cells were measured at different time points following treatment with benznidazole or nifurtimox. PBMCs were stimulated with a T. cruzi lysate (empty symbols) or a peptide pool from the trans-sialidase protein family (black symbols). The plots show representative data for single subjects. Time 0 indicates the assay point just prior to a 30-day treatment course. (A) Parasite-specific T-cell responses became undetectable after treatment followed by a rebound. (B) Parasite-specific T-cell responses decreased after treatment. (C) Previously undetectable cytokine-producing T cells prior to treatment became detectable after treatment. (D) The frequencies of cytokine producing T cells did not change relative to pretreatment. Subjects Ch257 and Ch244 were treated with nifurtimox, whereas the remaining patients received benznidazole.
T-cell responses specific for a set of HLA-I-restricted trans-sialidase peptides, which have been demonstrated as targets of CD8+ T-cell responses in chronic Chagas disease (10), were also measured in 28 children. These assays revealed that T. cruzi-specific CD8+ T-cell responses are modified after antiparasitic treatment (Figure 1A, Ch279 and Ch302; Figure 1B, Ch278; Figure 1C, Ch55, Ch244, and Ch256). Children treated with nifurtimox showed a similar pattern of T-cell responses compared to that of children treated with benznidazole (Table 2, Figure 1C, Ch257 and Ch244).
Cytokine and Chemokine Production Following Anti-T. cruzi Treatment
Next, we assessed the effect of anti-T. cruzi treatment on cytokine and chemokine production.
Cytokine concentrations representative of adaptive type 1, adaptive type 2, regulatory and proinflammatory profiles as well as chemokines were measured in the supernatant of PBMCs in response to stimulation with T. cruzi lysate or media alone. Prior to treatment, T. cruzi-infected children showed a mixed type1/type2/Treg profile, with increased levels of TNF-α, IL-6, and IL-10 compared with those of uninfected children (Figures 2A–C). T. cruzi-infected children also had increased concentrations of the inflammatory cytokines IL-1β (Figure 2D) and IL-8 (Figure 2E) and the chemokines IP-10, MCP-1 and MIG (Figures 2F–H) compared with those of the uninfected group. In addition, although the differences were not statistically significant, higher levels of RANTES were observed in T. cruzi-infected children than those in uninfected controls (Figure 2I). Among T. cruzi-infected subjects, no differences were observed between T. cruzi-stimulated cells and those stimulated with media alone, except for IL-1β, IP-10, and MIG (Figures 2D,F,H). In the experimental conditions assessed, IL-2, IL-4, IL-17, and IL-12p were not detected (data not shown).
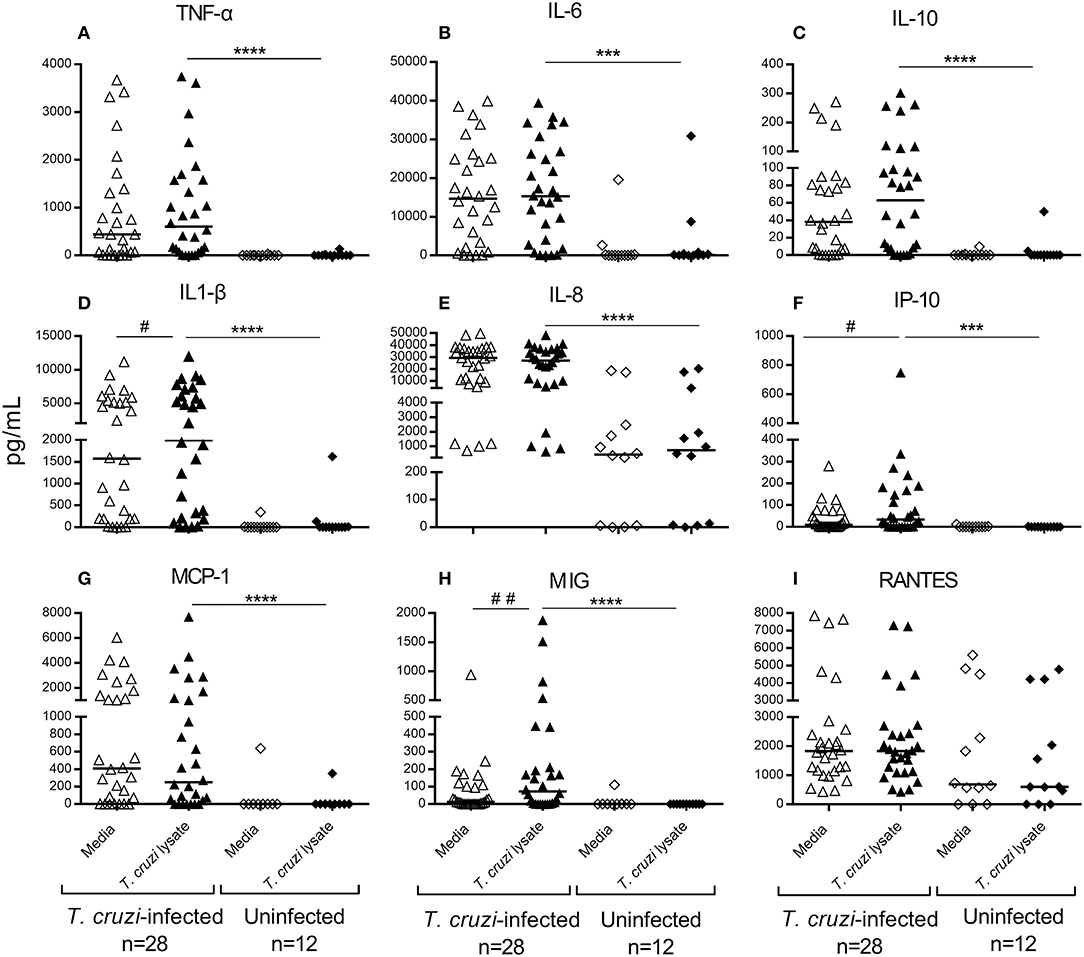
Figure 2. Cytokine and chemokine production in untreated children in early stages of Chagas disease. Type 1 (A), Type 2 (B), T-regulatory (C) and inflammatory cytokines (D,E) and chemokines (F–I) were measured in the supernatants of T. cruzi-stimulated or unstimulated PBMCs from T. cruzi-infected and uninfected children. Each circle represents the levels of cytokines/chemokines for each individual subject. Horizontal lines indicate median values. ##P < 0.001 and #P < 0.05, vs. T. cruzi lysate-stimulated wells of children seropositive for T. cruzi infection by Wilcoxon signed rank test. ****P < 0.0001 and ***P < 0.001, vs. T. cruzi lysate-stimulated wells of uninfected subjects by the Mann-Whitney U-test.
Because a positive correlation was found between the number of IFN-γ-producing cells and the concentration of cytokines and chemokines (Supplementary Table 1), the analysis of the changes of these molecules posttreatment was performed separately between patients who had positive IFN-γ-ELISPOT responses and those who had IFN-γ-producing cell levels below the background levels prior to treatment. Figure 3 shows the median concentration changes of the set of cytokines and chemokines evaluated. Among patients with positive baseline IFN-γ responses, the levels of IP-10 (Figure 3A), MCP-1 (Figure 3B) and MIG (Figure 3C) significantly decreased between 12 and 36 months posttreatment, whereas the levels of IL-1β, IL-6, IL-8, IL-10, TNF-α, and RANTES did not change posttreatment (data not shown). In contrast, among subjects with baseline IFN-γ-producing cell levels below background frequencies, the levels of IL-1β, IL-6, IL-8, IL-10, MCP-1, and TNF-α increased at 12 months posttreatment and decreased thereafter (Figures 3D–I), whereas those of IP-10, MIG and RANTES remained unaltered posttreatment (data not shown). These changes in posttreatment profiles were similar between T. cruzi-stimulated samples and those derived from PBMCs with media alone.
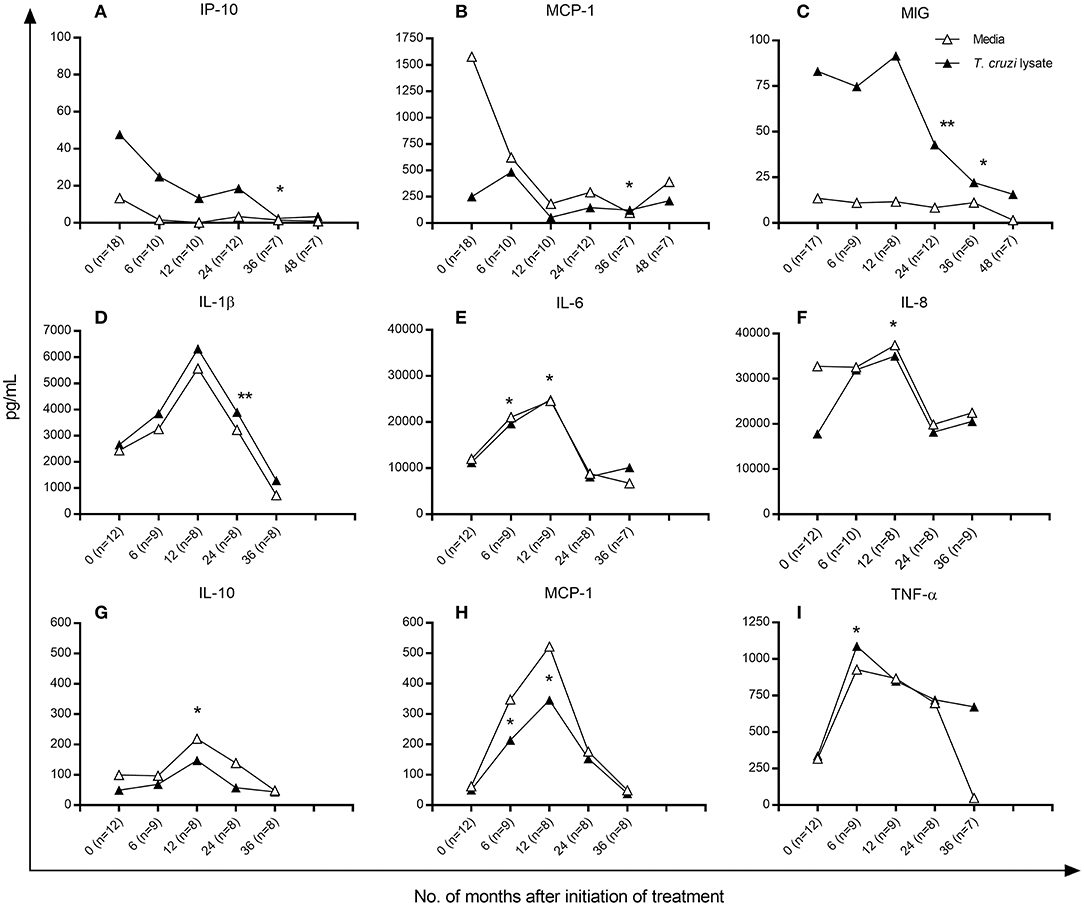
Figure 3. Cytokine and chemokine production in T. cruzi-infected children following anti-T. cruzi therapy. Cytometric bead analysis was used to measure the concentrations of Type 1, Type 2 and T-regulatory cytokines and chemokines in the supernatants of T. cruzi-stimulated (black symbols) and unstimulated (empty symbols) PBMCs at different time points following treatment with benznidazole or nifurtimox. Median values over time are shown for each variable measured. (A–C) Subjects with positive baseline IFN-γ ELISPOT responses as indicated in the Materials and Methods. (D–I) Subjects with baseline IFN-γ-producing cells under background levels. Changes from baseline (time 0) were evaluated by a linear mixed model for repeated measures. **P < 0.001 and *P < 0.05, vs. pretreatment values.
Differentiation, Activation, and Antigen-Experienced Profile of Total T Cells in Treated Children
We then evaluated whether antiparasitic therapy may revert the high degree of activation and differentiation of T cells observed in T. cruzi-infected children (6, 7, 14). Total effector (CD45RA+CD28− and CD45RA+CCR7−CD62L−), activated (HLA-DR+) and antigen-experienced (CD45−KLRG-1+CD127−) CD4+ T cells were decreased after treatment with benznidazole or nifurtimox, reaching values similar to those of uninfected subjects within 24 months of follow-up (Figures 4A–D, Supplementary Figure 1). A decline in TCR stimulation posttreatment was also evidenced by an increase in the frequency of total CD4+ T cells expressing CD127, which serves as the IL-7 receptor (Figure 4E), and a decrease in CD4+CD127− T cells (Figure 4F). Etiological treatment also induced a reduction in effector (CD45RA+CD27−) (Figure 5A), effector memory (CD45RA−CD127−) (Figure 5B), CD127− (Figure 5C) and activated (HLA−DR+) (Figure 5D) CD8+ T cells.
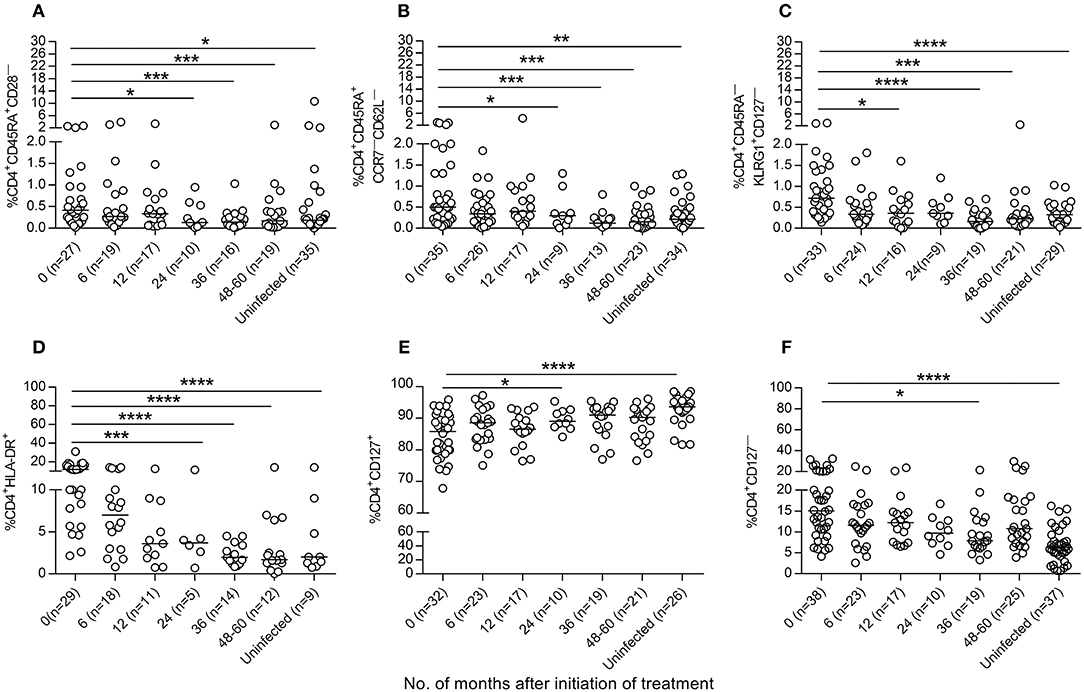
Figure 4. Phenotypic profiles of CD4+T cells in T. cruzi-infected children following drug treatment. PBMCs were stained with the indicated monoclonal antibodies and analyzed by flow cytometry. Lymphocytes were gated based on forward scattering and side scattering. CD4+ T cells were then selected and analyzed for the different T-cell phenotypes. (A–F) Each point represents the percentage of CD4+ T cells expressing a particular phenotype for single subjects prior to and after treatment. Median values are represented by horizontal lines. ****P < 0.0001; ***P < 0.001, **P < 0.01, and *P < 0.05. Changes posttreatment from baseline (time 0) were evaluated by a linear mixed model for repeated measures.
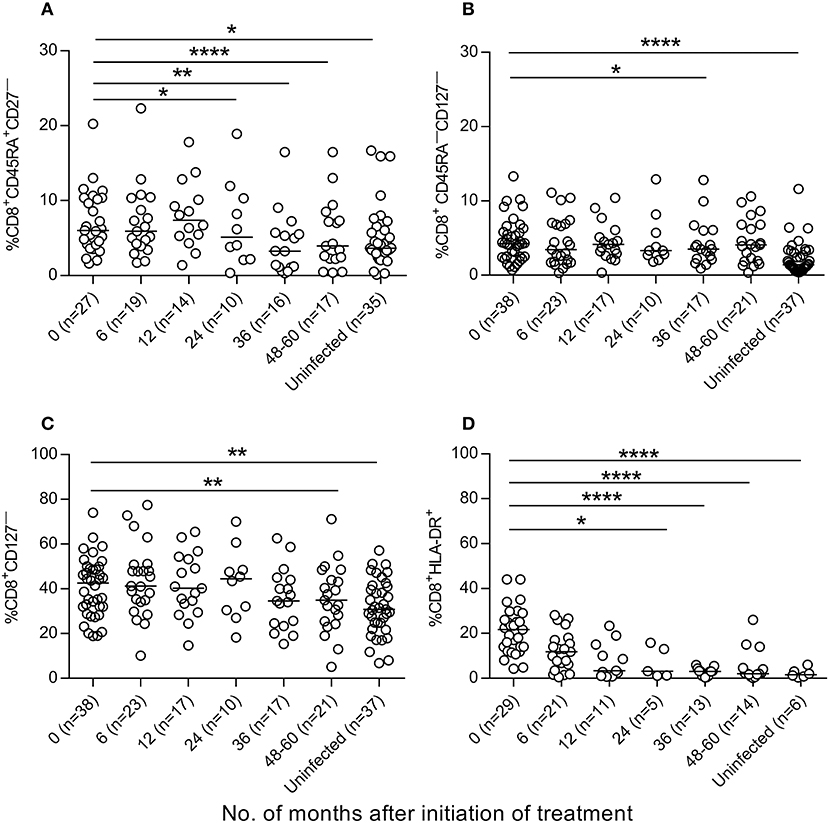
Figure 5. Phenotypic profiles of CD8+ T cells in T. cruzi-infected children following T. cruzi therapy. PBMCs were stained with the indicated monoclonal antibodies and analyzed by flow cytometry. Lymphocytes were gated based on forward scattering and side scattering. CD8+ T cells were then selected and analyzed for the different T-cell phenotypes. (A–D) each point represents the percentage of CD8+ T cells expressing a particular phenotype for single subjects prior to and after treatment. Median values are represented by horizontal lines. ****P < 0.0001; **P < 0.01, and *P < 0.05. Changes posttreatment from baseline (time 0) were evaluated by a linear mixed model for repeated measures.
T. cruzi-Specific Humoral Responses and Parasite Burden Following Anti-T. cruzi Treatment
Prior to treatment, all children were seropositive by the three conventional tests. T. cruzi-specific antibodies declined at 6 months following treatment (Table 3) and at the end of follow-up, 10 out of 52 (10.23%) treated children (i.e., nine children treated with benznidazole and one child treated with nifurtimox) were seronegative in at least 2 of 3 tests, with a median time of seroreversion of 48 months (range 12–60 months posttreatment), (Table 3).
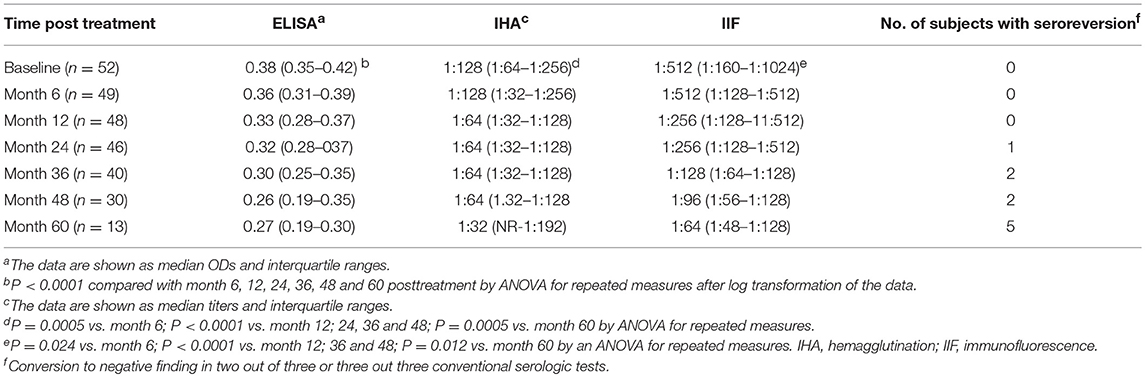
Table 3. Antibody titers by conventional serologic tests in T. cruzi-infected children treated with benznidazole or nifurtimox.
Prior to treatment, the sera from the 45 T. cruzi-infected children evaluated using a Luminex-based multiplex assay (4, 5, 12) recognized five proteins on average (ranging from 2 to 8 proteins). At 6 months following treatment, 17 of the 45 patients (37.3%) showed significant decreases in one or more proteins (2.8 proteins on average) (Figure 6, Supplementary Figure 2).
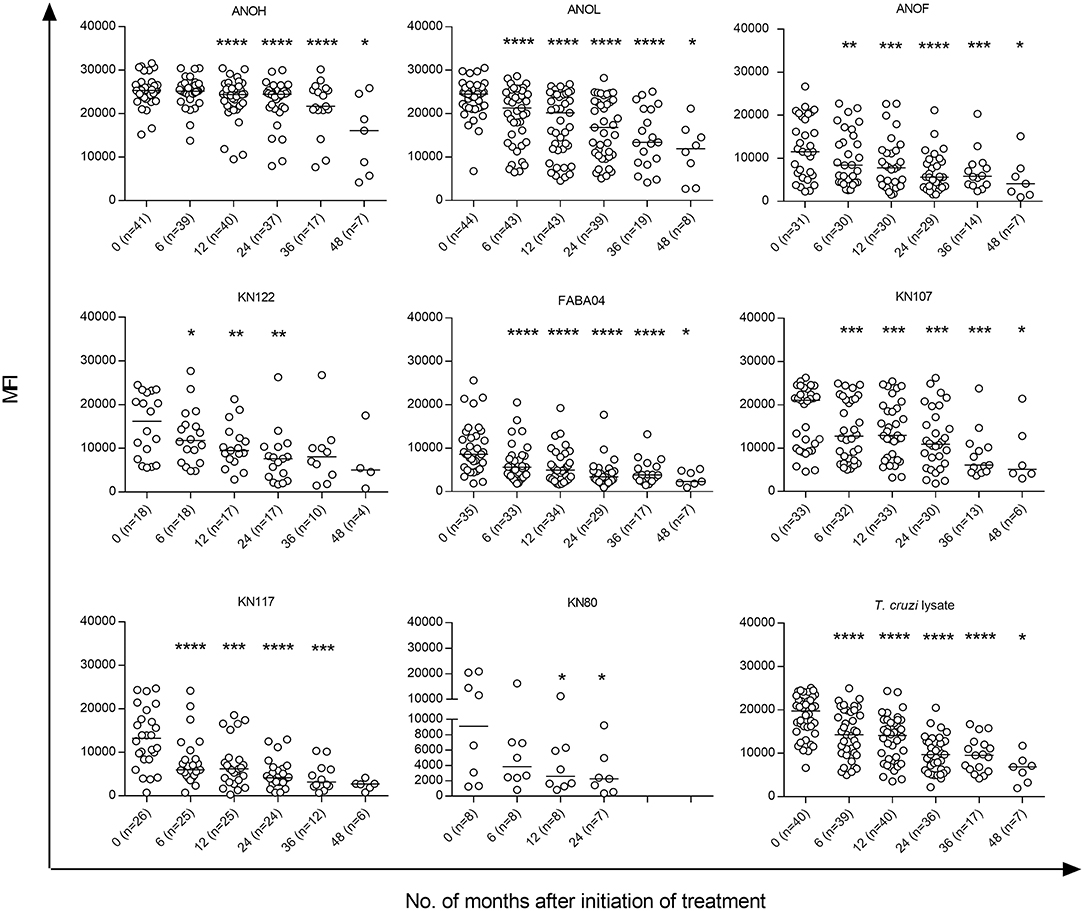
Figure 6. T. cruzi-specific humoral responses measured by the multiplex technique in children at early stages of chronic Chagas disease treated with benznidazole or nifurtimox. Serum specimens obtained at the indicated time points were screened using a bead array-based multiplex serological assay with recombinant T. cruzi proteins, as described in the Materials and Methods. Each point represents the mean fluorescence intensity (MFI) for reactive proteins for each individual analyzed prior to (time 0) and at several times points posttreatment. Median values are represented by horizontal lines. Decreases of T. cruzi-specific antibodies from baseline (time 0) were determined using ANOVA for repeated measures. ****P < 0.0001; ***P < 0.001; **P < 0.01, and *P < 0.05 vs. time 0.
Although the infection status of the subjects prior to treatment was not determined, quantitative PCR (qPCR) was used to detect T. cruzi DNA as a measure of treatment failure in posttreatment blood samples from 46 of the 52 children treated with benznidazole or nifurtimox. One (20 subjects) or two (26 subjects) blood samples were tested at a 1-year interval with a median follow-up of 48 months (ranging from 24 to 60 months) posttreatment. In all cases, the blood samples were negative for T. cruzi DNA.
Posttreatment Changes in Immune Parameters Associated With Declining Serologic Response
We then evaluated the association between treatment-dependent changes in the profile of cytokines, chemokines and T-cell phenotypes and the distribution of the serologic treatment outcomes. Considering reversion to negative serology and significant decreases in antibody titers by at least two conventional serologic tests and the multiplex assay, the subjects were classified as those with declining T. cruzi-specific antibody levels (i.e., effectively treated) or those with sustained antibody responses (i.e., ineffectively treated). The kinetics of the changes in IFN-γ-secreting cells as well as those of IL-1β, IL-6, IL-8, IL-10, MCP-1, TNF-α, and effector T cells following treatment were different between patients with dissimilar serologic treatment outcomes. A marked decline in IFN-γ-producing cells (Tables 2, 4) and in the frequency of effector CD4+CD45RA+CD28− T cells after therapy was associated with effective treatment (Table 4). As indicated by the estimate value, at 36 months posttreatment, the number of IFN-γ-producing cells was 1.74-fold higher in patients with ineffective treatment than that in effectively treated patients. IL-1β, IL-6, IL-8, IL-10, MCP-1, and TNF-α presented different kinetics associated with an effective treatment outcome, with an increase at 12 months posttreatment, followed by a decline. Children with ineffective treatment showed milder changes in these biomarkers (Table 4).
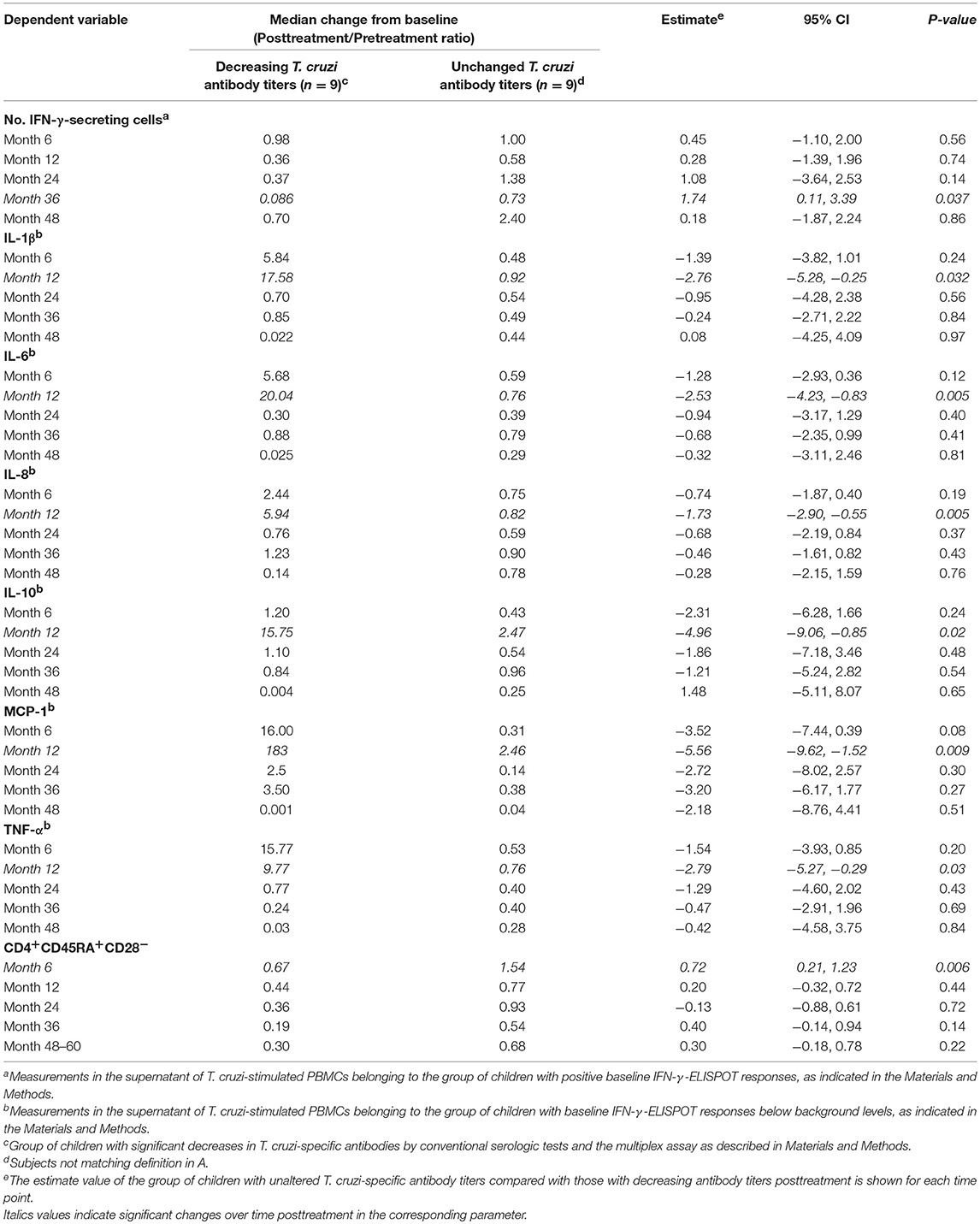
Table 4. Mixed model analysis of cytokine, chemokine and T cell-phenotype profiles for the association with serologic treatment response in children in the early stages of chronic Chagas disease following therapy with benznidazole or nifurtimox.
Baseline Immune Profiles Associated With Declining Serologic Response Posttreatment
To explore whether pretreatment immunological parameters were associated with serologic treatment outcomes, a univariate and multivariate regression analysis that was conducted using the reduction in the levels of T. cruzi-specific antibodies as the outcome revealed that younger age during treatment, lower TNF-α levels and lower percentages of effector memory CD4+ T cells were associated with effective treatment (Table 5). Although not statistically significant, higher frequencies of activated and differentiated memory CD4+ T cells, higher IL-10 levels and lower numbers of IFN-γ-producing cells were associated with ineffective treatment (Table 5).
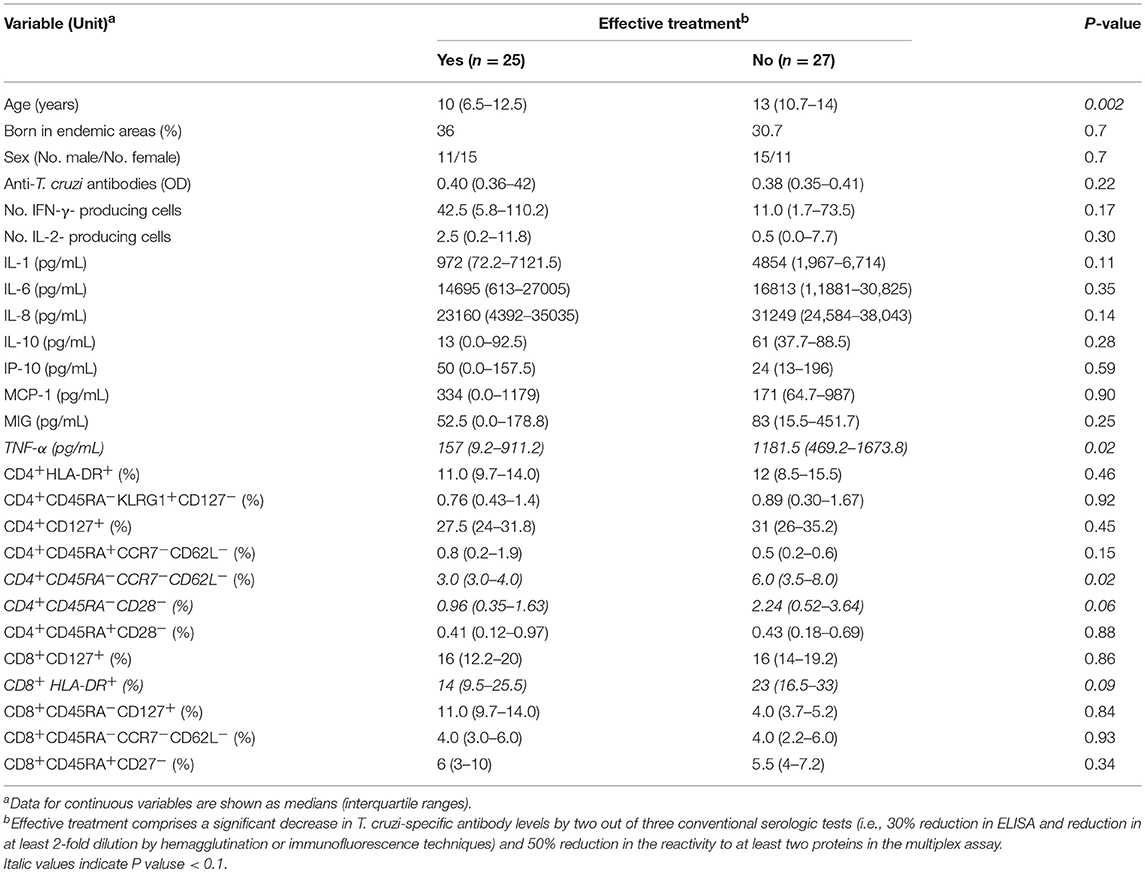
Table 5. Univariate analysis of baseline factors associated with serologic response in children in the early stages of chronic Chagas disease treated with benznidazole or nifurtimox.
When PC analysis was applied, we identified two distinct principal components (Supplementary Figure 3). PC1 was characterized by cytokines of the adaptive type 1, adaptive type 2 and T regulatory profiles, chemokines and effector CD4+ and CD8+ T-cell populations (accounting for 40.43% of the total variance in the data); PC2 comprised memory CD4+ and CD8+ T cells with different degree of differentiation, total activated CD4+ and CD8+ T cells, and IFN-γ-producing cells in response to T. cruzi antigens (accounting for 24.20% of total variance in the data). A logistic regression analysis constructed with immune parameters selected by the PC analysis (Supplementary Figure 3) showed that effector memory (CD45RA−CCR7−CD62L−) CD4+ T cells were an independent variable associated with effective treatment (P = 0.023, odds ratio = 0.60, and 95% CI = 0,39–0,93).
Discussion
Despite extensive efforts, early metrics of treatment efficacy in clinical practice for the management of chronic Chagas disease are still lacking. Treatment outcomes are considered to be better (i.e., more cures) and easier to assess in children, as titers of anti-T. cruzi antibodies decline more rapidly after effective treatment (1, 2, 3). Therefore, the present study focused on treatment outcomes specifically in children followed up for 60 months posttreatment with benznidazole or nifurtimox. We identified a broad range of cytokines and chemokines as well as T-cell phenotypes differentially modified in children presenting declining serologic responses indicative of successful treatment.
Anti-T. cruzi treatment in children induced significant changes in the frequencies of T. cruzi-responsive IFN-γ-producing cells similar to those observed in adult patients (4, 5). A posttreatment decrease to background levels in the number of IFN-γ-producing cells, followed by a rebound, was also observed in treated children. We previously showed that rebound CD4+ IFN-γ+ T cells are enriched in costimulatory functions compared with those present prior to treatment, consistent with the suppression of antigen load (5). Notably, changes in T-cell responses, either decreases or transient increases, were highly correlated with an early decline in T. cruzi-specific antibody levels, supporting the usefulness of monitoring parasite-specific T-cell responses during treatment. Likewise, the transient increase in the levels of the proinflammatory cytokines IL-1β, IL-6, IL-8, and TNF-α along with the transient increase in the levels of IL-10 and MCP-1 in patients with low baseline levels of these molecules were associated with a successful outcome of anti-T. cruzi therapy. The decline in the levels of MIG, IP-10 and MCP-1 following treatment supports a potential cessation in the recruitment of T cells and monocytes at target tissues.
Anti-T. cruzi therapy also induced a reconstitution of the phenotype of total CD4+ and CD8+ T cells with a clear decrease in the number of effector, activated, effector memory and memory T cells with low proliferative capacity, as well as an increase in the number of T cells expressing the α chain of the IL-7 receptor (i.e., the CD127 marker). The decreased expression of the IL-7 receptor and the presence of highly differentiated T cells are distinct features of T-cell exhaustion (15, 16). The responsiveness of T cells to IL-7 is essential for the maintenance of memory T cells (16). Chronically T. cruzi-infected adults have a predominance of monofunctional T. cruzi-specific T-cell responses and a perturbed Il-7/IL-7R pathway (9, 17). Although T. cruzi-infected children have polyfunctional parasite-specific T-cell responses compared with those of adults, the total T-cell compartment shows signs of T-cell exhaustion (7). Recently, it has been shown that inflammation perturbs the IL-7 axis, promoting T-cell exhaustion (18). Therefore, a lessened inflammatory environment following treatment would account for a more resting immune status of total T cells and the increase in the CD4+CD127+ T-cell levels observed herein. Of note, the decline in effector CD4+CD45RA+CD28− T-cell levels was associated with a decline in T. cruzi-specific antibody levels, further indicating a favorable treatment outcome in these children.
Sather Velar et al. showed that in children, treatment with benznidazole led to a high activation status of circulating monocytes (19) and a shift toward a type 1-modulated profile with increased CD8+IFN-γ+ counterbalanced by CD4+IL-10+ T cells (20). Other studies in children have also reported a decrease in the serum levels of IFN-γ (21) and soluble adhesion molecules (22) after treatment with benznidazole. An improvement in antigen-specific CD8+ T-cell responses (23) and a decline in activated T-cells have also been shown in adult patients treated with benznidazole (24, 25).
The rate of seroconversion to negative reactivity by conventional serologic tests was in the same range as that reported by other studies in a similar cohort of Argentinean benznidazole-treated children (2, 26, 27), but lower than that observed in Colombian children treated with nifurtimox (26). In addition, the rate of seroreversion to negative reactivity in children was not different from those reported in previous studies of adult patients treated with benznidazole, with a median follow-up of 36 months (i.e., 19% in children vs. 21% in adults) (27). However, a significant decrease in antibody levels at 6 months posttreatment by the multiplex assay and by conventional serologic tests was detected in children, an observation not generally found in adult patients (27). By the end of follow-up, the proportion of children with a decrease in T. cruzi-specific antibody levels by the multiplex assay reached 87.5% compared with the 68.7% of adult patients detected in our previous study (27). This faster decline in parasite-specific antibody levels, indicative of a favorable outcome, could explain the higher cure rates, determined by seroconversion to negative findings (i.e., 43–64%), reported for 5- to 14-year-old children with long-term follow-up (28, 29). Undetectable qPCR confirms that treatment failure was not evident in the present study group.
Because of the faster decline in antibody levels following benznidazole administration in children compared with that in adult subjects, we were able to set up clear outcomes of treatment in the present study group and explore the association between baseline measures of the different parameters evaluated and the outcome of the treatment. Effective treatment was observed in younger children in association with fewer signs of T-cell exhaustion and lower baseline levels of proinflammatory cytokines. Notably, the frequency of total effector memory (CD62L−CCR7−) CD4+ T cells was an independent indicator of successful treatment.
There may be several explanations for the association of a less-exhausted T-cell profile and successful treatment. In first place, a less-exhausted immune status in children might be accompanied by fewer survival niches for long-lived plasma cells, which might be responsible for maintaining the levels of circulating antibodies for long term, even in the absence of antigen stimulation (30, 31). The duration of these plasma cells survival niches and, consequently, of their resident plasma cells was likely terminated by the resolution of the focus. It is also likely that because T-cell responses in children are not exhausted (6, 7), the immune system has a chance to contribute to treatment efficacy, whereas in adults (9, 10, 32), there is a reduced chance for this contribution. In the experimental infection, benznidazole therapy was less efficacious in treating immunosuppressed mice (33, 34), and adjunct therapy with recombinant IL-12 enhanced the efficacy of benznidazole (35). However, further experimental studies are necessary to answer this question.
Collectively, this work documents that effective treatment outcomes in T. cruzi-infected children are accompanied by changes in parasite-specific T-cell responses and T-cell phenotypes supporting the beneficial effect of treatment in the early chronic phase of the infection.
Author Contributions
SAL, RT, and RV: designed research studies; MCA, AD, GC, HS, MAN, MC, MGA, JB, LF, KS, RV, and SAL: performed experiments, acquired data, analyzed data; MN: statistics; AS, MF, MGA, and BL: Patient care; SAL, RT, and MCA: drafted the manuscript; SAL: provided funding.
Funding
This work was supported by grants from The National Institutes of Health [Grants R01AI083188 and R01A I110346 to SAL]; the Ministerio de Salud de la Provincia de Buenos Aires, Argentina; and the Ministerio de Salud de la Nación, Argentina. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript. SAL, MCA, JB, and LF are members of the Scientific Career, Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET), Argentina. MAN and MC are CONICET Ph.D. fellows.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We thank the patients of the Instituto Nacional de Parasitología Dr. Mario Fatala Chaben and Hospital Eva Perón, Argentina, who provided blood samples, the Diagnostic Department of the Instituto Nacional de Parasitología Dr. Mario Fatala Chaben, Argentina, for serological tests and blood extractions, Pablo Viotti for data management, Claudia Nose for technical assistance with figures and Dr. Mario Chen of the DMID International Clinical Studies Support Center (ICSSC) at FHI 360, for biostatistics advice.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fimmu.2018.01958/full#supplementary-material
References
1. Altcheh J, Corral R, Biancardi MA, Freilij H. Anti-F2/3 antibodies as cure marker in children with congenital Trypanosoma cruzi infection. Medicina (B Aires) (2003) 63:37–40.
2. Sosa Estani S, Segura EL, Ruiz AM, Velazquez E, Porcel BM, Yampotis C. Efficacy of chemotherapy with benznidazole in children in the indeterminate phase of Chagas' disease. Am J Trop Med Hyg. (1998) 59:526–9. doi: 10.4269/ajtmh.1998.59.526
3. Sosa-estani S, Viotti R, Segura EL. Therapy, diagnosis and prognosis of chronic Chagas disease: insight gained in Argentina. Mem Inst Oswaldo Cruz (2009) 104(Suppl. 1):167–80. doi: 10.1590/S0074-02762009000900023
4. Laucella SA, Mazliah DP, Bertocchi G, Alvarez MG, Cooley G, Viotti R, et al. Changes in Trypanosomacruzi-specific immune responses after treatment: surrogate markers of treatment efficacy. Clin Infect Dis. (2009) 49:1675–84.doi: 10.1086/648072
5. Alvarez MG, Bertocchi G, Cooley G, Albareda MC, Viotti R, Perez-Mazliah DE, et al. Treatment success in Trypanosomacruzi infection is predicted by early changes in serially monitored parasite-specific T and B cell responses. PLoS Negl Trop Dis. (2016) 10:e0004657. doi: 10.1371/journal.pntd.0004657
6. Albareda MC, Olivera GC, De Rissio AM, Postan M. Assessment of CD8(+) T cell differentiation in Trypanosoma cruzi-infected children. Am J Trop Med Hyg. (2010) 82:861–4. doi: 10.4269/ajtmh.2010.09-0604
7. Albareda MC, De Rissio AM, Tomas G, Serjan A, Alvarez MG, Viotti R, et al. Polyfunctional T cell responses in children in early stages of chronic Trypanosoma cruzi infection contrast with monofunctional responses of long-term infected adults. PLoS Negl Trop Dis. (2013) 7(12):e2575. doi: 10.1371/journal.pntd.0002575
8. World Health Organization. Control of Chagas disease. World Health Organ Tech Rep Ser. (2012) 975:1–116.
9. Laucella SA, Postan M, Martin D, Hubby Fralish B, Albareda MC, Alvarez MG, et al. Frequency of interferon-gamma-producing T cells specific for Trypanosoma cruzi inversely correlates with disease severity in chronic human Chagas disease. J Infect Dis. (2004) 189:909–18. doi: 10.1086/381682
10. Alvarez MG, Postan M, Weatherly DB, Albareda MC, Sidney J, Sette A, et al. HLA class I-T cell epitopes from trans-sialidase proteins reveal functionally distinct subsets of CD8+ T cells in chronic Chagas disease. PLoS Negl Trop Dis. (2008) 2:e288. doi: 10.1371/journal.pntd.0000288
11. Lalvani A, Hill AV. Cytotoxic T-lymphocytes against malaria and tuberculosis: from natural immunity to vaccine design. Clin Sci. (1998) 95:531–8. doi: 10.1042/cs0950531
12. Cooley G, Etheridge RD, Boehlke C, Bundy B, Weatherly DB, Minning T, et al. High throughput selection of effective serodiagnostics for Trypanosoma cruzi infection. PLoS Negl Trop Dis. (2008) 2:e316. doi: 10.1371/journal.pntd.0000316
13. Bua J, Volta BJ, Perrone AE, Scollo K, Velázquez EB, Ruiz AM, et al. How to improve the early diagnosis of Trypanosoma cruzi infection: relationship between validated conventional diagnosis and quantitative DNA amplification in congenitally infected children. PLoS Negl Trop Dis. (2013) 7:e2476. doi: 10.1371/journal.pntd.0002476
14. Vitelli-Avelar DM, Sathler-Avelar R, Massara RL, Borges JD, Lage PS, Lana M, et al. Are increased frequency of macrophage-like and natural killer (NK) cells, together with high levels of NKT and CD4+CD25high T cells balancing activated CD8+ T cells, the key to control Chagas' disease morbidity? Clin Exp Immunol. (2006) 145:81–92. doi: 10.1111/j.1365-2249.2006.03123.x
15. Akbar AN, Henson SM. Are senescence and exhaustion intertwined or unrelated processes that compromise immunity? Nat Rev Immunol. (2011) 11:289–95. doi: 10.1038/nri2959
16. Wherry EJ, Kurachi M. Molecular and cellular insights into T-cell exhaustion. Nat Rev Immunol. (2015) 15:486–99. doi: 10.1038/nri3862
17. Albareda MC, Perez-Mazliah D, Natale MA, Castro-Eiro M, Alvarez MG, Viotti R, et al. Perturbed T cell IL-7 receptor signaling in chronic Chagas disease. J Immunol. (2015) 194:3883–9. doi: 10.4049/jimmunol.1402202
18. Shive CL, Clagett B, McCausland MR, Mudd JC, Funderburg NT, Freeman ML, et al. Inflammation perturbs the il-7 axis, promoting senescence and exhaustion that broadly characterize immune failure in treated HIV infection. J Acquir Immune. Defic Syndr. (2016) 71:483–92. doi: 10.1097/QAI.0000000000000913
19. Sathler-avelar R, Vitelli-Avelar DM, Massara RL, de Lana M, Pinto Dias JC, Teixeira-Carvalho A, et al. Etiological treatment during early chronic indeterminate Chagas disease incites an activated status on innate and adaptive immunity associated with a type 1-modulated cytokine pattern. Microbes Infect. (2008) 10:103–13. doi: 10.1016/j.micinf.2007.10.009
20. Sathler-avelar R, Vitelli-Avelar DM, Massara RL, Borges JD, Lana M, Teixeira-Carvalho A, et al. Benznidazole treatment during early-indeterminate Chagas' disease shifted the cytokine expression by innate and adaptive immunity cells toward a type 1-modulated immune profile. Scand J Immunol. (2006) 64:554–63. doi: 10.1111/j.1365-3083.2006.01843.x
21. Cutrullis RA, Moscatelli GF, Moroni S, Volta BJ, Cardoni RL, Altcheh JM, et al. Benzonidazole therapy modulates interferon-γ and M2 muscarinic receptor autoantibody responses in Trypanosoma cruzi-infected children. PLoS ONE (2011) 6:e27133. doi: 10.1371/journal.pone.0027133
22. Laucella SA, Segura EL, Riarte A, Sosa ES. Soluble platelet selectin (sP-selectin) and soluble vascular cell adhesion molecule-1 (sVCAM-1) decrease during therapy with benznidazole in children with indeterminate form of Chagas' disease. Clin Exp Immunol. (1999) 118:423–7. doi: 10.1046/j.1365-2249.1999.01070.x
23. Mateus J, Pérez-Antón E, Lasso P, Egui A, Roa N, Carrilero B, et al. Antiparasitic treatment induces an improved cd8+ t cell response in chronic chagasic patients. J Immunol. (2017) 198:3170–80. doi: 10.4049/jimmunol.1602095
24. Perez-Mazliah DE, Alvarez MG, Cooley G, Lococo BE, Bertocchi G, Petti M, et al. Sequential combined treatment with allopurinol and benznidazole in the chronic phase of Trypanosoma cruzi infection: a pilot study. J Antimicrob Chemother. (2013) 68:424–37. doi: 10.1093/jac/dks390.
25. Vallejo A, Monge-Maillo B, Gutiérrez C, Norman FF, López-Vélez R, Pérez-Molina JA. Changes in the immune response after treatment with benznidazole versus no treatment in patients with chronic indeterminate Chagas disease. Acta Trop. (2016) 164:117–24. doi: 10.1016/j.actatropica.2016.09.010
26. Bianchi F, Cucunubá Z, Guhl F, González NL, Freilij H, Nicholls RS, et al. Follow-up of an asymptomatic Chagas disease population of children after treatment with nifurtimox (Lampit) in a sylvatic endemic transmission area of Colombia. PLoS Negl Trop Dis. (2015) 9:e0003465. doi: 10.1371/journal.pntd.0003465
27. Viotti R, Vigliano C, Alvarez MG, Lococo B, Petti M, Bertocchi G, et al. Impact of aetiological treatment on conventional and multiplex serology in chronic Chagas disease. PLoS Negl Trop Dis. (2011) 5:e1314. doi: 10.1371/journal.pntd.0001314
28. Sosa-Estani S, de Bravo BH, de Bizzoto LH, Canil S, Cura EN, Segura EL. Evolución serológica a largo plazo en niños infectados por Trypanosoma cruzi que cursan fase clínica indeterminada, tratados con benznidazol. Available online at: http://www.fac.org.ar/fec/chagas2/llave/md8/md804/sosaes.htm. (Accessed November 21, 2017).
29. Streiger ML, del Barco ML, Fabbro DL, Arias ED, Amicone NA. Longitudinal study and specific chemotherapy in children with chronic Chagas' disease, residing in a low endemicity area of Argentina. Rev Soc Bras Med Trop. (2004) 37:365–75.
30. Kunkel EJ, Butcher EC. Plasma-cell homing. Nat Rev Immunol. (2003) 3:822–9. doi: 10.1038/nri1203
31. Tiburzy B, Szyska M, Iwata H, Chrobok N, Kulkarni U, Hirose M, et al. Persistent autoantibody-production by intermediates between short-and longlived plasma cells in inflamed lymph nodes of experimental epidermolysis bullosa acquisita. PLoS ONE (2013) 8:e83631. doi: 10.1371/journal.pone.0083631
32. Albareda MC, Olivera GC, Laucella SA, Alvarez MG, Fernandez ER, Lococo B, et al. Chronic human infection with Trypanosoma cruzi drives CD4+ T cells to immune senescence. J Immunol. (2009) 183:4103–08. doi: 10.4049/jimmunol.0900852
33. Toledo MJO, Machado GBN, Pereira MES, Brener Z. Results of treatment in mice immunosuppressed inoculated with different Trypanosoma cruzi strains. Mem Inst Oswaldo Cruz (1991) 86:237.
34. Romanha AJ, Alves RO, Murta SM, Silva JS, Ropert C, Gazzinelli RT. Experimental chemotherapy against Trypanosoma cruzi infection: essential role of endogenous interferon-gamma in mediating parasitologic cure. J Infect Dis. (2002) 186:823–8. doi: 10.1086/342415
35. Michailowsky V, Murta SM, Carvalho-Oliveira L, Pereira ME, Ferreira LR, Brener Z, et al. Interleukin-12 enhances in vivo parasiticidal effect of benznidazole during acute experimental infection with a naturally drug-resistant strain of Trypanosoma cruzi. antimicrob Agents Chemother. (1998) 42:2549–56.
Keywords: Trypanosoma cruzi, T cells, benznidazole, nifurtimox, pediatric infection
Citation: Albareda MC, Natale MA, De Rissio AM, Fernandez M, Serjan A, Alvarez MG, Cooley G, Shen H, Viotti R, Bua J, Castro Eiro MD, Nuñez M, Fichera LE, Lococo B, Scollo K, Tarleton RL and Laucella SA (2018) Distinct Treatment Outcomes of Antiparasitic Therapy in Trypanosoma cruzi-Infected Children Is Associated With Early Changes in Cytokines, Chemokines, and T-Cell Phenotypes. Front. Immunol. 9:1958. doi: 10.3389/fimmu.2018.01958
Received: 27 April 2018; Accepted: 08 August 2018;
Published: 13 September 2018.
Edited by:
Jason Paul Gigley, University of Wyoming, United StatesReviewed by:
Phileno Pinge-Filho, Universidade Estadual de Londrina, BrazilThomas Jacobs, Bernhard-Nocht-Institut für Tropenmedizin (BMITM), Germany
Copyright © 2018 Albareda, Natale, De Rissio, Fernandez, Serjan, Alvarez, Cooley, Shen, Viotti, Bua, Castro Eiro, Nuñez, Fichera, Lococo, Scollo, Tarleton and Laucella. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Susana A. Laucella, c2xhdWNlbGxhQHlhaG9vLmNvbQ==