 Ángel J. Zamora-Ramírez
Ángel J. Zamora-Ramírez Moisés Santillán
Moisés Santillán- Centro de Investigacion y de Estudios Avanzados del IPN (Cinvestav), Unidad Monterrey, Apodaca, Mexico
There is growing evidence that mitochondrial dynamics play a role in mitochondrial function quality control. Recent studies have discovered that physical activity can aid in maintaining a healthy mitochondrial population and may even counteract the mitochondrial damage caused by obesity by modifying mitochondrial dynamics processes like mitogenesis, fission, and mitophagy. However, the underlying mechanisms and how they influence the various processes of mitochondrial dynamics have not been completely clarified. This study seeks to make an initial effort to fill this gap by utilizing mathematical modeling to investigate how modifications to the processes involved in mitochondrial dynamics can aid in preserving a healthy mitochondrial population under conditions of accelerated damage. We found by analyzing the model stationary behavior that fission and mitophagy are strictly required for a healthy mitochondrial population, that increasing the rate of mitochondrial damage has a negative impact on the population of functional mitochondria, and that such damage can be mitigated to some extent by increasing fission and/or mitophagy rates. These findings are finally discussed in the context of the benefits of physical activity for mitochondrial population health in obese individuals.
1. Introduction
Mitochondria are membrane-bound cell organelles that generate the majority of the chemical energy required to power the biochemical reactions of the cell. They are, however, much more than that, as they are involved in other critical processes such as apoptosis, calcium homeostasis, and heme synthesis [1].
In living cells, mitochondria form a dynamic reticulum that is constantly remodeled by fusion and fission events. These processes are referred to as mitochondrial dynamics [2], along with mitochondrial replication (mitogenesis) and degradation (mitophagy). There is growing evidence that mitochondrial dynamics are important in the quality control of mitochondrial function [3–5]. According to this evidence, fission followed by selective fusion segregates dysfunctional mitochondria and allows mitophagy to remove them.
There is a complex relationship between mitochondrial dysfunction and obesity [6, 7]. Obesity, in short, causes a chronic inflammatory state that increases the production of reactive oxygen species (ROS) and provokes oxidative stress. In turn, oxidative stress causes mitochondrial damage, which leads to mitochondrial dysfunction. Furthermore, an abundance of nutrients (which is typically the cause of obesity) can overwhelm the Krebs cycle and the mitochondrial respiratory chain, impairing mitochondrial function. Finally, dysfunctional mitochondria produce an abundance of ROS, exacerbating the inflammatory process in obesity.
Physical exercise, in conjunction with lifestyle changes, is widely recognized as the most effective non-pharmacological intervention for improving cardiorespiratory fitness and managing chronic diseases. Recent research has shown that exercise can counteract the negative effects of obesity on mitochondria and foster the growth of a healthy mitochondrial population by altering processes such as mitogenesis, fission, and mitophagy [8–13]. However, the underlying mechanisms and their impact on the mitochondrial dynamics processes have not been fully elucidated. This study seeks to make an initial effort to fill this gap by utilizing mathematical modeling to investigate how modifications to the processes involved in mitochondrial dynamics can aid in preserving a healthy mitochondrial population under conditions of accelerated damage.
Numerous works have developed computational and mathematical models to address various problems related to mitochondrial dynamics; see Tam et al. [14], Patel et al. [4], Kowald and Klipp [1], and Kornick et al. [15], for example. We are particularly interested in the agent-based model developed by Patel et al. [4] because it specifically addresses the question of how mitochondrial dynamics can be optimized for quality control in mitochondrial networks. The model presented here incorporates the same regulatory mechanisms as the Patel et al. [4] model, but instead of modeling individual mitochondria, it employs ordinary differential equations to track the dynamics of mitochondrial populations. Although this approach implies a lower level of detail (especially in terms of spatial information), it has the advantage of allowing its stationary solutions to be studied analytically.
2. Methods
2.1. Brief review of the mitochondrial dynamics processes
Apart from replicating through mitogenesis and being degraded through processes such as mitophagy, mitochondria are constantly undergoing fission and fusion in order to adapt their morphology to the cellular environment and energetic demands [2]. That is, mitochondrial dynamics are governed by two sets of opposing processes: mitochondrial fusion and fission, as well as mitochondrial biogenesis and degradation; with other processes such as intracellular transport also playing an important role. Overall, mitochondrial dynamics processes maintain homeostasis, regulate mitochondrial form, volume, and function, and are increasingly recognized as critical components of the cellular stress response [16].
Mitochondrial fusion is the physical merging of two or more previously distinct mitochondria's outer and then inner mitochondrial membranes. Fusion results in networks with continuous membranes and a matrix lumen. As a result, each network's constituents share solutes, metabolites, and proteins [3]. Recent in vitro fluorescence microscopy studies [3] have also shown that fusion preferentially involves mitochondria with high electrochemical potential across their inner membrane (Ψm). All of this suggests that fusion is a mechanism by which intact mitochondria can supplement a damaged one and possibly restore its activity, preserving metabolic efficiency.
Mitochondrial fission is the splitting of a single mitochondrial body into two. According to in vitro studies, approximately 30% of fission events resulted in a statistically significant difference in the membrane potentials of the two daughter cells [3]. The mechanism underlying this asymmetry is unknown, but it has a significant impact on the relative capacities for ATP and reactive oxygen species (ROS) production of the two daughter mitochondria [17].
Mitophagy is the selective degradation of mitochondrial by autophagy. It frequently follows damage or stress events because it targets depolarized mitochondria for digestion and elimination [18].
Mitogenesis, also known as mitochondrial biogenesis, is the process by which new mitochondria are formed through the growth and division of existing mitochondria [19]. This process is strictly governed by intracellular or extracellular signals that communicate an energy imbalance caused by increased energy demand and/or decreased energy production.
According to Twig et al. [3], mitochondrial fusion-fission and autophagy form a quality-control axis that maintains mitochondrial health by segregating dysfunctional mitochondria into a separate pool of mitochondria that are unable to fuse and undergo autophagy.
2.2. Mathematical modeling
Based on the previous discussion, we propose the following conceptual model for mitochondrial dynamics. First, we assume that the total mitochondrial volume density (the percentage of muscle fibber volume occupied by mitochondria) can be classified into four different groups: healthy or little damaged (H), moderately damaged (M), severely damaged (S) and fused (F) mitochondria. Given that cell division is energetically demanding, dysfunctional (severely damaged) mitochondria replicate at a much smaller rate than functional ones, and the persistence or their damaged genome can cause severe problems. However, we assume for the sake of simplicity that only healthy and moderately damaged mitochondria can proliferate. Concomitantly, the model takes into consideration that the volume density of healthy and moderately damaged mitochondria increases due to mitogenesis. The Lotka-Volterra model for interspecific competition between species, in which all four mitochondria subpopulations compete for resources, is employed to model mitogenesis. Mitochondria are subject to constant damage due to multiple factors, like oxidative stress. This process is modeled as linear fluxes from compartment H to compartment M, and from compartment M to compartment S. Severely damaged mitochondria are degraded via mitophagy; a process that is also assumed to be linear. Given that fusion is Ψm-dependent (with Ψm the potential across the mitochodrial inner membrane), we assume that only healthy and moderately damaged mitochondria may fuse. The rate at which fusion takes place is assumed to be proportional to the mitochondrial volume density. To account for the fact that mitochondria randomly exchange material while fused, we assume that fission gives rise to equal amounts of healthy, moderately damaged and severely damaged mitochondria. The rate at which fission occurs is assumed to be directly proportional to the volume density of fussed mitochondria.
The mitochondrial dynamics clockwork depicted in the formed paragraph (which is schematically represented in Figure 1) can be translated into the following set of Ordinary Differential Equations (ODEs):
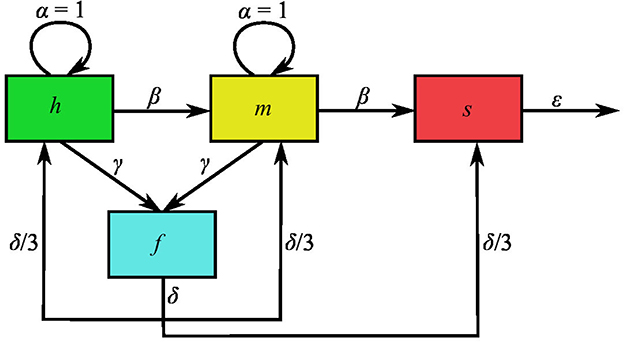
Figure 1. Schematic representation of the mathematical model variables and interactions. Variables h, m, s, and f denote the volume density of healthy, moderately damaged, severely damaged and fused mitochondria, normalized to the carrying capacity, K. Moreover, the intrinsic-mitogenesis, damaging, fusion, fission, and mitophagy rate are respectively represented by parameters α, β, γ, δ, and ϵ. These parameters are normalized to the intrinsic mitogenesis rate, which is why α = 1.
With N = H + M + S + F. In the equations above, H, M, S, and F, respectively, stand for the volume density of healthy or little damaged, moderately damaged, severely damaged, and fused mitochondria, while N denotes the total mitochondrial volume density. The first terms in the right hand side of Equations (1), (2) represent mitogenesis flows into the populations of healthy and moderately damaged mitochondria. The expressions for these flows implicitly assume that mitochondrial growth is logistic and that all mitochondrial subpopulations compete for resources with the same efficiency—see for instance [20], Section 3.5. Accordingly, parameter a represents the intrinsic mitogenesis rate (i.e. the rate of mitogenesis when resources are abundant), while K is the volume-density carrying capacity (the maximum total mitochondrial volume density cells can sustain). Finally, taking into consideration that the rest of the flows accounted for in Equations (1)–(4) are linear, the meaning of the corresponding parameters is as follows. Parameter b is the mitochondrial damaging rate, c is the fusion rate for healthy and moderately damaged mitochondria, d is the fission rate, and e is the mitophagy rate.
It is convenient to work with non-dimensional variables and parameters to simplify the analysis of the model dynamic behavior. To this end, define the following normalized variables:
and parameters:
With this, Equations (1)–(4) can be rewritten as
Where x′ denotes the derivative of variable x with respect to time t′. Notice that variables h, m, s, and f correspond to the volume density of healthy, moderately damaged, severely damaged and fused mitochondria, normalized to the carrying capacity, K. Moreover, parameters β, γ, δ and ϵ, respectively, stand for the damaging, fusion, fission and mitophagy rates, normalized to the intrinsic mitogenesis rate.
3. Results
3.1. No-fission and no-mitophagy particular cases
It is straightforward to verify that is a steady state of the ordinary differential equation system defined in Equations (5)–(8).
To find other positive fixed points, let us start analyzing two specific cases: no fission (δ = 0) and no mitophagy (ϵ = 0).
In the no fission case, Equations (5)–(8) become
Let (h*, m*, d*, f*) denote a stationary solution. Then, it must satisfy:
Under the assumption that h* > 0, Equation (9) implies that 1 − n* = β + γ. However, when this last result is substituted into Equation (10), it yields h* = 0. Furthermore, substitution into Equations (12), (11) demonstrates that m* = s* = 0, while f can take arbitrary values that are ultimately determined by the initial condition. In practice, having only a population of fussed mitochondria is unachievable and so the trivial steady state is the only one available for the system in the no fission case.
Consider now the no mitophagy case (ϵ = 0). Under this assumption, the ordinary differential equation (ODE) system given by Equations (5)–(8) becomes
The equations determining the steady states in this case are
Since all model parameters are positive, it follows from Equation (15) that m*, f* = 0. These results, together with Equation (16) further imply that h* = 0, while the value of s* is determined by the initial condition. In practice, solely having severely damaged mitochondria is physiologically unattainable. Hence no other steady state than the trivial one exists in the no mitophagy case.
3.2. Nontrivial steady state behavior
We have seen that in the absence of fission and/or mitophagy, the only available steady state in practice is the trivial one. This suggests that such processes are required for the maintenance of a functional mitochondial network. To gain a better understanding of the significance of these parameters, let us look for positive fixed points in the general normalized model.
The algebraic equations determining the stationary solution of the ODE system given by Equations (5)–(8) are:
By solving for f* in Equation (20) and substituting into Equations (17), (18) we get a couple of equations involving variables h*, m* and f*. If, for the sake of algebraic manipulation n* is regarded as a constant parameter, such equations result to be a complete set of linear homogeneous equations for variable h* and m*. Hence, nontrivial solutions only exist when the determinant of the coefficient matrix equals zero. By doing the corresponding algebra, this last constrains yields a equation for n* whose positive solution is:
Furthermore, by taking into consideration that the nontrivial solutions of the previously described homogeneous equation system are of the form
we get:
By substituting this last result into Equation (20) and solving for f* we obtain:
Finally, substitution of this last result into Equation (19) allows us to solve for s* to get:
If we now substitute Equations (21)–(24) into the definition of n* (n* = h* + m* + s* + f*), we can solve for h* to obtain an expression for this variable in terms of the model parameters. Further substitution into Equations (22)–(24) allows us to do the same for variables m*, s* and f*. The resulting algebraic expressions are omitted for the sake of brevity. Nonetheless the can be straightforwardly obtained and they constitute a nontrivial positive solution for the complete model normalized ODE system—Equations (5)–(8).
It is not hard to verify that this solution is consistent with the special cases studied in the former subsection. That is, h* = m* = s* = 0 in the absence of fission (δ = 0), whereas f* can take any value. However, one can argue from biological considerations that is has to be f* = 0 (no fused mitochondria can be obtained without healthy of moderately damaged ones). Similarly, h* = m* = f* = 0 without mitophagy (ϵ = 0), while the value of s* is arbitrary from a mathematical standpoint, but null from biological considerations.
Another interesting result is that, according to Equation (21), the total mitochondrial volume density is only a function of the damaging (β) and fusion (γ) rates (normalized to the mitogenesis rates). In other words, it is independent of fission and mitophagy. Thus, our model predicts that modifying the rates of fission (δ) or mitophagy (ϵ) would change the way mitochondria distribute among the various populations, without affecting the total amount (see Figure 3).
The way n* depends on parameters β and γ is illustrated in Figure 2. Notice that n* is a decreasing function of β, while in general the curves n* vs. β are concave with a single maximum. Further observe that n* attains positive values in the interval [0, 3] when β = 0—see Equation (21), but this interval shrinks as the value of β increases.
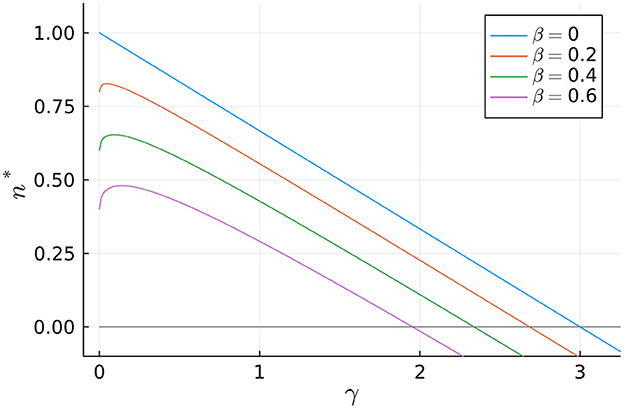
Figure 2. Dependence of the normalized total mitochondrial volume density, n*, on the normalized fusion rate, γ, for different values of the normalized aging rate, β.
To simplify the parametric analysis of the model, we make the assumption that, given the value of β, parameter γ takes on the value that maximizes the total mitochondrial volume density, n*. By performing the necessary mathematical calculations, we can straightforwardly show that this assumption imposes the following restriction:
With this, n* is only a function of β, while the value of variables h*, m*, s* and f* depends on parameters β, δ and ϵ.
Figure 3 depicts the impact of fission and mitophagy on the distribution of mitochondrial volume density across compartments. Specifically, the steady-state values of variables h, m, f, and s (normalized to the total volume density, n) are plotted against parameters ϵ and δ for a specific value of β. Observe that when the fission rate (δ) or the mitophagy rate (ϵ) increases, there is an increase in the volume densities of healthy (h*) and moderately damaged (m*) mitochondria. Further note that this effect is more significant for moderately damaged mitochondria. On the other hand, it can be observed that the volume density of fused mitochondria remains almost constant regardless of the mitophagy rate, but it rapidly decreases with increasing fission rate. In contrast, the volume density of severely damaged mitochondria remains almost constant regardless of δ, but rapidly decreases with increasing ϵ.
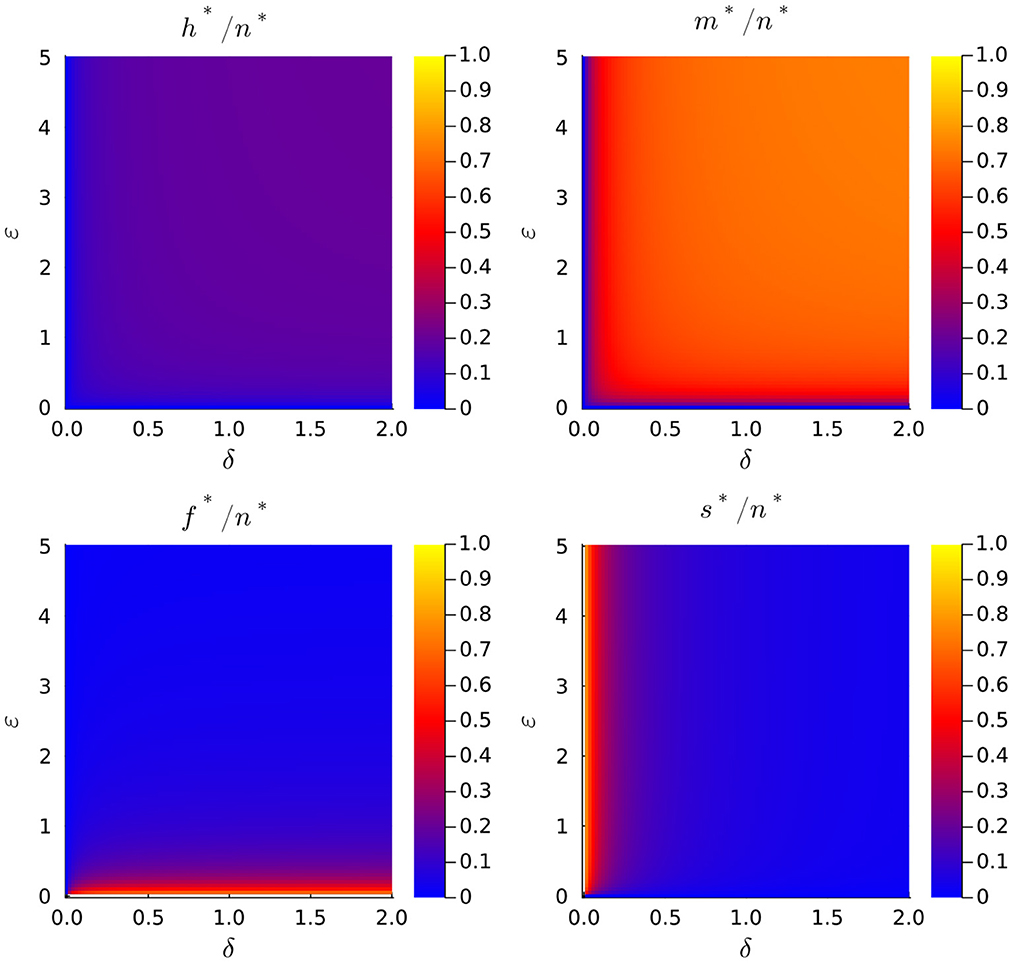
Figure 3. Dependence of the normalized total mitochondrial volume density, n*, on the normalized fission rate, γ, for different values of the normalized aging rate, β.
Of particular interest are the values of h* and m* because these variables represent the volume density of functional mitochondria. In this study, we do not regard fused mitochondria as functional, as they represent a temporary state involving the exchange of mitochondrial components. Since changes of parameters δ and ϵ do not affect the value of n* but only the way the total mitochondrial volume density distributes among different subpopulations, is it important to investigate the effect these parameters have upon the fractions of healthy, h*/n*, and moderately damaged mitochondria, h*/n*. It is possible to prove that both quantities are monotonic decreasing functions of β and monotonic increasing functions of δ and ϵ, but for simplicity we refer the reader to Figure 4 to discuss this behavior. There, we plot the fraction of functional mitochondrial volume density, which defined as Φ = (h* + m*)/n*, as a function of parameters δ and ϵ, for a couple of different values of parameter β.
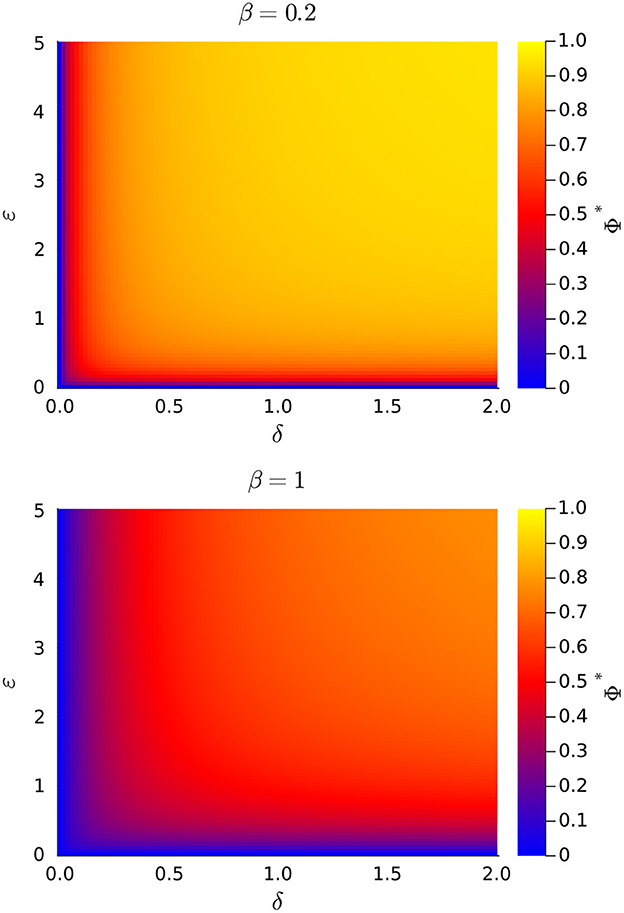
Figure 4. Plots of the fraction of stationary functional mitochondrial volume density (Φ* = f* + m*) vs. the rates of mitochondrial fission (δ) and mitophagy (ϵ), for a couple of values of the damaging rate (β). All rate are normalized to the intrinsic mitogenesis rate.
Note in Figure 4 that Φ = 0 when either δ = 0 or ϵ = 0, in agreement with previous results. This confirms that, according to our model, mitochondrial fission and mitophagia are essential for maintaining a functional mitochondrial volume density. Further observe that Φ decreases as β increases, for all values of δ = 0 and ϵ = 0. That is, increasing the rate of mitochondrial damaging not only causes a decrement of the total mitochondrial volume density, but also diminishes the fraction of functional mitochondria. Finally, the value of Φ augments when δ and/or ϵ, are increased. Indeed, by increasing the rates of mitochondrial fission and/or mitophagy, it is possible to at least partially revert the decrease of ϕ originated by an increment of the mitochondrial damaging rate.
4. Further discussion and concluding remarks
Mathematical models can be classified as either explanatory or predictive. Explanatory modeling seeks to uncover causal relationships between input variables and outcomes, whereas the goal of predictive modeling is to generate precise quantitative predictions. Explanatory models generally require a small number of parameters and are simple enough to be studied in depth, preferably analytically. Predictive models, on the other hand, are typically much more detailed, have a large number of parameters to estimate, and can generally only be handled numerically. The model presented here, in our opinion, is a good example of the first category of models. It takes into account the fundamental regulatory elements of mitochondrial dynamics, which is consistent with the idea that it plays an important role in the quality control of mitochondrial function, yet it is relatively simple. In fact, the model stationary states and their dependence on model parameters could be studied analytically thanks to its simplicity. The following are the main findings of such an analysis:
• Fission and mitophagy are strictly necessary for a healthy mitochondrial population. Functional mitochondria are eventually eliminated if these processes are not present.
• Increasing the rate of mitochondrial damage has a negative impact on the population of functional mitochondria. However, the damage can be mitigated to some extent by increasing fission and/or mitophagy rates.
We want to highlight the second of the previously listed results because it may help us understand how, by modifying mitochondrial dynamics parameters, physical exercise may help to (at least partially) counteract the damage caused in mitochondria by affections such as obesity.
The existing literature generally agrees that mitochondrial dynamics processes play a crucial role in maintaining mitochondrial health. However, there are conflicting findings on the specific contributions of different processes, particularly in the case of fusion and fission. With this in mind, our findings support previous research indicating that exercise promotes mitophagy, which in turn has a beneficial effect on mitochondrial health, as described in some recent reviews [11, 12]. Previous studies have shown that the knockdown of gene Drp1, which promotes mitochondrial fission, leads to increased markers of muscle regeneration, denervation, fibrosis, and oxidative stress, as well as decreased mitochondrial respiration. These findings are in line with our own, which suggest that a reduced mitochondrial fission can have a negative impact on mitochondrial health [21].
Data availability statement
The original contributions presented in the study are included in the article/supplementary material, further inquiries can be directed to the corresponding author.
Author contributions
ÁZ-R contributed to the model development, performed the mathematical analysis, participated in the discussion, and contributed to the manuscript writing. MS conceived the project, contributed to the model development, supervised the mathematical analysis, participated in the discussion, and contributed to the manuscript writing. All authors contributed to the article and approved the submitted version.
Funding
This work was partially financed by Conacyt-Mexico under Grant: CDF-19-568462.
Acknowledgments
MS was grateful to the Applied Physics Department at University of Salamanca for generously hosting as a visiting professor while this project was performed.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher's note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
1. Kowald A, Klipp E. Mathematical models of mitochondrial aging and dynamics. In: Progress in Molecular Biology and Translational Science. New York, NY: Elsevier (2014). p. 63–92.
2. Leduc-Gaudet JP, Hussain SNA, Barreiro E, Gouspillou G. Mitochondrial dynamics and mitophagy in skeletal muscle health and aging. Int J Mol Sci. (2021) 22:8179. doi: 10.3390/ijms22158179
3. Twig G, Elorza A, Molina AJA, Mohamed H, Wikstrom JD, Walzer G, et al. Fission and selective fusion govern mitochondrial segregation and elimination by autophagy. EMBO J. (2008) 27:433–46. doi: 10.1038/sj.emboj.7601963
4. Patel PK, Shirihai O, Huang KC. Optimal dynamics for quality control in spatially distributed mitochondrial networks. PLoS Comput Biol. (2013) 9:e1003108. doi: 10.1371/journal.pcbi.1003108
5. Liesa M, Shirihai OS. Mitochondrial dynamics in the regulation of nutrient utilization and energy expenditure. Cell Metab. (2013) 17:491–506. doi: 10.1016/j.cmet.2013.03.002
6. de Mello AH, Costa AB, Engel JDG, Rezin GT. Mitochondrial dysfunction in obesity. Life Sci. (2018) 192:26–32. doi: 10.1016/j.lfs.2017.11.019
7. Lahera V, de las Heras N, López-Farré A, Manucha W, Ferder L. Role of mitochondrial dysfunction in hypertension and obesity. Curr Hypertension Rep. (2017) 19(2). doi: 10.1007/s11906-017-0710-9
8. Chen PB, Yang JS, Park Y. Adaptations of skeletal muscle mitochondria to obesity, exercise, and polyunsaturated fatty acids. Lipids. (2018) 53:271–8. doi: 10.1002/lipd.12037
9. Arribat Y, Broskey NT, Greggio C, Boutant M, Alonso SC, Kulkarni SS, et al. Distinct patterns of skeletal muscle mitochondria fusion, fission and mitophagy upon duration of exercise training. Acta Physiol. (2018) 225:e13179. doi: 10.1111/apha.13179
10. Trewin A, Berry B, Wojtovich A. Exercise and mitochondrial dynamics: keeping in shape with ROS and AMPK. Antioxidants. (2018) 7:7. doi: 10.3390/antiox7010007
11. Wu NN, Tian H, Chen P, Wang D, Ren J, Zhang Y. Physical exercise and selective autophagy: benefit and risk on cardiovascular health. Cells. (2019) 8:1436. doi: 10.3390/cells8111436
12. Tanaka T, Nishimura A, Nishiyama K, Goto T, Numaga-Tomita T, Nishida M. Mitochondrial dynamics in exercise physiology. Pflügers Archiv - Eur J Physiol. (2019) 472:137–53. doi: 10.1007/s00424-019-02258-3
13. Wahwah N, Kras KA, Roust LR, Katsanos CS. Subpopulation-specific differences in skeletal muscle mitochondria in humans with obesity: insights from studies employing acute nutritional and exercise stimuli. Am J Physiol Endocrinol Metabol. (2020) 318:E538–53. doi: 10.1152/ajpendo.00463.2019
14. Tam ZY, Gruber J, Halliwell B, Gunawan R. Mathematical modeling of the role of mitochondrial fusion and fission in mitochondrial DNA maintenance. PLoS ONE. (2013) 8:e76230. doi: 10.1371/journal.pone.0076230
15. Kornick K, Bogner B, Sutter L, Das M. Population dynamics of mitochondria in cells: a minimal mathematical model. Front Phys. (2019) 7:146. doi: 10.3389/fphy.2019.00146
16. Meyer JN, Leuthner TC, Luz AL. Mitochondrial fusion, fission, and mitochondrial toxicity. Toxicology. (2017) 391:42–53. doi: 10.1016/j.tox.2017.07.019
17. Nicholls DG, Ward MW. Mitochondrial membrane potential and neuronal glutamate excitotoxicity: mortality and millivolts. Trends Neurosci. (2000) 23:166–74. doi: 10.1016/S0166-2236(99)01534-9
18. Kim I, Rodriguez-Enriquez S, Lemasters JJ. Selective degradation of mitochondria by mitophagy. Arch Biochem Biophys. (2007) 462:245–53. doi: 10.1016/j.abb.2007.03.034
19. Rabelo ACS. Baccharis trimera phytochemicals and the mitochondria. In: Mitochondrial Physiology and Vegetal Molecules. New York, NY: Elsevier (2021). p. 377–93.
Keywords: mitochondrial dynamics, mitochondrial maintenance, nonlinear dynamics, obesity, exercise
Citation: Zamora-Ramírez JÁ and Santillán M (2023) Effects of fission and mitophagy on mitochondrial functionality under accelerated damage conditions: A mathematical modeling approach. Front. Appl. Math. Stat. 9:1082928. doi: 10.3389/fams.2023.1082928
Received: 07 November 2022; Accepted: 15 March 2023;
Published: 29 March 2023.
Edited by:
Yasser Aboelkassem, University of Michigan-Flint, United StatesReviewed by:
Vicente Lahera, Complutense University of Madrid, SpainStephen T. Kinsey, University of North Carolina Wilmington, United States
Copyright © 2023 Zamora-Ramírez and Santillán. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Moisés Santillán, bXNhbnRpbGxhbkBjaW52ZXN0YXYubXg=