 Fériel Bouzid
Fériel Bouzid Fabienne Brégeon2,3
Fabienne Brégeon2,3 Isabelle Poncin
Isabelle Poncin Stéphane Canaan
Stéphane Canaan- 1Aix-Marseille Université, CNRS, EIPL IMM FR3479, Marseille, France
- 2Aix Marseille Université, URMITE, UMR CNRS 7278, IRD 198, INSERM 1095, IHU Méditerranée Infection, Marseille, France
- 3Service des Explorations Fonctionnelles Respiratoires, Centre Hospitalo-Universitaire Nord, Pôle Cardio-Vasculaire et Thoracique, Assistance Publique Hôpitaux de Marseille, Marseille, France
Adipose tissues were shown to host Mycobacterium tuberculosis which is persisting inside mature adipocytes. It remains unknown whether this holds true for Mycobacterium canettii, a rare representative of the M. tuberculosis complex responsible for lymphatic and pulmonary tuberculosis. Here, we infected primary murine white and brown pre-adipocytes and murine 3T3-L1 pre-adipocytes and mature adipocytes with M. canettii and M. tuberculosis as a positive control. Both mycobacteria were able to infect 18–22% of challenged primary murine pre-adipocytes; and to replicate within these cells during a 7-day experiment with the intracellular inoculums being significantly higher in brown than in white pre-adipocytes for M. canettii (p = 0.02) and M. tuberculosis (p = 0.03). Further in-vitro infection of 3T3-L1 mature adipocytes yielded 9% of infected cells by M. canettii and 17% of infected cells by M. tuberculosis (p = 0.001). Interestingly, M. canettii replicated and accumulated intra-cytosolic lipid inclusions within mature adipocytes over a 12-day experiment; while M. tuberculosis stopped replicating at day 3 post-infection. These results indicate that brown pre-adipocytes could be one of the potential targets for M. tuberculosis complex mycobacteria; and illustrate differential outcome of M. tuberculosis complex mycobacteria into adipose tissues. While white adipose tissue is an unlikely sanctuary for M. canettii, it is still an open question whether M. canettii and M. tuberculosis could persist in brown adipose tissues.
Introduction
In mammals, brown adipose tissue (BAT) and white adipose tissue (WAT) are mainly composed of mature adipocytes, pre-adipocytes and immune cells (Ouchi et al., 2011). WAT and BAT differ in cell morphology, tissue distribution and physiological functions (Gomez-Hernandez et al., 2016): WAT constitutes the main energy reserve of the organism while BAT ensures thermogenesis in hibernating animals and newborns (Cannon and Nedergaard, 2004; Gomez-Hernandez et al., 2016) and remains metabolically active in adults (Nedergaard et al., 2007).
Adipose tissues have been previously supposed to act as long-term sanctuaries sheltering Mycobacterium tuberculosis, the major agent of human tuberculosis worldwide (Neyrolles et al., 2006; Kim et al., 2011; Agarwal et al., 2014, 2016; Rastogi et al., 2016). In vitro experiments were conducted using murine 3T3-L1 mature adipocytes (Neyrolles et al., 2006; Kim et al., 2011; Rastogi et al., 2016), and 3T3-L1 pre-adipocytes (Neyrolles et al., 2006), but also with murine primary pre-adipocytes and mature adipocytes obtained from WAT (Agarwal et al., 2014). In immune-competent mice, intravenously and intra-nasally inoculated M. tuberculosis disseminates in WAT of visceral, subcutaneous, peri-renal and mesenteric adipose depots (Agarwal et al., 2014, 2016). In these models, the burden of M. tuberculosis in WAT depots stagnated or decreased with time (Agarwal et al., 2014). Indeed, compelling data show that M. tuberculosis adopts in mature adipocytes and WAT a non-replicating, dormant state characterized by an arrest of multiplication, accumulation of intra-cytosolic lipid inclusions (ILIs), resistance to anti-mycobacterial drugs (Neyrolles et al., 2006; Agarwal et al., 2014) and up-regulation of dosR and icl genes (Rastogi et al., 2016). The infection of mature adipocytes was therefore proposed as a suitable model to study the accumulation of neutral lipids into ILIs within mycobacteria (Santucci et al., 2016).
Among the M. tuberculosis complex (MTBC), Mycobacterium canettii is a peculiar member specifically diagnosed in dozens of tuberculosis patients with reported contacts to the Horn of Africa (Aboubaker Osman et al., 2015). M. canettii differs from the other members of the MTBC by processing a larger 4.48 ± 0.05 Mb mosaic genome and producing cordless and smooth-looking mycobacteria (Gutierrez et al., 2005; Koeck et al., 2011; Supply et al., 2013; Boritsch et al., 2016a) with an ability of intra-species horizontal gene transfer (Boritsch et al., 2016b). M. canettii infection mainly presents as lymph node and pulmonary tuberculosis (Koeck et al., 2011; Aboubaker Osman et al., 2015). However, M. canettii pulmonary tuberculosis is unique in being seemingly non-contagious (Koeck et al., 2011).
In order to further examine the in-vitro interactions of M. canettii with natural adipose tissues, we developed in-vitro experimental models using primary murine white and brown pre-adipocytes; and further investigated the interactions between M. canettii and murine 3T3-L1 pre-adipocytes and mature adipocytes using M. tuberculosis H37Rv as a positive control.
Materials and Methods
Mycobacteria
M. tuberculosis H37Rv and M. canettii CIPT 140010059 were used in this study. Mycobacteria were cultivated in Middelbrook 7H10 (Becton Dickinson, Le Pont de Claix, France) supplemented with 10% oleic acid-albumin-dextrose-catalase (OADC) (Becton Dickinson). Fluorescent M. canettii CIPT 140010059 mCherry (CSURP3621) was constructed by transforming the pMV261 mCherry vector (gift from L. Kremer, CPBS, Montpellier, France) into the M. canettii strain (Alibaud et al., 2011). Fluorescent strains were cultured in Middlebrook 7H10 broth supplemented with 10% OADC and 50 μg/mL kanamycin (Sigma). Prior to infection, mycobacteria were resuspended in phosphate buffered saline (PBS), shaken on a vortex mixer for 10 min with glass ball to disperse clumps and centrifuged for 1 min at 300 g to remove residual clumps. The supernatant was then dispersed by expelling the suspension 10 times through a sterile 25-gauge needle attached to a 1-mL syringe. Calibration was then performed according to Mcfarland standard confirmed by counting mycobacteria after Ziehl-Neelsen staining (RAL diagnostics, Martillac, France). All experiments using these mycobacteria were performed in a biosafety level 3 laboratory of the Faculté de Médecine, Aix-Marseille Université, France.
Primary Murine Pre-adipocyte Culture
The experiments conducted on mice was approved by the Institutional Animal Care and Use Committee of Aix-Marseille University “C2EA-14,” France and registered by the “Ministère de l'Enseignement Supérieur et de la Recherche” under reference n° 2015092415474605. Mice were handled according to the rules of Décret N° 2013–118, Février 7, 2013, France. Inguinal WAT and inter-scapular BAT were collected from ten 6-week-old Balb/cByj mice. Pre-adipocytes cells were isolated from tissues as described elsewhere (Aune et al., 2013) and then cultured in complete culture medium Dulbecco's Modified Eagle Medium DMEM/F12 (Invitrogen, France) supplemented with 10% heat-inactivated fetal bovine serum (FBS) and were plated at a concentration of 5 × 104 cells/ml/well in 12-well plates and incubated at 37°C under a 5% CO2 atmosphere.
3T3-L1 Cell Lines
Murine embryonic fibroblasts 3T3-L1 (ATCC, lot 62158491) were cultured in DMEM (Invitrogen, France) supplemented with 10% heat-inactivated FBS at 37°C under a 5% CO2 atmosphere. Prior to confluence, cells were plated at 5 × 104 cells/well in a 12-well sterile plates and incubated as described above. Six days after plating, pre-adipocytes reach confluency and stop dividing. Adipocyte differentiation was initiated upon confluency by adding 1 μg/mL insulin (Sigma), 0.5 mM isobuthyl-methylxanthine (Sigma), and 1 μM dexamethasone (sigma) into the culture medium. After 48 h of induction, the medium was replaced by DMEM-10% FBS containing 1 μg/mL of insulin and maintained each 2 days in order to achieve a full differentiation as previously described (Neyrolles et al., 2006). Adipocyte differentiation was checked by BODIPY 493/503 staining (Sigma) of lipid droplets over the 2-week experiments.
Infection
Pre-adipocytes and mature adipocytes were infected with a multiplicity of infection (MOI) of 1 per cell at day 6 after plating and at day 10 post-induction respectively. Accordingly, 1 × 105 cells per well were infected with 1 × 105 mycobacteria. After 4-h incubation, cells were washed 3 times with serum-free DMEM to remove all extracellular bacteria. Only mature adipocytes were then incubated with 200 μg/mL amikacin for 2 h and then washed thrice. The last wash was plated to validate the correct elimination of extracellular mycobacteria. The inoculated cultures were incubated with fresh complete medium up to 7 days for primary cultures and 12 days for 3T3-L1 cell lines at 37°C under a 5% CO2 atmosphere. At various time points, infected cells were lysed in PBS containing 0.1% Triton X-100 and the lysate was plated at 10-fold dilutions from 10−1 to 10−4 onto Middlebrook 7H10 agar plates incubated at 37°C under a 5% CO2 atmosphere. The number of colonies enumerated after 20-day of incubation was used to estimate the number of colony-forming units (CFUs) per 105 cells. All experiments were conducted in triplicate: three independent plates the same day for primary cell infection experiments and three replicates at different days with different bacterial cultures for 3T3-L1 cells infection experiments.
Intracellular Localization of Mycobacteria
Ziehl-Neelsen staining (RAL diagnostics, Martillac, France) was performed on infected cells (MOI 1:1 and 5:1) at day 3 p.i. Observations were acquired using the slide scanner Axio Scan.Z1 (Zeiss) (Magnification 20X) with a color Hitachi tri-CCD (1,800 × 1,200 pixels) camera. The number of intracellular mycobacteria per one cell was recorded on image acquisition by ImageJ software.
M. canettii mCherry was used to localize intracellular bacteria using confocal microscopy. Infected adipocytes (MOI 10) were fixed using 4% formaldehyde at day 3 p.i. Lipid droplets were stained with BODIPY 493/503 (Sigma). The images were acquired using a confocal microscope (Zeiss, Spinning disk with a Yokogawa head and an emCCD camera 512 × 512 pixels) at magnification 63X/1.4 oil objective.
Processing for Electron Microscopy
Cells were fixed at room temperature with 2.5% glutaraldehyde in Na-cacodylate buffer 0.1 M (pH 7.2) containing 0.1 M sucrose, 5 mM CaCl2, and MgCl2 5 mM, washed with complete cacodylate buffer and postfixed for 1 h at room temperature with 1% osmium tetroxide in the same buffer without sucrose (de Chastellier, 2008). They were washed with buffer, scratched gently, concentrated in agar up to 2% with cacodylate buffer and processed for 1 h at room temperature with 1% uranyl acetate in maleate buffer. The samples were dehydrated in a graded series of ethanol solutions and gradually incorporated in Spurr resin. Thin sections (80 nm thick) were stained with 1% uranyl acetate in distilled water and then with lead citrate before being observed by electron microscopy.
Nile Red Staining
Adipocytes were infected with M. canettii at MOI 5:1 on 12-well plate. At day 7 p.i. all wells were lysed in 500 μL water containing 0.1% Triton X-100. The pooled lysate was sonicated and centrifuged at 9,000 g for 25 min. The M. canettii cells were washed thrice with 0.1% Triton X-100 and the pellet was resuspended in 500 μL of PBS containing 0.05% Tween 80. Nile red staining was performed as previously described (Christensen et al., 1999). The stained smears were subjected to high resolution confocal (Zeiss AiryScan head, magnification 63X, Numerical aperture = 1.4), with higher resolution (1.6X) than conventional confocal.
Lactate Dehydrogenase (LDH) Assay
LDH, a soluble cytoplasmic enzyme released into extracellular space when the cell membrane is damaged, was used as an estimator of cell lysis (Korzeniewski and Callewaert, 1983). Culture supernatants of uninfected or infected mature adipocytes with M. canettii or M. tuberculosis (MOI 1) were collected at day 3, 7, and 12 post-inoculation. The release of LDH from negative controls and infected cells was measured spectrophotometrically at 340 nm on a Cobas 8000 (Roche, Meylan, France).
Statistical Analyses
Statistics were performed using the SigmaPlot13 software. The distribution of the variables was assessed statistically with the Shapiro-Wilk test. The equality of the variance was checked using the Brown-Forsythe test. All data were normally distributed and were expressed using means ± standard deviation. The statistical significance was performed using the Student T-test. The Chi-square test was used to compare rates and proportions with Yates correction. A p <0.05 was regarded as significant.
Results
M. canettii and M. tuberculosis within Primary Murine Pre-adipocytes
Pre-adipocytes purified from murine inguinal WAT and inter-scapular BAT were ex vivo inoculated with M. canettii or M. tuberculosis for 4 h p.i. For all experiments, extracellular mycobacteria were removed by extensive washing with serum-free medium, 4 h p.i. At day 3 p.i. Ziehl-Neelsen staining was performed to assess the ability of mycobacteria to infect primary murine pre-adipocytes. While negative control cells remained free of detectable mycobacteria, microscopic observations clearly showed intracellular M. canettii and M. tuberculosis in white and brown pre-adipocytes (Figure 1).
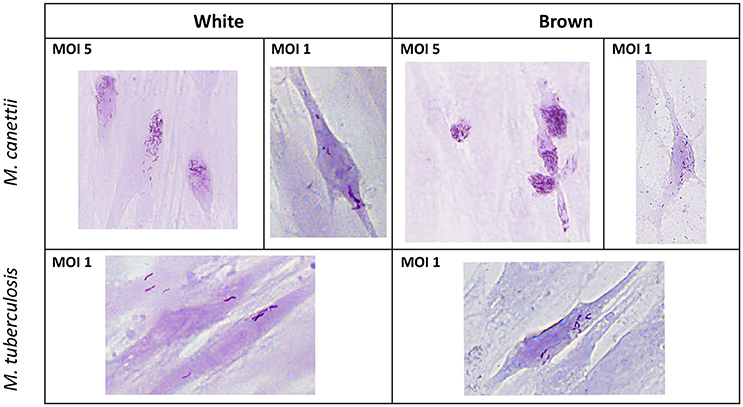
Figure 1. Ziehl-Neelsen staining performed on infected primary murine pre-adipocytes. White and brown pre-adipocytes obtained from ex vivo culture from mouse adipose depots were infected for 4 h with M. canettii CIPT 140010059 or M. tuberculosis H37Rv and stained at day 3 post-inoculation. Microscopic observations at 20x magnification showed infection of white and brown pre-adipocytes by M. canettii upper panel and by M. tuberculosis H37Rv lower panel.
The ratio of infected cells relative to the total number of observed cells as well as the average number of intracellular bacilli per one cell were measured on a random selection of 10 microscope fields on stained infected cells at day 3 p.i. (see Supplementary Data S1). White pre-adipocytes infected at MOI 1:1 yielded 19% and 18% of infected cells with 9 ± 6 M. canettii bacilli per cell and 7 ± 5 M. tuberculosis organisms per cell, respectively. Likewise, 22 and 19% of brown pre-adipocytes were infected with 16 ± 7 M. canettii organisms/cell and 11 ± 4 M. tuberculosis organisms/cell, respectively. The ratio of infection did not significantly differ between brown and white pre-adipocytes (p = 0.7 for M. canettii and p = 0.8 for M. tuberculosis, χ2 test) but the number of bacilli in brown pre-adipocytes was significantly higher than in white pre-adipocytes (p = 0.02 for M. canettii and p = 0.03 for M. tuberculosis).
In order to confirm these observations and to study the viability of bacilli into the infected cells, the number of intracellular mycobacteria was quantified by scoring CFUs/105 cells after plating infected cell lysates with M. canettii or M. tuberculosis at MOI 1:1 at different time intervals (Figure 2) (see Supplementary Data S1). At day 0, white pre-adipocytes had internalized 15% (1.5 × 104 ± 9 × 103 CFUs) and 10% (9 × 103 ± 4 × 103 CFUs) of the initial 1 × 105 inoculum of M. canettii and M. tuberculosis, respectively. As for brown pre-adipocytes, 45% (4.5 × 104 ± 2.3 × 103 CFUs) of the M. canettii inoculum and 25% (2.47 × 104 ± 4 × 103 CFUs) of the M. tuberculosis inoculum was phagocytized. By scoring CFUs, intracellular M. canettii and M. tuberculosis organisms were more abundant in primary brown pre-adipocytes than in white pre-adipocytes (p = 0.01 for M. canettii and p = 0.008 for M. tuberculosis) which is consistent with our microscopic observations.
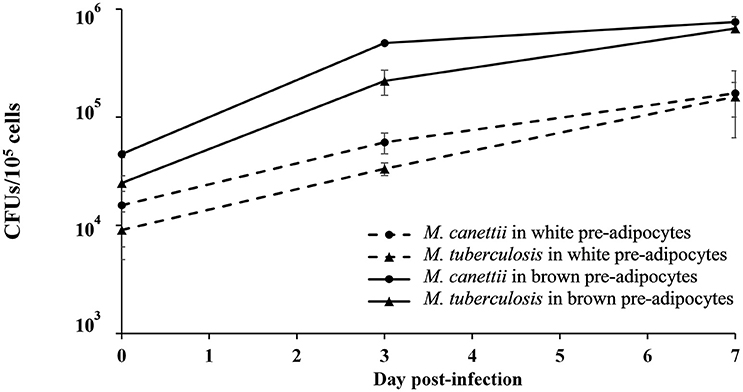
Figure 2. Number of colony-forming units (CFUs/105 cells) of M. canettii and M. tuberculosis in murine white and brown pre-adipocytes over the 7-day experiments. Primary murine pre-adipocytes were infected at MOI 1:1 with M. canettii CIPT 140010059 or M. tuberculosis H37Rv for 4 h. At day 0, 3, and 7 post-inoculation, CFUs were scored by plating serial dilutions of cell lysates after washing three times to remove of extracellular mycobacteria. Results shown are the mean of three independent experiments and bars indicate + standard deviation. A similar intracellular kinetic was observed for the two mycobacteria in both type cells.
A similar intracellular behavior was observed for M. canettii and M. tuberculosis during the 7-day experiment in primary white and brown pre-adipocytes: both mycobacteria survived and multiplied with approximately one log gain.
Interactions between M. canettii and 3T3-L1 Pre-adipocytes and Mature Adipocytes
M. canettii Is Internalized into 3T3-L1 Pre-adipocytes and Mature Adipocytes
To explore M. canettii interactions with 3T3-L1 pre-adipocytes and mature adipocytes compare to M. tuberculosis (Neyrolles et al., 2006; Kim et al., 2011), the quantification of the ratio of infected cells/total cells and the number of intracellular bacilli/cell were performed on a random selection of 10 microscope fields after Ziehl-Neelsen staining of infected cells at day 3 p.i. (see Supplementary Data S1). For pre-adipocytes, 20% of cells were infected with 7 ± 5 M. canettii organisms/cell while 40% of cells were infected with 12 ± 6 M. tuberculosis organisms/cell which is significantly higher compared to M. canettii infection (p <0.001, χ2 test). Under the same conditions, only 9% and 17% of adipocytes were infected with M. canettii and M. tuberculosis, respectively. The ratio of infected cells was significantly higher for pre-adipocytes compared to mature adipocytes (p = 0.002 for M. canettii and p <0.001 for M. tuberculosis, χ2 test).
To confirm the intracellular location of M. canettii within 3T3-L1 mature adipocytes, cells were inoculated with M. canettii CIPT 140010059 mCherry (CSURP3621) at MOI = 10. Three days p.i. infected cells were treated with BODIPY 493/503 stain and fixed. Confocal microscopy observations showed the presence of 15 ± 6 intracellular M. canettii inside adipocytes, as compared to negative controls which remained free of detectable mCherry-fluorescence (Figure 3A). We noticed that approximately 30% of intracellular M. canettii organisms were localized in close contacts with cytoplasmic lipid bodies (LBs) in 100% of the observed adipocytes (Figure 3A white squares).
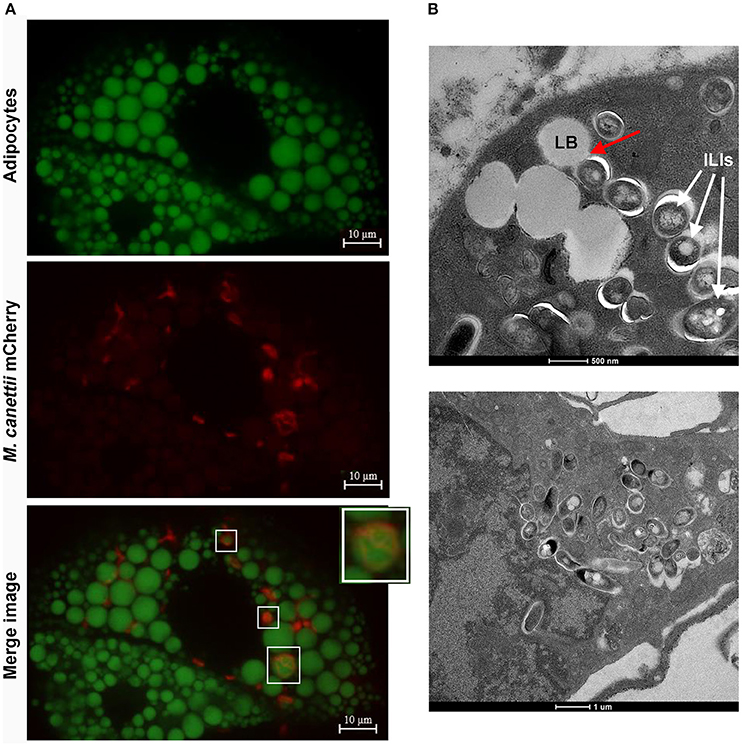
Figure 3. M. canettii infection of 3T3-L1 mature adipocytes. (A) Confocal images of 3T3-L1 mature adipocytes infected with M. canettii. Adipocytes were infected at MOI 10:1 with M. canettii mCherry for 4 h. At day 3 post-inoculation, extracellular mycobacteria were washed off, infected cells were fixed in 4% formaldehyde and the lipid droplets of adipocytes were stained using BODIPY 493/503. Confocal images showed intracellular location of M. canettii mCherry and 30% of intracellular bacilli were next to lipid bodies (White squares). (B) Electron microscopy observations of 3T3-L1 mature adipocytes infected with M. canettii. Adipocytes were infected at MOI 5:1 with M. canettii for 4 h. At day 3 post-inoculation, the supernatant was removed and infected cells were fixed for electron microscopy. M. canettii organisms were inside adipocytes in close contact with adipocytes LBs as indicated by the red arrow and accumulated ILIs (upper panel). Pre-adipocytes that failed to reach full differentiation were infected also with M. canettii (lower panel).
Further electron microscopy observation of adipocytes infected at a MOI 5:1 for 3 days demonstrated the presence of M. canettii within adipocytes in close contact with adipocyte LBs, corroborating observations performed by confocal microscopy (Figure 3B, upper panel). Intracellular M. canettii bacilli were loaded with small electron-dense vesicles resembling ILIs (Figure 3B, upper panel). We also found that some pre-adipocytes that failed to reach full differentiation were infected by M. canettii which had weakly accumulated ILIs (Figure 3B, lower panel).
Intracellular Outcome of Internalized M. canettii
Using the 3T3-L1 continuous cell line which is more robust and easier to manipulate than primary cells, it was possible to compare the successive steps of infection over 12 days in three independent experiments conducted at different days, comparing pre-adipocytes and mature adipocytes. After thorough dispersal of the mycobacterial aggregate, cells were infected with M. tuberculosis, used as positive control, or M. canettii at a MOI 1:1 for 4 h followed by extensive elimination of extracellular mycobacteria. All results were compared with non-infected cells manipulated in parallel, which remained negative in culture during the 12-day experiment. At various time points 0, 3, 7, and 12 days p.i. cells were lysed and viable bacterial content was measured by plating cell lysates onto Middelbrook agar 7H10 and scoring CFUs/105 cells (Figure 4) (see Supplementary Data S1).
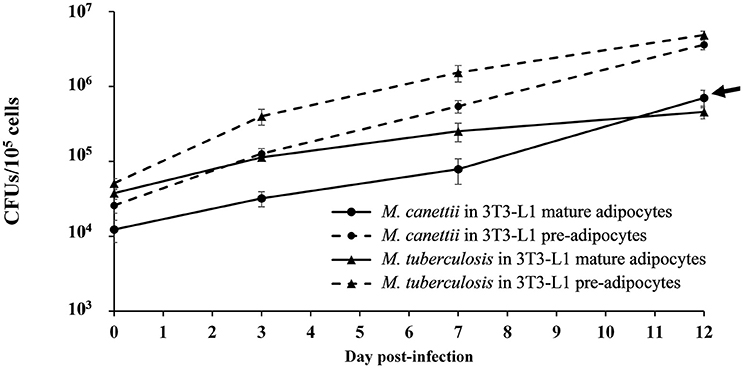
Figure 4. Intracellular kinetic of M. canettii CIPT 140010059 and M. tuberculosis H37Rv in 3T3-L1 pre-adipocytes and adipocytes during 12-day experiments. Cells were infected at MOI 1:1 with M. canettii or M. tuberculosis for 4 h. The number of bacterial colony forming units (CFU) was determined at the indicated time points (0, 3, 7, and 12 day p.i.). A similar intracellular kinetic was observed for M. canettii and M. tuberculosis inside pre-adipocytes. However, in mature adipocytes M. tuberculosis stopped its replication between day 3 and 12 day p.i. while M. canettii continued to replicate (arrow). Data shown are the mean ± standard deviation of three independent experiments.
As for internalization, at day 0, pre-adipocytes contained 25% of the initial M. canettii inoculum (2.5 × 104 ± 5 × 103 CFUs) and 50% of the initial M. tuberculosis inoculum (5 × 104 ± 8 × 103 CFUs). In mature adipocyte infection experiments, 12% (1.2 × 104 ± 4 × 103 CFUs) of the infecting M. canettii inoculum and 37% of the infecting M. tuberculosis inoculum (3.7 × 104 ± 4 × 103 CFUs) was internalized. Initial infection of mature adipocytes was significantly lower with M. canettii (p = 0.001) than with M. tuberculosis, as observed with infected pre-adipocytes (p = 0.006), but numbers of intracellular mycobacteria from day 0 were significantly higher in pre-adipocytes than in mature adipocytes (p = 0.01 for M. canettii and p = 0.03 for M. tuberculosis).
As for intracellular behavior, both mycobacteria continuously replicated into pre-adipocytes during the 12-day experiment and gained two logs between day 0 and day 12 (Figure 4). In mature adipocytes, M. tuberculosis and M. canettii showed two different kinetics: intracellular M. tuberculosis inoculum stabilized between day 3 and 12 p.i. (4 ± 3.66 CFUs gain), whereas M. canettii replicated with a 2 × 101 ± 2.5 × 101 CFUs gain between day 3 and 12 p.i. which is significantly higher than M. tuberculosis (p = 0.01) (Figure 4) but this load remains lower than in pre-adipocytes (Figure 4).
The arrest of M. tuberculosis's replication inside adipocytes has been correlated with a heavy accumulation of ILIs leading to a dormant state (Neyrolles et al., 2006; Kim et al., 2011; Agarwal et al., 2014). To investigate whether this holds true for M. canettii, intra-adipocyte bacilli were extracted at day 7 p.i. after Triton X-100 cell lysis and sonication. While Middlebrook 7H10 agar-grown M. canettii were free of detectable ILIs, Nile-red staining showed that intracellular M. canettii yielded ILIs (Figure 5). Intracellular M. canettii presented variable numbers of small ILIs corresponding to 60% ILI+1 and 40% ILI+2 profiles which were previously described (Caire-Brandli et al., 2014).
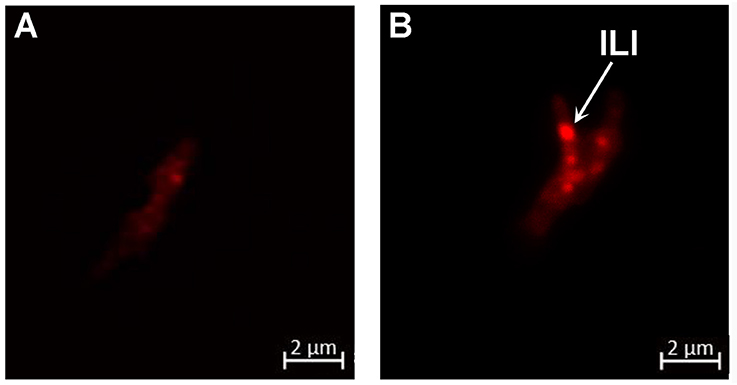
Figure 5. M. canettii moderately accumulates ILIs inside adipocytes. Adipocytes infected with M. canettii at MOI 5:1 were lysed at day 7 p.i. Intracellular mycobacteria were separated from cellular debris by lysis in water containing 0.1% Triton X-100 and sonication followed by several washes. Nile red staining was performed on (A) cultured M. canettii used as negative control and (B) intra-adipocyte M. canettii. Confocal observations at 63x magnification showed a free cytoplasm of cultured M. canettii and ILIs seen as bright red spots in the cytoplasm of adipocyte-dwelling M. canettii (white arrow). These results are representative of 100 observed mycobacteria in different microscopic fields.
Intracellular M. canettii Induce Mature Adipocyte Cytolysis
Microscopic examination indicated that M. canettii infection yielded no cytolysis on mature 3T3-L1 adipocytes up to day 7 p.i. but cellular debris were observed at day 12 p.i. while a smaller amount of cellular debris was observed in uninfected control cells. Microscopic observation of M. tuberculosis-infected cells clearly showed cellular debris at day 7 p.i. A cytolytic effect of mycobacteria was further investigated by measuring the lactate-dehydrogenase (LDH) in the supernatant of infected and non-infected control adipocytes (see Supplementary Data S1). We observed a progressive increase of the LDH concentration in the three culture supernatants. However, the increase of LDH concentration was significantly higher in M. canetti-infected adipocytes than in negative control adipocytes at day 3 p.i. (p = 0.002) and day 12 p.i. (p <0.001) (Figure 6). In M. tuberculosis-infected cells, high rates of LDH concentration measured throughout the 12-day experiment were significantly higher compared to uninfected cells and M. canettii infected cells at each time point (Figure 6). Positive and high correlation was noted between the rates of LDH release and the count of CFUs in the case of M. canettii and M. tuberculosis infection (r = 0.90 for M. canettii and r = 0.88 for M. tuberculosis).
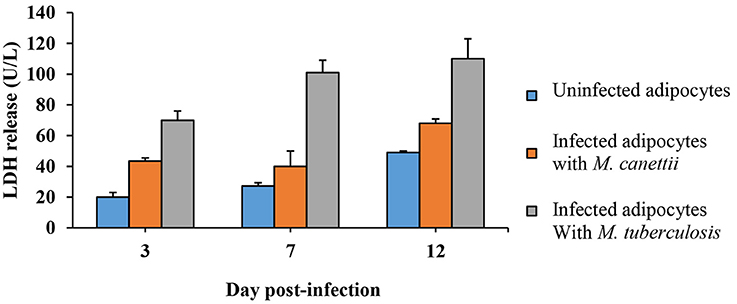
Figure 6. Quantitative analysis of LDH release. Culture supernatants of infected adipocytes were collected at day 3, 7, and 12 post-inoculation. LDH concentration was assessed in uninfected adipocytes (blue), M. canettii infected cells (orange), and M. tuberculosis H37Rv infected cells (gray). Data shown are the mean + standard deviation of three independent measurements.
Discussion
Data here reported indicate that M. canettii is able to infect, survive and replicate in primary murine white and brown pre-adipocytes which are constantly detected in adipose tissues (Cawthorn et al., 2012) and 3T3-L1 pre-adipocytes and mature adipocytes after in-vitro and ex-vivo experimental infection. Differentiation of 3T3-L1 cells was induced so as to obtain in vitro model of mature white adipocytes (Morrison and McGee, 2015). Data were authenticated by their reproducibility over a triplicate experiment in the presence of negative controls. All experiments with M. tuberculosis, here used as a positive control, were in agreement with those previously reported (Neyrolles et al., 2006; Kim et al., 2011; Agarwal et al., 2014). We extended for the first time these observations to brown pre-adipocytes.
Mature adipocytes were infected by M. canettii at a lower infectivity than M. tuberculosis (Neyrolles et al., 2006; Kim et al., 2011). M. tuberculosis binds to adipocytes through scavenger receptors (Neyrolles et al., 2006) but assessing whether this also holds true for M. canettii was beyond the scope of our study. Further, M. tuberculosis stopped replicating in mature adipocytes in agreement with its previously demonstrated dormant state inside mature adipocytes (Neyrolles et al., 2006; Kim et al., 2011; Agarwal et al., 2014; Rastogi et al., 2016). Unexpectedly, M. canettii continuously replicated and accumulated small ILIs. ILIs, previously reported in M. tuberculosis (McKinney et al., 2000; Daniel et al., 2004; Deb et al., 2006; Peyron et al., 2008; Russell et al., 2009), M. bovis BCG (Low et al., 2009, 2010), M. leprae (Mattos et al., 2010, 2011) and M. smegmatis (Garton et al., 2002; Dhouib et al., 2011) are here observed for the first time in M. canettii. Accordingly, the ILI+1/ILI+2 and not ILI+3 profiles of intracellular M. canettii agree with its replicating-state (Caire-Brandli et al., 2014). However, the experimental model used here did not enable us to observe potential long-term dormancy of M. canettii in mature adipocytes.
The primary and 3T3-L1 pre-adipocytes also phagocytized M. canettii and M. tuberculosis as previously shown with other particles (Cousin et al., 1999). More precisely, the amount of intracellular mycobacteria was significantly higher in brown than in white pre-adipocytes, corroborating previous observation of the earlier deposition of M. bovis in the BAT after intravenous administration in mice (Mauss and Levy, 1972). In vivo, the rich vascular network of BAT (Shimizu et al., 2014) could facilitate the tissue infection by M. canettii. Furthermore, the burden of intracellular mycobacteria was higher in pre-adipocytes than in mature adipocytes. Pre-adipocytes are fibroblast-like cells lacking cytoplasmic lipid droplets thus capable of rapidly phagocytosing (Hoffman and Dow, 2016); while mature adipocytes have lost this phagocytic activity (Cousin et al., 1999). Likewise, foamy macrophages loaded with LBs lost the ability to phagocytize mycobacteria compared to undifferentiated non-foamy macrophages (Peyron et al., 2008). These data indicate that pre-adipocytes are preferential targets for MTBC mycobacteria.
In conclusion, our results offer new pieces of information regarding the subtle interplay between MTBC mycobacteria and their hosts: infection of pre-adipocytes and adipocytes is a probable common feature of the MTBC at large, being shared by distantly related M. tuberculosis and M. canettii (Gutierrez et al., 2005), nevertheless, the intracellular outcome depends on the mycobacteria. While WAT is an unlikely sanctuary for M. canettii, the question remains open as for BAT. In the current quest for a yet unknown animal reservoir for M. canettii, our observations suggest that any mammal harboring BAT is a candidate reservoir, suggesting new field of investigations.
Author Contributions
SC and MD, conceived and supervised the experiments; Fé.B, performed the experiments; Fa.B, conducted the mice experiment; IP, conducted the electron microscopic observations; PW, was helpful for all confocal observations; SC and MD, and Fé.B analyzed the data and drafted the manuscript. All authors reviewed and approved the manuscript.
Funding
This study was financially supported by URMITE, IHU Méditerranée Infection, Marseille, France; CNRS and by the A*MIDEX project (n°ANR-11-IDEX-0001-02) funded by the “Investissements d'Avenir” French Government program, managed by the French National Research Agency (ANR).
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We thank the Biochemistry Laboratory–(Hospital of la Timone, Marseille, France) directed by Prof. Régis GUIEU for the LDH assay. The authors acknowledge Pablo Gluschankof who contributed to review the manuscript and Farid Soltani who contributed to analyze the slide scans of stained cells.
Supplementary Material
The Supplementary Material for this article can be found online at: http://journal.frontiersin.org/article/10.3389/fcimb.2017.00189/full#supplementary-material
References
Aboubaker Osman, D., Bouzid, F., Canaan, S., and Drancourt, M. (2015). Smooth tubercle bacilli: neglected opportunistic tropical pathogens. Front. Public Health 3:283. doi: 10.3389/fpubh.2015.00283
Agarwal, P., Khan, S. R., Verma, S. C., Beg, M., Singh, K., Mitra, K., et al. (2014). Mycobacterium tuberculosis persistence in various adipose depots of infected mice and the effect of anti-tubercular therapy. Microbes Infect. 16, 571–580. doi: 10.1016/j.micinf.2014.04.006
Agarwal, P., Pandey, P., Sarkar, J., and Krishnan, M. Y. (2016). Mycobacterium tuberculosis can gain access to adipose depots of mice infected via the intra-nasal route and to lungs of mice with an infected subcutaneous fat implant. Microb. Pathog. 93, 32–37. doi: 10.1016/j.micpath.2016.01.004
Alibaud, L., Rombouts, Y., Trivelli, X., Burguiere, A., Cirillo, S. L., Cirillo, J. D., et al. (2011). A Mycobacterium marinum TesA mutant defective for major cell wall-associated lipids is highly attenuated in Dictyostelium discoideum and zebrafish embryos. Mol. Microbiol. 80, 919–934. doi: 10.1111/j.1365-2958.2011.07618.x
Aune, U. L., Ruiz, L., and Kajimura, S. (2013). Isolation and differentiation of stromal vascular cells to beige/brite cells. J. Vis. Exp. 73:50191. doi: 10.3791/50191
Boritsch, E. C., Frigui, W., Cascioferro, A., Malaga, W., Etienne, G., Laval, F., et al. (2016a). pks5-recombination-mediated surface remodelling in Mycobacterium tuberculosis emergence. Nat. Microbiol. 1:15019. doi: 10.1038/nmicrobiol.2015.19
Boritsch, E. C., Khanna, V., Pawlik, A., Honore, N., Navas, V. H., Ma, L., et al. (2016b). Key experimental evidence of chromosomal DNA transfer among selected tuberculosis-causing mycobacteria. Proc. Natl. Acad. Sci. U.S.A. 113, 9876–9881. doi: 10.1073/pnas.1604921113
Caire-Brandli, I., Papadopoulos, A., Malaga, W., Marais, D., Canaan, S., Thilo, L., et al. (2014). Reversible lipid accumulation and associated division arrest of Mycobacterium avium in lipoprotein-induced foamy macrophages may resemble key events during latency and reactivation of tuberculosis. Infect. Immun. 82, 476–490. doi: 10.1128/IAI.01196-13
Cannon, B., and Nedergaard, J. (2004). Brown adipose tissue: function and physiological significance. Physiol. Rev. 84, 277–359. doi: 10.1152/physrev.00015.2003
Cawthorn, W. P., Scheller, E. L., and MacDougald, O. A. (2012). Adipose tissue stem cells meet preadipocyte commitment: going back to the future. J. Lipid Res. 53, 227–246. doi: 10.1194/jlr.R021089
Christensen, H., Garton, N., Horobin, R., Minnikin, D., and Barer, M. (1999). Lipid domains of mycobacteria studied with fluorescent molecular probes. Mol. Microbiol. 31, 1561–1572. doi: 10.1046/j.1365-2958.1999.01304.x
Cousin, B., Munoz, O., Andre, M., Fontanilles, A. M., Dani, C., Cousin, J. L., et al. (1999). A role for preadipocytes as macrophage-like cells. FASEB J. 13, 305–312.
Daniel, J., Deb, C., Dubey, V. S., Sirakova, T. D., Abomoelak, B., Morbidoni, H. R., et al. (2004). Induction of a novel class of diacylglycerol acyltransferases and triacylglycerol accumulation in Mycobacterium tuberculosis as it goes into a dormancy-like state in culture. J. Bacteriol. 186, 5017–5030. doi: 10.1128/JB.186.15.5017-5030.2004
Deb, C., Daniel, J., Sirakova, T. D., Abomoelak, B., Dubey, V. S., and Kolattukudy, P. E. (2006). A novel lipase belonging to the hormone-sensitive lipase family induced under starvation to utilize stored triacylglycerol in Mycobacterium tuberculosis. J. Biol. Chem. 281, 3866–3875. doi: 10.1074/jbc.M505556200
de Chastellier, C. (2008). EM analysis of phagosomes. Methods Mol. Biol. 445, 261–285. doi: 10.1007/978-1-59745-157-4_17
Dhouib, R., Ducret, A., Hubert, P., Carriere, F., Dukan, S., and Canaan, S. (2011). Watching intracellular lipolysis in mycobacteria using time lapse fluorescence microscopy. Biochim. Biophys. Acta 1811, 234–241. doi: 10.1016/j.bbalip.2011.01.001
Garton, N., Christensen, H., Minnikin, D., Adegbola, R., and Barer, M. (2002). Intracellular lipophilic inclusions of mycobacteria in vitro and in sputum. Microbiology 148, 2951–2958. doi: 10.1099/00221287-148-10-2951
Gomez-Hernandez, A., Beneit, N., Diaz-Castroverde, S., and Escribano, O. (2016). Differential role of adipose tissues in obesity and related metabolic and vascular complications. Int. J. Endocrinol. 2016:1216783. doi: 10.1155/2016/1216783
Gutierrez, M. C., Brisse, S., Brosch, R., Fabre, M., Omais, B., Marmiesse, M., et al. (2005). Ancient origin and gene mosaicism of the progenitor of Mycobacterium tuberculosis. PLoS Pathog. 1:e5. doi: 10.1371/journal.ppat.0010005
Hoffman, A. M., and Dow, S. W. (2016). Concise review: stem cell trials using companion animal disease models. Stem Cells 34, 1709–1729. doi: 10.1002/stem.2377
Kim, J. S., Ryu, M. J., Byun, E. H., Kim, W. S., Whang, J., Min, K. N., et al. (2011). Differential immune response of adipocytes to virulent and attenuated Mycobacterium tuberculosis. Microbes Infect. 13, 1242–1251. doi: 10.1016/j.micinf.2011.07.002
Koeck, J. L., Fabre, M., Simon, F., Daffe, M., Garnotel, E., Matan, A. B., et al. (2011). Clinical characteristics of the smooth tubercle bacilli 'Mycobacterium canettii' infection suggest the existence of an environmental reservoir. Clin. Microbiol. Infect. 17, 1013–1019. doi: 10.1111/j.1469-0691.2010.03347.x
Korzeniewski, C., and Callewaert, D. M. (1983). An enzyme-release assay for natural cytotoxicity. J. Immunol. Methods 64, 313–320. doi: 10.1016/0022-1759(83)90438-6
Low, K. L., Rao, P. S., Shui, G., Bendt, A. K., Pethe, K., Dick, T., et al. (2009). Triacylglycerol utilization is required for regrowth of in vitro hypoxic nonreplicating Mycobacterium bovis bacillus Calmette-Guerin. J. Bacteriol. 191, 5037–5043. doi: 10.1128/JB.00530-09
Low, K. L., Shui, G., Natter, K., Yeo, W. K., Kohlwein, S. D., Dick, T., et al. (2010). Lipid droplet-associated proteins are involved in the biosynthesis and hydrolysis of triacylglycerol in Mycobacterium bovis bacillus Calmette-Guerin. J. Biol. Chem. 285, 21662–21670. doi: 10.1074/jbc.M110.135731
Mattos, K. A., D'Avila, H., Rodrigues, L. S., Oliveira, V. G., Sarno, E. N., Atella, G. C., et al. (2010). Lipid droplet formation in leprosy: toll-like receptor-regulated organelles involved in eicosanoid formation and Mycobacterium leprae pathogenesis. J. Leukoc. Biol. 87, 371–384. doi: 10.1189/jlb.0609433
Mattos, K. A., Lara, F. A., Oliveira, V. G., Rodrigues, L. S., D'Avila, H., Melo, R. C., et al. (2011). Modulation of lipid droplets by Mycobacterium leprae in Schwann cells: a putative mechanism for host lipid acquisition and bacterial survival in phagosomes. Cell. Microbiol. 13, 259–273. doi: 10.1111/j.1462-5822.2010.01533.x
Mauss, H., and Levy, F. M. (1972). Involvement of adipose tissue in experimental tuberculosis of the mouse. Pathol. Microbiol. 38, 333–345.
McKinney, J. D., Honer zu Bentrup, K., Munoz-Elias, E. J., Miczak, A., Chen, B., Chan, W. T., et al. (2000). Persistence of Mycobacterium tuberculosis in macrophages and mice requires the glyoxylate shunt enzyme isocitrate lyase. Nature 406, 735–738. doi: 10.1038/35021074
Morrison, S., and McGee, S. L. (2015). 3T3-L1 adipocytes display phenotypic characteristics of multiple adipocyte lineages. Adipocyte 4, 295–302. doi: 10.1080/21623945.2015.1040612
Nedergaard, J., Bengtsson, T., and Cannon, B. (2007). Unexpected evidence for active brown adipose tissue in adult humans. Am. J. Physiol. Endocrinol. Metab. 293, E444–E452. doi: 10.1152/ajpendo.00691.2006
Neyrolles, O., Hernandez-Pando, R., Pietri-Rouxel, F., Fornes, P., Tailleux, L., Payan, J. A., et al. (2006). Is adipose tissue a place for Mycobacterium tuberculosis persistence? PLoS ONE 1:e43. doi: 10.1371/journal.pone.0000043
Ouchi, N., Parker, J. L., Lugus, J. J., and Walsh, K. (2011). Adipokines in inflammation and metabolic disease. Nat. Rev. Immunol. 11, 85–97. doi: 10.1038/nri2921
Peyron, P., Vaubourgeix, J., Poquet, Y., Levillain, F., Botanch, C., Bardou, F., et al. (2008). Foamy macrophages from tuberculous patients' granulomas constitute a nutrient-rich reservoir for M. tuberculosis persistence. PLoS Pathog. 4:e1000204. doi: 10.1371/journal.ppat.1000204
Rastogi, S., Agarwal, P., and Krishnan, M. Y. (2016). Use of an adipocyte model to study the transcriptional adaptation of Mycobacterium tuberculosis to store and degrade host fat. Int. J. Mycobacteriol. 5, 92–98. doi: 10.1016/j.ijmyco.2015.10.003
Russell, D. G., Cardona, P. J., Kim, M. J., Allain, S., and Altare, F. (2009). Foamy macrophages and the progression of the human tuberculosis granuloma. Nat. Immunol. 10, 943–948. doi: 10.1038/ni.1781
Santucci, P., Bouzid, F., Smichi, N., Poncin, I., Kremer, L., De Chastellier, C., et al. (2016). Experimental models of foamy macrophages and approaches for dissecting the mechanisms of lipid accumulation and consumption during dormancy and reactivation of Tuberculosis. Front. Cell. Infect. Microbiol. 6:122. doi: 10.3389/fcimb.2016.00122
Shimizu, I., Aprahamian, T., Kikuchi, R., Shimizu, A., Papanicolaou, K. N., MacLauchlan, S., et al. (2014). Vascular rarefaction mediates whitening of brown fat in obesity. J. Clin. Invest. 124, 2099–2112. doi: 10.1172/JCI71643
Keywords: tuberculosis, Mycobacterium canettii, Mycobacterium tuberculosis, adipose tissues, adipocyte
Citation: Bouzid F, Brégeon F, Poncin I, Weber P, Drancourt M and Canaan S (2017) Mycobacterium canettii Infection of Adipose Tissues. Front. Cell. Infect. Microbiol. 7:189. doi: 10.3389/fcimb.2017.00189
Received: 23 February 2017; Accepted: 02 May 2017;
Published: 17 May 2017.
Edited by:
Anthony Baughn, University of Minnesota, United StatesReviewed by:
Siouxsie Wiles, University of Auckland, New ZealandBrian Weinrick, Albert Einstein College of Medicine, United States
Murugesan V. S. Rajaram, Ohio State University at Columbus, United States
Copyright © 2017 Bouzid, Brégeon, Poncin, Weber, Drancourt and Canaan. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Stéphane Canaan, c3RlcGhhbmUuY2FuYWFuQGltbS5jbnJzLmZy