 Muhammad Shahid1†
Muhammad Shahid1† Jianfang Wang2†
Jianfang Wang2† Xiaolong Gu1
Xiaolong Gu1 Wei Chen1
Wei Chen1 Tariq Ali1
Tariq Ali1 Jian Gao1Dandan Han1Rui Yang3
Jian Gao1Dandan Han1Rui Yang3 Séamus Fanning4*
Séamus Fanning4* Bo Han1*
Bo Han1*- 1College of Veterinary Medicine, China Agricultural University, Beijing, China
- 2Beijing Key Laboratory of Traditional Chinese Veterinary Medicine, Beijing University of Agriculture, Beijing, China
- 3Beijing Key Laboratory for Agricultural Application and New Technique, Beijing University of Agriculture, Beijing, China
- 4UCD-Centre for Food Safety, School of Public Health, Physiotherapy and Sports Science, University College Dublin, Dublin, Ireland
Prototheca zopfii infections are becoming global concerns in humans and animals. Bovine protothecal mastitis is characterized by deteriorating milk quality and quantity, thus imparting huge economic losses to dairy industry. Previous published studies mostly focused on the prevalence and characterization of P. zopfii from mastitis. However, the ultrastructural pathomorphological changes associated with apoptosis in bovine mammary epithelial cells (bMECs) are not studied yet. Therefore, in this study we aimed to evaluate the in vitro comparative apoptotic potential of P. zopfii genotype-I and -II on bMECs using flow cytometry, scanning electron microscopy (SEM), and transmission electron microscopy (TEM). The results showed fast growth rate and higher adhesion capability of genotype-II in bMECs as compared with genotype-I. The viability of bMECs infected with P. zopfii genotype-II was significantly decreased after 12 h (p < 0.05) and 24 h (p < 0.01) in comparison with control cells. Contrary, genotype-I couldn't show any significant effects on cell viability. Moreover, after infection of bMECs with genotype-II, the apoptosis increased significantly at 12 h (p < 0.05) and 24 h (p < 0.01) as compared with control group. Genotype-I couldn't display any significant effects on cell apoptosis. The host specificity of P. zopfii was also tested in mouse osteoblast cells, and the results suggest that genotype-I and -II could not cause any significant apoptosis in these cell lines. SEM interpreted the pathomorphological alterations in bMECs after infection. Adhesion of P. zopfii with cells and further disruption of cytomembrane validated the apoptosis caused by genotype-II under SEM. While genotype-1 couldn't cause any significant apoptosis in bMECs. Furthermore, genotype-II induced apoptotic manifested specific ultrastructure features, like cytoplasmic cavitation, swollen mitochondria, pyknosis, cytomembrane disruption, and appearance of apoptotic bodies under TEM. The findings of the current study revealed that genotype-II has the capability to invade and survive within the bMECs, thus imparting significant damages to the mammary cells which result in apoptosis. This study represents the first insights into the pathomorphological and ultrastructure features of apoptosis in bMECs induced by P. zopfii genotype-II.
Introduction
Bovine mastitis is an inflammatory condition of mammary gland which is characterized by pathological, physiological, and bacteriological changes in the udder that affects the milk quality and quantity (Sharma et al., 2011). Different pathogens are involved in bovine mastitis which invades the udder through the teat canal in milking animals. These pathogens proliferate into the mammary gland and produce harmful effects which consequences in inflammation of udder (Seegers et al., 2003; Halasa et al., 2007). Protothecosis, mostly caused by Prototheca zopfii, clinically appears in the form of mastitis in dairy cattle (Ahrholdt et al., 2012). Bovine mammary protothecosis leads to subclinical and clinical bovine mastitis. In acute clinical form, protothecal mastitis is generally characterized by elevated body temperature (up to 40°C), pain, hot edema of the udder, loss of appetite, and reluctance to move. The chronic form of protothecal mastitis is accompanied by slight pain, hard tissue consistency with pasty edema in the udder, as well as pronounced decrease in milk production and there is elevated somatic cell count, especially macrophages, which may even lead to culling of cow and ultimately result in high economic losses (Wawron et al., 2013). The somatic cells are mainly cells of the immune system which mostly include leukocytes (75%), that is neutrophils, macrophages, lymphocytes, polymorphonuclear cells, and the disrupted mammary epithelial cells (25%) (Pillai et al., 2001). The leukocytes are part of natural defense mechanism to fight the infection and to help in the repair of damaged tissues. The epithelial cells of mammary gland are normally sloughed off and renewed, but in infection the number considerably increase. Somatic cells count is an important indicator of intramammary infection (IMI) as well as the quality of milk for human consumption (Schukken et al., 2003).
P. zopfii, a fungus like alga, is divided into two genotypes (type-I and -II) according to biochemical, serological, and genetic assays (Roesler et al., 2006). Additionally, P. zopfii genotype-II, P. wickerhamii, and P. blaschkeae have been mainly associated with bovine mastitis (Marques et al., 2008; Capra et al., 2014); while, P. wickerhamii and P. cutis are mostly related to human diseases (Lass-Florl and Mayr, 2007; Satoh et al., 2010). Protothecosis is also important from public health point of view, as P. zopfii is usually associated with bovine mastitis which can be transferred to human being through consumption of contaminated milk (Bozzo et al., 2014). Cutaneous or disseminated infection and olecranon bursitis are main form of protothecosis in human being (Lass-Florl and Mayr, 2007).
Bovine udder tissue is the main target site of protothecal infection and ascending infection can lead to long-term involvement. Chronic mastitis caused by P. zopfii is dominated by macrophages in bovine mammary gland (Cheville et al., 1984). Their study reported that mammary gland infected with P. zopfii showed different degenerative stages of algal cells contained by macrophages in the interstitial spaces, seclude between alveolar epithelial cells and lumen of alveoli under electron microscopy (Cheville et al., 1984). Recent protothecal bovine mastitis outbreaks have been reported from China (Gao et al., 2012; Chang et al., 2013; Shahid et al., 2016), Canada (Pieper et al., 2012), Iran (Zaini et al., 2012), Japan (Sobukawa et al., 2012), Mexico (Mayorga et al., 2012), Poland (Jagielski et al., 2011), and Romania (Bouari et al., 2011); hence, it led to considerable economic losses in dairy herds. The findings of these studies investigated that genotype-II is the causative agent of bovine mastitis. However, there is still a paucity of published literature about the ultrastructure features of apoptosis in bMECs after P. zopfii exposure.
Apoptosis in cells after infection or any injury is characterized by typical ultrastructural features, such as shrinkage of cell, chromatin condensation, pyknosis, nuclear fragmentation, and appearance of apoptotic bodies (Gajewska et al., 2005). The invasion and survival of P. zopfii type-II in bMECs and its role in apoptosis is not comprehensively studied yet. Previous published studies regarding protothecal mastitis caused by P. zopfii are mostly focused on prevalence, molecular characterization of P. zopfii (Jagielski et al., 2010; Gao et al., 2012; Chang et al., 2013; Bozzo et al., 2014; Shahid et al., 2016), and on the determination of some immunogenic proteins and virulence determinants of P. zopfii genotype-II (Irrgang et al., 2015). Therefore, the current study was designed with the objective to evaluate the ultrastructural features of apoptosis and to study the comparative apoptotic potentials of P. zopfii genotype-I and -II on bMECs. To the best of our knowledge, this is the first in vitro study unveiling the ultrastructural features of apoptosis in bMECs infected with P. zopfii genotype-I and -II.
Materials and Methods
Isolation and Identification of P. zopfii
P. zopfii genotype-I and -II were previously isolated from milk samples of dairy cows suffering from bovine mastitis and were stored in our laboratory at College of Veterinary Medicine, China Agricultural University, Beijing (Gao et al., 2012). The strains were revived on sabouraud dextrose agar (SDA; Difco™, Becton Dickison, Sparks MD, USA) at 37°C for 72 h and were used to assess their ability to invade and to evaluate their apoptotic potential in bovine mammary epithelial cells (bMECs). Prior to each experiment, fresh P. zopfii type-I and -II suspensions were used as described formerly (Shahid et al., 2016). P. zopfii was very carefully handled and all the experimental steps were carried out in biological safety cabinets, and we ensured the biosafety level 2 (BSL 2) precautions according to guidelines of Centers of Disease Control and Prevention (CDC).
Cell Morphology of P. zopfii
Colonies of the P. zopfii genotype-I and -II were resuspended in phosphate buffered saline with pH 7.4 (PBS) and evaluated using an optical microscope (CKX 41SF Olympus, Japan).
Colonial Morphology
P. zopfii genotype-I and II were cultivated on SDA to observe the morphological characteristics of colonies, the cultured plates were incubated at 37°C under aerobic condition for 5–7 days.
Exponential Growth Curve
The exponential growth curve features of P. zopfii were analyzed in triplicate. The strains of genotype-I and -II were inoculated into 10 mL sabouraud dextrose broth (SDB; Difco™, Becton Dickison, Sparks MD, USA) in 15 mL centrifuge tube incubated at 37°C on a rotary shaker (150 rpm). Algal growth was determined by counting CFU per 4 h for first 3 days, after that CFU was counted per 24 for 7–8 days on SDA.
Cell Culture
Primary bMECs were isolated and characterized as described previously (Liu et al., 2014). Cells were cultivated in growth medium containing Dulbecco's modified Eagle's (DMEM)/F12 medium (HyClone, USA) supplemented with 10% Fetal Bovine Serum (FBS; Gibco, Grand island, NY, USA) and antimicrobial agents (100 U/mL penicillin, 100 mg/mL streptomycin and 1 mg/mL amphotericin B) in cell culture plates (Corning, Corning, NY, USA). Cells were incubated in 5% CO2 at 37°C, and cells from 2–8 passages were used for experiments.
Infection Experiments
For infection experiments, P. zopfii genotype-I and -II were grown on SDA for 48 h at 37°C, single colony was sub-cultivated into SDB for 72 h, after that P. zopfii were collected and suspended in DMEM/F12. Finally, the concentrations genotype-I and -II was adjusted to 5 × 105 CFU/mL for all experiments.
Adhesion Capability and Survival of P. zopfii Genotypes
Adhesion capability and time course of the survival of P. zopfii genotypes-I and -II in bMECs was determined as described previously (Pereyra et al., 2016; Chen et al., 2017). The bMECs were infected with genotypes-I and –II at a 5:1 multiplicity of infection (MOI; ratio of P. zopfii to bMECs). To study the adhesion capability and survival of P. zopfii genotypes-I and -II in bMEC cells, samples were taken at 1, 4, 8, 12, and 24 h and cultured on SDA after infection.
Cell Viability Assay in bMECs
MTT cell proliferation assay kit (Trevigen, Gaithersburg, MD, USA) was used to assess the cell viability according to Verma et al. (2010) with slight modification. The bMECs at a density of 1 × 105 cells per well were cultured in 96-well plates. After being treated as mentioned above, the cells were washed three times with PBS (pH 7.2), and cells were incubated with 100 μL medium and 10 μL of the activated MTT solution at 37°C for 4 h, after that treated with 50 μL DMSO (Sigma-Aldrich), and the absorption was measured by microplate reader (SpectraMax 190, Molecular Devices Corporation, Sunnyvale, CA, USA) at 570 nm. Cell viability was determined as the percentages (%) of the control.
Apoptosis in bMECs
The infected bMECs with P. zopfii genotype-I and -II were stained with FITC annexin V/PI apoptosis detection kit (Becton Dickinson, Franklin Lakes, NJ, USA) as described previously (Liu et al., 2014). Apoptosis rate was expressed using a FACSAria flow cytometry (Becton Dickinson, USA).
Detection of Bax Expression in Infected bMECs Using Western Blotting
To further validate the apoptosis in bMECs caused by P. zopfii genotypes, Bax protein expression was detected by western blot analysis. The bMECs were infected with P. zopfii genotype-I and -II for 0, 4, 12, and 24 h at 37°C with 5% CO2. Total protein was extracted from protothecal infected cells with protein extraction kit (KeyGEN, Nanjing, China). The protein quantification was carried out using bicinchoninic acid (BCA) protein assay kit (Beyotime, Haimen, China). Equivalent proteins from each sample were separated by sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) and transferred to a polyvinylidene difluoride membrane (PVDF; Millipore, MA, USA). Subsequently, the membranes were blocked in 5% bovine serum albumin (BSA; Gibco) and incubated with specific primary antibody for Bax (1:200, Santa, USA) and β-actin (1:1,000, Cell Signaling Technology, USA) overnight at 4°C followed by incubation with HRP-conjugated secondary antibody (1:5,000) for 1 h at room temperature. Finally, the bands were visualized using an enhanced chemiluminescence system (ECL, Beyotime, Haimen, China). The results were normalized to β-actin using Image J (National Institutes of Mental Health, Bethesda, MD, USA).
Apoptosis in Mouse Osteoblast Cells
To explore the host cells specificity of P. zopfii, we infected the mouse osteoblast cell line MC3T3-E1 with P. zopfii genotype-I and -II. The MC3T3-E1 was purchased from American Type Culture Collection (ATCC, Rockville, MD, USA). These cell lines were maintained in minimal essential medium (α-MEM medium; Gibco) in the presence of 10% (v/v) fetal bovine serum (FBS; Gibco) and antimicrobials (100 U/mL penicillin and 100 U/mL streptomycin), incubated at 37°C in the presence of 5% CO2. The infection protocol and flow cytometry assay was same as described earlier.
Scanning Electron Microscopy (SEM) and Transmission Electron Microscopy (TEM) of bMECs
The infected cells with P. zopfii genotype-I and -II were washed three times with phosphate buffer saline (PBS; pH 7.2) and the cells were fixed with 2.5% glutaraldehyde (Novon Scientific, China; pH 7.4) as described previously (Chen et al., 2017). Then cells were dehydrated by a graded ethanol from 30 to 100%, dried by critical-point drying method, then gold-coated with E-1010 Ion Sputter Coater (Hitachi, Japan). Finally, the changes in bMECs were observed by SEM (TESCAN 5136, Czech Republic). TEM was used for ultrastructure analysis of bMECs infected with P. zopfii genotype-I and -II. The bMECs were cultured and challenged as mentioned above and after washing with PBS cells were fixed with 2% glutaraldehyde-1% paraformaldehyde solution (Sinopharm Chemical Reagent Co., Ltd., China; pH 7.2) for 45 min at room temperature. After washing with PBS, the fixed cells were harvested with a rubber scraper (Fisher Scientific, Nepean, ON, Canada) in 1.5 mL microcentrifuge tubes. Further steps in the processing of the TEM samples were conducted at Electron Microscopy Unit of Beijing University of Agriculture, China. Briefly, the pellet was washed 5–7 times with phosphate buffer saline (pH 7.2), post-fixed in a 0.5% osmium tetroxide for 2 h, washed 5–7 times with PBS. After five times washing with PBS, the pellet was dehydrated with graded alcohol series (30, 50, 70, 80, 90, 95, and 100%). The pellet was embedded in 50/50 LR white embedding resin (Electron Microscopic Sciences, Hatfield, PA, USA) and pure ethanol solution for 1 h followed by a pure resin solution for overnight at 4°C and incubated for 1 h at 60°C for polymerization. Thin slices (100 nm) were cut by an Ultracutmicrotome (Leica EM, Germany) with a glass knife. Sections were positioned on copper grid, stained with 2% uranyl acetate and lead citrate and were viewed on a transmission electron microscopy (HITACHI H-7650, Japan) at 80 kV. The imaging was done using a 4 K Gatan CCD camera using the iTEM software.
Statistical Analysis
Statistical evaluation for triplicate experiments was carried out using student's t-test. Data are presented as the mean values ± standard deviation. p < 0.05 was considered as significant, while p < 0.01 was highly significant.
Results
Morphological Characteristics of P. zopfii
Under optical microscope the cells of P. zopfii genotype-I were spherical in shape whereas that of genotype-II were oval, elliptical, and smaller in size than genotype-I. Figures 1A,B indicates the different phases of microbial growth, having numerous stem cells called sporangia, forming sporangiospores or endospores by internal division. Under the electron microscopy, there were different morphological feature of genotype-II and genotype-I sporangia. Genotype-II revealed typical features of sporangia with spherical to oval, containing sporangiospores as shown in Figure 2A; while type-I showed compact, round shaped and twice in size of type-II (Figure 2B).
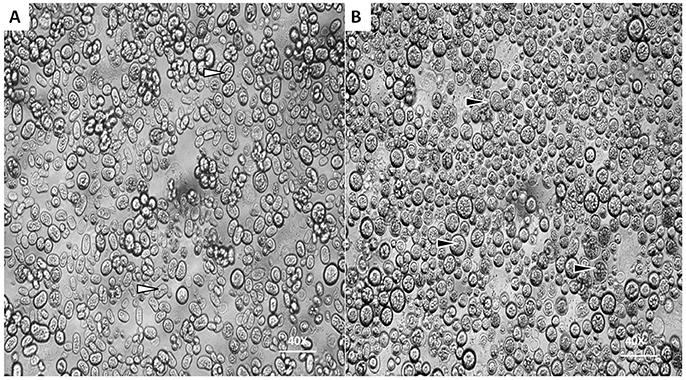
Figure 1. Morphological characteristics of P. zopfii under optical microscope. (A,B) Showing the different cells sizes of P. zopfii genotype-I (black arrow) and genotype-II (white arrow) with internally dividing cells with endospores (40x).
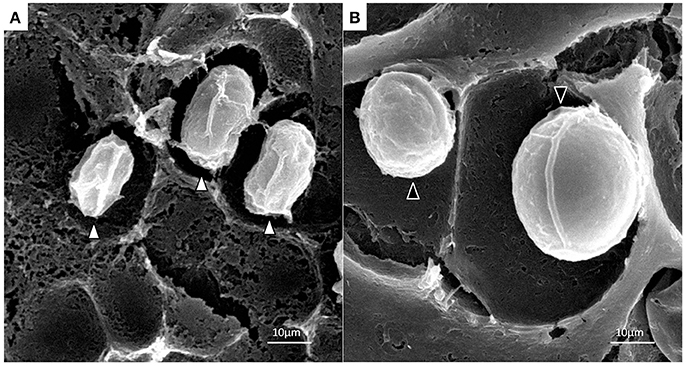
Figure 2. Scanning electron micrographs of P. zopfii genotype-II (A) and -I (B) cells. Indicating the difference in sporangial size and shape of genotype-I and -II.
Colonial Morphology
The colonies of both P. zopfii showed typical morphological characteristics like in case of genotype-II grayish white in color, with a central protrusion with granular surface and serrated shape (Figures 3A,C), while genotype-I displayed the creamy white colonies with bulging smooth surface (Figures 3B,D).
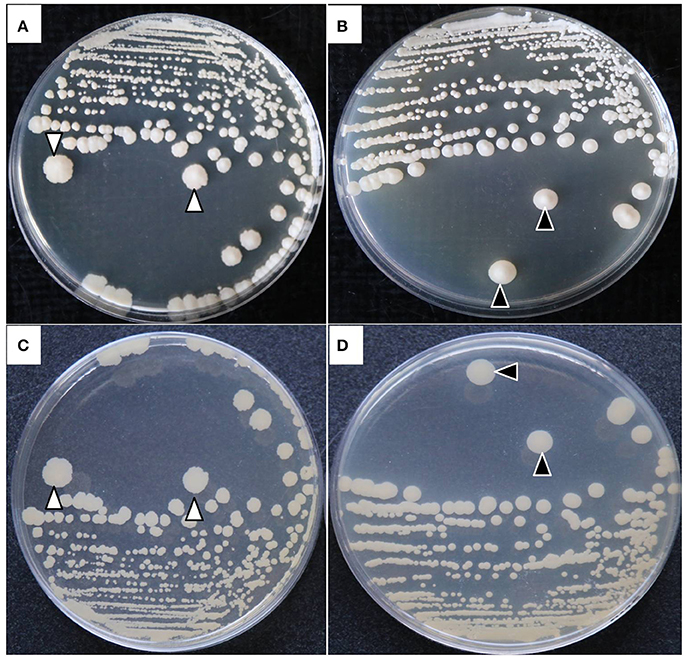
Figure 3. Colonial morphological characteristics of P. zopfii genotype-I and -II. (A) Type-II with central irregular protrusion like granular surface and serrated borders, (B) Type-I with smooth bulging surface at the center of colony. (C,D) Back view of the colonies.
Exponential Growth Curve
The exponential growth of P. zopfii is shown in Figure 4. According to growth curve of CFU, genotype-I and -II entered in logarithmic growth phase after 30 and 12 h and achieved the peak level at 144 and 96 h, respectively. This indicated that the genotype-II has faster growth kinetics than genotype-I.
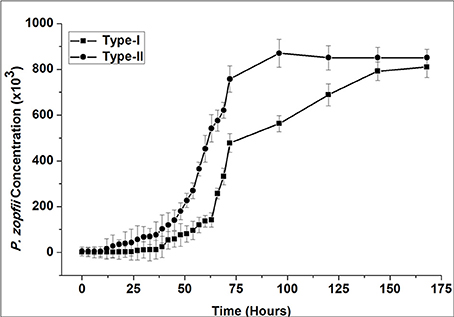
Figure 4. Exponential growth curve of P. zopfii genotype-I and -II. P. zopfii genotype-I and -II were initially grown on sabouraud dextrose agar (SDA; Difco™), sub-cultivated into sabouraud dextrose broth for 72 h. Exponential growth curve, as determined from colony forming unit, showed genotype-I and -II entered in logarithmic growth phase after 30 and 12 h and achieved the peak level at 144 and 96 h, respectively.
Survival of P. zopfii Genotypes-I and -II in bMECs
Morphological changes in the bMECs monolayers following infection with genotype-I and -II are shown in Figures 5A–G. In case of genotype-I infection, the bMECs appeared similar to the control (uninfected cells); whereas type-II showed slight detachment from bMECs monolayer after 12 h (Figure 5F) and affected the integrity of monolayer cells after 24 h, as obvious from bMECs detachment in Figure 5G. The results depicted that the numbers of P. zopfii genotypes-I and -II increased steadily with passage of time as elaborated in Figure 5H. The genotype-II exhibited stronger adhesion capacity as compared to type-I.

Figure 5. Adhesion of P. zopfii and its effect on bMECs morphology at different time of infection. (A) control, (B,C) 4 h, (D,E) 12 h, and (F,G) 24 h. Black arrowheads show the type-II, while white arrowheads show type-I cells adhesion. (B,C,E,F) There were no significant changes in bMECs, (D) Depicting slight cells morphological changes in type-II at 12 h, (G) Showing disruption of monolayer of bMECs in type-II at 24 h. (H) Time course of the survival and adhesive capability of P. zopfii in bMECs that increased significantly with passage of time. Data indicated as Mean ± SD of three independent experiments. **p < 0.01.
Cell Viability
The results of the MTT assay revealed that viability of cells infected with P. zopfii genotype-II was significantly decreased after 12 and 24 h as compared with the uninfected cells (Figure 6), the cell viability values significantly declined from 85.3 ± 3.7% (p < 0.05) to 48.17 ± 10.38% (p < 0.01), respectively. In contrast, genotype-I didn't show significant effect on cell viability as compared with control.
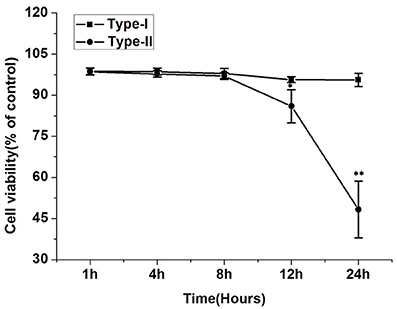
Figure 6. Effect of P. zopfii on the cells viability of bMECs by MTT assays. The viability of bMECs infected with P. zopfii genotype-II for 12 and 24 h was significantly decreased in comparison to uninfected group. Results are presented as Mean ± SD. *p < 0.05, **p < 0.01.
Apoptosis in bMECs
The results of flow cytometry assays showed significant changes in cell profile after exposure to P. zopfii genotype-II for 12 and 24 h. These results revealed that genotype-II significantly induced apoptosis in bMECs. After exposure, genotype-II showed high rise in apoptosis in comparison with control group as apoptotic rate was increased from 14.91 ± 5.54% (p < 0.05) to 63.83 ± 23.28% (p < 0.01) at 12 to 24 h, respectively. While, in case of genotype-I, there was non-significant effect on cells apoptosis (Figures 7A,B).
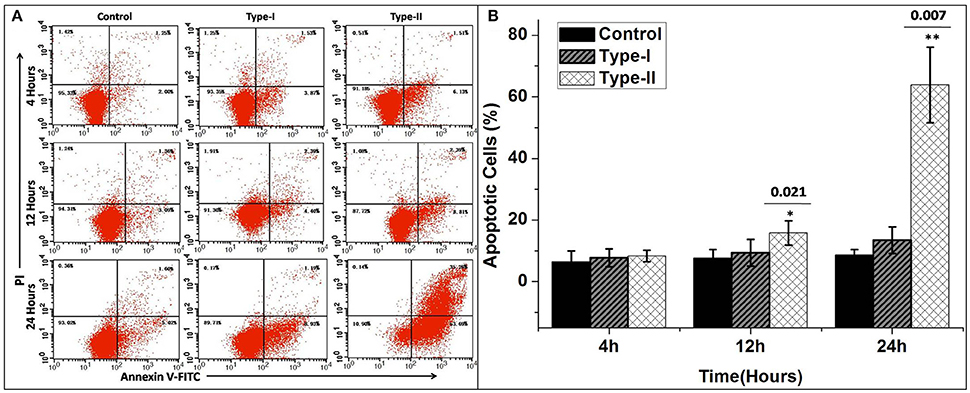
Figure 7. Apoptosis of bMECs induced by P. zopfii. Cells distributions were examined on basis of annexin V binding and propidium iodide (PI) dye uptake. The florescence was measured by flow cytometry. (A) Demonstrates two dimensional scatter plots of annexin V vs. PI. (B) Percentage of apoptotic cells. Control (uninfected group), type-I (5 × 105 CFU/ml) infection, type-II (5 × 105 CFU/ml) infection for 4, 12, and 24 h. Data of triplicate experiments are shown as Mean ± SD. *p < 0.05, **p < 0.01.
Bax Protein Expression in bMECs after P. zopfii Genotype -I and -II Infection
The results of Bax protein expression, using western blotting, in bMECs infected with P. zopfii genotype-I and -II are shown in Figure 8. Genotype-I showed non-significant effects on the expression Bax in bMECs as compared to control (i.e., at 0 h of infection). However, bMECs infected with genotype-II for 4, 12, and 24 h resulted in significant (p < 0.01) increase in Bax level (Figures 8A,B). This also validates the apoptotic effects of genotype-II on bMECs.
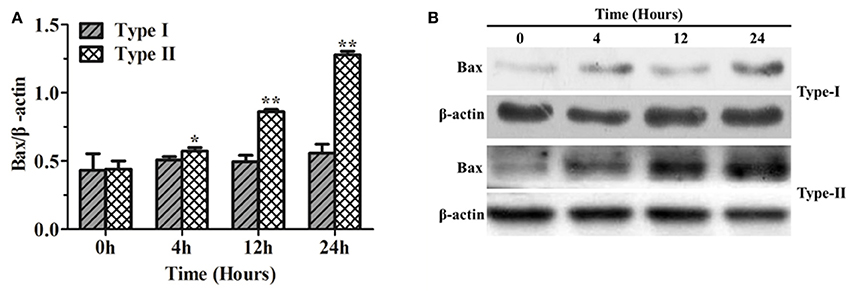
Figure 8. Bax protein expression in bMECs infected with Prototheca zopfii by western blot analysis. (A) P. zopfii genotype-II induces Bax protein expression. bMECs were treated with P. zopfii genotype-I and -II at 4, 12, and 24 h. (B) Bax protein levels were assessed by western blot analysis. Data are presented as mean ± SD of three separate experiments. *p < 0.05, **p < 0.01.
Apoptosis in Mouse Osteoblast Cells (MC3T3-E1)
Figure 9 shows the apoptosis rate in MC3T3-E1 cells infected with P. zopfii genotype-I and -II for 0, 4, 12, and 24 h. The results elaborated non-significant difference in apoptosis rate in MC3T3-E1 cells at different time intervals (Figures 9A,B). This might suggest that P. zopfii genotype-I and -II didn't possess specificity toward these cell lines.
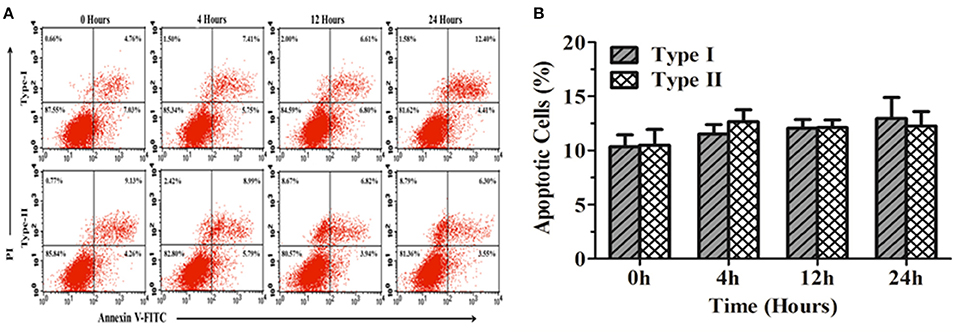
Figure 9. Apoptosis of mouse osteoblast cells (MC3T3-E1) induced by P. zopfii. (A) Represents two dimensional scatter plots of annexin V vs. PI. (B) Percentage of apoptotic cells. Control (0 h), genotype-I (5 × 105 CFU/mL) infection, type-II (5 × 105 CFU/ml) infection for 4, 12, and 24 h.
Ultrastructural Changes in bMECs Infected with P. zopfii
Under electron microscopy there were no morphological changes in uninfected bMECs (Figure 10A). In genotype-I infected group, there was also no prominent pathological alterations in the structure of cells; however the genotype-I have the adhesive capability with bMECs without damaging the cell integrity, this adhesion of genotype-I increased with the passage of time (Figures 10B–I). Whereas, genotype-II has showed adhesion without any structural alteration at 4 h as shown in Figure 11B. There were marked morphological changes in bMECs after exposure at 12–24 h. At 12 h, strong adhesion to bMECs was observed, which increased with passage of time, and disruption of cytomembrane initiated, this indicates the start of apoptosis (Figures 11C–F). At 24 h of exposure, the adhesion of genotype-II further increased (Figure 11G) and the cytomembrane was totally damaged (Figures 11H,I). In addition, the extent of cell disruption, the disappearance of microvilli and rupture of cellular membrane were prominent in the genotype-II exposure at 24 h (Figures 11G–I).
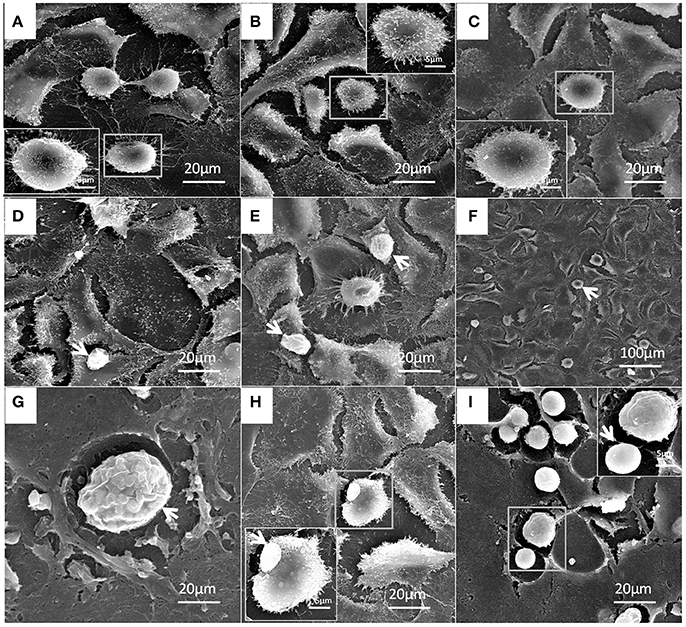
Figure 10. Patho-morphological alterations in bMECs after infection of P. zopfii genotype-I under SEM. (A) Control, (B) 4 h, and (C) 12 h showing no effect on cytomembrane of bMECs. (D,E) Cells adhesion of type-I with cells (white arrow) at 12 h. (F–I) Type-I infected epithelial cells at 24 h is depicting no morphological changes (white arrow). (G) Sporangia of type-I with many sporangiospores attached with the cells (white arrow). (H) Type-I attached with the cells (white arrow). (I) Different developmental stages of sporangia and sporangiospores (white arrow).
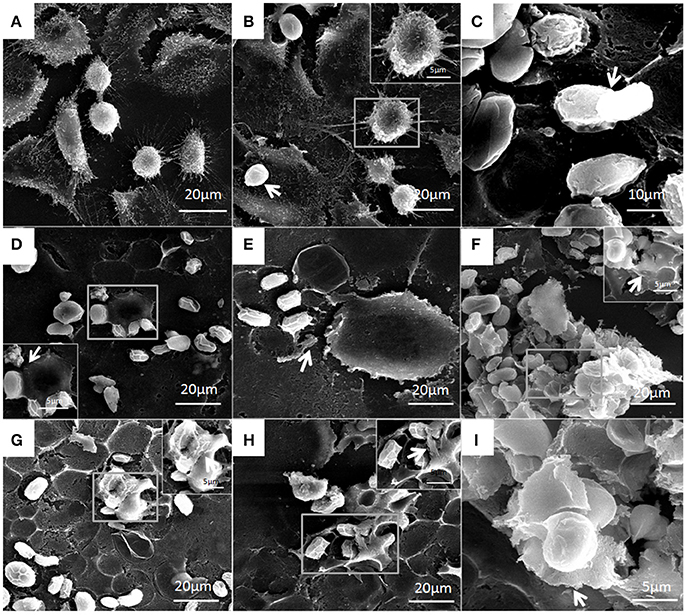
Figure 11. Structural features of bMECs after infection of P. zopfii genotype-II observed under SEM at 4, 12, and 24 h. (A) Control, (B) Shows no effect on cytomembrane but type-II shows the cells adhesion at 4 h (white arrow), and (C) Indicates the cells adhesion of sporangiospores and sporangia at 12 h (white arrow). (D,E) Depicts the start of apoptotic effect on cells at 12 h. (F) Represents the rapture of sporangia and shows sporopollenin (white arrow) after maturation and release of sporangiospores at 24 h. (G–I) Indicates the degree of cells disruption, disappearance of microvilli, and no more intact of cellular membrane (white arrow) at 24 h.
The ultrastructural features of bMECs as observed under TEM demonstrated various stages of P. zopfii genotype-II infection and its cytopathic and apoptotic effects in comparison with genotype-I. The normal bMECs maintained in DMEM with 10% FBS exhibited normal structure like intact cytoplasm and organelles (including intact mitochondria and endoplasmic reticulum structures) as in Figures 12A–C. After the adhesion of genotype-II at 4 h, slight increase in size of mitochondria was noted (Figure 12D), whereas, some epithelial cells extended around the adherent algae with the appearance of pseudopod like structures (Figure 12E). At 12 h, there were vacuolization in cytoplasm that originated from the swelling of endoplasmic reticulum as shown in Figure 12F and some of the mitochondria were swollen (Figure 12I). The genotype-II sporangiospores enclosed within the endocytic vacuoles were also seen in cells (Figures 12G–H). At 24 h post-infection, as elaborated in Figures 13B–I, all ultrastructural characteristics of apoptosis were confronted in the infected bMECs, such as cytoplasmic cavitation (Figures 13C,D,G), pyknosis (Figure 13D), cytomembrane disruption (Figures 13D,E), and appearance of apoptotic bodies (Figures 13G,H) as compared to control group (Figure 13A). The monolayer of bMECs was damaged and the released algae were seen near the remnant of disrupted epithelial cells as shown in Figure 13E at 24 h. The swollen mitochondrial structures and mitochondrial disruption were also observed (Figures 13F,H,I), and the cytoplasm was replaced by phagocytic vacuoles (Figure 13I) at 24 h after P. zopfii genotype-II infection. The average increase in mitochondrial size was 1.11, 1.69, and 2.63 μm at 4, 12, and 24 h post infection, respectively, as compared to the mitochondrial size of 0.98 μm of the control bMECs. Whereas, some epithelial cells having large vacuoles without any algae were observed, these vacuoles were the swelling of the endoplasmic reticulum (Figures 13F–H).
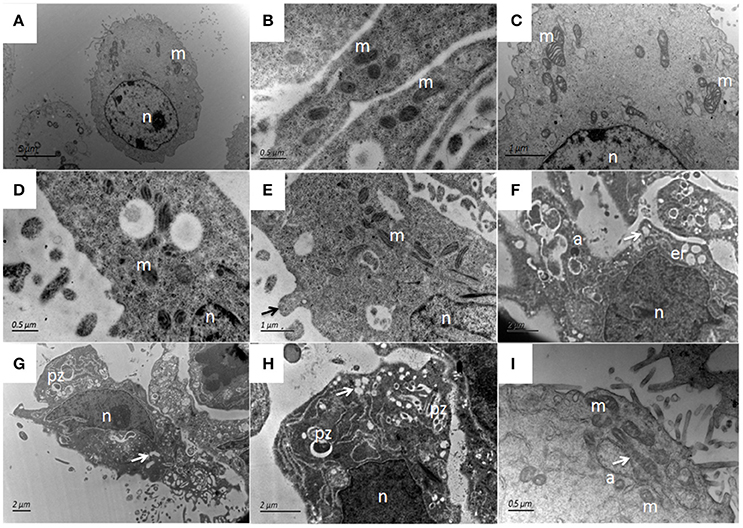
Figure 12. Ultrastructural pathological changes in bMECs after 4 h and 12 h infection of P. zopfii type-II under TEM. (A–C) Control bMECs. (D) Shows slight increase in size of mitochondria in bMECs at 4 h and (E) Depicts bulging (pseudopod) of bMECs toward P. zopfii type-II at 4 h (black arrow). (F–H) Vacuolization in cytoplasm at 12 h (white arrow) originated from the swelling of endoplasmic reticulum and mitochondria; with condense nucleus and appearance of apoptotic bodies. (G,H) Engulfment of algae by cells. (I) Showing higher magnification with swollen mitochondria (white arrow). pz, P. zopfii; n, nucleus; m, mitochondrion; er, endoplasmic reticulum; a, apoptotic bodies.
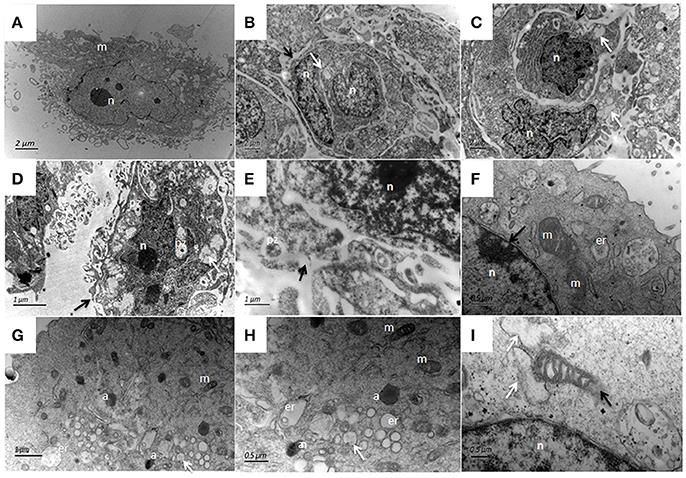
Figure 13. Patho-morphological changes of bMECs after 24 h infection with P. zopfii type-II under TEM. (A) Non-infected cells that show abundant mitochondria in cytoplasm, (B,C) Disruption of cytomembrane (black arrow), pyknosis of nucleus, swelling of endoplasmic reticulum as cytoplasmic vacuoles (white arrow). (D,E) Changes in cytoplasmic features, P. zopfii enclosed within the cytoplasmic vacuoles, the chromatin condensation, fragmentation, and cell membrane breakage (black arrow). (F) higher magnification indicating swollen mitochondria with crista degeneration and expansion of perinuclear space (black arrow). (G) Appearance of apoptotic bodies and swelling of endoplasmic reticulum as cytoplasmic vacuole. (H) higher magnification with apoptotic bodies. (I) Mitochondria disruption (black arrow) and cytoplasm replaced by phagocytic vacuoles (white arrow). pz, P. zopfii; n, nucleus; m, mitochondrion; er, endoplasmic reticulum; a, apoptotic bodies.
In P. zopfii genotype-I exposure, there was no marked ultrastructural changes in the bMECs at 4 h (Figure 14C) and 12 h (Figure 14D), similar like control cells (Figures 14A,B). While at 24 h post-infection the cytoplasmic membrane remained intact (Figures 14E,F,I), with slight vacuolization with enclosed algae (Figures 14G,H).
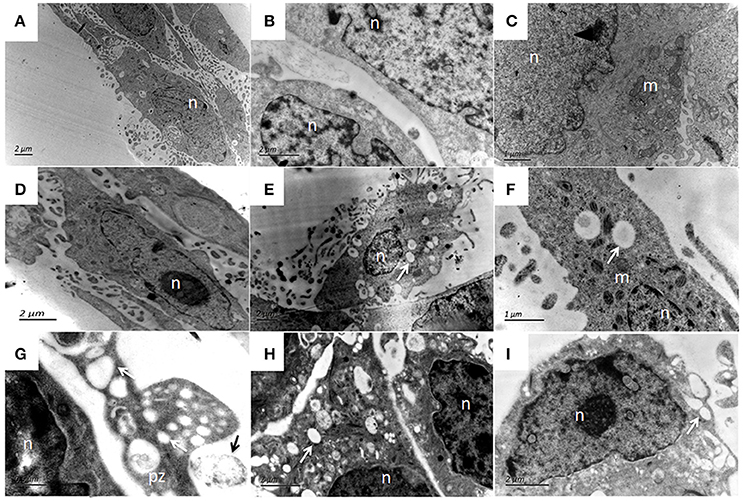
Figure 14. Ultrastructural features of bMECs after P. zopfii genotype-I infection at different time interval under TEM. (A,B) Uninfected cells with mitochondria, intact cytoplasmic, and nuclear membrane. (C) Cells at 4 h and (D) Cells at 12 h indicating no morphological changes in bMECs. (E–I) TEM of P. zopfii type-I infected cells at 24 h showing cytoplasm cavitation (white arrow), (G,H) Sporangiospores enclosed within the vacuoles, pseudopod like bulging surrounding sporangiospores and (G) Adherence of algae (black arrow) to epithelial cells surface. pz, P. zopfii; n, nucleus; m, mitochondrion; er, endoplasmic reticulum.
Discussion
Chronic bovine mastitis caused by P. zopfii is increasingly reported in dairy herds and is liable for huge economic losses in dairy herds (Bozzo et al., 2014). Protothecosis is also important from public health point of view, as P. zopfii could be transmitted to humans through contaminated milk and cause intestinal infection like enteritis (Melville et al., 1999). The present study reported the ultrastructural features of apoptosis and to study the cytotoxic and apoptotic potentials of P. zopfii genotype-I and -II on bMECs. It also elucidated the morphological changes in bMECs after P. zopfii genotype-II in vitro infection under scanning electron microscope, which explored the relationship of the genotypes with the etiology of protothecal bovine mastitis and demonstrated that genotype-II caused severe apoptosis in bMECs in time dependent manner. As SEM depicted the adhesion of P. zopfii to the host cells which is the most important step in the infection process followed by colonization of the pathogen within the host cells. This is in agreement with previous studies (Melchior et al., 2006; Akers et al., 2015), which explored the adhesion and biofilm formation of P. zopfii associated with mastitis. Here, we comprehensively studied, for very first time, the pathomorphological and ultrastructure features of apoptosis in bMECs infected with P. zopfii genotype-II. Although, chronic protothecal mastitis causes increase in the somatic cell counts which is predominated by macrophages (Cheville et al., 1984); however, bMECs are of special importance in the mammary gland and its defense mechanism. These bMECs are at front line defense, play an important role in immune response of the mammary gland and can quickly respond against the invading pathogens (Gilbert et al., 2013). Thus, keeping the importance of bMECs in the udder health and the increasing prevalence of protothecal mastitis, this study was designed. Kwiecinski also recommended further studies for detail mechanism of apoptotic effects and control of protothecosis (Kwiecinski, 2015).
The results of the present study confirmed that P. zopfii genotype-II have higher apoptotic potential in bMECs. It is in agreement with the reports of Moller et al. in Germany (Moller et al., 2007), Aouay et al. in Belgium (Aouay et al., 2008), Osumi et al. in Japan (Osumi et al., 2008), Jagielski et al. in Poland (Jagielski et al., 2010), and Shahid et al. in China (Shahid et al., 2016). We determined Bax protein expression, by western blotting, to further validate the apoptosis in bMECs infected with P. zopfii genotypes. Our results showed that P. zopfii genotype-II for 4, 12, and 24 h resulted in a significant (p < 0.01) increase of Bax level. These finding supported our previous report that P. zopfii genotype-II induced apoptosis through reactive oxygen species (ROS) generation (Shahid et al., 2017). Increasing ROS level might activate the mitochondrial Bax that lead to apoptosis in bMECs after P. zopfii genotype-II infection. Bax is a proapoptotic factor of the Bcl-2 family proteins, and it plays an important role in the mitochondrial apoptosis pathway (Li et al., 2013). In normal living cells, Bax exists predominantly in the cytosol, and during apoptosis it migrates to the membrane of mitochondria (Yu et al., 2008). Our findings support these results that increase level of Bax expression in P. zopfii genotype-II infection induced apoptosis. In addition, the host specificity of genotype-II was also tested in mouse osteoblast cell lines (MC3T3-E1) and the results revealed that both genotype-I and -II could not cause any significant apoptosis in these cells. This might suggest that genotype-II does not have specificity toward the MC3T3-E1 cell lines.
In the current study, we examined ultrastructural variations in bMECs after P. zopfii genotype-I and -II exposure. P. zopfii genotype-II may proliferate in the bMECs and this proliferative stage may survive for some time following phagocytosis that was demonstrated by the observation of intracellular sporangia after 12 h of infection. Whereas, in genotype-I only few sporangia were observed inside the epithelial cells after 24 h of infection. The ultrastructure variations noted in the current study are compatible with the morphological features of apoptosis and para-apoptosis (Asher et al., 1995). Asher et al. (1995) were the first who described the paraptosis, which is characterized by cytoplasmic vacuolization due to swollen mitochondria and endoplasmic reticulum. Moreover, Sperandio et al. explained the form of para-apoptosis, called paraptosis, with similar ultrastructure features by cellular characteristics and response to inhibitors of apoptosis (Sperandio et al., 2000).
In the natural infection of protothecosis, it was observed that macrophages are sites for proliferation of P. zopfii and survives for some time after phagocytosis as observation of intact intracellular sporangia (Jensen et al., 1998). In an experimental infection of Prototheca in the BALB/c mice demonstrated that cells with whole organism have spindle shaped and/or oval nuclei with abundant dilated rough endoplasmic reticulum. Cells near the completion of pathogen digestion, presented irregular and/or independent nuclei with often one or two distinct nucleoli and also the cytoplasm was often replaced by phagocytic vacuoles (Horiuchi and Masuzawa, 1995). Electron microscopy of mammary gland from cows infected with P. zopfii elaborated that the macrophages with algae were distinctly enlarged due to primarily from reduplication of the Golgi complexes and its associated vesicles. This study also described that intracellular P. zopfii was degenerated and comprised of intact cell wall profiles that contained membrane fragment without nuclei and cytoplasmic organelles (Cheville et al., 1984). This is in line with the current findings that P. zopfii genotype-II survives within the bMECs and cause pathogenic effects, such as cytoplasmic cavitation, swollen mitochondrial structures, pyknosis, cytomembrane disruption, and appearance of apoptotic bodies.
In our study, the results of cell viability, apoptosis, scanning electron microscopy, and transmission electron microscopy depicted that both genotype-I and -II were persistent in bMECs, but the in vitro pathogenic effect of genotype-II were more profound as previously reported in vivo infection (Chang et al., 2013).
It is concluded that P. zopfii genotype-II have the capability to invade and survive within the bMECs, causing pathomorphological alterations associated with apoptosis in infected bMECs. Our findings support the previous observations that P. zopfii genotype-II is the causative agents of bovine mastitis (Jensen et al., 1998; Morandi et al., 2016). To date, there is no treatment to profoundly eliminate chronic mastitis in dairy cow herds; therefore, further investigative study should be carried out to understand the pathogenesis of chronic mastitis caused by P. zopfii genotype-II.
Author Contributions
Designed the research: BH and MS; performed the experiments: MS, JW, WC, TA, XG, DH, and RY; analyzed the molecular data: MS and together with JG; Wrote the paper: MS with collaboration of JW and RY; and revised and corrected the manuscript: SF. All authors have read and approved the final manuscript.
Funding
This research was supported by the Chinese Twelfth “Five-year” National Science and Technology Support Project (No. 2012BAD12B03), National Education Ministry's Major Project (No. 313054), Specialized Research Fund for the Doctoral Program of Higher Education (SRFDP) State Education Ministry (No. 20120008110042), High-end Foreign Experts Recruitment Program (No. GDT20141100043), and the National Natural Science Foundation of China (No. 3151101034 and No. 31572587).
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
Ahrholdt, J., Murugaiyan, J., Straubinger, R. K., Jagielski, T., and Roesler, U. (2012). Epidemiological analysis of worldwide bovine, canine and human clinical Prototheca isolates by PCR genotyping and MALDI-TOF mass spectrometry proteomic phenotyping. Med. Mycol. 50, 234–243. doi: 10.3109/13693786.2011.597445
Akers, K. S., Cardile, A. P., Wenke, J. C., and Murray, C. K. (2015). Biofilm formation by clinical isolates and its relevance to clinical infections. Adv. Exp. Med. Biol. 830, 1–28. doi: 10.1007/978-3-319-11038-7_1
Aouay, A., Coppee, F., Cloet, S., Cuvelier, P., Belayew, A., Lagneau, P. E., et al. (2008). Molecular characterization of Prototheca strains isolated from bovine mastitis. J. Mycol. Med. 18, 224–227. doi: 10.1016/j.mycmed.2008.10.002
Asher, E., Payne, C. M., and Bernstein, C. (1995). Evaluation of cell death in EBV-transformed lymphocytes using agarose gel electrophoresis, light microscopy and electron microscopy. II. Induction of non-classic apoptosis (“para-apoptosis”) by tritiated thymidine. Leuk. Lymphoma 19, 107–119. doi: 10.3109/10428199509059664
Bouari, M. C., Fit, N., Rapuntean, S., Nadas, G., Gal, A., Bolfa, P., et al. (2011). In vitro evaluation of the antimicrobial properties of some plant essential oils against clinical isolates of Prototheca spp. Rom. Biotechnol. Lett. 16, 6146–6152.
Bozzo, G., Bonerba, E., Di Pinto, A., Bolzoni, G., Ceci, E., Mottola, A., et al. (2014). Occurrence of Prototheca spp.in cow milk samples. New Microbiol. 37, 459–464.
Capra, E., Cremonesi, P., Cortimiglia, C., Bignoli, G., Ricchi, M., Moroni, P., et al. (2014). Simultaneous identification by multiplex PCR of major Prototheca spp. isolated from bovine and buffalo intramammary infection and bulk tank. Lett. Appl. Microbiol. 59, 642–647. doi: 10.1111/lam.12326
Chang, R., Yang, Q., Liu, G., Liu, Y., Zheng, B., Su, J., et al. (2013). Treatment with gentamicin on a murine model of protothecal mastitis. Mycopathologia 175, 241–248. doi: 10.1007/s11046-013-9628-1
Chen, W., Liu, Y., Zhang, L., Gu, X., Liu, G., Shahid, M., et al. (2017). Nocardia cyriacigeogica from bovine mastitis induced in vitro apoptosis of bovine mammary epithelial cells via activation of mitochondrial-caspase pathway. Front. Cell. Infect. Microbiol. 7:194. doi: 10.3389/fcimb.2017.00194
Cheville, N. F., McDonald, J., and Richard, J. (1984). Ultrastructure of Prototheca zopfii in bovine granulomatous mastitis. Vet. Pathol. 21, 241–248. doi: 10.1177/030098588402100312
Gajewska, M., Gajkowska, B., and Motyl, T. (2005). Apoptosis and autophagy induced by TGF-b1 in bovine mammary epithelial BME-UV1 cells. J. Physiol. Pharmacol. 56, 143–157.
Gao, J., Zhang, H. Q., He, J. Z., He, Y. H., Li, S. M., Hou, R. G., et al. (2012). Characterization of Prototheca zopfii associated with outbreak of bovine clinical mastitis in herd of Beijing, China. Mycopathologia 173, 275–281. doi: 10.1007/s11046-011-9510-y
Gilbert, F. B., Cunha, P., Jensen, K., Glass, E. J., Foucras, G., Robert-Granie, C., et al. (2013). Differential response of bovine mammary epithelial cells to Staphylococcus aureus or Escherichia coli agonists of the innate immune system. Vet. Res. 44, 1–23. doi: 10.1186/1297-9716-44-40
Halasa, T., Huijps, K., Osteras, O., and Hogeveen, H. (2007). Economic effects of bovine mastitis and mastitis management. Vet. Q. 29, 18–31. doi: 10.1080/01652176.2007.9695224
Horiuchi, Y., and Masuzawa, M. (1995). Electron microscopic observations of epithelioid cell granulomas experimentally induced by prototheca in the skin of mice. J. Dermatol. 22, 643–649. doi: 10.1111/j.1346-8138.1995.tb03891.x
Irrgang, A., Murugaiyan, J., Weise, C., Azab, W., and Roesler, U. (2015). Well-known surface and extracellular antigens of pathogenic microorganisms among the immunodominant proteins of the infectious microalgae Prototheca zopfii. Front. Cell Infect. Microbiol. 5:67. doi: 10.3389/fcimb.2015.00067
Jagielski, T., Lassa, H., Ahrholdt, J., Malinowski, E., and Roesler, U. (2011). Genotyping of bovine Prototheca mastitis isolates from Poland. Vet. Microbiol. 149, 283–287. doi: 10.1016/j.vetmic.2010.09.034
Jagielski, T., Lassa, H., Ahrholdt, J., Roesler, U., and Malinowski, E. (2010). Molecular characterization of Polish Prototheca zopfii mastitis isolates and first isolation of Prototheca blaschkeae in Poland. Polish J. Vet. Sci. 13, 725–729. doi: 10.2478/v10181-010-0025-0
Jensen, H. E., Aalbaek, B., Bloch, B., and Huda, A. (1998). Bovine mammary protothecosis due to Prototheca zopfii. Med. Mycol. 36, 89–95. doi: 10.1080/02681219880000151
Kwiecinski, J. (2015). Biofilm formation by pathogenic Prototheca algae. Lett. Appl. Microbiol. 61, 511–517. doi: 10.1111/lam.12497
Lass-Florl, C., and Mayr, A. (2007). Human Protothecosis. Clin. Microbiol. Rev. 20, 230–242. doi: 10.1128/CMR.00032-06
Li, Z., Meng, J., Xu, T. J., Qin, X. Y., and Zhou, X. D. (2013). Sodium selenite induces apoptosis in colon cancer cells via Bax-dependent mitochondrial pathway. Eur. Rev. Med. Pharmacol. Sci. 17, 216–2171.
Liu, Y., Chen, W., Ali, T., Alkasir, R., Yin, J., Liu, G., et al. (2014). Staphylococcal enterotoxin H induced apoptosis of bovine mammary epithelial cells in vitro. Toxin 6, 3552–3567. doi: 10.3390/toxins6123552
Marques, S., Silva, E., Kraft, C., Carvalheira, J., Videira, A., Huss, V. A., et al. (2008). Bovine mastitis associated with Prototheca blaschkae. J. Clin. Microbiol. 46, 1941–1945. doi: 10.1128/JCM.00323-08
Mayorga, J., Barba-Gomez, J. F., Verduzco-Martinez, A. P., Munoz-Estrada, V. F., and Welsh, O. (2012). Protothecosis. Clin. Dermatol. 30, 432–436. doi: 10.1016/j.clindermatol.2011.09.016
Melchior, M. B., Vaarkamp, H., and Fink-Gremmels, J. (2006). Biofilms: a role in recurrent mastitis infections? Vet. J. 171, 398–407. doi: 10.1016/j.tvjl.2005.01.006
Melville, P. A., Watanabe, E. T., Benites, N. R., Ribeiro, A. R., Silva, J. A., Garino Junior, F., et al. (1999). Evaluation of the susceptibility of Prototheca zopfii to milk pasteurization. Mycopathologia 146, 79–82. doi: 10.1023/A:1007005729711
Moller, A., Truyen, U., and Roesler, U. (2007). Prototheca zopfii genotype 2: the causative agent of bovine protothecal mastitis? Vet. Microbiol. 120, 370–374. doi: 10.1016/j.vetmic.2006.10.039
Morandi, S., Cremonesi, P., Capra, E., Silvetti, T., Decimo, M., Bianchini, V., et al. (2016). Molecular typing and differences in biofilm formation and antibiotic susceptibilities among Prototheca strains isolated in Italy and Brazil. J. Diary Sci. 99, 6436–6445. doi: 10.3168/jds.2016-10900
Osumi, T., Kishimoto, Y., Kano, R., Maruyama, H., Onozaki, M., Makimura, K., et al. (2008). Prototheca zopfii genotypes isolated from cow barns and bovine mastitis in Japan. Vet. Microbiol. 131, 419–423. doi: 10.1016/j.vetmic.2008.04.012
Pereyra, E. A., Picech, F., Renna, M. S., Baravalle, C., Andreotti, C. S., Russi, R., et al. (2016). Detection of Staphylococcus aureus adhesion and biofilm-producing genes and their expression during internalization in bovine mammary epithelial cells. Vet. Microbiol. 183, 69–77. doi: 10.1016/j.vetmic.2015.12.002
Pieper, L., Godkin, A., Roesler, U., Polleichtner, A., Slavic, D., Leslie, K. E., et al. (2012). Herd characteristics and cow-level factors associated with Prototheca mastitis on dairy farms in Ontario, Canada. J. Dairy Sci. 95, 5635–5644. doi: 10.3168/jds.2011-5106
Pillai, S. R., Kunze, E., Sordillo, L. M., and Jayarao, B. M. (2001). Application of differential inflammatory cell count as a tool to monitor udder health. J. Dairy Sci. 84, 1413–1420. doi: 10.3168/jds.S0022-0302(01)70173-7
Roesler, U., Moller, A., Hensel, A., Baumann, D., and Truyen, U. (2006). Diversity within the current algal species Prototheca zopfii: a proposal for two Prototheca zopfii genotypes and description of a novel species, Prototheca blaschkeae sp. nov. Int. J. Syst. Evol. Microbiol. 56, 1419–1425. doi: 10.1099/ijs.0.63892-0
Satoh, K., Ooe, K., Nagayama, H., and Makimura, K. (2010). Prototheca cutis sp. nov., a newly discovered pathogen of Protothecosis isolated from inflamed human skin. Int. J. Syst. Evol. Microbiol. 60, 1236–1240. doi: 10.1099/ijs.0.016402-0
Schukken, Y. H., Wilson, D. J., Welcome, F., Garrison-Tikofsky, L., and Gonzalez, R. N. (2003). Monitoring udder health and milk quality using somatic cell counts. Vet. Res. 34, 579–596. doi: 10.1051/vetres:2003028
Seegers, H., Fourichon, C., and Beaudeau, F. (2003). Production effects related to mastitis and mastitis economics in dairy cattle herds. Vet. Res. 34, 475–491. doi: 10.1051/vetres:2003027
Shahid, M., Ali, T., Zhang, L., Hou, R., Zhang, S., Ding, L., et al. (2016). Characterization of Prototheca zopfii genotypes isolated from cases of bovine mastitis and cow barns in China. Mycopathologia 181, 185–195. doi: 10.1007/s11046-015-9951-9
Shahid, M., Gao, J., Zhou, Y., Liu, G., Ali, T., Deng, Y., et al. (2017). Prototheca zopfii isolated from bovine mastitis induced oxidative stress and apoptosis in bovine mammary epithelial cells. Oncotarget 8, 31938–31947. doi: 10.18632/oncotarget.16653
Sharma, N., Singh, N. K., and Bhadwal, M. S. (2011). Relationship of somatic cell count and mastitis: an overview. Asian Aust. J. Anim. Sci. 24, 429–438. doi: 10.5713/ajas.2011.10233
Sobukawa, H., Yamaguchi, S., Kano, R., Ito, T., Suzuki, K., Onozaki, M., et al. (2012). Molecular typing of Prototheca zopfii from bovine mastitis in Japan. J. Dairy Sci. 95, 4442–4446. doi: 10.3168/jds.2011-5101
Sperandio, S., de Belle, I., and Bredesen, D. E. (2000). An alternative, nonapoptotic form of programmed cell death. Proc. Natl. Acad. Sci. U.S.A. 97, 14376–14381. doi: 10.1073/pnas.97.26.14376
Verma, A., Prasad, K. N., Singh, A. K., Nyati, K. K., Gupta, R. K., and Paliwal, V. K. (2010). Evaluation of the MTT lymphocyte proliferation assay for the diagnosis of neurocysticercosis. J. Microbiol. Methods 81, 175–178. doi: 10.1016/j.mimet.2010.03.001
Wawron, W., Bochniarz, M., Piech, T., Lopuszyński, W., and Wysocki, J. (2013). Outbreak of protothecal mastitis in a herd of dairy cows in Poland. Bull. Vet. Inst. Pulawy 57, 335–339. doi: 10.2478/bvip-2013-0058
Yu, C., Friday, B. B., Yang, L., Atadja, P., Wigle, D., Sarkaria, J., et al. (2008). Mitochondrial Bax translocation partially mediates synergistic cytotoxicity between histone deacetylase inhibitors and proteasome inhibitors in glioma cells. Neuro Oncol. 10, 309–319. doi: 10.1215/15228517-2007-063
Keywords: P. zopfii, bovine mastitis, bMECs, apoptosis, SEM, TEM
Citation: Shahid M, Wang J, Gu X, Chen W, Ali T, Gao J, Han D, Yang R, Fanning S and Han B (2017) Prototheca zopfii Induced Ultrastructural Features Associated with Apoptosis in Bovine Mammary Epithelial Cells. Front. Cell. Infect. Microbiol. 7:299. doi: 10.3389/fcimb.2017.00299
Received: 01 March 2017; Accepted: 19 June 2017;
Published: 13 July 2017.
Edited by:
Amal O. Amer, The Ohio State University, Department of Internal Medicine, United StatesReviewed by:
Jianwu Pei, American Animal Health, Inc., United StatesNicholas A. Wallace, Kansas State University, United States
Copyright © 2017 Shahid, Wang, Gu, Chen, Ali, Gao, Han, Yang, Fanning and Han. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Séamus Fanning, c2Zhbm5pbmdAdWNkLmll
Bo Han, aGFuYm9AY2F1LmVkdS5jbg==
†These authors have contributed equally to this work.