 Carlos M. Suligoy1
Carlos M. Suligoy1 Santiago M. Lattar1
Santiago M. Lattar1 Mariángeles Noto Llana1Cintia D. González1
Mariángeles Noto Llana1Cintia D. González1 Lucía P. Alvarez1
Lucía P. Alvarez1 D. Ashley Robinson2Marisa I. Gómez1
D. Ashley Robinson2Marisa I. Gómez1 Fernanda R. Buzzola1
Fernanda R. Buzzola1 Daniel O. Sordelli1*
Daniel O. Sordelli1*- 1Instituto de Investigaciones en Microbiología y Parasitología Médica, University of Buenos Aires - CONICET, Buenos Aires, Argentina
- 2Department of Microbiology and Immunology, University of Mississippi Medical Center, Jackson, MS, United States
Selection pressures exerted on Staphylococcus aureus by host factors may lead to the emergence of mutants better adapted to the evolving conditions at the infection site. This study was aimed at identifying the changes that occur in S. aureus exposed to the host defense mechanisms during chronic osteomyelitis and evaluating whether these changes affect the virulence of the organism. Genome assessment of two S. aureus isolates collected 13 months apart (HU-85a and HU-85c) from a host with chronic osteomyelitis was made by whole genome sequencing. Agr functionality was assessed by qRT-PCR. Isolates were tested in a rat model of osteomyelitis and the bacterial load (CFU/tibia) and the morphometric osteomyelitic index (OI) were determined. The ability of the isolates to trigger the release of proinflammatory cytokines was determined on macrophages in culture. Persistence of S. aureus within the host resulted in an agrC frameshift mutation that likely led to the observed phenotype. The capacity to cause bone tissue damage and trigger proinflammatory cytokines by macrophages of the agr-deficient, unencapsulated derivative (HU-85c) was decreased when compared with those of the isogenic CP8-capsulated parental strain (HU-85a). By comparison, no significant differences were found in the bacterial load or the OI from rats challenged with isogenic Reynolds strains [CP5, CP8, and non-typeable (NT)], indicating that lack of CP expression alone was not likely responsible for the reduced capacity to cause tissue damage in HU-85c compared with HU-85a. The production of biofilm was significantly increased in the isogenic derivative HU-85c. Lack of agr-dependent factors makes S. aureus less virulent during chronic osteomyelitis and alteration of the agr functionality seems to permit better adaptation of S. aureus to the chronically infected host.
Introduction
Staphylococcus aureus is an ubiquitous opportunistic pathogen that can infect, replicate and persist in humans thus making this species a worldwide threat to public health. Although S. aureus may colonize mucosal surfaces of healthy humans with unnoticeable or mild clinical features, it has the invasive potential to generate diverse life threatening infections. S. aureus is one of the most prevalent and difficult-to-eradicate pathogens causing prosthetic device-associated osteomyelitis (Tong et al., 2015). In addition to the widespread emergence of methicillin-resistant S. aureus (MRSA) (Prestinaci et al., 2015), the control of S. aureus infections is hampered by the evolution of S. aureus with low-level vancomycin resistance (Howden et al., 2008).
The S. aureus genome carries a vast array of genes coding for virulence and immune evasion factors. Many of these genes are conserved in the S. aureus genome and display broad functionality with considerable redundancy (Tong et al., 2015). It is believed that initially the ability of S. aureus to regulate expression of certain virulence factors permits its adaptation to defined microenvironments in the infected host. Once S. aureus is well established at the infected tissue, and the infection becomes refractory to antibiotic treatment, certain regulatory traits may be fixed by spontaneous mutations occurring during chronic infection (Tuchscherr et al., 2010), likely due to selection pressure exerted by a vast number of yet undefined host factors. These variants are more suitable to evade immune defense mechanisms than the parental infecting wild type and are able to generate chronic infection refractory to antibiotic treatment, not necessarily associated to bacterial antibiotic resistance. A few phenotypic features such as loss of capsule (types 5 and 8) expression, loss of SSR repeats in the protein A Xr region and small colony variant (SCV) emergence have been recognized and may be considered endpoints in the short term evolution of S. aureus in the chronically infected host (Proctor et al., 2006; Lattar et al., 2009; Garofalo et al., 2012; Das et al., 2016). The present study was designed to investigate the main changes that occurred in S. aureus in a chronically infected patient persisting over a period exceeding 1 year.
Materials and Methods
Bacterial Strains
S. aureus clinical isolates HU-85a and HU-85c were collected as the initial isolate and 13 months later, respectively, from the same infection site (right tibia) of a 20-year-old man with chronic osteomyelitis (Hospital de Clínicas “General San Martín,” Universidad de Buenos Aires, Argentina) (Lattar et al., 2012). Species was confirmed by a species-specific PCR (Martineau et al., 1998). S. aureus strain Reynolds (CP5) and its isogenic derivatives Reynolds CP8 and Reynolds NT (Watts et al., 2005) were provided by Dr. Jean C. Lee (Division of Infectious Diseases, Department of Medicine, Brigham and Women's Hospital and Harvard Medical School, Boston, MA, USA). All strains were kept frozen in trypticase soy broth (TSB) with 20% glycerol at −20°C and S. aureus was routinely cultured at 37°C for 24 h on Columbia agar supplemented with 2% NaCl. To prepare bacterial inocula S. aureus was cultured on Columbia salt agar and incubated at 37°C for 24 h. Bacterial cells were harvested and suspended to the appropriate density in saline.
The Osteomyelitis Rat Model
Outbred Wistar adult rats weighing 250–350 g were purchased from local vendors and were kept at the vivarium of the Instituto de Investigaciones en Microbiología y Parasitología Médica (IMPaM), Buenos Aires. Animal care was in accordance with the recommendations of the guidelines set forth by: the 11 report of the BVAAWF/FRAME/RSPCA/UFAW Joint Working Group on Refinement (Hawkins et al., 2011). The animal research protocol utilized in this study was approved by the “Comité Institucional para el Uso y Cuidado de los Animales de Laboratorio,” through resolution N° 2269 issued on September 17, 2014 by the “Consejo Directivo de la Facultad de Medicina, Universidad de Buenos Aires,” Argentina. In order to produce bone infection, rats were anesthetized with ketamine/xylazine and a 5 μl suspension containing 1 × 106 CFU of bacteria suspended in fibrin glue (Tissucol kit 1 ml, Baxter Argentina-AG Vienna, Austria) was injected in the left tibia as describe previously (Lattar et al., 2014). Groups of rats were sacrificed 14 weeks after intratibial challenge by exposure to CO2. Both left and right tibias were removed and the morphometric osteomyelitic index (OI) was assessed as detailed previously (Lattar et al., 2014). Afterwards 1 cm bone segments involving the infected zone were sectioned, crushed and homogenized in sterile mortars. Homogenates were quantitatively cultured overnight on trypticase soy agar (TSA) and the number of CFU was determined. To validate the results from the rat model experiments the correlation between the bacterial load and the OI was tested in the experiments. Phenotypic expression of α- and β-haemolysin was performed by evaluating the production of the haemolysis halo in rabbit and goat blood agar (α- and β-haemolysin, respectively). CP production was evaluated by colony immunoblot on TSA plates as described previously (Lee et al., 1990).
Biofilm Formation
Biofilm formation was quantitatively assessed according to a procedure routinely performed in our laboratory (Dotto et al., 2017). Briefly, S. aureus suspensions were placed in sterile 96-well polystyrene microtiter plates. After 24 h incubation at 37°C, each culture final optical density at 595 nm (named ODG) was measured in a microplate reader (Multiskan EX, Thermo Electron Corp., Waltham, MA, USA). The culture medium was then removed and plates were washed twice with phosphate buffered saline (PBS). The biofilms were fixed with 100% methanol, stained with 0.5% crystal violet and washed with tap water. After addition of 30% glacial acetic acid biofilm biomasses were measured by reading the optical density at 595 nm (ODB). The intensity of crystal violet staining was expressed relative to the final culture density measured prior to the biofilm assay (biofilm: ODB)/ODG) and termed “biofilm” in the text for the sake of clarity. Agr-deficient strain S. aureus SA113 (Iordanescu and Surdeanu, 1976; Periasamy et al., 2012), which is a robust biofilm producer, was included in the experiments.
Real Time Quantitative Reverse Transcription (qRT) PCR
Bacterial RNA was extracted from S. aureus cultures in TSB harvested at the post-exponential phase, using Trizol Reagent® (Invitrogen Life Technologies), according to the manufacturer's protocol. RNA was subjected to DNAse treatment using a RQ1 RNAse free DNAse (Promega). cDNA synthesis was performed with an ImProm-II™ Reverse Transcriptase kit (Promega). Quantitative RT-PCR was performed using the SYBR Green PCR Master Mix (Applied Biosystems) equipment and kits. cDNA was subjected to Real time PCR using the following primer pairs: rnaIII-Fw 5′-TTC ACT GTG TCG ATA ATC CA−3′, rnaIII-Rv 5′-TGA TTT CAA TGG CAC AAG AT-3′ (Vaudaux et al., 2002); 16S-Fw 5′-GAT CAG CAT GCT ACG GTG AA-3′ and 16S-Rv 5′-ACC TTC CGA TAC GGC TAC CT-3′. Cycling conditions were: 95°C for 10 min followed by 45 cycles of 95°C for 10 s, 55°C for 10 s and 72°C for 15 s, and 1 cycle of 40°C for 30 s. The 16S gene was used to normalize data. The number of copies of each sample transcript was determined with the aid of the 7500 system SDS software (Applied Biosystems). The (−ΔCT) value represents the difference in threshold cycle (Ct) between the target and control (16S) genes (Livak and Schmittgen, 2001).
Primary Cell Cultures and Cytokine Detection
Mouse peritoneal macrophages were obtained from BALB/c mice by peritoneal lavage. Adherent cells were selected after plating the peritoneal cells in 96-well tissue culture plates. Confluent cells were stimulated 24 h later with the different bacterial strains (2–2.5 × 109 CFU/well). Mouse IL-6 and TNFα were determined quantitatively at 24 h after stimulation by ELISA using specific antibody pairs (Beckton-Dickinson) as described previously (Giai et al., 2016).
DNA Extraction, Whole Genome Sequencing, and Analysis
Genomic DNA was extracted from isolates according to a standard protocol (Pitcher et al., 1989). DNA libraries were prepared using Nextera XT (Illumina, San Diego, CA, USA) and whole genome sequencing was performed using Illumina HiSeq pair-end sequencing with 500 cycles (Instituto Nacional de Tecnología Agropecuaria, Castelar, Buenos Aires, Argentina). FASTQ reads were processed with Trimmomatic (Bolger et al., 2014) to remove bases from the trailing end that fall below a PHRED score of 30. Short reads were assembled using SPAdes v3.10.1 (Bankevich et al., 2012). Contigs less than 500 bp and 30X coverage were discarded. Resulting contigs were ordered using Mauve 2.4.0 (Darling et al., 2010) and strain MRSA252 as reference. The genome was annotated using Prokka 1.12 (Seemann, 2014). Variant calling was performed using Snippy v3.2 (Seemann, 2015). The sequences reported in this paper have been deposited in the NCBI Sequence Read Archive under Bioproject number PRJNA414566.
Statistical Analysis
Groups of data were statistically compared with the Mann-Whitney test for non-parametrics. P < 0.05 were considered statistically significant. Multiple comparisons (three groups of rats) were performed by the Kruskal-Wallis test for non-parametrics. The correlation analysis of CFU vs. OI data was performed with the Spearman test for non-parametrics. Data from in vitro cytokine and biofilm production was analyzed using the Student t-test. The Prism GraphPad software (version 5.0) was used for all statistical analysis.
Results
Isolate Main Features
The characterization of S. aureus isolates HU-85a and HU-85c revealed that both were MSSA, ST188, CC1, spa type t189 and agr type I, suggesting that these longitudinally collected isolates shared a recent common ancestor at the infection site. Since HU-85a was the initial isolate and HU-85c was collected 13 months later from the same infection site, isolate HU-85a is considered to be more reflective of the “parent” isolate. Phenotypic analysis revealed that HU-85c had lost not only the capacity to express capsular polysaccharide 8 (CP8) but also α- and β-haemolysin, compared with HU-85a. These phenotypic features were stable since HU-85c did not regain CP8 or haemolysin expression upon six passages on TSA or blood agar.
Agr Functionality
To test whether HU-85c was an agr-deficient derivative, the expression of RNAIII from the agr locus was assessed by qRT-PCR. The results showed that the relative level of RNAIII expression was significantly decreased in HU-85c when compared with HU-85a (Figure 1).
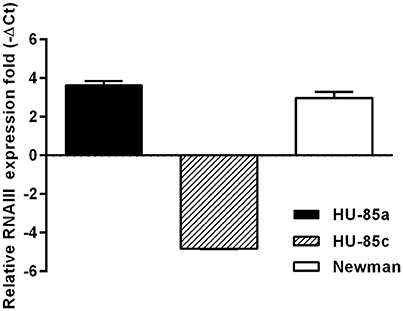
Figure 1. Real Time PCR of agr transcripts from strains HU-85a and HU-85c. Strain Newman was used as reference. Changes in gene expression are shown as normalized mean fold change −ΔCt ± SEM. Data were normalized to 16S expression. The data represent the mean of duplicate measurements from 3 independent experiments.
Sequence Analysis
Comparative sequence analysis of HU-85a and HU-85c revealed two insertions that result in frameshifts in genes coding proteins: one in the known virulence gene regulator agrC and the other in the tgt gene encoding Queuine tRNA-ribosyltransferase, which intervenes in tRNA modification. The lesion in the agr locus consisted of a C insertion in position 471 of agrC resulting in a truncated product. In addition, there were a few other mutations, which are described in Table 1.
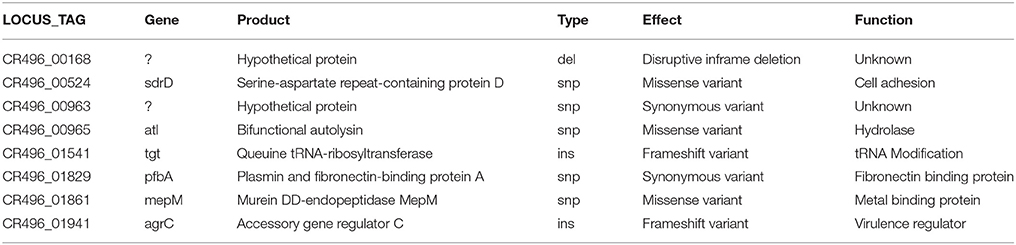
Table 1. Mutations detected by comparative sequence analysis of isolate HU-85a and its isogenic isolate HU-85c.
S. aureus Virulence in an Osteomyelitis Rat Model
The virulence of the two S. aureus isolates was evaluated in the rat model of osteomyelitis through determination of the bacterial load and the OI. Groups of rats infected with parent isolate HU-85a (agr+) were compared with rats infected with the derivative isolate HU-85c (agr-deficient). The bacterial load in the rat tibias and the OI of HU-85c by 14 weeks were significantly lower than those of HU-85a (Figure 2). No unencapsulated or non-haemolytic colonies were recovered from the bones of rats infected with HU-85a by 14 weeks after intratibial challenge. Histopathological analysis revealed that derivative isolate HU-85c caused less tissue damage when compared with the parental isolate HU-85a (Figure 3).
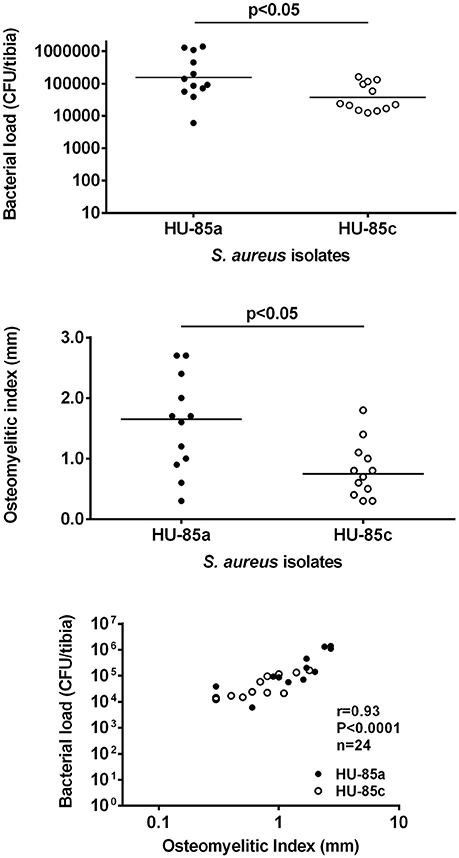
Figure 2. CFU/tibia (upper) and OI (middle) from rats infected with strains HU-85a and HU-85c. Bacterial loads and OI were measured 14 weeks after intratibial challenge. The sample size was n = 9 (HU-85a) and n = 12 (HU-85c), in both panels. The scattergram bars in the upper and middle panels represent the medians. The levels of significance are shown on the charts (Mann-Whitney test). The lower panel shows the correlation of the CFU/bone counts and the OI of all the rats included in the experiments shown in the upper and middle panels (experiment quality control).
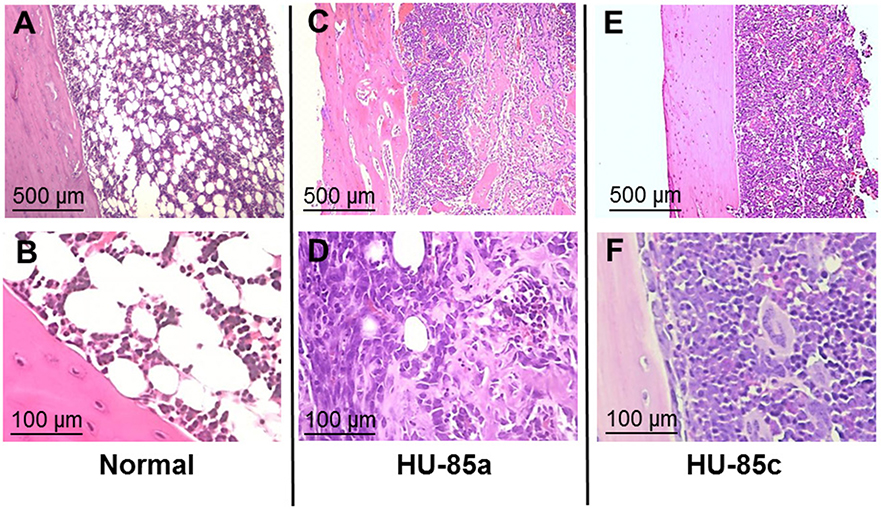
Figure 3. Hematoxylin-eosin-stained sections of bone after intratibial challenge with 106 CFU of S. aureus HU-85a or HU-85c. The strain obtained earlier in the study (HU-85a; C,D) induced marked alterations that include bone cortex with irregular border/necrosis and bone marrow substituted by inflammatory infiltrates and invaded by trabecular structures. The strain obtained later in the study (HU-85c; E,F) induced less pathological changes, including significant inflammatory infiltrate composed of mononuclear cells and reduction of fat cells while the bone cortex exhibited conserved regular borders. (A,B) are stained sections of control, unchallenged tibias that exhibit normal bone marrow with fat tissue and cells, and a bone cortex with laminar structure.
Whereas the loss of CP8 expression by S. aureus HU-85c was due to agrC mutation, the loss of CP is still a trait associated with chronic osteomyelitis (Lattar et al., 2009). In order to ascertain the role of CP expression loss in the osteomyelitis rat model, groups of rats were infected with S. aureus strain Reynolds CP5 and its isogenic CP8 and NT derivatives. The results revealed no significant differences among groups (Supplementary Figure 1). Taken together, these results indicate that the sole loss of CP5 (or CP8) expression by S. aureus may not be solely responsible for the reduced virulence of S. aureus in the rat osteomyelitis model.
Assessment of biofilm expression revealed that the mutation in the agrC resulted in a significant increase in biofilm formation by isolate HU-85c compared with isolate HU-85a (Figure 4). Reference strain S. aureus SA113, which is an agr-deficient derivative of S. aureus strain NCTC8325, produced the expected robust biofilm levels.
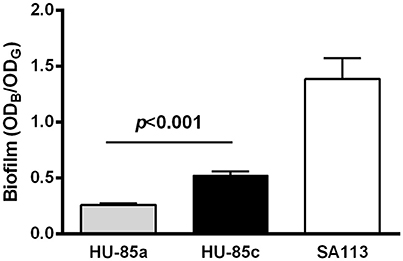
Figure 4. Biofilm formation by isolates HU-85a and HU-85c. S. aureus strain SA113 was included as reference. Each bar represents the arithmetic media ± SEM from 4 to 6 wells from 3 independent experiments. Biofilm formation values correspond to the optical density at 595 nm of crystal violet (ODB) measured relative to the final culture density of the bacterial growth (ODG) after 24 h incubation. The increase in biofilm formation by isolate HU-85c compared with HU-85a was significant (Student t-test).
Cytokine Production by Macrophages
Long exposure of S. aureus to the infected human bone microenvironment may select for additional phenotypic changes. We examined whether the two isolates differed in ability to induce pro-inflammatory cytokine expression. A significant reduction in TNF-α and IL-6 induction was observed in macrophages in primary culture stimulated with HU-85c (agr+, CP8), compared with cells stimulated with strain HU-85a (agr-deficient, NT) (Figure 5). These results indicate that the S. aureus isolate that persisted in the host with chronic osteomyelitis exhibited a reduction in its capacity to trigger an inflammatory response.
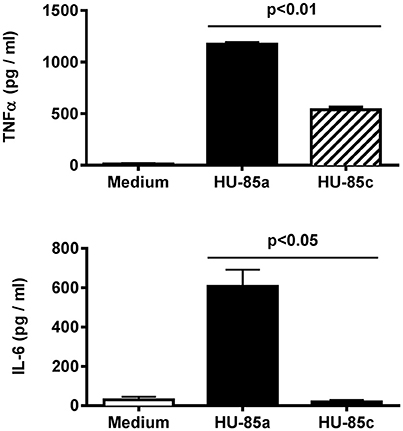
Figure 5. Production of TNF-α (upper) and IL-6 (bottom) by adherent mouse peritoneal macrophages stimulated with S. aureus HU-85a (agr+) or HU-85 (agr-deficient). Each bar represents cytokine concentrations obtained in representative experiments performed in triplicate. Comparisons of cytokine concentrations produced by stimulation with the wild-type compared with stimulation with the agr-deficient derivative were analyzed using the Student t-test.
Discussion
Genomic and phenotypic assessment of isolates HU-85a and HU-85c obtained 13 months apart from a patient with osteomyelitis revealed a frameshift mutation in the HU-85c agrC and a significant reduction in the RNAIII expression by HU-85c compared with HU-85a, which explain the loss of CP8, α- and β-hemolysin expression by HU-85c. Our finding agrees with those of Fischer et al., who showed in a study on S. aureus isolates from different sources that all non-encapsulated S. aureus isolated from human infections are agr deficient (Fischer et al., 2014). However, in S. aureus strains of the USA300 and USA500 backgrounds, which are not closely related to the ST188 strain studied here, the loss of capsular expression is due to mutations in the capsular genes rather than in a regulator of capsule (Boyle-Vavra et al., 2015). The S. aureus genome is capable of adapting to environmental changes, which can lead to significant phenotypic diversification among clinical isolates (Goerke and Wolz, 2004, 2010; Takeuchi et al., 2005). A high genetic variability of the agr locus has been described in S. aureus (Dufour et al., 2002) and attributed to balancing selection (Thomas et al., 2012). Traber and coworkers have shown that agr-defective variants arise by mutation during the course of infection (Traber et al., 2008). It was also reported that 9% of healthy human subjects are colonized with agr-defective strains (Shopsin et al., 2008) and that 22% of S. aureus isolates from chronically infected patients with atopic dermatitis had an agr mutant-like phenotype (Soong et al., 2015). In a recent study, it was shown that mutation in the agr locus occurs at a different rate in S. aureus of dissimilar clonal types (Recker et al., 2017). From the virulence standpoint, a mutation of the agr induces not only the lack of CP5(8) expression but also the lack of expression of secreted virulence factors (Novick, 2003), which resulted in reduced virulence in a model of septic arthritis (Blevins et al., 2002). Regulation of virulence factor expression by the agr system also seems to play an essential role in the adaptation of S. aureus to persistence at intracellular niches (Grundmeier et al., 2010). It has been shown that common single nucleotide polymorphisms that abrogate production of α-haemolysin and interfere with signaling of the agr reduced the virulence of contemporary hospital-associated MRSA and clinical MSSA included in S. aureus CC30. The authors concluded that loss of RNAIII expression, in addition to other defined mutations, made S. aureus less virulent to mice and provided the bacteria with suitable niche-adaptation (McGavin et al., 2012). In the present report we describe that loss of agr expression can occur in chronically infected patients with S. aureus osteomyelitis. Similarly, the reduction or total loss of agr functionality has been previously described in S. aureus from the cystic fibrosis patient lung (Goerke et al., 2000; Kahl et al., 2003). This association between persistence of S. aureus in the infected host and loss of agr expression supports the hypothesis that a non-functional agr may provide an advantage to S. aureus by defining a phenotype able to better adapt to persistence in the host.
Long-term persistence in the host leads to changes in the S. aureus genome. In a study on 1163 S. aureus genomes, Young and coworkers described one of these genome changes, i.e., the emergence of variants in protein altering genes responding to rsp, agr and host-derived antimicrobial peptides (Young et al., 2017). The authors further suggest that these changes emerge as a result of disease-associated, short-term, within-the-host selection pressures. Whereas in our study the factor responsible for S. aureus HU-85c positive selection is obscure it is interesting to note that S. aureus evolved from a capsulated into a non-encapsulated phenotype due to mutation in a regulator. Although diverse mechanisms account for loss of CP5(8) expression (Cocchiaro et al., 2006) so far only antibodies to CP5(8) have been pointed out as responsible for exerting selective pressure that lead to the emergence of stable NT S. aureus during experimental infection (Tuchscherr et al., 2008). In any event, it is speculated that adaptation of S. aureus for persistence at the infection site is multifactorial, as much as virulence, and that further research is needed to identify the specific host factors that exert selective pressure leading to the emergence of bacterial variants better adapted to persistence.
A major strategy displayed by S. aureus to support unfavorable conditions is to adopt the biofilm lifestyle, which plays a relevant role in chronic persistent infections, such as prosthetic implant associated infection (Archer et al., 2011). The importance of biofilm formation in S. aureus adaptation to the host to cause severe infection has been strengthened in a recent review article (Dastgheyb and Otto, 2015). In the present study it is shown that during infection mutation in the agrC led to significantly increased biofilm production. It is known that Agr negatively controls biofilm formation and that agr mutant strains of S. aureus produce thicker biofilms (Vuong et al., 2000). Since agr mutants are frequently isolated from biofilm-covered prosthetic implants, it can be assumed that robust biofilms confer an advantage to bacteria during chronic infection. Agr mutants, therefore, likely represent a dead end of infection, because they lack the capacity to disseminate within the body or establish infection in other hosts (Periasamy et al., 2012).
The progression of S. aureus osteomyelitis from acute into chronic may be benefited from evolution within the host from an aggressive phenotype, perhaps better adapted to cause bone damage at the start of the infection process into a less aggressive phenotype better adapted to persistence and responsible for chronic infection refractory to antibiotic treatment. The role of the evolutionary change in S. aureus during the progression of infection from acute into chronic has gained interest over the past few years. Expression of traits that permit adaptation of S. aureus to the host may initially be due to regulatory mechanisms but, in the long run, these traits may be fixed by mutation and may be considered endpoints in microevolution. One of these endpoints in S. aureus microevolution within the host is the stable SCV, which emerge during infection (Proctor et al., 2006; Tuchscherr et al., 2015). In vitro studies with host cells revealed that SCVs are phenotypes that hold low virulence, but are particularly adapted to the intracellular environment for long-term persistence (Tuchscherr et al., 2010). Interestingly, all SCV known so far are agr deficient thus failing to produce agr-regulated factors (Fraunholz and Sinha, 2012). Another endpoint of microevolution within the host is the loss of CP5(8) expression (Lattar et al., 2009). The loss of short sequence repeats (SSRs) in the protein A polymorphic region Xr is yet another endpoint, which results in the reduction of the S. aureus ability to trigger inflammation. In addition, the loss of SSRs was significantly higher in S. aureus persisting at the infection site in patients with chronic osteomyelitis or cystic fibrosis compared with patients with acute infection (Garofalo et al., 2012). In the present study, persistence of S. aureus in the bone and decreased ability to cause tissue damage in a rat model of osteomyelitis was associated with reduced capacity of S. aureus to cause inflammation, in this case through decreased triggering of proinflammatory cytokine release by macrophages. Therefore, the loss of RNAIII expression due to an agrC mutation and the reduction in virulence of the resulting variant makes functional impairment of agr another endpoint in microevolution within the host.
Conclusion
Our study demonstrates that S. aureus was able to adapt to the bone during chronic infection. Stable agr mutants arising during human chronic osteomyelitis were less able to trigger inflammation, and to cause bone damage as ascertained in the rat model of osteomyelitis. Lack of agr-dependent factors turned S. aureus less virulent and therefore it is suggested that mutations that alter the agr functionality seem to permit better adaptation of S. aureus to infection niche, making the reduction of agr functionality yet another endpoint in S. aureus microevolution within the chronically infected host.
Author Contributions
FB, MG, and DS, conceived and designed the experiments; MN and SL, performed experiments involving the osteomyelitis rat model and data analysis; CS, DR, and DS, performed sequencing experiments and data analysis; LA and CS, performed qRT PCR experiments; MG and CG, performed cytokine experiments and data analysis; DS and CS, wrote the manuscript; FB, DR, and MG, critically revised the manuscript; DS, MG, DR, and FB, procured funding; All authors read and approved the final manuscript.
Funding
This study was partially supported by grants from the Agencia Nacional de Promoción de la Ciencia y la Tecnología (ANPCYT), Argentina, PICT 2013-00941 to DS, PICT 2013-1233 to MG; CONICET (PIP 11220150100310) to FB; Secretaría de Ciencia y Técnica, Universidad de Buenos Aires (UBACyT), Argentina, #20020100100347 to DS; and NIH grant R01-GM080602 to DR.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
The authors thank Dr. Jean C. Lee, Division of Infectious Diseases, Department of Medicine, Brigham and Women's Hospital and Harvard Medical School, Boston, MA, USA, for providing S. aureus strains Reynolds (CP5) and its isogenic derivatives Reynolds (CP8) and Reynolds (NT). The authors also thank Lorena Medina for her dedicated technical assistance.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fcimb.2018.00018/full#supplementary-material
Supplementary Figure 1. CFU/tibia (upper) and OI (middle) from rats infected with isogenic Reynolds CP5, Reynolds CP8 and Reynolds NT. Bacterial loads and OI were measured 14 weeks after intratibial challenge. The sample size was n = 14 (Reynolds CP5), n = 12 (Reynolds CP8), and n = 11 (Reynolds NT) in both panels. The scattergram bars in the upper and middle panels represent the medians. Differences among bars were not significant (Kruskal-Wallis test). The lower panel shows the correlation of the CFU/bone counts and the OI of all the rats included in the experiments shown in the upper and middle panels. The CFU/bone counts correlated with the OI, indicating the good quality of the results.
References
Archer, N. K., Mazaitis, M. J., Costerton, J. W., Leid, J. G., Powers, M. E., and Shirtliff, M. E. (2011). Staphylococcus aureus biofilms: properties, regulation and roles in human disease. Virulence 2, 445–459. doi: 10.4161/viru.2.5.17724
Bankevich, A., Nurk, S., Antipov, D., Gurevich, A. A., Dvorkin, M., Kulikov, A. S., et al. (2012). SPAdes: a new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 19, 455–477. doi: 10.1089/cmb.2012.0021
Blevins, J. S., Beenken, K. E., Elasri, M. O., Hurlburt, B. K., and Smeltzer, M. S. (2002). Strain-dependent differences in the regulatory roles of sarA and Agr in Staphylococcus aureus. Infect. Immun. 70, 470–480. doi: 10.1128/IAI.70.2.470-480.2002
Bolger, A. M., Lohse, M., and Usadel, B. (2014). Trimmomatic: a flexible trimmer for Illumina sequence data. Bioinformatics 30, 2114–2120. doi: 10.1093/bioinformatics/btu170
Boyle-Vavra, S., Li, X., Alam, M. T., Read, T. D., Sieth, J., Cywes-Bentley, C., et al. (2015). USA300 and USA500 clonal lineages of Staphylococcus aureus do not produce a capsular polysaccharide due to conserved mutations in the cap5 locus. mBio 6:e02585-14. doi: 10.1128/mBio.02585-14
Cocchiaro, J. L., Gomez, M. I., Risley, A., Solinga, R., Sordelli, D. O., and Lee, J. C. (2006). Molecular characterization of the capsule locus from non-typeable Staphylococcus aureus. Mol. Microbiol. 59, 948–960. doi: 10.1111/j.1365-2958.2005.04978.x
Darling, A. E., Mau, B., and Perna, N. T. (2010). Progressivemauve: multiple genome alignment with gene gain, loss and rearrangement. PLoS ONE 5:e11147. doi: 10.1371/journal.pone.0011147
Das, S., Lindemann, C., Young, B. C., Muller, J., Österreich, B., Ternette, N., et al. (2016). Natural mutations in a Staphylococcus aureus virulence regulator attenuate cytotoxicity but permit bacteremia and abscess formation. Proc. Natl. Acad. Sci. U.S.A. 113, E3101–E3110. doi: 10.1073/pnas.1520255113
Dastgheyb, S. S., and Otto, M. (2015). Staphylococcal adaptation to diverse physiologic niches: an overview of transcriptomic and phenotypic changes in different biological environments. Future Microbiol. 10, 1981–1995. doi: 10.2217/fmb.15.116
Dotto, C., Serrat, A. L., Cattelan, N., Barbagelata, M. S., Yantorno, O. M., Sordelli, D. O., et al. (2017). The active component of aspirin, salicylic acid, promotes Staphylococcus aureus biofilm formation in a PIA-dependent manner. Front. Microbiol. 8:4. doi: 10.3389/fmicb.2017.00004
Dufour, P., Jarraud, S., Vandenesch, F., Greenland, T., Novick, R. P., Bes, M., et al. (2002). High genetic variability of the agr locus in Staphylococcus species. J. Bacteriol. 184, 1180–1186. doi: 10.1128/jb.184.4.1180-1186.2002
Fischer, J., Lee, J. C., Peters, G., and Kahl, B. C. (2014). Acapsular clinical Staphylococcus aureus isolates lack agr function. Clin. Microbiol. Infect. 20, O414–O417. doi: 10.1111/1469-0691.12429
Fraunholz, M., and Sinha, B. (2012). Intracellular Staphylococcus aureus: live-in and let die. Front. Cell. Infect. Microbiol. 2:43. doi: 10.3389/fcimb.2012.00043
Garofalo, A., Giai, C., Lattar, S., Gardella, N., Mollerach, M., Kahl, B. C., et al. (2012). The length of the Staphylococcus aureus protein a polymorphic region regulates inflammation: impact on acute and chronic infection. J. Infect. Dis. 206, 81–90. doi: 10.1093/infdis/jis311
Giai, C., Gonzalez, C. D., Sabbione, F., Garofalo, A., Ojeda, D., Sordelli, D. O., et al. (2016). Staphylococcus aureus induces shedding of IL-1RII in monocytes and neutrophils. J. Innate Immun. 8, 284–298. doi: 10.1159/000443663
Goerke, C., Campana, S., Bayer, M. G., Döring, G., Botzenhart, K., and Wolz, C. (2000). Direct quantitative transcript analysis of the agr regulon of Staphylococcus aureus during human infection in comparison to the expression profile in vitro. Infect. Immun. 68, 1304–1311. doi: 10.1128/IAI.68.3.1304-1311.2000
Goerke, C., and Wolz, C. (2004). Regulatory and genomic plasticity of Staphylococcus aureus during persistent colonization and infection. Int. J. Med. Microbiol. 294, 195–202. doi: 10.1016/j.ijmm.2004.06.013
Goerke, C., and Wolz, C. (2010). Adaptation of Staphylococcus aureus to the cystic fibrosis lung. Int. J. Med. Microbiol. 300, 520–525. doi: 10.1016/j.ijmm.2010.08.003
Grundmeier, M., Tuchscherr, L., Brück, M., Viemann, D., Roth, J., Willscher, E., et al. (2010). Staphylococcal strains vary greatly in their ability to induce an inflammatory response in endothelial cells. J. Infect. Dis. 201, 871–880. doi: 10.1086/651023
Hawkins, P., Morton, D. B., Burman, O., Dennison, N., Honess, P., Jennings, M., et al. (2011). A guide to defining and implementing protocols for the welfare assessment of laboratory animals: eleventh report of the BVAAWF/FRAME/RSPCA/UFAW joint working group on refinement. Lab. Anim. 45, 1–13. doi: 10.1258/la.2010.010031
Howden, B. P., Smith, D. J., Mansell, A., Johnson, P. D., Ward, P. B., Stinear, T. P., et al. (2008). Different bacterial gene expression patterns and attenuated host immune responses are associated with the evolution of low-level vancomycin resistance during persistent methicillin-resistant Staphylococcus aureus bacteraemia. BMC Microbiol. 8:39. doi: 10.1186/1471-2180-8-39
Iordanescu, S., and Surdeanu, M. (1976). Two restriction and modification systems in Staphylococcus aureus NCTC8325. J. Gen. Microbiol. 96, 277–281. doi: 10.1099/00221287-96-2-277
Kahl, B. C., Becker, K., Friedrich, A. W., Clasen, J., Sinha, B., Von Eiff, C., et al. (2003). Agr-dependent bacterial interference has no impact on long-term colonization of Staphylococcus aureus during persistent airway infection of cystic fibrosis patients. J. Clin. Microbiol. 41, 5199–5201. doi: 10.1128/JCM.41.11.5199-5201.2003
Lattar, S. M., Llana, M. N., Denoël, P., Germain, S., Buzzola, F. R., Lee, J. C., et al. (2014). Protein antigens increase the protective efficacy of a capsule-based vaccine against Staphylococcus aureus in a rat model of osteomyelitis. Infect. Immun. 82, 83–91. doi: 10.1128/IAI.01050-13
Lattar, S. M., Tuchscherr, L. P. N., Caccuri, R. L., Centrón, D., Becker, K., Alonso, C. A., et al. (2009). Capsule expression and genotypic differences among Staphylococcus aureus isolates from patients with chronic or acute osteomyelitis. Infect. Immun. 77, 1968–1975. doi: 10.1128/IAI.01214-08
Lattar, S. M., Tuchscherr, L. P. N., Centrón, D., Becker, K., Predari, S. C., Buzzola, F. R., et al. (2012). Molecular fingerprinting of Staphylococcus aureus isolated from patients with osteomyelitis in Argentina and clonal distribution of the cap5(8) genes and of other selected virulence genes. Eur. J. Clin. Microbiol. Infect. Dis. 31, 2559–2566. doi: 10.1007/s10096-012-1596-8
Lee, J. C., Liu, M.-J., Parsonnet, J., and Arbeit, R. D. (1990). Expression of type 8 capsular polysaccharide and production of toxic shock syndrome toxin 1 are associated among vaginal isolates of Staphylococcus aureus. J. Clin. Microbiol. 28, 2612–2615.
Livak, K. J., and Schmittgen, T. D. (2001). Analysis of relative gene expression data using Real-time quantitative PCR and the 2–ΔΔCT Method. Methods 25, 402–408. doi: 10.1006/meth.2001.1262
Martineau, F., Picard, F. J., Roy, P. H., Ouellette, M., and Bergeron, M. G. (1998). Species-specific and ubiquitous-DNA-based assays for rapid identification of Staphylococcus aureus. J. Clin. Microbiol. 36, 618–623.
McGavin, M. J., Arsic, B., and Nickerson, N. N. (2012). Evolutionary blueprint for host- and niche-adaptation in Staphylococcus aureus clonal complex CC30. Front. Cell. Infect. Microbiol. 2:48. doi: 10.3389/fcimb.2012.00048
Novick, R. P. (2003). Autoinduction and signal transduction in the regulation of Staphylococcal virulence. Mol. Microbiol. 48, 1429–1449. doi: 10.1046/j.1365-2958.2003.03526.x
Periasamy, S., Joo, H. S., Duong, A. C., Bach, T. H. L., Tan, V. Y., Chatterjee, S. S., et al. (2012). How Staphylococcus aureus biofilms develop their characteristic structure. Proc. Natl. Acad. Sci. U.S.A. 109, 1281–1286. doi: 10.1073/pnas.1115006109
Pitcher, D. G., Saunders, N. A., and Owen, R. J. (1989). Rapid extraction of bacterial genomic DNA with guanidium thiocyanate. Lett. Appl. Microbiol. 8, 151–156. doi: 10.1111/j.1472-765X.1989.tb00262.x
Prestinaci, F., Pezzotti, P., and Pantosti, A. (2015). Antimicrobial resistance: a global multifaceted phenomenon. Pathog. Glob. Health 109, 309–318. doi: 10.1179/2047773215Y.0000000030
Proctor, R. A., von Eiff, C., Kahl, B. C., Becker, K., McNamara, P., Herrmann, M., et al. (2006). Small colony variants: a pathogenic form of bacteria that facilitates persistent and recurrent infections. Nat. Rev. Microbiol. 4, 295–305. doi: 10.1038/nrmicro1384
Recker, M., Laabei, M., Toleman, M. S., Reuter, S., Saunderson, R. B., Blane, B., et al. (2017). Clonal differences in Staphylococcus aureus bacteraemia-associated mortality. Nat. Microbiol. 2, 1381–1388. doi: 10.1038/s41564-017-0001-x
Seemann, T. (2014). Prokka: rapid prokaryotic genome annotation. Bioinformatics 30, 2068–2069. doi: 10.1093/bioinformatics/btu153
Seemann, T. (2015). Snippy: Fast Bacterial Variant Calling from NGS Reads. Available online at: https://github.com/tseemann/snippy
Shopsin, B., Drlica-Wagner, A., Mathema, B., Adhikari, R. P., Kreiswirth, B. N., and Novick, R. P. (2008). Prevalence of Agr dysfunction among colonizing Staphylococcus aureus strains. J. Infect. Dis. 198, 1171–1174. doi: 10.1086/592051
Soong, G., Paulino, F., Wachtel, S., Parker, D., Wickersham, M., Zhang, D., et al. (2015). Methicillin-resistant Staphylococcus aureus adaptation to human. MBio 6, 1–14. doi: 10.1128/mBio.00289-15
Takeuchi, F., Watanabe, S., Baba, T., Yuzawa, H., Ito, T., Morimoto, Y., et al. (2005). Whole-genome sequencing of Staphylococcus haemolyticus uncovers the extreme plasticity of its genome and the evolution of human-colonizing staphylococcal species. J. Bacteriol. 187, 7292–7308. doi: 10.1128/JB.187.21.7292-7308.2005
Thomas, J. C., Godfrey, P. A., Feldgarden, M., and Robinson, D. A. (2012). Candidate targets of balancing selection in the genome of Staphylococcus aureus. Mol. Biol. Evol. 29, 1175–1186. doi: 10.1093/molbev/msr286
Tong, S. Y., Davis, J. S., Eichenberger, E., Holland, T. L., and Fowler, V. G. (2015). Staphylococcus aureus infections: epidemiology, pathophysiology, clinical manifestations, and management. Clin. Microbiol. Rev. 28, 603–661. doi: 10.1128/CMR.00134-14
Traber, K. E., Lee, E., Benson, S., Corrigan, R., Cantera, M., Shopsin, B., et al. (2008). Agr function in clinical Staphylococcus aureus isolates. Microbiology 154, 2265–2274. doi: 10.1099/mic.0.2007/011874-0
Tuchscherr, L., Bischoff, M., Lattar, S. M., Noto Llana, M., Pförtner, H., Niemann, S., et al. (2015). Sigma factor sigB is crucial to mediate Staphylococcus aureus adaptation during chronic infections. PLoS Pathog. 11:e1004870. doi: 10.1371/journal.ppat.1004870
Tuchscherr, L., Heitmann, V., Hussain, M., Viemann, D., Roth, J., von Eiff, C., et al. (2010). Staphylococcus aureus small-colony variants are adapted phenotypes for intracellular persistence. J. Infect. Dis. 202, 1031–1040. doi: 10.1086/656047
Tuchscherr, L. P., Buzzola, F. R., Alvarez, L. P., Lee, J. C., and Sordelli, D. O. (2008). Antibodies to capsular polysaccharide and clumping factor a prevent mastitis and the emergence of unencapsulated and small-colony variants of Staphylococcus aureus in mice. Infect. Immun. 76, 5738–5744. doi: 10.1128/IAI.00874-08
Vaudaux, P., Francois, P., Bisognano, C., Kelley, W. L., Lew, D. P., Schrenzel, J., et al. (2002). Increased expression of clumping factor and fibronectin-binding proteins by hemB mutants of Staphylococcus aureus expressing small colony variant phenotypes. Infect. Immun. 70, 5428–5437. doi: 10.1128/IAI.70.10.5428-5437.2002
Vuong, C., Saenz, H. L., Götz, F., and Otto, M. (2000). Impact of the Agr quorum-sensing system on adherence to polystyrene in Staphylococcus aureus. J. Infect. Dis. 182, 1688–1693. doi: 10.1086/317606
Watts, A., Ke, D., Wang, Q., Pillay, A., Lee, J. C., and Nicholson-weller, A. (2005). Staphylococcus aureus strains that express serotype 5 or serotype 8 capsular polysaccharides differ in virulence. Infect Immun. 73, 3502–3511. doi: 10.1128/IAI.73.6.3502-3511.2005
Keywords: Staphylococcus aureus, adaptation, infection, agr, osteomyelitis, chronic, biofilm, capsular polysaccharide
Citation: Suligoy CM, Lattar SM, Noto Llana M, González CD, Alvarez LP, Robinson DA, Gómez MI, Buzzola FR and Sordelli DO (2018) Mutation of Agr Is Associated with the Adaptation of Staphylococcus aureus to the Host during Chronic Osteomyelitis. Front. Cell. Infect. Microbiol. 8:18. doi: 10.3389/fcimb.2018.00018
Received: 26 November 2017; Accepted: 12 January 2018;
Published: 02 February 2018.
Edited by:
Martin John McGavin, University of Western Ontario, CanadaReviewed by:
Mark Smeltzer, University of Arkansas for Medical Sciences, United StatesYan Q. Xiong, David Geffen School of Medicine at UCLA, United States
Copyright © 2018 Suligoy, Lattar, Noto Llana, González, Alvarez, Robinson, Gómez, Buzzola and Sordelli. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Daniel O. Sordelli, ZHNvcmRlbGxpQHlhaG9vLmNvbQ==