 Ascel Samba-Louaka1
Ascel Samba-Louaka1 Etienne Robino1
Etienne Robino1 Thierry Cochard2Maxime Branger2
Thierry Cochard2Maxime Branger2 Vincent Delafont1Willy Aucher1Wilfrid Wambeke2
Vincent Delafont1Willy Aucher1Wilfrid Wambeke2 John P. Bannantine3
John P. Bannantine3 Franck Biet2*
Franck Biet2* Yann Héchard1*
Yann Héchard1*- 1Université de Poitiers, Laboratoire Ecologie et Biologie des Interactions, UMR Centre National de la Recherche Scientifique 7267, Equipe Microbiologie de l'Eau, Poitiers, France
- 2Institut National de la Recherche Agronomique, Université de Tours, UMR1282, Infectiologie et Santé Publique, Nouzilly, France
- 3National Animal Disease Center, Agricultural Research Service, United States Department of Agriculture, Ames, IA, United States
Mycobacterium avium subsp. paratuberculosis is responsible for paratuberculosis in animals. This disease, leading to an inflammation of the gastrointestinal tract, has a high impact on animal health and an important economic burden. The environmental life cycle of M. avium subsp. paratuberculosis is poorly understood and several studies suggest that free-living amoebae (FLA) might be a potential environmental host. FLA are protozoa found in water and soil that are described as reservoirs of pathogenic and non-pathogenic bacteria in the environment. Indeed, bacteria able to survive within these amoebae would survive phagocytosis from immune cells. In this study, we assessed the in vitro interactions between several strains of M. avium subsp. paratuberculosis and Acanthamoeba castellanii. The results indicate that the bacteria were able to grow within the amoeba and that they can survive for several days within their host. To explore the presence of M. avium subsp. paratuberculosis in environmental amoebae, we sampled water from farms positive for paratuberculosis. A M. avium subsp. paratuberculosis strain was detected within an environmental amoeba identified as related to the poorly described Rosculus genus. The bacterial strain was genotyped, showing that it was similar to previous infectious strains isolated from cattle. In conclusion, we described that various M. avium subsp. paratuberculosis strains were able to grow within amoebae and that these bacteria could be found on farm within amoebae isolated from the cattle environment. It validates that infected amoebae might be a reservoir and vector for the transmission of M. avium subsp. paratuberculosis.
Introduction
Mycobacterium avium subsp. paratuberculosis (Map), the etiologic agent of paratuberculosis also called Johne's disease, induces extensive inflammation in the gastrointestinal tract of ruminants. This inflammation prevents absorption of nutrients, which leads to a chronic and progressive weight loss despite eating habits. The hallmark of advance stage disease in cattle is copious diarrhea, appearance of ribs and other skeletal bones. Paratuberculosis is responsible for economic losses up to $1.5 billion annually to the dairy industries on the five continents (Ott et al., 1999; Sweeney et al., 2012). The management of this epidemic, is particularly difficult because efficient prophylactic tools are lacking. There is no treatment and the vaccine available is not very effective and not widely used because it compromises the diagnosis of bovine tuberculosis. Serological diagnostic tools lack specificity and are not adapted to early diagnosis of an infection, although this is beginning to change with the discovery of new antigens (Li et al., 2017). Another factor complicating the management of this epidemic involves the shedding of the bacteria in the environment where it is able to survive. Map is a genetically homogenous subspecies of M. avium, especially among bovine, human and wildlife isolates. However, two prominent lineages of Map have emerged following molecular strain typing and comparative genomic analysis—ovine (Map-S) and bovine (Map-C) strains (Biet et al., 2012). Initially, the Map-S and Map-C type strains were distinguished based on their molecular fingerprints using IS1311 polymorphism, MLSSR typing and hsp65 sequencing. However, beyond genetic typing, there are observed phenotypic differences including differences in their pigmentation, growth characteristics and lipopeptide structures in their cell walls (Bannantine et al., 2017). The environmental life cycle of Map is still poorly understood and at least one recent publication suggests that free-living amoebae (FLA) might be a potential environmental host (Salgado et al., 2015).
Free-living amoebae (FLA) are protozoa found in the same environmental niches as Map, including water and soil (Rodríguez-Zaragoza, 1994). FLA mainly feed on bacteria in the environment and digest them by phagocytosis. They thus play, along with other grazing protists, a key role in shaping the bacterial community composition in the environment (Jürgens and Matz, 2002). However, it has been repeatedly shown over the last decades that some bacteria may resist this digestion and persist or even grow within amoebae (Greub and Raoult, 2004). Importantly, these resistant bacteria are generally also more resistant to phagocytic immune cells, such as macrophages. This is well documented for Legionella pneumophila and for various Mycobacterium species in interaction with Acanthamoeba castellanii. The latter is the most common species in water and the best known FLA. As a consequence, FLA are considered as a training ground for pathogenic bacteria, including some Mycobacterium species (Molmeret et al., 2005; Salah et al., 2009). The fate of internalized mycobacteria can drastically vary according to the species, ranging from digestion (e.g., M. bovis BCG) and survival without replication (e.g., M. tuberculosis) to intra amoebal multiplication (e.g., M. abscessus or M. chelonae) (Drancourt, 2014). However, as described in this paper, most of the interactions were studied in vitro and only few studies have reported mycobacteria detected inside amoebae isolated from the environment (White et al., 2010; Amissah et al., 2014). Our group has previously described such interaction in drinking water (Delafont et al., 2014). Importantly, intra-amoebal growth of Mycobacterium would lead to improve its virulence (Cirillo et al., 1997).
The interaction between Map and amoebae has been poorly described. It was first reported in 2006 through a study showing in vitro that a strain of Map was ingested by Acanthamoeba and resisted digestion for at least 24 days (Whan et al., 2006). Four years later a short communication described the co-occurrence of Map and FLA in the soil (White et al., 2010). Amoebae and Map were found in the same soil samples and an amoeba strain was isolated harboring an intracellular acid-fast stained bacterium. Mura et al. (2006) described in vitro that Map persisted for up to four years in presence of Acanthamoeba (Mura et al., 2006). Recently, Map were found within amoebae isolated from soil after application of cattle slurry spiked with Map (Salgado et al., 2015). However, these studies used a very limited range of strains and no study has demonstrated clearly the presence of Map in environmental FLA.
Our study was aimed to assess the infectious potential of several Map strains including the two main genetic lineages of Map, C and S types toward A. castellanii. We also investigated whether Map-infected FLA might be found in the environment surrounding infected cattle.
Materials and Methods
Cultivation of Microorganisms
Bacterial strains used in this study are listed in Table 1. Mycobacterial strains were grown at 37°C in Sauton medium or Middlebrook 7H9 broth (Difco Laboratories, Detroit, MI) with 0.2% glycerol and albumin-dextrose-catalase enrichment medium (ADC, Becton Dickinson, Le Pont de claix, France). M. avium subsp. paratuberculosis cultures were supplemented with 2 mg l−1 mycobactin J (Allied Monitor). Bacteria were harvested at mid-log phase and kept frozen (−80°C) in aliquots until use. Recombinant M. avium subsp. paratuberculosis, strain K-10 expressing the Green fluorescent protein (GFP) - and luciferase described in Lefrançois et al. (2011) was grown in the Middlebrook 7H10 medium supplemented with ADC, 0.2 mg ml-1 mycobactin and 50 mg ml−1 of hygromycin. Escherichia coli strain K12 was grown in LB liquid medium (Lysogeny Broth: 10 g/L NaCl, 5 g/L yeast extract, 10 g/L tryptone), and incubated for 24 h at 37°C, under agitation (180 rpm).
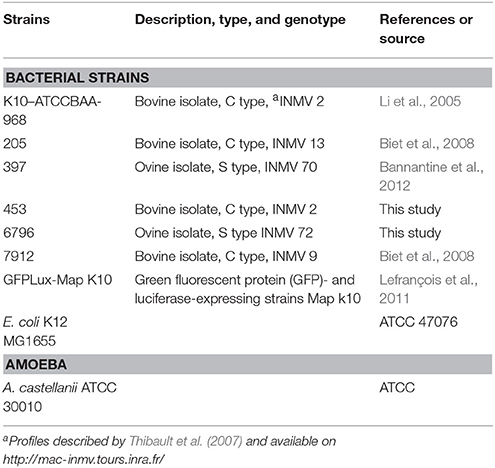
Table 1. Bacterial and amoebal strains.
The free-living amoeba A. castellanii ATCC 30010 was grown in PYG liquid medium (ATCC 712; 20 g/L proteose peptone, 1 g/L yeast extract, 1 g/L sodium citrate, 0.1 M D-glucose, 0.4 mM CaCl2, 4 mM MgSO4, 2.5 mM Na2HPO4, 2.5 mM KH2PO4, 50 μM Fe(NH4)2(SO4)2, pH 6.5) in 25 or 75 cm2 flask. A. castellanii cultures were incubated at 30°C for 3–4 days, until a cell confluency of 80–90% was reached.
Isolation of Environmental Amoebae
Environmental water samples were collected in March 2016 from two farms located in “Indre et Loire” and reported to be contaminated with Map since more than 5 years. Two 1-L samples were taken for each herd from the drinking troughs. The samples were filtered through a 5 μm nitrocellulose membrane (N3771-100EA, Sigma-Aldrich). The membranes were placed onto NNA plates (Non Nutrient Agar, 15 g/L) seeded with live E. coli strain K12. The plates were incubated at 30°C and examined daily under phase-contrast microscopy. The presence of amoebal cells is characterized by the formation of a migration front, representing the amoebal movement upon agar plates, seeking for food. Amoebae were recovered by scrapping the agar plate at the migration front for collection or DNA extraction.
A. castellanii Infection by Different Genotypes of Map
A. castellanii cells from fully grown flasks were rinsed once using PAS buffer [Page's Amoeba Saline; 1 g/L sodium citrate, 0.4 mM CaCl2, 4 mM MgSO4, 2.5 mM Na2HPO4, 2.5 mM KH2PO4, 50 μM Fe(NH4)2(SO4)2, pH 6.5], and resuspended in a mixture of PAS/PYG (1:1) supplemented with 0.005% triton X-100, to avoid mycobacteria clumping in the medium. Trophozoites cells were harvested and detached by vigorously tapping the flask. The cell suspension was adjusted to a concentration of 1.25 × 105 cells/mL, and 2 mL of the suspension distributed in each well of a 12 well plates. Mycobacteria cells were dispersed by 3 passages through a 18 Gauge needle, followed by a centrifugation step at 200 g for 5 min. The suspensions were subsequently quantified by optical density (OD) at 600 nm, using an equivalence of 6 × 108 mycobacteria per mL for 1 OD unit. Mycobacteria suspensions were used to infect A. castellanii at an MOI (multiplicity of infection) of 10, i.e., 10 bacteria per amoeba cell. The infected cultures were then centrifuged at 500 g for 10 min to facilitate the infection, and incubated for 1 h at 30°C. The supernatant was then discarded and directly replaced with a fresh mixture of PAS/PYG (1:1), 0.005% triton X-100, supplemented with amikacin at 10 μM. Timepoints at 1, 24, 48, and 72 h post infection at 30°C were selected to estimate presence or growth of Mycobacteria by qPCR. To estimate the growth of A. castellanii, cells were harvested at different time points and counted using plastic counting slides FastRead 102 (Biosigma).
The localization of bacteria within amoeba was visualized using a recombinant rGFP-Map strain K-10 (Table 1) previously described (Lefrançois et al., 2011). Infected cultures were analyzed by fluorescence microscopy (BX41, Olympus).
Total Genomic DNA Extraction
Cells from infection experiments as well as from environmental amoebae isolated as described above, were used for DNA extraction. Amoeba cells (4 × 104 - 2 × 106) and associated mycobacteria were harvested and resuspended in 500 μL of PAS buffer. Cells were lysed by bead-beating in tubes containing 500 mg of small diameter glass beads (100 μm) and 4 glass beads of 2 mm diameter (Sigma) using Fastprep apparatus for 30 s (speed 5 m/s). The suspension was then processed for DNA extraction using NucleoSpin Microbial DNA (Macherey-Nagel), following manufacturer recommendations for bacterial DNA extraction.
Detection and Quantification of Mycobacteria by Quantitative PCR (qPCR)
The presence of bacterial genomic DNA was assessed by qPCR performed in a final volume of 10 μL, including 2 μL of extracted DNA, 2 μL of 5x Master Mix LightCycler® FastStart DNA Masterplus SYBR Green I (Roche), a set of primer pairs (Table 2), at a concentration of 0.5 μM each, and water in sufficient quantity for 10 μL. Experiments were carried out using a LightCycler 1.5 thermocycler (Roche), consisting in an initial denaturation step at 95°C for 10 min, 45 cycles of denaturation at 95°C for 10 s, annealing for 10 s at a temperature according to the primer pairs used, and elongation at 72°C for 20 s. Fusion curves were collected for each experiment. Quantification of DNA was performed by calculating fold changes using the 2−ΔCt method. ΔCt corresponds to the difference in cycle threshold (Ct) between a specific time point and the condition at 1 h.
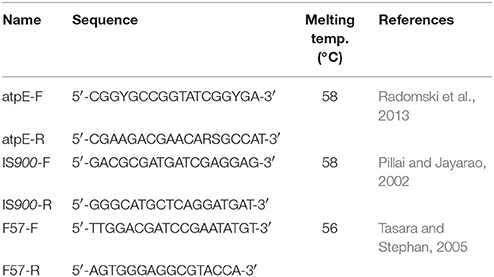
Table 2. Oligonucleotide primers.
Multiple-Locus Variable Number Tandem Repeat Analysis (MLVA) Typing of Environmental Mycobacteria, Using MIRU-VNTR Markers
The method has been described previously (Thibault et al., 2007). The PCR mixture was composed as follows using the Go Taq Flexi DNA polymerase (Promega). Five microliters from DNA solution were added to a final volume of 25 μL containing 0.1 μL of Go Taq Flexi DNA polymerase (5 U/μL), 5 μl of betaine (Sigma), or 1 μl of dimethyl sulfoxide (Sigma); 0.2 mM (each) dATP, dCTP, dGTP, and dTTP (Promega); 5 μL of 5 × PCR buffer supplied by the manufacturer; 1 μM of primers see (Thibault et al., 2007); and 1.5 mM of MgCl2. The reactions were carried out using a TC-520 thermal cycler (Techne). PCR conditions were as follows: 1 cycle of 5 min at 94°C; 30 cycles of 30 s at 94°C, 30 s at 55, 58, 60, 64°C according to MIRU-VNTR searched, and 30 s at 72°C; 1 cycle of 7 min at 72°C. To detect differences in repeat numbers, the PCR products were analyzed by electrophoresis using 1.5% agarose gels (agarose electrophoresis grade; Invitrogen).
Amoebal Identification
The identification of the amoeba isolated from the water troughs was performed through sequencing of a 18S rRNA gene amplicon. From the total genomic DNA extract, a PCR was performed using the universal F566 and R1200 eukaryotic primers, targeting the 18S rRNA gene (Hadziavdic et al., 2014). The amplicon was sequenced and the sequence compared to the nucleotide nr database using BLASTn.
Evaluation of Phagosomal Acidification
Mycobacteria suspension prepared as described previously were pre-labeled using pHrodoTM Red succinimidyl ester (ThermoFischer Scientific) following manufacturer recommendations, but excluding the methanol washing step. Briefly, bacterial suspensions were incubated for 1 h in 0.1 M sodium bicarbonate buffer containing 20 μM pHrodoTM Red succinimidyl ester, in the dark. Bacteria were then pelleted by centrifugation at 12,000 g for 5 min, and washed twice in PAS buffer supplemented with 0.005% triton X-100. A. castellanii monolayers were infected by labeled mycobacteria at a MOI of 10. After 4 h of incubation, trophozoites were detached, fixed with 2% paraformaldehyde for 10 min in the dark. Cells were pelleted by centrifugation at 800 g for 10 min, and resuspended in a 30 μL of SlowFade Diamonds antifade mountant (ThermoFischer Scientific) with Hoechst at a concentration of 400 ng/mL. Samples were examined using an epifluorescence microscope (BX41, Olympus).
Statistics
Results were statistically analyzed using GraphPad Prism software. Multiple comparison was performed using Kruskal–Wallis analysis followed by a Dunn's post-test. Differences with p < 0.05 were considered statistically significant.
Results
Map Strains Grow within A. castellanii
In order to see whether Map strains would behave differently within amoeba, we investigated infection of A. castellanii by different genotype and genetic lineage of Map strains. A. castellanii were co-cultured with the reference strain (K10), two cattle strains (205, 7912), and two sheep strains (397, 6796). The fate of Map internalized by A. castellanii was determined by a qPCR based bacterial quantification assay, at 1, 24, 48, and 72 h post infection. We found that all Map persisted and even grew within A. castellanii (Figure 1A). There were no significant differences in growth rate between the strains (p = 0.4441) even though the K10 strain tended to be slightly slower. The fate of infected A. castellanii was also monitored throughout the infection, indicating that the host proliferation was not significantly modified (p = 0.8939) by the presence of Map in comparison with an uninfected A. castellanii culture (Figure 1B). Taken together, these results suggest that Map can successfully infect and grow within A. castellanii without any deleterious effect on its host.
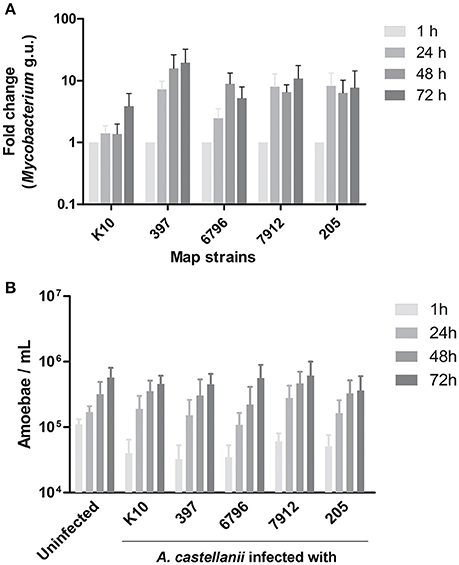
Figure 1. A. castellanii is permissive to both ovine (S) and bovine (C) Map. (A) A. castellanii were infected with ovine (397, 6796) and bovine (K10, 7912, 205) strains of Map at a MOI of 10. Presence of Mycobacterium spp. was quantified (Mycobacterium genome unit) through qPCR amplification of the atpE gene, 1, 24, 48, and 72 h post-infection. Results represents the mean (±SEM) of four independent experiments (p = 0.4441). Results are normalized on the condition 1 h. (B) Uninfected and infected A. castellanii were counted 1, 24, 48, and 72 h post-infection. Results represents the mean (±SEM) of four independent experiments (p = 0.8939).
Map Resides within Phagosomal Compartments
To localize Map within A. castellanii, amoebae were infected with a Map K10 strain expressing a green fluorescent protein (GFP). Microscopy analysis of infected A. castellanii showed colocalization between the Map K10-GFP and A. castellanii 7 days after infection (Figure 2A). This figure also suggests that the bacteria were found inside A. castellanii vesicles. Later observations allowed to visualize A. castellanii cysts but no GFP signal from the K10-GFP strain was detected, suggesting that in our conditions Map K10 was not present within the cysts. We have also assessed the acidification of Map-containing intracellular compartments in the first hours of infection. Before infection, the Map K10-GFP strain was stained with the pHrodo dye, which fluoresces in a pH-dependent manner. The results show that a fraction of intracellular Map was labeled in red, indicating that they were found within acidified vacuoles, likely phagosomes, while other Map were not labeled (Figure 2B). These observations suggest that Map are phagocytised by A. castellanii and resides, at least transiently, within acidified vacuoles.
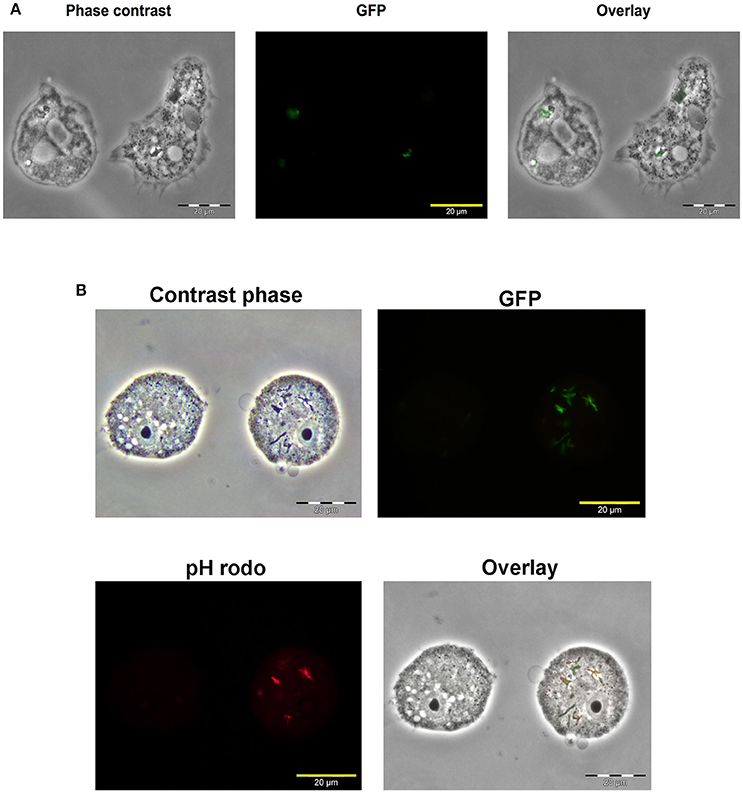
Figure 2. Map K10 persists within A. castellanii for up to 7 days. (A) A. castellanii were co-cultured with a K10-GFP map strain at a MOI of 10. Mycobacteria localization within amoebae trophozoites was assessed by epifluorescence microscopy 7 days after the initial infection. (B) A. castellanii were infected with K10-GFP labeled with a pHrodo dye at a MOI of 10. At 4 h post-infection, infected cells were analyzed through epifluorescence microscopy to reveal the Map K10 strain and acidification of bacteria through GFP and red signals.
Map Is Found within an Environmental Amoeba in Infected Farm
As no study had isolated environmental FLA infected by Map so far, we investigated the potential Map-amoeba association in water from the drinking troughs in herds naturally infected by Map. From the two farms enrolled in this study, only one sample was positive for the presence of cultivable amoebae. Total genomic DNA extracted from the amoebal culture was used to detect the presence of Map DNA using a Map-specific qPCR assay, targeting the IS900 and F57 genes (Table 3). The successful amplification of the specific genomic target highlighted the presence of Map in association with the isolated amoebal culture. The Map strains K10, 397, and 7912 were used as positive control and M. llatzerenze as negative control for all PCR-based assays (Table 3). We have previously established a database containing isolated and genotyped Map strains from feces of infected cattle (Holbert et al., 2015). Therefore, we genotyped the Map-positive DNA from the amoeba culture, using MLVA genotyping and compare it to our database. The result of the MLVA genotyping based on the eight classical loci MIRU VNTR (Thibault et al., 2007) gave a profile INMV 2 identical to that of the 453 strain isolated from feces of cattle (Table 4).

Table 3. Map-specific qPCR targeting the atpE, IS900 and F57 genes.
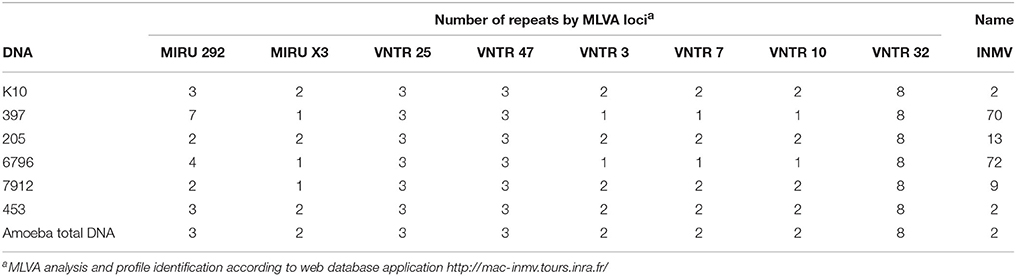
Table 4. MLVA genotyping of Map strains.
Furthermore, in order to identify the amoeba isolate, we tried to subculture the strain but no subculture was observed. Thus, we performed a PCR targeting a portion of 18S rRNA coding gene. Sequencing of purified amplicon (1265 nucleotides, GenBank MG859939), indicated that the most closely related sequence to the isolate was Rosculus ithacus, a cercozoan amoeba, although the sequence identity was moderate (78%). Therefore, this amoebal isolate is likely to represent a newly described genus among the sainouroid clade.
Discussion
A growing number of studies underlines the important role of amoebae in the carrying of pathogenic and non-pathogenic mycobacteria, as well as in providing an environmental source of mycobacterial infection. On a microbiological level, the question of on farm disease transmission is particularly relevant because of the characteristics of Map biology. Indeed the disease transmission happens through ingestion of Map then during the disease progression, animals excrete bacilli in their environment, ranging from a few and discontinuous bacterial excretions in asymptomatic phases to very high levels and continuously shedding at clinical stage of the disease (Magombedze et al., 2017). This means that even in the silent phases of the disease the animals can excrete Map in the environment and in particular in the water points of the herds. In this context, it was interesting to investigate whether Map can be hosted and survive in amoeba and if water points of infected herds could be a source of contamination of Map mediated by FLA. Some other pathogenic mycobacteria were able to survive in soil and this environment might be a source of infection (Ghodbane et al., 2014).
In this study, the five selected strains (C- and S-type) not only survived but also showed a growth trend within A. castellanii for at least 72 h. In previous studies the growth of Map was not so high in the first days post-infection. This difference could be due to the Acanthamoeba strains and the infection protocols that were used in each study (Mura et al., 2006; Whan et al., 2006). For example, the co-cultures were centrifuged to increase efficiency and better synchronize the infection process. These differences might also explain the higher levels of bacteria per amoeba we observed. Moreover, we demonstrated that Map can persist for at least 7 days within A. castellanii. Within Map there is little genetic variability nevertheless the evolutionary pattern deciphered by Turenne et al. (2006) and Alexander et al. (2009) showed that the Map sub-species evolved from a common ancestor M. avium subsp. hominissuis into two different genetic lineages associated with a host preference. We speak of a lineage of S-type for sheep and C-type for cattle. In this study the strains were selected for their difference in terms of genotype and to include representative S and C-type. Altogether these results suggest that amoeba may serve as a significant phagocytic model to further investigate comparable or distinct phenotypic traits obtained by each lineage S and C-type of Map.
We also observed that the growth of A. castellanii was not adversely affected by infection with any of the Map strains. This indicates that Map had no deleterious effects on amoebal viability. These results suggest that interactions between these two types of microorganisms could very likely happen in the environment. This stability could indeed favor Map resilience in the environment, providing a shelter against harsh conditions, as well as a potential substrate source for mycobacterial multiplication (Caire-Brändli et al., 2014; Barisch et al., 2015; Delafont et al., 2017).
Microscopy studies indicate that Map was localized within phagosomes and that some bacteria were found in acidified environment while others were not. This suggests that Map could block, to a certain extent, the normal acidification process of A. castellanii phagosomes. Inhibition of phagosomal acidification has been observed with other Mycobacterium species both in amoebae and macrophages (Kuehnel et al., 2001; Rohde et al., 2007; Delafont et al., 2017). Although much of the cell biology has been done on mycobacteria-macrophage interactions, further investigations would be required to better understand the fate of Map after its internalization in amoeba.
Amoeba-Map association in the environment has been poorly documented. To our knowledge, only one short communication has reported that amoeba isolated from soil were positive for Map-specific PCR but neither the Map nor the amoebae strains were characterized (White et al., 2010). In our study, two water troughs were selected for sampling, as those were providing the water source to distinct herds previously declared as infected by Map. We were able to recover cultivable FLA from one of the water troughs, confirming their presence in this environment. From the cultivated amoeba, the presence of Map DNA was detected using two specific primer sets by qPCR. This finding represents a strong argument in favor of interactions between amoebae and Map in environmental conditions. MLVA genotyping confirmed the identification of the species Map. The MLVA profile of the DNA was identical to that of an isolated strain from feces of a Map naturally infected cattle in the same farm (Holbert et al., 2015). These results not only confirm the presence of FLA harboring Map in the environment of animals on farm, but the identical genotypes recovered may also suggest a link between residence in amoeba and ability to infect cattle. Further investigations could be done to isolate the Map strains and improve the discrimination of genotype by combination of genotyping methods to ascertain the strains isolated from amoebae and animals are strictly identical. These results indicate that the strains might persist and circulate in the herd. They further suggest that fresh drinking water sources on farm may be important for healthy cows and that the problem of Johne's disease may be difficult to solve using herd management practices alone.
In addition, the identification of the amoebal isolate indicated that its closest sequence corresponds to R. ithacus. The latter is a coprophilic amoeba belonging to the sainouroid clade, as part of the Cercozoan radiation. Interestingly, protozoa from this clade were also identified in cow and sheep dung in a previous study (Bass et al., 2016). It is therefore not surprising to have identified such an amoeba from a drinking through providing cattle. However, we were not able to subculture this amoebal strain for some reason even after several try. It suggests that the culture method that was used is not fully efficient to subculture any FLA.
In conclusion, our study showed that various Map strains were able to persist and grow within A. castellanii, without observable deleterious effects on the host. A screening of environment surrounding Map-infected herds allowed to recover a free-living amoeba, representative of the sainouroid clade, from a drinking trough. This environmental amoeba harbored a Map strain that was genotyped. This study is bringing additional pieces of evidence reinforcing the role of amoebae in the persistence, and potential transmission, of pathogenic mycobacteria, including Map. This work should stimulate further studies focused on the characterization of environmental interactions between amoebae and pathogenic mycobacteria.
Author Contributions
All the authors have substantial contributions to the conception or design of the work; or the acquisition, analysis, or interpretation of data for the work; drafting the work or revising it critically for important intellectual content and final approval of the version to be published.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We would like to thank Manon Duforestel for her technical help and Laure Malherbe Duluc, GDS veterinarian responsible for the epidemiological surveillance of infected herds. YH laboratory is partly financed by the European Union and the region of Nouvelle Aquitaine through the “Habisan program” (CPER-FEDER).
References
Alexander, D. C., Turenne, C. Y., and Behr, M. A. (2009). Insertion and deletion events that define the pathogen Mycobacterium avium subsp. paratuberculosis. J. Bacteriol. 191, 1018–1025. doi: 10.1128/JB.01340-08
Amissah, N. A., Gryseels, S., Tobias, N. J., Ravadgar, B., Suzuki, M., Vandelannoote, K., et al. (2014). Investigating the role of free-living amoebae as a reservoir for Mycobacterium ulcerans. PLoS Negl. Trop. Dis. 8:e3148. doi: 10.1371/journal.pntd.0003148
Bannantine, J. P., Etienne, G., Laval, F., Stabel, J. R., Lemassu, A., Daffé, M., et al. (2017). Cell wall peptidolipids of Mycobacterium avium: from genetic prediction to exact structure of a nonribosomal peptide. Mol. Microbiol. 105, 525–539. doi: 10.1111/mmi.13717
Bannantine, J. P., Wu, C. W., Hsu, C., Zhou, S., Schwartz, D. C., Bayles, D. O., et al. (2012). Genome sequencing of ovine isolates of Mycobacterium avium subspecies paratuberculosis offers insights into host association. BMC Genomics 13:89. doi: 10.1186/1471-2164-13-89
Barisch, C., Paschke, P., Hagedorn, M., Maniak, M., and Soldati, T. (2015). Lipid droplet dynamics at early stages of Mycobacterium marinum infection in Dictyostelium. Cell. Microbiol. 17, 1332–1349. doi: 10.1111/cmi.12437
Bass, D., Silberman, J. D., Brown, M. W., Pearce, R. A., Tice, A. K., Jousset, A., et al. (2016). Coprophilic amoebae and flagellates, including Guttulinopsis, Rosculus and Helkesimastix, characterise a divergent and diverse rhizarian radiation and contribute to a large diversity of faecal-associated protists. Environ. Microbiol. 18, 1604–1619. doi: 10.1111/1462-2920.13235
Biet, F., Bay, S., Thibault, V. C., Euphrasie, D., Grayon, M., Ganneau, C., et al. (2008). Lipopentapeptide induces a strong host humoral response and distinguishes Mycobacterium avium subsp. paratuberculosis from M. avium subsp. avium. Vaccine 26, 257–268. doi: 10.1016/j.vaccine.2007.10.059
Biet, F., Sevilla, I. A., Cochard, T., Lefrançois, L. H., Garrido, J. M., Heron, I., et al. (2012). Inter- and intra-subtype genotypic differences that differentiate Mycobacterium avium subspecies paratuberculosis strains. BMC Microbiol. 12:264. doi: 10.1186/1471-2180-12-264
Caire-Brändli, I., Papadopoulos, A., Malaga, W., Marais, D., Canaan, S., Thilo, L., et al. (2014). Reversible lipid accumulation and associated division arrest of Mycobacterium avium in lipoprotein-induced foamy macrophages may resemble key events during latency and reactivation of tuberculosis. Infect. Immun. 82, 476–490. doi: 10.1128/IAI.01196-13
Cirillo, J. D., Falkow, S., Tompkins, L. S., and Bermudez, L. E. (1997). Interaction of Mycobacterium avium with environmental amoebae enhances virulence. Infect. Immun. 65, 3759–3767.
Delafont, V., Mougari, F., Cambau, E., Joyeux, M., Bouchon, D., Héchard, Y., et al. (2014). First evidence of amoebae-mycobacteria association in drinking water network. Environ. Sci. Technol. 48, 11872–11882. doi: 10.1021/es5036255
Delafont, V., Samba-Louaka, A., Cambau, E., Bouchon, D., Moulin, L., and Héchard, Y. (2017). Mycobacterium llatzerense, a waterborne Mycobacterium, that resists phagocytosis by Acanthamoeba castellanii. Sci. Rep. 7:46270. doi: 10.1038/srep46270
Drancourt, M. (2014). Looking in amoebae as a source of mycobacteria. Microb. Pathog. 77, 119–124. doi: 10.1016/j.micpath.2014.07.001
Ghodbane, R., Mba Medie, F., Lepidi, H., Nappez, C., and Drancourt, M. (2014). Long-term survival of tuberculosis complex mycobacteria in soil. Microbiology 160, 496–501. doi: 10.1099/mic.0.073379-0
Greub, G., and Raoult, D. (2004). Microorganisms resistant to free-living amoebae. Clin. Microbiol. Rev. 17, 413–433. doi: 10.1128/CMR.17.2.413-433.2004
Hadziavdic, K., Lekang, K., Lanzen, A., Jonassen, I., Thompson, E. M., and Troedsson, C. (2014). Characterization of the 18S rRNA gene for designing universal eukaryote specific primers. PLoS ONE 9:e87624. doi: 10.1371/journal.pone.0087624
Holbert, S., Branger, M., Souriau, A., Lamoureux, B., Ganneau, C., Richard, G., et al. (2015). Interferon gamma response to Mycobacterium avium subsp. paratuberculosis specific lipopentapeptide antigen L5P in cattle. Res. Vet. Sci. 102, 118–121. doi: 10.1016/j.rvsc.2015.07.017
Jürgens, K., and Matz, C. (2002). Predation as a shaping force for the phenotypic and genotypic composition of planktonic bacteria. Antonie Van Leeuwenhoek 81, 413–434. doi: 10.1023/A:1020505204959
Kuehnel, M. P., Goethe, R., Habermann, A., Mueller, E., Rohde, M., Griffiths, G., et al. (2001). Characterization of the intracellular survival of Mycobacterium avium ssp. paratuberculosis: phagosomal pH and fusogenicity in J774 macrophages compared with other mycobacteria. Cell Microbiol. 3, 551–566. doi: 10.1046/j.1462-5822.2001.00139.x
Lefrançois, L. H., Pujol, C., Bodier, C. C., Teixeira-Gomez, A. P., Drobecq, H., Rosso, M. L., et al. (2011). Characterization of the Mycobacterium avium subsp. paratuberculosis laminin-binding/histone-like protein (Lbp/Hlp) which reacts with sera from patients with Crohn's disease. Microbes Infect 13, 585–594. doi: 10.1016/j.micinf.2011.02.002
Li, L., Bannantine, J. P., Campo, J. J., Randall, A., Grohn, Y. T., Katani, R., et al. (2017). Identification of sero-reactive antigens for the early diagnosis of Johne's disease in cattle. PLoS ONE 12:e0184373. doi: 10.1371/journal.pone.0184373
Li, L., Bannantine, J. P., Zhang, Q., Amonsin, A., May, B. J., Alt, D., et al. (2005). The complete genome sequence of Mycobacterium avium subspecies paratuberculosis. Proc. Natl. Acad. Sci. U.S.A. 102, 12344–12349. doi: 10.1073/pnas.0505662102
Magombedze, G., Shiri, T., Eda, S., and Stabel, J. R. (2017). Inferring biomarkers for Mycobacterium avium subsp. paratuberculosis infection and disease progression in cattle using experimental data. Sci. Rep. 7:44765. doi: 10.1038/srep44765
Molmeret, M., Horn, M., Wagner, M., Santic, M., and Abu Kwaik, Y. (2005). Amoebae as training grounds for intracellular bacterial pathogens. Appl. Environ. Microbiol. 71, 20–28. doi: 10.1128/AEM.71.1.20-28.2005
Mura, M., Bull, T. J., Evans, H., Sidi-Boumedine, K., McMinn, L., Rhodes, G., et al. (2006). Replication and long-term persistence of bovine and human strains of Mycobacterium avium subsp. paratuberculosis within Acanthamoeba polyphaga. Appl. Environ. Microbiol. 72, 854–859. doi: 10.1128/AEM.72.1.854-859.2006
Ott, S. L., Wells, S. J., and Wagner, B. A. (1999). Herd-level economic losses associated with Johne's disease on US dairy operations. Prev. Vet. Med. 40, 179–192. doi: 10.1016/S0167-5877(99)00037-9
Pillai, S. R., and Jayarao, B. M. (2002). Application of IS900 PCR for detection of Mycobacterium avium subsp. paratuberculosis directly from raw milk. J. Dairy Sci. 85, 1052–1057. doi: 10.3168/jds.S0022-0302(02)74165-9
Radomski, N., Roguet, A., Lucas, F. S., Veyrier, F. J., Cambau, E., Accrombessi, H., et al. (2013). atpE gene as a new useful specific molecular target to quantify Mycobacterium in environmental samples. BMC Microbiol. 13:277. doi: 10.1186/1471-2180-13-277
Rodríguez-Zaragoza, S. (1994). Ecology of free-living amoebae. Crit. Rev. Microbiol. 20, 225–241. doi: 10.3109/10408419409114556
Rohde, K., Yates, R. M., Purdy, G. E., and Russell, D. G. (2007). Mycobacterium tuberculosis and the environment within the phagosome. Immunol. Rev. 219, 37–54. doi: 10.1111/j.1600-065X.2007.00547.x
Salah, I. B., Ghigo, E., and Drancourt, M. (2009). Free-living amoebae, a training field for macrophage resistance of mycobacteria. Clin. Microbiol. Infect. 15, 894–905. doi: 10.1111/j.1469-0691.2009.03011.x
Salgado, M., Alfaro, M., Salazar, F., Badilla, X., Troncoso, E., Zambrano, A., et al. (2015). Application of cattle slurry containing Mycobacterium avium subsp. paratuberculosis (MAP) to grassland soil and its effect on the relationship between MAP and free-living amoeba. Vet. Microbiol. 175, 26–34. doi: 10.1016/j.vetmic.2014.09.022
Sweeney, R. W., Collins, M. T., Koets, A. P., McGuirk, S. M., and Roussel, A. J. (2012). Paratuberculosis (Johne's disease) in cattle and other susceptible species. J. Vet. Intern. Med. 26, 1239–1250. doi: 10.1111/j.1939-1676.2012.01019.x
Tasara, T., and Stephan, R. (2005). Development of an F57 sequence-based real-time PCR assay for detection of Mycobacterium avium subsp. paratuberculosis in milk. Appl. Environ. Microbiol. 71, 5957–5968. doi: 10.1128/AEM.71.10.5957-5968.2005
Thibault, V. C., Grayon, M., Boschiroli, M. L., Hubbans, C., Overduin, P., Stevenson, K., et al. (2007). New variable-number tandem-repeat markers for typing Mycobacterium avium subsp. paratuberculosis and M. avium strains: comparison with IS900 and IS1245 restriction fragment length polymorphism typing. J. Clin. Microbiol. 45, 2404–2410. doi: 10.1128/JCM.00476-07
Turenne, C. Y., Semret, M., Cousins, D. V., Collins, D. M., and Behr, M. A. (2006). Sequencing of hsp65 distinguishes among subsets of the Mycobacterium avium complex. J. Clin. Microbiol. 44, 433–440. doi: 10.1128/JCM.44.2.433-440.2006
Whan, L., Grant, I. R., and Rowe, M. T. (2006). Interaction between Mycobacterium avium subsp. paratuberculosis and environmental protozoa. BMC Microbiol. 6:63. doi: 10.1186/1471-2180-6-63
Keywords: paratuberculosis, amoebae, water, infection, Mycobacterium, Rosculus
Citation: Samba-Louaka A, Robino E, Cochard T, Branger M, Delafont V, Aucher W, Wambeke W, Bannantine JP, Biet F and Héchard Y (2018) Environmental Mycobacterium avium subsp. paratuberculosis Hosted by Free-Living Amoebae. Front. Cell. Infect. Microbiol. 8:28. doi: 10.3389/fcimb.2018.00028
Received: 31 October 2017; Accepted: 23 January 2018;
Published: 09 February 2018.
Edited by:
Thierry Soldati, Université de Genève, SwitzerlandReviewed by:
Marina Santic', University of Rijeka, CroatiaOtmane Lamrabet, Université de Genève, Switzerland
Fabienne Misguich, Versailles Saint-Quentin-en-Yvelines University, France
Copyright © 2018 Samba-Louaka, Robino, Cochard, Branger, Delafont, Aucher, Wambeke, Bannantine, Biet and Héchard. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Franck Biet, ZnJhbmNrLmJpZXRAaW5yYS5mcg==
Yann Héchard, eWFubi5oZWNoYXJkQHVuaXYtcG9pdGllcnMuZnI=