 José Pedro Lopes1,2,3*
José Pedro Lopes1,2,3* Marios Stylianou1,2,3
Marios Stylianou1,2,3 Emelie Backman1,2,3
Emelie Backman1,2,3 Sandra Holmberg1,2,3Maria Ekoff4
Sandra Holmberg1,2,3Maria Ekoff4 Gunnar Nilsson4
Gunnar Nilsson4 Constantin F. Urban1,2,3*
Constantin F. Urban1,2,3*- 1Department of Clinical Microbiology, Umeå University, Umeå, Sweden
- 2Umeå Centre for Microbial Research, Umeå University, Umeå, Sweden
- 3Laboratory for Molecular Infection Medicine Sweden, Umeå University, Umeå, Sweden
- 4Immunology and Allergy Division, Department of Medicine Solna, Karolinska Institutet, Karolinska University Hospital, Stockholm, Sweden
Cryptococcosis, caused by the basidiomycete Cryptococcus neoformans, is a life-threatening disease affecting approximately one million people per year worldwide. Infection can occur when C. neoformans cells are inhaled by immunocompromised people. In order to establish infection, the yeast must bypass recognition and clearance by immune cells guarding the tissue. Using in vitro infections, we characterized the role of mast cells (MCs) in cryptococcosis. We found that MCs recognize C. neoformans and release inflammatory mediators such as tryptase and cytokines. From the latter group MCs released mainly CCL-2/MCP-1, a strong chemoattractant for monocytic cells. We demonstrated that supernatants of infected MCs recruit monocytes but not neutrophils. During infection with C. neoformans, MCs have a limited ability to kill the yeast depending on the serotype. C. neoformans, in turn, modulates the lifespan of MCs both, by presence of its polysaccharide capsule and by secreting soluble modulators. Taken together, MCs might have important contributions to fungal clearance during early stages of cryptocococis where these cells regulate recruitment of monocytes to mucosal tissues.
Introduction
Cryptococcus neoformans is an encapsulated yeast, ubiquitous in the environment (Goldman et al., 2001). Infections with C. neoformans are commonly diagnosed in immunocompromised patients (Singh et al., 2008; Pappas, 2013; Ventura Aguiar et al., 2014) and with the onset of the HIV/AIDS epidemic, cryptococcosis became a leading cause of death in these patients mainly due to fatal meningoencephalitis (Park et al., 2009). The infectious process begins when the host inhales aerosolized cryptococcal cells. Some particles find their way into the alveolus where the resident cells of the innate immune system attempt to clear the infection. A large body of research shows an important role for alveolar macrophages in anti-cryptococcal defense by detecting and engulfing fungal spores (Giles et al., 2009; Mansour et al., 2014). Non-professional phagocytes, such as eosinophils and mast cells (MCs) have received less attention, although one report describes effector contribution of eosinophils and interaction of MCs and C. neoformans (Feldmesser et al., 1997). MCs contribute to immune defense against Aspergillus fumigatus and Candida albicans (Urb et al., 2009; Trevisan et al., 2014) and orchestrate a specific and temporal response by inducing inflammatory processes in the tissue (Lopes et al., 2015; Moretti et al., 2017). To induce inflammation, MCs take advantage of their ability to recognize fungi using pattern recognition receptors (PRR) (Yang and Marshall, 2009; Agier et al., 2018) which in turn triggers the release of granular vesicles containing a broad spectrum of inflammatory mediators, cytokines, and growth factors (Mukai et al., 2018).
Our hypothesis is that the outcome of cryptococcal infection is dependent on the initial interaction with resident immune cells of the mucosal tissue, such as MCs. We, therefore, decided to explore, the interaction of MCs and C. neoformans using in vitro infections of both, a cell line and primary mast cells. We further used capsular and acapsular C. neoformans strains to determine the importance of capsule to the infection outcome.
Materials and Methods
Microbial Culture Conditions
The strains used in this study were DSM 11959 (C. neoformans var. grubii), B-3501 (C. neoformans var. neoformans), and acapsular mutant cap59 (Zaragoza et al., 2008). In all cases, microbial cells were incubated overnight at 30°C in synthetic complete dropout medium + 2% glucose. A fresh subculture was inoculated before each experiment at 30°C for 3 h.
Immune Cell Culture Conditions
Human mast cell line 1 (HMC-1) and cord blood-derived mast cells (CBMC) were maintained and isolated as previously described (Lopes et al., 2015). Prior to any assays, MCs were primed with 25 nM PMA (12-myristate-13-acetate, Sigma-Aldrich) for 15 min at 37 °C, subsequently centrifuged for 10 min at 300 × g and re-suspended in RPMI without fetal calf serum or antibiotics. For spleen tyrosine kinase (Syk)-blocking experiments, MCs were blocked with piceatannol 10 μM for (Invivogen) for 15 min at 37°C. MCs were washed once before infection to remove excessive inhibitor.
Blood was obtained from the blood bank of the Norrland University Hospital Umeå anonymously following recommendations from the local ethical committee—Regionala etikprövningsnämnden i Umeå. Cord-blood donations were obtained from umbilical cord blood collected from normal, full-term deliveries at the Karolinska University Hospital. All donors were healthy donors and participants provided informed consent. Neutrophils were isolated from venous blood as previously described (Hosseinzadeh et al., 2012) and monocytes were isolated from buffy coats. Briefly, buffy coat blood was layered over equal amounts of Histopaque (ThermoScientific) and centrifuged at 900 rpm for 60 min. Leukocytes were collected and washed in PBS. Cells were then counted and diluted to 1 × 106 cells/ml in RPMI and added to cell culture flask. After 24 h non-adherent cells were removed and the cell culture flask washed with fresh RPMI. Adherent cells were collected using a cell scraper.
MC Granular Content Release
N-acetyl-beta-D-hexosaminidase and tryptase release was measured in HMC-1 cells (1 × 105 cells/well) infected in a 96-well plate with C. neoformans or left untreated. After 1 h, 60 μL of supernatant was transferred to a 96-well plate and mixed with an equal volume of 7.5 mM of p-nitrophenyl-N-acetyl-β-D-glucosaminide dissolved in 80 mM of citric acid, pH 4.5. The mixture was incubated shaking for 2 h at 37°C. After incubation, 120 μL of glycine (0.2 M, pH 10.7) was added and absorbance was measured in a plate reader (Fluostar, BMG Labtech). Tryptase was measured using the mast cell degranulation kit (Millipore) according to the manufacturer's instructions.
For cytokine release, MCs (1 × 106 cells/well) were infected with C. neoformans (MOI 5) for 20 h in 24-well plates at 37°C and 5% CO2. C neoformans only and uninfected MCs served as controls. At endpoint, the plate was centrifuged and supernatants collected and stored at −80°C. Supernatants were tested for the release of the following cytokines: IL-1β, IL-1ra, IL-2, IL-4, IL-5, IL-6, IL-7, IL-8, IL-9, IL-10, IL-12, IL-13, IL-15, IL-16, IL-17, IL-18, IL-5, Eotaxin, FGF Basic, G-CSF, GM-CSF, IFN-γ, IP-10, MCP-1, MIP-1a, PDGF-BB, MIP-1β, RANTES, TNF-α, VEGF, IL-1 α, IL-2RA, IL-12, CTACK, GROα, HGF, IFN-α2, LIF, MCP-3, M-CSF, MIF, MIG, β-NGF, SCF, SCGF-β, SDF-1 α, TNF-β, TRAIL using a Bio-Plex human cytokine assay (Bio-Rad Laboratories). Cytokine fold change was calculated as the ratio between infected condition and uninfected control. Additionally, MCP-1 release was confirmed using Human MCP-1 ELISA max deluxe (Biolegend) according to manufacturer's instructions.
Fungal Killing
MCs (1 × 105 cell/well) were infected with C. neoformans (MOI 5) in a 24-well plate. After 3 h, supernatants were plated in YEPD plates in triplicate. The fungal killing was measured as the number of fungal colonies forming units (CFU).
Chemotaxis
Using a transwell system, as previously described (Lopes et al., 2015), the chemotactic migration of neutrophils (5 × 105 cells/well) and monocytes (5 × 105 cells/well) was tested with supernatants of MCs (1 × 105 cell/well) infected with C. neoformans (MOI 5), uninfected controls or the equivalent number of fungal cells. Neutrophil migration was accessed for 30 min and monocyte migration for 90 min. Tested supernatants were collected and stored similarly as described for cytokine release.
Phagocytosis
MCs phagocytosis of capsular and acapsular C. neoformans yeasts was performed according to Alanio et al. (2011) with slight modifications. Yeast cells (5 × 106 /ml cells) were stained with IgG-Alexa-488 dye (A11006 Invitrogen) for 20 min and then centrifuged at 3,000 g for 10 min. Mast cells (1 × 106 cells/ml) and C. neoformans were seeded to a flat-bottomed 24-well plate and incubated in a CO2 incubator at 37°C for 1 and 2 h. At the end-point, the mixture was collected to 1.5 ml eppendorf tubes and IgG-Alexa-633 dye (A21086 Invitrogen) was added for 20 min. The tubes were then centrifuged at 1,500 g for 10 min and the pellet re-suspended in cold deionized water for 10 min. The tubes were centrifuged at 1,500 g for 10 min and the pellet re-suspended in 1x PBS and measured using a FACS Accuri C6 instrument (BD Biosciences) with post-acquisition data analysis with Accuri C6 analysis. Phagocytized cryptococci were determined as the % of IgG-Alexa-488 dye high/IgG-Alexa-633 low cells in the FACS plot.
Cell Death
MCs (5 × 104 cells/well) were infected with C. neoformans (MOI 5) and host cell death was quantified using a sytox green-based (ThermoScientific) cell viability assay as described previously (Lopes et al., 2015). To measure the effect of cryptococcal mediators released into solution during delay of MC death we incubated C. neoformans for 20 h in RPMI medium. Cultures were centrifuged at 3,000 rpm for 10 min and the supernatants collected. Conditioned media containing 5, 10, 25, or 50% of overnight supernatants + fresh RPMI was incubated with MCs in the presence of sytox green.
Statistical Analysis
All data are shown as mean ± SD. Significance in degranulation and cytokine release experiments was analyzed by multi-comparison using one-way ANOVA with Bonferroni post-test. Cell death experiments were analyzed applying two-way ANOVA with Bonferroni's post-test. Syk-inhibition and fungal killing experiment significances were analyzed using unpaired t-test with Welch correction. For all analyses, p-values < 0.05 were considered statistically significant.
Results
C. neoformans Triggers MC Degranulation
Secretion of granules containing inflammatory molecules is known to be one of the primary mechanisms employed by MCs. Degranulation elicits inflammatory reactions and recruits other immune cells (Beghdadi et al., 2011; Dudeck et al., 2018). MCs have recently been shown to degranulate in response to a component of the fungal cell, β-glucan, with a unique response pattern with included release of histamine and β-hexosaminidase but not LTC4, IL-6, or CCL2 (Barbosa-Lorenzi et al., 2017). We first assessed the degranulation of MCs in response to C. neoformans using tryptase and β-hexosaminidase quantification in supernatants of in vitro infection (Figures 1A,B). C. neoformans triggered MCs to release of tryptase and β-hexosaminidase in a dose-dependent manner. At MOI 1, MCs degranulated tryptase 30% above control levels (Figure 1A). Degranulation of β-hexosaminidase increased by 20% at MOI 1 and 50% at MOI 5 above control levels (Figure 1B).
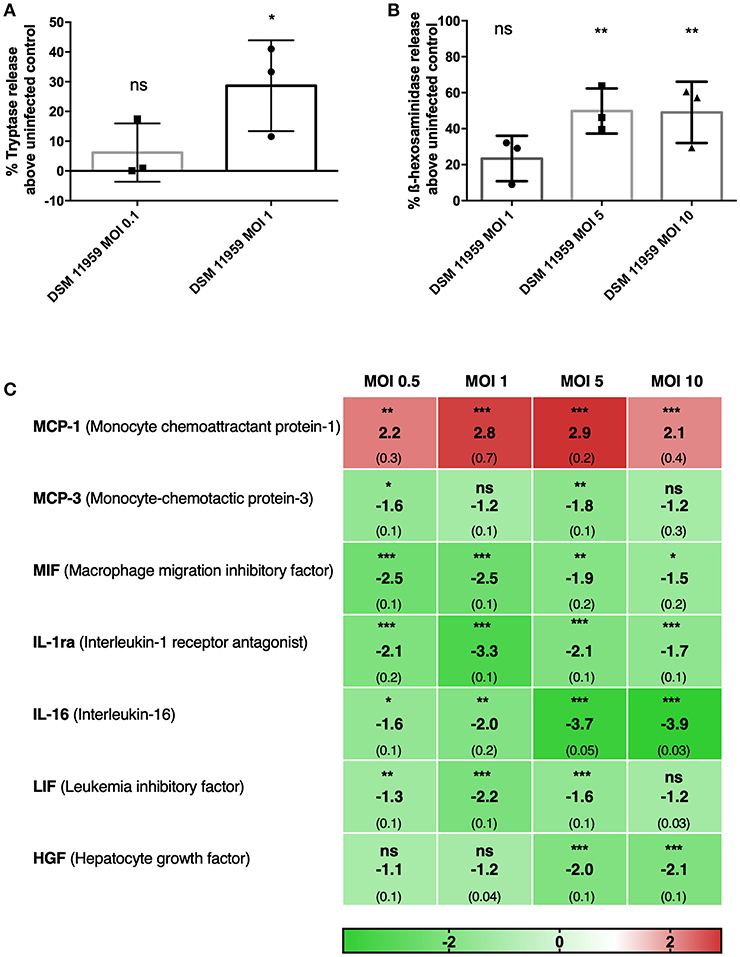
Figure 1. MCs show direct action against C. neoformans and release granular proteins. (A) MCs released tryptase after 6 h of infection with C. neoformans (MOI 1). Data from n = 3 (3). (B) MCs released ß-hexosaminidase after 1 h infection with C. neoformans (MOI 5 and 10). Data from n = 3 (3). (C) Fold change of a total of 48 cytokines were measured in supernatants of MCs infected with C. neoformans at different MOIs. Seven cytokines were differentially expressed and only MCP-1 was upregulated. Fold change was calculated as the ratio between infected condition and uninfected control. Data from n = 2 (4). For all analyses significance was analyzed by multi-comparison using one-way ANOVA with Bonferroni post-test. *P ≤ 0.05; **P ≤ 0.01; ***P ≤ 0.001; ns, not significant.
To get a grip on effector molecules released from MCs released upon C. neoformans infection, we collected supernatants after 20 h of incubation and performed multiplex cytokine assays for 48 different molecules. Surprisingly, from the 48 tested cytokines, only MCP-1 was released upon C. neoformans stimulation (Figure 1C). MCP-1 is one of the key chemokines that regulate migration and infiltration of monocytes/macrophages to the tissues in response to infection (Deshmane et al., 2009). Since macrophages play an important role in anti-cryptococcal defense, we used MCP-1 secretion as a relevant readout for subsequent assays assessing MC activation upon encounter with C. neoformans. Cytokines released to a significantly lower extent than without stimulus encompassed MIF, MCP-3, IL-1ra, IL-16, LIF, and HGF.
To investigate if the release of MC mediators could contribute to the clearance of C. neoformans, we infected MCs with C. neoformans for 3 h and quantified the viability of yeast cells by plating (Figure 2A). We tested wild-type strain DSM 11959 and saw no reduction on the fungal load after incubation with MCs. To exclude if the presence of the capsule could disturb the killing, we used acapsular mutant cap59 and parental strain B3501. The viability of both strains was reduced approximately by 3-fold but surprisingly no statistical difference between the killing of the capsular strain and the acapsular mutant was seen. We propose that the difference between the two wild-type strains might arise from different susceptibility between different C. neoformans serotypes—DSM 11959 (serotype A) and B3501 (serotype D).
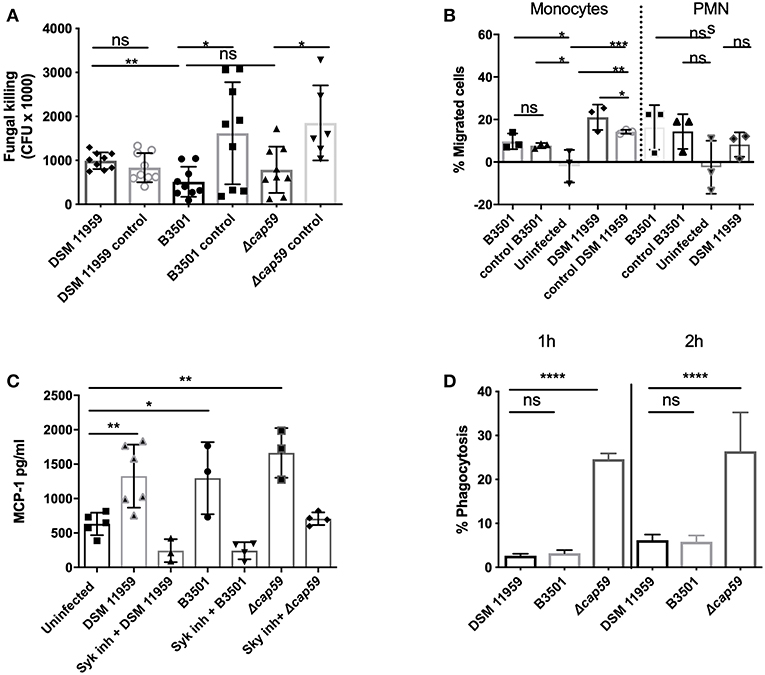
Figure 2. MC recognition of C. neoformans was mediated by Syk signaling and chemoattracts monocytes. (A) MCs induced killing was serotype dependent. Serotype A (DSM 11959) resisted killing by MCs whereas serotype D (B3501) and acapsular mutant cap59 showed some level of susceptibility. Data from n = 9. (B) Monocytes but not neutrophils were recruited by MCs after C. neoformans infection. The graph depicts the percentage of migration of neutrophil or monocyte migration over time toward supernatants of MCs infected with C. neoformans. (C) MCs secreted MCP-1 in a Syk dependent manner after 20 h infection with C. neoformans (MOI 5). MCP-1 was measured by ELISA in supernatants of HMC-1 infected with C. neoformans (MOI 5). Data from n = 3 (3). Syk was inhibited by piceatannol 10 μM after infection with HMC-1 infection with C. neoformans strains. (D) Acapsular mutants induce significantly more phagocytosis after 1 and 2 h infection. MC phagocytosis of C. neoformans was determined by FACS as the percentage of IgG-Alexa-488 dye high/IgG-Alexa-633 low Cryptococcus cells. Data from n = 4. Significance was analyzed by multi-comparison using one-way ANOVA with Bonferroni post-test in (B,D). For (A,C) significance was analyzed using unpaired t-test with Welch correction. *P ≤ 0.05; **P ≤ 0.01; ***P ≤ 0.001; ****P ≤ 0.0001; ns, not significant.
We conclude that MCs sense soluble components of C. neoformans and respond in a direct manner by releasing granular effectors and cytokines. The antifungal potential of these responses is dependent on the serotype of encountered C. neoformans strains.
Monocytes but Not Neutrophils Are Chemoattracted in Response to Infection of MCs
Since MCs can modulate the recruitment of immune cells, we followed up on the ability of the chemokines secreted from infected MCs to induce migration of neutrophils and monocytes. In a functional migration assay we fluorescently labeled neutrophils or monocytes, added them to the upper compartment of a transwell system and allowed them to migrate toward the lower compartment containing supernatants from MCs infected with C. neoformans. In agreement with the exclusive induction of MCP-1 release, supernatants of infected MCs were chemotactic for monocytes but not for neutrophils, as only monocytes showed increased migration toward the lower compartment as compared to control samples (Figure 2B). Specific migration in response to the infection supernatants doubled as compared to migration toward supernatants from fungal cells alone. This indicates that MC degranulation triggered by C. neoformans selectively induced monocyte recruitment to the site of infection.
MCs Recognize C. neoformans in a Syk-Dependent Manner
MCs use PRR to recognize fungal pathogen-associated molecular patterns (PAMPs). Most commonly, PRR ligands are fungal cell wall components, such as β-glucan. C-type lectin receptors (CLRs) are PRRs particularly important for the recognition of fungal PAMPs, for instance, CLR dectin-1 binds β-glucan (Shiokawa et al., 2017). Ligand binding to the C-type lectin receptors—dectin-1 and dectin-2 and mincle—triggers recruitment and phosphorylation of spleen tyrosine kinase (Syk) (Plato et al., 2013).
To test the role of CLRs in MC recognition of C. neoformans we blocked Syk activation in MCs by using specific inhibitor piceatannol using MCP-1 release as a readout (Oliver et al., 1994). Syk inhibition reduced MCP-1 release to basal levels suggesting that MC recognition of C. neoformans and release of MCP-1 is dependent on the signaling cascades involving CLRs and Syk (Figure 2C).
C. neoformans Capsule Is Protective Against MC Phagocytosis
The polysaccharide capsule is an essential virulence factor and plays an important role in the interaction of this fungus with host immune recognition and response pathways (Alspaugh, 2015). In order to study the effect of the capsule of C. neoformans during MC phagocytosis, we incubated C. neoformans with MCs in a 24-well microtiter plate for 1 h and 2 h (Figure 2C). All C. neoformans cells were pre-labeled before infection and extracellular cells were additionally labeled at the end of the experiment. After successful MC lysis and FACS analysis, the percentage of intracellular yeast (single-labeled cells) could be determined.
Percentage of phagocytosis of cells from the capsular strain was 2% (DSM 11959) and 3% (B3501) at 1 h increasing to 5% (DSM 11959) and 7% (B3501) at 2 h. Cells from the acapsular mutant strain on the other hand where phagocytosed at much higher rates −25% at 1 h with an increase to 27% at 2 h. We concluded that the C. neoformans capsule protects against MC phagocytosis.
Both Capsule and Soluble Factors of the C. neoformans Membrane Contribute to MC Survival
To quantify MC death induced by C. neoformans we used a microplate-based fluorescence assay and the membrane-impermeable fluorescent DNA dye sytox. Fluorescent signal by DNA staining only occurs in cells with compromised plasma membranes. Cell death was calculated as the percentage of lysis control. Unexpectedly, both capsular C. neoformans strains promoted MC survival as uninfected MCs died half as much than infected cells (Figure 3A). Infection with acapsular mutants caused higher levels of death in MCs that were nevertheless not distinct from the death rate of uninfected controls (Figure 3A). We obtained similar results using primary MCs (Figure 3B). We concluded that C. neoformans capsule was important for modulation of the MC lifespan albeit not being the only factor responsible for that.
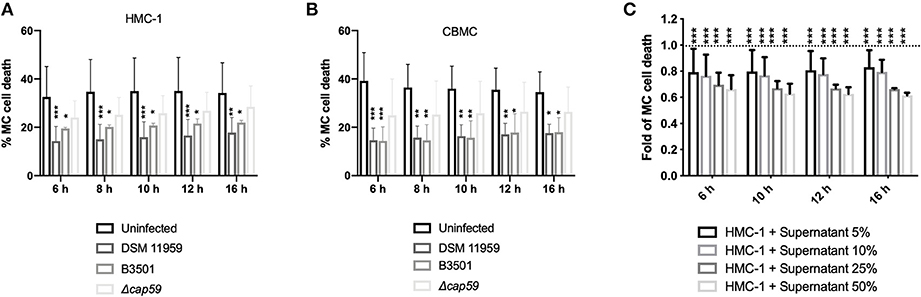
Figure 3. C. neoformans infection increase MC survival. (A) Infection with capsular C. neoformans increased HMC-1 survival. Infections were performed using MOI of 5. Data from n = 6 (3). (B) Infection with capsular C. neoformans (MOI 5) increased viability of primary MCs. Data from n = 4 (3). (C) Secreted components from C. neoformans overnight cultures induced MC survival. Plotted is the delayed death of resting MCs incubated with increasing concentrations of supernatants of an overnight culture of C. neoformans plotted as fold change in relation to untreated MCs (dashed line). For all analyses significance was analyzed by multi-comparison using two-way ANOVA with Bonferroni post-test. *P ≤ 0.05; **P ≤ 0.01; ***P ≤ 0.001; ****P ≤ 0.0001; ns, not significant.
C. neoformans is known to secrete vesicles into a culture containing virulence factors and other cargo (Wolf et al., 2014). We reasoned that fungal secretion of extracellular factors could additionally help postpone MC death. To test our hypothesis, we incubated supernatants from an overnight culture of C. neoformans in different dilutions with fresh medium and incubated with resting MCs (Figure 3C). Increasing concentration of fungal conditioned supernatants partially delayed MC death. When using 50% fungal conditioned media MC cell death was decreased to 65% of uninfected baseline.
Overall, we concluded that MC lifespan is controlled by both by capsule presence and by the release of soluble fungal components.
Discussion
C. neoformans is one of the most important causative agents of cryptococcosis and one of the major fungal pathogens. Yearly, C. neoformans is responsible for over 180 000 deaths worldwide affecting especially populations in Sub-Sahara (Rajasingham et al., 2017). Cryptococcosis is strongly associated with groups at risk to acquire severe opportunistic infections, such as HIV/AIDS patients (Rodrigues, 2016; Molloy et al., 2017).
Here, we explored the role of MCs in antifungal immunity against C. neoformans. MCs are important actors to maintain the equilibrium in host mucosa with significant contributions to symptoms caused by hypersensitivity and inflammatory reactions. Owing to the variety of expressed receptors MCs can carry out effector and regulatory tasks reacting to external stimuli (Halova et al., 2018). MCs are present in the oral mucosa of the nasal cavity and of the lungs and are therefore some of the most common cells to interact with C. neoformans. Using an in vitro model, we studied the interaction between MCs and C. neoformans. We applied different approaches to qualitatively and quantitatively assess this interplay.
Quantification of released mediators known to be produced by MCs in order to orchestrate immune reactions (Fukuishi et al., 2014; Metcalfe et al., 2016), such as β-hexosaminidase, tryptase, and cytokines, revealed that MCs recognized and responded to C. neoformans (Figures 1A,B).
Analysis of the cytokine multiplex showed that out of 48 cytokines only 7 were differentially secreted upon C. neoformans infection. From those only MCP-1 was released in a dose-dependent manner, whereas the release of six other cytokines and chemokines was reduced. C. neoformans infection has been shown to stimulate the production of MCP-1 in the lungs (Huffnagle et al., 1995) and failure to induce MCP-1 resulted in a diminished cell-mediated inflammatory response in vivo and diminished cryptococcal clearance (Huffnagle et al., 1995; Levitz et al., 1997). Three other cytokines are involved in leukocyte recruitment: MIF and MCP-3 are important for the recruitment of monocytic cells, whereas IL-16 is important for lymphocyte migration. The remaining three - IL-1ra, LIF, and HGF—are important in the regulation of inflammation. If the cytokine downregulation can be attributed to the direct action of the fungus remains to be determined but C. neoformans capsule polysaccharide has been previously implicated in cytokine downregulation in human monocytes (Vecchiarelli et al., 1995) while C. gattii downregulates the expression of antimicrobial peptide, cathelicidin, in human PBMCs (Herkert et al., 2018). Interestingly, in a previous study, we measured the response of MCs to C. albicans and detected a very different pattern of cytokine release (Lopes et al., 2018). Hence, MCs seem to orchestrate cytokine and chemokine responses dependently of the encountered fungal pathogen.
We tested the chemotactic potential of MC supernatants from infected cells. These supernatants induced migration of macrophages but not of neutrophils (Figure 2B). Neutrophils, besides being effective killers of fungi, constitute the second line of defense against C. neoformans behind alveolar macrophages (Mambula et al., 2000; Chiller et al., 2002) which are essential for fungal clearance during cryptococcosis (Hardison et al., 2012; Campuzano and Wormley, 2018). The pathogen has however evolved strategies to survive and replicate inside the macrophages and even use the host cell as a Trojan horse for dissemination (Geunes-Boyer et al., 2009; Casadevall, 2010; Davis et al., 2015). Our data suggest that C. neoformans also exploits MC responses in this regard since monocytic cells were selectively recruited.
Recognition of C. neoformans by MCs was dependent on Syk signaling (Figure 2C). This is in good agreement with a recent report describing non-opsonic uptake of cryptoccoci to be dependent on CLRs and its adaptor molecule Syk (Lim et al., 2018). Syk signaling is also know to be required for chemotaxis, helping in the reorganization of the actin cytoskeleton in response to chemokines (Park and Cox, 2011).
Antifungal activity of MCs was dependent on the fungal serotype since one wild-type strain was virtually resistant against MCs (Figure 2A). Alanio et al. showed that different clinical isolates of C. neoformans evoked varying macrophage responses contributing to different disease outcomes (Alanio et al., 2011). Notably, the presence of the capsule made no difference to the killing activity of MCs (Figure 2A). The killing of C. neoformans by neutrophils is also not influenced by the size or presence of the capsule but rather by the melanin-producing capacity of the fungus (Qureshi et al., 2011).
Moreover, C. neoformans infection delayed MC death as demonstrated by using both a cell line and primary cells (Figures 3A,B). Delaying host cell apoptosis is a mechanism used by other microbes to allow time for replication and cell-to-cell spread (Clifton et al., 1998). In this context, the immunomodulatory effect could reflect the ability of the fungus to augment immune responses mediated by MCs. Increased monocytic migration might allow the fungal cell to subsequently exploit incoming monocytic cells as a reservoir for dissemination. However, inhibition of apoptosis leading to in vitro survival of immune cells in response to β-glucan may also augment immune defense (Garcia-Valtanen et al., 2017).
The delay of MC death is larger when using capsular strains indicating a contribution of the polysaccharide capsule. Nevertheless, acapsular strains had similar death rates as uninfected MCs suggesting a complementary mechanism of modulation of MC lifespan (Figures 3A,B). We, therefore, decided to explore, if released fungal mediators could additionally contribute to delayed cellular death of MCs. Indeed, secretion of soluble mediators as culture supernatants helped to additionally mediate the effect (Figure 3C). The release of extracellular vesicles containing virulence factors is a described approach used by the fungus to modulate immune cell function (Rodrigues et al., 2007; Oliveira et al., 2010). During replication of C. neoformans in macrophages, accumulation of vesicles in the cytoplasm occurs (Tucker and Casadevall, 2002; Davis et al., 2015), which have immune modulatory effects (Ellerbroek et al., 2004; Bielska et al., 2018). Besides containing the major capsular polysaccharide, glucuronoxylomannan (GXM), other molecules such as sterylglucosidase are also present in these vesicles. Vesicles enriched in both components have been shown to delay the acute lethality C. neoformans infection (Colombo et al., 2019). Identification of the responsible soluble factors is currently under investigation.
Taken together, we show a new role for MCs during cryptococcosis. MCs influence the mounting of innate immune responses and can contribute to clearance. C. neoformans, in turn, might exploit the immune response aiming for dissemination. These findings highlight the complexity of MC and C. neoformans interaction. Further studies are needed to characterize the function of MCs during cryptococcosis in more detail, to find new targets for improved disease prevention strategies.
Data Availability
The raw data supporting the conclusions of this manuscript will be made available by the authors, without undue reservation, to any qualified researcher.
Ethics Statement
Blood was obtained from the blood bank of the Norrland University Hospital Umeå anonymously following recommendations from the local ethical committee—Regionala etikprövningsnämnden i Umeå. Cord-blood donation were obtained from umbilical cord blood collected from normal, full-term deliveries at the Karolinska University Hospital. All donors were healthy donors and participants provided informed consent.
Author Contributions
CU conceived and designed the study. JL, MS, EB, and SH performed the experiments and analyzed the data. ME and GN provided materials and expertise. JL and CU wrote the manuscript.
Funding
This study was funded by grants to CU from the Medical Faculty Umeå University (316-886-10) and the Swedish Research Council VR-M 2017-01681 and VR-M 2014-2281. JL acknowledges grant support from J. C. Kempes Minnes Stipendiefond. The funders had no role in study design, data collection, and analysis, decision to publish, or preparation of the manuscript.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We would like to thank Oscar Zaragoza for the B3501 and cap59 strains.
References
Agier, J., Pastwinska, J., and Brzezinska-Błaszczyk, E. (2018). An overview of mast cell pattern recognition receptors. Inflamm. Res. 67, 737–746. doi: 10.1007/s00011-018-1164-5
Alanio, A., Desnos-Ollivier, M., and Dromer, F. (2011). Dynamics of Cryptococcus neoformans-macrophage interactions reveal that fungal background influences outcome during cryptococcal meningoencephalitis in humans. MBio 2:e00158–11. doi: 10.1128/mBio.00158-11
Alspaugh, J. A. (2015). Virulence mechanisms and Cryptococcus neoformans pathogenesis. Fungal Genet. Biol. 78, 55–58. doi: 10.1016/j.fgb.2014.09.004
Barbosa-Lorenzi, V. C., Peyda, S., Scheynius, A., Nilsson, G., and Lunderius-Andersson, C. (2017). Curdlan induces selective mast cell degranulation without concomitant release of LTC4, IL-6 or CCL2. Immunobiology 222, 647–650. doi: 10.1016/j.imbio.2016.12.001
Beghdadi, W., Madjene, L. C., Benhamou, M., Charles, N., Gautier, G., Launay, P., et al. (2011). Mast cells as cellular sensors in inflammation and immunity. Front. Immunol. 2:37. doi: 10.3389/fimmu.2011.00037
Bielska, E., Sisquella, M. A., Aldeieg, M., Birch, C., O'donoghue, E. J., and May, R. C. (2018). Pathogen-derived extracellular vesicles mediate virulence in the fatal human pathogen Cryptococcus gattii. Nat. Commun. 9:1556. doi: 10.1038/s41467-018-03991-6
Campuzano, A., and Wormley, F. L. (2018). Innate immunity against Cryptococcus, from recognition to elimination. J. Fungi 4:E33. doi: 10.3390/jof4010033
Casadevall, A. (2010). Cryptococci at the brain gate: break and enter or use a Trojan horse? J. Clin. Invest. 120, 1389–1392. doi: 10.1172/JCI42949
Chiller, T., Farrokhshad, K., Brummer, E., and Stevens, D. A. (2002). Effect of granulocyte colony-stimulating factor and granulocyte-macrophage colony-stimulating factor on polymorphonuclear neutrophils, monocytes or monocyte-derived macrophages combined with voriconazole against Cryptococcus neoformans. Med. Mycol. 40, 21–26. doi: 10.1080/mmy.40.1.21.26
Clifton, D. R., Goss, R. A., Sahni, S. K., Van Antwerp, D., Baggs, R. B., Marder, V. J., et al. (1998). NF-kappa B-dependent inhibition of apoptosis is essential for host cellsurvival during Rickettsia rickettsii infection. Proc. Natl. Acad. Sci. U.S.A. 95, 4646–4651. doi: 10.1073/pnas.95.8.4646
Colombo, A. C., Rella, A., Normile, T., Joffe, L. S., Tavares, P. M., De, S. Araújo G. R., et al. (2019). Cryptococcus neoformans glucuronoxylomannan and sterylglucoside are required for host protection in an animal vaccination model. MBio 10:18. doi: 10.1128/mBio.02909-18
Davis, M. J., Eastman, A. J., Qiu, Y., Gregorka, B., Kozel, T. R., Osterholzer, J. J., et al. (2015). Cryptococcus neoformans-induced macrophage lysosome damage crucially contributes to fungal virulence. J. Immunol. 194, 2219–2231. doi: 10.4049/jimmunol.1402376
Deshmane, S. L., Kremlev, S., Amini, S., and Sawaya, B. E. (2009). Monocyte chemoattractant protein-1 (MCP-1): an overview. J. Interferon Cytokine Res. 29, 313–326. doi: 10.1089/jir.2008.0027
Dudeck, J., Froebel, J., Kotrba, J., Lehmann, C. H. K., Dudziak, D., Speier, S., et al. (2018). Engulfment of mast cell secretory granules on skin inflammation boosts dendritic cell migration and priming efficiency. J. Allergy Clin. Immunol. 143, 1849–1864.e4. doi: 10.1016/j.jaci.2018.08.052
Ellerbroek, P. M., Walenkamp, A. M., Hoepelman, A. I., and Coenjaerts, F. E. (2004). Effects of the capsular polysaccharides of Cryptococcus neoformans on phagocyte migration and inflammatory mediators. Curr. Med. Chem. 11, 253–266. doi: 10.2174/0929867043456188
Feldmesser, M., Casadevall, A., Kress, Y., Spira, G., and Orlofsky, A. (1997). Eosinophil-Cryptococcus neoformans interactions in vivo and in vitro. Infect. Immun. 65, 1899–1907.
Fukuishi, N., Murakami, S., Ohno, A., Yamanaka, N., Matsui, N., Fukutsuji, K., et al. (2014). Does beta-hexosaminidase function only as a degranulation indicator in mast cells? The primary role of beta-hexosaminidase in mast cell granules. J. Immunol. 193, 1886–1894. doi: 10.4049/jimmunol.1302520
Garcia-Valtanen, P., Guzman-Genuino, R. M., Williams, D. L., Hayball, J. D., and Diener, K. R. (2017). Evaluation of trained immunity by beta-1, 3 (d)-glucan on murine monocytes in vitro and duration of response in vivo. Immunol. Cell. Biol. 95, 601–610. doi: 10.1038/icb.2017.13
Geunes-Boyer, S., Oliver, T. N., Janbon, G., Lodge, J. K., Heitman, J., Perfect, J. R., et al. (2009). Surfactant protein D increases phagocytosis of hypocapsular Cryptococcus neoformans by murine macrophages and enhances fungal survival. Infect. Immun. 77, 2783–2794. doi: 10.1128/IAI.00088-09
Giles, S. S., Dagenais, T. R., Botts, M. R., Keller, N. P., and Hull, C. M. (2009). Elucidating the pathogenesis of spores from the human fungal pathogen Cryptococcus neoformans. Infect. Immun. 77, 3491–3500. doi: 10.1128/IAI.00334-09
Goldman, D. L., Khine, H., Abadi, J., Lindenberg, D. J., Pirofski, La., Niang, R., et al. (2001). Serologic evidence for Cryptococcus neoformans infection in early childhood. Pediatrics 107:E66. doi: 10.1542/peds.107.5.e66
Halova, I., Rönnberg, E., Draberova, L., Vliagoftis, H., Nilsson, G. P., and Draber, P. (2018). Changing the threshold-Signals and mechanisms of mast cell priming. Immunol. Rev. 282, 73–86. doi: 10.1111/imr.12625
Hardison, S. E., Herrera, G., Young, M. L., Hole, C. R., Wozniak, K. L., and Wormley, F. L. Jr. (2012). Protective immunity against pulmonary cryptococcosis is associated with STAT1-mediated classical macrophage activation. J. Immunol. 189, 4060–4068. doi: 10.4049/jimmunol.1103455
Herkert, P. F., Dos Santos, J. C., Hagen, F., Ribeiro-Dias, F., Queiroz-Telles, F., Netea, M. G., et al. (2018). Differential in vitro cytokine induction by the species of Cryptococcus gattii complex. Infect. Immun. 86:e00958–17. doi: 10.1128/IAI.00958-17
Hosseinzadeh, A., Messer, P. K., and Urban, C. F. (2012). Stable Redox-Cycling Nitroxide Tempol Inhibits NET Formation. Front. Immunol. 3:391. doi: 10.3389/fimmu.2012.00391
Huffnagle, G. B., Strieter, R. M., Standiford, T. J., Mcdonald, R. A., Burdick, M. D., Kunkel, S. L., et al. (1995). The role of monocyte chemotactic protein-1 (MCP-1) in the recruitment of monocytes and CD4+ T cells during a pulmonary Cryptococcus neoformans infection. J. Immunol. 155, 4790–4797.
Levitz, S. M., North, E. A., Jiang, Y., Nong, S. H., Kornfeld, H., and Harrison, T. S. (1997). Variables affecting production of monocyte chemotactic factor 1 from human leukocytes stimulated with Cryptococcus neoformans. Infect. Immun. 65, 903–908.
Lim, J., Coates, C. J., Seoane, P. I., Garelnabi, M., Taylor-Smith, L. M., Monteith, P., et al. (2018). Characterizing the mechanisms of nonopsonic uptake of cryptococci by macrophages. J. Immunol. 200, 3539–3546. doi: 10.4049/jimmunol.1700790
Lopes, J. P., Stylianou, M., Backman, E., Holmberg, S., Jass, J., Claesson, R., et al. (2018). Evasion of immune surveillance in low oxygen environments enhances Candida albicans virulence. MBio 9:e02120–18. doi: 10.1128/mBio.02120-18
Lopes, J. P., Stylianou, M., Nilsson, G., and Urban, C. F. (2015). Opportunistic pathogen Candida albicans elicits a temporal response in primary human mast cells. Sci. Rep. 5:12287. doi: 10.1038/srep12287
Mambula, S. S., Simons, E. R., Hastey, R., Selsted, M. E., and Levitz, S. M. (2000). Human neutrophil-mediated nonoxidative antifungal activity against Cryptococcus neoformans. Infect. Immun. 68, 6257–6264. doi: 10.1128/IAI.68.11.6257-6264.2000
Mansour, M. K., Reedy, J. L., Tam, J. M., and Vyas, J. M. (2014). Macrophage cryptococcus interactions: an update. Curr. Fungal Infect. Rep. 8, 109–115. doi: 10.1007/s12281-013-0165-7
Metcalfe, D. D., Pawankar, R., Ackerman, S. J., Akin, C., Clayton, F., Falcone, F. H., et al. (2016). Biomarkers of the involvement of mast cells, basophils and eosinophils in asthma and allergic diseases. World Allergy Organ J. 9:7. doi: 10.1186/s40413-016-0094-3
Molloy, S. F., Chiller, T., Greene, G. S., Burry, J., Govender, N. P., Kanyama, C., et al. (2017). Cryptococcal meningitis: a neglected NTD? PLoS Negl. Trop. Dis. 11:e0005575. doi: 10.1371/journal.pntd.0005575
Moretti, S., Renga, G., Oikonomou, V., Galosi, C., Pariano, M., Iannitti, R. G., et al. (2017). A mast cell-ILC2-Th9 pathway promotes lung inflammation in cystic fibrosis. Nat. Commun. 8:14017. doi: 10.1038/ncomms14017
Mukai, K., Tsai, M., Saito, H., and Galli, S. J. (2018). Mast cells as sources of cytokines, chemokines, and growth factors. Immunol. Rev. 282, 121–150. doi: 10.1111/imr.12634
Oliveira, D. L., Freire-De-Lima, C. G., Nosanchuk, J. D., Casadevall, A., Rodrigues, M. L., and Nimrichter, L. (2010). Extracellular vesicles from Cryptococcus neoformans modulate macrophage functions. Infect. Immun. 78, 1601–1609. doi: 10.1128/IAI.01171-09
Oliver, J. M., Burg, D. L., Wilson, B. S., Mclaughlin, J. L., and Geahlen, R. L. (1994). Inhibition of mast cell Fc epsilon R1-mediated signaling and effector function by the Syk-selective inhibitor, piceatannol. J. Biol. Chem. 269, 29697–29703.
Pappas, P. G. (2013). Cryptococcal infections in non-HIV-infected patients. Trans. Am. Clin. Climatol. Assoc. 124, 61–79.
Park, B. J., Wannemuehler, K. A., Marston, B. J., Govender, N., Pappas, P. G., and Chiller, T. M. (2009). Estimation of the current global burden of cryptococcal meningitis among persons living with HIV/AIDS. AIDS 23, 525–530. doi: 10.1097/QAD.0b013e328322ffac
Park, H., and Cox, D. (2011). Syk regulates multiple signaling pathways leading to CX3CL1 chemotaxis in macrophages. J. Biol. Chem. 286, 14762–14769. doi: 10.1074/jbc.M110.185181
Plato, A., Willment, J. A., and Brown, G. D. (2013). C-type lectin-like receptors of the dectin-1 cluster: ligands and signaling pathways. Int. Rev. Immunol. 32, 134–156. doi: 10.3109/08830185.2013.777065
Qureshi, A., Grey, A., Rose, K. L., Schey, K. L., and Del Poeta, M. (2011). Cryptococcus neoformans modulates extracellular killing by neutrophils. Front. Microbiol. 2:193. doi: 10.3389/fmicb.2011.00193
Rajasingham, R., Smith, R. M., Park, B. J., Jarvis, J. N., Govender, N. P., Chiller, T. M., et al. (2017). Global burden of disease of HIV-associated cryptococcal meningitis: an updated analysis. Lancet Infect. Dis. 17, 873–881. doi: 10.1016/S1473-3099(17)30243-8
Rodrigues, M. L. (2016). Funding and innovation in diseases of neglected populations: the paradox of cryptococcal meningitis. PLoS Negl. Trop. Dis. 10:e0004429. doi: 10.1371/journal.pntd.0004429
Rodrigues, M. L., Nimrichter, L., Oliveira, D. L., Frases, S., Miranda, K., Zaragoza, O., et al. (2007). Vesicular polysaccharide export in Cryptococcus neoformans is a eukaryotic solution to the problem of fungal trans-cell wall transport. Eukaryot. Cell 6, 48–59. doi: 10.1128/EC.00318-06
Shiokawa, M., Yamasaki, S., and Saijo, S. (2017). C-type lectin receptors in anti-fungal immunity. Curr. Opin. Microbiol. 40, 123–130. doi: 10.1016/j.mib.2017.11.004
Singh, N., Alexander, B. D., Lortholary, O., Dromer, F., Gupta, K. L., John, G. T., et al. (2008). Pulmonary cryptococcosis in solid organ transplant recipients: clinical relevance of serum cryptococcal antigen. Clin. Infect. Dis. 46, e12–18. doi: 10.1086/524738
Trevisan, E., Vita, F., Medic, N., Soranzo, M. R., Zabucchi, G., and Borelli, V. (2014). Mast cells kill Candida albicans in the extracellular environment but spare ingested fungi from death. Inflammation 37, 2174–2189. doi: 10.1007/s10753-014-9951-9
Tucker, S. C., and Casadevall, A. (2002). Replication of Cryptococcus neoformans in macrophages is accompanied by phagosomal permeabilization and accumulation of vesicles containing polysaccharide in the cytoplasm. Proc. Natl. Acad. Sci. U.S.A. 99, 3165–3170. doi: 10.1073/pnas.052702799
Urb, M., Pouliot, P., Gravelat, F. N., Olivier, M., and Sheppard, D. C. (2009). Aspergillus fumigatus induces immunoglobulin E-independent mast cell degranulation. J. Infect. Dis. 200, 464–472. doi: 10.1086/600070
Vecchiarelli, A., Retini, C., Pietrella, D., Monari, C., Tascini, C., Beccari, T., et al. (1995). Downregulation by cryptococcal polysaccharide of tumor necrosis factor alpha and interleukin-1 beta secretion from human monocytes. Infect. Immun. 63, 2919–2923.
Ventura Aguiar, P., Lopes, V., Martins, L. S., Santos, J., Almeida, M., Pedroso, S., et al. (2014). Cryptococcal infection in non-HIV immunosuppressed patients - Three case reports in a nephrology setting. Med. Mycol. Case Rep. 3, 14–16. doi: 10.1016/j.mmcr.2013.11.003
Wolf, J. M., Espadas-Moreno, J., Luque-Garcia, J. L., and Casadevall, A. (2014). Interaction of Cryptococcus neoformans extracellular vesicles with the cell wall. Eukaryot. Cell 13, 1484–1493. doi: 10.1128/EC.00111-14
Yang, Z., and Marshall, J. S. (2009). Zymosan treatment of mouse mast cells enhances dectin-1 expression and induces dectin-1-dependent reactive oxygen species (ROS) generation. Immunobiology 214, 321–330. doi: 10.1016/j.imbio.2008.09.002
Zaragoza, O., Chrisman, C. J., Castelli, M. V., Frases, S., Cuenca-Estrella, M., Rodríguez-Tudela, J. L., et al. (2008). Capsule enlargement in Cryptococcus neoformans confers resistance to oxidative stress suggesting a mechanism for intracellular survival. Cell Microbiol. 10, 2043–2057. doi: 10.1111/j.1462-5822.2008.01186.x
Keywords: innate immunity, Cryptococcus neoformans, mast cells, monocytes, monocyte chemoattractant protein 1/CCL-2, fungi
Citation: Lopes JP, Stylianou M, Backman E, Holmberg S, Ekoff M, Nilsson G and Urban CF (2019) Cryptococcus neoformans Induces MCP-1 Release and Delays the Death of Human Mast Cells. Front. Cell. Infect. Microbiol. 9:289. doi: 10.3389/fcimb.2019.00289
Received: 02 April 2019; Accepted: 26 July 2019;
Published: 13 August 2019.
Edited by:
Rebecca Drummond, University of Birmingham, United KingdomReviewed by:
Carolina Coelho, University of Exeter, United KingdomAlexandre Alanio, Pariss Diderot University, France
Copyright © 2019 Lopes, Stylianou, Backman, Holmberg, Ekoff, Nilsson and Urban. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: José Pedro Lopes, cGVkcm8ubG9wZXNAdW11LnNl; Constantin F. Urban, Y29uc3RhbnRpbi51cmJhbkB1bXUuc2U=