 Guilherme Loss1†
Guilherme Loss1† Patricia Martins Simões2,3†
Patricia Martins Simões2,3† Florent Valour3,4
Florent Valour3,4 Marina Farrel Cortês5
Marina Farrel Cortês5 Luiz Gonzaga1
Luiz Gonzaga1 Marine Bergot3
Marine Bergot3 Sophie Trouillet-Assant6
Sophie Trouillet-Assant6 Jêrome Josse3
Jêrome Josse3 Alan Diot3
Alan Diot3 Emiliano Ricci3
Emiliano Ricci3 Ana Tereza Vasconcelos1
Ana Tereza Vasconcelos1 Frédéric Laurent2,3*
Frédéric Laurent2,3*- 1Laboratório Nacional de Computação Científica, Rio de Janeiro, Brazil
- 2National Reference Center for Staphylococci - Hospices Civils de Lyon, IAI-Department of Clinical Microbiology, Northern Hospital Group, Lyon, France
- 3Centre International de Recherche en Infectiologie (CIRI), Lyon, France
- 4Hospices Civils de Lyon, Infectious Diseases Department, Northern Hospital Group, Lyon, France
- 5Institute of Microbiology Professor Paulo de Góes, Federal University of Rio de Janeiro, Rio de Janeiro, Brazil
- 6Hospices Civils de Lyon, Joint Research Unit HCL-BioMerieux, Centre Hospitalier Lyon Sud, Pierre-Benite, France
Small colony variants (SCV) of Staphylococcus aureus have been reported as implicated in chronic infections. Here, we investigated the genomic and transcriptomic changes involved in the evolution from a wild-type to a SCV from in a patient with prosthetic joint infection relapse. The SCV presented a stable phenotype with no classical auxotrophy and the emergence of rifampicin resistance. Whole Genome Sequencing (WGS) analysis showed only the loss of a 42.5 kb phage and 3 deletions, among which one targeting the rpoB gene, known to be the target of rifampicin and to be associated to SCV formation in the context of a constitutively active stringent response. Transcriptomic analysis highlighted a specific signature in the SCV strain including a complex, multi-level strategy of survival and adaptation to chronicity within the host including a protection from the inflammatory response, an evasion of the immune response, a constitutively activated stringent response and a scavenging of iron sources.
Introduction
Staphylococcus aureus is among the most important human pathogens, associated with a wide variety of manifestations, ranging from benign superficial skin infections to life-threatening conditions, such as endocarditis, necrotizing pneumonia, and sepsis (Tong et al., 2015). Within the plethora of more severe conditions caused by S. aureus, bone and joint infections (BJIs) are caused by the invasion and progressive destruction of bone tissues and cartilages, mostly targeting long bones and native or prosthetic joints. Although BJI-related mortality is low, the development of these infections is burdened by a high morbidity with poor long-term functional outcome.
Despite the combination of an adequate surgical management with a prolonged course of antimicrobial chemotherapy, S. aureus BJIs are associated with a high propensity of chronicization and relapse (Osmon et al., 2013; Tande et al., 2014). Indeed, many clinical observational studies have demonstrated that S. aureus BJIs can persist asymptomatically before relapse occurs months or years after the initial episode, even in immunocompetent hosts (Greer and Rosenberg, 1993; Bosse et al., 2005; Stevens et al., 2007).
This capacity of S. aureus to persist and remain clinically quite indolent (Rogers et al., 2009) relies on alternative lifestyles adopted by the bacteria to protect itself from an inhospitable environment (Proctor et al., 2014). Among these lifestyles, the Small Colony Variants (SCV) phenotype can potentially be responsible for the chronicization of the infection (Garzoni and Kelley, 2009; Tande et al., 2014; Kahl et al., 2016), offering a better resistance to the challenge posed by the host defenses (Abel et al., 2011; Ou et al., 2016) and to antibiotic therapies (von Eiff et al., 1998; Massey et al., 2001; Massey and Peacock, 2002).
These SCVs variants constitute a naturally quasi-dormant, slow-growing subpopulation of bacteria. They harbor distinctive phenotypic and pathogenic traits such as formation of smaller colonies on agar plates, a quiescent metabolism, reduced haemolytic and coagulase activities, decreased carbohydrate utilization, low virulence potential, and elevated antibiotic resistance (Proctor et al., 2006; Johns et al., 2015). Recent work has reported that external environmental stress factors (reactive oxygen species, low pH, cationic peptides, and limited nutrition) can trigger the emergence of SCVs (Tuchscherr et al., 2010, 2011; Bui et al., 2015).
Environmental stressors induce the emergence of phenotypically distinct types of SCVs. Most of them represent a transient state that can, under favorable conditions, revert to a wild-type (WT) or a different phenotype (distinct from both the progenitor and the SCV), thus leading to the relapse that can be clinically observed. Others are due to permanent genetic changes, and are consequently stable, irreversible SCVs (von Eiff et al., 1998; Massey et al., 2001; Proctor et al., 2006; Tuchscherr et al., 2011; Onyango et al., 2013).
While phenotypic reversion occurs rapidly and is thought to circumvent any lasting fitness cost, permanent genetic alteration seems to depend largely on the nature of the environmental stress. Nevertheless, numbers of genetic changes are prevalent in SCV and related to auxotrophies, such as hemin, menadione, thiamine, thymidine, and/or unsaturated fatty acids (Proctor et al., 1995; Kohler et al., 2003; Seggewiss et al., 2006; Tuchscherr et al., 2011). More recently, these genetic changes have been associated with the persistent active stringent response coupled to relA and rpoB mutations (Gao et al., 2010). Novel mechanisms for the development of persistent organisms and SCVs have been reported in the last years and include changes in RNA processing, stringent response, toxin-antitoxin systems, ribosome protein L6 (RplF), or cold shock protein B (CspB) (for a review see: Proctor et al., 2014).
The detection and identification of SCVs in routine laboratories and their accurate studies in research laboratories are challenges not overcome yet. Indeed, the majority of studies conducted on SCVs use laboratory-generated mutant S. aureus strains rather than naturally derived SCVs. Recently Bui et al. published the first genomic comparison and transcriptomic analysis of a clinical “wild type” S. aureus isolate with its corresponding stable SCV. The latter was induced experimentally by a prolonged exposure of the parental form to a simulated inflammatory environment (methylglyoxal) and a steady-state growth conditions with low nutrients (Bui and Kidd, 2015). However, as for many previous works, the main drawback of this work is the lack of comparison between clinical acute and relapse/chronic SCV forms isolated from the same patient. Moreover, recent publications have revealed a number of mutations that result in SCVs, but most of these have not been studied in clinical isolates.
Here, we provide the first detailed description of the genetic and transcriptomic changes involved in the evolution of a natural wild type isolate into a stable SCV isolate in the clinical context of a relapsing prosthetic joint infection (PJI).
Results and Discussion
Isolation of S. aureus From Acute and Relapse Episodes of a Prosthetic Joint Infection: Clinical History, Antibiogram, and First Characterization
Two S. aureus clinical isolates were obtained from a patient who benefited from the implantation of a left total knee arthroplasty (TKA), see Figure 1. Six years after the implantation, the patient presented an acute haematogenous Streptococcus dysgalactiae PJI which, after an attempt for a conservative treatment, demonstrated signs of a persistent infection. TKA re-implantation occurred in March 2004 with an uneventful immediate postoperative course. Ten months later, the patient was admitted to the emergency department for acute fever associated with a painful and swollen left knee. All preoperative bacteriological samples yielded growth of a multi-susceptible Staphylococcus aureus (WT, isolate ST20130944). Treatment was interrupted in October 2005 after a 7 month course of antimicrobial therapy. However, the patient developed PJI relapse in June 2006, with no general symptoms. All the bacteriological samples made at the time of the surgery were positive to both S. dysgalactiae and S. aureus. The latter demonstrated the characteristics of a stable small colony variant (SCV, isolate ST20130945), as shown in Figure 1.
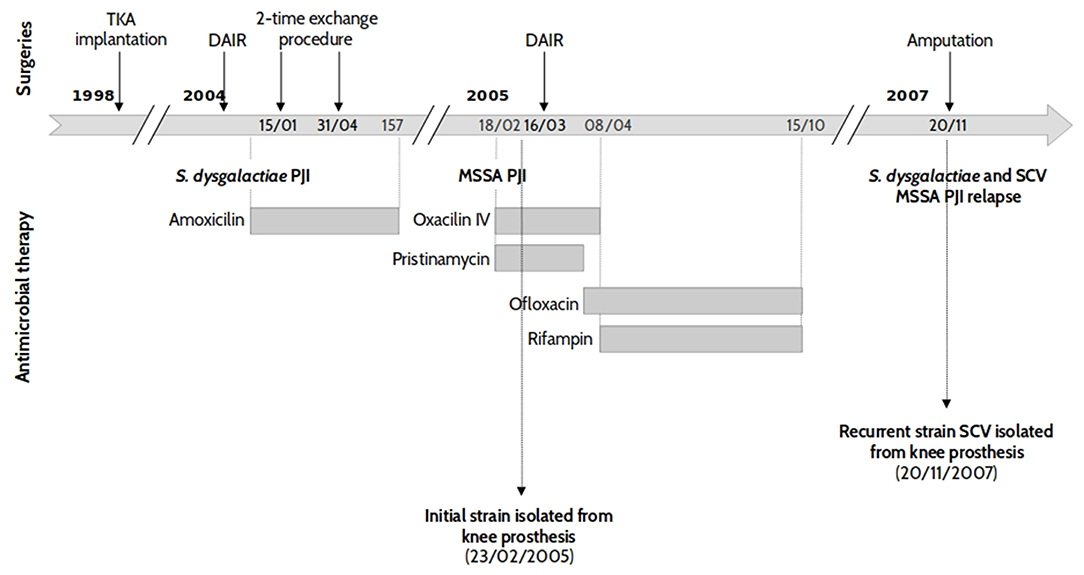
Figure 1. Patient history case and chronology of isolate collection. Two strains of S. aureus (SA) sensitive to methicillin (MSSA) were isolated from an 83 year-old patient with left knee prosthetic joint infection. The first isolate (strain ST20130944) was collected during the first, acute SA infection, while the second was obtained 2 years and 9 months later during a recurrent episode.
The S. aureus isolates from both the initial and relapse episodes were isolated from the same site of infections (knee prosthesis) and were, initially, characterized by DNA microarray and Protein A gene (spa) typing. Both strains presented the same DNA microarray profile with an assignment to CC30-MSSA [PVL-] and spa typing t12. These results suggest that the same S. aureus strain was responsible for both PJI episodes which were separated by 2 years and 9 months. Routine susceptibility tests were also performed to check if novel resistance phenotypes had emerged. Both the initial WT-strain (ST20130944) and relapse-SCV (ST20130945) isolates showed resistance to penicillin and tetracycline. The relapse SCV strain presented an additional resistance to rifampin.
Exploration of the SCV Phenotype
Several morphological differences have been described for SCVs variants including haemolytic irregularities, pigmentation, variation in colony size, slower growth rate, and differences in cell wall composition and thickness (Baddour and Christensen, 1987; Kahl, 2003; Onyango et al., 2013). In our case, both initial-WT and relapse-SCV strains were haemolytic and pigmented (Figure 2A). Regarding the colony size, the relapse-SCV isolate was 8-fold smaller than the initial isolate with a mean colony area ± standard deviation of 0.46 ± 0.1 mm2 vs. 3.8 ± 0.9 mm2 (p ≤ 0.01). Growth rate experiments demonstrated that the relapse-SCV isolate grows slower than the initial-WT isolate with a 2-fold increased doubling time (mean doubling time ± standard deviation of 83.7 ± 3.1 min vs. 39.5 ± 0.3 min, p ≤ 0.01, respectively). This difference is mainly due to the lag phase of the stable SCV strain which is 2 h longer than that of the initial, WT strain (data not shown).
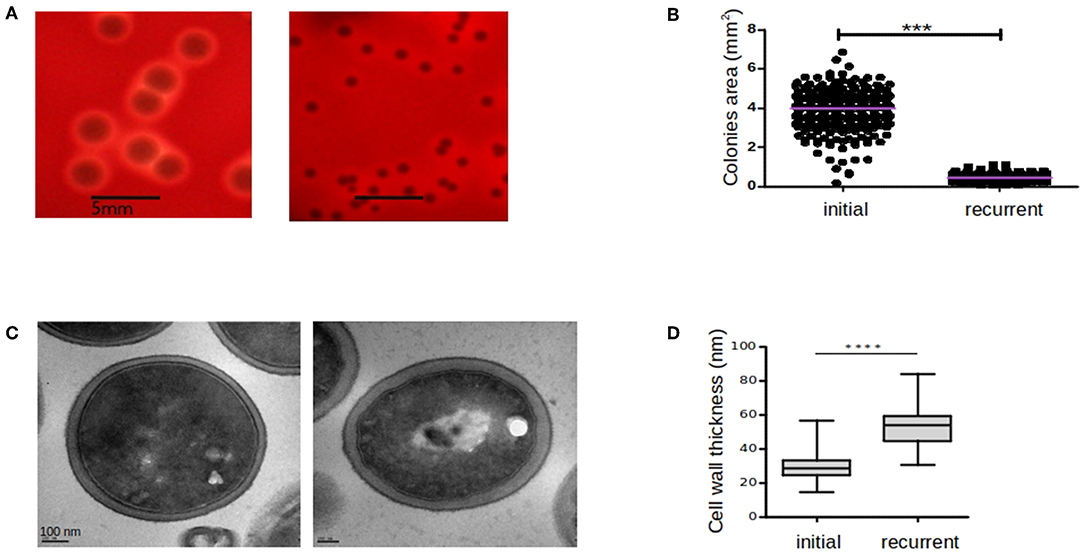
Figure 2. Comparison of colony sizes, hemolytic activities, and cell wall thickness between initial and recurrent infections. The initial-WT (ST20130944) and recurrent-SCV (ST20130945) strains were cultured on blood agar plates and hemolytic activity (A) and colony areas (B) were assessed after 48 h incubation at 37°C. Cell-wall thickness of ST20130944 (left hand-side) and ST20130945 (right hand-side) strain photographed by TEM (C) and comparison of cell-wall thickness of initial and recurrent strains (D). Colony areas (mm2) were measured among three independent experiments and compared using the non-parametric exact Wilcoxon-Mann-Whitney test with alpha < 0.05. Thickness measurements (mm) were taken twice per cell in 30 cells per condition and compared using the non-parametric exact Wilcoxon-Mann-Whitney test with alpha < 0.05. Horizontal bars denote the 1st, 2nd (median), and 3rd quartiles. ***p < 0.001 and ****p < 0.0001.
Cell wall thickness of both the initial-WT and relapse-SCV strains, measured using transmission electron microscopy, revealed that the relapse-SCV strain had a significantly thicker cell wall (mean cell wall thickness ± standard deviation = 53.78 nm ± 1.4) compared to the initial-WT phenotype (29.63 nm ± 0.9, p ≤ 0.001) (Figure 2B). Together these results corroborate the SCV phenotype of the relapse isolate.
Experiments to highlight auxotrophy for menadione, hemin, NAD+ or thymidine (Kahl, 2003, 2005; Proctor et al., 2006; Sendi and Proctor, 2009) were negative. No reversion of the SCV phenotype to the normal phenotype was observed when the relapse-SCV isolate was cultured in the presence of supplemented disks.
Genomic Comparison of Initial-WT and Stable Relapse- SCV S. aureus Isolates
Once the genetic backgrounds of both WT and SCV isolates were confirmed to be identical by DNA microarray analysis and spa typing, whole genome sequencing of both isolates was performed to assess if the SCV phenotype can be related to genetic differences. The fully closed chromosome was obtained for both the initial-WT and relapse-SCV clinical isolates (Table 1). In silico MLST confirmed assignation of both strains to ST30 (CC30) and confirmed the presence of an identical virulence profile (Table S1).

Table 1. SMRT mapping statistical summary (Post-filtering).
The phenotypic resistance to penicillin and tetracycline was corroborated by the presence of (i) a conjugative Tn916-like transposon, in both initial-WT and stable relapse-SCV strains, which carries the tetM gene coding for the resistance to tetracycline; (ii) a Tn552 transposon harboring the BlaI, BlaR, and BlaZ components of the inducible S. aureus β-lactamase (bla operon); and (iii) a smaller non-conjugative Tn3-like transposon which carries a second tetracycline (tet) resistance gene inserted at the end of an integrated plasmid which confers resistance to the heavy metals arsenic and cadmium.
Presence/Absence of Genes
A striking difference between the initial-WT and relapse-SCV isolates was the loss of a ~42.5 kb prophage in the genome of the stable relapse-SCV strain. Detailed analysis revealed that it was a φSa2 which shares some homology to the 46 kb φSa2 (MRSA252) (Holden et al., 2004) prophage. Like its MRSA252 counterpart, this prophage is inserted immediately downstream from a conserved lipoprotein coding gene and it does not contain any obvious virulence determinants either than a second copy of the enterotoxin A gene (sea), see Table S1.
Detection of Nucleotide Variants
To assess the integral genomic differences between the initial-WT and the relapse-SCV, we further compared the genomes of both strains by mapping the reads of the relapse-SCV isolate against the closed genome of the initial-WT isolate. This allows for the identification of single nucleotide polymorphisms (SNP) and small insertions or deletions (INDELs). Only 5 SNPs were identified, of which 2 were intergenic and 3 were intragenic, with only 1 introducing a silent mutation. The remaining 2 intragenic SNP concerned 2 distinct hypothetical proteins (Table 2). Small deletions or insertions were observed in the SCV isolate (Table 2). Small INDELs (1 bp) inducing truncated proteins, due to a frameshift change, were observed in sasA (coding for the surface protein A) which plays a role in the adhesion to host cells, and in glpD coding for an aerobic glycerol-3-phosphate dehydrogenase, which participates in biosynthetic pathways of secondary metabolites and in iron metabolism (Haley and Skaar, 2012). Deletions leading to the suppression of amino acids without the emergence of truncated protein forms were observed for an alpha-beta hydrolase (9 bp INDEL) and a putative serine protease (6 bp INDEL). More interestingly, a 9 bp (3 aa, LysGlyPro488−490) non-truncating deletion was found within the rpoB gene encoding a DNA-dependent RNA polymerase which is the target of rifampin. Multiple SNP mutations in the rpoB gene have been previously described and associated to an emergence of resistance to rifampicin (Wichelhaus et al., 2002; O'Neill et al., 2006; Cui et al., 2010; Gao et al., 2013). This previously unreported mutation is coherent with the resistant profile observed in the recurrent, stable SCV isolate. Moreover, a point mutation (H481Y) in the RpoB protein has already been reported to play a major role on virulence, persistence, bacterial gene expression, and innate immune evasion of a clinical, stable SCV isolate (Gao et al., 2013). However, this point mutation in rpoB co-occurs with a point mutation in the relA gene (Gao et al., 2010, 2013), a key player in the synthesis pathway of the intracellular signaling molecule guanosine 3′,5′ bis(diphosphate), i.e., the alarmone ppGpp involved in the stringent response (Godfrey et al., 2002; Dalebroux et al., 2010; Wolz et al., 2010). Although we did not detect any mutation in neither relA gene nor in the two other small (p)ppGpp synthetases reported in S. aureus (relP and relQ) at the genomic level, only the point mutation in rpoB gene could explain the SCV phenotype of our recurrent strain.
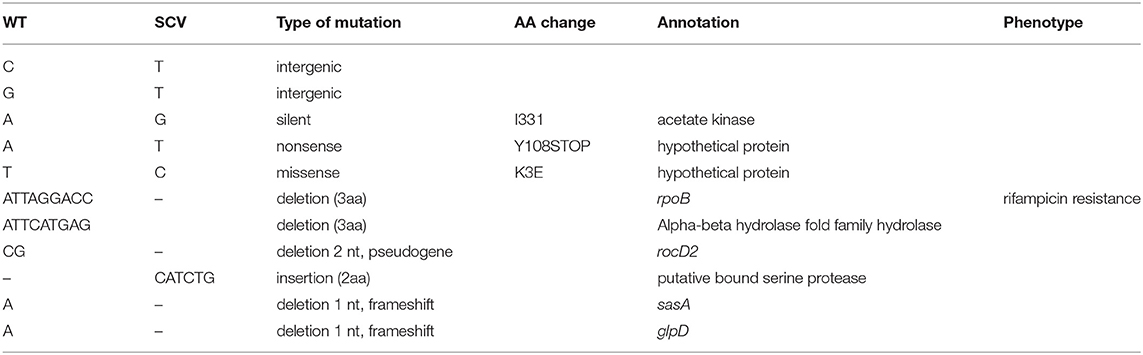
Table 2. Nucleotide variations observed between initial, WT, and recurrent, stable SCV strains.
Methylome
We pursued our analysis by comparing the methylome of the initial-WT and stable relapse-SCV strains to check if the SCV phenotype could be correlated with an epigenetic regulation of gene expression.
Out of a total of 1195 N6-Methyladenosine m6A motifs detected, 19 m6A methylated motifs were distinct between the initial-WT and the stable relapse-SCV (1.5% of differences in methylation motifs) with the initial-WT having all 19 motifs methylated while the relapse-SCV strain had none of the 19 motifs methylated. One of these motifs was located in an intergenic region and 7 methylations were found to be located in phage proteins located within the φSa2 region exclusively found in the initial-WT strain. The remainder 12 differentially methylated regions concerned genes coding for transposases (n = 2), exported proteins (n = 2), membrane proteins (n = 2), yolD family protein (n = 1), penicillinase biosynthesis (n = 1), thioredoxin, an important antioxidant protein (n = 1), a thiamine-phosphate synthase related to thiamine/vitamin B1 biosynthetic process (n = 1) and the translation factor SUA5 which is associated with double-stranded RNA binding biological pathways (n = 1), see Table S2.
Despite a differential pattern in the methylation status of the thiamine-phosphate synthase, no auxotrophism was observed in the stable SCV isolate. This seems to suggest that the observed methylation status does not play an essential role in the stable SCV phenotype.
Nonetheless, considering these differential methylation patterns, in particular, those affecting the response to oxidative stress, the vitamin B1 pathway and the double-stranded RNA pathways, we next thought to confirm if these methylation patterns correlated with changes in overall transcript expression of the SCV isolate.
Transcriptomic Comparison of Initial WT and Stable SCV Relapse S. aureus Isolates
Messenger RNA sequencing revealed 101 genes strongly down-regulated (Log Fold Change < −2) and 61 strongly up-regulated (Log Fold Change > 2) (see Table S3).
Down Regulated Genes
The most strongly down-regulated genes are involved in the arginine and ornithine catabolism [arginine deiminase (ADI) pathway]. This pathway permits S. aureus to get energy from arginine sources in glucose-free and oxygen-free conditions and leads to the production of ammonia (Makhlin et al., 2007). Other metabolic pathways such as the pyrimidine metabolism pathway, alternative carbon sources transport (phosphotransferase system PTS), carbon catabolism via the glycolytic pathway, tricarboxilic acid (TCA) pathway or the pentose phosphate pathway are also downregulated in relapse-SCV isolate compared to the initial-WT (Figure 3 and Table S3). Down-regulation of these various energy pathways may lead to ATP depletion, which have been reported in persister formation in S. aureus (Conlon et al., 2016). Other important down-regulated genes are involved in the cell wall regulon vraRS and the operon vraABCX, an ABC transport system under vraRS control. Modulation of vraRS expression can impact the emergence of cell-wall active antibiotic resistance in S. aureus (Galbusera et al., 2011;McCallum et al., 2011).
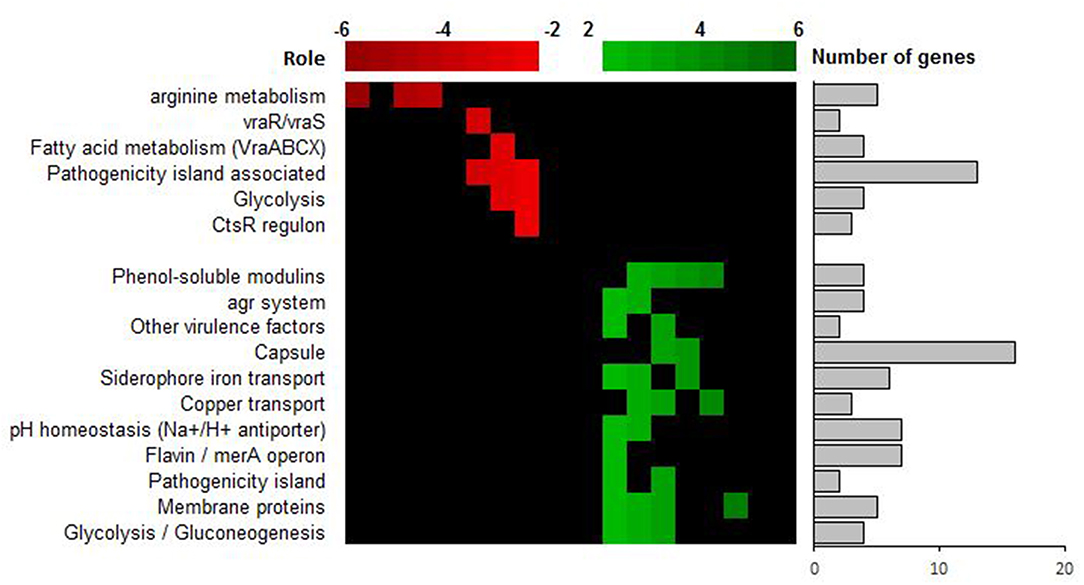
Figure 3. Transcriptional analysis of SCV strain ST20130945 and parental strain ST20130944. Up-regulated genes (in green) are differentially up-regulated in ST20130945 compared to the parent strain ST20130944, and the down-regulated genes (in red) are differentially down-regulated in ST20130945 compared to ST20130944. The heat map analysis highlights the role of the genes for which the differential expression was highest.
Down-regulation of energy pathway and vraRS and is in line with the emergence of the SCV phenotype and the switch from acute to chronic and persistence phenotype observed in clinical isolates.
Up-regulated Genes
The most surprising result corresponds to an upregulation of the agr locus and agr-regulated virulence genes such as delta-toxin (hld), PSM alpha 1 to 3, this transcriptomic profile was validated in vitro by an increased cytotoxicity for relapse-SCV compared to initial-WT isolate in an osteoblastic cytotoxicity test (Figure S2). Indeed, such pattern is classically associated with virulence and acute infections due to high release of staphylococcal toxins. In previous studies, we reported that intracellular release of PSM favors the cytotoxicity and that deficiency of delta-toxin is correlated with the chronicity of BJI (Rasigade et al., 2013; Valour et al., 2015). This observed up-regulation of virulence factors is in contradiction with the accepted paradigm of SCV colonies formation as quasi-dormant, low virulent subpopulations of bacteria.
Of note, we observed a concomitant up-regulation of genes related to the capsule formation (cap operon). Similar up-regulation of the cap operon (biofilm formation), the agr and alpha PSMs has already been reported in a clinical SCV strain, when compared to its WT, parental strain, and it was demonstrated that transcriptomic changes were due to a constitutively activated stringent response (Gao et al., 2013).
This high expression of phenol soluble modulins (PSM alpha and delta-hemolysin) was observed throughout the mid-exponential and (early and late) stationary phase of the stable SCV strain which further emphasizes the hypothesis of a permanently active stringent response (Figure S3). This is in contrast with the “classical” expression profile observed for the WT strain which has a high RNA III (regulatory mRNA transcript encoding the delta-toxin and part of the agr locus) during the exponential phase and a decreased expression on the stationary phase.
The transcriptomic pattern of the stable, SCV strain is in agreement with the 9 bp mutation in rpoB, the relapse-SCV phenotype and permanent stringent stress response due to a multitude of different environmental stress conditions. Moreover, it has also been demonstrated that the stringent response is activated upon internalization of S. aureus in human polymorphonuclear neutrophils (PMNs) with the intracellular psm expression contributing to survival after phagocytosis (Geiger et al., 2012).
The high expression of virulence factors by the stable-SCV strain is interesting as this strain was isolated in a context of a relapsing infection. The expression profile of virulence genes found in the stable-SCV strain is typically associated with S. aureus strains isolated from acute infections. Our results and the stability of the SCV phenotype observed for the relapse isolate might be associated to the “reactivation” status of a bacterial strain which has adapted to evade the host's immune response via a genetic imprinting (rpoB mutation) that lead to a permanent phenotype change (SCV phenotype) without compromising the strain's virulent “potential” (expression of toxins like PSM alpha, delta toxin and its major regulator, the agr locus). Moreover, the over expression of PSM seems pertinent as: (1) on one side they allow S. aureus to evade neutrophils destruction, which are the first professional phagocytic cells to be recruited into the site of bone infections; (2) on the other side, if S. aureus has persisted indolent via an intracellular lifestyle within osteoblast, it allows the destruction of its cellular host and the release of S. aureus into the extracellular bone and joint environment where it can pursue the destruction of bone tissue characteristic of BJIs. This dual potential (virulence vs. persisting, indolent status) might be achieved by a permanent activation and regulation of the stringent.
Other up-regulated genes include genes coding for ABC transporters and a multitude of genes related to the detoxification of oxidative stress associated reactive electrophilic species (RES), such as glyoxal and methylglyoxal, and/or reactive oxygen species (ROS) as well as cell wall associated lipoteichoic acid and lipid biosynthesis, glycolysis/gluconeogenesis and pH homeostasis (Figure 3 and Table S3). Again, this is interesting as (i) ABC transporters play a major role in nutrient acquisition and adaptation to rapidly changing host micro-environments (metal scavenging, amino-acids scavenging and detoxification) and (ii) reactive electrophilic species such as glyoxal and methylglyoxal are toxic chemical stressors present in the host-pathogen milieu due, in part, to the oxidative burst of activated macrophages and neutrophils and/or cellular metabolism (methylglyoxal is a highly toxic, reactive aldehyde produced as a by-product from triose-phosphate intermediates during glycolysis) (Ferguson et al., 1998; Beavers and Skaar, 2016; Imber et al., 2018). The concomitant over-expression of both ABC transporters and other detoxification genes suggests a rapid adaptation to a reactive immune system and/or changes in the pathogen's micro-environment (pH homeostasis, high concentration of toxic chemical stressors in the site of infection).
The absence of a significant up or down-regulation in the expression levels of the differentially methylated genes, in the relapse-SCV isolate, seems to suggest that the slight difference observed in the methylome of the initial-WT and relapse-SCV strains might be due to a more important role of methylation in the recognition of self and non-self DNA than in gene regulation in the evolution from acute to chronic and persistent S. aureus infections.
The analysis of the expression of small non-coding RNAs (ncRNA) revealed a Log Fold change (LFC) increase of 13 for the expression of the sprG1 ncRNA, the toxin of class I toxin-antitoxin (TA) system, in the stable SCV (Table S4) The activity of type I TA toxins, most of which are small hydrophobic proteins corrupting cell envelopes, is controlled by an RNA antitoxin that inhibits translation and/or promotes degradation of toxin mRNA (Schuster and Bertram, 2016). It has been shown that sprG1, when over-expressed in S. aureus strain N315, causes cell death of S. aureus but also of competing bacteria and, most importantly, of erythrocytes (Pinel-Marie et al., 2014). This is interesting as the overexpression of sprG1 is concomitant with the up regulation of high affinity iron recovery siderophores (aerobactin iron transport system, see Figure 3 and Table S3). This seems to point to a highly fine-tuned regulation of iron uptake via a dual mechanism of (1) erythrocytes' killing and (2) rescuing of newly released iron using high affinity siderophores. Iron is one of the major enzymatic co-factors in biological systems. Its acquisition is a critical determinant in staphylococcal pathogenesis and restriction to iron access is one of the host's mechanisms to resist to infection and improve its own inflammatory response (Bullen et al., 2005; Wessling-Resnick, 2010; Haley and Skaar, 2012). The ability of our relapse-SCV isolate to evade the host strategies of iron sequestration might be of major importance for its persistence within the host and evasion of the host immune system.
Conclusion
Our stable, relapse-SCV isolate is a non-classical SCV that seems to have adopted a complex, multi-level strategy of survival and adaptation to chronicity and/or reactivation within the host. Its phenotype seems to be independent from the previously described hemin, thymidine or menadione auxotrophisms and dependent from an activated stringent response. The most obvious reason for the persistence of the strain is its rpoB 9bp mutation that correlates perfectly with phenotypic resistance to rifampicin and might also be responsible for a permanent activation of the stringent response, via its messenger molecule (p)ppGpp, even if we did not identify any mutation in relA or the other known (p)ppGpp synthetases (relP and relQ).
Thus, our clinical, stable relapse-SCV isolate seems to protect itself from the inflammatory response/oxidative stress by a plethora of pathways, including (i) RES and ROS reduction, riboflavin and NADH+ formation, osmoprotectants, chaperones and ABC transporters over-expression and reduced metabolism; (ii) it over expresses alpha PSM and capsule biosynthesis genes which are known to favor evasion from the immune response by impairing phagocytosis and reducing phagocytic killing; (iii) it seems to have a constitutively activated stringent response with inhibited DNA replication and a post-transcriptional inhibition via decreased ribosomal assembly and consequent inhibition of cell growth; and (iv) a down-regulation of the cell wall regulon VraRS associated to resistance to cell wall disrupting antibiotics and human cationic antimicrobial peptides (CAMPs) (Pietiäinen et al., 2009). Put together, our data strongly suggests an adaptation to chronicity, where the host immune response and antibiotic pressure are a permanent environmental stress leading to the permanent activation of the stringent response.
To further elucidate these in silico results more work is underway to pinpoint the modulator(s) in the stringent response pathway that are behind this stable SCV phenotype.
Methods
Ethics Statement
The clinical portion of this study was conducted with the approval of the French SouthEast ethics committee (no. CAL201121 for clinical isolates). In accordance with the French legislation, written informed patient consent was not required for any part of the study.
Bacterial Strains Characterization and Growth Conditions
The two clinical Staphylococcus aureus isolates, collected from the acute phase [wildtype (WT) isolate ST 20130944] and the chronic phase [stable small-colony-variant (SCV) isolate ST20130945] of a BJI in the same patient, were stored at −80°C in cryotubes. Prior to all experiments, the isolates were first sub-cultivated on Columbia agar supplemented with 5% sheep blood (COS; BioMérieux) at 37°C for 24 h after thawing.
To confirm that the same bacteria was at the origin of the first, acute event and the second, relapsing event, both clinical isolates were genotyped using diagnostic DNA microarrays, identibac S. aureus Genotyping (Alere) used for this study, as well as related procedures and protocols, have been previously described in detail (Monecke et al., 2009). The assigning of isolates to CCs was determined by the comparison of hybridization profiles with those previously characterized by using multilocus sequence typing reference strains (Monecke et al., 2009). S. aureus spa-typing (Staphylococcus aureus Protein A) was also performed as previously described (Jarraud et al., 2002).
Antimicrobial Susceptibility Testing (AST)
The AST profiles of the isolates were performed using the standard agar diffusion technique following the recommendations of the French Committee for Antimicrobial Susceptibility Testing (CA-SFM). MICs were interpreted in terms of susceptibility according to the 2016 recommendations of the EUCAST. The Minimal inhibitory concentrations for rifampin were confirmed by Etest as recommended by manufacturers.
Colony Size Measurements
The two clinical S. aureus isolates (WT and SCV) were subcultivated in BHI incubated for 16 h at 36°C. After a dilution in fresh BHI of the overnight cultures to OD600 = 0.5, 100 microL of serial dilutions were inoculated in both Trypticase Soy Agar (TSA) and COS plates. After overnight incubation at 36°C, the plates were photographed, and the wild-type and SCV colonies were analyzed using an automated process in which a high-resolution picture of each plate was taken and analyzed by means of the image analysis software ImageJ (W. S. Rasband, National Institutes of Health, Bethesda, MD) (Cai et al., 2011) with a customized macro involving color thresholding, watershed algorithm, and particle analysis, in order to extract the distribution of colony areas. The WT colony area was defined as the median area of all colonies (because the median is robust to outliers, the presence of SCVs did not influence significantly this measure) while the SCVs colonies were defined as colonies with an area less than one-fifth of the median area according to the usual operational definition. Experiments were performed in triplicate.
Auxotrophy Complementation Test
Disk diffusion assays were used to determine the auxotrophic phenotype of the stable SCV isolate for hemin, NAD, thymidine, menadione using an adaptation of the protocol previously described by Kahl et al. (1998). Briefly, colonies sub-cultivated overnight in COS were resuspended in a phosphate buffered saline (PBS, pH = 7.4) solution to 0.5 McF and 200 MicroL of bacterial suspension were spread on either Mueller-Hinton (MH) agar (BioMérieux) or Triptone/casein/Peptone (COS) (BioMérieux) media. For hemin and NAD/factor auxotrophy factor X, V and X+ V, standard disks (Sigma Aldrich, Germany) were used. For thymidine and menadione (Sigma Aldrich, Germany) auxotrophy testing disks were prepared by impregnation of 15 mL of thymidine at 100 mg/mL or menadione at 10 mg/mL (Sigma Aldrich, Germany). To determine single auxotrophism, impregnated disks were put on the MH agar surface with an inoculum of SCV isolate similar to the inoculum used for AST. auxotrophism was determined as positive if a zone of normal growth surrounding the impregnated disks was detected after 1–24 h of incubation.
Growth Rate Determination
For the characterization of the growth dynamics of both wild type and stable SCV S. aureus isolates, a starter culture was prepared by inoculating colonies into BHI grown for 16 h at 36°C. Growth dynamics was performed in fresh brain-heart infusion (BHI) medium at 36°C, after dilution of overnight cultures to OD600 = 1.0. A thermostated microplate reader (TECAN M200 Infinite Pro) was used to follow bacterial growth by measuring OD600 every 15 min for 24 h. As controls, specific wells were inoculated with medium only. Experiments were performed in triplicate.
Transmission Electron Microscopy
Both WT and stable SCV strains were prepared for TEM as described elsewhere (Gounon, 2002). and photographed by means of a JEOL 1400 microscope (Tokyo, Japan). Cell wall thickness was measured in 32 cells of each sample with nearly equatorially cut surfaces (two measures per cell) by means of the DigitalMicrographTM software (Gatan, Inc.). Cell-wall thicknesses (in nm) were compared using Student's t-test. The significance threshold was set at 0.05. Analyses were performed with GraphPad PRISM (version 5.0.3.477).
Genomic DNA Preparation and Whole Genome Sequencing (WGS)
DNA extractions were performed using the QIACube R automate (Qiagen) according to the manufacturer's recommendations. DNA was eluted in molecular grade water to final concentration ≥200 ng/μL. Ion PGM sequencing—Eight 200-base-read gDNA fragment libraries were constructed using 100 ng of DNA from each sample. The libraries were prepared according to Ion XpressTM Plus gDNA Fragment Library Preparation protocol. For DNA fragmentation it was used Ion ShearTM Plus Reagents. The barcoded libraries were quantified and pooled together in even amounts. Emulsion PCR (emPCR) was carried out in the Ion OneTouchTM 2 System. The libraries were sequenced in one Ion 318TM Chip v2 using an Ion PGMTM System. For read quality assessment FastQC tool (http://www.bioinformatics.babraham.ac.uk/projects/fastqc/) was used and FASTX-Toolkit v0.0.13 (http://hannonlab.cshl.edu/fastx_toolkit/) for trimming. Single-molecule real-time (SMRT) sequencing was also performed to obtain closed genomes for both initial and recurrent strains. DNA extractions were performed using a NucleoBond® PC 100 (Macherey-Nagel) according to the manufacturer's recommendations. Sequencing for de novo assembly was performed using PacBio RS II (Menlo Park, CA, USA). High molecular weight DNA was sheared in a Covaris g-TUBE (Covaris, Woburn, MA, USA) to obtain 20 kb fragments. After shearing, the DNA size distribution was checked using a Fragment Analyzer (Advanced Analytical Technologies, Ames, IA, USA), and 5 μg of the sheared DNA was used to prepare a SMRTbell library with PacBio SMRTbell Template Prep Kit 1 (Pacific Biosciences, Menlo Park, CA, USA) according to the manufacturer's recommendations. The resulting library was size selected using a BluePippin system (Sage Science, Inc. Beverly, MA, USA) for molecules larger than 11 kb. The recovered library was sequenced using 1 SMRT cell with P6-C4 chemistry and MagBeads with a PacBio RSII system (Pacific Biosciences, Menlo Park, CA, USA) at 240 min movie length. SMRT Mapping statistics are reported in Table 1. For de novo assembly, the PacBio module “RS_HGAP_Assembly.2” in SMRTpipeline version v2.3.0 was used for continuous long reads (CLR) after polishing and error correction with Quiver, as described previously (Chin et al., 2013). DNA methylation was determined using the RS_Modification_and_Motif_Analysis protocol within SMRT Portal v2.30, with a standardized in silico false positive error of ~1%. Only motifs with a mean modification quality value (QV) >50 and a mean coverage of >100X were validated as being modified (https://github.com/PacificBiosciences/Bioinformatics-Training/wiki/Methylome-Analysis-Technical-Note). For each strain, one polished contigs corresponding to the chromosome was obtained. Circularization of both contigs was achieved by manual comparison and removal of regions of overlaps.
Assembly and Annotation
The gene predictions were performed using the SABIA (System for Automated Bacterial Integrated Annotation) service (Almeida et al., 2004). Further annotations were manually inspected, then checked also with SABIA.
SNP Analysis
Ion torrent reads with at least 30 nucleotides in length were aligned with bowtie2 (Langmead and Salzberg, 2012) using “very sensitive” preset option, and one mismatch per seed region. The resultant alignment files were filtered with samtools v0.1.19 package (Li et al., 2009; Li, 2011) for mapping quality (MQ) ≥30. The SNP calling was performed with samtools mpileup and GATK (McKenna et al., 2010) tools. The PCR duplicates were next identified and marked using Picard (v. 1.51) MarkDuplicates tool (broadinstitute.github.io/picard/).The samtools mpileup parameters were minimum base quality (-Q) > 7 and skipping INDEL calling with average per-sample depth (-L) > 1,000. This was followed by filtering with bcftools SNP calling, and filtering with vcftools varFilter setting the minimum coverage (-d) of 1 and maximum (-D) of 200 reads. The GATK tool parameters were used following the Best Practice Variant Detection manual (https://www.broadinstitute.org/gatk/guide/best-practices). Three steps of calibration were performed until the number of SNPs called was stabilized. The resulting Variant Call Files (VCFs) were annotated with SnpEff tool V 4.1 (http://snpeff.sourceforge.net/). All SNPs called were manually inspected with the Integrative Genomics Viewer (IGV) tool (Robinson et al., 2011). For the manual validation of SNPs we performed the mapping of reads from the recurrent isolates into the respective initial isolate assembled genome (both SMRT and Ion Torrent reads). SNPs of “high quality” were validated using the following criteria: (1) minimum coverage ≥ 20 reads; (2) ≥75% of the reads supporting the variant call; (3) no INDELS due to homopolimeric regions 5 nts upstream or downstream to the variant call; (4) INDELS in polymeric regions were discarded (due to the limitations of the Ion Torrent technique). Manual validation of INDELS was performed using the cross match tool from phrap-consed package (http://www.phrap.org/phredphrapconsed.html). All INDELS and SNPs predicted in protein coding regions (CDS) were validated by Sanger sequencing, using primers designed especially for the amplification of the polymorphic regions.
Detection of Presence/absence of ORFs
Was performed with the Bidirectional Best Hit (BBH) approach using BLASTp. All BLASTp searches were performed with an e-value cut off of E = 1e-05, query coverage of 90% and positives of 90%. Once the BBH pairs have been determined, we constructed the similarity clusters as follows: A cluster was defined as a set of genes in which every gene has a BBH with at least another element. Manual curation was then performed to validate the presence/absence of ORFs using the Mauve Progressive alignment algorithm (Darling et al., 2010) and the clustering analysis results.
Detection of Mobile Genetic Elements
Distinct tools were used to characterize mobile genetic elements (MGE) such as prophages, transposons and genomic islands: PHAST (Zhou et al., 2011), Issaga (https://www-is.biotoul.fr/). Comparison with known S. aureus genomes showed a very high homology of both ST30 strains with the genome of strain MRSA252 (Table 2 and Figure S1), a well-characterized hospital acquired ST36 (CC30) MRSA strain representative of the highly successful epidemic EMRSA-16 clone (Holden et al., 2004). Manual curation of MGE was performed by comparison with closely related and well-annotated genome of strain MRSA252.
RNA Preparation and Whole Transcriptome Sequencing (RNA-seq)
Overnight bacterial cultures were used to inoculate a BHI broth and incubated at 36°C with gyratory shaking at 200 rpm. Bacteria were harvested after 10 h to reach the post-exponential growth phase. Culture aliquots of 1 mL were centrifuged at 13,000 g, and the pellets were washed with 1 mL of 10 mM Tris buffer and adjusted to an optical density at 600 nm (OD600) of 1, corresponding to approximately 109 S. aureus cells/mL. One mL of adjusted and washed bacterial suspension was centrifuged at 13,000 g, and the pellets were treated with lysostaphin (Sigma-Aldrich) at a final concentration of 200 mg/L. The total RNA of the pellets was then purified using the RNeasy Plus Mini Kit (Qiagen) according to the manufacturer's instructions. Removal of rRNAs was performed using the Ribo Zero kit (Illumina), following the manufacturer's instructions, and each RNA sample was suspended in 30 μL of RNA storage solution. The purity and concentration of each ribosomal depleted RNA was controlled by Bioanalyzer (Agilent) using the RNA Nano Chips, and quantity was assessed using the ND-8000 (NanoDrop Technologies). Complementary RNA libraries were prepared following the previously published protocol (Heyer et al., 2015) and quality control of the DNA colony template library was performed by cloning an aliquot into a TOPO plasmid and sequencing 8/10 clones. Samples were sequenced using a 1 ×50 bp strategy in a Hiseq2500 (Illumina, CA) sequencer. Each RNAseq library was first quality checked with the FASTQC tool (www.bioinformatics.babraham.ac.uk/projects/fastqc/) and further trimmed with Trimmomatic tool (Bolger et al., 2014) with head crop of 7 nt and minimum length of 15 nt. The resulting reads were mapped in respective S. aureus strain with bowtie2 tool (Langmead and Salzberg, 2012) using default parameters, excepted to one mismatch per seed region. The resulting mapping files were processed by HTSeq tool (Anders et al., 2013), in order to extract the raw counts of reads per gene. The Differential gene expression (DGE) analysis were carried out using the edgeR package (Robinson et al., 2010) from R programming language, as described in Anders et al. (2013). Genes were considered as differentially expressed (DEGs) when the following conditions were met: they were statistically significant as indicated by an FDR value lower than 0.05 (FDR is the False Discovery Rate, a correction of the p-value to account for multiple simultaneous tests) and had a change in transcript abundance of at least two (in either direction). Visual inspection of the alignment was performed using the IGV tool (Robinson et al., 2011).
Statistical Analysis
For colony size experiments, cell wall size experiments and cytotoxicity assay, non-parametric exact Wilcoxon-Mann-Whitney tests with alpha < 0.05 were carried out to determine the significance of the results (GraphPad PRISM v 5.0.3.477).
Data Availability Statement
The genomes of the initial WT-strain (ST20130944) and relapse-SCV (ST20130945) were submitted to the NCBI BioProject (http://www.ncbi.nlm.nih.gov/bioproject) under BioProject accessions: PRJNA497221 (strain ST20130945) and PRJNA497214 (strain ST20130944), respectively.
Author Contributions
PS and GL performed all bioinformatic analysis and wrote the main manuscript. FV, JJ, AD, AV, and FL contributed to the main manuscript writing. ER and GL performed the RNA Seq analysis. FV prepared Figure 1. PS, ST-A, MC, and JJ performed the experiments and prepared Figures 2, 3, and Supplementary Data. All authors reviewed the manuscript.
Funding
MB was funded by the Fondation pour la Recherche Medicale (FRM) grant ING20160435683. This work was partially suppoted by Coordination of Superior Level Staff Improvement (CAPES) - Brasil - Finance Code 001. ATRV supported by CNPq #303170/2017-4 and Faperj #E-26/202.826/2018 and GLM CAPES #23038.010041/2013-13.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fcimb.2019.00363/full#supplementary-material
References
Abel, J., Goldmann, O., Ziegler, C., Höltje, C., Smeltzer, M. S., Cheung, A. L., et al. (2011). Staphylococcus aureus evades the extracellular antimicrobial activity of mast cells by promoting its own uptake. J. Innate Immun. 3, 495–507. doi: 10.1159/000327714
Almeida, L. G. P., Paixão, R., Souza, R. C., da Costa, G. C., Almeida, D. F., and de Vasconcelos, A. T. R. de (2004). A new set of bioinformatics tools for genome projects. Genet. Mol. Res. GMR 3, 26–52. Available online at: http://www.funpecrp.com.br/gmr/year2004/vol1-3/SCv0002_full_text.htm
Anders, S., McCarthy, D. J., Chen, Y., Okoniewski, M., Smyth, G. K., Huber, W., et al. (2013). Count-based differential expression analysis of RNA sequencing data using R and Bioconductor. Nat. Protoc. 8, 1765–1786. doi: 10.1038/nprot.2013.099
Baddour, L. M., and Christensen, G. D. (1987). Prosthetic valve endocarditis due to small-colony staphylococcal variants. Rev. Infect. Dis. 9, 1168–1174. doi: 10.1093/clinids/9.6.1168
Beavers, W. N., and Skaar, E. P. (2016). Neutrophil-generated oxidative stress and protein damage in Staphylococcus aureus. Pathog. Dis. 74:ftw060. doi: 10.1093/femspd/ftw060
Bolger, A. M., Lohse, M., and Usadel, B. (2014). Trimmomatic: a flexible trimmer for Illumina sequence data. Bioinformatics 30, 2114–2120. doi: 10.1093/bioinformatics/btu170
Bosse, M. J., Gruber, H. E., and Ramp, W. K. (2005). Internalization of bacteria by osteoblasts in a patient with recurrent, long-term osteomyelitis. A case report. J. Bone Joint Surg. Am. 87, 1343–1347. doi: 10.2106/00004623-200506000-00022
Bui, L. M., Turnidge, J. D., and Kidd, S. P. (2015). The induction of Staphylococcus aureus biofilm formation or Small Colony Variants is a strain-specific response to host-generated chemical stresses. Microbes Infect. 17, 77–82. doi: 10.1016/j.micinf.2014.09.009
Bui, L. M. G., and Kidd, S. P. (2015). A full genomic characterization of the development of a stable Small Colony Variant cell-type by a clinical Staphylococcus aureus strain. Infect. Genet. Evol. 36, 345–355. doi: 10.1016/j.meegid.2015.10.011
Bullen, J. J., Rogers, H. J., Spalding, P. B., and Ward, C. G. (2005). Iron and infection: the heart of the matter. FEMS Immunol. Med. Microbiol. 43, 325–330. doi: 10.1016/j.femsim.2004.11.010
Cai, Z., Chattopadhyay, N., Liu, W. J., Chan, C., Pignol, J.-P., and Reilly, R. M. (2011). Optimized digital counting colonies of clonogenic assays using ImageJ software and customized macros: comparison with manual counting. Int. J. Radiat. Biol. 87, 1135–1146. doi: 10.3109/09553002.2011.622033
Chin, C.-S., Alexander, D. H., Marks, P., Klammer, A. A., Drake, J., Heiner, C., et al. (2013). Nonhybrid, finished microbial genome assemblies from long-read SMRT sequencing data. Nat. Methods 10, 563–569. doi: 10.1038/nmeth.2474
Conlon, B. P., Rowe, S. E., Gandt, A. B., Nuxoll, A. S., Donegan, N. P., Zalis, E. A., et al. (2016). Persister formation in Staphylococcus aureus is associated with ATP depletion. Nat. Microbiol. 1:16051. doi: 10.1038/nmicrobiol.2016.51
Cui, L., Isii, T., Fukuda, M., Ochiai, T., Neoh, H., Camargo, I. L., et al. (2010). An RpoB mutation confers dual heteroresistance to daptomycin and vancomycin in Staphylococcus aureus. Antimicrob. Agents Chemother. 54, 5222–5233. doi: 10.1128/AAC.00437-10
Dalebroux, Z. D., Svensson, S. L., Gaynor, E. C., and Swanson, M. S. (2010). ppGpp conjures bacterial virulence. Microbiol. Mol. Biol. Rev. 74, 171–199. doi: 10.1128/MMBR.00046-09
Darling, A. E., Mau, B., and Perna, N. T. (2010). progressiveMauve: multiple genome alignment with gene gain, loss and rearrangement. PloS ONE 5:e11147. doi: 10.1371/journal.pone.0011147
Ferguson, G. P., Totemeyer, S., MacLean, M. J., and Booth, I. R. (1998). Methylglyoxal production in bacteria: suicide or survival? Arch. Microbiol. 170, 209–218. doi: 10.1007/s002030050635
Galbusera, E., Renzoni, A., Andrey, D. O., Monod, A., Barras, C., Tortora, P., et al. (2011). Site-specific mutation of Staphylococcus aureus VraS reveals a crucial role for the VraR-VraS sensor in the emergence of glycopeptide resistance. Antimicrob. Agents Chemother. 55, 1008–1020. doi: 10.1128/AAC.00720-10
Gao, W., Cameron, D. R., Davies, J. K., Kostoulias, X., Stepnell, J., Tuck, K. L., et al. (2013). The RpoB H481Y rifampicin resistance mutation and an active stringent response reduce virulence and increase resistance to innate immune responses in Staphylococcus aureus. J. Infect. Dis. 207, 929–939. doi: 10.1093/infdis/jis772
Gao, W., Chua, K., Davies, J. K., Newton, H. J., Seemann, T., Harrison, P. F., et al. (2010). Two novel point mutations in clinical Staphylococcus aureus reduce linezolid susceptibility and switch on the stringent response to promote persistent infection. PLoS Pathog. 6:e1000944. doi: 10.1371/journal.ppat.1000944
Garzoni, C., and Kelley, W. L. (2009). Staphylococcus aureus: new evidence for intracellular persistence. Trends Microbiol. 17, 59–65. doi: 10.1016/j.tim.2008.11.005
Geiger, T., Francois, P., Liebeke, M., Fraunholz, M., Goerke, C., Krismer, B., et al. (2012). The stringent response of Staphylococcus aureus and Its Impact on Survival after phagocytosis through the induction of intracellular PSMs expression. PLoS Pathog. 8:e1003016. doi: 10.1371/journal.ppat.1003016
Godfrey, H. P., Bugrysheva, J. V., and Cabello, F. C. (2002). The role of the stringent response in the pathogenesis of bacterial infections. Trends Microbiol. 10, 349–351. doi: 10.1016/S0966-842X(02)02403-4
Gounon, P. (2002). 28 Electron microscopy in cellular microbiology: ultrastructural methods for the study of cell surface antigens and pathogenic host—cell interactions. Methods Microbiol. 31, 531–557. doi: 10.1016/S0580-9517(02)31029-8
Greer, R. B., and Rosenberg, A. E. (1993). A 69-year-old woman with a sclerotic lesion of the femur and pulmonary nodules. N. Engl. J. Med. 328, 422–428. doi: 10.1056/NEJM199302113280609
Haley, K. P., and Skaar, E. P. (2012). A battle for iron: host sequestration and Staphylococcus aureus acquisition. Microbes Infect. 14, 217–227. doi: 10.1016/j.micinf.2011.11.001
Heyer, E. E., Ozadam, H., Ricci, E. P., Cenik, C., and Moore, M. J. (2015). An optimized kit-free method for making strand-specific deep sequencing libraries from RNA fragments. Nucleic Acids Res. 43:e2. doi: 10.1093/nar/gku1235
Holden, M. T., Feil, E. J., Lindsay, J. A., Peacock, S. J., Day, N. P., Enright, M. C., et al. (2004). Complete genomes of two clinical Staphylococcus aureus strains: evidence for the rapid evolution of virulence and drug resistance. Proc. Natl. Acad. Sci. U.S.A. 101, 9786–9791. doi: 10.1073/pnas.0402521101
Imber, M., Loi, V. V., Reznikov, S., Fritsch, V. N., Pietrzyk-Brzezinska, A. J., Prehn, J., et al. (2018). The aldehyde dehydrogenase AldA contributes to the hypochlorite defense and is redox-controlled by protein S-bacillithiolation in Staphylococcus aureus. Redox Biol. 15, 557–568. doi: 10.1016/j.redox.2018.02.001
Jarraud, S., Mougel, C., Thioulouse, J., Lina, G., Meugnier, H., Forey, F., et al. (2002). Relationships between Staphylococcus aureus genetic background, virulence factors, agr groups (alleles), and human disease. Infect. Immun. 70, 631–641. doi: 10.1128/IAI.70.2.631-641.2002
Johns, B. E., Purdy, K. J., Tucker, N. P., and Maddocks, S. E. (2015). Phenotypic and genotypic characteristics of small colony variants and their role in chronic infection. Microbiol. Insights 8, 15–23. doi: 10.4137/MBI.S25800
Kahl, B., Herrmann, M., Everding, A. S., Koch, H. G., Becker, K., Harms, E., et al. (1998). Persistent infection with small colony variant strains of Staphylococcus aureus in patients with cystic fibrosis. J. Infect. Dis. 177, 1023–1029. doi: 10.1086/515238
Kahl, B. C. (2003). Thymidine-dependent small-colony variants of Staphylococcus aureus exhibit gross morphological and ultrastructural changes consistent with impaired cell separation. J. Clin. Microbiol. 41, 410–413. doi: 10.1128/JCM.41.1.410-413.2003
Kahl, B. C. (2005). Thymidine-dependent Staphylococcus aureus small colony variants are associated with extensive changes in regulator and virulence gene expression profiles. Infect. Immun. 73, 4119–4126. doi: 10.1128/IAI.73.7.4119-4126.2005
Kahl, B. C., Becker, K., and Löffler, B. (2016). Clinical significance and pathogenesis of staphylococcal small colony variants in persistent infections. Clin. Microbiol. Rev. 29, 401–427. doi: 10.1128/CMR.00069-15
Kohler, C., von Eiff, C., Peters, G., Proctor, R. A., Hecker, M., and Engelmann, S. (2003). Physiological characterization of a heme-deficient mutant of Staphylococcus aureus by a proteomic approach. J. Bacteriol. 185, 6928–6937. doi: 10.1128/JB.185.23.6928-6937.2003
Langmead, B., and Salzberg, S. L. (2012). Fast gapped-read alignment with Bowtie 2. Nat. Methods 9, 357–359. doi: 10.1038/nmeth.1923
Li, H. (2011). A statistical framework for SNP calling, mutation discovery, association mapping and population genetical parameter estimation from sequencing data. Bioinformatics 27, 2987–2993. doi: 10.1093/bioinformatics/btr509
Li, H., Handsaker, B., Wysoker, A., Fennell, T., Ruan, J., Homer, N., et al. (2009). The sequence alignment/map format and SAMtools. Bioinformatics 25, 2078–2079. doi: 10.1093/bioinformatics/btp352
Makhlin, J., Kofman, T., Borovok, I., Kohler, C., Engelmann, S., Cohen, G., et al. (2007). Staphylococcus aureus ArcR controls expression of the arginine deiminase operon. J. Bacteriol. 189, 5976–5986. doi: 10.1128/JB.00592-07
Massey, R. C., Buckling, A., and Peacock, S. J. (2001). Phenotypic switching of antibiotic resistance circumvents permanent costs in Staphylococcus aureus. Curr. Biol. 11, 1810–1814. doi: 10.1016/S0960-9822(01)00507-3
Massey, R. C., and Peacock, S. J. (2002). Antibiotic-resistant sub-populations of the pathogenic bacterium Staphylococcus aureus confer population-wide resistance. Curr. Biol. 12, R686–R687. doi: 10.1016/S0960-9822(02)01205-8
McCallum, N., Meier, P. S., Heusser, R., and Berger-Bächi, B. (2011). Mutational analyses of open reading frames within the vraSR operon and their roles in the cell wall stress response of Staphylococcus aureus. Antimicrob. Agents Chemother. 55, 1391–1402. doi: 10.1128/AAC.01213-10
McKenna, A., Hanna, M., Banks, E., Sivachenko, A., Cibulskis, K., Kernytsky, A., et al. (2010). The genome analysis toolkit: a MapReduce framework for analyzing next-generation DNA sequencing data. Genome Res. 20, 1297–1303. doi: 10.1101/gr.107524.110
Monecke, S., Luedicke, C., Slickers, P., and Ehricht, R. (2009). Molecular epidemiology of Staphylococcus aureus in asymptomatic carriers. Eur. J. Clin. 28, 1159–1165. doi: 10.1007/s10096-009-0752-2
O'Neill, A. J., Huovinen, T., Fishwick, C. W. G., and Chopra, I. (2006). Molecular genetic and structural modeling studies of Staphylococcus aureus RNA polymerase and the fitness of Rifampin resistance genotypes in relation to clinical prevalence. Antimicrob. Agents Chemother. 50:298. doi: 10.1128/AAC.50.1.298-309.2006
Onyango, L. A., Hugh Dunstan, R., Roberts, T. K., Macdonald, M. M., and Gottfries, J. (2013). Phenotypic variants of staphylococci and their underlying population distributions following exposure to stress. PloS ONE 8:e77614. doi: 10.1371/journal.pone.0077614
Osmon, D. R., Berbari, E. F., Berendt, A. R., Lew, D., Zimmerli, W., Steckelberg, J. M., et al. (2013). Diagnosis and management of prosthetic joint infection: clinical practice guidelines by the Infectious Diseases Society of America. Clin. Infect. Dis. 56, e1–e25. doi: 10.1093/cid/cis966
Ou, J. J., Drilling, A. J., Cooksley, C., Bassiouni, A., Kidd, S. P., Psaltis, A. J., et al. (2016). Reduced innate immune response to a Staphylococcus aureus small colony variant compared to its wild-type parent strain. Front. Cell. Infect. Microbiol. 6:187. doi: 10.3389/fcimb.2016.00187
Pietiäinen, M., François, P., Hyyryläinen, H.-L., Tangomo, M., Sass, V., Sahl, H.-G., et al. (2009). Transcriptome analysis of the responses of Staphylococcus aureus to antimicrobial peptides and characterization of the roles of vraDE and vraSR in antimicrobial resistance. BMC Genomics 10:429. doi: 10.1186/1471-2164-10-429
Pinel-Marie, M.-L., Brielle, R., and Felden, B. (2014). Dual toxic-peptide-coding Staphylococcus aureus RNA under antisense regulation targets host cells and bacterial rivals unequally. Cell Rep. 7, 424–435. doi: 10.1016/j.celrep.2014.03.012
Proctor, R. A., Kriegeskorte, A., Kahl, B. C., Becker, K., Löffler, B., and Peters, G. (2014). Staphylococcus aureus Small Colony Variants (SCVs): a road map for the metabolic pathways involved in persistent infections. Front. Cell. Infect. Microbiol. 4:99. doi: 10.3389/fcimb.2014.00099
Proctor, R. A., van Langevelde, P., Kristjansson, M., Maslow, J. N., and Arbeit, R. D. (1995). Persistent and relapsing infections associated with small colony variants of Staphylococcus aureus. Clin. Infect. Dis. 20, 95–102. doi: 10.1093/clinids/20.1.95
Proctor, R. A., von Eiff, C., Kahl, B. C., Becker, K., McNamara, P., Herrman, M., et al. (2006). Small colony variants: a pathogenic form of bacteria that facilitates persistent and recurrent infections. Nat. Rev. Microbiol. 4, 295–305. doi: 10.1038/nrmicro1384
Rasigade, J.-P., Trouillet-Assant, S., Ferry, T., Diep, B. A., Sapin, A., Lhoste, Y., et al. (2013). PSMs of hypervirulent Staphylococcus aureus act as intracellular toxins that kill infected osteoblasts. PloS ONE 8:e63176. doi: 10.1371/journal.pone.0063176
Robinson, J. T., Thorvaldsdóttir, H., Winckler, W., Guttman, M., Lander, E. S., Getz, G., et al. (2011). Integrative genomics viewer. Nat. Biotechnol. 29, 24–26. doi: 10.1038/nbt.1754
Robinson, M. D., McCarthy, D. J., and Smyth, G. K. (2010). edgeR: a Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 26, 139–140. doi: 10.1093/bioinformatics/btp616
Rogers, K. L., Fey, P. D., and Rupp, M. E. (2009). Coagulase-negative staphylococcal infections. Infect. Dis. Clin. North Am. 23, 73–98. doi: 10.1016/j.idc.2008.10.001
Schuster, C. F., and Bertram, R. (2016). Toxin-antitoxin systems of Staphylococcus aureus. Toxins 8:E140. doi: 10.3390/toxins8050140
Seggewiss, J., Becker, K., Kotte, O., Eisenacher, M., Yazdi, M. R. K., Fischer, A., et al. (2006). Reporter metabolite analysis of transcriptional profiles of a Staphylococcus aureus strain with normal phenotype and its isogenic hemB mutant displaying the small-colony-variant phenotype. J. Bacteriol. 188, 7765–7777. doi: 10.1128/JB.00774-06
Sendi, P., and Proctor, R. A. (2009). Staphylococcus aureus as an intracellular pathogen: the role of small colony variants. Trends Microbiol. 17, 54–58. doi: 10.1016/j.tim.2008.11.004
Stevens, Q. E. J., Seibly, J. M., Chen, Y. H., Dickerman, R. D., Noel, J., and Kattner, K. A. (2007). Reactivation of dormant lumbar methicillin-resistant Staphylococcus aureus osteomyelitis after 12 years. J. Clin. Neurosci. 14, 585–589. doi: 10.1016/j.jocn.2005.12.006
Tande, A. J., Osmon, D. R., Greenwood-Quaintance, K. E., Mabry, T. M., Hanssen, A. D., and Patel, R. (2014). Clinical characteristics and outcomes of prosthetic joint infection caused by small colony variant staphylococci. mBio 5:e01910-14. doi: 10.1128/mBio.01910-14
Tong, S. Y. C., Davis, J. S., Eichenberger, E., Holland, T. L., and Fowler, V. G. (2015). Staphylococcus aureus infections: epidemiology, pathophysiology, clinical manifestations, and management. Clin. Microbiol. Rev. 28, 603–661. doi: 10.1128/CMR.00134-14
Tuchscherr, L., Heitmann, V., Hussain, M., Viemann, D., Roth, J., von Eiff, C., et al. (2010). Staphylococcus aureus small-colony variants are adapted phenotypes for intracellular persistence. J. Infect. Dis. 202, 1031–1040. doi: 10.1086/656047
Tuchscherr, L., Medina, E., Hussain, M., Völker, W., Heitmann, V., Niemann, S., et al. (2011). Staphylococcus aureus phenotype switching: an effective bacterial strategy to escape host immune response and establish a chronic infection. EMBO Mol. Med. 3, 129–141. doi: 10.1002/emmm.201000115
Valour, F., Trouillet-Assant, S., Riffard, N., Tasse, J., Flammier, S., Rasigade, J.-P., et al. (2015). Antimicrobial activity against intraosteoblastic Staphylococcus aureus. Antimicrob. Agents Chemother. 59, 2029–2036. doi: 10.1128/AAC.04359-14
von Eiff, C., Lindner, N., Proctor, R. A., Winkelmann, W., and Peters, G. (1998). Development of gentamicin-resistant small colony variants of Staphylococcus aureus after implantation of gentamicin beads in osteomyelitis as a possible cause of recurrence. Z Orthop Ihre Grenzgeb 136, 268–271. doi: 10.1055/s-2008-1054234
Wessling-Resnick, M. (2010). Iron homeostasis and the inflammatory response. Annu. Rev. Nutr. 30, 105–122. doi: 10.1146/annurev.nutr.012809.104804
Wichelhaus, T. A., Böddinghaus, B., Besier, S., Schäfer, V., Brade, V., and Ludwig, A. (2002). Biological cost of Rifampin resistance from the perspective of Staphylococcus aureus. Antimicrob. Agents Chemother. 46, 3381–3385. doi: 10.1128/AAC.46.11.3381-3385.2002
Wolz, C., Geiger, T., and Goerke, C. (2010). The synthesis and function of the alarmone (p)ppGpp in firmicutes. Int. J. Med. Microbiol. 300, 142–147. doi: 10.1016/j.ijmm.2009.08.017
Keywords: Staphylococcus aureus, small colony variant, prosthetic joint infection, chronic infection, rifampicin resistance
Citation: Loss G, Simões PM, Valour F, Cortês MF, Gonzaga L, Bergot M, Trouillet-Assant S, Josse J, Diot A, Ricci E, Vasconcelos AT and Laurent F (2019) Staphylococcus aureus Small Colony Variants (SCVs): News From a Chronic Prosthetic Joint Infection. Front. Cell. Infect. Microbiol. 9:363. doi: 10.3389/fcimb.2019.00363
Received: 13 June 2019; Accepted: 07 October 2019;
Published: 22 October 2019.
Edited by:
Sarah Vreugde, University of Adelaide, AustraliaReviewed by:
Stephen Peter Kidd, University of Adelaide, AustraliaShuai Le, Army Medical University, China
Copyright © 2019 Loss, Simões, Valour, Cortês, Gonzaga, Bergot, Trouillet-Assant, Josse, Diot, Ricci, Vasconcelos and Laurent. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Frédéric Laurent, ZnJlZGVyaWMubGF1cmVudEB1bml2LWx5b24xLmZy
†These authors have contributed equally to this work