 McKinlee M. Salazar
McKinlee M. Salazar Mônica T. Pupo
Mônica T. Pupo Amanda M. V. Brown
Amanda M. V. Brown- 1Department of Biological Sciences, Texas Tech University, Lubbock, TX, United States
- 2School of Pharmaceutical Sciences of Ribeirão Preto, University of São Paulo, Ribeirão Preto, Brazil
Interactions between insect symbionts and plant pathogens are dynamic and complex, sometimes involving direct antagonism or synergy and sometimes involving ecological and evolutionary leaps, as insect symbionts transmit through plant tissues or plant pathogens transition to become insect symbionts. Hemipterans such as aphids, whiteflies, psyllids, leafhoppers, and planthoppers are well-studied plant pests that host diverse symbionts and vector plant pathogens. The related hemipteran treehoppers (family Membracidae) are less well-studied but offer a potentially new and diverse array of symbionts and plant pathogenic interactions through their distinct woody plant hosts and ecological interactions with diverse tending hymenopteran taxa. To explore membracid symbiont–pathogen diversity and co-occurrence, this study performed shotgun metagenomic sequencing on 20 samples (16 species) of treehopper, and characterized putative symbionts and pathogens using a combination of rapid blast database searches and phylogenetic analysis of assembled scaffolds and correlation analysis. Among the 8.7 billion base pairs of scaffolds assembled were matches to 9 potential plant pathogens, 12 potential primary and secondary insect endosymbionts, numerous bacteriophages, and other viruses, entomopathogens, and fungi. Notable discoveries include a divergent Brenneria plant pathogen-like organism, several bee-like Bombella and Asaia strains, novel strains of Arsenophonus-like and Sodalis-like symbionts, Ralstonia sp. and Ralstonia-type phages, Serratia sp., and APSE-type phages and bracoviruses. There were several short Phytoplasma and Spiroplasma matches, but there was no indication of plant viruses in these data. Clusters of positively correlated microbes such as yeast-like symbionts and Ralstonia, viruses and Serratia, and APSE phage with parasitoid-type bracoviruses suggest directions for future analyses. Together, results indicate membracids offer a rich palette for future study of symbiont–plant pathogen interactions.
Introduction
Many insects that host microbial symbionts can also vector significant plant pathogens (Hogenhout et al., 2008a; Hogenhout et al., 2008b; Scholthof et al., 2011; Mansfield et al., 2012; Weintraub et al., 2019; Shafiq et al., 2020), but investigating the co-occurrence and forces underlying insect symbiont and plant pathogen infection presents a challenge. Some data suggest there could be an energetic cost of hosting symbionts, which might reduce an insect’s plant pathogen vectoring ability, while conversely, the benefits of symbionts might provide energy or metabolic advantages that increase plant pathogen transmission (Heck, 2018; Gonella et al., 2019). In developing a theory to predict the outcomes of these interactions, part of the challenge is their complexity. Insect symbioses tend to be richly multipartite, including many species of bacteria and fungi along with their phages and viruses. This complexity involves many insect body sites (gut, salivary glands, fat body, hemolymph, bacteriomes) and external ecological interactions that mediate symbiont and pathogen exchange. As a final surprising dimension of complexity in the dynamics of insect symbionts and plant pathogens, several studies show that insect symbionts can transfer through plant tissues and plant pathogens can evolve readily into insect symbionts (Chrostek et al., 2017; Martinson et al., 2020).
Hemipterans—especially aphids, whiteflies, psyllids, leafhoppers (Cicadellidae), and planthoppers (Fulgoromorpha)—are the most well-studied vectors of plant pathogens, but few studies have investigated treehoppers (Membracidae). Whereas related Auchenorrhyncha hemipterans typically host two bacteria in their symbiont organs (bacteriomes) that cooperatively synthesize missing essential amino acids not present in their plant-sap diet (Wu et al., 2006; Bennett and Moran, 2013; Douglas, 2016; Mao et al., 2017), there are exceptions. Some species have lost one of the obligate symbionts and gained a replacement symbiont (Sudakaran et al., 2017; Matsuura et al., 2018; Bell-Roberts et al., 2019). Other Hemiptera may have gained numerous symbionts. For example, light microscopy studies show Brazilian membracids host at least 28 diverse symbiotic microbes with up to six microbial species cohabiting a bacteriome (Rau, 1943; Müller, 1962; Buchner, 1965). It is unclear which of these microbes are primary or obligate from the host’s perspective, or secondary (facultative), functioning to enable their hosts to survive biotic or abiotic stresses or thrive in particular niches (Oliver et al., 2012; White et al., 2013; Oliver et al., 2014; Guidolin et al., 2018; Santos-Garcia et al., 2018; Lemoine et al., 2020). Many membracids further depend on additional behavioral symbioses with ants, bees, and wasps (Delabie, 2001; Godoy et al., 2006; Ibarra-Isassi and Oliveira, 2018; Klimes et al., 2018; Canedo-Júnior et al., 2019; dos Santos et al., 2019), which feed on the membracids’ secreted honeydew which, in turn, contains additional symbiotic microbes (Leroy et al., 2011; Fischer et al., 2015; Calcagnile et al., 2019; Shamim et al., 2019). However, there are few molecular studies of membracid symbioses. To date, there is one genomic study of a dual-symbiosis in a membracid (Mao et al., 2017) and one 16S rRNA and microscopy-based study showing three or four interacting microbial symbionts in two membracids (Kobiałka et al., 2019). Both studies, however, focused on membracids from the temperate climates, whereas membracid taxonomy, ecology, and microbiota are richest in the neotropics (Buchner, 1965; Cryan et al., 2000; Cryan et al., 2004; Deitz and Wallace, 2012; Hu et al., 2019).
A few membracids in North America are considered significant crop pests—specifically members of tribe Ceresini, such as Spissistilus, Ceresa, and Stictocephala (alfalfa and buffalo treehoppers) which cause swelling on stems or cuts that can facilitate infections in soybean and alfalfa (Meisch and Randolph, 1965; Bailey, 1975); however, in the neotropics, an estimated 18 genera of treehoppers are found as crop pests (Godoy et al., 2006). One study suggests Ceresa in Argentina may vector the witches’ broom phytoplasma (ArAWB) (Grosso et al., 2014). Further studies suggest membracids vector significant plant viruses: Micrutalis (tribe Micrutalini) vectors the viral pseudo-curly top disease (TPCTV) of tomatoes (Mead, 1986; Briddon et al., 1996) and Spissistilus vectors a closely related DNA virus, Grapevine red blotch-associated virus (GRBaV) (Bahder et al., 2016). Membracids’ roles as vectors of plant pathogens are not as well-studied as those of aphids, psyllids, and leafhoppers, which are more commonly found as crop pests. However, their predominance on woody plants makes membracids prime candidates worth investigating as possible vectors for phytoplasmas specific to woody plants such as 16SrIII, 16SrX (apple proliferation), or ESFY (European stone fruit yellows) (Wilson and Weintraub, 2007). Some studies suggest hemipteran secondary symbionts may travel between insects by passage through plants (Chrostek et al., 2017; Li et al., 2018; Pons et al., 2019), including Wolbachia, Rickettsia, Candidatus Cardinium, Serratia, and Symbiopectobacterium (formerly BEV), but these plant–insect–microbe exchanges have not been studied in membracids.
The current study sought to characterize symbionts and potential plant pathogens in membracids, including neotropical species, examining co-occurrence and correlations in abundance as an initial survey of this underexamined group. Our approach used shotgun metagenomics of the membracid bacteriome and surrounding tissues and hemolymph, searched for matches to major bacterial plant pathogens (Mansfield et al., 2012) including phytoplasmas (Hogenhout et al., 2008b), and DNA plant viruses (Shafiq et al., 2020), and an array of bacteriophage (Duron, 2014; Rouïl et al., 2020). Results showed our set of membracids hosted at least nine types of potential plant pathogens, in addition to 12 primary and secondary endosymbionts, along with numerous bacteriophages, and other viruses and parasites. Although our study focused on bacteriomes and surrounding hemolymph, we detected traces of phytoplasmas and spiroplasmas with no indication of plant viruses and we detected significant correlation between subsets of microbes present. Furthermore, we identified divergent Brenneria plant pathogen-like organisms, potential transitional strains of Arsenophonus-like symbionts, Ralstonia and Ralstonia-type phages, Serratia, and APSE-type phages and bracoviruses, indicating membracids may be host to a rich array of symbiont–plant pathogen interactions.
Materials and Methods
Insect Collection and Bacteriome Dissection
To obtain a phylogenetically diverse collection of membracids, sampling was performed over several years in the US and Brazil (permits and registration: SISBIO 46555-6; SisGen A350676 and R848FAD). Adult insects were collected by sweep net or by inspection of branches with capture in large plastic zip-closure bags. Sample collection site details are shown in Supplementary Table S1. Insects were preserved at -80°C prior to dissection in Brazil or the U.S. Insects were photographed and identified morphologically to genus or species prior to dissection. Microdissection to extract bacteriomes was performed on insects, one at a time, on ice trays with forceps and micron pins washed with bleach and 70–95% ethanol between each insect. For dissections, insects were placed in 100 µl of sterile phosphate buffered saline. Under 20–60X magnification, the posterior portion of the abdomen (~last three segments) was removed with forceps and a dissection needle, then tissue containing the bacteriomes was removed with micron pins and placed in labeled tubes, pooling several individuals collected at a single site together, to increase DNA yield (number of pooled individuals per sample: BM11 = 6, BM13-1 = 8, BM13-2 = 4, BM4 = 8, BM43 = 5, BM44 = 8, BM50 = 3, BM51 = 2, BM53 = 4, BM56 = 8, BM59 = 5, BM65 = 1, BM69 = 4, Cer = 4, Ent = 2, Gar = 8, MemA = 10, MemE = 12, MemM =16, Pub = 3). Due to the small size of bacteriomes, dissection was performed conservatively, allowing the inclusion of small amounts of surrounding abdominal hemolymph and host tissues including fat body cells.
DNA Extraction and Illumina Library Preparation and Sequencing
To isolate and sequence DNA from pooled bacteriome tissues, we used either the Qiagen DNeasy Blood & Tissue Kit (Valencia, CA) or the Qiagen AllPrep DNA/RNA/miRNA Kit (Valencia, CA) following the manufacturer’s directions. DNA quantity and quality were assessed on the Nanodrop spectrophotometer. Library preparation for samples ‘Cer’, ‘Ent’, ‘Gar’, and ‘Pub’ was performed as described previously (Brown et al., 2014). For all other samples, libraries were prepared as follows: approximately 0.2 to 1 µg of DNA was used with the QIAseq FX DNA Library Kit (Valencia, CA) following the manufacturer’s directions except with modified fragmentation times and AMPure bead concentrations optimized to target 450–550 bp inserts. Library quality and quantity was assessed on the Agilent 2200 TapeStation. Libraries were normalized and pooled before sequencing on Illumina HiSeq, with 150 PE cycles performed at Genewiz, Inc (NJ).
Sequence Assembly
To assemble reads for analysis, reads were filtered and trimmed using Trimmomatic v.0.38 (Bolger et al., 2014) and overlaps in paired reads were identified and joined together using Pear v0.9.11 (Zhang et al., 2014). Filtered paired and merged reads were de novo assembled with metaSPAdes v.3.13.0 (Bankevich et al., 2012; Nurk et al., 2017) using error correction and kmers (-k 25,33,43,53,65,87,101,115). Assembly quality and basic statistics were evaluated using Quast v5.0.1 (Gurevich et al., 2013).
Database Searches
To identify microbes within the samples and confirm insect taxonomic identification, assembled scaffolds were processed by a three-step blast pipeline, using custom scripts. Briefly, to increase the speed of large blastn searches, scaffolds were first subjected to blastn in BLAST+ v2.10.1 (Camacho et al., 2009) against small custom target databases: a database of cytochrome oxidase (COI) genes from hemipterans, a database of 16S rRNA sequences from select bacteria including a wide range of endosymbionts, a database of 18S rRNA genes from fungi including a wide range of yeast-like symbionts, a database of phytoplasma/mycoplasma genomes downloaded from GenBank (NCBI; National Center for Biotechnology Information), and a virus and phage genome database compiled from the widest possible range of viral and phage genomes from NCBI. Samtools (Li et al., 2009) faidx was then used to extract the blast hit regions matching each custom database. These hit regions were then subjected to a second blastn search against all sequences in the nt database, extracting taxonomic data with the resulting hits. These hits were then filtered for the top blast match per scaffold, using a simple grep, to extract separately the full-length scaffolds corresponding to the desired hits matching taxonomic groups of interest (e.g., ‘bacteria’, ‘fungi’, ‘viruses’, ‘bugs’, ‘eukaryotes’, and ‘mycoplasmas’). Finally, the full scaffolds extracted above were subjected to a final blastn against the nt database to confirm that each scaffold matched the organismal clade previously identified.
Abundance and Correlation Analysis
To assess the relative abundance and correlation between symbionts, plant pathogens, and other microbes within the membracids, we performed several filtering and analysis steps. First, we selected only scaffolds with top blastn hit to bacteria, fungi, insects, or viruses, removing any contaminants with high blastn sequence identity to common human microbiota or viruses. Next, we selected only scaffolds with blast hit length >40 bp and evalue >0.03 and bitscore >52. We then removed hits below 83% hit identity to the target (except with virus hits, for which we set a lower threshold of >70%). We also removed hits <48 bp length, except for phytoplasma and viruses, which we kept at the >40 bp threshold. To estimate and normalize coverage, kmer coverage was converted to absolute coverage with the equation C = (CK R)/(R-K+1), where C is total coverage, CK is kmer coverage, K is the length of kmers, and R is read length. For hits to the same species within a sample, coverages were added (i.e., combining variants) to assess total abundance. Final absolute coverages for all relevant hits were normalized based on the host insect’s COI gene coverage. Abundance and presence/absence were calculated, normalizing for within-sample abundance after COI normalization, plotting results using the function ‘heatmap’ in R. Spearman rank correlation was calculated for a matrix including all blastn hits, organized by taxa. Spearman rho values and p-values were calculated and plotted using several R packages: ‘Hmisc’ v4.5-0 (Harrell Miscellaneous) program ‘rcorr’ which calculates a matrix of Spearman’s rho rank correlation coefficients for all pairs of columns for non-missing elements, using midranks for ties, specifying method = “spearman”, corrplot with order = “hclust”, hclust.method = “average”, and for plotting, using packages ‘tidyr’, ‘tibble’, ‘ggplots2’, ‘corrplot’. P-value were corrected for multiple testing using the Benjamini and Hochberg (1995) method (FDR) in R with ‘p.adjust’ “BH”. Matching hits were classified into predicted categories (primary or secondary symbionts, entomopathogens or viruses, or potential pathogenic or beneficial plant associated microbes) based on metadata with matching references in NCBI (e.g., host insect or plant source and keywords indicating its role) combined with literature surveys on matching species or strains. Because some matches, such as Pantoea, Enterobacter, and Serratia can be potentially insect-associated, but are most often studied and reported as plant-associated, for the purposes of this study, we classified these as putative or potential plant pathogens. Similarly, those matching strains with possible beneficial functions or more rarely, other functions, were classified for this study as “potentially beneficial” plant-associated bacteria. These classifications are by no means definitive, but instead serve as a tentative best assessment of potential functional class.
Phylogenetic Analysis
To confirm the taxonomic identity and evolutionary place of the membracids sampled, we extracted partial cytochrome oxidase I (COI) sequences from our scaffolds and performed phylogenetic analyses with other membracid and outgroup sequences downloaded from GenBank. While the COI mitochondrial locus is not ideal for inferring deep phylogenetic relationships in the Membracidae, it is useful as an abundant marker likely to produce sufficient coverage for samples with lower sequencing depth and provides databases of additional species for comparison. Resolving deep relationships among major membracid clades was not the primary goal of this study. COI sequences were aligned with Mafft v1.0.4 (Katoh and Standley, 2013) within the Geneious Prime v2020.0.4 (Biomatters, Ltd) suite. Maximum likelihood phylogenetic analysis was performed using RAxML v4.0 (Stamatakis, 2014) with the GTR Gamma nucleotide model, with rate heterogeneity alpha estimated, and with rapid bootstrapping and search for the best-scoring ML tree (-f a -x 1) with 100 replicates. Bayesian inference phylogenetic analysis was also performed on the same alignment block using MrBayes v2.2.4 (Huelsenbeck and Ronquist, 2001; Ronquist et al., 2012) with substitution model GTR+G with 4 categories, and Markov chain Monte Carlo settings of: chain length 1,100,000, 4 heated chains, heated chain temp 0.2, subsampling frequency 200, Burn-in length 100,000, with random seed 31,569, and priors with unconstrained branch lengths GammaDir (1,0.1,1,1), checking for convergence with minESS >200. Phylogenies were displayed using FigTree v1.4.4 (http://tree.bio.ed.ac.uk/software/), and image annotations were added in Adobe Illustrator.
To confirm microbial (bacterial and fungal) sequence identities and relationships, phylogenetic analyses were performed on 16S and 18S/28S rRNA regions extracted from our scaffolds, combined with the top 250 to 500 blastn hits from GenBank. Alignments and phylogenetic analyses were performed as described above for the COI region.
Results
Membracid Microbiota Diversity and Abundance
In total, from the 20 sequenced treehopper samples comprising 8,609,965 scaffolds of >500 bp length adding to 8,708,441,348 bp of assembled sequence of which 5,539 scaffolds were over 10,000 bp and 189 scaffolds were over 50,000 bp with largest scaffold length 642,820 bp (see Supplementary Table S2), blastn top matches revealed over 133 potential strains or variants of bacteria, fungi, viruses, and parasites. There were 12 major groups of primary or secondary symbionts, with Arsenophonus, Candidatus Sulcia and Candidatus Nasuia (hereafter denoted simply Sulcia and Nasuia), Rickettsia, Candidatus Sodalis (hereafter denoted Sodalis), and Bombella sp. being the most abundant, based on 16S rRNA gene hits, in this order (Figure 1). Much less abundant symbionts included Burkholderia, Wolbachia, Yeast-like symbionts/Ophiocordyceps, Candidatus Hamiltonella (hereafter denoted Hamiltonella), Candidatus Gullanella (hereafter denoted Gullanella), and Sulfuriferula sp. There were 11 groups of potential entomopathogens including putative endogenous nudivirus, with the latter being most relatively abundant, based on viral genome hits, followed by Iridovirus Liz-CrlV, and entomopoxviruses (Figure 1). There were nine groups of bacteriophages, with Wolbachia phage WO and Hamiltonella-type APSE phages being most common, followed by Sodalis phage and Ralstonia phage (Figure 1). There were trace levels of parasitoid wasps, but relatively high levels of the Cotesia-type bracovirus (Figure 1). Among potential plant pathogens and plant-associated microbes, the most abundant groups were Brenneria-like and Pectobacterium-like strains, followed by Pantoea agglomerans, Spiroplasma spp., Candidatus Phytoplasma species (hereafter denoted Phytoplasma spp.), and Serratia sp. However, the Spiroplasma and Phytoplasma abundances were calculated based on genome-wide hits, because we did not find significant long 16S rRNA hits to either of these mycoplasma groups (Figure 1). Additional notable plant pathogens found include a distant match to the pathogenic fungi Claviceps africana and a highly similar match to the plant pathogen Ralstonia solanacearum.
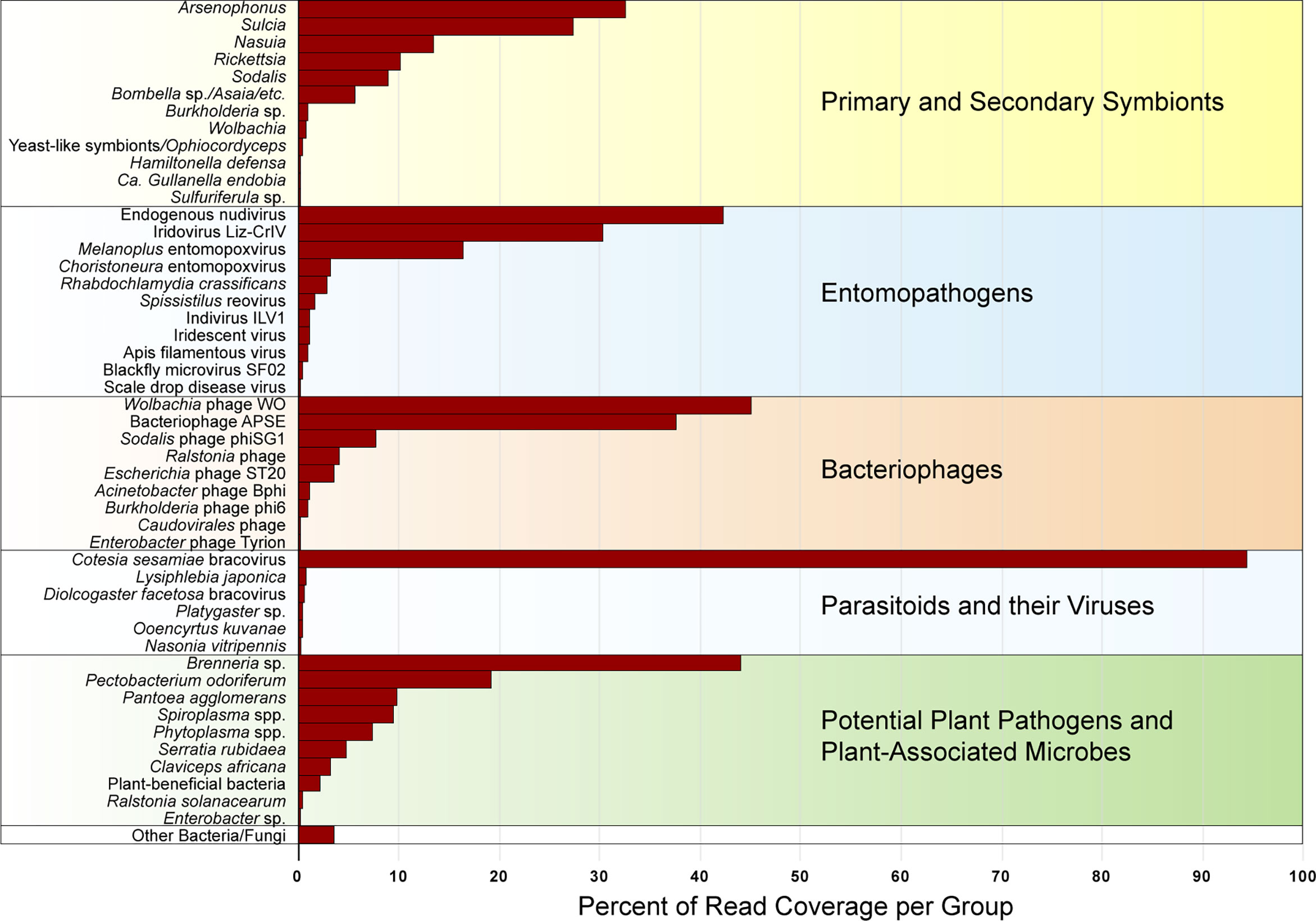
Figure 1 Relative abundance of primary and secondary membracid symbionts, entomopathogens, bacteriophages, parasitoids and their viruses, and potential plant pathogens (classified based on predominant function of closest blast hit), combined for all membracids, expressed as percentage of read coverage per each group shown in the figure, after normalizing between samples based on membracid cytochrome oxidase I (COI) gene coverage, calculated as a percentage of the read coverage for assembly scaffolds with blast hits to 16S rRNA genes (for bacteria), 18S rRNA genes (for fungi), or genomes for viruses and mycoplasmas or spiroplasmas.
Relative Abundance of Potential Primary and Secondary Symbionts in Membracids
Based on morphological identification, combined with COI blastn searches and phylogenetic analysis, our sampled treehoppers included 19 samples within family Membracidae, and one sample in a sister-family Aetalionidae, together comprising 8 tribes, 13 genera, and 16 species. The membracids sampled fell within subfamilies Centrotinae, Smiliinae, and Membracinae, in tribes Gargarini, Ceresini, Polyglyptini, Amastrini, Micrutalini, Aconophorini, and Membracini, most of which were supported clades in both Bayesian and Maximum Likelihood analyses (Figure 2). Presence/absence and abundance of microbes and pathogens within these samples shows a varied pattern of primary or secondary symbionts, entomopathogens, entomoviruses, bacteriophages, parasitoids and viruses, and possible plant pathogens (Figure 2). All species hosted the primary symbiont, Sulcia, with the average coverage of 282X, but with Sulcia occurring at varying abundances relative to other organisms (Figure 2). Phylogenetic analyses suggested these Sulcia symbionts formed a strongly supported monophyletic clade with similar phylogenetic topology to that of their hosts (Supplementary Figure S1). The second primary symbiont, betaproteobacteria Nasuia, appeared to be missing entirely for four species (Aetalion reticulatum, Micrutalis calva, Guyaquila tenuicornis, and Calloconophora sp.) (Figure 2). Remaining membracid Nasuia strains formed a well-supported monophyly with relationships that appear similar to those of the host (Supplementary Figure S2).
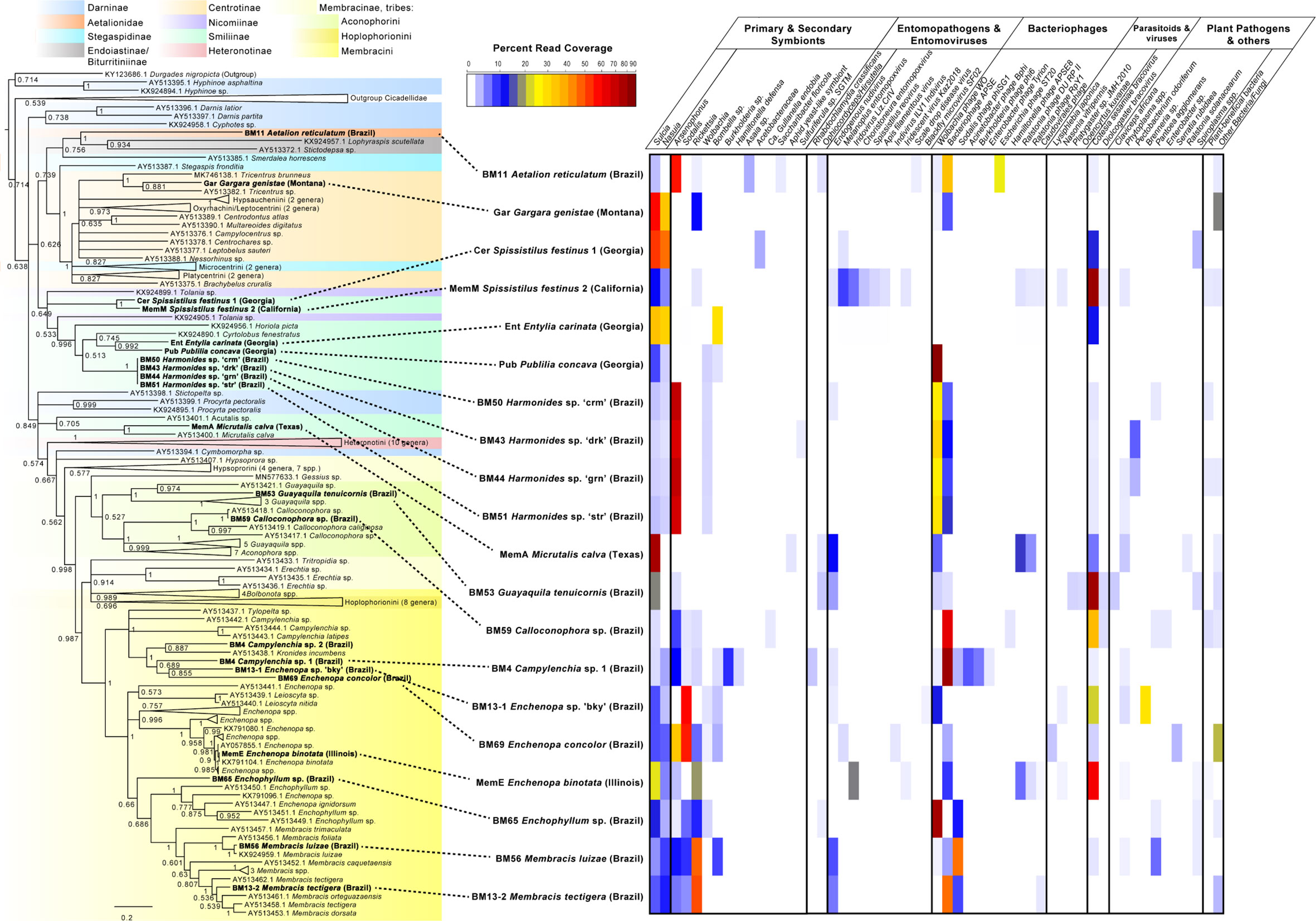
Figure 2 Phylogenetic tree and abundance plot of symbionts and plant pathogens of membracids in this study. Phylogeny is based on 956 aligned positions of the partial cytochrome oxidase I (COI) gene, generated using Bayesian 50% majority rule in MrBayes with GTR+G with 4 rate categories model, and showing posterior probabilities on branches (a similar topology with similar support was generated by RAxML GTR+Gamma with 100 bootstrap replicates). Specimens included in this study are shown in bold font. The presence and abundance plot is based on blastn hits to assembly scaffolds of symbionts, microbes, and pathogens, is depicted in the abundance heatmap as read coverage normalized as a percentage of the read coverage per scaffolds with blast hits from each sample, shown in color legend. Categories in abundance plot were classified based on predominant function of closest blast hit, and include putative primary and secondary membracid symbionts (bacterial and fungal); putative entomopathogens (including bacteria and fungi) and entomoviruses; bacteriophages, parasitoid wasps, and their viruses; potential fungal and bacterial plant pathogens; other bacteria and fungi.
Common or abundant secondary or perhaps primary replacement symbionts included Arsenophonus, Sodalis, Rickettsia, Wolbachia, and Bombella sp. Rarer or occasional symbionts were Burkholderia, Hamiltonella, Gullanella, Acetobacteraceae/Saccharibacter, and Sulfuriferula sp. Phylogenetic analysis of Arsenophonus 16S rRNA (Figure 3 and Supplementary Figure S3) showed most of these isolates clustered within a supported clade, with no monophyletic sub-clade for those strains from membracids; however, many samples contained more than one distinct sequence of the 16S rRNA gene. None of our sequences clustered with either of the two plant pathogenic groups of Arsenophonus-like organisms, Candidatus Phlomobacter fragariae and the ‘SMC proteobacterium isolates’, renamed Ca. Arsenophonus phytopathogenicus (Figure 3). Similarly, our sequences did not group with the Aschnera or ‘ALO-3’ clades comprising proposed obligate endosymbionts. One sample (BM59 Calloconophora sp.) included a more distantly placed Arsenophonus-like variant that clustered close to the adelgid endosymbiont clade Candidatus Hartigia pinicola. Fewer samples hosted Sodalis-like 16S rRNA sequences, but these formed no clear monophyly for strains from membracids, and several sequence variants were found within some samples (Supplementary Figure S4). Rickettsia formed two distinct clades: one clustered exclusively with hemipteran-host Rickettsia, and the other clade having polytomy with widely diverged insect hosts (Supplementary Figure S5). The Wolbachia sequences in these samples fell into supergroups A and B, widespread in insects (Supplementary Figure S6). Acetobacteraceae, including Bombella sp., Asaia sp., and Saccharibacter sp. were found in several samples at high abundances. Phylogenetic analyses of representative 16S rRNA gene sequences showed Asaia sp. in Aetalion reticulatum and a distinct clade of Bombella sp. in six membracid samples (Supplementary Figure S7). Five samples hosted yeast-like symbionts or potentially entomopathogenic fungi in the Ophiocordyceps-like groups, forming four distinct clusters, phylogenetically (Supplementary Figures S8, S9).
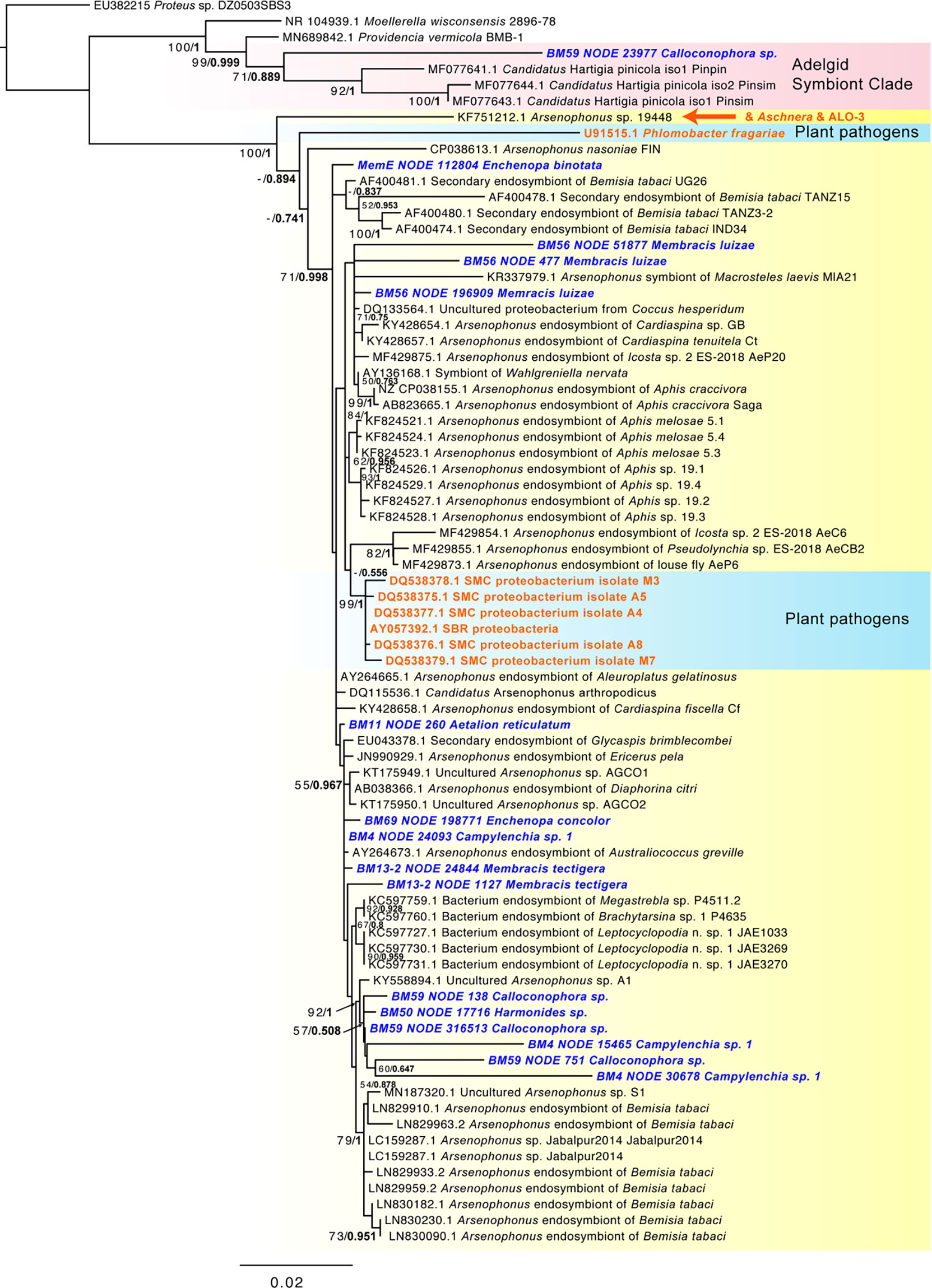
Figure 3 Phylogeny of Arsenophonus-like sequence based on 1,562 aligned positions of the 16S rRNA gene for sequences from GenBank and membracid samples in this study (bold blue font). Maximum likelihood phylogeny reconstruction was performed in RAxML GTR+Gamma with 100 bootstrap replicates. Supported nodes that were obtained from Bayesian 50% majority rule analysis in MrBayes with GTR+G with 4 rate categories are shown as values on branches as ML bootstrap/Bayesian posterior, with the latter values in bold font. Various Arsenophonus-like groups organisms are highlighted with color shading (yellow – most secondary symbiont Arsenophonus, red – clade with adelgid symbionts, blue – plant pathogenic organisms). The position of the obligate symbiont Aschnera and ALO-3 clade is indicated in orange font. A phylogeny including additional shorter scaffolds is shown in Supplementary Figure S3.
Closely related samples (i.e., in the same species or same genus) shown in Figure 2 generally had more similar symbiont patterns, e.g., Harmonides sp., Enchenopa spp., and Membracis spp. This pattern was similar for bacteriophages, but not for entomopathogens and entomoviruses, except endogenous nudiviruses. Among bacteriophages, Wolbachia phage WO and the Hamiltonella-derived bacteriophage APSE were highly abundant in many samples, followed by Sodalis phage phiSG1. Parasitoid wasp-like reads were rare and at low abundance, but wasp bracoviruses (Cotesia-like and Diolcogaster-like) were common, occurring in all but four samples, and the Cotesia-like bracovirus was highly abundant in many samples.
Presence of Potential Plant-Pathogenic Microbes and Viruses, and Plant-Beneficial Microbes
Possible plant pathogens were identified at relatively low levels (Figure 2). One sample (Guyaquila tenuicornis) carried a fungal isolate distantly matched to the fungal plant pathogen Claviceps africana. Eleven samples carried one or more hits to Phytoplasma spp. or Spiroplasma spp. (Table 1). However, despite these scaffolds’ top blastn similarity to these mycoplasmas or spiroplasmas, many of the hits were short, and no 16S rRNA matches to either Phytoplasma spp. or Spiroplasma spp. were found. Several other putative plant pathogens were detected, based on 16S matches: Pectobacterium sp., Brenneria sp., Pantoea agglomerans, Enterobacter sp., Serratia rubidaea, and Ralstonia solanacearum. While many of the Pectobacteriaceae hits were low-coverage or consisted of short 16S rRNA matches, two samples (BM44 & BM43 Harmonides sp.) had high coverage scaffolds (>100X) that clustered consistently with Brenneria sp. in both maximum likelihood and Bayesian phylogenetic analysis (Figure 4), although with low bootstrap support and Bayesian posterior values. None of the Pectobacteriaceae hits in our samples clustered with the Candidatus Symbiopectobacterium group.
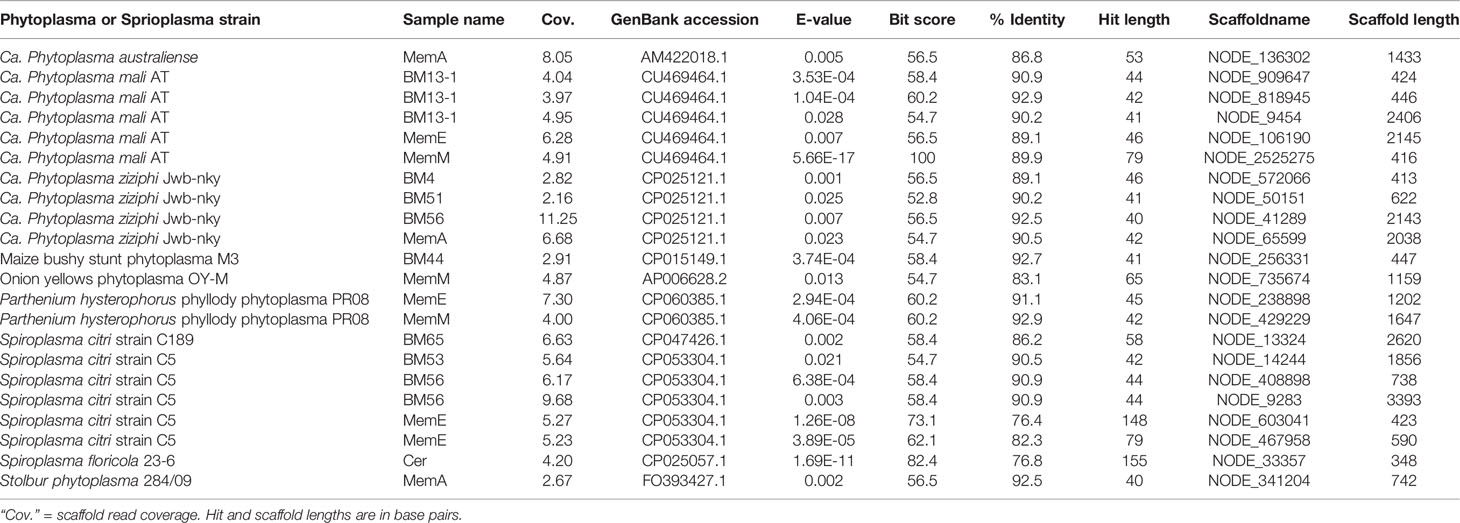
Table 1 Phytoplasma and Spiroplasma species and strains to which our membracid samples’ assembly scaffolds had top highest blastn matches, sorted in alphabetical order of strain names.
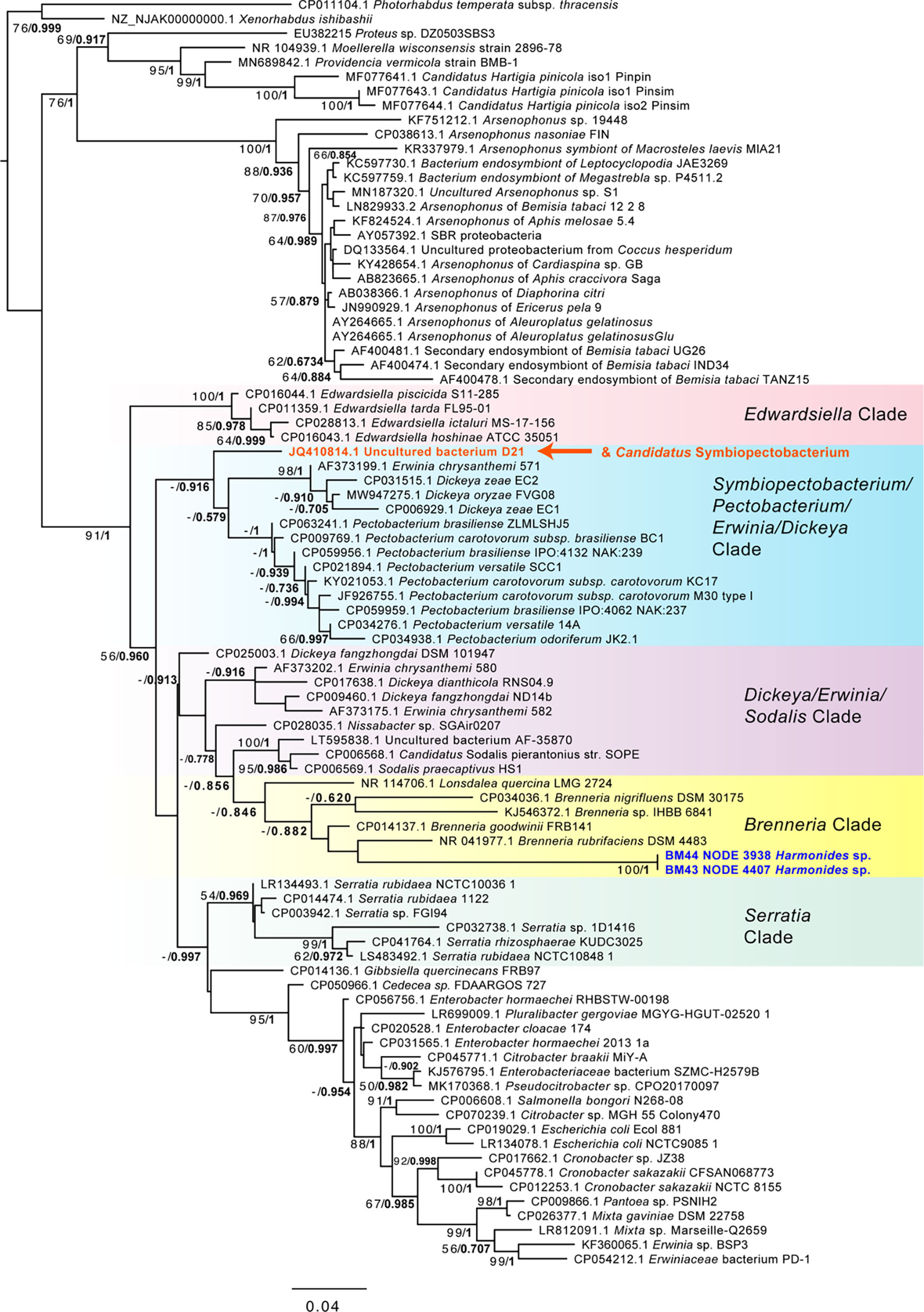
Figure 4 Phylogeny of Enterobacteriales including Pectobacteriaceae and Brenneria-like sequences based on 1,556 aligned positions of the 16S rRNA gene for sequences from GenBank and two membracid samples in this study with highest coverage (bold blue font). Maximum likelihood phylogeny reconstruction was performed in RAxML GTR+Gamma with 100 bootstrap replicates. Supported nodes that were obtained from Bayesian 50% majority rule analysis in MrBayes with GTR+G with 4 rate categories are shown as values on branches as ML bootstrap/Bayesian posterior, with the latter values in bold font. Various Pectobacteriaceae groups are highlighted with color gradient shading. The position of the Candidatus Symbiopectobacterium clade, comprising members with independent secondary endosymbiosis in insects and nematodes, is indicated in orange font.
Several of the bacteriophages detected in these data (Figure 2) appeared to be specific to these plant pathogens—particularly Ralstonia phages DU RP II and RpY1, and Enterobacter phage Tyrion. Many samples (12) were found to host other bacteria and fungi, based on 16S rRNA and 18S rRNA matches, for example, we found several hits matching typically plant-protective or plant-beneficial bacteria (e.g., in the groups Proteus spp., Acinetobacter, Pseudomonas, and Rhizobiales). Specific Pseudomonas matches were to strains naturally associated with potato, corn, and grass. In several samples, these unidentified (i.e., low blast similarity to named species) microbes were highly abundant. No plant viruses were found despite explicit searches against numerous DNA virus groups in our databases, including the Geminiviruses which are transmitted by a wide range of hemipterans and are often circulative in the insect.
Correlations in Abundances of Possible Plant-Pathogens, Symbionts, Other Organisms, and Viruses
Correlation analyses showed a range of positive and negative Spearman rho values for relative abundances of symbionts, plant pathogens, and other organisms and viruses within our membracids (Figure 5). The correlation plot of the rho values shows clusters of positively associated taxa (boxes in Figure 5), with larger clusters having putative plant pathogen taxa (dark orange font in Figure 5) for the Pectobacterium sp. + Wolbachia/Arsenophonus, Phytoplasma spp. +Bombella/Burholderia/Hamiltonella, Enterobacter sp. + Gullanella, and Ralstonia sp. + Aphid yeast-like symbiont. Within these groups, numerous entomopathogens and entomovirus taxa and phage were clustered.

Figure 5 Spearman correlation heatmap of symbionts, plant pathogens, and other microbes and viruses in membracids. Abundances were normalized to the membracid’s cytochrome oxidase (COI) gene coverage before analysis of correlation. Spearman’s rho R-values are depicted with blue to orange shading, from negative to positive. Statistically significant p-values after FDR correction using the Benjamini and Hochberg (1995) method, showing values <0.05 are represented with dark orange dots. Putative plant pathogen names are depicted in dark orange bold font, and putative primary or secondary membracid symbiont names are depicted in black font.
Statistical support for Spearman rho, after Benjamini and Hochberg (1995) correction, produced several strongly positively associated taxa (high R-values) with p-values < 0.05 (Tables 2, 3). Among correlations with putative plant-pathogenic taxa (Table 2), Brenneria sp. was significantly correlated with insect microvirus and a parasitoid, Enterobacter sp. was significantly associated with Gullanella and various phage and bracoviruses, Ralstonia sp. was associated with Aphid yeast-like symbionts and Ralstonia-type phages, Serratia sp. was associated with Indivirus and parasitoids, and Phytoplasma spp. was associated with Hamiltonella, Burkholderia, and several phages.
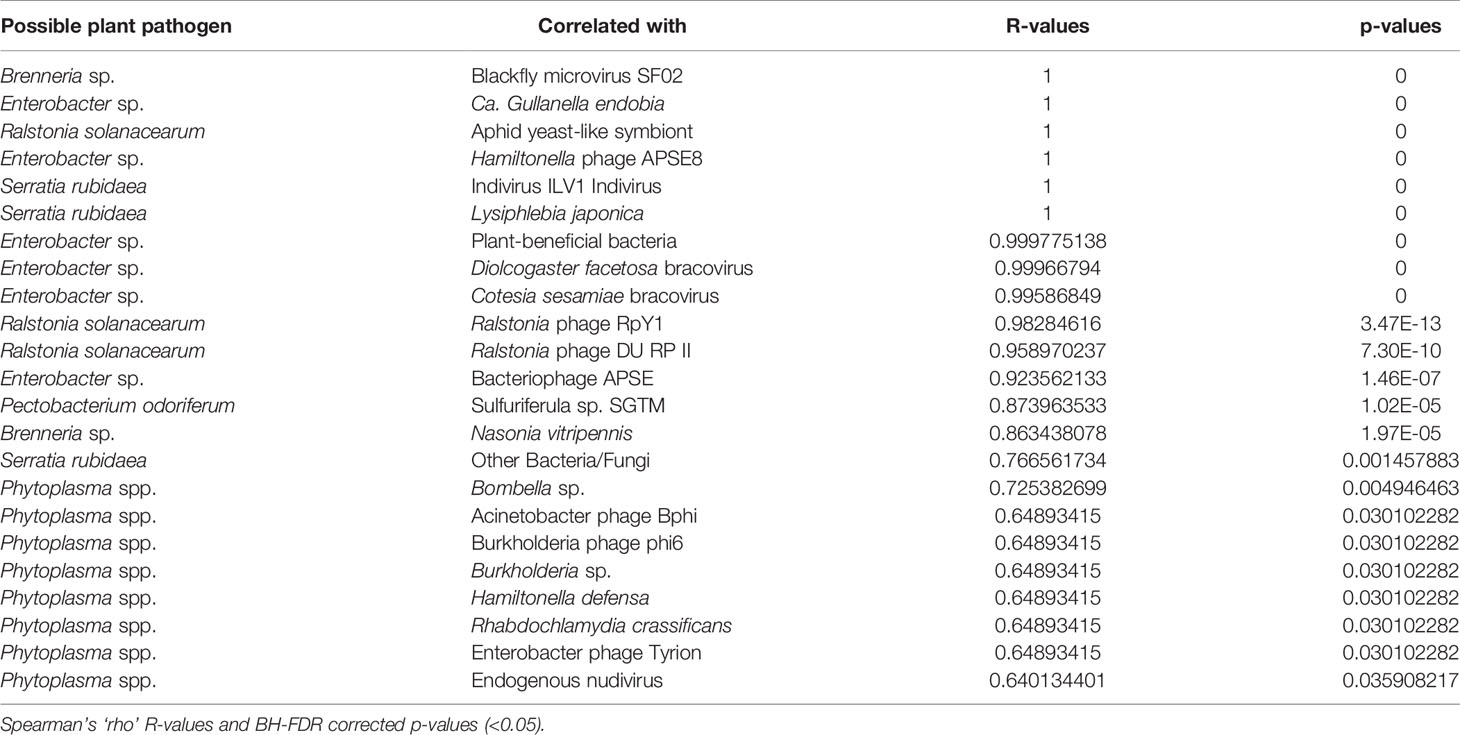
Table 2 Statistically significant correlations between relative abundances of putative plant-pathogens (classified based on predominant function of closest blast hit) and other organisms and viruses in membracids.
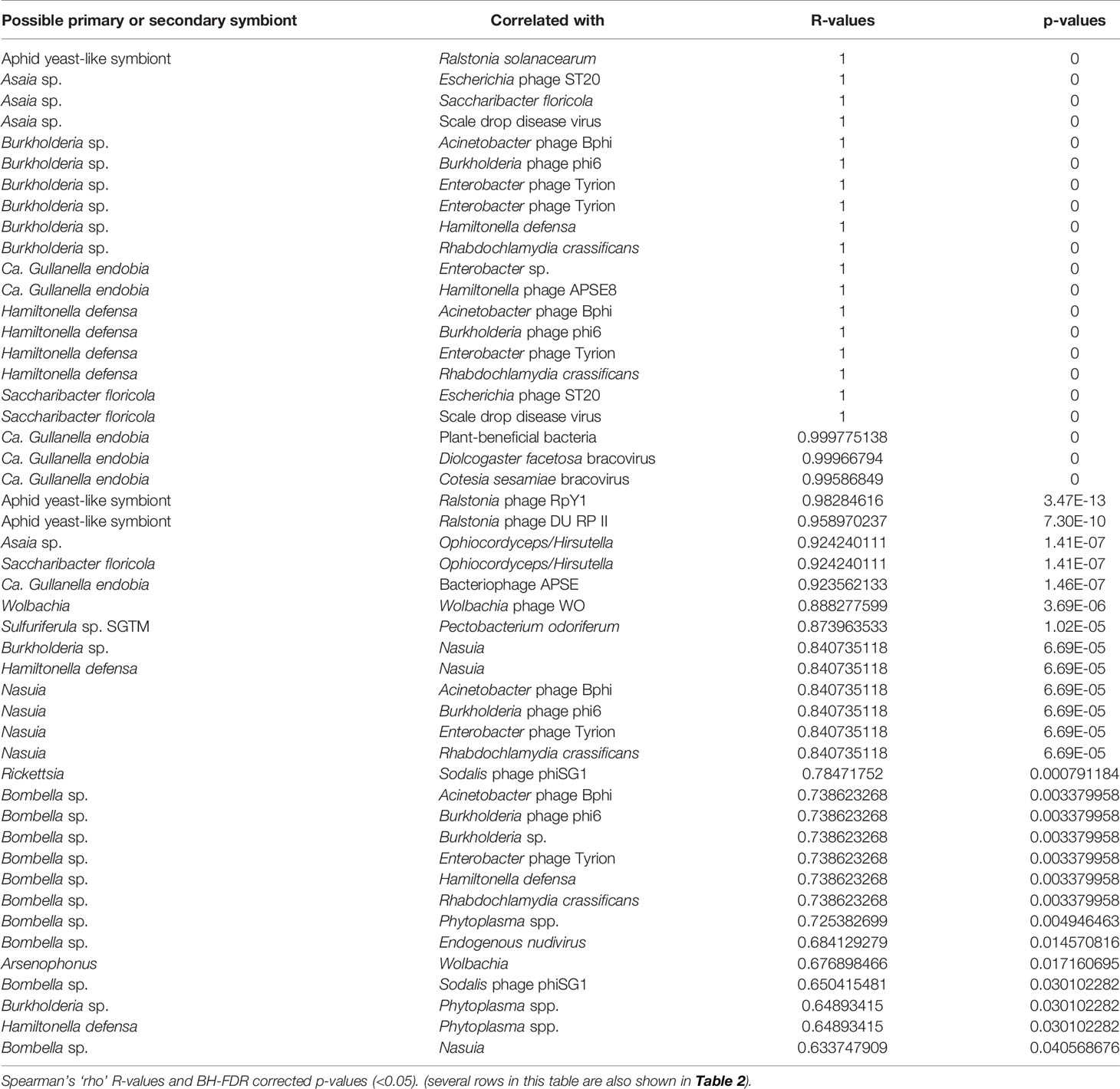
Table 3 Statistically significant correlations between relative abundances of primary and secondary symbionts of membracids and other organisms and viruses in the samples.
Among positively correlated primary and secondary symbionts and other taxa (Table 3), many of these associations were between bacterial symbionts and their presumptive phages. Other associations with symbionts were described previously, in Table 2 and in clustered boxes described for Figure 5.
Discussion
As a step toward discovering how symbionts impact vectoring of plant pathogens in an underexplored group of phloem-feeding insects, we performed this metagenomic sequencing study on membracids. We found, as expected based on historical studies of membracids from Brazil (Rau, 1943; Buchner, 1965), that these insects host a rich collection of primary and putative secondary symbionts, suggesting this group may be a model group for studying complex microbial interactions. Furthermore, from just 16 insect species using community metagenomics and a rapid blast pipeline, we found 12 potential symbiont clades, 9 groups of bacteriophages, 9 putative plant pathogen groups, and many other viruses and parasites, suggesting our approach could be promising if applied on a broader scale. We included membracid root taxa (Aetalion) (Cryan et al., 2000; Cryan et al., 2004; Costa, 2009; Dietrich et al., 2017; Evangelista et al., 2017; Skinner et al., 2020), taxa reported previously as symbiont-rich (Enchophyllum) (Rau, 1943), known virus-vectoring taxa (Micrutalis), and major crop pests (Spissistilus and Ceresa).
Putative plant pathogens included the ‘soft rot’ group of Enterobacteriales, Brenneria sp. and Pectobacterium sp., as well as other gammaproteobacteria such as Enterobacter sp., Pantoea agglomerans, and Serratia sp. Amongst these, Brenneria was most remarkable: Brenneria spp. are not previously known to be transmitted by insects, yet we found sequence matches that occurred at high coverage in two samples (Harmonides sp. membracids). Despite solid phylogenetic placement next to plant pathogenic Brenneria strains, the membracid Brenneria-like 16S rRNA gene sequences were highly divergent, suggesting an increased evolutionary rate, as is common in endosymbionts. However, without further study, we can only speculate on the features of this new Brenneria variant based on the group in which it is found. Importantly, Brenneria are relatives to three of the ‘top 10’ ranked plant pathogens Erwinia, Dickeya, and Pectobacterium (Mansfield et al., 2012) and are pathogens causing numerous diseases (cankers) of woody plants, including the deep bark canker of walnut (Brenneria rubrifasciens) and acute oak decline (Brenneria goodwinii) (Hauben et al., 1998; Bakhshi Ganje et al., 2021). They are noted for producing several unique compounds, such as the red pigment rubrifacine that may contribute to its virulence by inhibiting electron transport in mitochondria. Brenneria species also use sucrose to synthesize levan-type fructans for storage and defense (Liu et al., 2017), which may be of interest in hemipterans whose phloem diet is dominated by sucrose (Shaaban et al., 2020). The Brenneria strain did not, however, closely group with the broadly symbiotic group, Symbiopectobacterium, which includes recently evolved symbionts that independently colonized tissues of various arthropods and nematodes from plant pathogenic ancestors (Martinson et al., 2020; Vallino et al., 2021). Thus, the Brenneria-like sequences may reflect another independent case of a transition from plant pathogen to insect symbiont. Conversely, these sequences could simply be plant pathogens vectored by membracids.
Correlation analyses showed positive associations between the Brenneria and Pectobacterium strains and an insect microvirus, Sulfuriferula, and a parasitoid, but there was no other statistically supported association, suggesting no obvious interaction between primary or secondary symbionts and these Pectobacteriaceae. The Serratia strain found in this study was also correlated with a virus (Indivirus) and parasitoids. In contrast to the Brenneria strains, the Serratia strain showed high similarity (98% 16S rRNA) to Serratia rubidaea, a widespread plant pathogen. Notably, some Serratia species seem to circulate in both plants and hemipterans (Pons et al., 2019), for example, Serratia symbiotica, which likely helps its host digest plant proteins by secreting proteases (Skaljac et al., 2019). However, the Serratia strain occurred in one sample as a short scaffold and so any further analysis would require more data. Pantoea, a different enterobacterial plant pathogen that can be found in various settings including in insects (Walterson and Stavrinides, 2015), was found here only at very low coverage as a short ~85 bp match; therefore, it was not analyzed further. Similarly, while we found a sequence with 98.3% 16S identity to Ralstonia solanacearum, one of the top 10 plant pathogens, we found this in only a single sample (MemA Micrutalis calva from Texas). This Ralstonia was positively correlated with Ralstonia-type phages, as might be expected; however, Ralstonia-type phages were also found in two other U.S. samples (MemE Enchenopa binotata from Illinois and MemM Spissistilus festinus 2 from California), suggesting possible undetected Ralstonia in these samples. Phages of plant pathogens are increasingly becoming of interest for possible biocontrol of bacterial plant disease (Buttimer et al., 2017; Álvarez et al., 2019).
Phytoplasma species are wall-less phloem-infecting plant pathogenic bacteria that require both hemipteran insects and plants in their life cycles and occur in a wide range of woody plants; as such, they might be expected to occur in membracids. Their transmission and life cycle traits, and interactions with existing symbiont have been characterized in leafhoppers (Cicadellidae) and planthoppers (Hogenhout et al., 2008b; Ishii et al., 2013; Weintraub et al., 2019). Because Membracidae is a clade nested within the polyphyletic Cicadellidae (Skinner et al., 2020), authors have speculated that membracids might be expected to be important phytoplasma vectors (Wilson and Weintraub, 2007). Phytoplasmas appear most abundant in tropical and subtropical regions and one study from South America indicated a phytoplasma occurred in the membracid Ceresa (Grosso et al., 2014). Thus, it was surprising to find no 16S rRNA matches to Phytoplasma in these membracids, despite an initial search database of >5000 Phytoplasma 16S genes. Although we found eight samples with scaffolds having short matches to the large Phytoplasma genomic databases (including 1820 genomes), these are not strongly convincing that these membracids vector phytoplasmas. Although the path of these bacteria is through the stylet, intestine, hemolymph, and salivary glands (Weintraub et al., 2019), we expected that our bacteriome-focused dissections and sequencing depth would incidentally include phytoplasmas if they are present. With the caution that Phytoplasma vector status is ambiguous in these data, we note that there were some significantly positive associations in abundance between phytoplasmas and Bombella sp., phage, and other symbionts, suggesting perhaps ecologically common sources of these bacteria or bacterial fragments. Within order Hemiptera, Spiroplasma species reported thus far only from leafhoppers (Weintraub et al., 2019) where they can be either plant pathogens or be vertically transmitted as secondary symbionts, but we found short and low-similarity genome matches to this group as we did for phytoplasmas.
Whereas most viruses of plants have single-stranded RNA genomes and therefore would not be detected in this DNA sequence-based study, we searched for a wide range of DNA plant viruses, expecting to potentially discover some of these, especially those vectored by hemipterans including treehoppers (Mead, 1986; Briddon et al., 1996; Bahder et al., 2016; Varsani et al., 2017; Shafiq et al., 2020). We did not find any geminiviruses, including those related to Topocurvirus which includes the Micrutalis-vectored pseudo-curly top virus, TPCTV, or Grablovirus which includes the Spissistilus-vectored Grapevine red blotch-associated, GRBaV. Although most Geminiviridae are persistent or semipersistent and circulative in their hosts (Shafiq et al., 2020), therefore potentially found in the hemolymph or tissue surrounding the bacteriomes in this study, these are ssDNA viruses that form a dsDNA intermediate in the plant host but may not form a dsDNA phage in the insect, unless they are propagative. For most geminiviruses, it is not clear if they are propagative in the host. We also did not detect Caulimoviridae, which are dsDNA reverse transcribing viruses mostly transmitted by a range of hemipterans (Shafiq et al., 2020), although it is unclear how many of these viruses are circulating and propagative in the insects, suggesting perhaps these plant viruses could be vectored by these treehoppers but not easily detected by these methods.
Among the most abundant presumed secondary symbionts, we found these membracids to be dominated by the genera Arsenophonus, Rickettsia, Sodalis, and Bombella, each of which has members that can be found within plants or causing pathogenicity to plants (Crotti et al., 2016; Chrostek et al., 2017; Gonella et al., 2019). In general, secondary symbionts are diverse functionally, often enabling their hosts to survive a wide range of biotic or abiotic stresses (Gottlieb et al., 2008; Oliver et al., 2012; White et al., 2013; Oliver et al., 2014; Su et al., 2015; Sudakaran et al., 2015; Guidolin et al., 2018; Santos-Garcia et al., 2018; Lemoine et al., 2020). The prevalence, abundance, and phylogenetic analyses presented herein provide some hints, and many open questions, about how these bacteria function in these samples. The high abundance of Arsenophonus in these data has several possible explanations: its 16S rRNA gene occurs in multiple copies per genome (Sorfová et al., 2008) rather than as a single copy as for many endosymbionts including primary symbionts Sulcia and Nasuia, and remarkably Arsenophonus might occur as an abundant hypersymbiont living nested within the cells of the primary symbiont Sulcia (Kobiałka et al., 2016), in which case it could occur at high copy number. Regardless, the abundant and phylogenetically dispersed place of most of these Arsenophonus-like sequences suggest the pattern typical of facultative symbionts (Nováková et al., 2009). The lack of similarity to plant-pathogenic Arsenophonus-like organisms (P. fragariae and Ca. Arsenophonus phytopathogenicus, formerly ‘SMC proteobacteria’) suggests it is unlikely that these strains play this role. Similarly, the lack of similarity to the two probable obligate Arsenophonus-like organisms, Aschnera and ALO-3 (Duron, 2014; Santos-Garcia et al., 2018), suggests no evidence for this role in the sampled membracids. However, the discovery of one variant that clusters at the root of the adelgid symbiont clade (Ca. Hartigia pinicola) as sister to outgroup pathogens suggests a potentially distinct or perhaps parasite-to-commensal transitional function in this organism. Additionally, the typical Arsenophonus strains appear to have at the root of the tree a variant (accession KF751212.1) symbiotic in Stomaphis spp., which are hemipterans specializing on stems and roots of trees, raising the question of this diet as ancient in the Arsenophonus hosts. Ultimately, multi-locus Arsenophonus phylogenomics will be important in uncovering these relationships more accurately, particularly because the 16S rRNA tends to multi-copy in this group (Sorfová et al., 2008) along with comparative omics analysis and detailed microscopy to understand these symbionts, particularly given the observation that Arsenophonus can live within the cells of the primary symbiont Sulcia (Kobiałka et al., 2016).
Our finding of distinct strains of the symbiotic acetic acid bacteria (Acetobacteraceae).
Bombella (formerly Candidatus Parasaccharibacter apium), Asaia, and Saccharibacter floricola, is a novel finding and of special interest in the sugar metabolism of these insects which secrete sugary honeydew to engage trophobiosis with ants and bees. Saccharibacter floricola (Jojima et al., 2004) specifically is bee-associated (Smith et al., 2020). Asaia and Bombella spp. occur in tropical plants and can be plant growth promoters (Crotti et al., 2016), whereas in insects they can stimulate the innate immune system, increase the rate of larval development, and provide insecticide resistance (Chouaia et al., 2010; Chouaia et al., 2012; Mitraka et al., 2013; Comandatore et al., 2021). They can transmit horizontally and vertically, crossing from the gut to the hemolymph and eggs. As symbionts, they likely play a major role in metabolizing sugars to acids (Dong and Zhang, 2006; Crotti et al., 2010); thus, we hypothesize this group to be potentially very important in the observed strong associations between ants and bees and the seven samples of membracids in which we found them. Based on the level of sequence divergence between the membracid Bombella isolates and other Bombella sp. together with the fact that most of the Bombella sequences were similar or identical, we speculate that there may be horizontal transfer of Bombella either between these membracids and their tending hymenopteran insects, or with plants.
Other symbionts exhibited patterns typical for their respective groups, as they occur in other Hemiptera. For example, Rickettsia was dispersed amongst samples and the phylogenetic tree as a typical facultative, partially vertically transferred symbiont, with similar sequences within a host-species. In most cases, Rickettsia is considered parasitic, but there is also evidence that some strains may confer survival benefits (Hendry et al., 2014). Sodalis showed a similar pattern in these data, with phylogenetically dispersed strains, consistent with multiple environmental to secondary symbiont transitions. However, two 16S rRNA gene sequences matching Sodalis, both from the membracid Membracis tectigera (BM13-2), displayed significant sequence divergence, which could suggest a transition in these strains from secondary to primary endosymbiont (Toju et al., 2010), or possibly pseudogenization of 16S rRNA gene copies. Although we found several matches to other symbionts (Wolbachia, Burkholderia, Hamiltonella, Gullanella, and Sulfuriferula sp.), the most notable of these was a high coverage 99.02% 16S rRNA match to a Burkholderia strain that is endophytic, living within the tissues of palm leaves. The strain, initially named Burkholderia sp. JS23, was re-named Chitinasiproducens palmae (Madhaiyan et al., 2020). Its occurrence at high coverage in this membracid sample is a mystery, but interestingly, this same Burkholderiaceae clade includes the Mycoavidus bacteria, which are endohyphal bacteria of the fungus Mortierella elongata (Ohshima et al., 2016).
As expected, all sampled membracids hosted Sulcia, which almost certainly serves to synthesize amino acids missing from the bugs’ phloem diet, but four samples were missing the betaproteobacteria co-symbiont Nasuia, that normally cooperatively synthesizes the remaining amino acids (Bennett and Moran, 2013; Douglas, 2016; Mao et al., 2017). Given the scope of the present study, it remains unclear whether other bacteria or yeast have become replacement symbionts in samples that are missing Nasuia. However, it is noteworthy that we discovered four distinct sequences matching Ophiocordyceps/Hamiltonaphis-like symbionts, three of which occurred in species that were missing Nasuia (BM11 Aetalion reticulum, MemA Micrutalis calva, and BM53 Guyaquila tenuicornis). Numerous studies from aphids, leafhoppers, and other hemipterans suggest that yeast-like symbionts are common and can emerge as co-symbionts or replacement symbionts (Fukatsu and Ishikawa, 1996; Suh et al., 2001; Sacchi et al., 2008; Nishino et al., 2016; Meseguer et al., 2017; Matsuura et al., 2018).
In addition to the Ralstonia phage discussed previously, two other groups of phages were highly abundant: the Hamiltonella-type APSE phages and Wolbachia phage WO, and in general, we found correlation in abundances between bacteria and their presumptive phages, as expected. The APSE phage, well-studied for its parasitoid-protective toxin in Hamiltonella defensa (Oliver et al., 2012; Oliver et al., 2014; Su et al., 2015), particularly in aphids and whiteflies (Rouïl et al., 2020), is commonly integrated into Arsenophonus through lateral gene transfer (Duron, 2014). The infrequency of Hamiltonella and widespread occurrence of APSE in these membracids suggest perhaps Arsenophonus serves as the APSE host in these species. Despite this potential protection from parasitoids conferred by the APSE phage, these membracids showed signs of parasitoid infection, particularly in the level of wasp-specific bracoviruses, especially Cotesia-type bracovirus. These dsDNA viruses act as mutualists for their wasp hosts, contributing to immune suppression of the parasitized insect once injected along with the wasp’s eggs. While bracoviruses are best-known from braconid wasps specifically parasitizing Lepidoptera, discoveries of these virus sequences in Hemiptera previously (Peng et al., 2011; Cheng et al., 2014) and in the present study raise interest in further study of these viruses and related Polydnaviridae in Hemiptera. The high bracovirus levels in these data could also arise from bracovirus horizontal gene transfers into the membracids, as has been observed in other hosts (Cheng et al., 2014; Chevignon et al., 2018). The low levels of parasitoid DNA compared with parasitoid virus DNA in these data could also be explained by our focus on bacteriome tissue, from which most parasitoid wasp DNA, if present, would likely be missed. Finally, despite the opposite biological effects of these two abundant viruses, APSE and bracoviruses, in our data, rather than being negatively correlated, they were significantly positively correlated.
In conclusion, genomic sequence analysis of this kind cannot directly predict insect vector capacity nor microbe pathogenicity; however, these genomic analyses can be invaluable for uncovering previously overlooked microbial associations. Although membracids have been long studied for their exceptional morphological traits, such as the elaborate pronotum, there is scant data on their microbial associations and vectoring potential. These results showed a rich array of microbes and viruses, including plant pathogens and potential allies, painting a preliminary picture of some critical taxa and interactions worth further research. Furthermore, this study generated a large amount of assembled genomic data with thousands scaffolds that are long enough for future in-depth analysis of gene content. We suggest future studies should investigate the prevalence, function, and mechanisms of these potentially interacting microbes, and potential vectored microbes identified here, such as the Brenneria-like bacteria, Serratia, Ralstonia, mycoplasmas or spiroplasmas, and various associated phages. Specifically, co-occurrence patterns leave uncertainty as to role and function of these microbes which should be addressed with phylogenomic analyses, FISH and TEM microscopy, comparative genomics and studies to assess possible HGTs and pseudogenes, and of course wherever possible also controlled infection experiments.
Data Availability Statement
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found below: NCBI accession: PRJNA733715, SAMN19458266-SAMN19458285.
Author Contributions
AB led the design of the study, developed bioinformatics code and pipelines, and drafted the manuscript. MP led the work and field collection in Brazil and assisted with student and laboratory infrastructure and specimen cataloging. MS led the library preparation on Brazilian specimens and assisted in writing the manuscript. All authors contributed to the article and approved the submitted version
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Acknowledgments
We thank Stephen Saltus and other undergraduates in the Brown lab for assistance with membracid field collection, dissection, DNA isolation, and library preparation. We thank John McCutcheon for assistance and support in sequencing of several membracid samples in Montana. Graduate funding support to MS was provided through TTU’s Distinguished Graduate Student Assistantship. We acknowledge undergraduate research support from TTU’s Undergraduate Research Scholars program for various undergraduates who assisted with the project. Support for sequencing, experiments, and analysis to AB were through startup funding from the Department of Biological Sciences at Texas Tech. Travel to Brazil was supported by a SPRINT-FAPESP award to AB and MP at Texas Tech University and University of São Paulo Ribeirão Preto (FAPESP #2016/50492-5). MP also acknowledges Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) and Coordenação de Aperfeiçoamento de Pessoal de Nível Superior - Brasil (CAPES) - Finance Code 001.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fcimb.2021.715998/full#supplementary-material
References
Álvarez, B., López, M. M., Biosca, E. G. (2019). Biocontrol of the Major Plant Pathogen Ralstonia solanacearum in Irrigation Water and Host Plants by Novel Waterborne Lytic Bacteriophages. Front. Microbiol. 10, 2813. doi: 10.3389/fmicb.2019.02813
Bahder, B. W., Zalom, F. G., Jayanth, M., Sudarshana, M. R. (2016). Phylogeny of Geminivirus Coat Protein Sequences and Digital PCR Aid in Identifying Spissistilus festinus as a Vector of Grapevine Red Blotch-Associated Virus. Phytopathology 106, 1223–1230. doi: 10.1094/PHYTO-03-16-0125-FI
Bailey, J. C. (1975). Three-Cornered Alfalfa Hoppers (Homoptera: Membracidae): Effect of Four Population Levels on Soybeans. J. Kans. Entomol. Soc. 48 (4), 519–520.
Bakhshi Ganje, M., Mackay, J., Nicolaisen, M., Shams-Bakhsh, M. (2021). Comparative Genomics, Pangenome, and Phylogenomic Analyses of Brenneria Spp., and Delineation of Brenneria izadpanahii Sp. Nov. Phytopathology 111, 78–95. doi: 10.1094/PHYTO-04-20-0129-FI
Bankevich, A., Nurk, S., Antipov, D., Gurevich, A. A., Dvorkin, M., Kulikov, A. S., et al. (2012). SPAdes: A New Genome Assembly Algorithm and its Applications to Single-Cell Sequencing. J. Comput. Biol. 19, 455–477. doi: 10.1089/cmb.2012.0021
Bell-Roberts, L., Douglas, A. E., Werner, G. D. A. (2019). Match and Mismatch Between Dietary Switches and Microbial Partners in Plant Sap-Feeding Insects. Proc. R. Soc B Biol. Sci. 286, 20190065. doi: 10.1098/rspb.2019.0065
Benjamini, Y., Hochberg, Y. (1995). Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. J. R. Stat. Soc. Series B Stat. Methodol. 57.1, 289–300
Bennett, G. M., Moran, N. A. (2013). Small, Smaller, Smallest: The Origins and Evolution of Ancient Dual Symbioses in a Phloem-Feeding Insect. Genome Biol. Evol. 5, 1675–1688. doi: 10.1093/gbe/evt118
Bolger, A. M., Lohse, M., Usadel, B. (2014). Trimmomatic: A Flexible Trimmer for Illumina Sequence Data. Bioinformatics 30, 2114–2120. doi: 10.1093/bioinformatics/btu170
Briddon, R. W., Bedford, I. D., Tsai, J. H., Markham, P. G. (1996). Analysis of the Nucleotide Sequence of the Treehopper-Transmitted Geminivirus, Tomato Pseudo-Curly Top Virus, Suggests a Recombinant Origin. Virology 219, 387–394. doi: 10.1006/viro.1996.0264
Brown, A. M. V., Huynh, L. Y., Bolender, C. M., Nelson, K. G., McCutcheon, J. P. (2014). Population Genomics of a Symbiont in the Early Stages of a Pest Invasion. Mol. Ecol. 23, 1516–1530. doi: 10.1111/mec.12366
Buttimer, C., McAuliffe, O., Ross, R. P., Hill, C., O’Mahony, J., Coffey, A. (2017). Bacteriophages and Bacterial Plant Diseases. Front. Microbiol. 8, 34. doi: 10.3389/fmicb.2017.00034
Calcagnile, M., Tredici, S. M., Talà, A., Alifano, P. (2019). Bacterial Semiochemicals and Transkingdom Interactions With Insects and Plants. Insects 10, 441. doi: 10.3390/insects10120441
Camacho, C., Coulouris, G., Avagyan, V., Ma, N., Papadopoulos, J., Bealer, K., et al. (2009). BLAST+: Architecture and Applications. BMC Bioinf. 10, 1–9. doi: 10.1186/1471-2105-10-421
Canedo-Júnior, E. O., Monteiro, Â.B., de Queiroz, A. C. M., Silva, G. S. (2019). Aphid-Tending Ants and Their Effects on Natural Enemies Used in the Biological Control. In: Natural Enemies of Insect Pests in Neotropical Agroecosystems. (pp. 523–533). Springer, Cham. doi: 10.1007/978-3-030-24733-1_42
Cheng, R.-L., Xi, Y., Lou, Y.-H., Wang, Z., Xu, J.-Y., Xu, H.-J., et al. (2014). Brown Planthopper Nudivirus DNA Integrated in Its Host Genome. J. Virol. 88, 5310–5318. doi: 10.1128/jvi.03166-13
Chevignon, G., Periquet, G., Gyapay, G., Vega-czarny, N., Musset, K., Drezen, J., et al. (2018). Cotesia congregata Bracovirus Circles Encoding PTP and Ankyrin Genes Integrate Into the DNA of Parasitized Manduca sexta Hemocytes. J. Virol. 92, 1–21. doi: 10.1128/JVI.00438-18
Chouaia, B., Rossi, P., Epis, S., Mosca, M., Ricci, I., Damiani, C., et al. (2012). Delayed Larval Development in Anopheles Mosquitoes Deprived of Asaia Bacterial Symbionts. BMC Microbiol. 12, 1–8. doi: 10.1186/1471-2180-12-S1-S2
Chouaia, B., Rossi, P., Montagna, M., Ricci, I., Crotti, E., Damiani, C., et al. (2010). Molecular Evidence for Multiple Infections as Revealed by Typing of Asaia Bacterial Symbionts of Four Mosquito Species. Appl. Environ. Microbiol. 76, 7444–7450. doi: 10.1128/AEM.01747-10
Chrostek, E., Pelz-Stelinski, K., Hurst, G. D. D., Hughes, G. L. (2017). Horizontal Transmission of Intracellular Insect Symbionts via Plants. Front. Microbiol. 8, 2237. doi: 10.3389/fmicb.2017.02237
Comandatore, F., Damiani, C., Cappelli, A., Ribolla, P. E. M., Gasperi, G., Gradoni, F., et al. (2021). Phylogenomics Reveals That Asaia Symbionts From Insects Underwent Convergent Genome Reduction, Preserving an Insecticide-Degrading Gene. MBio 12, 1–12. doi: 10.1128/mBio.00106-21
Costa, J. F. (2009). Membrácidos (Hemiptera: Membracidae) De Los Bosques Nublados Del Parque Nacional Del Manu (PNM), Cusco, Perú. Bol. del Mus Entomol. la Univ. del Val 10, 8–13.
Crotti, E., Chouaia, B., Alma, A., Favia, G., Bandi, C., Bourtzis, K., et al. (2016). “Acetic Acid Bacteria as Symbionts of Insects”, in Acetic Acid Bacteria. Eds. Matsushita, K., Toyama, H., Tonouchi, N., Okamoto-Kainuma, A., Springer, 121–142.
Crotti, E., Rizzi, A., Chouaia, B., Ricci, I., Favia, G., Alma, A., et al. (2010). Acetic Acid Bacteria, Newly Emerging Symbionts of Insects. Appl. Environ. Microbiol. 76, 6963–6970. doi: 10.1128/AEM.01336-10
Cryan, J. R., Wiegmann, B. M., Deitz, L. L., Dietrich, C. H. (2000). Phylogeny of the Treehoppers (Insecta: Hemiptera: Membracidae): Evidence From Two Nuclear Genes. Mol. Phylogenet. Evol. 17, 317–334. doi: 10.1006/mpev.2000.0832
Cryan, J. R., Wiegmann, B. M., Deitz, L. L., Dietrich, C. H., Whiting, M. F. (2004). Treehopper Trees: Phylogeny of Membracidae (Hemiptera: Cicadomorpha: Membracoidea) Based on Molecules and Morphology. Syst. Entomol. 29, 441–454. doi: 10.1111/j.0307-6970.2004.00260.x
Deitz, L. L., Wallace, M. S. (2012). Richness of the Nearctic Treehopper Fauna (Hemiptera: Aetalionidae and Membracidae). Zootaxa 26, 1–26. doi: 10.11646/zootaxa.3423.1.1
Delabie, J. H. C. (2001). Trophobiosis Between Formicidae and Hemiptera (Sternorrhyncha and Auchenorrhyncha): An Overview. Neotrop Entomol. 30, 501–516. doi: 10.1590/s1519-566x2001000400001
Dietrich, C. H., Allen, J. M., Lemmon, A. R., Lemmon, E. M., Takiya, D. M., Evangelista, O., et al. (2017). Anchored Hybrid Enrichment-Based Phylogenomics of Leafhoppers and Treehoppers (Hemiptera: Cicadomorpha: Membracoidea). Insect Syst. Divers. 1, 57–72. doi: 10.1093/isd/ixx003
Dong, L. Q., Zhang, K. Q. (2006). Microbial Control of Plant-Parasitic Nematodes: A Five-Party Interaction. Plant Soil 288, 31–45. doi: 10.1007/s11104-006-9009-3
dos Santos, C. F., Halinski, R., de Souza dos Santos, P. D., Almeida, E. A. B., Blochtein, B. (2019). Looking Beyond the Flowers: Associations of Stingless Bees With Sap-Sucking Insects. Sci. Nat. 106, 12. doi: 10.1007/s00114-019-1608-y
Douglas, A. E. (2016). How Multi-Partner Endosymbioses Function. Nat. Rev. Micro 14, 731–743. doi: 10.1038/nrmicro.2016.151
Duron, O. (2014). Arsenophonus Insect Symbionts are Commonly Infected With APSE, a Bacteriophage Involved in Protective Symbiosis. FEMS Microbiol. Ecol. 90, 184–194. doi: 10.1111/1574-6941.12381
Evangelista, O., Sakakibara, A. M., Cryan, J. R., Urban, J. M. (2017). A Phylogeny of the Treehopper Subfamily Heteronotinae Reveals Convergent Pronotal Traits (Hemiptera: Auchenorrhyncha: Membracidae). Syst. Entomol. 42, 410–428. doi: 10.1111/syen.12221
Fischer, C. Y., Lognay, G. C., Detrain, C., Heil, M., Grigorescu, A., Sabri, A., et al. (2015). Bacteria may Enhance Species Association in an Ant–Aphid Mutualistic Relationship. Chemoecology 25, 223–232. doi: 10.1007/s00049-015-0188-3
Fukatsu, T., Ishikawa, H. (1996). Phylogenetic Position of Yeast-Like Symbiont of Hamiltonaphis styraci (Homoptera, Aphididae) Based on 18S rDNA Sequence. Insect Biochem. Mol. Biol. 26, 383–388. doi: 10.1016/0965-1748(95)00105-0
Gonella, E., Tedeschi, R., Crotti, E., Alma, A. (2019). Multiple Guests in a Single Host: Interactions Across Symbiotic and Phytopathogenic Bacteria in Phloem-Feeding Vectors – A Review. Entomol. Exp. Appl. 167, 171–185. doi: 10.1111/eea.12766
Gottlieb, Y., Ghanim, M., Gueguen, G., Kontsedalov, S., Vavre, F., Fleury, F., et al. (2008). Inherited Intracellular Ecosystem: Symbiotic Bacteria Share Bacteriocytes in Whiteflies. FASEB J. 22, 2591–2599. doi: 10.1096/fj.07-101162
Grosso, T. P., Conci, L. R., Saavedra Pons, A. B., De Remes Lenicov, A. M. M., Virla, E. G. (2014). First Biological Data of Ceresa nigripectus (Hemiptera: Membracidae), a Common Treehopper on Alfalfa Crops in Argentina. Florida Entomol. 97, 1766–1773. doi: 10.1653/024.097.0452
Guidolin, A. S., Cataldi, T. R., Labate, C. A., Francis, F., Cônsoli, F. L. (2018). Spiroplasma Affects Host Aphid Proteomics Feeding on Two Nutritional Resources. Sci. Rep. 8, 1–13. doi: 10.1038/s41598-018-20497-9
Gurevich, A., Saveliev, V., Vyahhi, N., Tesler, G. (2013). QUAST: Quality Assessment Tool for Genome Assemblies. Bioinformatics 29, 1072–1075. doi: 10.1093/bioinformatics/btt086
Hauben, L., Moore, E. R. B., Vauterin, L., Steenackers, M., Mergaert, J., Verdonck, L., et al. (1998). Phylogenetic Position of Phytopathogens Within the Enterobacteriaceae. Syst. Appl. Microbiol. 21, 384–397. doi: 10.1016/S0723-2020(98)80048-9
Heck, M. (2018). Insect Transmission of Plant Pathogens: A Systems Biology Perspective. mSystems 3, e00168–e00117. doi: 10.1128/msystems.00168-17
Hendry, T. A., Hunter, M. S., Baltrus, D. A. (2014). The Facultative Symbiont Rickettsia Protects an Invasive Whitefly Against Entomopathogenic Pseudomonas syringae Strains. Appl. Environ. Microbiol. 80, 7161–7168. doi: 10.1128/AEM.02447-14
Hogenhout, S. A., Ammar, E. D., Whitfield, A. E., Redinbaugh, M. G. (2008a). Insect Vector Interactions With Persistently Transmitted Viruses. Annu. Rev. Phytopathol. 46, 327–359. doi: 10.1146/annurev.phyto.022508.092135
Hogenhout, S. A., Oshima, K., Ammar, E. D., Kakizawa, S., Kingdom, H. N., Namba, S. (2008b). Phytoplasmas: Bacteria That Manipulate Plants and Insects. Mol. Plant Pathol. 9, 403–423. doi: 10.1111/j.1364-3703.2008.00472.x
Huelsenbeck, J. P., Ronquist, F. (2001). MRBAYES: Bayesian Inference of Phylogenetic Trees. Bioinformatics 17, 754–755. doi: 10.1093/bioinformatics/17.8.754
Hu, K., Yuan, F., Dietrich, C. H., Yuan, X. Q. (2019). Structural Features and Phylogenetic Implications of Four New Mitogenomes of Centrotinae (Hemiptera: Membracidae). Int. J. Biol. Macromol. 139, 1018–1027. doi: 10.1016/j.ijbiomac.2019.08.064
Ibarra-Isassi, J., Oliveira, P. S. (2018). Indirect Effects of Mutualism: Ant–Treehopper Associations Deter Pollinators and Reduce Reproduction in a Tropical Shrub. Oecologia 186, 691–701. doi: 10.1007/s00442-017-4045-7
Ishii, Y., Matsuura, Y., Kakizawa, S., Nikoh, N., Fukatsua, T., Fukatsu, T., et al. (2013). Diversity of Bacterial Endosymbionts Associated With Macrosteles Leafhoppers Vectoring Phytopathogenic Phytoplasmas. Appl. Environ. Microbiol. 79, 5013–5022. doi: 10.1128/AEM.01527-13
Jojima, Y., Mihara, Y., Suzuki, S., Yokozeki, K., Yamanaka, S., Fudou, R. (2004). Saccharibacter floricola Gen. Nov., Sp. Nov., a Novel Osmophilic Acetic Acid Bacterium Isolated From Pollen. Int. J. Syst. Evol. Microbiol. 54, 2263–2267. doi: 10.1099/ijs.0.02911-0
Katoh, K., Standley, D. M. (2013). MAFFT Multiple Sequence Alignment Software Version 7: Improvements in Performance and Usability. Mol. Biol. Evol. 30, 772–780. doi: 10.1093/molbev/mst010
Klimes, P., Borovanska, M., Plowman, N. S., Leponce, M. (2018). How Common is Trophobiosis With Hoppers (Hemiptera: Auchenorrhyncha) Inside Ant Nests (Hymenoptera: Formicidae)? Novel Interactions From New Guinea and a Worldwide Overview. Myrmecological News 26, 31–45.
Kobiałka, M., Michalik, A., Świerczewski, D., Szklarzewicz, T. (2019). Complex Symbiotic Systems of Two Treehopper Species: Centrotus cornutus (Linnaeus 1758) and Gargara genistae (Fabricius 1775) (Hemiptera: Cicadomorpha: Membracoidea: Membracidae). Protoplasma. 257 (3), 819–831 doi: 10.1007/s00709-019-01466-z
Kobiałka, M., Michalik, A., Walczak, M., Junkiert, Ł., Szklarzewicz, T. (2016). Sulcia Symbiont of the Leafhopper Macrosteles laevis (Ribaut 1927) (Insecta, Hemiptera, Cicadellidae: Deltocephalinae) Harbors Arsenophonus Bacteria. Protoplasma 253, 903–912. doi: 10.1007/s00709-015-0854-x
Lemoine, M. M., Engl, T., Kaltenpoth, M. (2020). Microbial Symbionts Expanding or Constraining Abiotic Niche Space in Insects. Curr. Opin. Insect Sci. 39, 14–20. doi: 10.1016/j.cois.2020.01.003
Leroy, P. D., Sabri, A., Heuskin, S., Thonart, P., Lognay, G., Verheggen, F. J., et al. (2011). Microorganisms From Aphid Honeydew Attract and Enhance the Efficacy of Natural Enemies. Nat. Commun. 2, 1–7. doi: 10.1038/ncomms1347
Li, Q., Fan, J., Sun, J., Wang, M. Q., Chen, J. (2018). Plant-Mediated Horizontal Transmission of Hamiltonella defensa in the Wheat Aphid Sitobion miscanthi. J. Agric. Food Chem. 66, 13367–13377. doi: 10.1021/acs.jafc.8b04828
Li, H., Handsaker, B., Wysoker, A., Fennell, T., Ruan, J., Homer, N., et al. (2009). The Sequence Alignment/Map Format and SAMtools. Bioinformatics 25, 2078–2079. doi: 10.1093/bioinformatics/btp352
Liu, Q., Yu, S., Zhang, T., Jiang, B., Mu, W. (2017). Efficient Biosynthesis of Levan From Sucrose by a Novel Levansucrase From Brenneria goodwinii. Carbohydr. Polym. 157, 1732–1740. doi: 10.1016/j.carbpol.2016.11.057
Madhaiyan, M., See-Too, W. S., Ee, R., Saravanan, V. S., Wirth, J. S., Alex, T. H. H., et al. (2020). Chitinasiproducens palmae Gen. Nov., Sp. Nov., a New Member of the Family Burkholderiaceae Isolated From Leaf Tissues of Oil Palm (Elaeis guineensis Jacq.). Int. J. Syst. Evol. Microbiol. 70, 2640–2647. doi: 10.1099/ijsem.0.004084
Mansfield, J., Genin, S., Magori, S., Citovsky, V., Sriariyanum, M., Ronald, P., et al. (2012). Top 10 Plant Pathogenic Bacteria in Molecular Plant Pathology. Mol. Plant Pathol. 13, 614–629. doi: 10.1111/j.1364-3703.2012.00804.x
Mao, M., Yang, X., Poff, K., Bennett, G. (2017). Comparative Genomics of the Dual-Obligate Symbionts From the Treehopper, Entylia carinata (Hemiptera: Membracidae), Provide Insight Into the Origins and Evolution of an Ancient Symbiosis. Genome Biol. Evol. 9, 1803–1815. doi: 10.1093/gbe/evx134
Martinson, V. G., Gawryluk, R. M. R., Gowen, B. E., Curtis, C. I., Jaenike, J., Perlman, S. J. (2020). Multiple Origins of Obligate Nematode and Insect Symbionts by a Clade of Bacteria Closely Related to Plant Pathogens. Proc. Natl. Acad. Sci. U. S. A. 117, 31979–31986. doi: 10.1073/pnas.2000860117
Matsuura, Y., Moriyama, M., Łukasik, P., Vanderpool, D., Tanahashi, M., Meng, X.-Y., et al. (2018). Recurrent Symbiont Recruitment From Fungal Parasites in Cicadas. Proc. Natl. Acad. Sci. U. S. A. 115, 201803245. doi: 10.1073/pnas.1803245115
Mead, F. W. (1986). Micrutalis Treehoppers and Pseudo-Curly Top in Florida (Homoptera: Membracidae). Florida Dep Agric. Consum Serv Div. Plant Ind. Entomol. Circ. 283, 1–4.
Meisch, M. V., Randolph, N. M. (1965). Life-History Studies and Rearing Techniques for the Three-Cornered Alfalfa Hopper. J. Econ. Entomol. 58, 1057–1059. doi: 10.1093/jee/58.6.1057
Meseguer, A. S., Manzano-Marín, A., Coeur d’Acier, A., Clamens, A. L., Godefroid, M., Jousselin, E. (2017). Buchnera has Changed Flatmate But the Repeated Replacement of Co-Obligate Symbionts is Not Associated With the Ecological Expansions of Their Aphid Hosts. Mol. Ecol. 26, 2363–2378. doi: 10.1111/mec.13910
Mitraka, E., Stathopoulos, S., Siden-Kiamos, I., Christophides, G. K., Louis, C. (2013). Asaia Accelerates Larval Development of Anopheles gambiae. Pathog. Glob. Health 107, 305–311. doi: 10.1179/2047773213Y.0000000106
Müller, H. J. (1962). Neuere Vorstellungen Über Verbreitung Und Phylogenie Der Endosymbiosen Der Zikaden. Z. für Morphol und Ökologie der Tiere 51, 190–210. doi: 10.1007/BF00409635
Nishino, T., Tanahashi, M., Lin, C.-P., Koga, R., Fukatsu, T. (2016). Fungal and Bacterial Endosymbionts of Eared Leafhoppers of the Subfamily Ledrinae (Hemiptera: Cicadellidae). Appl. Entomol. Zool. 51, 465–477. doi: 10.1007/s13355-016-0422-7
Nováková, E., Hypša, V., Moran, N. A. (2009). Arsenophonus, an Emerging Clade of Intracellular Symbionts With a Broad Host Distribution. BMC Microbiol. 9, 1–14. doi: 10.1186/1471-2180-9-143
Nurk, S., Meleshko, D., Korobeynikov, A., Pevzner, P. A. (2017). MetaSPAdes: A New Versatile Metagenomic Assembler. Genome Res. 27, 824–834. doi: 10.1101/gr.213959.116
Ohshima, S., Sato, Y., Fujimura, R., Takashima, Y., Hamada, M., Nishizawa, T., et al. (2016). Mycoavidus cysteinexigens Gen. Nov., Sp. Nov., an Endohyphal Bacterium Isolated From a Soil Isolate of the Fungus Mortierella elongata. Int. J. Syst. Evol. Microbiol. 66, 2052–2057. doi: 10.1099/ijsem.0.000990
Oliver, K. M., Noge, K., Huang, E. M., Campos, J. M., Becerra, J. X., Hunter, M. S. (2012). Parasitic Wasp Responses to Symbiont-Based Defense in Aphids. BMC Biol. 10:11. doi: 10.1186/preaccept-4284799146146604
Oliver, K. M., Smith, A. H., Russell, J. A. (2014). Defensive Symbiosis in the Real World - Advancing Ecological Studies of Heritable, Protective Bacteria in Aphids and Beyond. Funct. Ecol. 28, 341–355. doi: 10.1111/1365-2435.12133
Peng, X., Zha, W., He, R., Lu, T., Zhu, L., Han, B., et al. (2011). Pyrosequencing the Midgut Transcriptome of the Brown Planthopper, Nilaparvata lugens. Insect Mol. Biol. 20, 745–762. doi: 10.1111/j.1365-2583.2011.01104.x
Pons, I., Renoz, F., Noël, C., Hance, T. (2019). Circulation of the Cultivable Symbiont Serratia symbiotica in Aphids is Mediated by Plants. Front. Microbiol. 10, 764. doi: 10.3389/fmicb.2019.00764
Rau, A. (1943). Symbiose Und Symbiontenerwerb Bei Den Membraciden (Homoptera-Cicadina). Aus dem Zool. Inst. der Univ. Leipzig 39, 369–522. doi: 10.1007/BF00684470
Ronquist, F., Teslenko, M., van der Mark, P., Ayres, D. L., Darling, A., Höhna, S., et al. (2012). MrBayes 3.2: Efficient Bayesian Phylogenetic Inference and Model Choice Across a Large Model Space. Syst. Biol. 61, 539–542. doi: 10.1093/sysbio/sys029
Rouïl, J., Jousselin, E., Coeur d’acier, A., Cruaud, C., Manzano-Marın, A. (2020). The Protector Within: Comparative Genomics of APSE Phages Across Aphids Reveals Rampant Recombination and Diverse Toxin Arsenals. Genome Biol. Evol. 12, 878–889. doi: 10.1093/gbe/evaa089
Sacchi, L., Genchi, M., Clementi, E., Bigliardi, E., Avanzati, A. M., Pajoro, M., et al. (2008). Multiple Symbiosis in the Leafhopper Scaphoideus titanus (Hemiptera: Cicadellidae): Details of Transovarial Transmission of Cardinium Sp. And Yeast-Like Endosymbionts. Tissue Cell 40, 231–242. doi: 10.1016/j.tice.2007.12.005
Santos-Garcia, Di., Juravel, K., Freilich, S., Zchori-Fein, E., Latorre, A., Moya, A., et al. (2018). To B or Not to B: Comparative Genomics Suggests Arsenophonus a Source of B Vitamins in Whiteflies. Front. Microbiol. 9, 2254. doi: 10.3389/fmicb.2018.02254
Scholthof, K. B. G., Adkins, S., Czosnek, H., Palukaitis, P., Jacquot, E., Hohn, T., et al. (2011). Top 10 Plant Viruses in Molecular Plant Pathology. Mol. Plant Pathol. 12, 938–954. doi: 10.1111/j.1364-3703.2011.00752.x
Shaaban, B., Seeburger, V., Schroeder, A., Lohaus, G. (2020). Sugar, Amino Acid and Inorganic Ion Profiling of the Honeydew From Different Hemipteran Species Feeding on Abies Alba and Picea Abies. PloS One 15, 1–17. doi: 10.1371/journal.pone.0228171
Shafiq, M., Qurashi, F., Mushtaq, S., Hussain, M., Hameed, A., Saleem Haider, M. (2020). “DNA Plant Viruses: Biochemistry, Replication, and Molecular Genetics,” in Applied Plant Virology: Advances, Detection, and Antiviral Strategies. Academic Press. 169–182. doi: 10.1016/b978-0-12-818654-1.00013-x
Shamim, G., Sharma, K. K., Ramani, R. (2019). Isolation and Identification of Culturable Bacteria From Honeydew of Indian Lac Insect, Kerria lacca (Kerr) (Hemiptera: Tachardiidae). Meta Gene 19, 10–14. doi: 10.1016/j.mgene.2018.09.010
Skaljac, M., Vogel, H., Wielsch, N., Mihajlovic, S., Vilcinskas, A. (2019). Transmission of a Protease-Secreting Bacterial Symbiont Among Pea Aphids via Host Plants. Front. Physiol. 10, 438. doi: 10.3389/fphys.2019.00438
Skinner, R. K., Dietrich, C. H., Walden, K. K. O., Gordon, E., Sweet, A. D., Podsiadlowski, L., et al. (2020). Phylogenomics of Auchenorrhyncha (Insecta: Hemiptera) Using Transcriptomes: Examining Controversial Relationships via Degeneracy Coding and Interrogation of Gene Conflict. Syst. Entomol. 45, 85–113. doi: 10.1111/syen.12381
Smith, E. A., Vuong, H. Q., Miller, D. L., Parish, A. J., McFrederick, Q. S., Newton, I. L. G. (2020). Draft Genome Sequences of Four Saccharibacter Sp. Strains Isolated From Native Bees. Microbiol. Resour. Announc 9, 9–10. doi: 10.1128/mra.00022-20
Sorfová, P., Skeríková, A., Hypsa, V. (2008). An Effect of 16S rRNA Intercistronic Variability on Coevolutionary Analysis in Symbiotic Bacteria: Molecular Phylogeny of Arsenophonus triatominarum. Syst. Appl. Microbiol. 31, 88–100. doi: 10.1016/j.syapm.2008.02.004
Stamatakis, A. (2014). RAxML Version 8: A Tool for Phylogenetic Analysis and Post-Analysis of Large Phylogenies. Bioinformatics 30, 1312–1313. doi: 10.1093/bioinformatics/btu033
Sudakaran, S., Kost, C., Kaltenpoth, M. (2017). Symbiont Acquisition and Replacement as a Source of Ecological Innovation. Trends Microbiol. 25, 375–390. doi: 10.1016/j.tim.2017.02.014
Sudakaran, S., Retz, F., Kikuchi, Y., Kost, C., Kaltenpoth, M. (2015). Evolutionary Transition in Symbiotic Syndromes Enabled Diversification of Phytophagous Insects on an Imbalanced Diet. ISME J. 9, 2587–2604. doi: 10.1038/ismej.2015.75
Suh, S. O., Noda, H., Blackwell, M. (2001). Insect Symbiosis: Derivation of Yeast-Like Endosymbionts Within an Entomopathogenic Filamentous Lineage. Mol. Biol. Evol. 18, 995–1000. doi: 10.1093/oxfordjournals.molbev.a003901
Su, Q., Oliver, K. M., Xie, W., Wu, Q., Wang, S., Zhang, Y. (2015). The Whitefly-Associated Facultative Symbiont Hamiltonella defensa Suppresses Induced Plant Defences in Tomato. Funct. Ecol. 29, 1007–1018. doi: 10.1111/1365-2435.12405
Toju, H., Hosokawa, T., Koga, R., Nikoh, N., Meng, X. Y., Kimura, N., et al. (2010). “Candidatus Curculioniphilus Buchneri”, a Novel Clade of Bacterial Endocellular Symbionts From Weevils of the Genus Curculio. Appl. Environ. Microbiol. 76, 275–282. doi: 10.1128/AEM.02154-09
Vallino, M., Rossi, M., Ottati, S., Martino, G., Galetto, L., Marzachì, C., et al. (2021). Bacteriophage-Host Association in the Phytoplasma Insect vector Euscelidius variegatus. Pathogens 10, 612. doi: 10.3390/pathogens10050612
Varsani, A., Roumagnac, P., Fuchs, M., Navas-Castillo, J., Moriones, E., Idris, A., et al. (2017). Capulavirus and Grablovirus: Two New Genera in the Family Geminiviridae. Arch. Virol. 162, 1819–1831. doi: 10.1007/s00705-017-3268-6
Walterson, A. M., Stavrinides, J. (2015). Pantoea: Insights Into a Highly Versatile and Diverse Genus Within the Enterobacteriaceae. FEMS Microbiol. Rev. 39, 968–984. doi: 10.1093/femsre/fuv027
Weintraub, P. G., Trivellone, V., Krüger, K. (2019). “The Biology and Ecology of Leafhopper Transmission of Phytoplasmas”. in Phytoplasmas: Plant Pathogenic Bacteria-II (pp. 27–51). Singapore: Springer. doi: 10.1007/978-981-13-2832-9_2
White, J. A., Giorgini, M., Strand, M. R., Pennacchio, F. (2013). “Arthropod Endosymbiosis and Evolution”, in Arthropod Biology and Evolution: Molecules, Development, Morphology, Berlin, Heidelberg: Springer. 441–477. doi: 10.1007/978-3-642-36160-9
Wilson, M. R., Weintraub, P. G. (2007). An Introduction to Auchenorrhyncha Phytoplasma Vectors. Bull. Insectology 60, 177–178.
Wu, D., Daugherty, S. C., Van Aken, S. E., Pai, G. H., Watkins, K. L., Khouri, H., et al. (2006). Metabolic Complementarity and Genomics of the Dual Bacterial Symbiosis of Sharpshooters. PloS Biol. 4, 1079–1092. doi: 10.1002/adsc.200505252
Keywords: membracid, treehopper, endosymbiont, plant pathogen, phage, metagenomics
Citation: Salazar MM, Pupo MT and Brown AMV (2021) Co-Occurrence of Viruses, Plant Pathogens, and Symbionts in an Underexplored Hemipteran Clade. Front. Cell. Infect. Microbiol. 11:715998. doi: 10.3389/fcimb.2021.715998
Received: 27 May 2021; Accepted: 30 July 2021;
Published: 26 August 2021.
Edited by:
Benjamin Makepeace, University of Liverpool, United KingdomReviewed by:
Davide Sassera, University of Pavia, ItalyAlison Ravenscraft, University of Texas at Arlington, United States
Copyright © 2021 Salazar, Pupo and Brown. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Amanda M. V. Brown, YW1hbmRhLm12LmJyb3duQHR0dS5lZHU=