 Breno Luiz Pimenta1,2
Breno Luiz Pimenta1,2 Tatiana Prata Menezes1,3*
Tatiana Prata Menezes1,3* Vitória Louise1,2,3Fernanda Carolina Ribeiro Dias4Bianca Alves Almeida Machado1,5Lais Ribeiro1,2
Vitória Louise1,2,3Fernanda Carolina Ribeiro Dias4Bianca Alves Almeida Machado1,5Lais Ribeiro1,2 Kelerson Mauro de Castro Pinto1,6
Kelerson Mauro de Castro Pinto1,6 Andre Talvani1,2,3*
Andre Talvani1,2,3*- 1Laboratory of Immunobiology of Inflammation, Department of Biological Sciences, Federal University of Ouro Preto, Ouro Preto, Minas Gerais, Brazil
- 2Infectious Diseases and Tropical Medicine Post-Graduate Program, School of Medicine, Federal University of Minas Gerais, Belo Horizonte, Minas Gerais, Brazil
- 3Health and Nutrition Post-Graduate Program, School of Nutrition, Federal University of Ouro Preto, Ouro Preto, Minas Gerais, Brazil
- 4Department of Veterinary, Federal Rural University of Pernambuco, Recife, Pernambuco, Brazil
- 5School of Medicine, Federal University of Ouro Preto, Ouro Preto, Minas Gerais, Brazil
- 6Laboratory of Physiology of Exercise, School of Physical Education, Federal University of Ouro Preto, Ouro Preto, Minas Gerais, Brazil
Introduction: Chagas’ disease is a tropical neglected illness caused by Trypanosoma cruzi and remains one of the most significant causes of morbidity and mortality in South and Central Americas. The disease is caused by a moderate to intense and persistent inflammatory response characterized by local upregulated expression and production of inflammatory mediators that favors the activation and recruitment of distinct cells of the immune system into different tissues to eliminate the parasites. Theracurmin is a curcumin’s derived formulation of nanoparticles. Its anti-inflammatory properties make this bioactive compound a mitigating factor in pathological cases after an overwhelming inflammatory response.
Methods: Our research focused on the testicular investigation in 28 mice infected by 103 trypomastigote forms of Colombian strain of T. cruzi and preventively treated with Theracurmin. The mice were treated with 30 mg/Kg of Theracurmin during the period of 30 days. At the 30th day post infection animals were euthanized, and its testicles were collected to morphological and immunological assays.
Results: The animals infected and treated with Theracurmin presented a reduction in the testicular levels of IL-15 and IL-6. The volume density (%) of the tunica propria was also higher in all infected animals, but Theracurmin decreased this parameter in the treated animals. In the intertubular area, the percentage of some intertubular components was decreased in the infected animals such as the percentage and volume of Leydig cells, connective tissue, and macrophages.
Discussion: Furthermore, our data pointed to the daily use of Theracurmin in the diet as a protective element of the testicular function.
1 Introduction
Chagas disease is caused by the flagellated protozoan Trypanosoma cruzi and affects 6 to 7 million people around the world (World Health Organization, 2022), being endemic from the south of Argentina and Chile though the north of Mexico (Pérez-molina and Molina, 2018). This parasite has several routes of infection, and its main routes are oral and vectorial (during the repast of the triatomine insect while feeding), beyond another ways of transmission such as blood transfusion (mainly in non-endemic countries), laboratory accidents, congenital and sexual transmission, which is supported by experimental studies (Pérez-molina and Molina, 2018; Almeida et al., 2019; Zapparoli et al., 2022).
In the rodents, T. cruzi has a different tissue tropism after its infection depending on factors such as the load and genetic background of the parasites and the genetic/immune response related to the mammalian host (Andrade et al., 2010; Medina et al., 2018). The T. cruzi tissue tropism is described as heart, skeletal muscle, and tissues from the gastric system (Esper et al., 2015; Weaver et al., 2019; Hossain et al., 2020). However, this parasite infects a diverse range of tissues, including those from reproductive tract (Almeida et al., 2019). However, in animals, T. cruzi was found in the testicles and its associated components, such as vas deferens, epididymis, seminal vesicle, prostate, and penis in the acute phase of the experimental infection (Lenzi et al., 1998) and, sexual transmission was described in mice by Martin et al. (2015). These studies opened new questions concerning the capacity of T. cruzi to surpass the male reproductive epithelial tissue and induce infection in the female organism. In the reproductive tissue, T. cruzi infects cells and cause an inflammation responsible for damages on the reproductive condition and/or the ease of dissemination of the protozoan among infected individuals.
The infection’s treatment using nitrocompounds such as benznidazole is more effective during the acute than in chronic phase and, this effect is also dependent on the genetic background of the protozoan and of the host, which defines its virulence, infectivity and pathogenesis (Cunha-Neto and Chevillard, 2014; Zingales and Bartholomeu, 2022). Since most of the cases are diagnosticated during the chronic phase and, at this time, the trypanocidal effectiveness of benznidazole is partial without reduction of cardiac clinical disturbances (Morillo et al., 2015), studies with new potential drugs or compounds are desirable to the development of a preventive and/or curative treatment. Potential natural/chemical compounds have been investigated to control parasite and the immune response in different infected tissues (De Paula Costa et al., 2016; Menezes et al., 2020).
Curcumin is a polyphenol compound of saffron and it has antioxidant, anti-inflammatory, antiangiogenic, anticarcinogenic and apoptosis regulative properties (Arbiser et al., 1998; Aggarwal et al., 2006; Kuttan et al., 2007; Chiu et al., 2009; Kang and Chen, 2009; Nagajyothi et al., 2013). Curcumin regulates different biological and molecular routes, modulating the synthesis of cytokines, chemokines, enzymes, genes, and transcription factors in distinct inflammatory conditions (Gupta et al., 2012; Gupta et al., 2013; Nakagawa et al., 2020). The use of curcumin as a therapeutic compound in the experimental infection of T. cruzi in murine model demonstrated to be effective inhibiting the cellular invasion regulating the LDL receptors in those cells and decreasing the parasitic load in the heart and liver (Yuan et al., 2008; Kang and Chen, 2009; Nagajyothi et al., 2013). Whereas curcumin has a low bioavailability, a new formulation was developed (Theracurmin) whose absorption was 27 higher than curcumin in humans (Nakagawa et al., 2020).
The present study aims to investigate the parasitological and inflammatory patterns in the testicular site of mice infected by the Colombian strain of the T. cruzi under preventively treatment with the new formulation of the Theracurmin.
2 Materials and methods
2.1 Ethical approval
All the methodologies performed in this study were in accordance with the standards of the National Council for Control of Animal Experimentation (CONCEA) and previously approved by the Animal Research Ethics Committee (CEUA) of the Federal University of Ouro Preto (UFOP), Ouro Preto, Minas Gerais, Brazil, under the protocol number 4487110520 (ID 000412).
2.2 Trypanosoma cruzi infection
For these experiments, we used the Colombian strains of T. cruzi, classified as T. cruzi I (Zingales, 2009). These strains were maintained by successive passages in Swiss mice at the Center of Animal Science, UFOP.
2.3 Animals, Trypanosoma cruzi infection and study design
Swiss male mice aged 7-9 weeks weighing approximately 20-40 g were used in this study. Animals (n = 28) were grouped as (i) uninfected (n=7), (ii) uninfected + Theracurmin (n=7), (iii) T. cruzi (n=7), and (iv) T. cruzi + Theracurmin (n=7). Animals were infected by an intraperitoneal injection of the Colombian strain of the parasite (1000 trypomastigotes/animal). Blood parasites were daily evaluated in infected mice according to Brener’s method (Brener, 1962). On day 30 of infection, the animals were euthanized, their testicles were removed and weighed. The left testes were collected for the immune assay and the right were process to histopathology analysis. The mice were housed and maintained at the Center of Animal Science at UFOP at climatized room with controlled luminosity conditions and temperature (22 ± 2°C).
2.4 Theracurmin treatment
One day before the infection, animals were submitted with a daily therapy, via gavage, with 30 mg/kg (Sasaki et al., 2011) of Theracurmin (CurcuminRich®, Natural Factors, Canada) during 30 days. Each capsule of the product had 30 mg of Theracurmin® (Theravalues, Tokyo, Japan). The individual dose preparation consisted in Theracurmin’s dilution in distilled water with 0.5% of carboxymethyl cellulose. Each animal received 300 μL of this mix and the non-treated groups received the solution vehicle at the same amount.
2.5 Testicle processing and histological analysis
Testicular samples of all animals were fixed in tamponed formalin for 24 h, dehydrated in ethanol, embedded in glycol methacrylate resin, and cut into 3 μm thick sections using glass knives (Leica Biosystems, Wetzlar, Germany). Histological slides collected in a semi-series were obtained using one out of every 40 sections to avoid evaluating the same histological area. The sections were stained with toluidine blue.
Digital images obtain with the photomicroscope of light field (Leica DM5000B, Germany) equipped with a digital camera (Leica MC170HD, Germany) were used for morphometric analysis. All images were analyzed using the Image J® software (National Institute of Health, USA).
2.6 Parameters of the tubules seminiferous and of the intertubular area
The volumetric proportions (%) of the tubular and intertubular area were estimated with the count of 266 dots in 10 aleatory fields, totaling 2660 dots for each animal in histological images captured with the 10x objective (Dias et al., 2019). The volume (mL) of each testicular component were estimated considering the percentage obtained multiplied by the testicular parenchyma volume. The volume of each component was estimated from the knowledge of its percentage within the testis and the knowledge of the testicular parenchyma volume. Since the mammalian testis density is around 1 (Tae et al., 2005), its weight was considered as the same as the volume. The volumetric proportions (%) of the intertubular components were estimated with the count of 1000 dots projected in captures images of the intertubular area with the 40x objective in different histological slides of each animal (Mouro et al., 2018). The quantified elements were connective tissue, macrophages, blood vessels, lymphatic space and Leydig cells (nucleus and cytoplasm). To calculate the relation between nucleus and cytoplasm of the Leydig cells, the percentage occupied by nucleus was divided by the percentage occupied by cytoplasm (Mouro et al., 2018). The volume (mL) of each intertubular component by the testis was calculated using the following formula: proportion of the element on the testis/(100 × parenchymal mass of one testis) (Russell et al., 1993).
2.7 Leydig cell parameters
The diameters of 30 Leydig cells nuclei were measured in each animal, choosing those with circular outline, perinuclear chromatin, and evident nucleoli. To calculate nuclear volume (NV), cytoplasmic volume (CV) and each Leydig cell volume (LCV, the following formulas were used: NV = 4/3 πR3 (R = nuclear radius); CV = % of cytoplasm x NV/% of nucleus; LCV = NV + CV (Russell et al., 1993).
The calculation of the volume that Leydig cells occupy in the testis was performed from the proportion of Leydig cells in the testicular parenchyma x parenchyma weight of one testicle/100. The volume that the Leydig cells occupy per gram of testis was obtained using the gross weight of the testicles. The total number of Leydig cells in the testes was calculated from the volume that the Leydig cells occupy in the testes (μm3)/volume of one Leydig cell (μm3). To calculate the total number of Leydig cells per gram of testis the following formula was used: volume that the Leydig cell occupies per gram of testis (μm3)/volume of one Leydig cell (μm3). The Leydigosomatic Index (LSI) was calculated by the formula: LSI = total volume of Leydig cell in the testicular parenchyma/BW x 100 (BW = body weight).
2.8 Immunoassay
Levels of CCL2, CXCL10, IL-6 and IL-15 were detected in the supernatant of the homogenized testicular tissues. For sample preparation, 30 mg of testicular tissues were macerated in 300 mL of phosphate buffered saline (PBS) and, after centrifugation at 13000 rpm, for 10 min at 4°C, the supernatant was collected (Souza et al., 2021). Inflammatory mediators were measured by enzyme-linked immunosorbent assay (ELISA) using a specific kit (Peprotech, NJ, USA) and were performed according to the manufacturer’s information. The absorbance values were measured using the Biotek ELx808 (California, USA) ELISA reader at 450 nm.
2.9 Statistical analysis
Data are expressed as mean ± standard error of means. Multiple groups were compared using one-way analysis of variance (ANOVA) followed by the Tukey-Kramer post-test. Non paired t test was used to evaluate two independent samples for parametric data. All analyses were performed using the Prism 8 software (GraphPad Software). Groups were considered statistically different at p < 0.05.
3 Results
Animals infected with T. cruzi and preventively treated with Theracurmin showed a significant reduction in the parasitic load of the circulating blood at the final of the treatment. Figure 1 represents the area under the curve indicating a significative difference between the circulating T. cruzi in both infected groups.
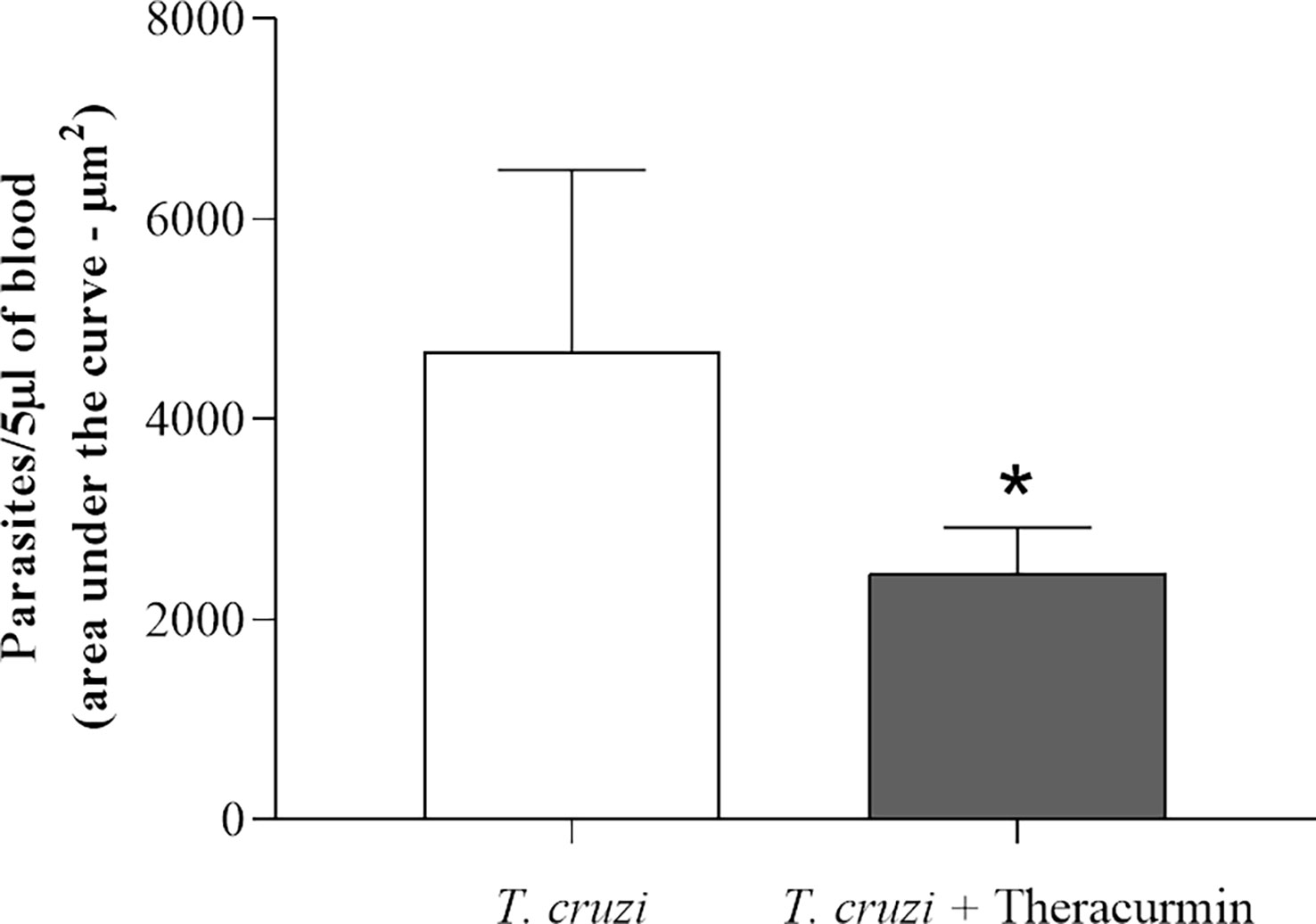
Figure 1 Area under the curve of the parasitemia of the T. cruzi infected group (n=7) and the T. cruzi + Theracurmin group (n=7). Data are expressed as mean ± SE. * shows significant difference (p = 0,0086).
Biometric data obtained from uninfected and T. cruzi-infected animals under treatment with Theracurmin is shown in Table 1. There was no difference at the body weight of the mice, such as at the albuginea’s weight. In contrast, the testicular weight and gonadosomatic index increased in uninfected animals under Theracurmin treatment when compared to those infected animals that received Theracurmin.
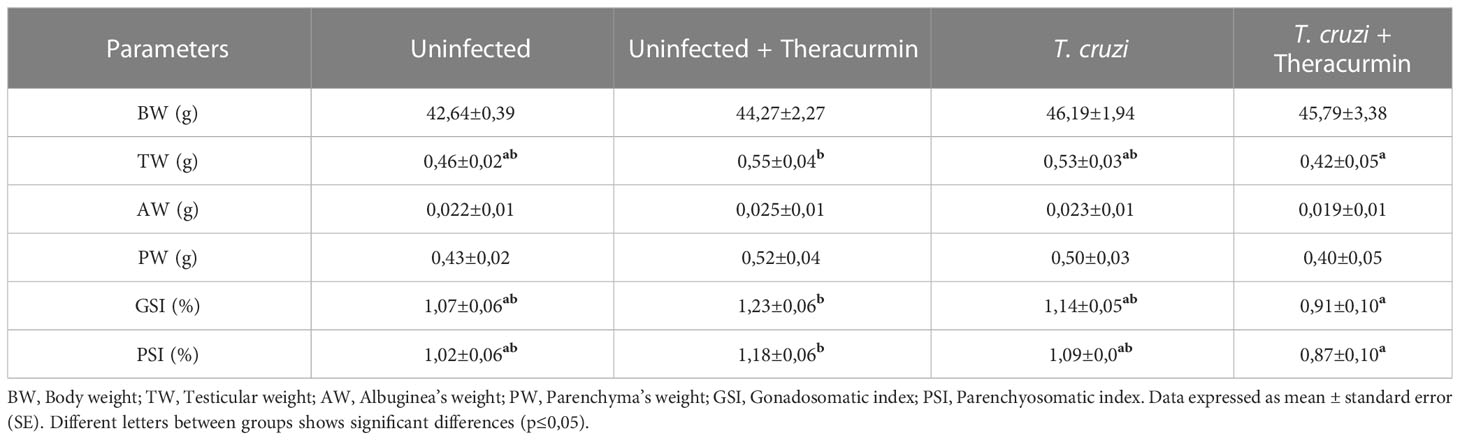
Table 1 Biometric and testicular parameters of mice infected with Trypanosoma cruzi and treated with Theracurmin.
In the histological analysis, there was not observed the presence of the amastigote nests in testicles, in the tubular, neither in the intertubular areas (Figure 2). In the morphometric analysis, there was an increase in the percentage of the tubules seminiferous and of the tunica propria in T. cruzi-infected animals when compared to those without infection, independent of the treatment with Theracurmin. However, in the infected groups under Theracurmin treatment, we observed a decrease in the percentage of the tunica propria when compared to the animals without the treatment. In addition, the tubulosomatic index raised in the uninfected animals under treatment and the Theracurmin was able to decrease the epitheliumsomatic index in the animals infected by T. cruzi (Table 2).
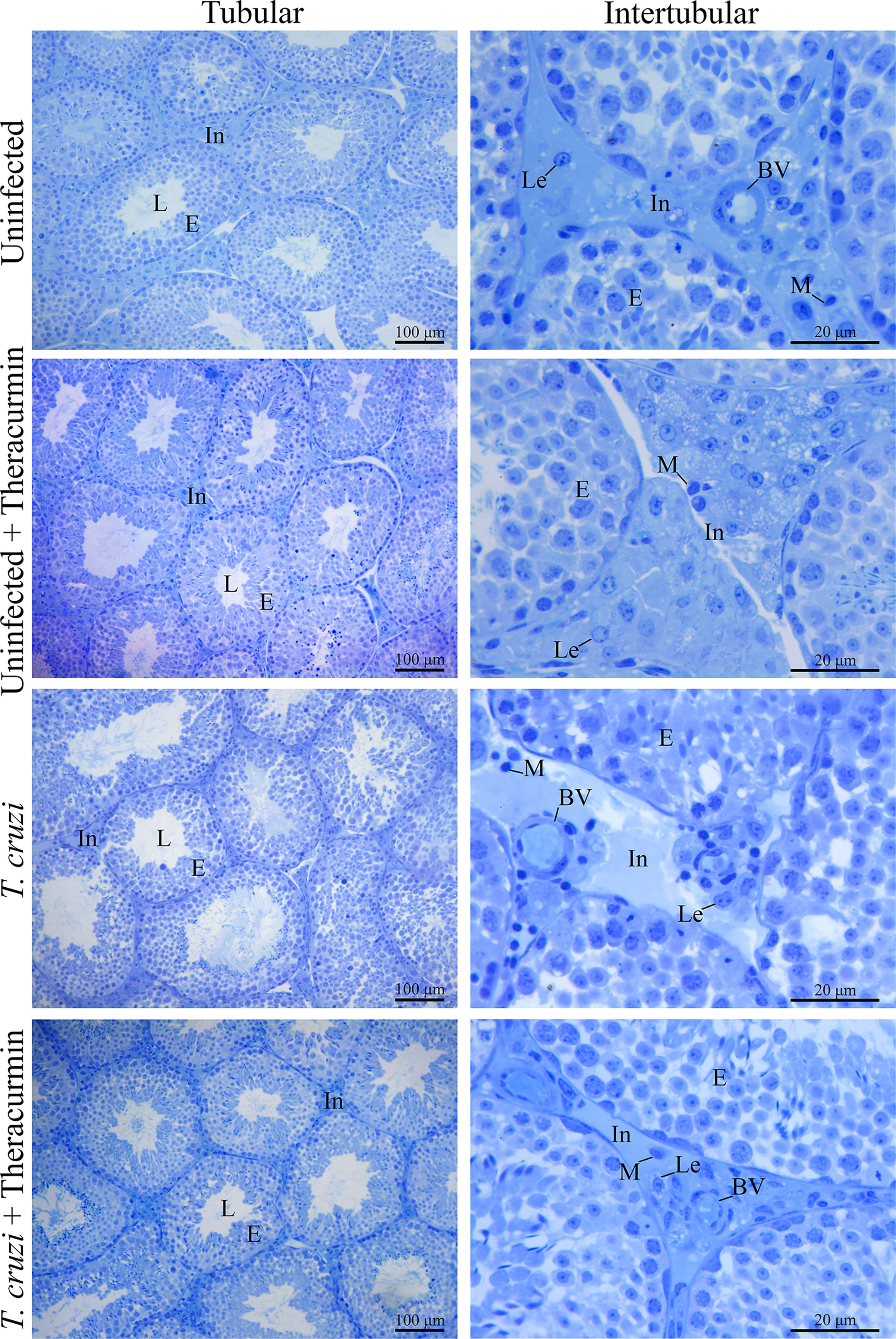
Figure 2 Histological images of testicles of Swiss mice infected with T. cruzi and treated with Theracurmin. Animals (n = 28) were grouped as (i) uninfected (n=7), (ii) uninfected + Theracurmin (n=7), (iii) T. cruzi (n=7), and (iv) T. cruzi + Theracurmin (n=7). At the second day after the beginning of the treatment, mice were intraperitoneally infected with 1000 trypomastigote forms of Colombian strain of T. cruzi. After 30 dpi testicles were fixed, embedded in glycol methacrylate resin, and cut into 3 mm thick sections. The sections were stained with toluidine blue. The volumetric proportions of the tubular and intertubular area were estimated with the count of 266 dots in 10 aleatory fields and, the volumetric proportions of the intertubular components were estimated with the count of 1000 dots projected in captured images of the intertubular area in different histological slides of each animal. In, Intertubular area; L, Lumen; E, Epithelium; Lc, Leydig cell; BV, Blood vessel; M, Macrophage.
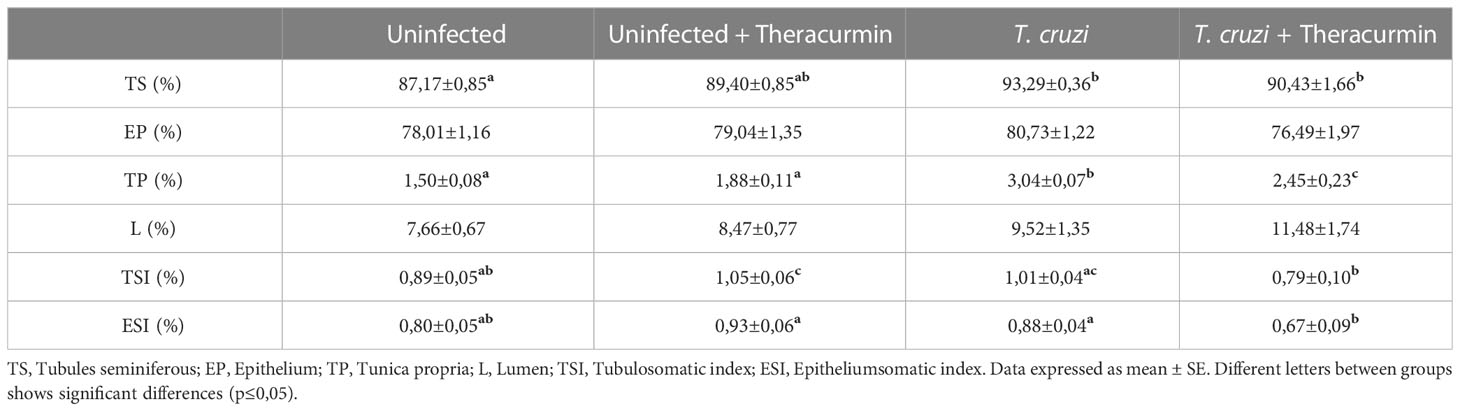
Table 2 Parameters of the tubules seminiferous of Swiss mice infected with T. cruzi and treated with Theracurmin.
The stereological analysis of the tubules seminiferous demonstrated an increase at the luminal diameter and luminal area in the animals infected by T. cruzi. Therefore, the tubules/epithelium ratio was higher in infected mice (Table 3).
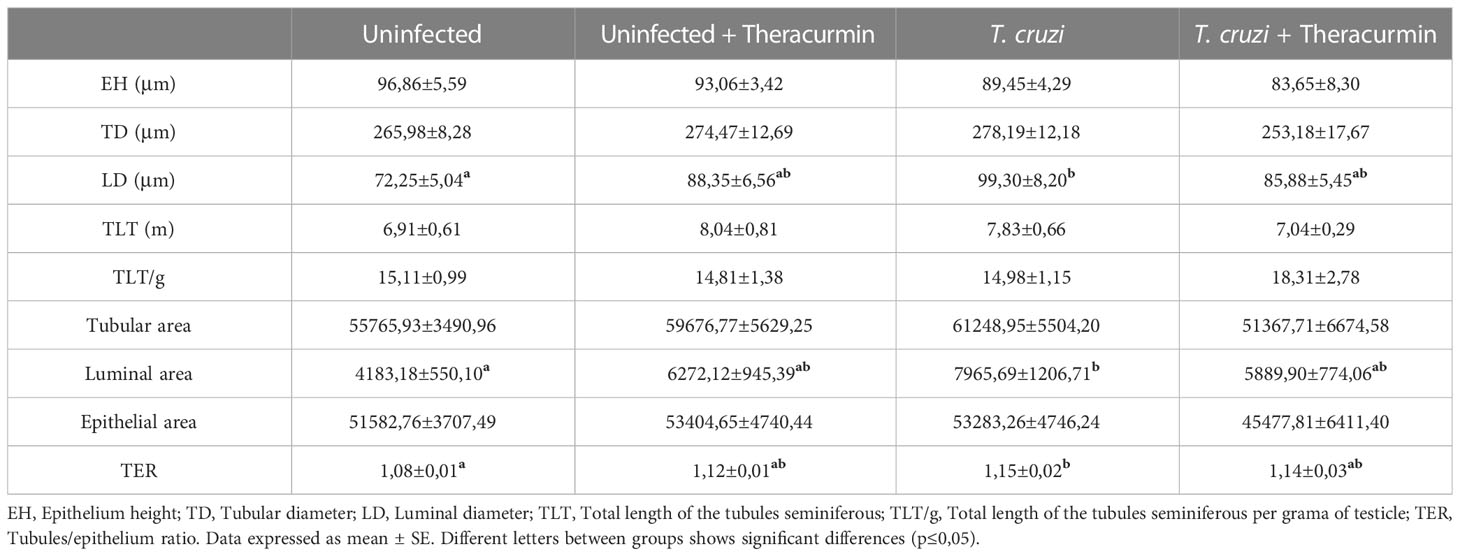
Table 3 Stereological analysis of the tubules seminiferous of Swiss mice infected with T. cruzi and treated with Theracurmin.
At the intertubular area, its percentual reduced significantly in the presence of T. cruzi and T. cruzi + Theracurmin when compared to the non-infected animals, while the Leydig nucleus percentage reduced only at the infected animals in comparison those uninfected. The Leydig cytoplasm and de Leydig cell percentages were decreased in the presence of T. cruzi, while the percentage of connective tissue was decreased in the presence of T. cruzi plus treatment with Theracurmin. The macrophages percentage were also smaller in the presence of T. cruzi when compared to the uninfected and untreated group (Table 4).
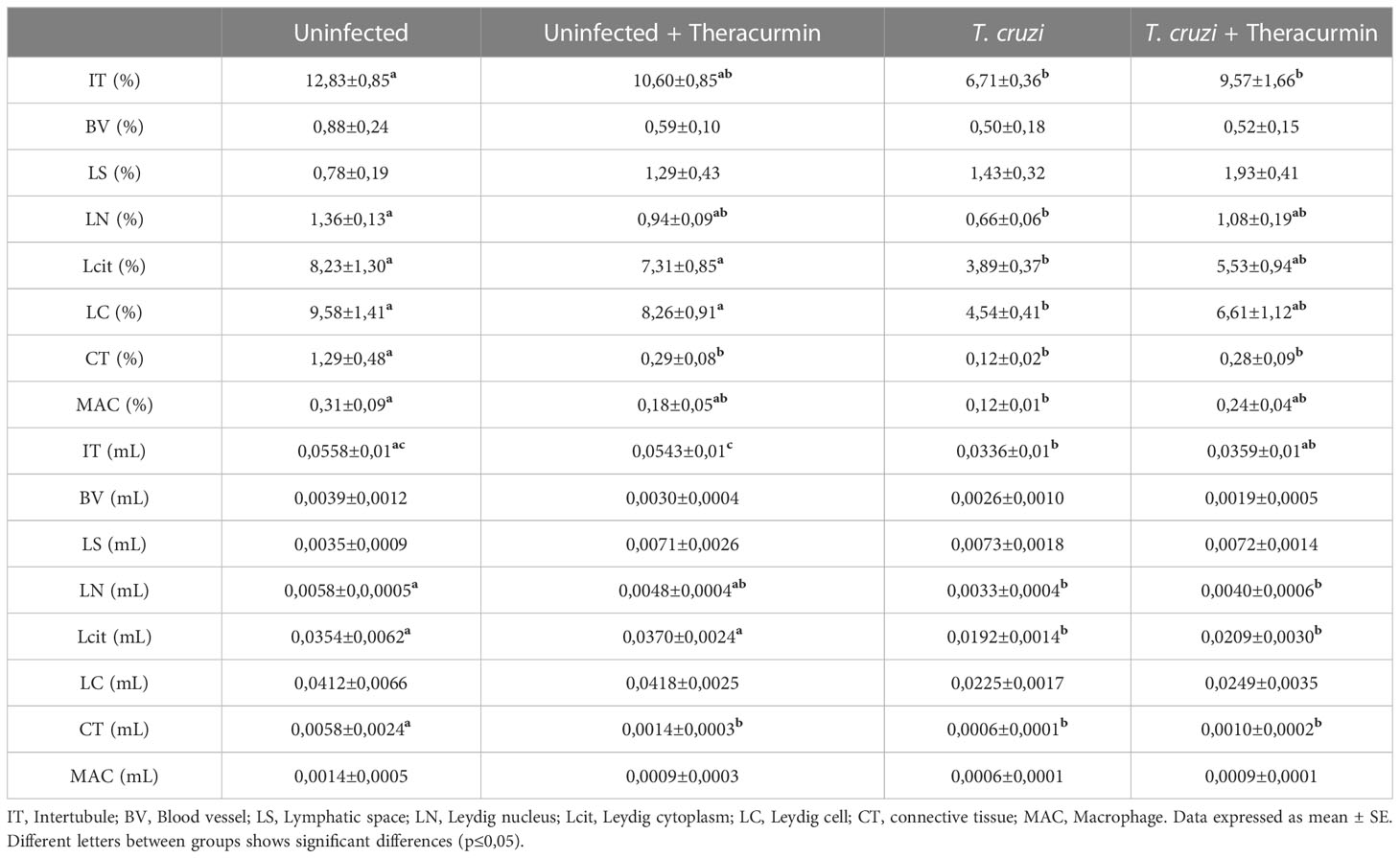
Table 4 Percentage and volume of the intertubular components in mice infected with T. cruzi and treated with Theracurmin.
The intertubular volume (mL) reduced in those animals infected by T. cruzi. These animals also had a smaller volume of Leydig nucleus and cytoplasm when compared to the non-infected animals. According to its percentage, the connective tissue volume was decreased in the presence of the parasite plus Theracurmin treatment (Table 4).
According to the morphometric and stereological parameters of the Leydig cells, its cytoplasmatic and cellular volumes were reduced in the presence of T. cruzi + Theracurmin when compared to the uninfected animals + Theracurmin. Furthermore, the Leydig cellular volume per testicle was reduced in the animals infected by the protozoan, while the Leydig cellular volume per gram of testicle were smaller only at the T. cruzi group, when compared to the non-infected animals. The number of Leydig cells per testicle also reduced in the infected animals when compared to the uninfected one. Even though the number of Leydig cells per testicle did not show difference between the infected groups, the Theracurmin treatment inhibit the decrease of the number of Leydig cells per gram of testicle. The leydigosomatic index were also reduced in the infected animals, when compared to those uninfected (Table 5).
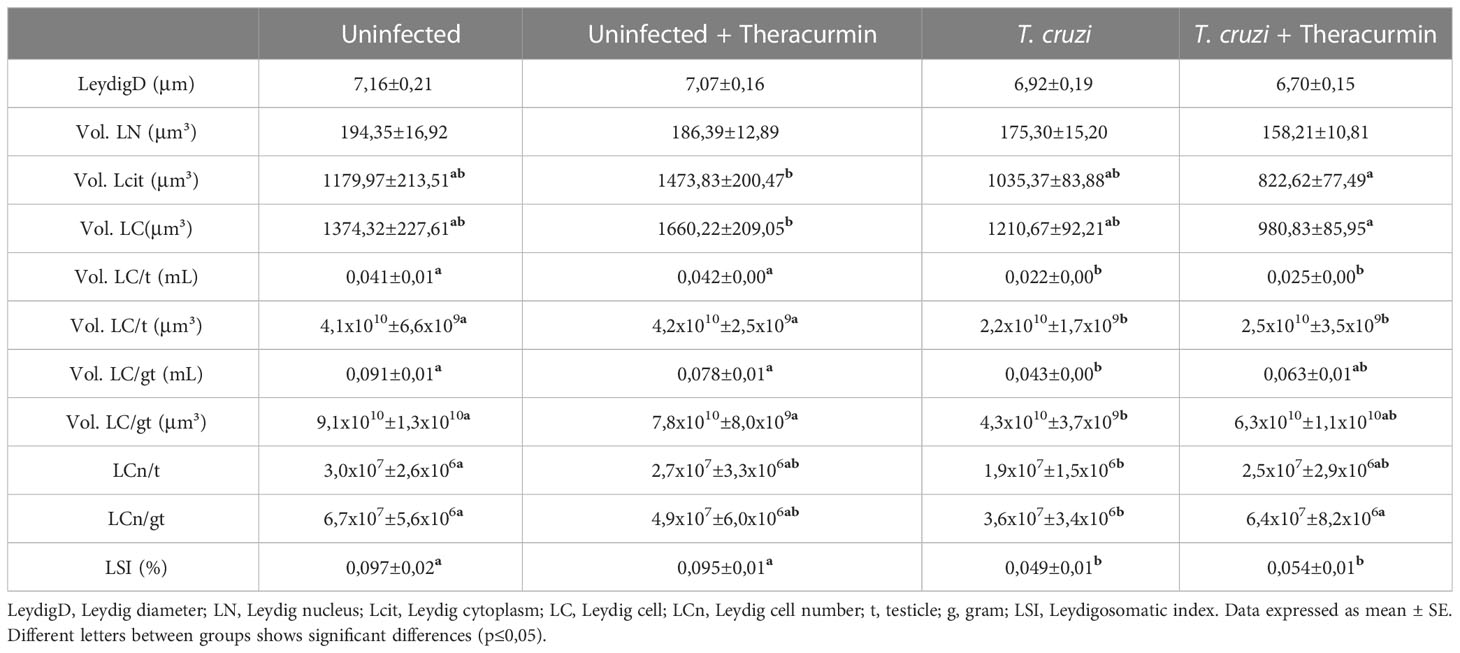
Table 5 Morphometric and stereological parameters of Leydig cells in mice infected with T. cruzi and treated with Theracurmin.
Finally, the concentration of CCL2, CXCL10 did not suffer any alterations in the animals infected with T. cruzi and/or treated with Theracurmin with in the testicular site (Figures 3A, B). Despite of that, the concentration of IL-15 and IL-6 shows that the T. cruzi have increased the production of those inflammatory markers, but the animals treated with Theracurmin were capable of reduce these cytokine levels in testicular area (Figures 3C, D).
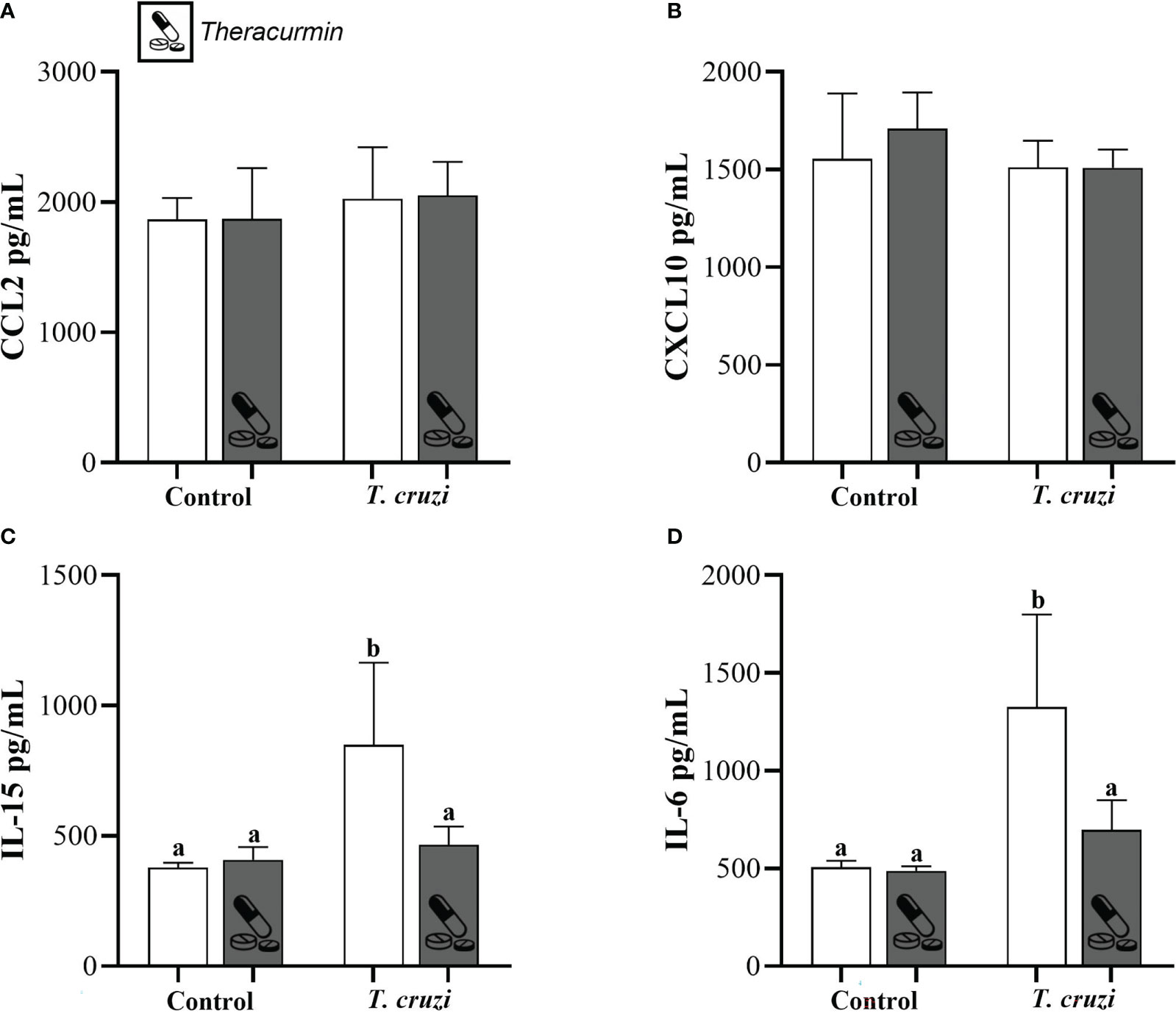
Figure 3 Production of CCL2 (A), CXCL10 (B), IL-15 (C), and IL-6 (D) at testicles of Swiss mice infected with T. cruzi and treated with Theracurmin. Animals (n = 28) were grouped as (i) uninfected (n=7), (ii) uninfected + Theracurmin (n=7), (iii) T. cruzi (n=7), and (iv) T. cruzi + Theracurmin (n=7). At the second day after the beginning of the treatment, mice were intraperitoneally infected with 1000 trypomastigote forms of Colombian strain of T. cruzi. After 30 dpi the concentration of these cytokines was evaluated by the enzymatic immunoassay method at the testicular macerated. Data expressed as mean ± SE. Different letters between groups shows significant differences (p ≤ 0,05).
4 Discussion
This present study aimed to verify if the preventive treatment with Theracurmin (highly bioavailable curcumin) was able to regulate the morphological and/or inflammatory parameters caused by the infection of T. cruzi in experimental model. Previously, curcumin was able to reduce the parasitemia in T. cruzi infected animals (Nagajyothi et al., 2013) which is explained by the inhibitory effect of curcumin at the transcription of low-density lipoprotein receptor (LDLr) (Yuan et al., 2008; Kang and Chen, 2009; Nagajyothi et al., 2013) as it is known that the T. cruzi utilizes these LDLr during its cellular invasion process (Nagajyothi et al., 2011).
The Colombian strain of T. cruzi has a positive tropism for muscular tissues, infecting myoid cells frequently (Melo and Brener, 1978; Camandaroba et al., 2001; Andrade et al, 2010). Research conducted by Carvalho et al. (2009) showed that the protozoan can infect the tunica propria cells of the tubule seminiferous and may have that preference because of contractible proteins responsible for the movement of non-motile sperm cells throughout the tubules (Lenzi et al., 1998). At this present study, we found a higher volumetric percentage and volume of the tunica propria cells in the presence of the T. cruzi, even though we did not find amastigotes of the parasite in the testicular site. Theracurmin may have a protective role in muscular-like cells and tissue (Hernández et al., 2021), resulting in a reduced cellular hyperplasia at the testicular tunica propria in treated mices. The infection by T. cruzi also induced a significant raise in the luminal diameter and area of the tubules seminiferous resulting in a higher tubule-epithelial ratio. The tubules seminiferous are composed of three main components: tunica propria, epithelium and lumen, this last structure is where the spermatozoa are released after spermatogenesis (Ross, 1974). This difference at the lumen may be the result of a compensatory effect due to the inflammatory stress caused by T. cruzi infection.
At this present study we found significant decrease in components of the intertubular area, such as the percentage of resident macrophages and the percentage and volume of the connective tissue during T. cruzi infection, corroborating the data in Mendis-Handagama et al. (1990), suggesting an alteration of the intertubular site caused by the inflammation at testicular site. The treatment with Theracurmin also led to a decrease in the percentage and volume of connective tissue probably because of the inhibition generated by the treatment in the genic expression of growth factors at the connective tissue causing a reduction at the extracellular matrix including type I collagen and fibronectin (Xu et al., 2003; Chen and Zheng, 2008).The Leydig cells and resident macrophages of the intertubular area of the testicles has an intimate relation as these macrophages are responsible for the release of biomarkers that affect directly at the diametric growth and development of the testosterone producers’ cells (Hales, 2007). In this context, the reduced presence of resident macrophages, as observed in the present study, may result in a subdevelopment of the Leydig cells. The T. cruzi is a crucial element to potentialize the local inflammatory response with its immunogenic molecules and, even in environment without inflammation, macrophages release important factors that contribute to the development of Leydig cells (Hales et al., 1999; Hales, 2002). The decreased in Leydig cell population may be due to an intense inflammation generated by the T. cruzi, reducing progressively the genic expression and function of these cells, as showed by Aldahhan et al. (2021) in rats with cryptorchids.
The infection caused by the protozoan T. cruzi is well known to cause an intense inflammatory response in a diverse range of tissues in the mammal host body with the release of inflammatory markers, such as cytokines and chemokines (Talvani and Teixeira, 2011). The curcumin has an anti-inflammatory and protective effects in tissues such as the heart and liver during the stress associated with chronic diseases, preserving their function, and regulating the release of inflammatory markers (Nagajyothi et al., 2013; Hernández et al., 2021). At the macerated of the testicles, Theracurmin treatment was able to reduce the concentration of the proinflammatory cytokines IL-6 and IL-15. Produced by the Sertoli cells, IL-6 has an important role at paracrine/autocrine regulation during spermatogenesis and steroidogenesis and presenting in higher concentrations during exacerbated inflammation (Hedger and Meinhardt, 2003; Rival et al., 2006). IL-15 has also presented higher concentration during microorganisms’ infection and may have an important role in innate and adaptative response at testicular site (Anastasiadou and Michailidis, 2016). Even though the concentrations of CCL2 and CXCL10 are normally higher in heart and blood during T. cruzi infection (De Araújo et al., 2020), our data did not show difference at testicular site. This fact reinforce that the inflammatory response conducted by the T. cruzi is compartmentalized and, in parts, defined by the genetic aspects of the parasite and its studied host.
In conclusion, our study demonstrated that the Colombian strain of the T. cruzi may cause immunological and punctual structural alterations at the testicular site in mice infected, mainly at the tunica propria, muscle layer that recovers the tubules seminiferous, and at the Leydig cells. Moreover, the preventive treatment with Theracurmin presented protective effects, regulating the synthesis of IL-6 and IL-15, but not CCL2 and CXCL10, in the testicular area. The relation between the T. cruzi infection and the testosterone production at the Leydig cells still needs to be researched for a bigger understanding of the damage caused by the protozoan at the testicular site and the relation with the spermatogenesis.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Ethics statement
It was approved by the Animal Research Ethics Committee (CEUA) of the Federal University of Ouro Preto (UFOP), Ouro Preto, Minas Gerais, Brazil, under the protocol number 4487110520.
Author contributions
The authors’ contributions were as follows: BP, TM, AT: conception, design, writing and final content; VS, FD, BM, LR and BP: perform the experiments; KP, TM, BP and AT: data analysis; AT: funding acquisition. All authors contributed to the article and approved the submitted version.
Funding
This research was supported by Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) (Protocol # 405946/2021-0), Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES), Fundação de Amparo à Pesquisa de Minas Gerais- (BPD-00754-22 and BPD-00754-22), Universidade Federal de Ouro Preto (UFOP, Brazil) and Universidade Federal de Minas Gerais (UFMG, Brazil).
Acknowledgments
AT (Process # 305634/2017-8) express their gratitude to the CNPq for the research productivity fellowship. The authors would like to acknowledge the Laboratório Multiusuários de Microscopia Avançada e Microanálise, Núcleo de Pesquisas em Ciências Biológicas, Universidade Federal de Ouro Preto, for providing the equipment and technical support for experiments involving brightfield. The authors also appreciated Prof. Sarah A. Auharek, PhD, (UFVJM, Brazil) scientific reading and valuable and profound comments concerning this manuscript.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Aggarwal, S., Kim, S. W., Cheon, K., Tabassam, F. H., Yoon, J.-H., Koo, J. S. (2006). Nonclassical action of retinoic acid on the activation of the cAMP response element-binding protein in normal human bronchial epithelial cells. Mol. Biol. Cell 17, 566–575. doi: 10.1091/mbc.e05-06-0519
Aldahhan, R. A., Stanton, P. G., Ludlow, H., de Kretser, D. M., Hedger, M. P. (2021). Experimental cryptorchidism causes chronic inflammation and a progressive decline in sertoli cell and leydig cell function in the adult rat testis. Reprod. Sci. 28, 2916–2928. doi: 10.1007/s43032-021-00616-0
Almeida, A. B., Araújo, P. F., Bernal, F. M., Rosa, A., de, C., Valente, S. A., et al. (2019). Sexual transmission of American trypanosomes from males and females to naive mates. J. Vis. Exp. 143, 1–12. doi: 10.3791/57985
Anastasiadou, M., Michailidis, G. (2016). Transcriptional changes of cytokines in rooster testis and epididymis during sexual maturation stages and Salmonella infection. Anim. Reprod. Sci. 171, 41–48. doi: 10.1016/j.anireprosci.2016.05.012
Andrade, L. O., Galvão, L. M. C., de Nazareth SL Meirelles, M., Chiari, E., Pena, S. D. J., Macedo, A. M. (2010). Differential tissue tropism of Trypanosoma cruzi strains: An in vitro study. Mem. Inst. Oswaldo Cruz 105, 834–837. doi: 10.1590/S0074-02762010000600018
Arbiser, J. L., Klauber, N., Rohan, R., Van Leeuwen, R., Huang, M. T., Fisher, C., et al. (1998). Curcumin is an in vivo inhibitor of angiogenesis. Mol. Med. 4, 376–383. doi: 10.1007/bf03401744
Brener, Z. (1962). Therapeutic activity and criterion of cure on mice experimentally infected with Trypanosoma cruzi. rev. Inst. Med. Trop. Sao Paulo 4, 389–396.
Camandaroba, E. L., Campos, R. F., Magalhães, J. B., Andrade, S. G. (2001). Clonal structure of Trypanosoma cruzi Colombian strain (biodeme type III): biological, isoenzymic and histopathological analysis of seven isolated clones. Rev. Soc Bras. Med. Trop. 34, 151–157. doi: 10.1590/S0037-86822001000200001
Carvalho, L. O. P., Abreu-Silva, A. L., Hardoim, D., de, J., Tedesco, R. C., Mendes, V. G., et al. (2009). Trypanosoma cruzi and myoid cells from seminiferous tubules: Interaction and relation with fibrous components of extracellular matrix in experimental chagas’ disease. Int. J. Exp. Pathol. 90, 52–57. doi: 10.1111/j.1365-2613.2008.00592.x
Chen, A., Zheng, S. (2008). Curcumin inhibits connective tissue growth factor gene expression in activated hepatic stellate cells in vitro by blocking NF-κB and ERK signalling. Br. J. Pharmacol. 153, 557–567. doi: 10.1038/sj.bjp.0707542
Chiu, J., Khan, Z. A., Farhangkhoee, H., Chakrabarti, S. (2009). Curcumin prevents diabetes-associated abnormalities in the kidneys by inhibiting p300 and nuclear factor-κB. Nutrition 25, 964–972. doi: 10.1016/j.nut.2008.12.007
Cunha-Neto, E., Chevillard, C. (2014). Chagas disease cardiomyopathy: Immunopathology and genetics. Mediators Inflamm. 1–11. doi: 10.1155/2014/683230
De Araújo, F. F., Lima Torres, K. C., Viana Peixoto, S., Pinho Ribeiro, A. L., Vaz Melo Mambrini, J., Bortolo Rezende, V., et al. (2020). CXCL9 and CXCL10 display an age-dependent profile in chagas patients: A cohort study of aging in bambui, Brazil. Infect. Dis. Poverty 9, 1–10. doi: 10.1186/s40249-020-00663-w
De Paula Costa, G., Lopes, L. R., Da Silva, M. C., Horta, A. L., Pontes, W. M., Milanezi, C. M., et al. (2016). Doxycycline and benznidazole reduce the profile of Th1, Th2, and Th17 chemokines and chemokine receptors in cardiac tissue from chronic Trypanosoma cruzi-infected dogs. Mediators Inflamm. 2016, 3694714 doi: 10.1155/2016/3694714
Dias, F. C. R., Martins, A. L. P., de Melo, F. C. S. A., do Carmo Cupertino, M., de Lucca Moreira Gomes, M., de Oliveira, J. M., et al. (2019). Hydroalcoholic extract of Pfaffia glomerata alters the organization of the seminiferous tubules by modulating the oxidative state and the microstructural reorganization of the mice testes. J. Ethnopharmacol. 233, 179–189. doi: 10.1016/j.jep.2018.12.047
Esper, L., Talvani, A., Pimentel, P., Teixeira, M. M., Machado, F. S. (2015). Molecular mechanisms of myocarditis caused by Trypanosoma cruzi. Curr. Opin. Infect. Dis. 28, 246–252. doi: 10.1097/QCO.0000000000000157
Gupta, S. C., Patchva, S., Aggarwal, B. B. (2013). Therapeutic roles of curcumin: Lessons learned from clinical trials. AAPS J. 15, 195–218. doi: 10.1208/s12248-012-9432-8
Gupta, S. C., Patchva, S., Koh, W., Aggarwal, B. B. (2012). Discovery of curcumin, a component of golden spice, and its miraculous biological activities. Clin. Exp. Pharmacol. Physiol. 39, 283–299. doi: 10.1111/j.1440-1681.2011.05648.x
Hales, D. B. (2002). Testicular macrophage modulation of leydig cell steroidogenesis. J. Reprod. Immunol. 57, 3–18. doi: 10.1016/S0165-0378(02)00020-7
Hales, D. B. (2007). “Regulation of leydig cell function as it pertains to the inflammatory response,” in The leydig cell in health and disease. Eds. Payne, A. H., Hardy, M. P. (Totowa, NJ: Humana Press), 305–321.
Hales, D. B., Diemer, T., Held Hales, K. (1999). Role of cytokines in testicular function. Endocrine 10, 201–217. doi: 10.1007/bf02738619
Hedger, M. P., Meinhardt, A. (2003). Cytokines and the immune-testicular axis. J. Reprod. Immunol. 58, 1–26. doi: 10.1016/S0165-0378(02)00060-8
Hernández, M., Wicz, S., Pérez Caballero, E., Santamaría, M. H., Corral, R. S. (2021). Dual chemotherapy with benznidazole at suboptimal dose plus curcumin nanoparticles mitigates Trypanosoma cruzi-elicited chronic cardiomyopathy. Parasitol. Int. 81, 1–9. doi: 10.1016/j.parint.2020.102248
Hossain, E., Khanam, S., Dean, D. A., Wu, C., Lostracco-Johnson, S., Thomas, D., et al. (2020). Mapping of host-parasite-microbiome interactions reveals metabolic determinants of tropism and tolerance in chagas disease. Sci. Adv. 6, 1–14. doi: 10.1126/sciadv.aaz2015
Kang, Q., Chen, A. (2009). Curcumin inhibits srebp-2 expression in activated hepatic stellate cells in vitro by reducing the activity of specificity protein-1. Endocrinology 150, 5384–5394. doi: 10.1210/en.2009-0517
Kuttan, G., Hari Kumar, K. B., Guruvayoorappan, C., Kuttan, R. (2007). “Antitumor, anti-invasion, and antimetastatic effects of curcumin. Adv. Exp. Med. Biol. 595, 173–184. doi: 10.1007/978-0-387-46401-5_6
Lenzi, H. L., Castelo-Branco, M. T. L., Pelajo-Machado, M., Oliveira, D. N., Gattass, C. R. (1998). Trypanosoma cruzi: compromise of reproductive system in acute murine infection. Acta Trop. 71, 117–129. doi: 10.1016/S0001-706X(98)00058-8
Martin, D. L., Lowe, K. R., McNeill, T., Thiele, E. A., Roellig, D. M., Zajdowicz, J., et al. (2015). Potential sexual transmission of Trypanosoma cruzi in mice. Acta Trop. 149, 15–18. doi: 10.1016/j.actatropica.2015.05.002
Medina, L., Castillo, C., Liempi, A., Herbach, M., Cabrera, G., Valenzuela, L., et al. (2018). Kemmerling, U Differential infectivity of two trypanosoma cruzi strains in placental cells and tissue. Acta Trop. 186, 35–40. doi: 10.1016/j.actatropica.2018.07.001
Melo, R. C., Brener, Z. (1978). Tissue tropism of different Trypanosoma cruzi strains. J. Parasitol. 64, 475–482. doi: 10.2307/3279787
Mendis-Handagama, C., Kerr, J., Kretser, D. (1990). Experimental cryptorchidism in the adult mouse: Qualitative and quantitative light microscopy morphology. J. Androl. 11, 539–547.
Menezes, A. P., Silva, M. L. da, Pereira, W. L., Costa, G.de P., Horta, A. L., et al. (2020). In vitro tripanocidal effect of 1,8-dioxooctahydroxanthenes (xanthenodiones) and tetraketones and improvement of cardiac parameters in vivo. J. Glob. Antimicrob. Resist. 22, 466–476. doi: 10.1016/j.jgar.2020.04.005
Morillo, C. A., Marin-Neto, J. A., Avezum, A., Sosa-Estani, S., Rassi, A., Jr., Rosas, F., et al. (2015). Randomized trial of benznidazole for chronic chagas' cardiomyopathy. N. Engl. J. Med. 373 (14), 1295–1306. doi: 10.1056/NEJMoa1507574
Mouro, V. G. S., Menezes, T. P., Lima, G. D. A., Domingues, R. R., Souza, A. C. F., Oliveira, J. A., et al. (2018). How bad is aluminum exposure to reproductive parameters in rats? Biol. Trace Elem. Res. 183, 314–324. doi: 10.1007/s12011-017-1139-3
Nagajyothi, F., Tanowitz, H. B., Zhao, D., Weiss, L. M. (2013). Curcumin treatment provides protection against Trypanosoma cruzi infection. Parasitol. Res. 110, 2491–2499. doi: 10.1007/s00436-011-2790-9.Curcumin
Nagajyothi, F., Weiss, L. M., Silver, D. L., Desruisseaux, M. S., Scherer, P. E., Herz, J., et al. (2011). Trypanosoma cruzi utilizes the host low density lipoprotein receptor in invasion. PloS Negl. Trop. Dis. 5. doi: 10.1371/journal.pntd.0000953
Nakagawa, Y., Mukai, S., Yamada, S., Murata, S., Yabumoto, H., Maeda, T., et al. (2020). The efficacy and safety of highly-bioavailable curcumin for treating knee osteoarthritis: A 6-month open-labeled prospective study. Clin. Med. Insights Arthritis Musculoskelet. Disord. 13, 1–8. doi: 10.1177/1179544120948471
Pérez-molina, J. A., Molina, I. (2018). Chagas disease. Lancet 391, 82–94. doi: 10.1016/S0140-6736(17)31612-4
Rival, C., Theas, S., Guazzone, V. A., Lustig, L. (2006). Interleukin-6 and IL-6 receptor cell expression in testis of rats with autoimmune orchitis. J. Reprod. Immunol. 70, 43–58. doi: 10.1016/j.jri.2005.10.006
Ross, M. H. (1974). The organization of the seminiferous epithelium in the mouse testis following ligation of the efferent ductules. A light microscopic study. Anat. Rec. 180, 565–579. doi: 10.1002/ar.1091800403
Russell, L. D., Ettlin, R. A., Hikim, A. P. S., Clegg, E. D. (1993). Histological and histopathological evaluation of the testis. Int. J. Androl. 16, 83. doi: 10.1111/j.1365-2605.1993.tb01156.x
Sasaki, H., Sunagawa, Y., Takahashi, K., Imaizumi, A., Fukuda, H., Hashimoto, T., et al. (2011). Innovative preparation of curcumin for improved oral bioavailability. Biol. Pharm. Bull. 34, 660–665. doi: 10.1248/bpb.34.660
Souza, D. M. S. de, Silva, M. C., Farias, S. E. B., Menezes, A. P. de J., Milanezi, C. M., Lúcio, K.de P., et al. (2021). Diet rich in lard promotes a metabolic environment favorable to Trypanosoma cruzi growth. Front. Cardiovasc. Med. 8. doi: 10.3389/fcvm.2021.667580
Tae, H. J., Jang, B. G., Ahn, D. C., Choi, E. Y., Kang, H. S., Kim, N. S., et al. (2005). Morphometric studies on the testis of Korean ring-necked pheasant (Phasianus colchicus karpowi) during the breeding and non-breeding seasons. Vet. Res. Commun. 29, 629–643. doi: 10.1007/s11259-005-3080-8
Talvani, A., Teixeira, M. M. (2011). Inflammation and chagas disease: Some mechanisms and relevance. Adv. Parasitol. 76, 171–194. doi: 10.1016/B978-0-12-385895-5.00008-6
Weaver, J. D., Hoffman, V. J., Roffe, E., Murphy, P. M., Herbert, D. R. (2019). Low-level parasite persistence drives vasculitis and myositis in skeletal muscle of mice chronically infected with Trypanosoma cruzi. Infect. Immun. 87, 81–119. doi: 10.1128/IAI.00081-19
World Health Organization (2022) Chagas disease (American trypanosomiasis). Available at: https://www.who.int/news-room/fact-sheets/detail/chagas-disease-(american-trypanosomiasis) (Accessed March 15, 2022).
Xu, J., Fu, Y., Chen, A. (2003). Activation of peroxisome proliferator-activated receptor-γ contributes to the inhibitory effects of curcumin on rat hepatic stellate cell growth. Am. J. Physiol. Liver Physiol. 285, 20–30. doi: 10.1152/ajpgi.00474.2002
Yuan, H., Kuang, S., Zheng, X., Ling, H., Yang, Y.-B., Yan, P.-K., et al. (2008). Curcumin inhibits cellular cholesterol accumulation by regulating SREBP-1/caveolin-1 signaling pathway in vascular smooth muscle cells1. Acta Pharmacol. Sin. 29, 555–563. doi: 10.1111/j.1745-7254.2008.00783.x
Zapparoli, D., Bertozzo, T. V., Alexandrino, M., Sanches, D. F., Aires, I. N., Manzini, S., et al. (2022). Commercially acquired açaí pulps contamination by Trypanosoma cruzi. Int. J. Food Microbiol. 363, 86. doi: 10.1016/j.ijfoodmicro.2021.109508
Zingales, B. (2009). A new consensus for Trypanosoma cruzi intraspecific nomenclature: Second revision meeting recommends TcI to TcVI. Mem. Inst. Oswaldo Cruz 104, 1051–1054. doi: 10.1590/S0074-02762009000700021
Keywords: Trypanosoma cruzi, inflammation, testicles, cytokines, theracurmin, Colombian strain
Citation: Pimenta BL, Menezes TP, Louise V, Dias FCR, Machado BAA, Ribeiro L, Pinto KMdC and Talvani A (2023) Protective effects of Theracurmin treatment during experimental infection of the Colombian strain of Trypanosoma cruzi at the testicular site. Front. Cell. Infect. Microbiol. 13:1143360. doi: 10.3389/fcimb.2023.1143360
Received: 12 January 2023; Accepted: 13 March 2023;
Published: 24 March 2023.
Edited by:
Juliana Perrone Bezerra De Menezes, Gonçalo Moniz Institute (IGM), BrazilReviewed by:
Jonilson Berlink Lima, Federal University of Western Bahia, BrazilSimone Macambira, Federal University of Bahia, Brazil
Núria Gironès, Centre for Molecular Biology Severo Ochoa, Spanish National Research Council (CSIC), Spain
Luciana Ribeiro Garzoni, Laboratory for Innovation in Therapies, Education and Bioproducts, Oswaldo Cruz Foundation (FIOCRUZ), Brazil
Copyright © 2023 Pimenta, Menezes, Louise, Dias, Machado, Ribeiro, Pinto and Talvani. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Andre Talvani, dGFsdmFuaUB1Zm9wLmVkdS5icg==; Tatiana Prata Menezes, dGF0aWFuYS5wcmF0YUB5YWhvby5jb20uYnI=