 José-Luis Mendívil-Giró
José-Luis Mendívil-Giró- Department of General and Hispanic Linguistics, University of Zaragoza, Zaragoza, Spain
The issue of whether languages adapt to their environment depends on our understanding of language, adaptation, and environment. I consider these three concepts from an internalist or biolinguistic point of view. If adaptation is defined as the result of the differential transmission of phenotypic traits by means of natural selection, then both natural species and languages are adapted. Recall that according to Darwin's own insight, the evolutionary mechanisms for species and languages are “curiously the same” (or “curiously parallel”). However, if the concept of adaptation entails that the environment is the essential source of the structure of evolving objects, then neither natural species nor languages can be said to be adapted to their environment. In the case of languages, I will argue that much of their structure is insensitive to historical change and, therefore, incapable of adaptation to the external environment. The immediate environment of languages is in fact internal to the mind/brain and is thus less variable than the social and physical environment in which people live. On the other hand, the dimensions of languages that are variable have such an indirect relation with the physical and social environment that the notion of adaptation to extra-linguistic reality can only be applied weakly, and then it is unable to explain the main patterns of linguistic structural diversity.
Introduction: Reasons for Skepticism
My aim here is to consider proposals that seek to explain the structure of languages in terms of adaptation to their physical and cultural environment, and to do so with a degree of skepticism. Ladd et al. characterize these proposals as “attempts to relate facts about language structure to facts about speakers and their environment—variables such as group size, geographical location, genetic makeup, and cultural expectations” (Ladd et al., 2015, p. 227). This is not, of course, to deny the inherent interest or value of such work (see current syntheses in Ladd et al., 2015; Lupyan and Dale, 2016). Actually, my critical position toward the claim that there is an influence of extralinguistic factors in the structure of languages is based on a restrictive conception of what is the structure of languages. Then, I neither reject nor question the works that detect (more or less robust) correlations between certain external factors and certain aspects of languages, but I argue that if we understand the structure of languages as it is done in the context of current syntactic theory (especially in the generativist domain), then the claim that the structure of languages can be explained as the result of an adaptation to environmental factors (social, physical, or otherwise) is misleading and inadequately simplifying. This is so because in the aforementioned tradition, the notion of “the structure of languages” transcends relatively superficial aspects (such as the morphological manifestation of certain grammatical categories or the variation in word order) and focuses on (essentially syntactic) formal structural aspects that underlie all languages and that, ultimately, define what is a possible human language.
Therefore, my skepticism arises from two principal claims: (i) the influence of the physical and cultural environment in which languages are developed has a limited scope for explaining the structure of languages, including their main patterns of typological variation, and (ii) such studies do not lead to a satisfying account of what a human language is, from a cognitive and biological perspective, but rather, they take us back to a traditional (and incomplete) view of language as a purely cultural phenomenon.
Comparing Languages and Species
Following August Schleicher, the first major linguist to address the analogy between languages and species suggested by Darwin, I will assume that “not a word of Darwin's need be changed here if we wish to apply this reasoning to languages” [Schleicher, 1863, p. 64 (I quote from the English translation included in Koerner, 1983)]. The reason for my assumption is that in both cases the evolving objects are historically modified natural objects. This identification allows us to say that the process of linguistic change and that of natural evolution are formally alike, although substantially different (for a review of different interpretations of the analogy between languages and species, see Mendívil-Giró, 2006, 2014).
Although various proposals for establishing the specific terms of the comparison have been suggested (e.g., Croft, 2000), the most appropriate one for my purpose is that formulated by Schleicher himself, in his review of the German edition of the Origin of Species:
“The species of a genus are what we call the languages of a family, the races of a species are with us the dialects of a language; the sub-dialects or patois correspond with the varieties of the species, and that which is characteristic of a person's mode of speaking corresponds with the individual” (Schleicher, 1863, p. 32).
What Schleicher calls “that which is characteristic of a person's mode of speaking” is the closest concept to the Chomskyan notion of I-language that could be formulated at that time. Chomsky's (1985) distinction between I-language and E-language was formulated to make clear that the object of study of linguistics as part of cognitive science is not an external object, a shared code or a social institution, but a property of a speaker's mind/brain. Adopting this point of view, I argue that in the comparison between linguistic change and natural evolution the appropriate terms for comparison are as follows: the equivalent of the natural organism (the individual) is the I-language, while the equivalent of the species is a set of similar I-languages (what is usually called a language). Thus, in this context, a language such as Spanish is simply the set of I-languages of Spanish-speaking people (i.e., of the people we identify as users of this way of speaking that we call Spanish), just as the natural species of tigers is nothing other than the set of organisms that we identify as tigers. In both cases the criterion of delimitation, based on similarity, is diffuse and somewhat arbitrary: the criterion of fertile breeding in natural species (Mayr, 1942), and the criterion of mutual intelligibility in languages (Dixon, 1997).
Central to this comparison is that both natural species and natural languages are groups of similar individuals. A natural species is made up of “sufficiently similar” individuals. An orang-utan and a human being have more in common than an orang-utan and a cow, but all three belong to different species. We know that the greater similarity between an orang-utan and a human is due to the fact that their common ancestor is far more recent (about 6 million years) than in the case of humans and cows, which goes back hundreds of millions of years. A “linguistic species” (i.e., a language in the normal use of the term) consists of “sufficiently similar” individuals (I-languages). Thus, the linguistic equivalent of the natural organism (e.g., a tiger) is each person's language organ (the I-language). The linguistic equivalent of the natural species (e.g., Panthera tigris) is the grouping of such language organs. And likewise Spanish and French are more alike than French and Russian, but all three are different languages. We know that the greater similarity between Spanish and French is due to the fact that their common ancestor is much more recent (about 1,500 years) than the ancestor they share with Russian (about 6,000 years).
If an I-language is a person's language organ (his/her faculty of language), there are not around 6,000 languages in the world, but billions, as many as there are people (in fact many more, given that bilingual people have more than one I-language). The only thing that can be said to exist, from an internalist, cognitive, point of view, are those billions of I-languages. All else (varieties, dialects, languages, families, etc.) are abstract constructs that we make by grouping I-languages according to their resemblances or their historical origins. The same is true in the biological realm: what exist are the emerging states of matter that we call life forms, the organisms (the billions of animals, plants, fungi, etc., living on the planet), whereas varieties, species, families, kingdoms, etc., are abstract constructs that we make on the basis of genetic and morphological similarity and historical origins.
And just as we would not say that tigers are manifestations or realizations of the species of tigers (which would have an independent existence), it is not appropriate to say that I-languages are manifestations or realizations of the Spanish or the Russian language (which would have an independent existence in grammars, in dictionaries or in social communities). The Chomskyan cognitive shift had as a central tenet the assertion that languages are not exclusively external, social objects that humans learn, use and transmit from generation to generation, but are in fact different (historically modified) states of the same language faculty, a specific attribute of human cognition. Similarly, natural organisms are different (historically modified) states of the same biochemical phenomenon: life (see Moreno and Mendívil-Giró, 2014 for a development of these ideas).
Comparable to natural evolution in biological organisms, then, is the process of linguistic change in human languages. The assumption that follows, hence, is that the process of language evolution (as a human faculty) is part of natural evolution, and not part of linguistic change. In other words, the process of linguistic change is one that affects (in historical time) the systems of knowledge we call I-languages, and has no relation to the evolutionary processes that could give rise (in geological time) to the faculty of language. To avoid the “unfortunate ambiguity” (cf. Hurford, 1992, p. 273) that expressions like language evolution have in English, I use the term linguistic change to refer to the process of historical change in languages, and I will reserve the term evolution for biological changes, including the evolutionary emergence of the language faculty (an issue that I will not discuss here). In this sense it is possible to affirm, following Berwick and Chomsky (2016, p. 92), that “languages change, but they do not evolve.” For arguments against the assumption that the process of linguistic change is part of the process of language evolution, see Mendívil-Giró (2016) and Longobardi (2003), who clearly distinguishes between historical adequacy and evolutionary adequacy in language sciences.
The parallelism between natural evolution and linguistic change in fact goes beyond the interesting similarities that Darwin (1871) observed, and persists in the relevant spheres of scholarship. Gould (2002) analyses in detail the controversy between adaptationist, externalist, and functionalist evolutionary theorists (using Gould's, 1996 characterization of neo-Darwinism) and, on the other hand, anti-neo-Darwinist theorists (such as Brian Goodwin, Stuart Kauffman, and Gould himself). In linguistics too there is also a parallel controversy, revolving around functionalist and non-functionalist theorists of language change (see Lass, 1997 for a detailed critical review, and for an argument against functional/adaptive models of linguistic change).
The impetus in the functionalist, adaptive approach to linguistic change is contemporary to the emergence and development of the Prague School of Linguistics (see Cercle Linguistique de Prague, 1929). I refer mainly to the conception of language as a social institution in the service of communication and to the preference for teleological explanations of linguistic change. It is relevant noting that the revival of teleological tendencies in the explanation of language change coincides in time and in orientation with the emergence in the twenties and thirties in the twentieth century of the Modern Synthesis of evolutionary theory. The new synthesis implies an inclination to consider natural selection as the only motive power of natural evolution, which implies the idea that every change must be adaptive. In my view, this trend corresponds to functionalist approaches to linguistic change and to the more recent tendency to consider languages as complex adaptive systems (Kirby, 1999).
Gould (1996) has described the fundamental difference between the neo-Darwinist model and its alternatives making use of the metaphor of the billiard ball against Galton's polyhedron. According to the neo-Darwinist point of view, an organism could be represented as a billiard ball in motion. Each time the cue hits the ball there is a variable movement. There is a free variation that goes in all directions. The cue hitting the ball would be natural selection, and the ball goes where selection drives it. This constitutes, in terms of Gould, an externalist, functionalist, and adaptationist evolutionary theory. By contrast, the anti-neo-Darwinist point of view presents the metaphor differently. The organism would be as a polyhedron resting on one of its facets. Once the cue hits it, the prospects for change are very constrained: it is a polyhedron, which has a certain internal structure that limits variation, so that certain options are more likely than others and some are impossible, however interesting that might be from an adaptive point of view.
Of course, this is not the place to review the long dispute over the meaning and implications of the term adaptation in evolutionary theory, nor to reiterate the debate on the channeling of previous history and the laws of nature “on which natural selection was privileged to work” (Kauffman, 1993, p. 643). However, it is important to note that by adopting a cognitive point of view in the study of languages one cannot ignore the strict restrictions that the human brain and cognition impose on the structural design of languages, independently of those aspects susceptible to historical change (and, therefore, candidates for possible processes of adaptation to the environment).
Gould characterized the controversy in evolutionary theory as follows:
“In what ways does the skewed and partial occupancy of the attainable morphospace of adaptive design record the operation of internal constraints (both negative limitations and positive channels), and not only the simple failure of unlimited number of unconstrained lineages to reach all possible position in the allotted time?” (Gould, 2002, p. 1053).
And both options have an equivalent view in current linguistic theory. The internist and formalist approach (characteristic of generative linguistics) conceives languages as systems of knowledge restricted in their range of variation by the structure of the human faculty of language (i.e., as Galton's polyhedrons). This view correlates with a uniformitarian conception of language diversity and with a restrictive conception of linguistic change. The externalist and functionalist approach (represented by cognitive-functional linguistics) conceives languages as external cultural objects that owe their structure to the adaptation to speakers' cognitive and communicative requirements (i.e., as billiard balls). This view correlates with a less constrained conception of linguistic change and with an emphasis on the diversity of languages (see Mendívil-Giró, 2012 for a review of this controversy).
I will argue that what we know about how, and how much, languages can change in time and in relation to the environment places us in the first scenario: i.e., one in which the human faculty of language strictly channels the aspects and components of languages that can vary in time and space.
But What Changes When Languages Change?
According to Hauser et al. (2002) influential model, the human language faculty could be conceived of as a complex system minimally integrated by three components: a conceptual-intentional (CI) system (related to meaning and interpretation), a sensory-motor (SM) system (related to the perception and production of linguistic signals), and a computational system (Narrow Syntax, responsible for the creation of the syntactic structure that underlies linguistic expressions, and ultimately for the compositionality and productivity of human language).
Following later developments of this model (Chomsky, 2007; Berwick and Chomsky, 2011, 2016), I will assume that the computational system has an asymmetrical relationship with the two “external” components (CI and SM), such that the computational system would be optimized for its interaction with the CI system, while the relationship with the SM system would be ancillary or secondary. See Figure 1.
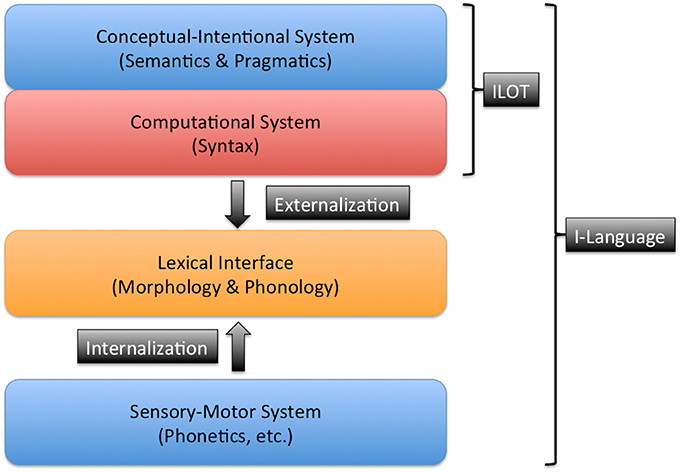
Figure 1. The structure of an I-language. For each component the traditional area of research is indicated. The main components of language are asymmetrically related. The conceptual-intentional (CI) system has a direct connection with the computational system and they form an internal language of thought (ILOT). This ILOT is connected with the sensory-motor (SM) system for language externalization. This connection is established through a lexical interface. The lexical interface changes historically during the process of transmission from generation to generation.
It is then implied that the computational system is coupled with the CI system to form an internal language of thought (ILOT), one that would be essentially homogeneous within the species, and the evolutionary design of which would not be for communication but for thought. Chomsky has suggested that from an evolutionary point of view “the earliest stage of language would have been just that: a language of thought, used internally” (Chomsky, 2007, p. 13).
The connection of the ILOT with the SM system is what would allow the “externalization” of language for interaction and communication with others. Since the connection of the ILOT with the externalization systems is posterior or secondary, it would be precisely within this process that the principal source of the structural diversity among human languages would emerge:
“Parameterization and diversity, then, would be mostly – possibly entirely – restricted to externalization. That is pretty much what we seem to find: a computational system efficiently generating expressions interpretable at the semantic/pragmatic interface, with Diversity resulting from complex and highly varied modes of externalization, which, furthermore, are readily susceptible to historical change” (Berwick and Chomsky, 2011, pp. 37–38).
The connection of the ILOT with the SM system is what allows the externalization of language and, incidentally, what causes the existence of different I-languages. The essential hypothesis is that the same ILOT underlies all languages, so that differences between them are not caused by differences in the CI, the computational, or even the SM systems (which would be biologically conditioned), but follow from differences in how the ILOT is connected to the SM system. Let us suppose, to simplify, that the interface between the ILOT and the sensorimotor system is a kind of “lexicon,” i.e., a repertoire of morpho-phonological formants that allow the externalization of the hierarchical syntactic-semantic representations (produced by the computational system in its interaction with the CI system) in the form of chains of morphemes and phonemes (or, if applicable, visual signs). The role of the lexical interface, then, is to transform abstract hierarchical structures into sequential structures legible at the sensorimotor system. A possible way to understand the format of this lexical interface would be in terms of the type of lexical entries postulated in so-called nanosyntax (Starke, 2009).
Such a model predicts that the diversity in I-languages is the result of variations in externalization, i.e., variations in the configuration of the lexical interface represented in Figure 1. As shown in the diagram, the development of language in an individual implies the learning (the internalization) of the “lexical” material necessary for communication, and it is exactly during this process that reanalyses can occur. A reanalysis is a mismatch in the grammar of two speakers between an internal representation and the linguistic expression produced by the SM system. It can be seen as the equivalent of genetic mutations in organisms.
Let us consider a simplified example: in present-day English the future is expressed as a phrase (I will love) whereas in Spanish it is expressed as a single word (Amaré). According to the model presented, the underlying syntactic structures of the two expressions are very similar (as well as their meaning), while the morphological (and phonological) structures are very different. However, what is now a bound morpheme in the Spanish future (-é) was an auxiliary verb in earlier stages of this language (derived from the vulgar Latin phrase amare habeo “I have to love,” an alternative to the classic Latin synthetic form amabo “I will love”). The transition from a phrase (main verb + auxiliary) to a word (root + affix) at some point in the historical evolution of Romance necessarily implied a process of reanalysis (a mutation). Hence, and again to simplify, we could say that for speaker S1 expression E has the underlying structure Verb+Aux, whereas for speaker S2 the same expression E has the underlying structure Root+Affix, i.e., speaker S2 reanalyses expression E, conferring on it a different underlying structure (Root+Affix) than that of speaker S1 (Verb+Aux). In a sense, then, the I-language of speaker S2 has a mutation, because the relationship between the elements of expression E and its underlying structure is different from that in the I-language of speaker S1. The listener (or the child acquiring a language) does not have immediate access to the syntactic structure or to the semantic representation underlying a given expression, but only to the sound wave that externalizes it. The task of the listeners (or learners) is to use their I-language (including their own lexical interface) to discover this structure by analyzing the sound wave received. In the ideal case, the structure that they get is identical to what the speaker had in mind. When this is not the case, we can say that reanalysis has occurred. So reanalysis is basically a decoding (or acquisition) error, and when this error (this “mutation”) is stabilized in the listener's I-language and is extended to other speakers, we say that there has been a linguistic change. The model predicts that changes happen in the lexical interface that materializes syntactic structures, not in the computational system itself. This view is coherent with the inertial theory of syntactic change (see Longobardi, 2001; Keenan, 2002).
On the other hand, in linguistic change, as in the case with natural evolution, one has to clearly differentiate the reasons why an innovation arises and the reasons why this innovation extends over a population over time. There are many factors that might lead, for example, to the introduction or elimination of a particular acoustic feature in a phonetic segment (from climatic conditions to the presence of speakers of other languages), but a linguistic change will only occur if that mutation extends to other individuals (I-languages), and this itself will only happen if the speakers imitate the speech of the innovators, and the innovations pass these on to subsequent generations. As Labov (1963) showed, the crucial factor in the selection of innovative variants, whether phonetic, morphological, lexical, or syntactic, is not functional efficiency or cost of execution, but social prestige. Some authors (e.g., Croft, 2000) argue that innovations are functional/adaptive, i.e., they have a teleological motivation. But as Lass notes, “unless a motivation is arbitrary, its implementation ought not to subject to contingent factors like age, sex, prestige, etc.” (Lass, 1997, p. 364).
Differences between languages (such as differences between natural species) are the result of change, but linguistic changes only occur in the most superficial dimension of languages, those that are exposed to learning from the environment and are susceptible to historical reanalysis. In the same way, biological evolution significantly alters the form and structure of organisms, but does not modify the biochemistry on which they are built, this remaining unchanged since the emergence of the first forms of life.
The Structural Typology of Languages Does Not Correlate with the Cultural Diversity of Speakers
Even assuming that externalization patterns are the only thing that changes historically in languages, it could still be argued that there is a great deal of room for variation and that, therefore, the structural diversity of languages could reflect processes of adaptation to the environment. Indeed, we know that notable variation in the structure of languages does exist, although the model proposed in Figure 1 would rule out the kind of weakly restricted variation which some authors continue to advocate (see Evans and Levinson, 2009; Mendívil-Giró, 2012 for a critique).
The lack of correlation between different linguistic types and different aspects of human cultures is a strong argument in favor of a restrictive vision of the notion of adaptation applied to human languages, and in favor of a non-exclusively cultural vision of what a language is.
The parameters of linguistic structural variation that have always caught the attention of typologists are those of a morphosyntactic nature (i.e., related to how the morphology of languages reflects the syntactic structure). There are languages with case marking morphemes, and languages without them; there are languages in which verbs are conjugated and agree with several arguments, and languages in which they do not; there are languages in which heads precede complements, and languages in which this happens in reverse; and there are languages in which interrogative words move to the front of sentences, and languages in which they do not (see Dryer and Haspelmath, 2013 for a general survey). Between each of the mentioned options there is a complex range of intermediate steps. For example, among the languages that morphologically mark grammatical relations between verbs and arguments (either with cases or with agreement), some follow the nominative-accusative pattern (formally grouping the subject and differentiating the direct object) and others the ergative-absolutive pattern (formally grouping the subject of the intransitive verb and the object, and differentiating the subject of the transitive verb). Yet there are also languages that are accusative in certain tenses/aspects and ergative in others (see Dixon, 1994). All such variation is compatible with the model set out in Figure 1, and a number of research programs are currently addressing the issues of structural typology based on differences in the externalization component (e.g. Richards, 2016).
What is relevant to us here is that, as Pinker (2007) has pointed out, “the non-universal, learned, variable aspects of language don't fit into any meaningful purposive narrative about the surrounding culture.” The causes of the changes that produce such variation are inherent to linguistic structure itself, and to the mechanism of change (reanalysis). To quote Pinker once more, these changes “aren't part of any symbolic or teleological plan of the culture.” Adapting Pinker's words to our example above, we can say that there are ergative languages and accusative languages, but there are no ergative cultures and accusative cultures. As Baker suggests, “indeed, there is no ecological regularity in how the major linguistic types are distributed around the world” (Baker, 2003, p. 350).
The assumption that there is a correlation between culture or worldview and the grammatical structure of languages is as old as reflections on language typology. In the past it was assumed that the degree of “cultural evolution” determined the degree of “linguistic evolution.” Thus, if we turn again to the case of ergativity, it was claimed that ergativity correlated with a lack of rationality: “What for us is a true cause is for primitive man merely an event involving mystical forces” or “savage man apparently feels that most events are not due to his own volition” (quoted by Seely, 1977, apud Dixon, 1994, p. 214). Dixon argues that by using the same data we could conclude that only speakers of ergative languages have a true notion of agency, since only these speakers formally identify the agentive argument; he concludes that, “in fact, there is no one-to-one correspondence between grammatical marking and mental view of the world” (Dixon, 1994, p. 214).
Even in more recent times, there is no shortage of (more sophisticated and reasonable) proposals about the existence of covariation between culture and grammar, especially relating grammatical complexity with cultural complexity, such as Swadesh (1971), Perkins (1988), or Everett (2005). Swadesh (1971) p. 49) mentions a correlation between inflectional categories and languages' geographical and social extension. But this correlation, if it really exists, does not reveal an adaptation of grammar to culture, but is probably a consequence of morphological simplification, typical of many so-called “world languages” (see section The Brain Internal Environment: Language Learning and Language Processing for discussion). Perkins (1988) proposes a correlation between grammatical complexity and cultural complexity. He surveys in 50 languages several morphological deictic features (tense, person, deictic affixes), syntactic devices related to the coding of reference (determiners, relatives, conjunctions), as well as a measure of cultural complexity (based on the size of settlements, the number of types of craft specialists, and social and political hierarchy depth). Perkins finds a strong correlation that would imply a kind of “linguistic evolution”: languages of complex cultures have few deictic affixes and many syntactic devices. However, Nichols applies her methods to these data and points out that these correlations “may actually reflect only accidentally coincident macroareal linguistic distributions and have no ultimate connection to cultural complexity” (Nichols, 1992, p. 317). (Everett, 2005) proposal on the cultural constraints in Pirahã's grammar is not statistically significant, and the proposed correlation itself has been questioned (see Nevins et al., 2009).
The most reasonable conclusion, therefore, is that there is no correlation between the structural diversity of languages and the cultural diversity of speakers. The fact that one language, for example Mohawk, has more morphological complexity than another, for example English, has no relation to the complexity of the culture in which those languages are spoken, or to the sophistication of its literary tradition, but simply depends on a chain of previous historical facts. The bound morphemes that characterize the complex morphology of many languages are the result of the historical reanalysis of ancient free words. Yet the almost invariable, morphologically simple words that characterize other languages are often the result of the loss of morphological complexity, also resulting from historical reanalysis. In both cases reanalyses, like genetic mutations, are blind and random processes, and Darwin's conclusions can be applied to them: “There seems to be no more design in the variability of organic beings, and in the action of natural selection, than in the course the wind blows” (Darwin, 1893/2000, p. 63).
This conclusion has a solid empirical support. Both Nichols (1992) and Nettle (1999) quantitatively analyse linguistic diversity in time and space and, although with different samples and methodologies, they reach similar conclusions: although there are social and geographical factors that correlate with linguistic diversity and with the density of languages, there is no correlation between typological structural diversity and external factors. As Nettle points out: “Structural diversity […] shows no overall pattern and no correlation with other types of diversity” (Nettle, 1999, p. 137).
Nettle suggests that some extralinguistic factors, such as the size of the speech community, could be related to the preservation of less frequent typological configurations (for example, OS word order, with the object preceding the subject). The argument is based on the assumption that infrequent types are less optimal in functional terms. This assertion is doubtful, because functional optimality is defined in relation to the greater or lesser frequency (I consider the relation between processing and grammar in section The Brain Internal Environment: Language Learning and Language Processing). If we ignore that problem, Nettle's suggestion is interesting. In this case the idea is that, as it happens in population genetics, the effects of random drift are greater when the population is small. But even in this case, it cannot be said that there is a correlation between linguistic types and extralinguistic factors, i.e., it cannot be said that small groups of speakers favor the evolution of certain linguistic types, nor that there is a causal relationship between a small group of speakers and the subject position in the sentence. Note that it could also be argued (what seems more likely) that the possible cause of the maintenance of an infrequent structural type in a given place is the isolation that defines small groups of speakers, isolation that would protect that group from the influence of speakers from other languages (word order is a grammatical feature very prone to diffusion; see Dixon, 1997). What this case shows is that the size of groups of speakers can influence the dynamics of linguistic changes, something perfectly coherent with the model presented here, but that does not allow to affirm that a certain structural feature (the OS order) is an adaptation to a certain type of linguistic context (the size of the community of speakers).
Nichols' (1992) conclusions on the historical evolution of linguistic diversity are also very relevant in this context:
“This survey has uncovered no evidence that human language in general has changed since the earliest stage recoverable by the method used here. There is simply diversity, distributed geographically. The only thing that has demonstrably changed since the first stage of humanity is the geographical distribution of diversity” (Nichols, 1992, p. 277).
If the generation of the structural diversity of languages were the result of adaptive processes to non-linguistic aspects (and not a continuous drift within a restricted design space) we should expect some kind of progression in the historical change of languages, such as we observe in other cultural institutions (politics, art, science, or technology), but this is not the case.
Although structural types of languages do not correlate with the types of societies and cultures that populate our planet, it is still possible to see how certain formal aspects of languages can be explained as processes of adaptation to the environment within the process of linguistic change. However, prior to this we need to determine what is understood by environment and what aspects of a language are sensitive to it.
What is The Environment to Which the Variable Parts of Languages Would Adapt?
So far I have assumed a generic notion of environment, as formulated in the leit Motiv of the Research Topic in which this contribution is included (“to explore the possibility that some aspects of the structure of languages may result from an adaptation to the natural and/or human-made environment”). I have shown that the claim that there is covariation between morphosyntactic typology and aspects of the environment (so defined) is empirically weak, something that is consistent with the prediction made by the presented model of what I-languages are, and what their margin of variation is.
The diagram in Figure 1 represents any I-language (i.e., the equivalent of a natural organism). As I have pointed out, it is obvious that every I-language has a variable component (the externalization component), therefore susceptible of adaptation to the environment (although to a lesser degree than it is assumed in models that conceive languages as purely cultural objects). But from this point of view, the notion of environment cannot be the same I have been using. What is the language external medium to which these variable parts could have adapted?
It is not a simple question. The structure of Figure 1 may be interpreted as a sandwich, so that only the outer layers would be susceptible to contact with the environment. Thus, we could consider that the CI and SM systems are “more external” than the computational system. The CI part of any language may be in contact with the rest of the conceptual system of people, so that it would then be expected that certain aspects of the physical, social, and cultural environment in which people develop and live can have an influence on the range of available concepts and notions. This would explain a relatively trivial aspect of the adaptation of languages to the environment, that of the substantive lexicon (Regier et al., 2016). In a culture with highly developed technology there will be words and phrases to denote scientific instruments, techniques, and concepts not found in languages spoken by hunter-gatherer communities, which, on the other hand, would have areas of the lexicon relating to wildly occurring food, animals, and methods of survival unrecognized in the languages of modern urban communities. Changes in culture, technology, and lifestyle often lead to changes in the lexical inventory that we require in everyday life. When a society moves from a rural to an industrialized life, the most widely used lexical inventory also changes. In this area, as pointed out by Ladd et al. (2015), several quantitative studies have shown that there is a correlation between environmental factors (latitude, ultra violet radiation) and the size of the lexical repertoire of color terms. But the differences in the type of conceptual elements that have specific lexical expression are not related to the morphosyntactic structure of languages. Indeed, languages spoken by supposedly simpler societies, hunter-gatherer societies, often have greater morphosyntactic complexity (greater “maturity” in the sense used by Dahl, 2004) than many European languages such as English or Romance languages.
On the other side of the sandwich, we have a sensory-motor system, which in oral languages corresponds to the vocal-auditory system. It is conceivable that certain aspects of the physical environment may bias the kind of sounds most used in some languages (see Everett et al., 2016), but again there would be very limited effects on the morphosyntactic structure of languages.
So, which environmental factors could have molded the historical drift of the morphosyntactic systems of languages? It is quite possible that such factors do not exist or have a weak effect, since the structural typology seems to be relatively isolated from the semantic and material dimension of languages and does not seem to fit them. But if we were to look for them, the place to start is within the brain.
The Brain Internal Environment: Language Learning and Language Processing
According to the model I have described here, the object of study, from a cognitive perspective, is not that of languages understood as social institutions, but the I-languages that reside in the minds/brains of individuals. In this context it is imperative that we recall that the only environment with which “mental organs” are in direct contact is the brain itself. If there is an “external” medium to which I-languages can adapt, it must be internal to the mind/brain.
It may be argued that many of the most notable changes that have been documented in the history of languages have contact with other languages as a crucial factor. And, indeed, it is indisputable that language contact has much more effect on linguistic phenotypes than the social or physical environment in which people live. But languages do not come into direct contact within the physical environment or in society, but only in the brains of speakers. Language A can only have influence on language B if the speaker of B has some kind of knowledge of language A. In our terms we could say that the development of a new lexical interface can affect the previous lexical interface, which can alter the linguistic emissions that the new generation of speakers will use to develop their own lexical interface.
Natural evolution is only possible thanks to the reproduction of organisms, and linguistic change is only possible thanks to the transmission of languages from generation to generation. Much of the structure of an I-language is transmitted from parents to children along with the rest of their biological endowment, but obviously the variable parts of language are learned (internalized) from environmental linguistic stimuli. As I have already noted, this is the phase in which mutations in the lexical interface can occur. These mutations, depending on their range of transmission, can give rise to linguistic changes and, ultimately, to what we see as a different language. The task of the child who learns a language is to reproduce in her mind/brain the lexical interface of her interlocutors, a typically insecure (“abductive,” cf. Andersen, 1973) procedure that is at the basis of linguistic change.
As Dahl (2004) has shown, the usual dynamics of linguistic change produce an increase in morphosyntactic complexity (maturity) up to a certain limit, and thereafter such complexity tends to be maintained. The degree of maturity of a language is measured in terms of the quantity of structures involving a previous derivational history, i.e., non-universal processes that can only be explained by long previous evolutionary chains, such as inflectional and derivative morphology, incorporation, the existence of phonological tone, case marking, or ergativity. However, we might note that according to the model presented in Figure 1 this natural increase in linguistic complexity actually amounts to an increase in the complexity of the lexical interface, not the whole language itself. In this sense, no languages are more complex than others, but there are languages with more complex lexical interfaces than others. This is an important difference. The notable grammatical differences between, on one extreme, Georgian and, on the other, Tok Pisin, do not imply differences in the deep layers of structure (basically the CI system and the computational system), but rather differences in the historical evolution of their externalization components. The proof of this is that the two languages serve their users in carrying out the same cognitive and communicative functions.
The initial intuition here is simple: the more prior uninterrupted history, the greater morphosyntactic complexity, and vice versa. In fact, McWhorter (2011) argues that the natural state of a language, i.e., when no drastic disturbances in its transmission from generation to generation have occurred, is “highly complex, to an extent that seems extreme to speakers of languages like English” (2011, p. 1). It seems clear that the brain of human children is able to internalize lexical interfaces as complex as those of Native American languages or Caucasus languages, typical examples of “mature” systems in Dahl's sense. Neither the brains of other organisms nor the brains of the majority of human adults are as efficient in the internalization of arbitrary systems of gender and noun classifiers, agreement patterns, or quirky cases (not to mention phonological systems). Consequently, McWhorter hypothesizes that whenever we find languages with low degrees of morphosyntactic complexity it is because such languages have been interrupted in their normal accumulation of complexity; i.e., languages with relatively low degrees of complexity “owe this state to second-language acquisition in the past” (McWhorter, 2011, p. 2). In this category we could include languages like English, Romance languages, Persian, Mandarin Chinese, and Indonesian. Compared to other, related languages (such as Sanskrit, Latin, Greek, or Baltic) these languages (which McWhorter calls Non-Hybrid Conventionalized Second-Language Varieties) are characterized by a loss of complexity that reveals evidence of widespread second-language learning in the past. In fact, Lupyan and Dale (2010) and Bentz and Winter (2013) present quantitative evidence showing that languages spoken by many second language speakers tend to have relatively small nominal case systems compared with languages with low proportions of L2 speakers. According to this model, creoles are extreme cases of the same phenomenon: “where complexity has been lost to a radical degree, we can assume that the language was born in a situation in which adult acquisition was universal” (McWhorter, 2011, p. 2). These cases of suboptimal transmission would therefore be clear examples in which the brains of adult learners have operated as an environmental factor to which some parts of languages have adapted.
Another brain internal potential source of modeling forces for morphosyntactic systems can be found in language use in real time (see Newmeyer, 2005, for a conciliatory synthesis on the division of labor between linguistic and processing principles in grammar development). The model I have presented stipulates that only the externalization component is subject to change and, therefore, to variation. It is therefore expected that processing principles (both in speech production and perception) have a remarkable role in the structure and dynamics of externalization systems (i.e., in the morphological mechanisms of syntax realization), precisely because these systems are relevant to the use of language for communication. In fact, language processing principles (see Hawkins, 2004, for a very explicit model) play their role by relating these two components (the computational system and the lexical interface of Figure 1).
Just by way of illustration, I will consider Bickel et al. (2015) regarding the development and persistence of ergative systems in relation to universal processing preferences. Using experimental evidence, Bickel et al. (2015) propose that there is a universal principle that favors the processing of an initial unmarked NP (in nominative or absolutive case) as an agent (as in John sold a car). When the rest of the sentence shows that this unmarked NP is not an agentive subject (as it would be in an ergative language, which marks the subjects of the transitive verbs), they observed an event-related potential (N-400) signaling a reanalysis of the role of the first NP (for example, as a patient argument). Bickel et al. hypothesize that this principle is “species-wide and independent of the structural affordances of specific languages” (Bickel et al., 2015, p. 2) and that, as such, “the principle favors the development and maintenance of case-marking systems that equate base-form cases with agents rather than with patients” (Bickel et al., 2015, p. 2), i.e., nominative-accusative systems over ergative-absolutive ones. Using a large database of linguistic changes in various language families (617 languages in total) they note that of the two possible historical changes, ergative > accusative or accusative > ergative, languages show a clear bias toward the former:
“Languages tend to avoid ergatives when they evolve over time: if a language has ergative case marking, it is more likely to lose than to keep it, and if a language lacks ergative case marking, it is unlikely to develop it. To be sure, ergative cases can arise and be maintained for a while, but the probabilities of this are always lower than the probabilities of avoiding ergatives” (Bickel et al., 2015, p. 18).
If Bickel et al.'s conclusions are correct, we would again have a clear example of how a language-external (but mind-internal) factor can condition the adaptation of languages in their processes of change. However, this also leads us to an important conclusion, one at the heart of our present discussion: even though a general principle of processing exerts a measurable pressure on linguistic systems, the inertia of the language's previous history is capable of overcoming it, showing that morphosyntactic structure is stubbornly resistant to external adaptive pressures, even though they are internal to the mind/brain and supposedly universal.
It is important to note that ergative systems are mature systems in Dahl's sense, which would also explain, at least in part, both the unequal statistical distribution of the two types of languages, and the historical bias documented by Bickel et al. The relevant fact for us here is that a language like Basque, which is fully ergative, shows no symptoms of maladjustment and remains fully functional for its users. More relevant still, there are processes of historical development of ergativity (otherwise, ergative languages would never have existed), which show that grammatical structure is largely immune to the influence of external (i.e., non-grammatical) factors. Actually, a recent synthesis of the research on processing costs of ergativity in Basque (Zawiszewski, 2017) concludes that there are no profound differences in the mechanisms underlying processing in languages with different case marking systems:
“In general, the electrophysiological pattern found when processing ergative case violations corresponds to that revealed during similar case violations in accusative languages (…) and thus indicate that the mechanisms underlying language comprehension are comparable across languages with a different case morphology.” (Zawiszewski, 2017, p. 706).
Conclusions
If we adopt McWhorter's theory, we could say that adult brains have influenced the historical development of some human languages to a decisive extent. From an externalist view of languages, it could be said that some languages have adapted to (non-flexible) mature brains, simplifying their historical accretions and rendering themselves easier to be learned. But from the internalist point of view, this statement is unsatisfactory. The externalist approach tends to identify languages with their lexical interfaces, and this identification, at least in part, is behind the different appreciation of the degree of adaptation of languages to the environment. From an internalist point of view, the notion of adaptation of languages to their environment is only acceptable in a weak sense. According to my argument, weak means that only relatively superficial aspects of languages can be explained as adaptations to extralinguistic reality. I do not intend to conclude that statements such as the following are incorrect:
“[L]inguistic differences, from sounds to grammars, may also reflect adaptations to different environments in which the languages are learned and used. The aspects of the environment that could shape language include the social, the physical, and the technological” (Lupyan and Dale, 2016, p. 1).
Of course, as reflected in the model of Figure 1, every language has a cultural component (internalized from the environment) that is susceptible to change and, therefore, to vary in relation to external factors (i.e., adaptation). However, statements like the previous one suggest that this process of adaptation is sufficient to explain the structure of languages and their typology, and that conclusion is what I have tried to put into question in this contribution.
Many and diverse external and internal factors have left their mark on languages, especially in their systems of externalization, but I do not believe that this in itself allows us to claim that the structure of languages is essentially a matter of adaptation to the environment.
Author Contributions
The author confirms being the sole contributor of this work and approved it for publication.
Funding
The present research has been funded by the Spanish AEI and Feder (EU) to grant FFI2017-82460-P.
Conflict of Interest Statement
The author declares that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
Baker, M. C. (2003). Linguistic differences and language design. Trends Cogn. Sci. 7, 349–353. doi: 10.1016/S1364-6613(03)00157-8
Bentz, C., and Winter, B. (2013). Languages with more second language learners tend to lose nominal case. Lang. Dyn. Change 3, 1–27. doi: 10.1163/22105832-13030105
Berwick, R. C., and Chomsky, N. (2011). “The Biolinguistic program: the current state of its development,” in The Biolinguistic Enterprise, eds A. M. Di Sciullo and C. Boeckx, (Oxford: Oxford University Press), 19–41.
Berwick, R. C., and Chomsky, N. (2016). Why Only Us. Boston, MA: The MIT Press. doi: 10.7551/mitpress/9780262034241.001.0001
Bickel, B., Witzlack-Makarevich, A., Choudhary, K. K., Schlesewsky, M., and Bornkessel-Schlesewsky, I. (2015). The neurophysiology of language processing shapes the evolution of grammar: evidence from case marking. PLoS ONE 10:e0132819. doi: 10.1371/journal.pone.0132819
Cercle Linguistique de Prague (1929). Thèses présentées au Premier Congrès des philologues slaves. Travaux Linguist. Prague 1, 5–29.
Chomsky, N. (2007). “Approaching UG from below,” in Interfaces + Recursion = language?” Chomsky's Minimalism and the View from Semantics, eds U. Sauerland, and H.-M. Gärtner (Berlin: Mouton de Gruyter), 1–30.
Dahl, Ö. (2004). The Growth and Maintenance of Linguistic Complexity. Amsterdam; Philadelphia, PA: John Benjamins.
Dixon, R. M. W. (1994). Ergativity. Cambridge: Cambridge University Press. doi: 10.1017/CBO9780511611896
Dixon, R. M. W. (1997). The Rise and Fall of Languages. Cambridge: Cambridge University Press. doi: 10.1017/CBO9780511612060
Dryer, M. S., and Haspelmath, M. (eds.). (2013). The World Atlas of Language Structures Online. Leipzig: Max Planck Institute for Evolutionary Anthropology. Available online at http://wals.info (accessed September 25 2017).
Evans, N., and Levinson, S. C. (2009). The myth of language universals: language diversity and its importance for cognitive science. Behav. Brain Sci. 32, 429–448. doi: 10.1017/S0140525X0999094X
Everett, C., Blasí, D. E., and Roberts, S. G. (2016). Language evolution and climate: the case of desiccation and tone. J. Lang. Evol. 1, 33–46. doi: 10.1093/jole/lzv004
Everett, D. L. (2005). Cultural Constraints on grammar and cognition in Pirahã. Curr. Anthropol. 46, 621–646. doi: 10.1086/431525
Gould, S. J. (1996). “The pattern of life's history,” in The Third Culture. Beyond the Scientific Revolution, ed J. Brockman (New York, NY: Simon & Schuster), 51–73.
Hauser, M. D., Chomsky, N., and Fitch, W. T. (2002). The faculty of language: what is it, who has it, and how it evolved? Science 298, 1569–1579. doi: 10.1126/science.298.5598.1569
Hurford, J. R. (1992). “An approach to the phylogeny of the language faculty,” in The Evolution of Human Languages, eds J. A. Hawkins and M. Gell-Mann (Reading MA: Addison-Wesley), 273–303.
Kauffman, S. A. (1993). The Origins of Order. Self-organization and Selection in Evolution. London: Oxford University Press.
Keenan, E. (2002). “Explaining the creation of reflexive pronouns in English,” in Studies in the History of the English Language: A Millennial Perspective, eds D. Minkova and R. Stockwell (Berlin; New York, NY: Mouton de Gruyter), 325–354. doi: 10.1515/9783110197143.3.325
Kirby, S. (1999). Function, Selection, and Innateness. The Emergence of Language Universals. Oxford: Oxford University Press.
Koerner, K. (ed.). (1983). Linguistics and Evolutionary Theory. Three Essays. Edited by A. Schleicher, E. Haeckel, and W. Bleek. Amsterdam; Philadelphia, PA: John Benjamins.
Labov, W. (1963). The social motivation of a sound change. Word 19, 273–309. doi: 10.1080/00437956.1963.11659799
Ladd, R. D., Roberts, S. G., and Dediu, D. (2015). Correlational studies in typological and historical linguistics. Annu. Rev. Linguist. 1, 221–241. doi: 10.1146/annurev-linguist-030514-124819
Longobardi, G. (2001). Formal syntax, diachronic Minimalism, and etymology: the history of French chez. Linguist. Inq. 32, 275–302. doi: 10.1162/00243890152001771
Longobardi, G. (2003). Methods in parametric linguistics and cognitive history. Linguist. Variat. Yearbook 3, 101–138. doi: 10.1075/livy.3.06lon
Lupyan, G., and Dale, R. (2010). Language structure is partly determined by social structure. PLoS ONE 5:e8559. doi: 10.1371/journal.pone.0008559
Lupyan, G., and Dale, R. (2016). Why are there different languages? The role of adaptation in linguistic diversity. Trends Cognit. Sci. 20, 649–660. doi: 10.1016/j.tics.2016.07.005
Mayr, E. (1942). Systematics and the Origin of Species from the Viewpoint of a Zoologist. New York, NY: Columbia University Press.
McWhorter, J. H. (2011). Linguistic Simplicity and Complexity. Why do Languages Undress? Boston, MA; Berlin: The Gruyter Mouton.
Mendívil-Giró, J.-L. (2006). “Languages and species: limits and scope of a venerable comparison,” in The Biolinguistic Turn. Issues on Language and Biology, eds J. Rosselló, and J. Martín (Barcelona: PPU), 82–118.
Mendívil-Giró, J.-L. (2012). “The myth of language diversity, in language, from a biological point of view,” eds C. Boeckx, M. C. Horno, and J. L. Mendívil-Giró (Newcastle: Cambridge Scholars), 85–133.
Mendívil-Giró, J.-L. (2014). What are languages? A biolinguistic perspective. Open Linguist. 1, 71–95. doi: 10.2478/opli-2014-0005
Mendívil-Giró, J.-L. (2016). El cambio lingüístico. Sus Causas, Mecanismos y Consecuencias. Madrid: Síntesis.
Moreno, J.-C., and Mendívil-Giró, J.-L. (2014). On Biology, History and Culture in Human Language. A Critical Overview. Sheffield: Equinox.
Nevins, A., Pesetsky, D., and Rodrigues, C. (2009). Pirahã exceptionality: a reassessment. Language 85, 355–404. doi: 10.1353/lan.0.0107
Newmeyer, F. J. (2005). Possible and Probable Languages. A Generative Perspective on Linguistic Typology. Oxford: Oxford University Press.
Nichols, J. (1992). Linguistic Diversity in Space and Time. Chicago, IL: University of Chicago Press.
Perkins, R. D. (1988). “The covariation of culture and grammar,” in Studies in Syntactic Typology, eds. M. Hammond, E. Moravcsik, and J. Wirth (Amsterdam; Philadelphia, PA: Benjamins), 359–378. doi: 10.1075/tsl.17.22per
Pinker, S. (2007). “Response to Recursion and Human Thought: Why the Pirahã Don't Have Numbers”, A Talk with Daniel L. Everett. Available online at: https://www.edge.org/conversation/recursion-and-human-thought (accessed September 25, 2017).
Regier, T., Carstensen, A., and Kemp, C. (2016). Languages support efficient communication about the environment: words for snow revisited. PLoS ONE 11:e0151138. doi: 10.1371/journal.pone.0151138
Richards, N. (2016). Contiguity Theory. Cambridge MA: The MIT Press. doi: 10.7551/mitpress/9780262034425.001.0001
Schleicher, A. (1863). Die Darwinsche Theorie und die Sprachwissenschaft. Weimar, H. Böhlau (Quoted from the 1869 English version Darwinism Tested by the Science of Language by J. C. Hotten included in Koerner, ed., 1–71).
Seely, J. (1977). An ergative historiography. Historiogr. Linguist. 4, 191–206. doi: 10.1075/hl.4.2.04see
Starke, M. (2009). Nanosyntax: a short primer to a new approach to language. Nordlyd 36, 1–6. doi: 10.7557/12.213
Keywords: language change, language evolution, adaptation, language typology, evolutionary theory, language diversity, faculty of language, I-language
Citation: Mendívil-Giró J-L (2018) Why Don't Languages Adapt to Their Environment? Front. Commun. 3:24. doi: 10.3389/fcomm.2018.00024
Received: 27 September 2017; Accepted: 28 May 2018;
Published: 14 June 2018.
Edited by:
Antonio Benítez-Burraco, Universidad de Sevilla, SpainReviewed by:
Giuseppe Longobardi, University of York, United KingdomSean Roberts, University of Bristol, United Kingdom
Copyright © 2018 Mendívil-Giró. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: José-Luis Mendívil-Giró, amxtZW5kaUB1bml6YXIuZXM=