



- Inserm UMR894 - Center for Psychiatry and Neuroscience, Université Paris Descartes, Sorbonne Paris Cité, Paris, France
Somatostatin is highly expressed in mammalian brain and is involved in many brain functions such as motor activity, sleep, sensory, and cognitive processes. Five somatostatin receptors have been described: sst1, sst2 (A and B), sst3, sst4, and sst5, all belonging to the G-protein-coupled receptor family. During the recent years, numerous studies contributed to clarify the role of somatostatin systems, especially long-range somatostatinergic interneurons, in several functions they have been previously involved in. New advances have also been made on the alterations of somatostatinergic systems in several brain diseases and on the potential therapeutic target they represent in these pathologies.
Neuroanatomy of Somatostatinergic System
Somatostatin Peptides in the Brain
Somatostatin14 (SRIF, somatotropin release inhibiting factor) was serendipitously discovered in 1972 by Roger Guillemin and colleagues who were aiming to purify and characterize growth hormone (GH)-releasing hormone from sheep hypothalamus (Brazeau et al., 1973). Soon thereafter, an N-terminally extended peptide, SRIF28, was purified from the gut. Both peptides, arising from a common propeptide encoded from a single gene, were found in the mammalian nervous system where SRIF14 is the predominant form (for review, see Epelbaum, 1986).
Two brain SRIF-related bioactive peptides have been discovered later. Cortistatin (CST) has been cloned in 1996 (De Lecea et al., 1996) and shares 11 amino acids with SRIF (Figure 1). CST peptides are predicted to occur as 14-AA or 17-AA short forms in rodents and humans, respectively, and a 29-AA extended form in both species. CST is mainly restricted to the cerebral cortex and the hippocampus in the central nervous system (CNS). CST has been implicated in several brain functions such as learning and memory, regulation of sleep/wakefulness rhythms and it is suspected to have an anticonvulsant activity (for review, see de Lecea, 2008). Recently, bioinformatics analyses of evolutionary conserved sequences identified neuronostatin, a 13-AA amidated peptide also encoded by the somatostatin gene. Mostly found in pancreas, spleen, and brain, it is involved in metabolic, cardiovascular, and neuronal functions (Samson et al., 2008).
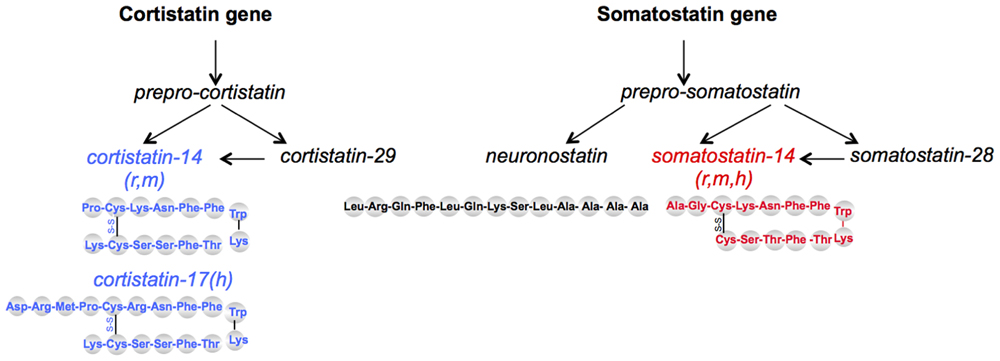
FIGURE 1. Schematic representation of somatostatin-related peptides. SRIF14 and CST14 come from two distinct genes but bind the five mammalian sst receptors. Neuronostatin is encoded by the same gene as SRIF but does not bind SRIF receptors; its effects seem mediated through the melanocortin system. r, rat; m, mouse; h, human.
Somatostatin induces many transduction mechanisms in transfected systems (for review, see Lahlou et al., 2004; Olias et al., 2004), but deciphering the physiological actions of the native receptors in situ remains an intense field of study. The last decade showed increasing progress in understanding the role of SRIF in brain functions using molecular, pharmacological, and behavioral approaches. The development of innovative molecular, genetic, and imaging tools now allows to go a step further and to assess the cellular contribution of SRIF-expressing cells in neuronal networks ex vivo and soon in vivo. In this review we will give an overview of the latest findings concerning SRIF systems in brain, report some recent data concerning their synaptic actions and their physiological roles within the brain, in normal or pathological conditions.
Somatostatinergic Networks in the Brain
Somatostatin is ubiquitously expressed in mammalian brain, including humans (Figure 2A). SRIF-immunoreactivity is found at high level in the mediobasal hypothalamus and median eminence, amygdala, preoptic area, hippocampus, striatum, cerebral cortex, sensory regions, and the brainstem (for review, see Epelbaum, 1986; Viollet et al., 2008).
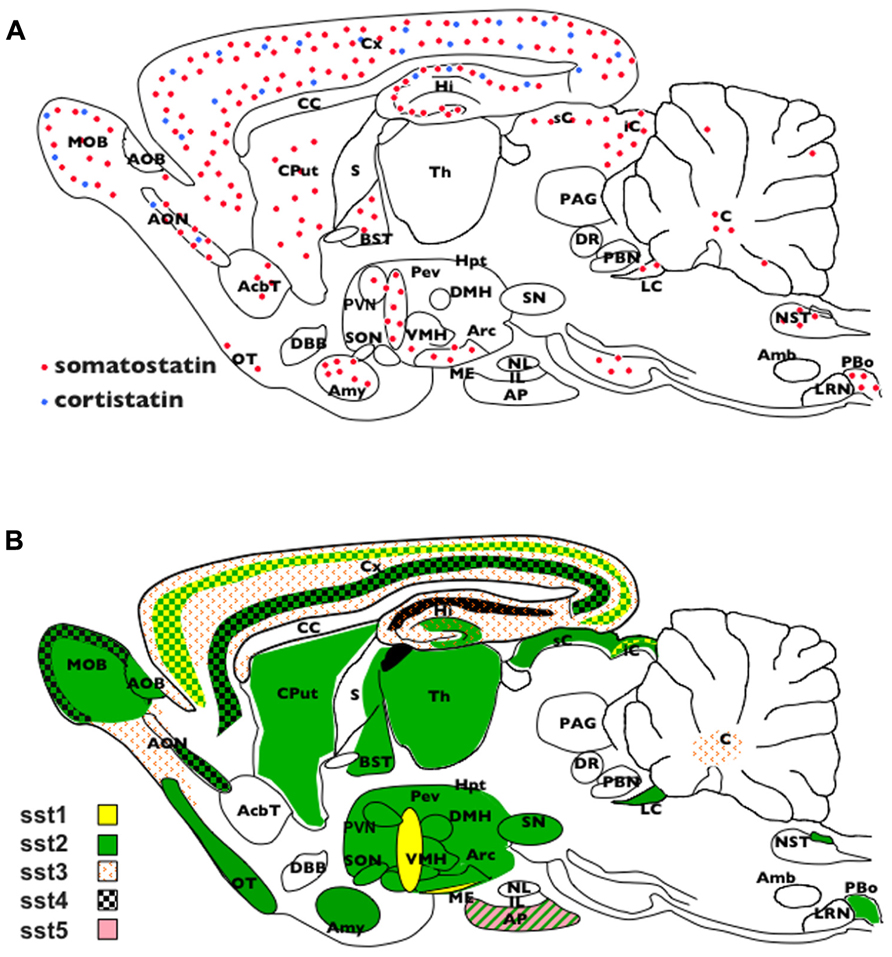
FIGURE 2. Schematic representation of somatostatin and cortistatin distribution in the mouse brain. (B) Schematic representation of somatostatin receptors distribution in the mouse brain. AcbT, nucleus accumbens; Amb, ambiguous nucleus; AP, anterior pituitary; AOB, accessory olfactory bulb; Amy, amygdala; Arc, arcuate nucleus; BNST, bed nucleus of the stria terminalis; CC, corpus callosum; CPut, caudate putamen; Cx, cortex; DBB, diagonal band of broca; DMH, dorsomedian hypothalamus; DR, dorsal raphe; iC, inferior colliculus; IL, intermediate lobe of the pituitary; Hi, hippocampus; Hpt, hypothalamus; LRN, lateral reticularis nucleus; ME, median eminence; MOB, main olfactory bulb; NL, neural lobe of the pituitary; NTS, nucleus of the solitary tract; OT, olfactory tubercle; PAG, periaqueductal gray; PBo, pre-Bötzinger nucleus; PeV, periventricular nucleus; PVN, paraventricular nucleus; sC, superior colliculus; S, septum; SON, supraoptic nucleus; SN, substantia nigra; Th, thalamus; VMH, ventromedian hypothalamus.
Somatostatin peptide colocalizes with gamma-aminobutyric acid (GABA), a major inhibitory neurotransmitter and labels mostly non-glutamatergic cells in the brain. In order to target SRIF interneurons in situ, most recent studies took advantage of rodent models expressing the green fluorescent protein (GFP) under the control of the GAD67 promoter to visualize GABAergic populations more easily. In the GIN (GFP-expressing inhibitory neurons) strain (Oliva et al., 2000) nearly all GFP cells stained for SRIF while in the GAD67-GFP strain, SRIF immunohistochemistry labels 37% of total GFP cells (Ma et al., 2006). The recent development of specific Cre recombinase and knock-in inducible driver lines for SRIF (Taniguchi et al., 2011) opens promising avenues to study SRIF functions at the cellular level combined to optogenetic and imaging tools.
Previous immunohistochemical and tracing studies have identified two main categories of SRIF neurons: those acting locally in a given structure within microcircuits (interneurons) and those projecting to a distant structure (long-projecting neurons). Nevertheless, recent data using GFP transgenic mice revisit previous anatomical records by demonstrating that some formerly called interneurons also belonged to the projecting neurons category. The different kinds of GABAergic interneurons are classified according to their molecular, physiological, and morphological properties. Immunohistochemical characterization of neuronal populations in rat cortex initially stated, based on calcium-binding proteins and peptides expressions that parvalbumin (PV), SRIF, calretinin, and cholecystokinin labeled four main non-overlapping chemical classes of interneurons (Xu et al., 2006). However, several studies later reported a significant colocalization of SRIF and calretinin in mouse brain (Xu et al., 2006; Kosaka and Kosaka, 2007; Lepousez et al., 2010a), pointing out species-dependent variations in the repertoire of calcium-binding proteins and neuropeptides. It seems that neuronal populations immunoreactive for calbindin or the neuropeptide Y strongly overlap with the somatostatinergic population in rats whereas calretinin is preferentially coexpressed with SRIF in the mouse.
Recent morphological and electrophysiological studies using GIN mice focused on SRIF-expressing populations in cortical circuits. In mouse cortex, parvalbumin- and SRIF-expressing neurons respectively constitute 40 and 30% of the total GABAergic neurons, calretinin being expressed in 50% of the somatostatinergic population (Rudy et al., 2011). The remaining cortical inhibitory interneurons, expressing ionotropic serotonergic receptor 5HT3a, include VIP- and NPY-positive subpopulations whose partial colocalization with SRIF has been reported (Gonchar et al., 2007; Xu et al., 2010; Rudy et al., 2011). SRIF-positive interneurons are homogeneously distributed in all cortical layers (2–6), as compared to PV-positive inhibitory interneurons that are concentrated in the upper part of the layer (Perrenoud et al., 2012).
The major class of SRIF interneurons, the Martinotti cells, have ascending axons that arborize and spread horizontally in layer 1, targeting the distal dendritic parts of excitatory pyramidal neurons (for review, see Viollet et al., 2008). Excitatory inputs onto Martinotti cells are generally strongly facilitating, allowing feedback inhibition of the excited pyramidal cell that increases as function of the rate and the duration of the presynaptic discharge (Kapfer et al., 2007; Silberberg and Markram, 2007). The relative distance between excitatory and interneurons inputs may also impact feedback selectivity and grade, inhibition being stronger for closer inputs. A recent study using a two-photon microscopy approach coupled to uncaged glutamate in cortical slices of GIN mouse mapped the inhibitory network between SRIF-positive interneurons and pyramidal cells at the single-cell resolution (Fino and Yuste, 2011). Whatever the pyramidal cell stimulated, it led to a dense innervation of the surrounding somatostatinergic interneurons, with activity related to the proximity of the cells. Notably, this inhibitory connectivity looked unspecific as all inhibitory interneurons were locally connected to every sampled pyramidal cells regardless whether these were connected among themselves or not. This dense circuit and the fact that somatostatinergic neurons electrically communicate via gap junctions (Ma et al., 2006; Hu et al., 2011) favors the hypothesis that the entire somatostatinergic population belongs to a same inhibitory cortical circuit, contradicting the hypothesis of specific inhibitory cortical subnetworks.
Additional classes of cortical SRIF inhibitory interneurons have been recently described according to their localization, intrinsic firing properties, expression of molecular markers, and connectivity (Gonchar et al., 2007; McGarry et al., 2010). On one hand, calretinin expression was proposed as a distinctive marker (Rudy et al., 2011), since its expression is associated to distinct neuronal morphology and connectivity in populations with distinct ontogenic origin (Sousa et al., 2009; Xu et al., 2010). On the other hand, two novel SRIF-positive subtypes were identified after cluster and principal component analysis of a whole range of morphological or electrophysiological parameters (McGarry et al., 2010). These cell types have some similarities to neurons labeled in a GABAergic-GFP strain distinct from GIN (X94 strain; Ma et al., 2006), such as the lack of expression in the layer 1, but they target different cortical layers. Future identification of their respective calcium-binding proteins and neuropeptides repertoire as well as their molecular phenotype will help to conciliate these independent classifications based on morphological and electrophysiological properties.
Somatostatin is found in most sensory systems, i.e., retina (Thermos, 2003; Cervia and Bagnoli, 2007). In the olfactory system, SRIF expression has also been described in sparse short-axon cells scattered in the deep part of the granule cell layer (the main site of intrinsic inhibitory neurons; Shipley and Ennis, 1996; Eyre et al., 2009) and in the peripheral glomerular layer (which receives sensory inputs) in some species (Hwang et al., 2004). Recently, a novel type of somatostatinergic interneurons has been described as predominant in the murine olfactory bulb and specific to this species (Lepousez et al., 2010a). SRIF-positive somata and dendritic fields are restricted to the layer of the olfactory bulb where intrinsic GABAergic interneurons and bulbar principal cells interact through dendrodendritic reciprocal synapses to initiate local gamma oscillations responsible for odor processing. Electron microscopy evidences suggest that SRIF-positive interneurons also establish reciprocal dendrodendritic synapses with the bulbar principal cells (mitral cells). SRIF-positive neurons have also been described downstream in the olfactory pathway; SRIF interneurons constitute a major GABAergic population in the pars principalis of the anterior olfactory nucleus and in the olfactory tubercle (Brunjes et al., 2011). In both piriform and entorhinal cortices, two cortical structures involved in the processing of odor coding, multipolar SRIF-positive interneurons displaying Martinotti-like morphological and electrical properties are found in the deep (Young and Sun, 2009; Saiz-Sanchez et al., 2010; Suzuki and Bekkers, 2010) and superficial (Saiz-Sanchez et al., 2010; Tahvildari et al., 2012) layers respectively.
As mentioned before, in addition to the somatostatinergic interneurons acting within microcircuits, long-projecting somatostatinergic cells have been described in several regions (Viollet et al., 2008). As glutamatergic pyramidal cells projections do, long-range inhibitory connections mediate communication between multiple brain areas. Long-range inhibitory terminals have a larger diameter and a thicker myelin layer than excitatory projection neurons, suggesting that inhibitory signal may precede the arrival of excitation in co-innervated cortical areas (Jinno et al., 2007). Long-range projecting SRIF-containing neurons are encountered in numerous brain areas (for review, see Viollet et al., 2008) such as the hippocampus (Jinno et al., 2007), the cerebral cortex (Tomioka et al., 2005), and the amygdala (McDonald et al., 2012). For instance, virtually all non-pyramidal neurons in the amygdala that have long-range projecting axons to the basal forebrain in the rat express SRIF (McDonald et al., 2012). Figure 3 represents the projections of all long-range somatostatinergic interneurons known to date in the brain.
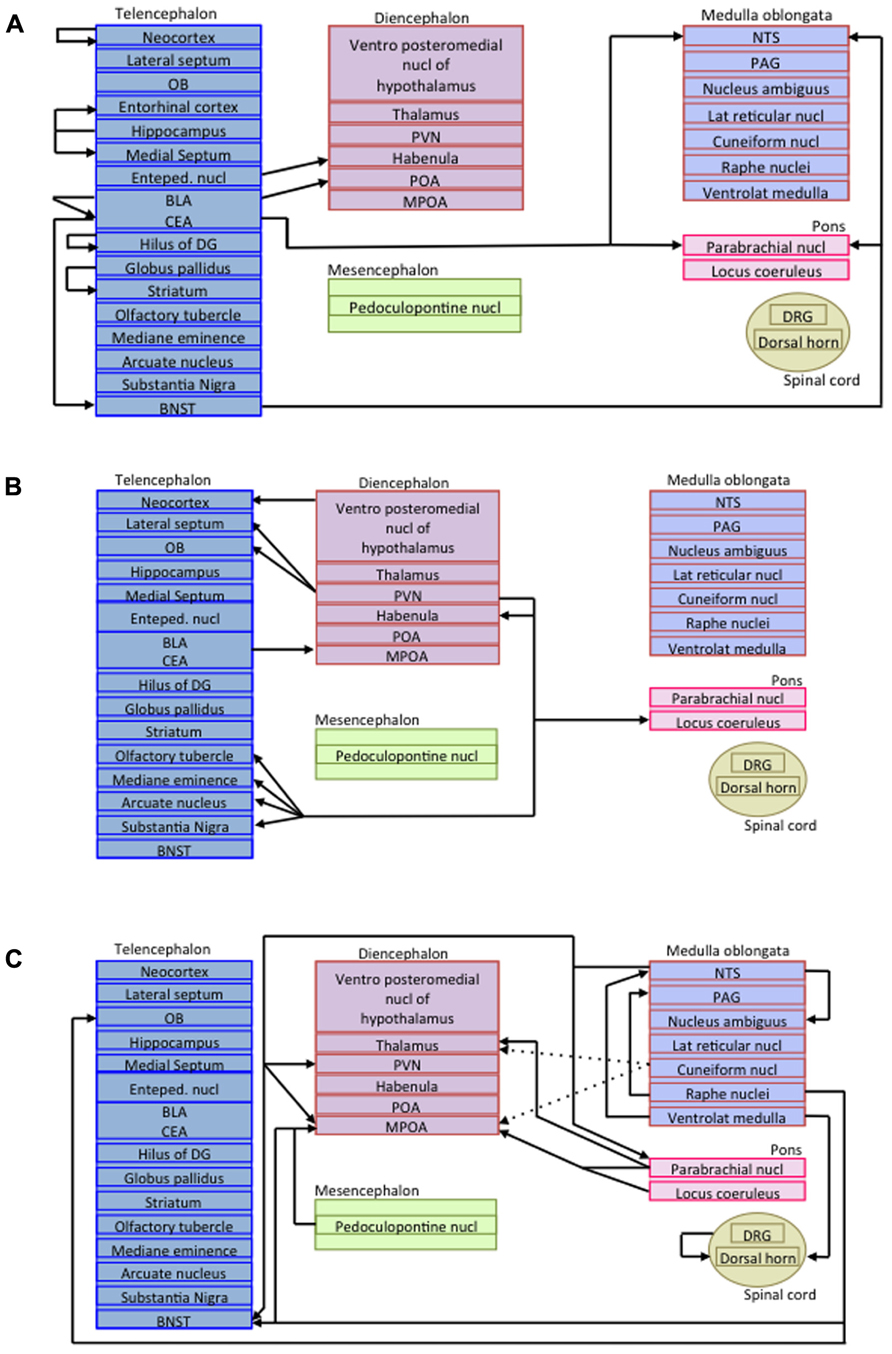
FIGURE 3. Schematic representations of long-range somatostatinergic interneurons in the central nervous system. (A) Telencephalic efferent projections to the rest of the brain. (B) Efferent projections arising from the diencephalon and projecting to the telencephalon and the pons. (C) Efferent projections arising from the mesencephalon, the medulla oblongata, the pons and the spinal cord. Enteped, entopeduncular; BLA, basolateral amygdala; BNST, bed nucleus of the stria terminalis; CEA, central amygdala; DG, dentate gyrus; DRG, dorsal root ganglia; MPOA, medial preoptic area; NTS, nucleus of the solitary tract; OB, olfactory bulb; PAG, periaqueductal gray; POA, preoptic area; PVN, paraventricular nucleus.
Somatostatin Receptors in the Central Nervous System
Autoradiographic studies characterized initially two SRIF binding site according to their affinity for the synthetic agonist octreotide and their pattern of expression. In the early 1990s, five receptors (sst1-5) belonging to the G-protein-coupled receptors (GPCRs) family were cloned and characterized from various species. Sequence homology is 39–57% among the five subtypes, each being highly conserved across species. They activate multiple intracellular targets (Olias et al., 2004) and display distinct internalization and dimerization properties (Csaba et al., 2012). Based on structural, pharmacological, and operational features, they are now divided into two groups displaying nanomolar affinity for both SRIF and CST: SRIF-1 (sst2, sst3, and sst5 receptors) and SRIF-2 (sst1 and sst4 receptors). Figure 2B represents the wide expression of SRIF receptors in the CNS. In contrast to most GPCRs, sst1-5 are unique because their gene coding sequence is devoid of introns. However, this does not preclude the generation of spliced variants such as a shorter isoform of mouse sst2, named sst2B, originating by the excision of a cryptic intronic sequence (Vanetti et al., 1992), and spliced variants of sst5 in human and rodents (Córdoba-Chacón et al., 2011). While some data suggested that CST also acts through the proadrenomedullin receptor MgrX2 or the ghrelin orexigenic peptide receptors, the existence of specific CST receptors has not been demonstrated (Siehler et al., 2008). Neuronostatin does not bind to SRIF receptors, but some of its effects seem mediated through the central melanocortin system (Yosten et al., 2011). Recent findings have shown that neuronostatin is involved in regulating depressive behavior and nociception (Yang et al., 2011a,b, 2012).
The extended distribution of sst2 receptors in the CNS together with studies using subtype selective SRIF analogs in both in vivo and in vitro experiments, suggested that these subtypes are the major players in the SRIF receptor family. They have broad inhibitory effects in many neuronal networks including cortex, hippocampus, limbic regions, and sensory systems (retina and olfactory system; Viollet et al., 2008; Lepousez et al., 2010a; Radojevic et al., 2011). The sst1 receptor may function as an autoreceptor in basal ganglia, hypothalamus, and sensory systems (Thermos et al., 2006), and in the hippocampus (de Bundel et al., 2010). sst3 receptors are localized to mature neuronal cilia in most brain regions (Stanić et al., 2008), and pharmacological or genetic blockade of sst3 have marked behavioral effects (Einstein et al., 2010). sst4 receptors are highly expressed in the olfactory bulb, cortex, and hippocampus, where their role remains to be clarified. In the mouse they modulate epileptic activity, whereas in the rat it seems that this effect is largely related to sst2 receptors. Hippocampal sst4 have also been involved in cognitive processes (Gastambide et al., 2009; Sandoval et al., 2011), functionally interacting with sst2 (Dutar et al., 2002; Gastambide et al., 2010). sst5 receptors mediate regulation of GH release and inhibit cell proliferation by SRIF/CST, mainly through sst2/sst5 receptors interaction. The detection of functional truncated forms of sst5 suggests that they could interfere in and modulate those interactions (Córdoba-Chacón et al., 2011).
Somatostatinergic Functions in the Brain
Neuronal Actions of SRIF
Presynaptic Mechanisms
Somatostatin, like other neuropeptides, can modulate CNS excitability via presynaptic mechanisms (Baraban and Tallent, 2004). In rat hippocampus and cortex, SRIF induces a presynaptic inhibition of excitatory neurotransmission leading to a decrease in glutamate release and in the amplitude of evoked synaptic responses (Ishibashi and Akaike, 1995; Boehm and Betz, 1997; Tallent and Siggins, 1997; Grilli et al., 2004). The SRIF-induced decrease in glutamate release is explained by an inhibition of excitatory transmission via a G-protein of the Gi/Go family and modulation of calcium channels. Indeed, SRIF selectively inhibits N-type Ca2+ channel via the picrotoxin-sensitive G(i)/G(o) protein. Somatostatin can also inhibit N-type Ca2+ channels in the dentate gyrus (Baratta et al., 2002). By these inhibitory effects on excitatory synaptic transmission, SRIF, co-released with GABA on dendritic shafts of principal neurons, increases and prolongs GABA effect. This presynaptic action on Ca2+ conductance could explain, at least in part, the inhibitory effect of SRIF on long-term potentiation in the mouse dentate gyrus (Baratta et al., 2002). Other studies suggest that presynaptic K+ channels modulation may also be involved in the SRIF inhibition of excitatory transmission (Tallent and Siggins, 1997). More precisions on the mechanisms have been given by Grilli et al. (2004), demonstrating on synaptosomal preparations from mouse cerebral cortex that activation of sst2 presynaptic receptors may inhibit the cAMP/PKA pathway stimulated by high potassium concentration, leading to a decrease of the evoked glutamate release. If in the hippocampus, cortex and also hypothalamus, the presynaptic effects of SRIF concern almost exclusively the excitatory transmission (Peineau et al., 2003), SRIF is also able to decrease GABA release in different brain structures, such as the rat basal forebrain (Momiyama and Zaborszky, 2006), the neostriatum (Lopez-Huerta et al., 2008), and the thalamus (Leresche et al., 2000). In the basal forebrain, SRIF presynaptically inhibits both GABA and glutamate release onto cholinergic neurons in a Ca2+-dependent way.
Postsynaptic Mechanisms
Effects of SRIF on intrinsic neuronal membrane properties are well documented. Somatostatin induces a membrane hyperpolarization resulting from the activation of two distinct K+ current, the voltage-sensitive K+ current or M-current (IM; Moore et al., 1988; Jiang et al., 2003), and a voltage-insensitive leak current (Schweitzer et al., 1998). In hippocampal CA1 pyramidal neurons, sst4 seems to be the receptor subtype that couples to IM (Qiu et al., 2008).
In medium spiny neostriatal neurons, SRIF produces a qualitative change in the firing pattern from a tonic regular to an interrupted “stuttering”-like pattern (Galarraga et al., 2007). These authors demonstrated that SRIF changes the firing pattern via sst2-subtype activation, which reduces the small conductance Ca2+-activated K+ currents (SK-channels) and activates large conductance g(K)Ca2+ (GK channels). These results highlight the fact that SRIF is a regulator of cellular function in the striatum. The numerous effects of SRIF on Ca2+ and K+ channel conductance in different structures are reviewed by Cervia and Bagnoli (2007).
A huge amount of literature has tried to define the pharmacological nature of SRIF effects, using agonists and antagonists of SRIF receptors or mice invalidated for receptor subtypes. Results are often controversial and are different in mice and rats (Aourz et al., 2011). Therefore, the classification of SRIF effects is complex and it is accentuated by the description of functional cooperation between different receptor subtypes sst2/sst3, sst2/sst4, sst3/sst4 (Moneta et al., 2002; Gastambide et al., 2010; Aourz et al., 2011). Recent publications suggest that sst3 and sst4 (but not sst1; de Bundel et al., 2010) have potent anticonvulsive properties (Aourz et al., 2011), and that sst2, the major receptor subtype involved in the anticonvulsant effect of SRIF in the hippocampus exerts a functional cooperation with sst3/sst4. In hippocampus, sst1 activation inhibits both NMDA- and AMPA-mediated responses but did not affect the inhibitory transmission (Cammalleri et al., 2009).
Srif-Containing Neurons are Involved in Physiological Functions
Interneurons
A large diversity of inhibitory interneurons is able to exert inhibition on specific compartments of principal cells. Among these populations is the dendrite-targeting SRIF-expressing interneuron located in oriens-lacunosum moleculare of the hippocampus. These SRIF-containing neurons are the only subtype of interneuron that reliably follows synaptic stimulation of the alveus in the theta frequency range via activation of their kainate receptors, suggesting that they play an important role in theta band frequency oscillations (Goldin et al., 2007). Spontaneous activities of inhibitory interneurons have been characterized and SRIF-containing neurons are described in the cortex and piriform cortex as regular-spiking (Kawaguchi and Kubota, 1998; Suzuki and Bekkers, 2010) or low-threshold spiking neurons (Goldberg et al., 2004), often opposed to the fast spiking PV-containing neurons. In the hippocampus, SRIF neurons are locked to the ascending phase of the theta cycle. However, using an optogenetic inhibition of different populations of interneurons, it was recently demonstrated that silencing SRIF interneurons increases burst firing of pyramidal cells without altering the theta phase of spikes (Royer et al., 2012). Applying optogenetic technique to animals trained to run head-fixed on a treadmill belt rich with visual-tactile stimuli, these authors provided evidence that the dendritic (but not somatic) inhibition of pyramidal neurons by SRIF interneurons is critical for controlling spike burst firing during active exploration. They concluded that perisomatic PV-targeting interneurons control the spikes’ theta phase while the dendrite-targeting SRIF interneurons control the rate of discharge. This is in agreement with the fact that dendritic but not somatic GABAergic inhibition is decreased in experimental epilepsy (Cossart et al., 2001). Combining optogenetic stimulation with in vivo two-photon imaging in the mouse visual cortex, Wilson et al. (2012) demonstrate that soma-targeting PV neurons regulate the gain of cortical response, while dendritic-targeting SRIF neurons shift response level and alter stimulus selectivity, leaving response gain unaffected.
Another demonstration of the role of SRIF interneurons in cellular function has been given recently (Gentet et al., 2012). In this study, SRIF neurons recorded in the barrel cortex of awake mice were tonically active during quiet wakefulness but they decreased their firing during whisker sensorimotor processing. This decrease in firing relieves the dendrites of excitatory pyramidal neurons from inhibition.
It is known that inhibitory neurons have diverse roles in physiological and synaptic function, based on their connectivity patterns and intrinsic properties. All the experiments described above demonstrated that SRIF interneurons have a prominent role in the regulation of distal dendrites excitability.
Long-range Projecting Neurons
The long-range projecting somatostatinergic non-pyramidal cells found in the hippocampus target the medial septum and the medial entorhinal cortex (Viollet et al., 2008; Melzer et al., 2012) and more specifically form inhibitory synapses on GABAergic interneurons of these areas. They coordinate activity between distant brain regions, contributing to the generation and the synchronization of rhythmic oscillatory activity in the hippocampus and entorhinal cortex (Melzer et al., 2012). They are therefore involved in spatial and temporal coding. Interestingly, early-generated GABA-containing hub neurons, dendrite-targeting interneurons, express preferentially SRIF and give long-range projecting neurons (Picardo et al., 2011). These superconnected hub cells are present early in the developing hippocampus. They develop a widespread axonal arborization and remain into adulthood. They play a key role in the control of the hippocampal giant depolarizing potentials as well as in the modulation of network dynamics. In the other brain areas, the precise contribution of these long-projecting SRIF neurons in the oscillatory activity still needs to be addressed.
Hypophysiotropic Neurons
Somatotropin release inhibiting factor was initially discovered as a neurohormone that inhibits GH secretion from anterior pituitary somatotroph cells. This function is exerted by hypophysiotropic neurons, located in the anterior periventricular hypothalamic nucleus, which project to the median eminence and release the peptide in the fenestrated capillaries of the hypothalamo–hypophyseal portal vessels; thus directly connecting the brain to the anterior pituitary. SRIF is also a potent inhibitor of many hormonal and exocrine secretions as well as an antiproliferative agent in normal and tumoral tissue (Epelbaum, 1986). SRIF analogs (octreotide and lanreotide) have potent inhibitory effects on hypersecretion, thereby alleviating the symptoms associated with neuroendocrine tumors. Furthermore, the antitumor potential of octreotide is now well documented. Pasireotide, a long-acting SRIF analog, has the advantage of targeting a wider range of SRIF receptors (subtypes 1, 2, 3, and 5) than the analogs previously used in clinical practice (which preferentially target subtype 2) and has a broader spectrum of activity (for review, see Bousquet et al., 2012).
Involvement of SRIF Systems in Sensory, Motor, and Cognitive Functions
Since SRIF systems are widely expressed in CNS, they are involved in numerous functions including nociceptive and vasoconstrictor properties. Here, we will present recent advances about the role of SRIF systems in autonomic responses (digestion, cardiac rate, and respiration) and motor functions as well as cognitive functions such as learning and memory and emotion (for review, see Viollet et al., 2008).
Somatostatinergic Involvement in Sensory Functions
Somatostatin and visual information. Somatostatinergic system is expressed in mammalian retina (for review, see Thermos, 2003; Casini et al., 2005; Cervia et al., 2008), where it is suspected to exert multiple actions on neurons and on retinal physiology. SRIF acts as a positive factor in the retina by regulating homeostasis and protecting neurons against damage. Both sst2 and sst5 somatostatinergic receptors are involved. Indeed, activation of sst2 protects the retina from ischemic insults ex vivo (Mastrodimou et al., 2005) and sst2 as well as sst5 receptor activation protect from excitotoxicity in vivo (Kiagiadaki and Thermos, 2008; Kiagiadaki et al., 2010; Kokona et al., 2012). The severity of angiogenic responses to hypoxia is correlated to the sst2 expression level in the retina (Dal Monte et al., 2007). Moreover, the sst2-preferring agonist octreotide prevents hypoxia-induced VEGF up-regulation (Dal Monte et al., 2009).
Somatostatinergic modulation of olfactory discrimination. Recent studies have shown that SRIF modulates olfactory processing in mice (Lepousez et al., 2010a,b). In mouse main olfactory bulb, SRIF is mainly concentrated in local GABAergic interneurons synaptically connected to the mitral cells by reciprocal dendrodendritic synapses. When activated by an odor, mitral cells synchronize and generate gamma oscillations of the local field potentials that are involved in olfactory processing. Pharmacological or genetic blockade of sst2 transmission in the olfactory bulb of awake animal selectively decreased the gamma oscillations power while pharmacological activation of sst2 had opposite effects. These treatments were respectively correlated to either impairment or improvement of odor discrimination performances of the pharmacologically injected animals. Thus, bulbar endogenous SRIF, presumably released from external plexiform layer interneurons, affects gamma oscillations through the dendrodendritic reciprocal synapse and contributes to olfactory processing.
Involvement of SRIF in Learning and Memory
It has been reported for decades that SRIF plays a role in learning and memory at different stages of information processing. The first studies investigating its role in cognition showed that intracerebroventricular administrations of SRIF improved learning in active avoidance tasks (Bollok et al., 1983; Vecsei et al., 1983; Vecsei and Widerlov, 1988) and prevented electroshock-induced amnesia in passive avoidance paradigms (Vecsei et al., 1983, 1984). Conversely, the depletion of SRIF in the brain by cysteamine (which depletes SRIF levels; Szabo and Reichlin, 1981) produced major memory deficits in passive avoidance (Bakhit and Swerdlow, 1986; Schettini et al., 1988; DeNoble et al., 1989). These studies revealed that SRIF is involved in the acquisition of information but other studies showed that cysteamine produced memory deficits not only when given before the training session but also within a critical time window (0–4 h) after acquisition, suggesting that SRIF plays a critical role in memory consolidation proces-sing (Haroutunian et al., 1987, 1989; Schettini et al., 1988; Vecsei et al., 1990).
The hippocampus is an essential structure in learning and memory (Jeneson and Squire, 2012), and is also a chosen site to study the effects of SRIF on learning and memory since injection of cysteamine impairs tasks requiring its integrity (DeNoble et al., 1989; Guillou et al., 1998). Surprisingly in the rodent hippocampus, both activation of SRIF receptors as well as depletion of SRIF contents generate hippocampal memory impairments. Indeed, microinjections of cysteamine, SRIF or CST directly into the hippocampus impaired hippocampal-dependent spatial learning (Guillou et al., 1993; Sanchez-Alavez et al., 2000; Lamirault et al., 2001; Mendez-Diaz et al., 2005; Gastambide et al., 2009). Consistent with these pharmacological results, transgenic mice overexpressing CST display a profound impairment of spatial learning (Tallent et al., 2005). Studies that investigated which SRIF receptor mediates SRIF memory effect showed that intra-hippocampal injections of the sst4 agonist, but not sst1, sst2, or sst3 agonists, dramatically impaired spatial memory formation (Gastambide et al., 2009). Importantly, these authors found that concomitantly to the impairment of spatial memory, an sst4 agonist also enhanced the use of striatum-dependent memory. Therefore, it was hypothesized that hippocampal sst4 controls the use of cognitive strategies by switching from hippocampus-based multiple associations to simple striatum-based behavioral response through a functional interaction with sst2 receptor (Gastambide et al., 2010). The precise cellular and molecular mechanisms involved in this functional interaction between sst2 and sst4 are not fully understood but some studies showed that sst4 mediates increases in glutamatergic excitability and bursting frequency, which were blocked by sst2 agonists or antagonists and were lacking in sst2 knockout (KO) mice (Moneta et al., 2002; Cammalleri et al., 2006). Therefore, sst4 is not the unique SRIF receptor in the hippocampus mediating SRIF memory effects as sst2 also modulates memory as previously suggested by Dutar et al. (2002).
Involvement of SRIF in the Control of Emotion
Somatostatin and its receptors are strongly expressed in the different nuclei of the amygdala (Hannon et al., 2002), a key brain structure involved in the emotional assessment of the environment (Schumann et al., 2011). Despite the extensive expression of SRIF systems in this area, the effects of SRIF on emotions have not yet been studied extensively. Nevertheless, some studies reported an involvement of SRIF systems in the control of emotion and anxiety. Indeed, a very recent work revealed that the pattern of activation of SRIF-positive interneurons was specific to the nuclei of the amygdala considered and also to the kind of stressor used (Butler et al., 2012). Moreover, SRIF has anxiolytic- and antidepressant-like effects (Engin et al., 2008b) that are associated with the suppression of the frequency of hippocampal theta activity, a neurophysiological signature common to most classes of anxiolytic drugs (i.e., benzodiazepines, selective 5-HT reuptake inhibitors, 5-HT1A agonists). These effects seem to be mediated by sst2 receptor since both intra-septal and intra-amygdala SRIF microinfusions induced anxiolytic effects that were completely reversed by selective sst2 receptor antagonist injection in these brain areas (Yeung and Treit, 2012). Additional evidence for a specific role of sst2 receptor came from the observation that a stressful experience is associated with an increase of sst2 mRNA levels within the amygdala (Nanda et al., 2008) and that mice lacking sst2 receptor display increased anxiety-like behaviors associated with increased pituitary ACTH levels, a main regulator of the stress response (Viollet et al., 2000).
Involvement of SRIF in Locomotion
An involvement for SRIF was also reported in motor functions. Increased motor activity was shown in rats receiving intracerebroventricular administration of SRIF (Havlicek et al., 1976) as well as in mice receiving unilateral striatal infusions of the peptide by retrodialysis (Hathway et al., 2004) and in animals receiving direct injections of SRIF in the nucleus accumbens (Raynor et al., 1993). Tashev et al. (2001) showed that SRIF modulated locomotor activity in biphasic manner. Indeed, shortly after SRIF striatal injection a decrease of locomotor activity is observed whereas later, the locomotor behavior is increased. Similar effects have been found after striatal injection of sst2 and sst4 agonists. On the other hand, genetic invalidation of sst2 receptor in two different strains of mice as well as SRIF null mice showed an impairment of motor functions (Viollet et al., 2000; Zeyda et al., 2001; Allen et al., 2003). But the role of SRIF in locomotion seems to be limited to fine motor control since these different lines of transgenic mice only develop impaired motor coordination in tasks that require a fine motor control and display normal levels of motor activity and coordination in undemanding tasks (Viollet et al., 2000; Zeyda et al., 2001; Allen et al., 2003).
Autonomic Responses
Somatotropin release inhibiting factor and its receptors are found in several medulla oblongata nuclei that control autonomic functions such as digestion, cardiac rate, and respiration (Llona and Eugenín, 2005; Spary et al., 2008; Viollet et al., 2008). In the preBötzinger complex (preBötC), a critical component of the respiratory rhythm generator that underlies mammalian breathing, SRIF is expressed in a subpopulation of glutamatergic neurokinin 1 receptor-positive neurons, a kind of neuron rhythmically active (Stornetta et al., 2003). Originating from the homeogene Dbx1 lineage, these cells are mandatory for breathing, since invalidation of the Dbx1 gene impaired their differentiation and disrupted respiratory rhythm generation in the preBötC (Bouvier et al., 2010; Gray et al., 2010). Acute silencing of somatostatinergic preBötC neurons increased respiratory rhythm, leading to persistent apnea (Tan et al., 2008). Similar effects were found in vitro after pharmacological blockade of sst2 transmission, while exogenous SRIF application decreased rhythms generation (Pantaleo et al., 2011; Ramírez-Jarquín et al., 2012). This demonstrated that the peptide exerts a tonic inhibitory control on the rythmogenic neurons in order to avoid deleterious overactivity, probably through cellular subdomain-specific inhibitory and excitatory synaptic contacts (Wei et al., 2012). The existence of long-range somatostatinergic projections to either contralateral PreBötC (Stornetta et al., 2003) or downstream premotor neurons (Tan et al., 2010) favors a neuromodulatory role for PreBötC SRIF (Llona and Eugenín, 2005), whose developmental impairment may be involved in human pathologies (Schwarzacher et al., 2011) such as the sudden infant death syndrome (Lavezzi and Matturri, 2008).
Somatostatinergic Networks in Pathological Conditions
In animals, an alteration of SRIF systems is observed during normal aging (Stanley et al., 2012) and pathological models of aging. In human a similar specific dysregulation is observed in normal pathological disorders such as some neurodegenerative and psychiatric diseases (Glorioso et al., 2011; Gleichmann et al., 2012).
Alzheimer’s Disease
Somatostatin has been involved in Alzheimer’s disease (AD) pathology for a number of years. Indeed, since the early 1980s, it is known that SRIF levels in cortex and hippocampus are decreased in AD patients (Davies et al., 1980). Later, it was demonstrated that the decline in SRIF concentrations in the CSF (Tamminga et al., 1987) or in the middle frontal gyrus (Dournaud et al., 1995) correlates with cognitive deficits. Using quantitative real-time PCR, a recent study confirmed this decrease of SRIF in the inferior, medial, and superior temporal lobe of AD patients (Gahete et al., 2010). Interestingly, SRIF concentrations were reported to be significantly lower in Alzheimer patients carrying the epsilon 4 allele of APOE (Grouselle et al., 1998), the main genetic risk factor described to date for late-onset AD (Genin et al., 2011). In addition, two different studies found in Finnish and Chinese patients that polymorphisms in the SST gene are associated with the risk of developing AD (Vepsalainen et al., 2007; Xue et al., 2009).
Regarding SRIF receptors, data are limited and controversial. Although all studies agreed on a decrease of SRIF receptors in AD, controversies appeared about the proportion, the localization, and receptor subtype specificity of this decrease. SRIF receptor quantification using quantitative real-time PCR in AD temporal lobe showed a decrease of sst1, sst3, and sst4 receptors whereas sst2 and sst5 receptors were unchanged (Gahete et al., 2010). Previously, an immunohistochemistry study reported a similar decrease of sst4 but showed a reduction in neuronal sst5 – and a modest decrease in sst2 –like immunoreactivity without any changes in sst1 immunoreactive neurons (Kumar, 2005). Surprisingly, in the same study, an increase of sst3 subtype was observed in AD cortex. A radioligand binding and functional study showed a general receptor decrease in AD brain (Beal et al., 1985). More specifically, receptors levels in the frontal and temporal cortex were reduced by approximately 50% of control values in AD patients while a 40% reduction was reported in the hippocampus and no significant changes were found in the cingulate cortex, postcentral gyrus, temporal pole, and superior temporal gyrus. Another radioligand binding study revealed that while the maximal binding capacity of the SRIF-1 receptor subtype (primarily sst2, and possibly sst5) is altered in frontal and temporal cortices, other putative cortical SRIF receptor classes (SRIF-2 sites, i.e., sst1 and sst4) are not as broadly affected (Krantic et al., 1992). Finally, a last study showed a significant decrease only in the frontal cortex, but not in other brain regions (Bergstrom et al., 1991). Because of the cholinergic hypothesis regarding AD etiology, it was concluded that the pattern of change of SRIF binding in AD cortex might be secondary to the degeneration of SRIF receptor-bearing cholinergic afferents arising from the nucleus basalis. In line with this idea, experiments in the literature demonstrate that the selective destruction of cholinergic neurons of the basal forebrain with intracerebroventricular injection of 192-IgG saporin produces an irreversible loss of SRIF-immunoreactive neurons in the hilus of the hippocampus (Jolkkonen et al., 1997) and in the cortex (Zhang et al., 1998). This last study shows a correlation between the intensity of acetylcholinesterase in the cortex and the number of remaining SRIF cells. These data highlight a trophic dependence of SRIF neurons on cholinergic inputs and are consistent with observations in AD and aging.
Although SRIF deficit is not correlated with the amyloid load in AD brain patients (Dournaud et al., 1995), SRIF was identified as a modulator that increases brain neprilysin activity, one of the main enzymes involved in Aβ degradation (Saito et al., 2005). Recently, it has been shown that neuropeptide pituitary adenylate cyclase-activating polypeptide slows down AD-like pathology and improves cognition in a transgenic mouse model of AD through the activation of SRIF-neprilysin cascade (Rat et al., 2011). In mouse primary embryonic neurons, SRIF concomitantly increased neprilysin activity and decreased Aβ42 in the culture medium and these effects were blocked by an sst5 antagonist (but also an agonist at sst1 and sst3 receptors; for review, see Epelbaum et al., 2009). Moreover, neprilysin activity was decreased by 50% and Aβ42 increased by a similar extent in SRIF KO mice (Saito et al., 2005). Such findings may have important implications for understanding the cellular mechanisms leading to AD and suggest that SRIF and its receptors are potential pharmacological targets for AD. Indeed, FK962, which promotes SRIF production in the brain, co-administrated with donepezil, an acetylcholinesterase inhibitor widely used to treat patients, enhances cognition in rat and has been proposed as an add-on therapy for AD (McCarthy et al., 2011). In addition, Rubio et al. (2012) recently suggested that SRIF and CST act as a protective agent against Aβ toxicity. However, in APP transgenic mouse models, data concerning SRIF-containing interneurons are contradictory. In the triple-transgenic model of AD, 3×Tg-AD, inhibitory neurotransmission is unchanged in the cerebral cortex and hippocampus (Gleichmann et al., 2012). In a APP/PS1 mouse model of AD, as soon as 6 months of age, a decrease in the number of oriens-lacunosum moleculare hilar perforant path-associated SRIF-positive interneurons was evidenced in the hippocampus, when no change was demonstrated for 21 additional mRNA markers tested (Ramos et al., 2006). In the APPswe/PS1dE9 mouse model, Aβ deposition disrupted cognitive circuits when the cholinergic and somatostatinergic systems remained relatively intact (Savonenko et al., 2005). Another study on this last model even found that, in most brain regions tested, SRIF concentrations were increased rather than decreased relative to controls (Horgan et al., 2007). Thus, the validity of a direct and major role for SRIF in the regulation of Aβ42 degradation remains to be further confirmed (Iwata et al., 2005). More recent studies, focusing on olfaction, an early-altered function in AD (Wilson et al., 2009), account for evidence of a relationship between Aβ pathology and SRIF alterations in the disease. Indeed, SRIF interneurons and receptors are selectively reduced by approximately 50% in the anterior olfactory nucleus of AD patients (Saiz-Sanchez et al., 2010). These authors suggested that SRIF decreases in AD might be linked with Aβ. Moreover, an increase in the levels of aggregated Aβ peptide is observed with aging in olfactory cortices of APP/PS1 transgenic mouse model of AD, and it is accompanied by a fall in numbers of SRIF-positive interneurons (Saiz-Sanchez et al., 2012).
Experiments from our laboratory demonstrated that intrahippocampal injections of Aβ in rats induced aberrant inhibitory septo-hippocampal network activity associated with an impairment of hippocampal memory processes (Villette et al., 2010). This effect can be explained by the selective loss of long-range hippocampo-septal projecting neurons population containing calbindin and SRIF (Villette et al., 2012). This population of SRIF neurons could be a favored target for Aβ, explaining the early decrease of SRIF observed in AD.
Somatotropin release inhibiting factor is not only interacting with Aβ42 in AD, it has also an effect on Tau phosphorylation. Rubio et al. (2008) indicated that in mouse cortex SRIF and CST induce Tau phosphorylation at Ser262, a site modified in AD (Wang et al., 2007), although with different kinetics. An sst2/sst4 interaction seems implicated in this process but the types of phosphatases that are involved remain to be determined. Moreover, in human apoE4 knock-in mice where Tau phosphorylation and intracellular neurofibrillary tangle-like deposits are detected (Huang et al., 2001; Harris et al., 2003; Brecht et al., 2004), Huang’s group showed that the number of SRIF-positive interneurons correlated inversely with the performance of these mice in a spatial memory task (Andrews-Zwilling et al., 2010).
Parkinson’s Disease
Alteration of SRIF levels is also observed in other neurodegenerative diseases. Indeed, decrease in SRIF levels has been described in demented Parkinson’s disease patients (Epelbaum et al., 1989) as well as in a unilateral 6-OHDA experimental mouse model of Parkinson’s disease (Nilsson et al., 2009). Recent data obtained in a rat model of Parkinsonism showed that an alteration of presynaptic modulation by SRIF after dopamine deprivation. This observation may underlie a homeostatic mechanism trying to compensate for the excitability imbalance between direct and indirect basal ganglia pathways found during Parkinson’s disease (Lopez-Huerta et al., 2012).
Major Depressive Disorder
Evidence in major depressive disorder (MDD) suggests an impaired excitation/inhibition balance that is potentially mediated by decreased GABA content (Levinson et al., 2010). More specifically, Sibille et al. (2011) reported a down-regulation of SRIF in the dorsolateral prefrontal cortex (PFC), the subgenual cingulate cortices (Tripp et al., 2011), and the amygdala (Guilloux et al., 2011) of MDD patients. Engin et al. (2008a) and Engin and Treit (2009) revealed an antidepressant effect of SRIF mediated by either sst2 or sst3 receptor and suggested that while SRIF itself is not appropriate for clinical use because of its short half-life and diverse range of effects (Pinter et al., 2006), a closely related SRIF derivative may have some potential for the pharmacological treatment of depression.
Schizophrenia
One of the most consistent findings in schizophrenia neuropathology is deficits in cortical inhibitory interneurons across multiple cortical regions (Hashimoto et al., 2008). It has been known for years that cerebral cortical concentrations of SRIF are reduced in schizophrenics (Roberts et al., 1983) as well as hippocampal concentration (Ferrier et al., 1983; Konradi et al., 2011). Moreover, Hashimoto et al. (2008) found that subjects with schizophrenia exhibited deficits in SRIF expression in the PFC, and this was further confirmed after global analysis from six previously published microarray studies (Perez-Santiago et al., 2012). A recent study suggested that this decrease of SRIF-positive inhibitory interneurons in the PFC may be related to changes in an inflammatory response pathway that are often observed in schizophrenics (Fillman et al., 2012). In addition, Beneyto et al. (2012) showed that SRIF neurotransmission in the PFC of subjects with schizophrenia is also altered at the postsynaptic level in a receptor subtype-, layer-, and cell type-specific manner. The expression of sst2, but not sst1, mRNA is preferentially lower in layers 5–6, and in larger, putative pyramidal neurons in those layers. These authors suggested converging pre- and postsynaptic mechanisms to reduce inhibitory neurotransmission in pyramidal neurons in the PFC, which could alter the synchronization of low frequency oscillations and disturb working memory performance in subjects with schizophrenia.
Epilepsy
Somatostatin is highly expressed in brain regions associated with seizures and has been implicated as playing a prominent role in epilepsy (Vezzani and Hoyer, 1999) based on the observation of an activity-dependent release of SRIF during seizures, the modulation of SRIF mRNA expression, peptide and receptors levels by seizures and the effect of SRIF and its analogs on seizures (Tallent and Qiu, 2008; Zafar et al., 2012). Temporal lobe epilepsy (TLE) is characterized by hippocampal sclerosis together with profound phenotypic changes of different classes of interneurons. Hilar SRIF interneurons undergo extensive degeneration in patients with hippocampal sclerosis (de Lanerolle et al., 1989; Robbins et al., 1991). Recently, this selective neurodegeneration has been linked to the specific enrichment of somatostatinergic neurons in striatum-enriched phosphatase, an enzyme that counteracts the MAPK neuroprotective pathway (Choi et al., 2007; Florio et al., 2008). SRIF receptors may represent potential therapeutic targets for TLE. Indeed, SRIF is released in characteristic conditions of seizures and SRIF and its analogs affect seizures (Vezzani and Hoyer, 1999; Buckmaster et al., 2002). However, information on the precise contribution of each SRIF receptor on the SRIF-induced inhibition of epileptiform activity is still limited. Although the sst2 receptor is likely to mediate the anticonvulsant effects of SRIF in rat hippocampus (Vezzani and Hoyer, 1999), observations in the mouse support a central role for sst4 (Moneta et al., 2002) and/or sst1 receptors (Cammalleri et al., 2004, 2006) in mediating SRIF inhibition of epileptiform activity. In a rodent model of cortical focal ischemia, sst2 is also activated while the infarct size is significantly reduced in sst2 KO mice (Stumm et al., 2004). However, recent data in rats showed that sst1 receptors do not appear to mediate the in vivo anticonvulsive effect of SRIF (de Bundel et al., 2010), whereas sst3 and sst4 mediate this effect through a functional interaction with sst2 receptor (Aourz et al., 2011).
Conclusion
Somatostatin systems are widely expressed in the different brain regions and are involved in numerous processes from sensory to cognitive functions, suggesting that they play major roles in brain functioning. These key roles are illustrated by the decrease of SRIF concentrations observed in neurodegenerative diseases such as AD and Parkinson’s disease but also in psychiatric diseases such as schizophrenia and MDD. From this perspective, SRIF systems represent a potential and challenging therapeutic target. Further studies need to be carried on to unravel the role of SRIF systems in all functions they have been implicated in.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
Supported by NeRF post-doctoral fellowship (to Guillaume Martel), ANR-10-MALZ-003-01 SOMADOLF and Fondation Recherche Plan Alzheimer.
References
Allen, J. P., Hathway, G. J., Clarke, N. J., Jowett, M. I., Topps, S., Kendrick, K. M., et al. (2003). Somatostatin receptor 2 knockout/lacZ knockin mice show impaired motor coordination and reveal sites of somatostatin action within the striatum. Eur. J. Neurosci. 17, 1881–1895.
Andrews-Zwilling, Y., Bien-Ly, N., Xu, Q., Li, G., Bernardo, A., Yoon, S. Y., et al. (2010). Apolipoprotein E4 causes age- and Tau-dependent impairment of GABAergic interneurons, leading to learning and memory deficits in mice. J. Neurosci. 30, 13707–13717.
Aourz, N., De Bundel, D., Stragier, B., Clinckers, R., Portelli, J., Michotte, Y., et al. (2011). Rat hippocampal somatostatin sst3 and sst4 receptors mediate anticonvulsive effects in vivo: indications of functional interactions with sst2 receptors. Neuropharmacology 61, 1327–1333.
Bakhit, C., and Swerdlow, N. (1986). Behavioral changes following central injection of cysteamine in rats. Brain Res. 365, 159–163.
Baraban, S. C., and Tallent, M. K. (2004). Interneuron Diversity Series: interneuronal neuropeptides – endogenous regulators of neuronal excitability. Trends Neurosci. 27, 135–142.
Baratta, M. V., Lamp, T., and Tallent, M. K. (2002). Somatostatin depresses long-term potentiation and Ca2+ signaling in mouse dentate gyrus. J. Neurophysiol. 88, 3078–3086.
Beal, M. F., Mazurek, M. F., Tran, V. T., Chattha, G., Bird, E. D., and Martin, J. B. (1985). Reduced numbers of somatostatin receptors in the cerebral cortex in Alzheimer’s disease. Science 229, 289–291.
Beneyto, M., Morris, H. M., Rovensky, K. C., and Lewis, D. A. (2012). Lamina- and cell-specific alterations in cortical somatostatin receptor 2 mRNA expression in schizophrenia. Neuropharmacology 62, 1598–1605.
Bergstrom, L., Garlind, A., Nilsson, L., Alafuzoff, I., Fowler, C. J., Winblad, B., et al. (1991). Regional distribution of somatostatin receptor binding and modulation of adenylyl cyclase activity in Alzheimer’s disease brain. J. Neurol. Sci. 105, 225–233.
Boehm, S., and Betz, H. (1997). Somatostatin inhibits excitatory transmission at rat hippocampal synapses via presynaptic receptors. J. Neurosci. 17, 4066–4075.
Bollok, I., Vecsei, L., and Telegdy, G. (1983). The effects of interaction between propranolol and somatostatin on the active avoidance behavior, open-field activity and electroconvulsive shock-induced amnesia of rats. Neuropeptides 3, 263–270.
Bousquet, C., Lasfargues, C., Chalabi, M., Billah, S. M., Susini, C., Vezzosi, D., et al. (2012). Clinical review: current scientific rationale for the use of somatostatin analogs and mTOR inhibitors in neuroendocrine tumor therapy. J. Clin. Endocrinol. Metab. 97, 727–737.
Bouvier, J., Thoby-Brisson, M., Renier, N., Dubreuil, V., Ericson, J., Champagnat, J., et al. (2010). Hindbrain interneurons and axon guidance signaling critical for breathing. Nat. Neurosci. 13, 1066–1074.
Brazeau, P., Vale, W., Burgus, R., Ling, N., Butcher, M., Rivier, J., et al. (1973). Hypothalamic polypeptide that inhibits the secretion of immunoreactive pituitary growth hormone. Science 179, 77–79.
Brecht, W. J., Harris, F. M., Chang, S., Tesseur, I., Yu, G. Q., Xu, Q., et al. (2004). Neuron-specific apolipoprotein e4 proteolysis is associated with increased tau phosphorylation in brains of transgenic mice. J. Neurosci. 24, 2527–2534.
Brunjes, P. C., Kay, R. B., and Arrivillaga, J. P. (2011). The mouse olfactory peduncle. J. Comp. Neurol. 519, 2870–2886.
Buckmaster, P. S., Otero-Corchon, V., Rubinstein, M., and Low, M. J. (2002). Heightened seizure severity in somatostatin knockout mice. Epilepsy Res. 48, 43–56.
Butler, R. K., White, L. C., Frederick-Duus, D., Kaigler, K. F., Fadel, J. R., and Wilson, M. A. (2012). Comparison of the activation of somatostatin- and neuropeptide Y-containing neuronal populations of the rat amygdala following two different anxiogenic stressors. Exp. Neurol. 238, 52–63.
Cammalleri, M., Cervia, D., Dal Monte, M., Martini, D., Langenegger, D., Fehlmann, D., et al. (2006). Compensatory changes in the hippocampus of somatostatin knockout mice: upregulation of somatostatin receptor 2 and its function in the control of bursting activity and synaptic transmission. Eur. J. Neurosci. 23, 2404–2422.
Cammalleri, M., Cervia, D., Langenegger, D., Liu, Y., Dal Monte, M., Hoyer, D., et al. (2004). Somatostatin receptors differentially affect spontaneous epileptiform activity in mouse hippocampal slices. Eur. J. Neurosci. 20, 2711–2721.
Cammalleri, M., Martini, D., Timperio, A. M., and Bagnoli, P. (2009). Functional effects of somatostatin receptor 1 activation on synaptic transmission in the mouse hippocampus. J. Neurochem. 111, 1466–1477.
Casini, G., Catalani, E., Dal Monte, M., and Bagnoli, P. (2005). Functional aspects of the somatostatinergic system in the retina and the potential therapeutic role of somatostatin in retinal disease. Histol. Histopathol. 20, 615–632.
Cervia, D., and Bagnoli, P. (2007). An update on somatostatin receptor signaling in native systems and new insights on their pathophysiology. Pharmacol. Ther. 116, 322–341.
Cervia, D., Casini, G., and Bagnoli, P. (2008). Physiology and pathology of somatostatin in the mammalian retina: a current view. Mol. Cell. Endocrinol. 286, 112–122.
Choi, Y. S., Lin, S. L., Lee, B., Kurup, P., Cho, H. Y., Naegele, J. R., et al. (2007). Status epilepticus-induced somatostatinergic hilar interneuron degeneration is regulated by striatal enriched protein tyrosine phosphatase. J. Neurosci. 27, 2999–3009.
Córdoba-Chacón, J., Gahete, M. D., Durán-Prado, M., Luque, R. M., and Castaño, J. P. (2011). Truncated somatostatin receptors as new players in somatostatin-cortistatin pathophysiology. Ann. N. Y. Acad. Sci. 1220, 6–15.
Cossart, R., Dinocourt, C., Hirsch, J. C., Merchan-Perez, A., De Felipe, J., Ben-Ari, Y., et al. (2001). Dendritic but not somatic GABAergic inhibition is decreased in experimental epilepsy. Nat. Neurosci. 4, 52–62.
Csaba, Z., Peineau, S., and Dournaud, P. (2012). Molecular mechanisms of somatostatin receptor trafficking. J. Mol. Endocrinol. 48, R1–R12.
Dal Monte, M., Cammalleri, M., Martini, D., Casini, G., and Bagnoli, P. (2007). Antiangiogenic role of somatostatin receptor 2 in a model of hypoxia-induced neovascularization in the retina: results from transgenic mice. Invest. Ophthalmol. Vis. Sci. 48, 3480–3489.
Dal Monte, M., Ristori, C., Cammalleri, M., and Bagnoli, P. (2009). Effects of somatostatin analogues on retinal angiogenesis in a mouse model of oxygen-induced retinopathy: involvement of the somatostatin receptor subtype 2. Invest. Ophthalmol. Vis. Sci. 50, 3596–3606.
Davies, P., Katzman, R., and Terry, R. D. (1980). Reduced somatostatin-like immunoreactivity in cerebral cortex from cases of Alzheimer disease and Alzheimer senile dementa. Nature 288, 279–280.
de Bundel, D., Aourz, N., Kiagiadaki, F., Clinckers, R., Hoyer, D., Kastellakis, A., et al. (2010). Hippocampal sst(1) receptors are autoreceptors and do not affect seizures in rats. Neuroreport 21, 254–258.
de Lanerolle, N. C., Kim, J. H., Robbins, R. J., and Spencer, D. D. (1989). Hippocampal interneuron loss and plasticity in human temporal lobe epilepsy. Brain Res. 495, 387–395.
de Lecea, L. (2008). Cortistatin – functions in the central nervous system. Mol. Cell. Endocrinol. 286, 88–95.
De Lecea, L., Criado, J. R., Prospero-Garcia, O., Gautvik, K. M., Schweitzer, P., Danielson, P. E., et al. (1996). A cortical neuropeptide with neuronal depressant and sleep-modulating properties. Nature 381, 242–245.
DeNoble, V. J., Hepler, D. J., and Barto, R. A. (1989). Cysteamine-induced depletion of somatostatin produces differential cognitive deficits in rats. Brain Res. 482, 42–48.
Dournaud, P., Delaere, P., Hauw, J. J., and Epelbaum, J. (1995). Differential correlation between neurochemical deficits, neuropathology, and cognitive status in Alzheimer’s disease. Neurobiol. Aging 16, 817–823.
Dutar, P., Vaillend, C., Viollet, C., Billard, J. M., Potier, B., Carlo, A. S., et al. (2002). Spatial learning and synaptic hippocampal plasticity in type 2 somatostatin receptor knock-out mice. Neuroscience 112, 455–466.
Einstein, E. B., Patterson, C. A., Hon, B. J., Regan, K. A., Reddi, J., Melnikoff, D. E., et al. (2010). Somatostatin signaling in neuronal cilia is critical for object recognition memory. J. Neurosci. 30, 4306–4314.
Engin, E., Stellbrink, J., Treit, D., and Dickson, C. (2008a). Anxiolytic and antidepressant effects of intracerebroventricularly administered somatostatin: behavioral and neurophysiological evidence. Neuroscience 157, 666–676.
Engin, E., Stellbrink, J., Treit, D., and Dickson, C. T. (2008b). Anxiolytic and antidepressant effects of intracerebroventricularly administered somatostatin: behavioral and neurophysiological evidence. Neuroscience 157, 666–676.
Engin, E., and Treit, D. (2009). Anxiolytic and antidepressant actions of somatostatin: the role of sst2 and sst3 receptors. Psychopharmacology (Berl.) 206, 281–289.
Epelbaum, J. (1986). Somatostatin in the central nervous system: physiology and pathological modifications. Prog. Neurobiol. 27, 63–100.
Epelbaum, J., Agid, F., Agid, Y., Beaudet, A., Bertrand, P., Enjalbert, A., et al. (1989). Somatostatin receptors in brain and pituitary. Horm. Res. 31, 45–50.
Epelbaum, J., Guillou, J. L., Gastambide, F., Hoyer, D., Duron, E., and Viollet, C. (2009). Somatostatin, Alzheimer’s disease and cognition: an old story coming of age? Prog. Neurobiol. 89, 153–161.
Eyre, M. D., Kerti, K., and Nusser, Z. (2009). Molecular diversity of deep short-axon cells of the rat main olfactory bulb. Eur. J. Neurosci. 29, 1397–1407.
Ferrier, I. N., Roberts, G. W., Crow, T. J., Johnstone, E. C., Owens, D. G., Lee, Y. C., et al. (1983). Reduced cholecystokinin-like and somatostatin-like immunoreactivity in limbic lobe is associated with negative symptoms in schizophrenia. Life Sci. 33, 475–482.
Fillman, S. G., Cloonan, N., Catts, V. S., Miller, L. C., Wong, J., Mccrossin, T., et al. (2012). Increased inflammatory markers identified in the dorsolateral prefrontal cortex of individuals with schizophrenia. Mol. Psychiatry. doi: 10.1038/mp.2012.110 [Epub ahead of print].
Florio, P., Abella, R., Marinoni, E., Di Iorio, R., Letizia, C., Meli, M., et al. (2008). Adrenomedullin blood concentrations in infants subjected to cardiopulmonary bypass: correlation with monitoring parameters and prediction of poor neurological outcome. Clin. Chem. 54, 202–206.
Gahete, M. D., Rubio, A., Duran-Prado, M., Avila, J., Luque, R. M., and Castano, J. P. (2010). Expression of Somatostatin, cortistatin, and their receptors, as well as dopamine receptors, but not of neprilysin, are reduced in the temporal lobe of Alzheimer’s disease patients. J. Alzheimers Dis. 20, 465–475.
Galarraga, E., Vilchis, C., Tkatch, T., Salgado, H., Tecuapetla, F., Perez-Rosello, T., et al. (2007). Somatostatinergic modulation of firing pattern and calcium-activated potassium currents in medium spiny neostriatal neurons. Neuroscience 146, 537–554.
Gastambide, F., Lepousez, G., Viollet, C., Loudes, C., Epelbaum, J., and Guillou, J. L. (2010). Cooperation between hippocampal somatostatin receptor subtypes 4 and 2: functional relevance in interactive memory systems. Hippocampus 20, 745–757.
Gastambide, F., Viollet, C., Lepousez, G., Epelbaum, J., and Guillou, J. L. (2009). Hippocampal SSTR4 somatostatin receptors control the selection of memory strategies. Psychopharmacology (Berl.) 202, 153–163.
Genin, E., Hannequin, D., Wallon, D., Sleegers, K., Hiltunen, M., Combarros, O., et al. (2011). APOE and Alzheimer disease: a major gene with semi-dominant inheritance. Mol. Psychiatry 16, 903–907.
Gentet, L. J., Kremer, Y., Taniguchi, H., Huang, Z. J., Staiger, J. F., and Petersen, C. C. (2012). Unique functional properties of somatostatin-expressing GABAergic neurons in mouse barrel cortex. Nat. Neurosci. 15, 607–612.
Gleichmann, M., Zhang, Y., Wood, W. H. III, Becker, K. G., Mughal, M. R., Pazin, M. J., et al. (2012). Molecular changes in brain aging and Alzheimer’s disease are mirrored in experimentally silenced cortical neuron networks. Neurobiol. Aging 33, 205.e1–205.e18.
Glorioso, C., Oh, S., Douillard, G. G., and Sibille, E. (2011). Brain molecular aging, promotion of neurological disease and modulation by sirtuin 5 longevity gene polymorphism. Neurobiol. Dis. 41, 279–290.
Goldberg, J. H., Lacefield, C. O., and Yuste, R. (2004). Global dendritic calcium spikes in mouse layer 5 low threshold spiking interneurones: implications for control of pyramidal cell bursting. J. Physiol. 558, 465–478.
Goldin, M., Epsztein, J., Jorquera, I., Represa, A., Ben-Ari, Y., Crepel, V., et al. (2007). Synaptic kainate receptors tune oriens-lacunosum moleculare interneurons to operate at theta frequency. J. Neurosci. 27, 9560–9572.
Gonchar, Y., Wang, Q., and Burkhalter, A. (2007). Multiple distinct subtypes of GABAergic neurons in mouse visual cortex identified by triple immunostaining. Front. Neuroanat. 1, 1–11.
Gray, P. A., Hayes, J. A., Ling, G. Y., Llona, I., Tupal, S., Picardo, M. C. D., et al. (2010). Developmental origin of preBötzinger complex respiratory neurons. J. Neurosci. 30, 14883–14895.
Grilli, M., Raiteri, L., and Pittaluga, A. (2004). Somatostatin inhibits glutamate release from mouse cerebrocortical nerve endings through presynaptic sst2 receptors linked to the adenylyl cyclase-protein kinase A pathway. Neuropharmacology 46, 388–396.
Grouselle, D., Winsky-Sommerer, R., David, J. P., Delacourte, A., Dournaud, P., and Epelbaum, J. (1998). Loss of somatostatin-like immunoreactivity in the frontal cortex of Alzheimer patients carrying the apolipoprotein epsilon 4 allele. Neurosci. Lett. 255, 21–24.
Guillou, J. L., Micheau, J., and Jaffard, R. (1993). Effects of intrahippocampal injections of somatostatin and cysteamine on spatial discrimination learning in mice. Psychobiology 21, 265–271.
Guillou, J. L., Micheau, J., and Jaffard, R. (1998). The opposite effects of cysteamine on the acquisition of two different tasks in mice are associated with bidirectional testing-induced changes in hippocampal adenylyl cyclase activity. Behav. Neurosci. 112, 900–908.
Guilloux, J. P., Douillard-Guilloux, G., Kota, R., Wang, X., Gardier, A. M., Martinowich, K., et al. (2011). Molecular evidence for BDNF- and GABA-related dysfunctions in the amygdala of female subjects with major depression. Mol. Psychiatry 17, 1130–1142.
Hannon, J. P., Petrucci, C., Fehlmann, D., Viollet, C., Epelbaum, J., and Hoyer, D. (2002). Somatostatin sst2 receptor knock-out mice: localisation of sst1-5 receptor mRNA and binding in mouse brain by semi-quantitative RT-PCR, in situ hybridisation histochemistry and receptor autoradiography. Neuropharmacology 42, 396–413.
Haroutunian, V., Kanof, P. D., and Davis, K. L. (1989). Interactions of forebrain cholinergic and somatostatinergic systems in the rat. Brain Res. 496, 98–104.
Haroutunian, V., Mantin, R., Campbell, G. A., Tsuboyama, G. K., and Davis, K. L. (1987). Cysteamine-induced depletion of central somatostatin-like immunoactivity: effects on behavior, learning, memory and brain neurochemistry. Brain Res. 403, 234–242.
Harris, F. M., Brecht, W. J., Xu, Q., Tesseur, I., Kekonius, L., Wyss-Coray, T., et al. (2003). Carboxyl-terminal-truncated apolipoprotein E4 causes Alzheimer’s disease-like neurodegeneration and behavioral deficits in transgenic mice. Proc. Natl. Acad. Sci. U.S.A. 100, 10966–10971.
Hashimoto, T., Arion, D., Unger, T., Maldonado-Aviles, J. G., Morris, H. M., Volk, D. W., et al. (2008). Alterations in GABA-related transcriptome in the dorsolateral prefrontal cortex of subjects with schizophrenia. Mol. Psychiatry 13, 147–161.
Hathway, G. J., Humphrey, P. P., and Kendrick, K. M. (2004). Somatostatin induces striatal dopamine release and contralateral turning behaviour in the mouse. Neurosci. Lett. 358, 127–131.
Havlicek, V., Rezek, M., and Friesen, H. (1976). Somatostatin and thyrotropin releasing hormone: central effect on sleep and motor system. Pharmacol. Biochem. Behav. 4, 455–459.
Horgan, J., Miguel-Hidalgo, J. J., Thrasher, M., and Bissette, G. (2007). Longitudinal brain corticotropin releasing factor and somatostatin in a transgenic mouse (TG2576) model of Alzheimer’s disease. J. Alzheimers Dis. 12, 115–127.
Hu, H., Ma, Y., and Agmon, A. (2011). Submillisecond firing synchrony between different subtypes of cortical interneurons connected chemically but not electrically. J. Neurosci. 31, 3351–3361.
Huang, Y., Liu, X. Q., Wyss-Coray, T., Brecht, W. J., Sanan, D. A., and Mahley, R. W. (2001). Apolipoprotein E fragments present in Alzheimer’s disease brains induce neurofibrillary tangle-like intracellular inclusions in neurons. Proc. Natl. Acad. Sci. U.S.A. 98, 8838–8843.
Hwang, I. K., Lee, J. C., Park, S.-K., An, S.-J., Lee, H. Y., Lee, Y.-B., et al. (2004). Age-related change of somatostatin-immunoreactive neurones in the main olfactory bulb of the rat. Anat. Histol. Embryol. 33, 59–63.
Ishibashi, H., and Akaike, N. (1995). Somatostatin modulates high-voltage-activated Ca2+ channels in freshly dissociated rat hippocampal neurons. J. Neurophysiol. 74, 1028–1036.
Iwata, N., Higuchi, M., and Saido, T. C. (2005). Metabolism of amyloid-beta peptide and Alzheimer’s disease. Pharmacol. Ther. 108, 129–148.
Jeneson, A., and Squire, L. R. (2012). Working memory, long-term memory, and medial temporal lobe function. Learn. Mem. 19, 15–25.
Jiang, N., Furue, H., Katafuchi, T., and Yoshimura, M. (2003). Somatostatin directly inhibits substantia gelatinosa neurons in adult rat spinal dorsal horn in vitro. Neurosci. Res. 47, 97–107.
Jinno, S., Klausberger, T., Marton, L. F., Dalezios, Y., Roberts, J. D., Fuentealba, P., et al. (2007). Neuronal diversity in GABAergic long-range projections from the hippocampus. J. Neurosci. 27, 8790–8804.
Jolkkonen, J., Kahkonen, K., and Pitkanen, A. (1997). Cholinergic deafferentation exacerbates seizure-induced loss of somatostatin-immunoreactive neurons in the rat hippocampus. Neuroscience 80, 401–411.
Kapfer, C., Glickfeld, L. L., Atallah, B. V., and Scanziani, M. (2007). Supralinear increase of recurrent inhibition during sparse activity in the somatosensory cortex. Nat. Neurosci. 10, 743–753.
Kawaguchi, Y., and Kubota, Y. (1998). Neurochemical features and synaptic connections of large physiologically-identified GABAergic cells in the rat frontal cortex. Neuroscience 85, 677–701.
Kiagiadaki, F., Savvaki, M., and Thermos, K. (2010). Activation of somatostatin receptor (sst 5) protects the rat retina from AMPA-induced neurotoxicity. Neuropharmacology 58, 297–303.
Kiagiadaki, F., and Thermos, K. (2008). Effect of intravitreal administration of somatostatin and sst2 analogs on AMPA-induced neurotoxicity in rat retina. Invest. Ophthalmol. Vis. Sci. 49, 3080–3089.
Kokona, D., Mastrodimou, N., Pediaditakis, I., Charalampopoulos, I., Schmid, H. A., and Thermos, K. (2012). Pasireotide (SOM230) protects the retina in animal models of ischemia induced retinopathies. Exp. Eye Res. 103, 90–98.
Konradi, C., Yang, C. K., Zimmerman, E. I., Lohmann, K. M., Gresch, P., Pantazopoulos, H., et al. (2011). Hippocampal interneurons are abnormal in schizophrenia. Schizophr. Res. 131, 165–173.
Kosaka, K., and Kosaka, T. (2007). Chemical properties of type 1 and type 2 periglomerular cells in the mouse olfactory bulb are different from those in the rat olfactory bulb. Brain Res. 1167, 42–55.
Krantic, S., Robitaille, Y., and Quirion, R. (1992). Deficits in the somatostatin SS1 receptor sub-type in frontal and temporal cortices in Alzheimer’s disease. Brain Res. 573, 299–304.
Kumar, U. (2005). Expression of somatostatin receptor subtypes (SSTR1-5) in Alzheimer’s disease brain: an immunohistochemical analysis. Neuroscience 134, 525–538.
Lahlou, H., Guillermet, J., Hortala, M., Vernejoul, F., Pyronnet, S., Bousquet, C., et al. (2004). Molecular signaling of somatostatin receptors. Ann. N. Y. Acad. Sci. 1014, 121–131.
Lamirault, L., Guillou, J. L., Micheau, J., and Jaffard, R. (2001). Intrahippocampal injections of somatostatin dissociate acquisition from the flexible use of place responses. Eur. J. Neurosci. 14, 567–570.
Lavezzi, A. M., and Matturri, L. (2008). Functional neuroanatomy of the human pre-Bötzinger complex with particular reference to sudden unexplained perinatal and infant death. Neuropathology 28, 10–16.
Lepousez, G., Csaba, Z., Bernard, V., Loudes, C., Videau, C., Lacombe, J., et al. (2010a). Somatostatin interneurons delineate the inner part of the external plexiform layer in the mouse main olfactory bulb. J. Comp. Neurol. 518, 1976–1994.
Lepousez, G., Mouret, A., Loudes, C., Epelbaum, J., and Viollet, C. (2010b). Somatostatin contributes to in vivo gamma oscillation modulation and odor discrimination in the olfactory bulb. J. Neurosci. 30, 870–875.
Leresche, N., Asprodini, E., Emri, Z., Cope, D. W., and Crunelli, V. (2000). Somatostatin inhibits GABAergic transmission in the sensory thalamus via presynaptic receptors. Neuroscience 98, 513–522.
Levinson, A. J., Fitzgerald, P. B., Favalli, G., Blumberger, D. M., Daigle, M., and Daskalakis, Z. J. (2010). Evidence of cortical inhibitory deficits in major depressive disorder. Biol. Psychiatry 67, 458–464.
Llona, I., and Eugenín, J. (2005). Central actions of somatostatin in the generation and control of breathing. Biol. Res. 38, 347–352.
Lopez-Huerta, V. G., Blanco-Hernandez, E., Bargas, J., and Galarraga, E. (2012). Presynaptic modulation by somatostatin in the rat neostriatum is altered in a model of Parkinsonism. J. Neurophysiol. 108, 1032–1043.
Lopez-Huerta, V. G., Tecuapetla, F., Guzman, J. N., Bargas, J., and Galarraga, E. (2008). Presynaptic modulation by somatostatin in the neostriatum. Neurochem. Res. 33, 1452–1458.
Ma, Y., Hu, H., Berrebi, A. S., Mathers, P. H., and Agmon, A. (2006). Distinct subtypes of somatostatin-containing neocortical interneurons revealed in transgenic mice. J. Neurosci. 26, 5069–5082.
Mastrodimou, N., Lambrou, G. N., and Thermos, K. (2005). Effect of somatostatin analogues on chemically induced ischaemia in the rat retina. Naunyn Schmiedebergs Arch. Pharmacol. 371, 44–53.
McCarthy, A. D., Owens, I. J., Bansal, A. T., Mctighe, S. M., Bussey, T. J., and Saksida, L. M. (2011). FK962 and donepezil act synergistically to improve cognition in rats: potential as an add-on therapy for Alzheimer’s disease. Pharmacol. Biochem. Behav. 98, 76–80.
McDonald, A. J., Mascagni, F., and Zaric, V. (2012). Subpopulations of somatostatin-immunoreactive non-pyramidal neurons in the amygdala and adjacent external capsule project to the basal forebrain: evidence for the existence of GABAergic projection neurons in the cortical nuclei and basolateral nuclear complex. Front. Neural Circuits 6:46. doi: 10.3389/fncir.2012.00046
McGarry, L. M., Packer, A. M., Fino, E., Nikolenko, V., Sippy, T., and Yuste, R. (2010). Quantitative classification of somatostatin-positive neocortical interneurons identifies three interneuron subtypes. Front. Neural Circuits 4:12. doi: 10.3389/fncir.2011.00012
Melzer, S., Michael, M., Caputi, A., Eliava, M., Fuchs, E. C., Whittington, M. A., et al. (2012). Long-range-projecting GABAergic neurons modulate inhibition in hippocampus and entorhinal cortex. Science 335, 1506–1510.
Mendez-Diaz, M., Irwin, L., Gomez-Chavarin, M., Jimenez-Anguiano, A., Cabeza, R., Murillo-Rodriguez, E., et al. (2005). Cortistatin modulates memory evocation in rats. Eur. J. Pharmacol. 507, 21–28.
Momiyama, T., and Zaborszky, L. (2006). Somatostatin presynaptically inhibits both GABA and glutamate release onto rat basal forebrain cholinergic neurons. J. Neurophysiol. 96, 686–694.
Moneta, D., Richichi, C., Aliprandi, M., Dournaud, P., Dutar, P., Billard, J. M., et al. (2002). Somatostatin receptor subtypes 2 and 4 affect seizure susceptibility and hippocampal excitatory neurotransmission in mice. Eur. J. Neurosci. 16, 843–849.
Moore, S. D., Madamba, S. G., Joels, M., and Siggins, G. R. (1988). Somatostatin augments the M-current in hippocampal neurons. Science 239, 278–280.
Nanda, S. A., Qi, C., Roseboom, P. H., and Kalin, N. H. (2008). Predator stress induces behavioral inhibition and amygdala somatostatin receptor 2 gene expression. Genes Brain Behav. 7, 639–648.
Nilsson, A., Falth, M., Zhang, X., Kultima, K., Skold, K., Svenningsson, P., et al. (2009). Striatal alterations of secretogranin-1, somatostatin, prodynorphin, and cholecystokinin peptides in an experimental mouse model of Parkinson disease. Mol. Cell. Proteomics 8, 1094–1104.
Olias, G., Viollet, C., Kusserow, H., Epelbaum, J., and Meyerhof, W. (2004). Regulation and function of somatostatin receptors. J. Neurochem. 89, 1057–1091.
Oliva, A. A. Jr., Jiang, M., Lam, T., Smith, K. L., and Swann, J. W. (2000). Novel hippocampal interneuronal subtypes identified using transgenic mice that express green fluorescent protein in GABAergic interneurons. J. Neurosci. 20, 3354–3368.
Pantaleo, T., Mutolo, D., Cinelli, E., and Bongianni, F. (2011). Respiratory responses to somatostatin microinjections into the Bötzinger complex and the pre-Bötzinger complex of the rabbit. Neurosci. Lett. 498, 26–30.
Peineau, S., Potier, B., Petit, F., Dournaud, P., Epelbaum, J., and Gardette, R. (2003). AMPA-sst2 somatostatin receptor interaction in rat hypothalamus requires activation of NMDA and/or metabotropic glutamate receptors and depends on intracellular calcium. J. Physiol. 546, 101–117.
Perez-Santiago, J., Diez-Alarcia, R., Callado, L. F., Zhang, J. X., Chana, G., White, C. H., et al. (2012). A combined analysis of microarray gene expression studies of the human prefrontal cortex identifies genes implicated in schizophrenia. J. Psychiatr. Res. 46, 1464–1474.
Perrenoud, Q., Rossier, J., Geoffroy, H., Vitalis, T., and Gallopin, T. (2012). Diversity of GABAergic interneurons in layer VIa and VIb of mouse barrel cortex. Cereb. Cortex. doi: 10.1093/cercor/bhs032 [Epub ahead of print].
Picardo, M. A., Guigue, P., Bonifazi, P., Batista-Brito, R., Allene, C., Ribas, A., et al. (2011). Pioneer GABA cells comprise a subpopulation of hub neurons in the developing hippocampus. Neuron 71, 695–709.
Pinter, E., Helyes, Z., and Szolcsanyi, J. (2006). Inhibitory effect of somatostatin on inflammation and nociception. Pharmacol. Ther. 112, 440–456.
Qiu, C., Zeyda, T., Johnson, B., Hochgeschwender, U., De Lecea, L., and Tallent, M. K. (2008). Somatostatin receptor subtype 4 couples to the M-current to regulate seizures. J. Neurosci. 28, 3567–3576.
Radojevic, V., Hanusek, C., Setz, C., Brand, Y., Kapfhammer, J. P., and Bodmer, D. (2011). The somatostatinergic system in the mammalian cochlea. BMC Neurosci. 12:89. doi: 10.1186/1471-2202-12-89
Ramírez-Jarquín, J. O., Lara-Hernández, S., López-Guerrero, J. J., Aguileta, M. A., Rivera-Angulo, A. J., Sampieri, A., et al. (2012). Somatostatin modulates generation of inspiratory rhythms and determines asphyxia survival. Peptides 34, 360–372.
Ramos, B., Baglietto-Vargas, D., Del Rio, J. C., Moreno-Gonzalez, I., Santa-Maria, C., Jimenez, S., et al. (2006). Early neuropathology of somatostatin/NPY GABAergic cells in the hippocampus of a PS1xAPP transgenic model of Alzheimer’s disease. Neurobiol. Aging 27, 1658–1672.
Rat, D., Schmitt, U., Tippmann, F., Dewachter, I., Theunis, C., Wieczerzak, E., et al. (2011). Neuropeptide pituitary adenylate cyclase-activating polypeptide (PACAP) slows down Alzheimer’s disease-like pathology in amyloid precursor protein-transgenic mice. FASEB J. 25, 3208–3218.
Raynor, K., Lucki, I., and Reisine, T. (1993). Somatostatin receptors in the nucleus accumbens selectively mediate the stimulatory effect of somatostatin on locomotor activity in rats. J. Pharmacol. Exp. Ther. 265, 67–73.
Robbins, R. J., Brines, M. L., Kim, J. H., Adrian, T., De Lanerolle, N., Welsh, S., et al. (1991). A selective loss of somatostatin in the hippocampus of patients with temporal lobe epilepsy. Ann. Neurol. 29, 325–332.
Roberts, G. W., Ferrier, I. N., Lee, Y., Crow, T. J., Johnstone, E. C., Owens, D. G., et al. (1983). Peptides, the limbic lobe and schizophrenia. Brain Res. 288, 199–211.
Royer, S., Zemelman, B. V., Losonczy, A., Kim, J., Chance, F., Magee, J. C., et al. (2012). Control of timing, rate and bursts of hippocampal place cells by dendritic and somatic inhibition. Nat. Neurosci. 15, 769–775.
Rubio, A., Perez, M., De Lecea, L., and Avila, J. (2008). Effect of cortistatin on tau phosphorylation at Ser262 site. J. Neurosci. Res. 86, 2462–2475.
Rubio, A., Sanchez-Mut, J. V., Garcia, E., Velasquez, Z. D., Oliver, J., Esteller, M., et al. (2012). Epigenetic control of somatostatin and cortistatin expression by beta amyloid peptide. J. Neurosci. Res. 90, 13–20.
Rudy, B., Fishell, G., Lee, S., and Hjerling-Leffler, J. (2011). Three groups of interneurons account for nearly 100% of neocortical GABAergic neurons. Dev. Neurobiol. 71, 45–61.
Saito, T., Iwata, N., Tsubuki, S., Takaki, Y., Takano, J., Huang, S. M., et al. (2005). Somatostatin regulates brain amyloid beta peptide Abeta42 through modulation of proteolytic degradation. Nat. Med. 11, 434–439.
Saiz-Sanchez, D., Ubeda-Banon, I., De La Rosa-Prieto, C., Argandona-Palacios, L., Garcia-Munozguren, S., Insausti, R., et al. (2010). Somatostatin, tau, and beta-amyloid within the anterior olfactory nucleus in Alzheimer disease. Exp. Neurol. 223, 347–350.
Saiz-Sanchez, D., Ubeda-Banon, I., De La Rosa-Prieto, C., and Martinez-Marcos, A. (2012). Differential expression of interneuron populations and correlation with amyloid-beta deposition in the olfactory cortex of an AbetaPP/PS1 transgenic mouse model of Alzheimer’s disease. J. Alzheimers Dis. 31, 113–129.
Samson, W. K., Zhang, J. V., Avsian-Kretchmer, O., Cui, K., Yosten, G. L., Klein, C., et al. (2008). Neuronostatin encoded by the somatostatin gene regulates neuronal, cardiovascular, and metabolic functions. J. Biol. Chem. 283, 31949–31959.
Sanchez-Alavez, M., Gomez-Chavarin, M., Navarro, L., Jimenez-Anguiano, A., Murillo-Rodriguez, E., Prado-Alcala, R. A., et al. (2000). Cortistatin modulates memory processes in rats. Brain Res. 858, 78–83.
Sandoval, K. E., Farr, S. A., Banks, W. A., Niehoff, M. L., Morley, J. E., Crider, A. M., et al. (2011). Chronic peripheral administration of somatostatin receptor subtype-4 agonist NNC 26-9100 enhances learning and memory in SAMP8 mice. Eur. J. Pharmacol. 654, 53–59.
Savonenko, A., Xu, G. M., Melnikova, T., Morton, J. L., Gonzales, V., Wong, M. P., et al. (2005). Episodic-like memory deficits in the APPswe/PS1dE9 mouse model of Alzheimer’s disease: relationships to beta-amyloid deposition and neurotransmitter abnormalities. Neurobiol. Dis. 18, 602–617.
Schettini, G., Florio, T., Magri, G., Grimaldi, M., Meucci, O., Landolfi, E., et al. (1988). Somatostatin and SMS 201-995 reverse the impairment of cognitive functions induced by cysteamine depletion of brain somatostatin. Eur. J. Pharmacol. 151, 399–407.
Schumann, C. M., Bauman, M. D., and Amaral, D. G. (2011). Abnormal structure or function of the amygdala is a common component of neurodevelopmental disorders. Neuropsychologia 49, 745–759.
Schwarzacher, S. W., Rüb, U., and Deller, T. (2011). Neuroanatomical characteristics of the human pre-Bötzinger complex and its involvement in neurodegenerative brainstem diseases. Brain 134, 24–35.
Schweitzer, P., Madamba, S. G., and Siggins, G. R. (1998). Somatostatin increases a voltage-insensitive K+ conductance in rat CA1 hippocampal neurons. J. Neurophysiol. 79, 1230–1238.
Shipley, M. T., and Ennis, M. (1996). Functional organization of olfactory system. J. Neurobiol. 30, 123–176.
Sibille, E., Morris, H. M., Kota, R. S., and Lewis, D. A. (2011). GABA-related transcripts in the dorsolateral prefrontal cortex in mood disorders. Int. J. Neuropsychopharmacol. 14, 721–734.
Siehler, S., Nunn, C., Hannon, J., Feuerbach, D., and Hoyer, D. (2008). Pharmacological profile of somatostatin and cortistatin receptors. Mol. Cell. Endocrinol. 286, 26–34.
Silberberg, G., and Markram, H. (2007). Disynaptic inhibition between neocortical pyramidal cells mediated by Martinotti cells. Neuron 53, 735–746.
Sousa, V. H., Miyoshi, G., Hjerling-Leffler, J., Karayannis, T., and Fishell, G. (2009). Characterization of Nkx6-2-derived neocortical interneuron lineages. Cereb. Cortex 19(Suppl. 1), i1–i10.
Spary, E. J., Maqbool, A., and Batten, T. F. C. (2008). Expression and localisation of somatostatin receptor subtypes sst1–sst5 in areas of the rat medulla oblongata involved in autonomic regulation. J. Chem. Neuroanat. 35, 49–66.
Stanić, D., Malmgren, H., He, H., Scott, L., Aperia, A., and Hökfelt, T. (2008). Developmental changes in frequency of the ciliary somatostatin receptor 3 protein. Brain Res. 1249, 101–112.
Stanley, E. M., Fadel, J. R., and Mott, D. D. (2012). Interneuron loss reduces dendritic inhibition and GABA release in hippocampus of aged rats. Neurobiol. Aging 33, 431.e1–13.
Stornetta, R. L., Rosin, D. L., Wang, H., Sevigny, C. P., Weston, M. C., and Guyenet, P. G. (2003). A group of glutamatergic interneurons expressing high levels of both neurokinin-1 receptors and somatostatin identifies the region of the pre-Bötzinger complex. J. Comp. Neurol. 455, 499–512.
Stumm, R. K., Zhou, C., Schulz, S., Endres, M., Kronenberg, G., Allen, J. P., et al. (2004). Somatostatin receptor 2 is activated in cortical neurons and contributes to neurodegeneration after focal ischemia. J. Neurosci. 24, 11404–11415.
Suzuki, N., and Bekkers, J. M. (2010). Distinctive classes of GABAergic interneurons provide layer-specific phasic inhibition in the anterior piriform cortex. Cereb. Cortex 20, 2971–2984.
Szabo, S., and Reichlin, S. (1981). Somatostatin in rat tissues is depleted by cysteamine administration. Endocrinology 109, 2255–2257.
Tahvildari, B., Wölfel, M., Duque, A., and Mccormick, D. A. (2012). Selective functional interactions between excitatory and inhibitory cortical neurons and differential contribution to persistent activity of the slow oscillation. J. Neurosci. 32, 12165–12179.
Tallent, M. K., Fabre, V., Qiu, C., Calbet, M., Lamp, T., Baratta, M. V., et al. (2005). Cortistatin overexpression in transgenic mice produces deficits in synaptic plasticity and learning. Mol. Cell. Neurosci. 30, 465–475.
Tallent, M. K., and Qiu, C. (2008). Somatostatin: an endogenous antiepileptic. Mol. Cell. Endocrinol. 286, 96–103.
Tallent, M. K., and Siggins, G. R. (1997). Somatostatin depresses excitatory but not inhibitory neurotransmission in rat CA1 hippocampus. J. Neurophysiol. 78, 3008–3018.
Tamminga, C. A., Foster, N. L., Fedio, P., Bird, E. D., and Chase, T. N. (1987). Alzheimer’s disease: low cerebral somatostatin levels correlate with impaired cognitive function and cortical metabolism. Neurology 37, 161–165.
Tan, W., Janczewski, W. A., Yang, P., Shao, X. M., Callaway, E. M., and Feldman, J. L. (2008). Silencing preBötzinger Complex somatostatin-expressing neurons induces persistent apnea in awake rat. Nat. Neurosci. 11, 538–540.
Tan, W., Pagliardini, S., Yang, P., Janczewski, W. A., and Feldman, J. L. (2010). Projections of preBötzinger complex neurons in adult rats. J. Comp. Neurol. 518, 1862–1878.
Taniguchi, H., He, M., Wu, P., Kim, S., Paik, R., Sugino, K., et al. (2011). A resource of Cre driver lines for genetic targeting of GABAergic neurons in cerebral cortex. Neuron 71, 995–1013.
Tashev, R., Belcheva, S., Milenov, K., and Belcheva, I. (2001). Behavioral effects of somatostatin microinjected into caudate putamen. Neuropeptides 35, 271–275.
Thermos, K. (2003). Functional mapping of somatostatin receptors in the retina: a review. Vision Res. 43, 1805–1815.
Thermos, K., Bagnoli, P., Epelbaum, J., and Hoyer, D. (2006). The somatostatin sst1 receptor: an autoreceptor for somatostatin in brain and retina? Pharmacol. Ther. 110, 455–464.
Tomioka, R., Okamoto, K., Furuta, T., Fujiyama, F., Iwasato, T., Yanagawa, Y., et al. (2005). Demonstration of long-range GABAergic connections distributed throughout the mouse neocortex. Eur. J. Neurosci. 21, 1587–1600.
Tripp, A., Kota, R. S., Lewis, D. A., and Sibille, E. (2011). Reduced somatostatin in subgenual anterior cingulate cortex in major depression. Neurobiol. Dis. 42, 116–124.
Vanetti, M., Kouba, M., Wang, X., Vogt, G., and Höllt, V. (1992). Cloning and expression of a novel mouse somatostatin receptor (SSTR2B). FEBS Lett. 311, 290–294.
Vecsei, L., Alling, C., and Widerlov, E. (1990). Comparative studies of intracerebroventricularly administered cysteamine and pantethine in different behavioral tests and on brain catecholamines in rats. Arch. Int. Pharmacodyn. Ther. 305, 140–151.
Vecsei, L., Bollok, I., and Telegdy, G. (1983). Intracerebroventricular somatostatin attenuates electroconvulsive shock-induced amnesia in rats. Peptides 4, 293–295.
Vecsei, L., Kiraly, C., Bollok, I., Nagy, A., Varga, J., Penke, B., et al. (1984). Comparative studies with somatostatin and cysteamine in different behavioral tests on rats. Pharmacol. Biochem. Behav. 21, 833–837.
Vecsei, L., and Widerlov, E. (1988). Effects of intracerebroventricularly administered somatostatin on passive avoidance, shuttle-box behaviour and open-field activity in rats. Neuropeptides 12, 237–242.
Vepsalainen, S., Helisalmi, S., Koivisto, A. M., Tapaninen, T., Hiltunen, M., and Soininen, H. (2007). Somatostatin genetic variants modify the risk for Alzheimer’s disease among Finnish patients. J. Neurol. 254, 1504–1508.
Vezzani, A., and Hoyer, D. (1999). Brain somatostatin: a candidate inhibitory role in seizures and epileptogenesis. Eur. J. Neurosci. 11, 3767–3776.
Villette, V., Poindessous-Jazat, F., Bellessort, B., Roullot, E., Peterschmitt, Y., Epelbaum, J., et al. (2012). A new neuronal target for beta-amyloid peptide in the rat hippocampus. Neurobiol. Aging 33, 1126.e1121–1114.
Villette, V., Poindessous-Jazat, F., Simon, A., Lena, C., Roullot, E., Bellessort, B., et al. (2010). Decreased rhythmic GABAergic septal activity and memory-associated theta oscillations after hippocampal amyloid-beta pathology in the rat. J. Neurosci. 30, 10991–11003.
Viollet, C., Lepousez, G., Loudes, C., Videau, C., Simon, A., and Epelbaum, J. (2008). Somatostatinergic systems in brain: networks and functions. Mol. Cell. Endocrinol. 286, 75–87.
Viollet, C., Vaillend, C., Videau, C., Bluet-Pajot, M. T., Ungerer, A., L’Heritier, A., et al. (2000). Involvement of sst2 somatostatin receptor in locomotor, exploratory activity and emotional reactivity in mice. Eur. J. Neurosci. 12, 3761–3770.
Wang, J. Z., Grundke-Iqbal, I., and Iqbal, K. (2007). Kinases and phosphatases and tau sites involved in Alzheimer neurofibrillary degeneration. Eur. J. Neurosci. 25, 59–68.
Wei, X.-Y., Zhao, Y., Wong-Riley, M. T. T., Ju, G., and Liu, Y.-Y. (2012). Synaptic relationship between somatostatin- and neurokinin-1 receptor-immunoreactive neurons in the pre-Bötzinger complex of rats. J. Neurochem. 122, 923–933.
Wilson, N. R., Runyan, C. A., Wang, F. L., and Sur, M. (2012). Division and subtraction by distinct cortical inhibitory networks in vivo. Nature 488, 343–348.
Wilson, R. S., Arnold, S. E., Schneider, J. A., Boyle, P. A., Buchman, A. S., and Bennett, D. A. (2009). Olfactory impairment in presymptomatic Alzheimer’s disease. Ann. N. Y. Acad. Sci. 1170, 730–735.
Xu, X., Roby, K. D., and Callaway, E. M. (2006). Mouse cortical inhibitory neuron type that coexpresses somatostatin and calretinin. J. Comp. Neurol. 499, 144–160.
Xu, X., Roby, K. D., and Callaway, E. M. (2010). Immunochemical characterization of inhibitory mouse cortical neurons: three chemically distinct classes of inhibitory cells. J. Comp. Neurol. 518, 389–404.
Xue, S., Jia, L., and Jia, J. (2009). Association between somatostatin gene polymorphisms and sporadic Alzheimer’s disease in Chinese population. Neurosci. Lett. 465, 181–183.
Yang, A. M., Ge, W. W., Lu, S. S., Yang, S. B., Su, S. F., Mi, Z. Y., et al. (2011a). Central administration of neuronostatin induces antinociception in mice. Peptides 32, 1893–1901.
Yang, A. M., Ji, Y. K., Su, S. F., Yang, S. B., Lu, S. S., Mi, Z. Y., et al. (2011b). Intracerebroventricular administration of neuronostatin induces depression-like effect in forced swim test of mice. Peptides 32, 1948–1952.
Yang, S. B., Yang, A. M., Su, S. F., Wang, H. H., Wang, N. B., and Chen, Q. (2012). Neuronostatin induces hyperalgesia in formalin test in mice. Neurosci. Lett. 506, 126–130.
Yeung, M., and Treit, D. (2012). The anxiolytic effects of somatostatin following intra-septal and intra-amygdalar microinfusions are reversed by the selective sst2 antagonist PRL2903. Pharmacol. Biochem. Behav. 101, 88–92.
Yosten, G. L., Pate, A. T., and Samson, W. K. (2011). Neuronostatin acts in brain to biphasically increase mean arterial pressure through sympatho-activation followed by vasopressin secretion: the role of melanocortin receptors. Am. J. Physiol. Regul. Integr. Comp. Physiol. 300, R1194–R1199.
Young, A., and Sun, Q.-Q. (2009). GABAergic inhibitory interneurons in the posterior piriform cortex of the GAD67-GFP mouse. Cereb. Cortex 19, 3011–3029.
Zafar, R., King, M. A., and Carney, P. R. (2012). Adeno associated viral vector-mediated expression of somatostatin in rat hippocampus suppresses seizure development. Neurosci. Lett. 509, 87–91.
Zeyda, T., Diehl, N., Paylor, R., Brennan, M. B., and Hochgeschwender, U. (2001). Impairment in motor learning of somatostatin null mutant mice. Brain Res. 906, 107–114.
Keywords: interneurons, SRIF, GPCR, sst, cortistatin, long-range, Alzheimer’s disease
Citation: Martel G, Dutar P, Epelbaum J and Viollet C (2012) Somatostatinergic systems: an update on brain functions in normal and pathological aging. Front. Endocrin. 3:154. doi: 10.3389/fendo.2012.00154
Received: 11 October 2012; Paper pending published: 06 November 2012;
Accepted: 20 November 2012; Published online: 06 December 2012.
Edited by:
Hubert Vaudry, University of Rouen, FranceCopyright: © 2012 Martel, Dutar, Epelbaum and Viollet. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits use, distribution and reproduction in other forums, provided the original authors and source are credited and subject to any copyright notices concerning any third-party graphics etc.
*Correspondence: Cécile Viollet, Inserm UMR894 - Center for Psychiatry and Neuroscience, Université Paris Descartes, Sorbonne Paris Cité, 2 ter rue d’Alésia, 75014 Paris, France. e-mail:Y2VjaWxlLnZpb2xsZXRAaW5zZXJtLmZy