 María Barrera-Esparza1,2Elizabeth Carreón-Torres3
María Barrera-Esparza1,2Elizabeth Carreón-Torres3 Angélica Saraí Jiménez-Osorio4Julieta Angel-García4Octavio Jiménez-Garza4Olga Rocío Flores-Chávez4Geu S. Mendoza-Catalán4
Angélica Saraí Jiménez-Osorio4Julieta Angel-García4Octavio Jiménez-Garza4Olga Rocío Flores-Chávez4Geu S. Mendoza-Catalán4 Diego Estrada-Luna4*
Diego Estrada-Luna4*- 1Coordinación de Unidades de Segundo Nivel, Oficina Central, Servicios de Salud del Instituto Mexicano del Seguro Social para el Bienestar (IMSS-BIENESTAR), Mexico City, Mexico
- 2Physics Institute, Universidad Nacional Autónoma de México (UNAM), Mexico City, Mexico
- 3Department of Molecular Biology, Instituto Nacional de Cardiologiía “Ignacio Chaívez”, Mexico City, Mexico
- 4Departament of Nursing, Instituto de Ciencias de la Salud, Universidad Autoínoma del Estado Hidalgo (UAEH), Hidalgo, Mexico
Fatty acids disorders may lead to insulin resistance, resulting in long-term oxidative stress and inflammatory processes, both mediated by adipose tissue. Either in normal condition or obesogenic status, adipose cells components play an important role in several physiological and metabolic conditions. It has been shown that bioactive compounds can influence the development of obesity and its pathological complications such as insulin resistance. In this study, we performed a network between bioactive compounds and adipose tissue vis-a-vis insulin resistance. We constructed a regulatory network of 62 adipocyte cell components that incorporates current evidence of cellular and molecular interactions involved in healthy and obesity states. The network incorporated information about inflammation pathways and inhibition of insulin signaling; insulin signaling and GLUT 4 translocation; triglycerides production; ATP production; M2 macrophages recruitment; adipogenesis and lipolysis as well as mitochondrial biogenesis. Our mathematical model showed a discernment between the impact of various bioactive substances on the transitions from health to obesity and vice versa. We found that anthocyanins, punicalagin, oleanolic acid, and NRG4 proved to be critical nodes in the transition from obesity to the healthy state, due to their switch-on potential to up-regulate the complex network resulting in a beneficial transition.
1 Introduction
Obesity is a complex, chronic degenerative disease from multifactorial origin, based on an imbalance in the absorption and use of calories (1) which is usually a risk factor for the development of cardiovascular diseases, some types of cancer, type 2 diabetes, neurodegenerative disorders and liver damage (2–5). Under regular conditions, adipose tissue produces several cytokines and hormones (adiponectin, leptin, resistin, adipokines, chemokines, etc.) which are important in lipid and metabolic homeostasis.
Either directly or indirectly, these biological factors interact with extrinsic and intrinsic factors, which are mainly involved in inflammatory processes, oxidative stress, and lipid profile, mediated adipose tissue. Nevertheless, in a normal condition or inflammation, obesogenic status, adipose cells components and other immune cells (such as M1 and M2 type) play an important role in several physiological and metabolic conditions. These cells exhibit distinct but complementary functions through different pro- or anti-inflammatories mediators such as Nuclear Factor kappa B (NFκ-B) (6), Toll-like receptors family (TLR2 and TLR4, mainly), tumoral necrosis factor-alpha (TNF-α) (7), peroxisome proliferator-activated receptor (PPAR) (8), glucose transporters (GLUT) (9), diverse interleukins (IL) (10), and hormones such as leptin, glucagon-like 1 (GLP1) and adiponectin (11) and Neuregulin 4 (NRG4). Likewise, these factors and obesity per se have been widely described to be directly implicated in the development of insulin resistance, which, in turn, unleashes and exerts diverse other metabolic disorders such as lipid profile dysregulation, mitochondrial dysfunction, lipoperoxidation, etc., leading to diabetes and cardiovascular diseases, mainly (12–17). In recent years, some factors that has been studied with the aim of correcting the abnormalities of glucose resistance include the effect of oxygenation of adipose tissue (18), reducing the levels of C-reactive protein (19), proving the efficacy of the consumption of glucagon-like peptide-1 receptor agonist (20), among others, nevertheless, the results are still heterogeneous. Furthermore, a considerable number of investigations have been performed involving adipose cells components in several biomodels to regulate these changes mentioned above, implementing diets, exercise and physical activity, and bioactive compounds (21–24) among others.
However, the importance of specific dietary composition can affect how food is absorbed and how it is stored, used or dissipated in adipose tissue. In this context, it has been shown that distinct bioactive compounds can influence obesity and its pathological complications such as insulin resistance, inflammation and metabolic syndrome in lipid turnover. Bioactive compounds such as phenolic compounds, triterpenes, anthocyanins, fructooligosaccharides, etc., can directly or indirectly modulate the signaling and expression of genes involved in regulating energy intake, lipid metabolism, adipogenesis in adipose tissue, thermogenesis, lipotoxicity, adipo/cytokine synthesis and inflammation (25–27). Thus, also at the intestinal level, higher concentrations of these molecules can be achieved, modulating the composition, action and function of the microbiome.
Some of this research has been focused on the development of clinical trials, experimental studies on biomodels and cell strains, as well as modelling mathematics to stimulate clinical conditions and to predict physiological patterns using medical physics techniques.
Application of mathematical models to predict biological molecule interactions, tissue behaviors or the natural course of disease (28–30) has provided the basis for comprehension and understanding of disease outcomes, response and progression, being an important field of scientific research for the development of clinical and experimental protocols. Several mathematical models based on systems biology have been previously considered (31–36) to elucidate the mechanisms underlying adipocytes’ homeostasis functions and alterations that drive them to transitions from health to obesogenic state and vice versa. We used a mathematical model based on a regulatory network composed of nodes representing molecular and cellular associations playing important interactions linked to the development of negative physiological pathways in adipose tissue during insulin resistance and obesity. In a first approach, a regulatory network can be characterized by a Boolean analysis, first proposed by Stuart Kauffman in the 60’s as a mathematical model of genetic regulation (37), where all the components only can have two values: zero (completely turned off or inhibited), or one (completely expressed or activated); these values depend on previous states of all nodes in the network. By considering a network formed by n nodes, the state of the node k at a time t+τ can be described by the mapping ; here, fk is determined by a logical proposition that satisfies Boole axiomatics. The stationary states of the mapping are determined by the condition (t), where this can define different cellular phenotypes, defined as obesity or health in this work.
Depending on their value, each component (node) of the network could affect the behavior of some other nodes or even the entire network. The final nodes values determine or can be associated with a physiological state of health or obesity. In a more realistic approach, regulatory networks can be characterized by considering values between zero and one and, once we establish the behavior and connections between nodes through differential equations, we can alter the physiological final state by modifying values of some critical nodes. One of the main objectives of this type of methodology is to identify the critical nodes that have the potential to change the final physiological state when we alter their values and to determine specific molecular factors that may influence different pathways either directly or combined, in order to be used as a predictive-methodological instrument in future experimental studies and clinical trials related to neurological conditions (38, 39).
This study aims to identify by theoretical predictions and mathematical modeling relevant bioactive compounds capable to modulate positive or negative the physiological mechanisms in adipose tissue under insulin resistance to guide experimental work and model biological systems more accurately.
2 Materials and methods
In this study, we performed a network between bioactive compounds and adipose tissue vis-a-vis insulin resistance. First, a literature review was performed in metabases including PUBMED, MEDLINE and SCOPUS, considering keywords such as “bioactive compounds” and “adipocyte tissue”, “insulin resistance biomodel” as well as the interaction and effects between them. Articles in English and Spanish were chosen as eligible literature considering randomized clinical and experimental studies published in the last five years. Subsequently, two authors independently reviewed each article in turn to discuss scientific evidence using three steps according to recommendations by Miller (40): a) Identifying the biological pathways linked to the clinical outcomes of the disease, i.e., the mechanism of pathogenesis. b) Distinguishing the dynamics of the bioactive compounds and adipocyte tissue and c) Describing how the dynamics of the bioactive compounds and adipocyte tissue modify the biological pathways leading to insulin resistance.
Based on the literature review mentioned above (41–70), we constructed a regulatory network of 62 adipocyte cell components (Figure 1) that incorporates current evidence of cellular and molecular interactions involved in healthy and obesity states (Supplementary Table S1). The network incorporates information about inflammation pathways and inhibition of insulin signaling; insulin signaling and GLUT 4 translocation; triglycerides production; ATP production; M2 macrophages recruitment; adipogenesis and lipolysis as well as mitochondrial biogenesis; which in presence of adipocyte failure, a condition common in obesity, leads to an increased level of fatty acids in muscle and liver tissue, resulting in a reduction of insulin response in various tissues and bioactive compounds.
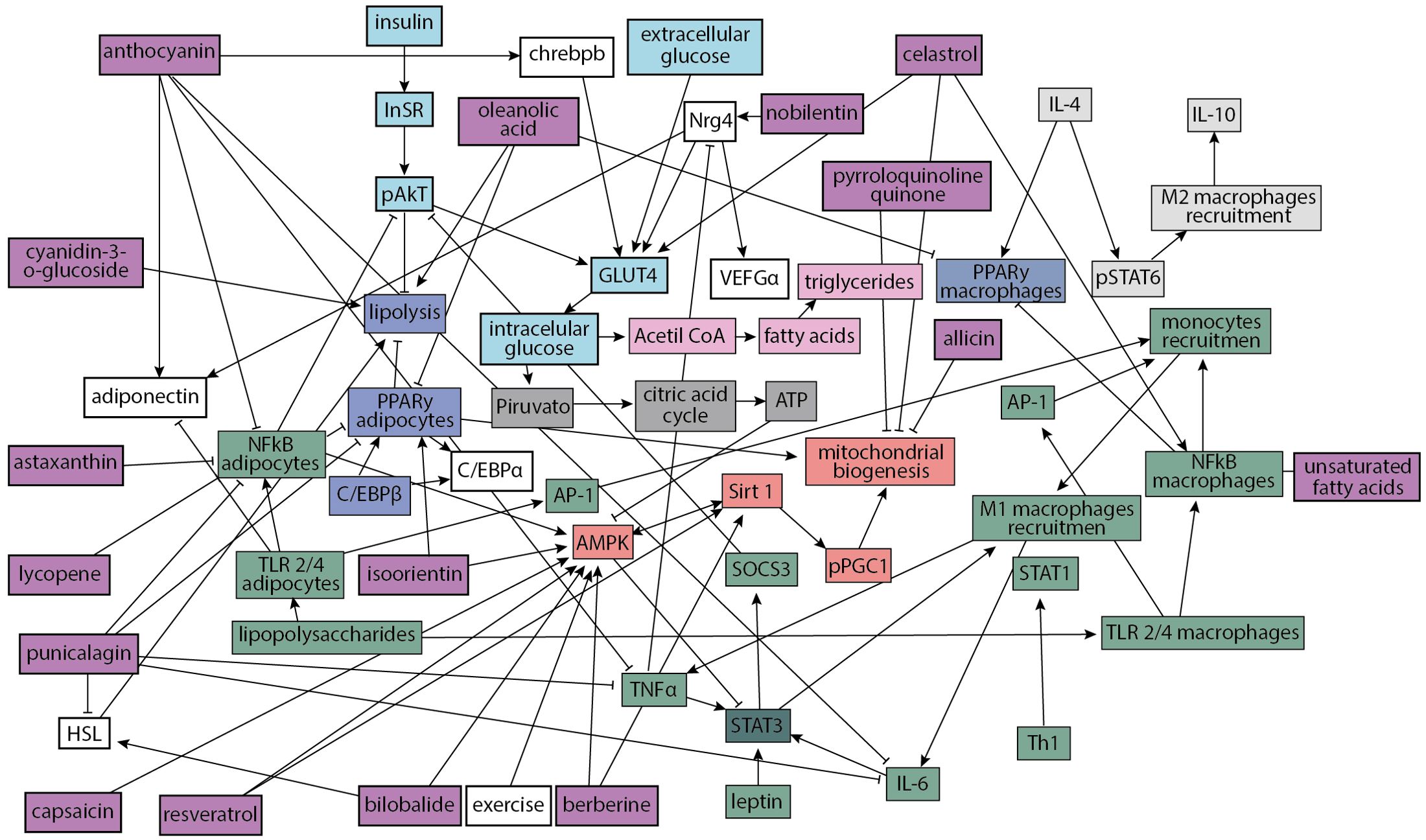
Figure 1. Adipocytes regulatory network. Inflammation pathways and inhibition of insulin signaling (green nodes); insulin signaling and GLUT 4 translocation (turquoise nodes); triglycerides production (pink nodes); ATP production (dark gray nodes); M2 macrophages recruitment (light gray nodes); adipogenesis and lipolysis (navy nodes); mitochondrial biogenesis (red nodes) and bioactive compounds (purple nodes). Interactions with final arrows represent a positive influence of one to another node, a node can “turn on” another node; meanwhile, interactions with red final bars indicate that a node can “turn off” or inhibit another node.
In Supplementary Table S1, we summarize the information of the literature review; for every node, we associated it with a value between zero and one for a state of health and obesity. Green cells correspond to node’s values greater than red ones (for example, anthocyanin has a higher expression level in health than in obesity). This table represents the attractors of the network: an attractor characterizes a steady state of values that remains stable over time while there are no disturbing external factors that change the final state. These attractors correspond to health and obesity states.
2.1 Mathematical description
Each node value can be seen as a variable that may vary from completely turned off (0) to completely expressed (1), and the degree to which an object exhibits a property p is given by a continuous expression µ[p], denoted as characteristic or membership function. The regulatory interactions are embraced by a characteristic function . Dynamical properties of the system were subsequently analyzed by means of a system of differential equations (32):
Where has a sigmoid structure and αk is a decay rate.
In this work, we studied the possible phase transitions between health and obesity by considering a set of ordinary differential equations for the expression levels of the network components. The characteristic function may be expressed by means of a function with a sigmoid structure:
Here, is a threshold value that renders true (expressed) if . The simplest assumption is to consider . Parameter b is a saturation rate for the change of the proposition from an unexpressed to a totally expressed state, being gradual for small b, and steep for large b. In this work we suppose that b=5.5.
For example, for the node insulin receptor, the differential equation that describe the expression level of the node over time is:
where =insulin, since the node insulin receptor only depends on the insulin node; b=5.5; =1 and is the expression level of the node insulin receptor at the time t.
2.2 Transitions between states
Dynamical analyses were performed to study the transitions from health to an obesity stage, by employing the continuous regime. This was analyzed with the Wolfram Mathematica computing system, using ordinary equations described in Supplementary Material 1. Each attractor in Supplementary Table S1 was in this step considered as a set of initial conditions of every node, and the analysis allowed to identify cases where the perturbation of a bioactive compound value induced a transition from the original to a different attractor, it was assumed that αk=1 for every node. This is equivalent to consider, for example, a person who is in an obesity stage characterized by a nutritional deficiency of bioactive compounds’ ingest and we examine if there will be a state transition of obesity to healthy state when the ingest of bioactive compounds increases. In results, the time function is a representation of the dynamic evolution of the nodes, where it is affected by the decay rate αk and by the saturation rate b. For small values of b, the change of the expression level of a node is gradual, while for large values of b, the node can change in a shorter time. On the other hand, if we increase the decay rate of a node, it will turn off in a short time. We show the time function in the results up to 20, because this is enough to exhibit a stable behavior of a node, where it can be turned on or turned off.
3 Results
3.1 Transition from obesity to health
In Figure 2, we observe an overall dynamic behavior reliable to a transition from disease (obesity) to health, where the inflammatory values decreased and the M2 macrophages recruitment increased, with a little increase of GLUT 4 expression and a steady state of M1 macrophages recruitment, triglycerides and ATP. This was achieved when all the bioactive compounds transited from small to big values.
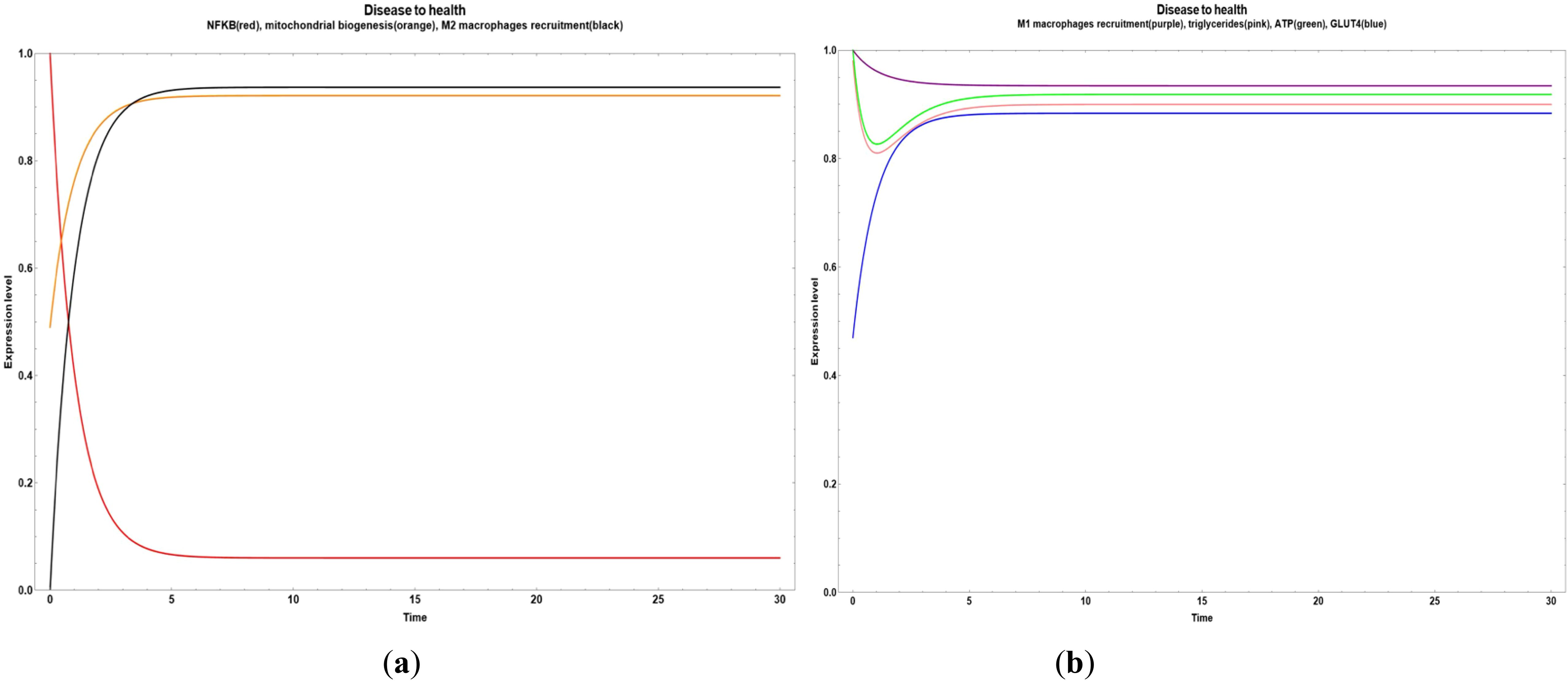
Figure 2. Transitions from disease to an improved health state when all the bioactive compounds are expressed inside of an obesity environment. (a) Dynamic transitions related to a decrease of NFκ-B in adipocytes and increase in mitochondrial biogenesis and M2 macrophages recruitment; (b) Dynamic transition showing an increase of GLUT 4 expression and a steady state of M1 macrophages recruitment, triglycerides and ATP.
3.1.1 Critical nodes: anthocyanin and punicalagin; neuroregulin 4 and oleanolic acid
We found some critical nodes driving the network final state from obesity to an improved state of health characterized by a reduced inflammatory value. For example, if we increase the anthocyanin and punicalagin values, we observe a drastic reduction in NF-κB (Figure 3a). On the other hand, if we increase only the neuroregulin 4 (NRG4) and the oleanolic acid, respectively, the final state transitions to a small reduction in inflammation, compared with the effect of turn on anthocyanin and punicalagin nodes, where they suppressed NF-κB, as it can be seen in Figures 3b and 4. However, the impact of NRG4 on mitochondrial biogenesis is clear, driving an evident increase when the NRG4 is turned on. In Figure 4, a marked increase in M2 macrophage recruitment occurred when the value of oleanolic acid is increased.
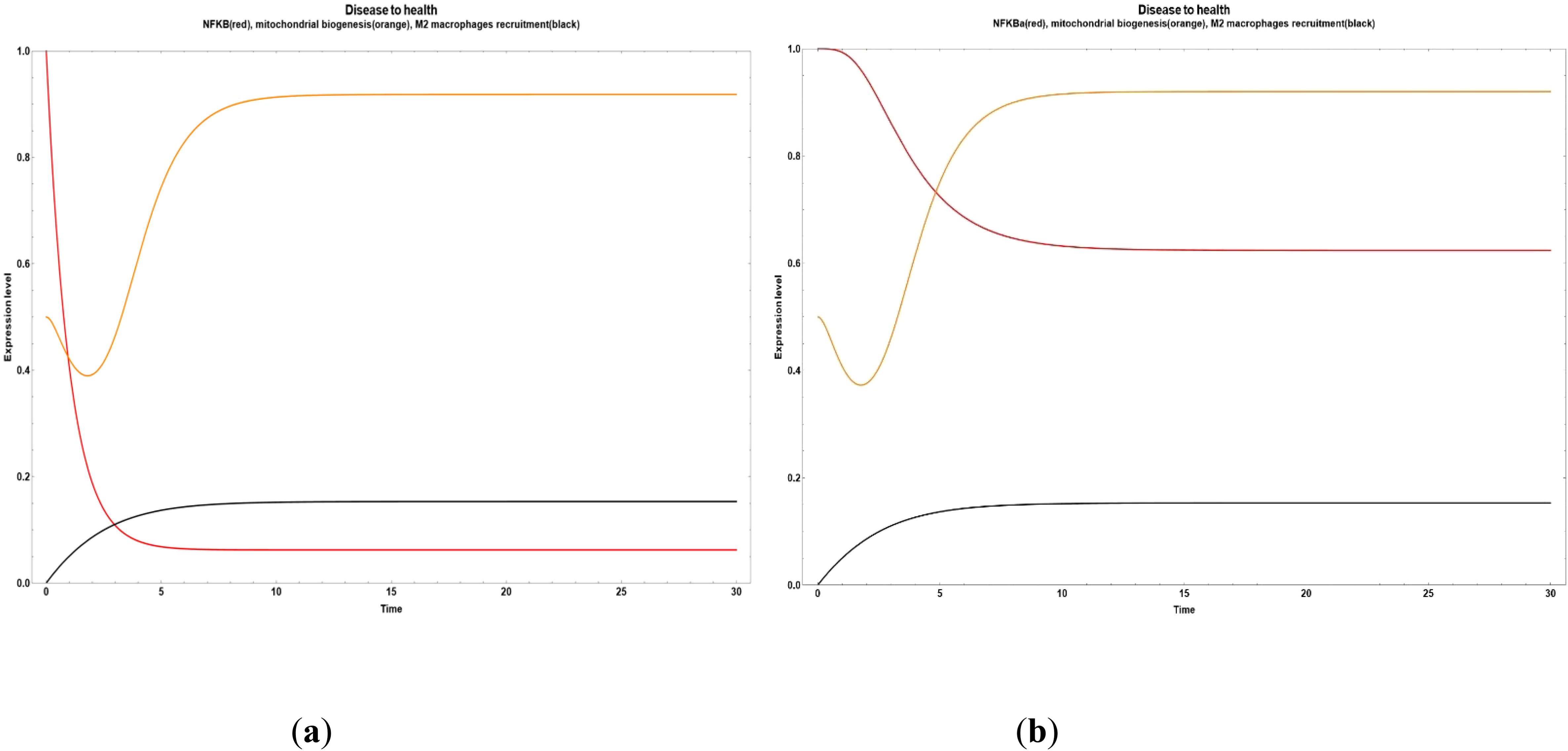
Figure 3. Transitions from disease to an improved health state when some bioactive compounds are expressed inside of an obesity environment. (a) Dynamic transitions related to an evident decrease of NFκ-B in adipocytes and a transient increase in mitochondrial biogenesis. In this scenario, we turned on the anthocyanin and punicalagin values (b) Dynamic transition showing a decrease of inflammation but not as big as in (a). This evolution is due to the NRG4 increased value.

Figure 4. Transition from disease to an improved health state when all the bioactive compounds are turned off, except for oleanolic acid.
3.2 Transition from health to obesity
In Figure 5, we can observe a pathway leading from health to disease with an early increase in NF-κB (Figure 5a) and M1 macrophages recruitment (Figure 5b). Both cases were stimulated when the values of all the network´s bioactive compounds were reduced.
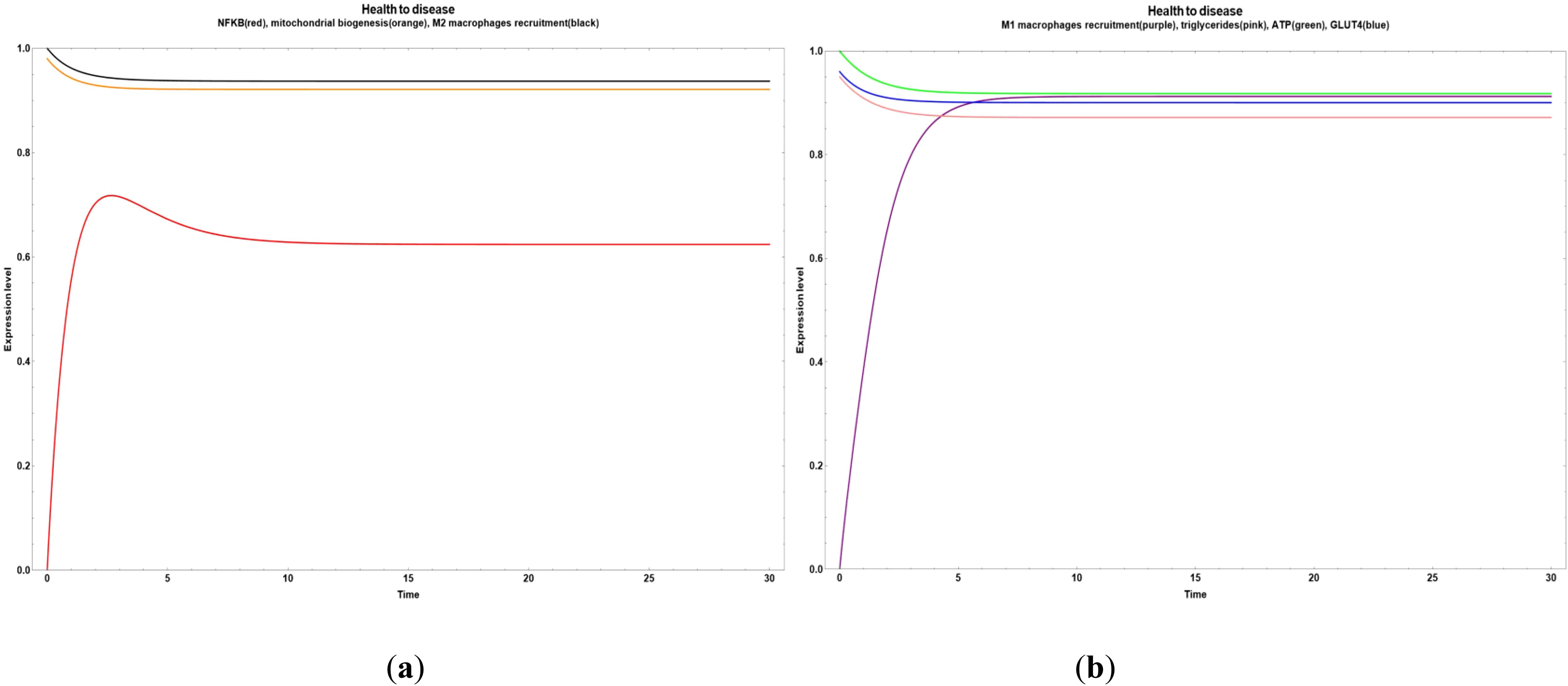
Figure 5. Transitions from health to disease state when all bioactive compounds are sub-expressed. (a) Dynamic transitions leading to an overexpression of NFκ-B and a steady expression of mitochondrial biogenesis and M2 macrophages recruitment. (b) Dynamic transition from a null to an evident M1 macrophages recruitment and a stable level of triglycerides, ATP and GLUT 4.
3.2.1 Critical nodes from health to disease
We found critical nodes driving the system from health to disease when their values turned on. In Figure 6, there is an evident increase in M1 macrophages recruitment when the node representing lipopolysaccharides increases its value. In this case, inflammation doesn’t suffer any change. Contrarily, if we modify the lipopolysaccharides value altogether with astaxanthin, anthocyanin, lycopene, punicalagin (suppressing their values), this condition promotes the NF-κB and M1 macrophages recruitment increase (Figures 7a, b).
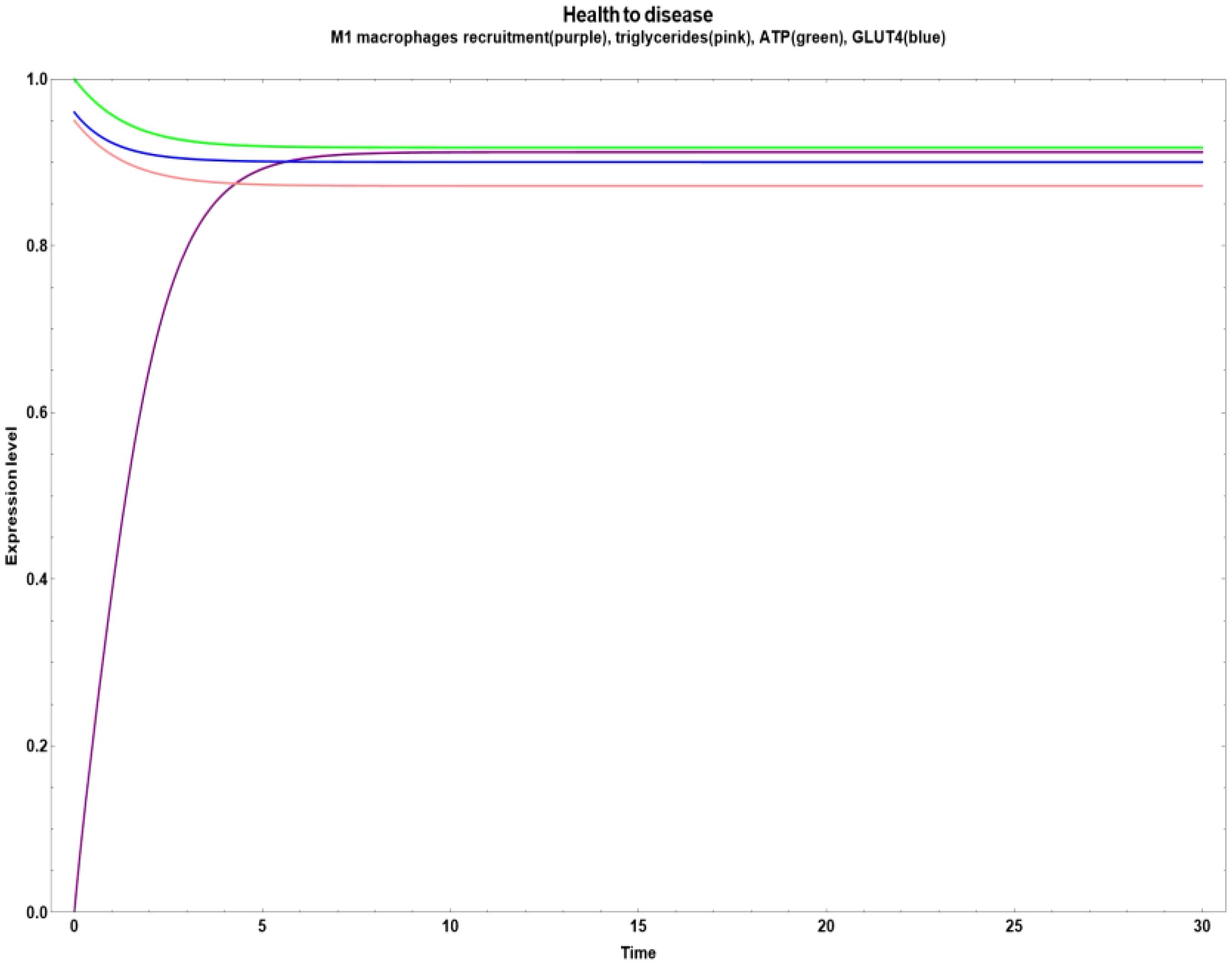
Figure 6. Transition from health to disease characterized by a boost in M1 macrophages recruitment caused by an increase in lipopolysaccharides.
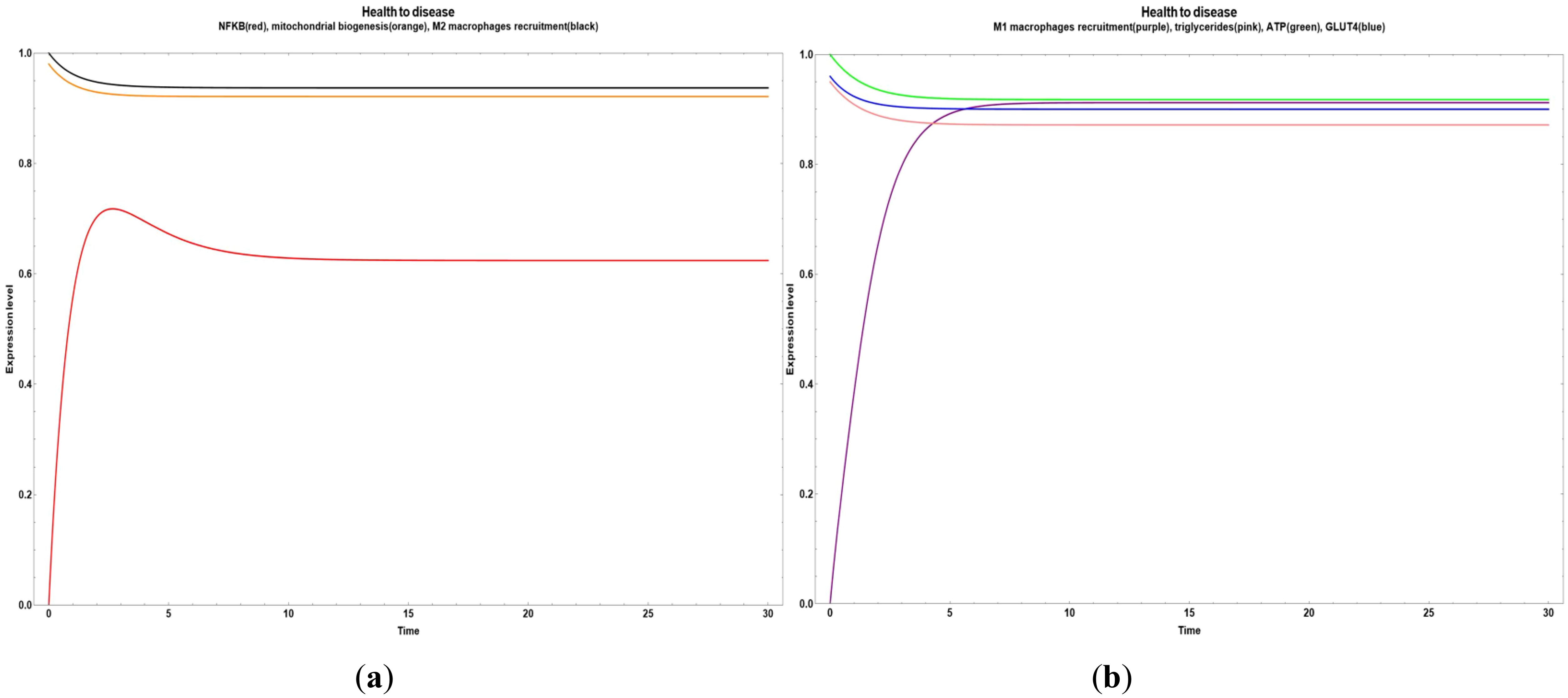
Figure 7. Transitions from health to disease are caused by a decrease in the astaxantin, anthocyanin, lycopene and punicalagin compounds, with a rise in lipopolysaccharides (all at the same time). (a) Increase in the NFκ-B inflammatory value. (b) Increase in the M1 macrophages recruitment with a constant behavior of triglycerides, ATP and GLUT 4.
Although the effect of the lipopolysaccharides increase without a decrease in the astaxanthin, anthocyanin, lycopene and punicalagin compounds does not affect the NFκ-B level, only increases the M1 macrophages recruitment; the increase of lipopolysaccharides while we reduce the astaxanthin, anthocyanin, lycopene and punicalagin levels produce a rise in NFκ-B in addition to the M1 macrophages recruitment.
3.3 Overall transitions
We resume the responsible critical nodes having the potential to induce a transition from obesity to health (Table 1) and from health to disease (Table 2).
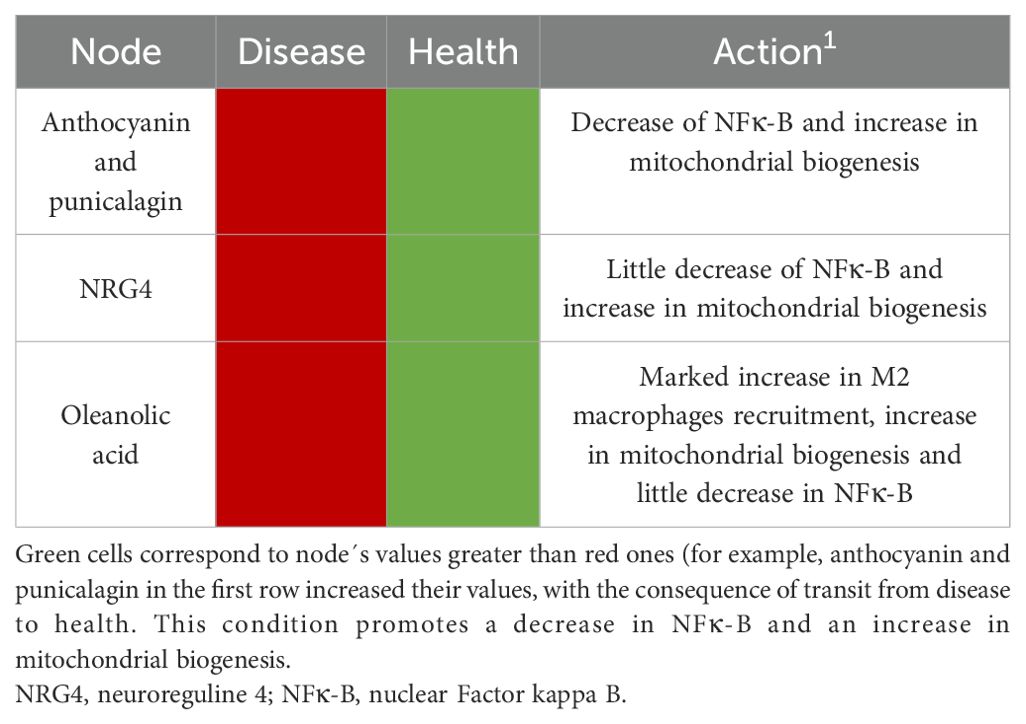
Table 1. Critical nodes that induce a transition from disease to health.

Table 2. Critical nodes that induce a transition from health to disease.
4 Discussion
Insulin resistance is a complex disease and a risk factor for the development of cardiometabolic diseases. The molecular mechanisms of insulin resistance in adipose tissue have been widely elucidated and used as a therapeutic target. The research of pharmacological interventions with the use of bioactive compounds also has been widely investigates with some molecular targets chosen for detailed analysis. However, the use of mathematical networks provides a tool for organizing large amounts of data in nodes to identify modules linked to a specific target that are reliably active only in the context of IR progression.
The study with mathematical models of the interactions (nodes and subnetworks) occurring between chronic-degenerative and infectious diseases with environmental, genetic and molecular factors (macromolecular complexes, enzymes, translation factors and genetic transcription) and, specifically, bioactive compounds or nutraceuticals, have been widely explored in recent decades by the global scientific community (71–74). These investigations have allowed us to understand the pathophysiological manifestations of diseases in different study models and the behavior of pharmacological and nutritional interventions in different pathologies, which can allow the construction of guides focused on new non-pharmacological treatments and the improvement or prevention in the evolution nature of chronic diseases (75–77). This has been widely reported in recent years, particularly in viral diseases, where the involvement and prediction of viral proteins, certain RNA strains, gene mutations or other transcription factors can promote disorder because of their connection and influence in turning on part of the system of these complex networks, and even identifying the initial cellular origin of the infectivity of viruses, providing significant information on the complex structures of interactions in these diseases (78–80).
We present a network constructed between obesity and different risk factors, based on the interactions of mitochondrial biogenesis, inflammatory pathways, insulin signaling and the expression of type 1 and 2 macrophages as well as different bioactive compounds. It has been reported that each of these elements could modulate or act as regulatory factors in adipose tissue.
Our analysis showed that the metabolic pathways commonly linked to obesity and insulin resistance (inflammation, oxidative stress, dysfunctional mitochondrial, etc.) exhibit a synergism with both extrinsic and intrinsic factors in a healthy and diseased state, such as anthocyanins and punicalagin. A possible explanation is that anthocyanins and punicalagin are a type of polyphenolic compounds with antioxidant and anti-inflammatory properties. Numerous pre-clinical and clinical studies have indicated that these polyphenolic compounds have significant efficacy in the prevention of malignant tumors, diabetes mellitus and cardiovascular diseases (81–84). A main source of these bioactive compounds is found in fresh foods such as blueberries, pomegranate, and cherries.
In order to know about the interdependence between adipose tissue response under insulin resistance and the dynamics in the network, we performed a simulation of all nodes associated with bioactive compounds reported to have an essential biological activity. Our analysis found that, when passing from an obesogenic to a healthy state, the main nodes capable of modulating the system by decreasing inflammatory processes regulated by cytokines and macrophages type 1, besides lipid homeostasis, were bioactive compounds such as anthocyanin and punicalagin, likewise oleanolic acid, and to adipokine, NRG4. Our results showed NRG4 as a main node that increases mitochondrial biogenesis, while not reduce inflammation drastically. However, altogether with oleanolic acid, the reduction of NF-κB is evident. On the other hand, when simulating from healthy to obesogenic state, significant changes in the network were the increase of NF-κB and the recruitment of type 1 macrophages with a shutdown of the nodes corresponding to bioactive compounds, whereas the critical node after the total resolutions in the system were the lipopolysaccharides.
Our mathematical model showed a discernment between the impact of various bioactive substances on the transitions from health to obesity and vice versa. In this context, of all the bioactive substances reported in our bibliographic research, our results established a hierarchy of importance between nodes to make a transition between final states associated with health or disease. We found that anthocyanins, punicalagin, oleanolic acid and NRG4 proved to be critical nodes in the transition from obesity to the healthy state, due to their individual switch-on potential to up-regulate the complex network resulting in beneficial transition. Historically, these nodes have been described in pre-clinical and clinical studies as anti-inflammatory, with antioxidant properties and positively influencing PPAR- γ pathway enhancing insulin response (85–87). There is evidence suggesting that oleanolic acid may improve inflammation and oxidative damage caused by diabetes (88, 89) since it improves the response to insulin (90). Additionally, anticancer, anti-inflammatory and antiatherogenic activities have been attributed to this molecule. In this context, in vitro studies with HepG2 cells, oleanolic acid can reduce lipid accumulation; a possible mechanism is through the negative regulation of gene transcription and protein expression of PPAR-γ, C/EBP-β and Sterol regulatory element-binding protein-1 (SREBP-1c). A crucial factor in regulating adipocyte differentiation, lipogenesis, and glucose metabolism gene expression is PPAR-γ (91). Likewise, SREBP1-C is an activator of PPAR-γ production and controls the expression of acid synthase. CCAAT-enhancing/binding protein β (C/EBPβ) plays a role in the induction of PPAR-γ and SREBP1-C. As a regulator of early adipocytes, as well as in maintaining the expression level in mature adipocytes. NRG4, is an adipokine, a secreted factor by brown adipose tissue, and its signaling protects hepatocytes from stress-induced injuries and attenuates those induced by a high-fat diet. Reduced NRG4 expression in adipose tissue and plasma levels has been associated with obesity and insulin resistance (92–95). NRG4 signaling has been implicated in the regulation of macrophage survival and function during intestinal inflammation (96, 97). Also, BAT-specific NRG4 deficiency accelerates vascular inflammation and adhesion responses, endothelial dysfunction and apoptosis and atherosclerosis in male mice.
On the other hand, another critical node from healthy to obesogenic state was LPS. In this context, bacterial endotoxins/LPS contributes to acute and chronic inflammations and triggers the innate immune response characterized by cytokine release and immune system activation. Changes in gut microbiota (a common condition in obesity) may induce endotoxemia processes produced by dysbiosis or disruption of the intestinal epithelial barrier which are important determinants involved in the pathophysiology of obesity in inflammatory processes and insulin resistance (98–100).
Network regulatory relations generate a cellular dynamic that portrays disease progression as a transit between attractors corresponding to health and obesity. The importance of the model used here was the identification of key nodes that have the potential to drive the final condition of obesity to a health state and vice versa. Findings of this study are limited by the adipocyte network design considered here, where we included inflammation pathways and inhibition of insulin signaling, insulin signaling and GLUT 4 translocation, triglycerides production, ATP production, M2 macrophages recruitment, adipogenesis and lipolysis, mitochondrial biogenesis and bioactive compounds. A mathematical model is never a complete representation of a biological system, even if it is complex; it is limited to identifying which component or components are responsible for a particular behavior. Therefore, one of the limitations of mathematical models is their reproducibility on a large scale. It is therefore crucial to have a biological system that allows us to investigate the predictions generated by this mathematical model.
One of the limitations of mathematical models is their large-scale reproducibility. Therefore, it is crucial to have a biological system that allows us to investigate the predictions generated by this mathematical model. In this sense, this network has an important role to elucidate or predict the activity of pathway endpoints (phenotypes) (101), drug targets (102) and crosstalk (103). The suggested application of the results of this network is the use of the nodes as potential targets for the design of new pharmacological strategies to transition from obesity to health. In the study design, the researcher should evaluate the M2/M1 macrophages recruitment and NF-kB expression.
Other pharmacological strategies could include the combination of anthocyanins with punicalagin to reduce proinflammatory states, measuring NF-κB in obesity models. In addition, the use of astaxanthin, anthocyanin, lycopene, and punicalagin should be investigated as a pretreatment to avoid the development of insulin resistance via LPS and M1 macrophages recruitment. However, this network may be complemented by inclusion of supplementary elements that may provide a more detailed description of the cellular mechanisms leading to obesity or an improved state of health.
5 Conclusion
This study uses a complex network composed of critical nodes which have been reported to interact both directly and indirectly during an obesogenic and insulin resistance process in order to select the nodes most important for the transition between a disease state and a healthy one and vice versa. Using this predictive mathematical model, it is possible to target treatments or molecular mechanisms in a short period of time. Controlled studies with the reported nodes are necessary to assess the effectiveness of the complex network and mathematical simulations.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Ethics statement
Ethical approval was not required for the study involving animals in accordance with the local legislation and institutional requirements because This is a study where the information was collected from previous published articles in order to describe the metabolic and molecular mechanisms and thus be able to design the complex network and the mathematical models.
Author contributions
MB-E: Conceptualization, Investigation, Methodology, Software, Writing – original draft, Writing – review & editing. EC-T: Formal Analysis, Methodology, Writing – original draft, Writing – review & editing. AJ-O: Formal Analysis, Methodology, Writing – original draft, Writing – review & editing. JA-G: Data curation, Methodology, Validation, Writing – review & editing. OJ-G: Data curation, Validation, Writing – review & editing. OF-C: Validation, Writing – review & editing. GM-C: Validation, Writing – review & editing. DE-L: Conceptualization, Funding acquisition, Investigation, Methodology, Validation, Writing – original draft, Writing – review & editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This research was funded by “Fondos Estatales Genéricos de la Universidad Autoínoma del Estado de Hidalgo”, Number PAO-2025-0394.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fendo.2025.1578552/full#supplementary-material
Supplementary Table 1 | We summarize the information of the literature review; for every node, we associated it with a value between zero and one for a state of health and obesity. Green cells correspond to node’s values greater than red ones (for example, anthocyanin has a higher expression level in health than in obesity). 1 The column health relates the initial values of every node in the case of health 2 The column obesity shows the common values in each node compared to the healthy state. HSL, hormone-sensitive lipase; NRG4, neuroreguline 4; VEGFα, vascular endothelial growth factor alpha; ChREBP-B, carbohydrate-responsive element-binding protein beta; Akt, protein kinase B; GLUT4, glucose transporter 4; ATP, adenosin triphosphate; TLR2/4, Toll-like receptor 2/4; NF-κB, nuclear Factor-kappaB; TNFα, tumor necrosis factor alpha STAT3, signal transducer and activator of transcription 3; SOCS3, suppressor of cytokine signaling 3; STAT1, signal transducer and activator of transcription 1; IL-6, interleukine 6; Th1, helper T cell 1; AP-1, AP-1, activator protein-1; C/EBPβ, CCAAT/enhancer‐binding protein beta; C/EBPα, CCAAT/enhancer‐binding protein alpha; PPAR-γ, peroxisome proliferator-activated receptor gamma; IL-4, interleukine 4; pSTAT6, phosphorylated signal transducer and activator of transcription 6; IL-10, interleukine 10; AMPK, AMP-activated protein kinase; SIRT1, silent information regulator sirtuin 1; pPGC1, phosphorylated peroxisome proliferator-activated receptor gamma coactivator.
References
1. Tanti J-F, Ceppo F, Jager J, Berthou F. Implication of inflammatory signaling pathways in obesity-induced insulin resistance. Front Endocrinol (Lausanne). (2012) 3:181. doi: 10.3389/fendo.2012.00181
2. Liu M, Xu X, Chen X, Guo Y, Zhang S, Lin Y, et al. Body weight time in target range and cardiovascular outcomes in adults with overweight/obesity and type 2 diabetes. Eur J Prev Cardiol. (2023) 30:1263–71. doi: 10.1093/eurjpc/zwad165
3. Xie W, Hong Z, Li B, Huang B, Dong S, Cai Y, et al. Influence of glucagon-like peptide-1 receptor agonists on fat accumulation in patients with diabetes mellitus and non-alcoholic fatty liver disease or obesity: A systematic review and meta-analysis of randomized control trials. J Diabetes Complications. (2024) 38:108743. doi: 10.1016/j.jdiacomp.2024.108743
4. Nikrad N, Farhangi MA, Fard Tabrizi FP, Vaezi M, Mahmoudpour A, Mesgari-Abbasi M. The effect of calorie-restriction along with thylakoid membranes of spinach on the gut-brain axis pathway and oxidative stress biomarkers in obese women with polycystic ovary syndrome: A randomized, double-blind, placebo-controlled clinical trial. J Ovarian Res. (2023) 16:216. doi: 10.1186/s13048-023-01288-x
5. Tzschaschel M, Friedl TWP, Schochter F, Schütze S, Polasik A, Fehm T, et al. Association between obesity and circulating tumor cells in early breast cancer patients. Clin Breast Cancer. (2023) 23:e345–53. doi: 10.1016/j.clbc.2023.05.011
6. Niersmann C, Röhrig K, Blüher M, Roden M, Herder C, Carstensen-Kirberg M. Increased release of proinflammatory proteins in primary human adipocytes and activation of the inflammatory NFĸB, P38, and ERK pathways upon omentin treatment. Obes Facts. (2020) 13:221–36. doi: 10.1159/000506405
7. Al Madhoun A, Kochumon S, Haddad D, Thomas R, Nizam R, Miranda L, et al. Adipose tissue caveolin-1 upregulation in obesity involves TNF-α/NF-κB mediated signaling. Cells. (2023) 12:1019. doi: 10.3390/cells12071019
8. Cataldi S, Aprile M, Melillo D, Mucel I, Giorgetti-Peraldi S, Cormont M, et al. TNFα Mediates inflammation-induced effects on PPARG splicing in adipose tissue and mesenchymal precursor cells. Cells. (2021) 11:42. doi: 10.3390/cells11010042
9. Gaugel J, Haacke N, Sehgal R, Jähnert M, Jonas W, Hoffmann A, et al. Picalm, a novel regulator of GLUT4-trafficking in adipose tissue. Mol Metab. (2024) 88:102014. doi: 10.1016/j.molmet.2024.102014
10. Frühbeck G, Gómez-Ambrosi J, Ramírez B, Mentxaka A, Rodríguez A, Becerril S, et al. Increased levels of interleukin-36 in obesity and type 2 diabetes fuel adipose tissue inflammation by inducing its own expression and release by adipocytes and macrophages. Front Immunol. (2022) 13:832185. doi: 10.3389/fimmu.2022.832185
11. Oster H. Adipose endothelin signaling-an unusual suspect linking obesity to insulin resistance. Acta Physiol (Oxf). (2024) 240:e14241. doi: 10.1111/apha.14241
12. Lungarotti F, Sebastiani M, Giacchi R. Value of total parenteral nutrition after total pancreatectomy. J Chir (Paris). (1985) 122:133–5.
13. Dasilva G, Lois S, Méndez L, Miralles-Pérez B, Romeu M, Ramos-Romero S, et al. Fish oil improves pathway-oriented profiling of lipid mediators for maintaining metabolic homeostasis in adipose tissue of prediabetic rats. Front Immunol. (2021) 12:608875. doi: 10.3389/fimmu.2021.608875
14. Khalil M, Shanmugam H, Abdallah H, John Britto JS, Galerati I, Gómez-Ambrosi J, et al. The potential of the mediterranean diet to improve mitochondrial function in experimental models of obesity and metabolic syndrome. Nutrients. (2022) 14:3112. doi: 10.3390/nu14153112
15. Patel D, Rathaur P, Parwani K, Patel F, Sharma D, Johar K, et al. In vitro, in vivo, and in silico analysis of synbiotics as preventive interventions for lipid metabolism in ethanol-induced adipose tissue injury. Lipids Health Dis. (2023) 22:49. doi: 10.1186/s12944-023-01809-z
16. Michicotl-Meneses MM, Thompson-Bonilla MDR, Reyes-López CA, García-Pérez BE, López-Tenorio II, Ordaz-Pichardo C, et al. Inflammation markers in adipose tissue and cardiovascular risk reduction by pomegranate juice in obesity induced by a hypercaloric diet in wistar rats. Nutrients. (2021) 13:2577. doi: 10.3390/nu13082577
17. Régnier M, Van Hul M, Roumain M, Paquot A, de Wouters d’Oplinter A, Suriano F, et al. Inulin increases the beneficial effects of rhubarb supplementation on high-fat high-sugar diet-induced metabolic disorders in mice: impact on energy expenditure, brown adipose tissue activity, and microbiota. Gut Microbes. (2023) 15:2178796. doi: 10.1080/19490976.2023.2178796
18. Crewe C, An YA, Scherer PE. The ominous triad of adipose tissue dysfunction: inflammation, fibrosis, and impaired angiogenesis. J Clin Invest. (2017) 127:74–82. doi: 10.1172/JCI88883
19. Ghalandari H, Askarpour M, Setayesh L, Ghaedi E. Effect of plum supplementation on blood pressure, weight indices, and C-reactive protein: A systematic review and meta-analysis of randomized controlled trials. Clin Nutr ESPEN. (2022) 52:285–95. doi: 10.1016/j.clnesp.2022.09.010
20. Alkhezi OS, Alahmed AA, Alfayez OM, Alzuman OA, Almutairi AR, Almohammed OA. Comparative effectiveness of glucagon-like peptide-1 receptor agonists for the management of obesity in adults without diabetes: A network meta-analysis of randomized clinical trials. Obes Rev. (2023) 24:e13543. doi: 10.1111/obr.13543
21. de Sousa Neto IV, Durigan JLQ, da Silva ASR, de Cássia Marqueti R. Adipose tissue extracellular matrix remodeling in response to dietary patterns and exercise: molecular landscape, mechanistic insights, and therapeutic approaches. Biol (Basel). (2022) 11:765. doi: 10.3390/biology11050765
22. Zhang Y-X, Ou M-Y, Yang Z-H, Sun Y, Li Q-F, Zhou S-B. Adipose tissue aging is regulated by an altered immune system. Front Immunol. (2023) 14:1125395. doi: 10.3389/fimmu.2023.1125395
23. de Melo DG, Anaruma CP, da Cruz Rodrigues KC, Pereira RM, de Campos TDP, Canciglieri RS, et al. Strength training alters the tissue fatty acids profile and slightly improves the thermogenic pathway in the adipose tissue of obese mice. Sci Rep. (2022) 12:6913. doi: 10.1038/s41598-022-10688-w
24. Medeiros CS, de Sousa Neto IV, Silva KKS, Cantuária APC, Rezende TMB, Franco OL, et al. The effects of high-protein diet and resistance training on glucose control and inflammatory profile of visceral adipose tissue in rats. Nutrients. (2021) 13:1969. doi: 10.3390/nu13061969
25. MaChado AM, da Silva NBM, de Freitas RMP, de Freitas MBD, Chaves JBP, Oliveira LL, et al. Effects of yacon flour associated with an energy restricted diet on intestinal permeability, fecal short chain fatty acids, oxidative stress and inflammation markers levels in adults with obesity or overweight: A randomized, double blind, placebo controlled clinical trial. Arch Endocrinol Metab. (2021) 64:597–607. doi: 10.20945/2359-3997000000225
26. Belza A, Frandsen E, Kondrup J. Body fat loss achieved by stimulation of thermogenesis by a combination of bioactive food ingredients: A placebo-controlled, double-blind 8-week intervention in obese subjects. Int J Obes (Lond). (2007) 31:121–30. doi: 10.1038/sj.ijo.0803351
27. Chyau C-C, Chu C-C, Chen S-Y, Duh P-D. The inhibitory effects of djulis (Chenopodium formosanum) and its bioactive compounds on adipogenesis in 3T3-L1 adipocytes. Molecules. (2018) 23:1780. doi: 10.3390/molecules23071780
28. Murphy RJ, Maclaren OJ, Simpson MJ. Implementing measurement error models with mechanistic mathematical models in a likelihood-based framework for estimation, identifiability analysis and prediction in the life sciences. J R Soc Interface. (2024) 21:20230402. doi: 10.1098/rsif.2023.0402
29. Arabidarrehdor G, Tivay A, Bighamian R, Meador C, Kramer GC, Hahn J-O, et al. Mathematical model of volume kinetics and renal function after burn injury and resuscitation. Burns. (2021) 47:371–86. doi: 10.1016/j.burns.2020.07.003
30. Quirouette C, Younis NP, Reddy MB, Beauchemin CAAA. Mathematical model describing the localization and spread of influenza A virus infection within the human respiratory tract. PloS Comput Biol. (2020) 16:e1007705. doi: 10.1371/journal.pcbi.1007705
31. Azpeitia E, Davila-Velderrain J, Villarreal C, Alvarez-Buylla ER. Gene regulatory network models for floral organ determination. Methods Mol Biol. (2014) 1110:441–69. doi: 10.1007/978-1-4614-9408-9_26
32. Barrera M, Hiriart M, Cocho G, Villarreal C. Type 2 diabetes progression: A regulatory network approach. Chaos. (2020) 30:093132. doi: 10.1063/5.0011125
33. Resendis-Antonio O, González-Torres C, Jaime-Muñoz G, Hernandez-Patiño CE, Salgado-Muñoz CF. Modeling metabolism: A window toward a comprehensive interpretation of networks in cancer. Semin Cancer Biol. (2015) 30:79–87. doi: 10.1016/j.semcancer.2014.04.003
34. Sedaghat AR, Sherman A, Quon MJA. Mathematical model of metabolic insulin signaling pathways. Am J Physiol Endocrinol Metab. (2002) 283:E1084–1101. doi: 10.1152/ajpendo.00571.2001
35. Steinway SN, Zañudo JGT, Ding W, Rountree CB, Feith DJ, Loughran TP, et al. Network modeling of TGFβ Signaling in hepatocellular carcinoma epithelial-to-mesenchymal transition reveals joint sonic hedgehog and wnt pathway activation. Cancer Res. (2014) 74:5963–77. doi: 10.1158/0008-5472.CAN-14-0225
36. Villarreal C, Padilla-Longoria P, Alvarez-Buylla ER. General theory of genotype to phenotype mapping: derivation of epigenetic landscapes from N-node complex gene regulatory networks. Phys Rev Lett. (2012) 109:118102. doi: 10.1103/PhysRevLett.109.118102
37. Kauffman SA. Metabolic stability and epigenesis in randomly constructed genetic nets. J Theor Biol. (1969) 22:437–67. doi: 10.1016/0022-5193(69)90015-0
38. Nevitt SJ, Sudell M, Cividini S, Marson AG, Tudur Smith C. Antiepileptic drug monotherapy for epilepsy: A network meta-analysis of individual participant data. Cochrane Database Syst Rev. (2022) 4:CD011412. doi: 10.1002/14651858.CD011412.pub4
39. Birkinshaw H, Friedrich CM, Cole P, Eccleston C, Serfaty M, Stewart G, et al. Antidepressants for pain management in adults with chronic pain: A network meta-analysis. Cochrane Database Syst Rev. (2023) 5:CD014682. doi: 10.1002/14651858.CD014682.pub2
40. Miller G, Chen E, Cole SW. Health psychology: developing biologically plausible models linking the social world and physical health. Annu Rev Psychol. (2009) 60:501–24. doi: 10.1146/annurev.psych.60.110707.163551
41. Bu S, Yuan CY, Xue Q, Chen Y, Cao F. Bilobalide suppresses adipogenesis in 3T3-L1 adipocytes via the AMPK signaling pathway. Molecules. (2019) 24:3503. doi: 10.3390/molecules24193503
42. Qiu K, Zhao Q, Wang J, Qi G-H, Wu S-G, Zhang H-J. Effects of pyrroloquinoline quinone on lipid metabolism and anti-oxidative capacity in a high-fat-diet metabolic dysfunction-associated fatty liver disease chick model. Int J Mol Sci. (2021) 22:1458. doi: 10.3390/ijms22031458
43. Abdillah AM, Yun JW. Capsaicin induces ATP-dependent thermogenesis via the activation of TRPV1/B3-AR/A1-AR in 3T3-L1 adipocytes and mouse model. Arch Biochem Biophys. (2024) 755:109975. doi: 10.1016/j.abb.2024.109975
44. Mazibuko-Mbeje SE, Ziqubu K, Dludla PV, Tiano L, Silvestri S, Orlando P, et al. Isoorientin ameliorates lipid accumulation by regulating fat browning in palmitate-exposed 3T3-L1 adipocytes. Metabol Open. (2020) 6:100037. doi: 10.1016/j.metop.2020.100037
45. Shan Y, Zhang S, Gao B, Liang S, Zhang H, Yu X, et al. Adipose tissue SIRT1 regulates insulin sensitizing and anti-inflammatory effects of berberine. Front Pharmacol. (2020) 11:591227. doi: 10.3389/fphar.2020.591227
46. de Ligt M, Bruls YMH, Hansen J, Habets M-F, Havekes B, Nascimento EBM, et al. Resveratrol improves ex vivo mitochondrial function but does not affect insulin sensitivity or brown adipose tissue in first degree relatives of patients with type 2 diabetes. Mol Metab. (2018) 12:39–47. doi: 10.1016/j.molmet.2018.04.004
47. Jonscher KR, Stewart MS, Alfonso-Garcia A, DeFelice BC, Wang XX, Luo Y, et al. Early PQQ supplementation has persistent long-term protective effects on developmental programming of hepatic lipotoxicity and inflammation in obese mice. FASEB J. (2017) 31:1434–48. doi: 10.1096/fj.201600906R
48. Charlot A, Morel L, Bringolf A, Georg I, Charles A-L, Goupilleau F, et al. Octanoic acid-enrichment diet improves endurance capacity and reprograms mitochondrial biogenesis in skeletal muscle of mice. Nutrients. (2022) 14:2721. doi: 10.3390/nu14132721
49. Ali U, Wabitsch M, Tews D, Colitti M. Effects of allicin on human simpson-golabi-behmel syndrome cells in mediating browning phenotype. Front Endocrinol (Lausanne). (2023) 14:1141303. doi: 10.3389/fendo.2023.1141303
50. Kang B, Kim CY, Hwang J, Jo K, Kim S, Suh HJ, et al. Punicalagin, a pomegranate-derived ellagitannin, suppresses obesity and obesity-induced inflammatory responses via the nrf2/keap1 signaling pathway. Mol Nutr Food Res. (2019) 63:e1900574. doi: 10.1002/mnfr.201900574
51. Luo Q, Wei Y, Lv X, Chen W, Yang D, Tuo Q. The effect and mechanism of oleanolic acid in the treatment of metabolic syndrome and related cardiovascular diseases. Molecules. (2024) 29:758. doi: 10.3390/molecules29040758
52. Sandoval V, Femenias A, Martínez-Garza Ú, Sanz-Lamora H, Castagnini JM, Quifer-Rada P, et al. Lyophilized maqui (Aristotelia Chilensis) berry induces browning in the subcutaneous white adipose tissue and ameliorates the insulin resistance in high fat diet-induced obese mice. Antioxidants (Basel). (2019) 8:360. doi: 10.3390/antiox8090360
53. Abu Bakar MH, Shariff KA, Tan JS, Lee LK. Celastrol attenuates inflammatory responses in adipose tissues and improves skeletal muscle mitochondrial functions in high fat diet-induced obese rats via upregulation of AMPK/SIRT1 signaling pathways. Eur J Pharmacol. (2020) 883:173371. doi: 10.1016/j.ejphar.2020.173371
54. Tomasello L, Pitrone M, Guarnotta V, Giordano C, Pizzolanti G. Irisin: A possible marker of adipose tissue dysfunction in obesity. Int J Mol Sci. (2023) 24:12082. doi: 10.3390/ijms241512082
55. Ardid-Ruiz A, Ibars M, Mena P, Del Rio D, Muguerza B, Bladé C, et al. Potential involvement of peripheral leptin/STAT3 signaling in the effects of resveratrol and its metabolites on reducing body fat accumulation. Nutrients. (2018) 10:1757. doi: 10.3390/nu10111757
56. Casado ME, Canelles S, Arilla-Ferreiro E, Frago LM, Barrios V. Changes in lipid metabolism enzymes in rat epididymal fat after chronic central leptin infusion are related to alterations in inflammation and insulin signaling. Int J Mol Sci. (2023) 24:7065. doi: 10.3390/ijms24087065
57. Schink A, Neumann J, Leifke AL, Ziegler K, Fröhlich-Nowoisky J, Cremer C, et al. Screening of herbal extracts for TLR2- and TLR4-dependent anti-inflammatory effects. PloS One. (2018) 13:e0203907. doi: 10.1371/journal.pone.0203907
58. Jia Y, Wu C, Kim Y-S, Yang SO, Kim Y, Kim J-S, et al. A dietary anthocyanin cyanidin-3-O-glucoside binds to PPARs to regulate glucose metabolism and insulin sensitivity in mice. Commun Biol. (2020) 3:514. doi: 10.1038/s42003-020-01231-6
59. Abbasian F, Alavi MS, Roohbakhsh A. Dietary carotenoids to improve hypertension. Heliyon. (2023) 9:e19399. doi: 10.1016/j.heliyon.2023.e19399
60. Wang H-H, Sun Y-N, Qu T-Q, Sang X-Q, Zhou L-M, Li Y-X, et al. Nobiletin prevents D-galactose-induced C2C12 cell aging by improving mitochondrial function. Int J Mol Sci. (2022) 23:11963. doi: 10.3390/ijms231911963
61. Lands B. Highly unsaturated fatty acids (HUFA) mediate and monitor food’s impact on health. Prostaglandins Other Lipid Mediat. (2017) 133:4–10. doi: 10.1016/j.prostaglandins.2017.05.002
62. Spaulding HR, Yan Z. AMPK and the adaptation to exercise. Annu Rev Physiol. (2022) 84:209–27. doi: 10.1146/annurev-physiol-060721-095517
63. Engin A. Adiponectin resistance in obesity: adiponectin leptin/insulin interaction. Adv Exp Med Biol. (2024) 1460:431–62. doi: 10.1007/978-3-031-63657-8_15
64. Akram M. Citric acid cycle and role of its intermediates in metabolism. Cell Biochem Biophys. (2014) 68:475–8. doi: 10.1007/s12013-013-9750-1
65. Berkoz M, Yalin S, Yildirim M, Yalın AE, Çömelekoğlu Ü. Punicalagin and punicalin suppress the adipocyte differentiation through the transcription factors. Acta Endocrinol (Buchar). (2021) 17:157–67. doi: 10.4183/aeb.2021.157
66. Pfeiffer SEM, Quesada-Masachs E, McArdle S, Zilberman S, Yesildag B, Mikulski Z, et al. Effect of IL4 and IL10 on a human in vitro type 1 diabetes model. Clin Immunol. (2022) 241:109076. doi: 10.1016/j.clim.2022.109076
67. Freese R, Basu S, Mutanen M. Green tea extract does not affect urinary markers of lipid peroxidation or thromboxane or nitric oxide synthesis during a high-linoleic acid diet in healthy females. Lipids. (1999) 34 Suppl:S317. doi: 10.1007/BF02562331
68. Blankenau R. Foster G. Mcgaw award winner. Defining and solving community problems. Parkland memorial hospital, dallas. Trustee. (1994) 47:14–5.
69. Zhang Y, Zhang Y, Gu W, Sun B. TH1/TH2 cell differentiation and molecular signals. Adv Exp Med Biol. (2014) 841:15–44. doi: 10.1007/978-94-017-9487-9_2
70. Tang QQ, Lane MD. Adipogenesis: from stem cell to adipocyte. Annu Rev Biochem. (2012) 81:715–36. doi: 10.1146/annurev-biochem-052110-115718
71. Fojo AT, Kendall EA, Kasaie P, Shrestha S, Louis TA, Dowdy DW. Mathematical modeling of “Chronic” Infectious diseases: unpacking the black box. Open Forum Infect Dis. (2017) 4:ofx172. doi: 10.1093/ofid/ofx172
72. Kulesza A, Couty C, Lemarre P, Thalhauser CJ, Cao Y. Advancing cancer drug development with mechanistic mathematical modeling: bridging the gap between theory and practice. J Pharmacokinet Pharmacodyn. (2024) 51:581–604. doi: 10.1007/s10928-024-09930-x
73. Awad SF, Toumi AA, Al-Mutawaa KA, Alyafei SA, Ijaz MA, Khalifa SAH, et al. Type 2 diabetes epidemic and key risk factors in Qatar: A mathematical modeling analysis. BMJ Open Diabetes Res Care. (2022) 10:e002704. doi: 10.1136/bmjdrc-2021-002704
74. Zhou J, Wang H, Ouyang Q. Mathematical modeling of viral infection and the immune response controlled by the circadian clock. J Biol Phys. (2024) 50:197–214. doi: 10.1007/s10867-024-09655-5
75. Zhang Y, Huo M, Zhou J, Xie S. PKSolver: an add-in program for pharmacokinetic and pharmacodynamic data analysis in microsoft excel. Comput Methods Programs BioMed. (2010) 99:306–14. doi: 10.1016/j.cmpb.2010.01.007
76. Cegarra L, Aguirre P, Nuñez MT, Gerdtzen ZP, Salgado JC. Calcium is a noncompetitive inhibitor of DMT1 on the intestinal iron absorption process: empirical evidence and mathematical modeling analysis. Am J Physiol Cell Physiol. (2022) 323:C1791–806. doi: 10.1152/ajpcell.00411.2022
77. Kadakia E, Shah L, Amiji MM. Mathematical modeling and experimental validation of nanoemulsion-based drug transport across cellular barriers. Pharm Res. (2017) 34:1416–27. doi: 10.1007/s11095-017-2158-7
78. Bösl K, Ianevski A, Than TT, Andersen PI, Kuivanen S, Teppor M, et al. Common nodes of virus-host interaction revealed through an integrated network analysis. Front Immunol. (2019) 10:2186. doi: 10.3389/fimmu.2019.02186
79. Dábilla N, Dolan PT. Structure and dynamics of enterovirus genotype networks. Sci Adv. (2024) 10:eado1693. doi: 10.1126/sciadv.ado1693
80. Zitzmann C, Dächert C, Schmid B, van der Schaar H, van Hemert M, Perelson AS, et al. Mathematical modeling of plus-strand RNA virus replication to identify broad-spectrum antiviral treatment strategies. PloS Comput Biol. (2023) 19:e1010423. doi: 10.1371/journal.pcbi.1010423
81. Rossi M, Garavello W, Talamini R, La Vecchia C, Franceschi S, Lagiou P, et al. Flavonoids and risk of squamous cell esophageal cancer. Int J Cancer. (2007) 120:1560–4. doi: 10.1002/ijc.22499
82. Khan J, Deb PK, Priya S, Medina KD, Devi R, Walode SG, et al. Dietary flavonoids: cardioprotective potential with antioxidant effects and their pharmacokinetic, toxicological and therapeutic concerns. Molecules. (2021) 26:4021. doi: 10.3390/molecules26134021
83. Giuliani ME, Bigossi G, Lai G, Marcozzi S, Brunetti D, Malavolta M. Marine compounds and age-related diseases: the path from pre-clinical research to approved drugs for the treatment of cardiovascular diseases and diabetes. Mar Drugs. (2024) 22:210. doi: 10.3390/md22050210
84. Sabadashka M, Hertsyk D, Strugała-Danak P, Dudek A, Kanyuka O, Kucharska AZ, et al. Anti-diabetic and antioxidant activities of red wine concentrate enriched with polyphenol compounds under experimental diabetes in rats. Antioxidants (Basel). (2021) 10:1399. doi: 10.3390/antiox10091399
85. Brum ISDC, Mafra D, Moreira LDSG, Teixeira KTR, Stockler-Pinto MB, Cardozo LFMDF, et al. Consumption of oils and anthocyanins may positively modulate PPAR-γ Expression in chronic noncommunicable diseases: A systematic review. Nutr Res. (2022) 105:66–76. doi: 10.1016/j.nutres.2022.06.004
86. Su S, Wu G, Cheng X, Fan J, Peng J, Su H, et al. Oleanolic acid attenuates PCBs-induced adiposity and insulin resistance via HNF1b-mediated regulation of redox and PPARγ Signaling. Free Radic Biol Med. (2018) 124:122–34. doi: 10.1016/j.freeradbiomed.2018.06.003
87. Shi L, Li Y, Xu X, Cheng Y, Meng B, Xu J, et al. Brown adipose tissue-derived nrg4 alleviates endothelial inflammation and atherosclerosis in male mice. Nat Metab. (2022) 4:1573–90. doi: 10.1038/s42255-022-00671-0
88. Iskender H, Dokumacioglu E, Terim Kapakin KA, Yenice G, Mohtare B, Bolat I, et al. Effects of oleanolic acid on inflammation and metabolism in diabetic rats. Biotech Histochem. (2022) 97:269–76. doi: 10.1080/10520295.2021.1954691
89. Pollier J, Goossens A. Oleanolic acid. Phytochemistry. (2012) 77:10–5. doi: 10.1016/j.phytochem.2011.12.022
90. Yang Y, Fan L, Jiang J, Sun J, Xue L, Ma X, et al. M2 macrophage-polarized anti-inflammatory microneedle patch for accelerating biofilm-infected diabetic wound healing via modulating the insulin pathway. J Nanobiotechnology. (2024) 22:489. doi: 10.1186/s12951-024-02731-x
91. Salmerón C, Riera-Heredia N, Gutiérrez J, Navarro I, Capilla E. Adipogenic gene expression in gilthead sea bream mesenchymal stem cells from different origin. Front Endocrinol (Lausanne). (2016) 7:113. doi: 10.3389/fendo.2016.00113
92. Dai Y-N, Zhu J-Z, Fang Z-Y, Zhao D-J, Wan X-Y, Zhu H-T, et al. Case-control study: association between serum neuregulin 4 level and non-alcoholic fatty liver disease. Metabolism. (2015) 64:1667–73. doi: 10.1016/j.metabol.2015.08.013
93. Li Y, Jin L, Jiang F, Yan J, Lu Y, Yang Q, et al. Mutations of NRG4 contribute to the pathogenesis of nonalcoholic fatty liver disease and related metabolic disorders. Diabetes. (2021) 70:2213–24. doi: 10.2337/db21-0064
94. Wang Y, Huang S, Yu P. Association between circulating neuregulin4 levels and diabetes mellitus: A meta-analysis of observational studies. PloS One. (2019) 14:e0225705. doi: 10.1371/journal.pone.0225705
95. Wang G-X, Zhao X-Y, Meng Z-X, Kern M, Dietrich A, Chen Z, et al. The brown fat-enriched secreted factor nrg4 preserves metabolic homeostasis through attenuation of hepatic lipogenesis. Nat Med. (2014) 20:1436–43. doi: 10.1038/nm.3713
96. Schumacher MA, Dennis IC, Liu CY, Robinson C, Shang J, Bernard JK, et al. NRG4-erbB4 signaling represses proinflammatory macrophage activity. Am J Physiol Gastrointest Liver Physiol. (2021) 320:G990–G1001. doi: 10.1152/ajpgi.00296.2020
97. Schumacher MA, Hedl M, Abraham C, Bernard JK, Lozano PR, Hsieh JJ, et al. ErbB4 signaling stimulates pro-inflammatory macrophage apoptosis and limits colonic inflammation. Cell Death Dis. (2017) 8:e2622. doi: 10.1038/cddis.2017.42
98. Rosendo-Silva D, Viana S, Carvalho E, Reis F, Matafome P. Are gut dysbiosis, barrier disruption, and endotoxemia related to adipose tissue dysfunction in metabolic disorders? Overview of the mechanisms involved. Intern Emerg Med. (2023) 18:1287–302. doi: 10.1007/s11739-023-03262-3
99. Ferraz LF, Caria CREP, Santos R. de C.; ribeiro, M.L.; gambero, A. Effects of systemic inflammation due to hepatic ischemia-reperfusion injury upon lean or obese visceral adipose tissue. Acta Cir Bras. (2022) 37:e370105. doi: 10.1590/acb370105
100. Rittig N, Bach E, Thomsen HH, Pedersen SB, Nielsen TS, Jørgensen JO, et al. Regulation of lipolysis and adipose tissue signaling during acute endotoxin-induced inflammation: A human randomized crossover trial. PloS One. (2016) 11:e0162167. doi: 10.1371/journal.pone.0162167
101. Béal J, Pantolini L, Noël V, Barillot E, Calzone L. Personalized logical models to investigate cancer response to BRAF treatments in melanomas and colorectal cancers. PloS Comput Biol. (2021) 17:e1007900. doi: 10.1371/journal.pcbi.1007900
102. Poret A, Guziolowski C. Therapeutic target discovery using boolean network attractors: improvements of kali. R Soc Open Sci. (2018) 5:171852. doi: 10.1098/rsos.171852
Keywords: complex networks, adipose tissue, insulin resistance, bioactive compounds, inflammation, oxidative stress
Citation: Barrera-Esparza M, Carreón-Torres E, Jiménez-Osorio AS, Angel-García J, Jiménez-Garza O, Flores-Chávez OR, Mendoza-Catalán GS and Estrada-Luna D (2025) Complex networks interactions between bioactive compounds and adipose tissue vis-à-vis insulin resistance. Front. Endocrinol. 16:1578552. doi: 10.3389/fendo.2025.1578552
Received: 17 February 2025; Accepted: 21 April 2025;
Published: 13 May 2025.
Edited by:
Ramith Ramu, JSS Academy of Higher Education and Research, IndiaReviewed by:
Sandeep Kumar Swain, ICMR-National Institute of Child Health and Development Research, IndiaReshma Martiz, JSS Academy of Higher Education and Research, India
Anuragh Singh, SRM University, India
Copyright © 2025 Barrera-Esparza, Carreón-Torres, Jiménez-Osorio, Angel-García, Jiménez-Garza, Flores-Chávez, Mendoza-Catalán and Estrada-Luna. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Diego Estrada-Luna, ZGVzdHJhZGFfbHVuYUB1YWVoLmVkdS5teA==