 Fernando Suárez-Sánchez1*
Fernando Suárez-Sánchez1* Evelyn Pérez-Ruiz1,2
Evelyn Pérez-Ruiz1,2 Claudia Ivonne Ramírez-Silva3Mario Antonio Molina-Ayala4
Claudia Ivonne Ramírez-Silva3Mario Antonio Molina-Ayala4 Sandra Rivera-Gutiérrez5Lizbel León-Solís5
Sandra Rivera-Gutiérrez5Lizbel León-Solís5 Lázaro García-Morales6Arturo Rodríguez-González4César Martínez-Ortiz4Luis Axiel Meneses-Tapia1
Lázaro García-Morales6Arturo Rodríguez-González4César Martínez-Ortiz4Luis Axiel Meneses-Tapia1 Miguel Cruz-López1
Miguel Cruz-López1- 1Medical and Biochemistry Research Unit, Specialty Hospital, Centro Médico Nacional Siglo XXI, Instituto Mexicano del Seguro Social, Mexico City, Mexico
- 2Faculty of Chemistry, Doctorate Program in Medical, Dental and Health Sciences, Clinical and Experimental Health Research, Universidad Nacional Autonoma de México, Mexico City, Mexico
- 3Department of Maternal, Child, and Adolescent Nutrition. Center for Nutrition and Health Research. National Institute of Public Health. Cuernavaca, Morelos, Mexico
- 4Diabetes and Obesity Clinic, Specialty Hospital, Centro Médico Nacional Siglo XXI, Instituto Mexicano del Seguro Social, Mexico City, Mexico
- 5Department of Microbiology, Escuela Nacional de Ciencias Biológicas, Instituto Politécnico Nacional, Mexico City, Mexico
- 6Department of Molecular Biomedicine, Centro de Investigación y de Estudios Avanzados del Instituto Politécnico Nacional, Mexico City, Mexico
Background: Severe obesity is associated with metabolic alterations and an increased risk of developing type 2 diabetes. Bariatric surgery, especially malabsorptive procedures, results in significant clinical improvements and induces changes in the gut microbiota composition. This study aimed to identify bacterial taxa associated with changes in body mass index (BMI) in patients undergoing bariatric surgery and to explore their relationship with nutrient intake.
Methods: Individuals with severe obesity were recruited prior to and following bariatric surgery. Fecal DNA was extracted and the V4 region of the 16S rRNA gene was sequenced. Quality control and taxonomic classification were performed using QIIME2 and the Greengenes database. Nutrient intake was assessed through a 7-day dietary recall. Anthropometric measurements and blood samples were collected to evaluate clinical variables. Statistical analyses were conducted using R software.
Results: Significant differences in gut microbiota diversity were observed post-bariatric surgery. The Shannon and Simpson diversity indices decreased significantly after surgery (p < 0.001). Beta diversity analysis (Bray-Curtis, Weighted and Unweighted UniFrac) also showed significant differences between pre- and post-surgery samples (p = 0.001). The abundance of a species within the genus Coprococcus was positively correlated with magnesium and thiamin intake in post-surgery patients (rho = 0.816, pFDR = 0.029 and rho = 0.812, pFDR = 0.029, respectively). Furthermore, Coprococcus sp. abundance was positive associated with BMI in pre-surgery individuals (p = 0.043) but negative associated with BMI in post-surgery individuals (p = 0.036). Several taxa within the order Clostridiales and microbial metabolic pathways involved in sugar degradation, acetate, thiamin (vitamin B1) and some amino acid production were enriched prior to surgery.
Conclusions: The abundance of a species of the genus Coprococcus showed an inverse relationship with BMI in pre-surgery and post-surgery individuals and correlated positively with magnesium and thiamin intake in patients who underwent a malabsorptive bariatric surgery. These findings suggest that optimizing micronutrient intake may enhance the beneficial effects of bariatric surgery on BMI by favorably modulating gut microbiota composition.
1 Introduction
Obesity is a disease characterized by the excessive accumulation of fat at several body sites. Fat deposits around the waist are particularly detrimental and are associated with the onset of other pathologies such as cardiovascular disease and type 2 diabetes (T2D) (1). A body mass index (BMI) of 40 or higher is classified as severe obesity (2). In addition to the metabolic alterations inherent to obesity, patients often experience mobility problems and everyday life is severely affected. In most cases, dietary and exercise interventions have limited impact on weight loss; therefore, other approaches, such as bariatric surgery, are employed (3). Although different types of bariatric surgery exist, the use of malabsorptive bariatric surgery has demonstrated particularly favorable outcomes in obese patients. The most common procedure is Roux-en-Y gastric bypass (RYGB) although one-anastomosis gastric bypass (OAGB) has also been used successfully. Both surgeries involve rearranging the intestines in a way that limits nutrients absorption by bypassing the proximal portion of the small intestine and facilitating the passage of poorly digested food to the distal small intestine and colon (1, 4). These changes contribute to modifications in gut bacterial diversity and balance. This new microbial equilibrium influences the capacity of the gut bacteria to perform biologically relevant functions, including the synthesis of essential vitamins, the production of signaling molecules and digestion of nutrients (5).
Prebiotics and probiotics have been utilized to modulate the abundance of gut bacteria in the host. For example, fiber intake has been widely recommended to promote the growth of short-chain fatty acid (SCFA)-producing bacteria (6). However, the benefits of specific bacteria should be evaluated in a context-dependent manner since some bacteria such as the SCFA-producing bacteria Coprococcus has been linked to both positive and negative traits (7–16). Although less is known about the role of other dietary micronutrients in shaping the gut flora, evidence suggests that vitamins, minerals and trace elements in the diet can alter gut bacteria composition leading to improvements of clinical variables (17, 18). Particularly, thiamin and magnesium have been linked to changes in the abundance of Bacteroides, Alistipes, Bacilli and Bifidobacterium as well as to improvements in blood glucose, medium-chain fatty acids and some metabolic syndrome parameters (17, 19, 20).
In this study, we identified a species of Coprococcus whose abundance was higher in patients with greater BMI after bariatric surgery. The response of Coprococcus sp. to dietary intake was subsequently investigated, and we identified two micronutrients (magnesium and thiamin) that correlated with its abundance in the gut. Given the essential role of micronutrients in microbiota composition and the altered nutrient absorption following bariatric surgery, understanding the nutrients-microbiota relationship is critical for post-operative dietary guidance. Changes in several other bacterial taxa and associated metabolic pathways were observed in patients who underwent malabsorptive bariatric surgery.
2 Material and methods
2.1 Study population
Participants were recruited at the Specialty Hospital, National Medical Center Century XXI, Mexican Social Security Institute. A total of 21 individuals, all classified as severely obese, were scheduled to undergo an elective malabsorptive bariatric surgery (Roux-en-Y gastric bypass [RYGB] or one-anastomosis gastric bypass [OAGB]). Patients were excluded if they had undergone a previous cholecystectomy or bariatric surgery at the time of the recruitment. Other exclusion criteria, either at recruitment or after surgery were having taken antibiotics within the previous three months or presenting signs of gastrointestinal infection. All patients signed an informed consent form prior to the collection of any blood and fecal samples, anthropometric measurements, or dietary intake interviews. This study was approved by the National Ethics Committee of the Mexican Social Security Institute and was conducted in accordance with the ethical guidelines of the Declaration of Helsinki.
2.2 Sample collection and diet
Blood samples were collected before and 109 ± 28 days after patients underwent a bariatric surgery. Patients were instructed to fast for 10 hrs. prior to sampling and glucose, triglycerides, total cholesterol, HDL, LDL and insulin concentrations were measured in serum. Fecal samples were received, aliquoted and stored at -70°C by the laboratory personnel the same days that the blood samples were drawn. Dietary intake during the previous seven days was assessed using a Semi-Quantitative Food Frequency Questionnaire (SFFQ). This tool was based on the National Health and Nutrition Survey SFFQ and adapted to include foods available in different seasons, such as fruits and vegetables, reflecting seasonal variation in food consumption (21, 22). The questionnaire included 191 food items divided into 16 sections.
2.3 Micro- and macronutrient estimations
Energy and nutrients intake were calculated using the methodology applied in the National Health and Nutrition Survey of Mexico (ENSANUT) (23). The nutrients database was built using items from the Mexican Food Database (MFD; BAM, acronym in Spanish: Base de datos de los Alimentos Mexicanos) version 1.1 which contains information on 1978 foods, recipes, and beverages (24). To improve the accuracy of the dietary intake estimation, nutrient retention factor (NRF) were applied according to the recommendation of the United States Department of Agriculture (USDA) and Bognar recommendations (25, 26). Data processing was performed in Stata v.14.0 (Stata Corporation).
2.4 DNA extraction and 16S rDNA sequencing
Fecal samples were collected by patients within 24 hrs. prior to delivery to laboratory personnel. Samples were aliquoted and stored at -80°C. DNA was extracted from 150–200 mg of stool using the QIAamp DNA Mini Kit (Qiagen) following the manufacturer's instructions, with the addition of a mechanical lysis step at the beginning of the extraction procedure. The mechanical lysis was performed with the TissueLyser LT equipment (Qiagen) for 5 minutes at 60 cycles/second. DNA integrity was assessed by 2% agarose gel electrophoresis and purity was determined by the 260/280 ratio using a BioTek Epoch™ Microplate Spectrophotometer (Agilent Technologies).
Library preparation and sequencing of the V4 hypervariable region of the bacterial 16S rDNA gene were performed following the methodology described by Kozich Westcott (27). Briefly, 20 ng of DNA, quantified with Qubit™ (Thermo Fisher Scientific) and the Quant IT™ dsDNA HS Assay Kit were used as a template for the amplification of the V4 region (25 amplification cycles) using the AccuPrime™ Pfx SuperMix high-fidelity DNA polymerase from Invitrogen™ (Thermo Fisher Scientific). Amplicons were purified with Beckman Coulter™ AMPure XP 1.8X beads (Life Sciences) and fragment size was confirmed using the 4200 TapeStation High Sensitivity DNA Reagent Kit (Agilent Technologies). DNA concentration was re-quantified with a Qubit™ instrument and the Quant IT™ dsDNA HS Assay Kit (Thermo Fisher Scientific) and the concentration was adjusted to 4 nM. Equimolar pooling was followed by a new quantification using the Qubit™ and the Quant IT™ dsDNA HS assay kit (Thermo Fisher Scientific) and denaturation with NaOH. Then, the DNA pooling was spiked with PhiX (PhiX Control Kit v3 Illumina). Paired-end DNA sequencing was performed on a MiSeq system (Illumina) with the MiSeq Reagent Kit V3 (600-cycle) (Illumina).
2.5 Data analysis
Sequencing quality control and identification of the amplicon sequence variants (ASVs) were performed with QIIME2 and the DADA2 package (28, 29). Features with less than 11 counts across all samples as well as features that appeared in only one sample were removed. Rarefaction with a sampling depth of 21,348 was performed. The rarefied table was later transformed to relative frequencies. Alpha and beta diversity were calculated in QIIME2 and the taxonomic assignation was performed using the Greengenes database (30). Statistical distribution of the variables was evaluated with the Shapiro–Wilk test. Phenotypic variables and alpha diversity were compared using paired Student’s t test or paired Wilcoxon signed ranked test depending on the type of statistical distribution. The abundance of one species of the genera Coprococcus (taxonomic assignation by QIIME2 was g_Coprococcus;s_ but hereafter referred as Coprococcus sp.) was stratified as “higher than” or “lower than” the median abundance in pre- and postsurgery groups. BMI was compared between these stratified groups using the Wilcoxon signed ranked test. PERMANOVA was employed to assess differences in Bray–Curtis, weighted and unweighted UniFrac data.
Pre- and post-surgery bacterial abundance differences were assessed with the paired Wilcoxon signed rank test. To account for multiple testing, p-values were adjusted using False Discovery Rate (FDR). Similarly, to control for false positives, FDR-adjusted p-values were calculated in Spearman correlations analysis of bacterial abundance with energy and nutrients intakes (macro- and micronutrients).
Functional inference was performed using Picrust2 to identify enriched metabolic pathways at the species level (31). Effect size was computed using the aldex function from the ALDEx2 package in R (version 4.3.0 [2023-04–21 ucrt]). All statistical analysis and plots were generated in R.
3 Results
3.1 Phenotypic characterization
Malabsorptive bariatric surgery improved several anthropometric and biochemical parameters. On average, patients lost 28 kg, which represents approximately 24% of their initial body weight. Most patients were reclassified as having class I obesity (30 ≤ BMI < 35) instead of severely obesity (BMI ≥ 40, their initial classification). Glucose, triglycerides, LDL and total cholesterol serum levels decreased after the surgery (Table 1).
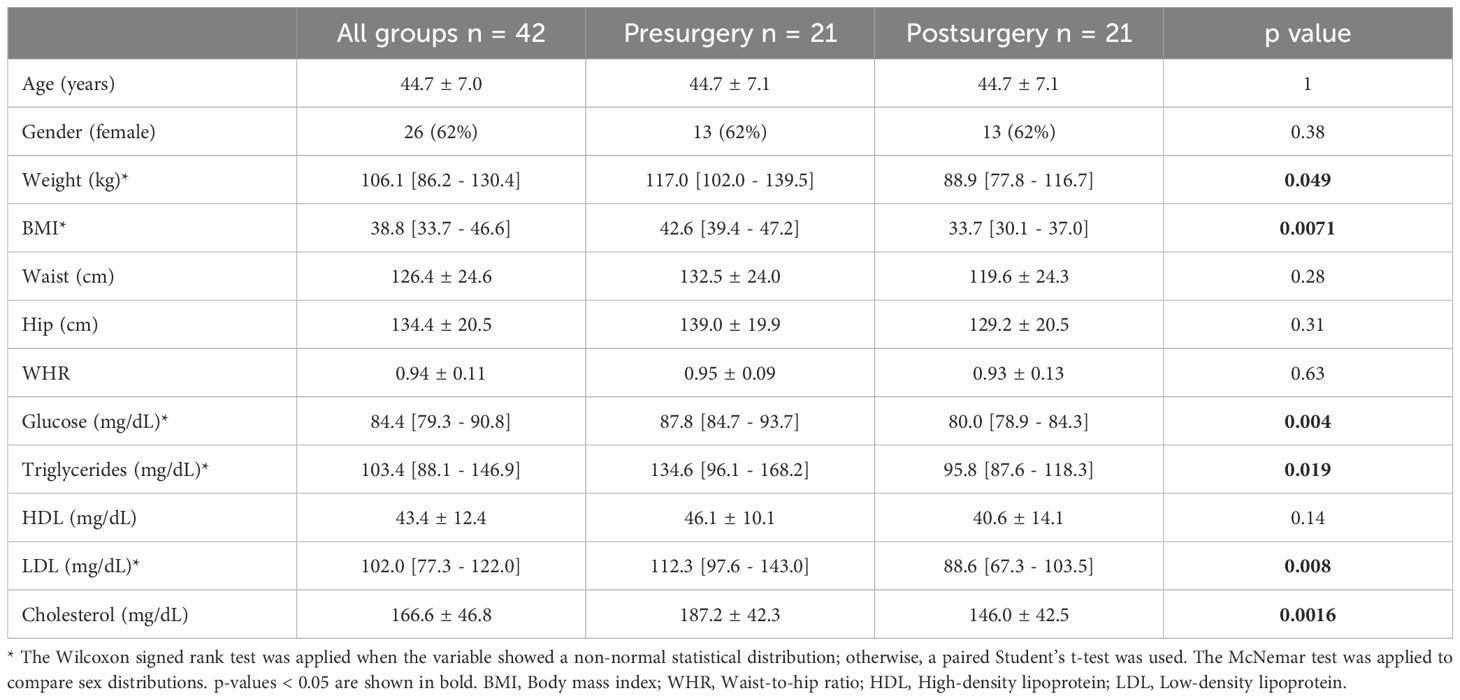
Table 1. Anthropometric and biochemical characteristics of the population.
3.2 Alpha and beta diversity
Microbiota richness assessed by the Chao1 index did not differ significantly when comparing samples collected before and after surgery (Figure 1a). However, the Shannon and Simpson indices showed significant differences between the two groups (p values of 0.0014 and 0.0029, respectively), with higher diversity index observed in the pre-surgery group (Figures 1c, e). Beta diversity was different for Bray–Curtis, weighted and unweighted UniFrac indices between the compared groups. In all three cases, the PERMANOVA p value was 0.001 (Figures 1b, d, f).
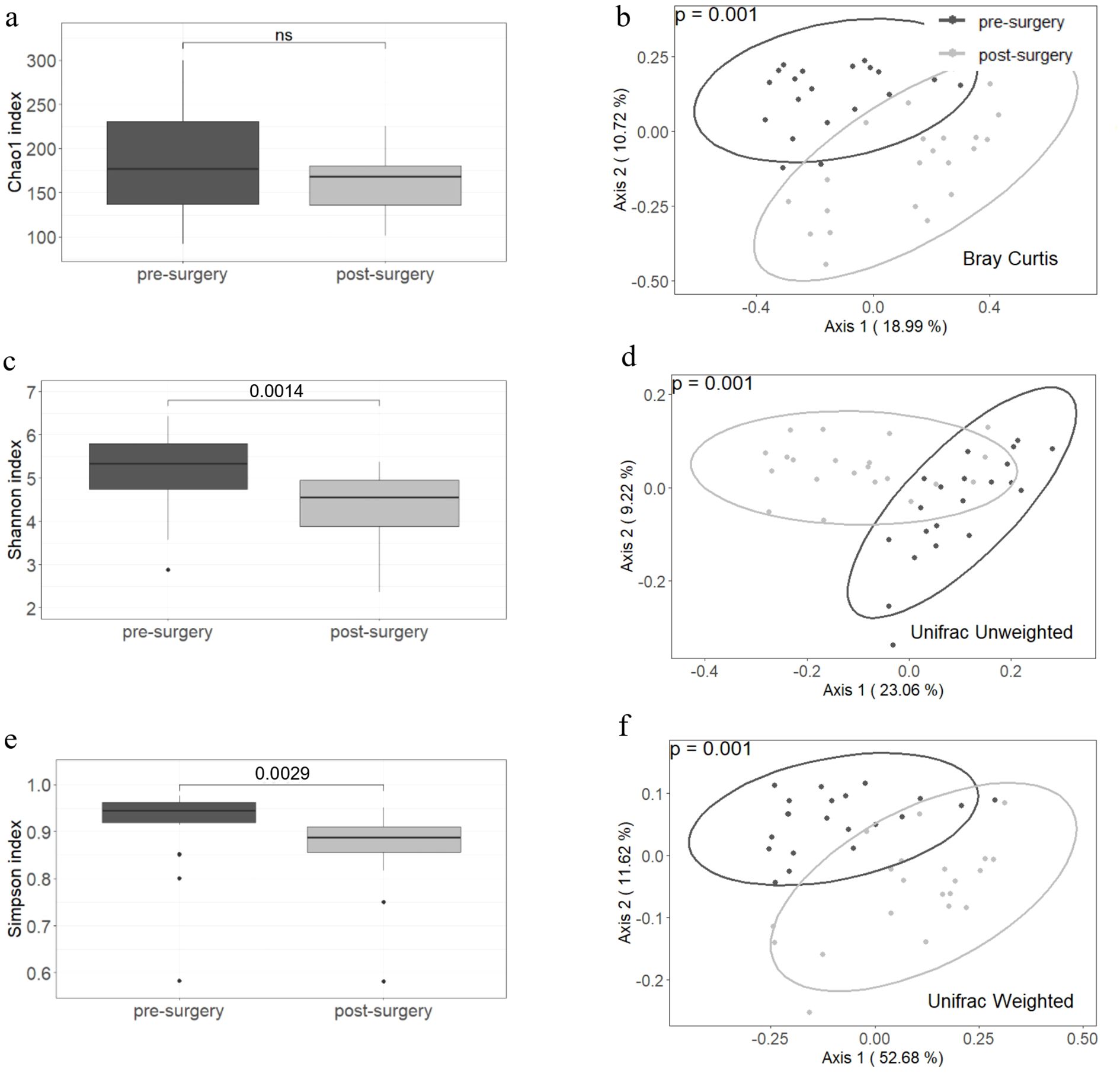
Figure 1. Alpha and beta diversity are different before and after a malabsorptive bariatric surgery. The Chao1 index (a) did not differ between groups but Shannon and Simpson indices (c, e) decreased after surgery. Beta diversity assessed by Bray-Curtis, Unweighted and Weighted Unifrac showed significant differences between pre- and post- surgery samples (b, d, f). Twenty-one individuals were included in each group. The paired Wilcoxon signed ranked-test was used to compare the alpha diversity between groups and PERMANOVA to assess statistical differences in beta diversity.
3.3 Correlation between microbiota and nutrients intake
Carbohydrates, proteins, and total energy intake showed a trend toward decreasing in the post-surgery group; however, the differences did not reach statistical significance. Magnesium and thiamin intake was positively correlated with the relative abundance of Coprococcus sp. in the post-surgery group (rho = 0.82, pFDR = 0.029; rho = 0.81, pFDR = 0.029, respectively) (Figures 2a, b) but not in obese individuals (data not shown). Further comparison of anthropometric and biochemical parameters between the low (≤ median) and high (> median) Coprococcus sp. abundance groups showed that BMI was greater in post-surgery individuals with higher Coprococcus sp. relative abundance (p = 0.036; Figure 2c). This relationship was reversed in the pre-surgery group where BMI decreased as Coprococcus sp. relative abundance increased (p = 0.043, Figure 2d). When the energy intake was compared by Coprococcus sp. abundance level, post-surgery patients in the higher-abundance group had greater energy intake (2066.4 kcal [1497.9 kcal – 2262.8 kcal] vs. 1379.6 kcal [1007.8 kcal – 1680.1 kcal], p = 0.024). No differences in energy intake was observed in the pre-surgery group.
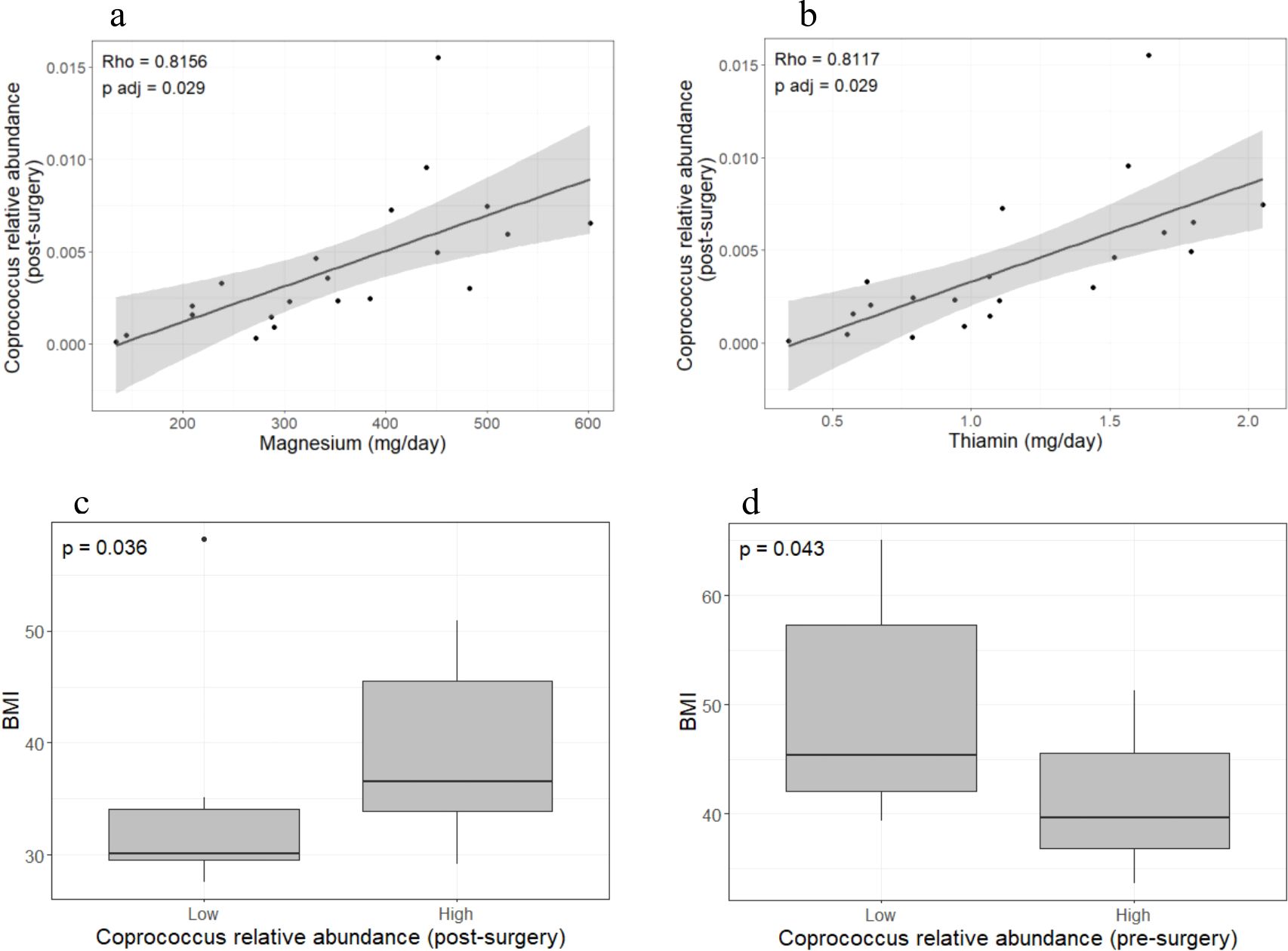
Figure 2. Coprococcus sp. abundance is related to micronutrients intake and BMI. Magnesium and thiamin intake was positively correlated with Coprococcus sp. abundance in the post-surgery group (a, b) but not in the pre-surgery group (data not shown). Patients with low Coprococcus sp. abundance (dichotomized into lower and higher quantiles) showed lower BMI in the post-surgery group (c) but the opposite was observed in the pre-surgery group (d). Twenty-one individuals were included in each group. Spearman correlation was used to assess the relationship between bacteria abundance and dietary intake. Due to the large number of bacteria and micronutrients analyzed, p-value were adjusted by FDR. Statistical differences in BMI between low- and high- abundance Coprococcus sp. groups were calculated with the Wilcoxon signed ranked-test. Analyses and plots were performed in R (version 4.3.0 [2023-04–21 ucrt]).
3.4 Changes in the microbiota before and after a bariatric surgery
Eighteen bacterial species (meeting the criteria of pFDR ≤ 0.05 and a median sequences count of 50 in the rarefied abundance data across the studied population) exhibited different relative abundances before and after a bariatric surgery (Figure 3). Most bacteria decreased in abundance after surgery (all belonging to the order Clostridiales). Only the abundance of Streptococcus, Megasphaera, Veillonella dispar and Veillonella parvula increased post-surgery. Bacteria that differed between groups belonged to the order Clostridiales and were clustered in three families: Lachnospiraceae, Ruminococcaceae and Veillonellaceae. At the phylum level, Bacteroidetes and Fusobacteria increased after surgery (0.32 [0.17-0.43] vs. 0.52 [0.42-0.60], pFDR = 0.014 and 0.0 [0.0-0.0] vs. 0.0 [0.0-0.02], pFDR = 0.002; respectively), whereas Firmicutes decreased (0.61 [0.49-0.73] vs. 0.38 [0.30-0.48], pFDR = 0.002) (Supplementary Data S1).
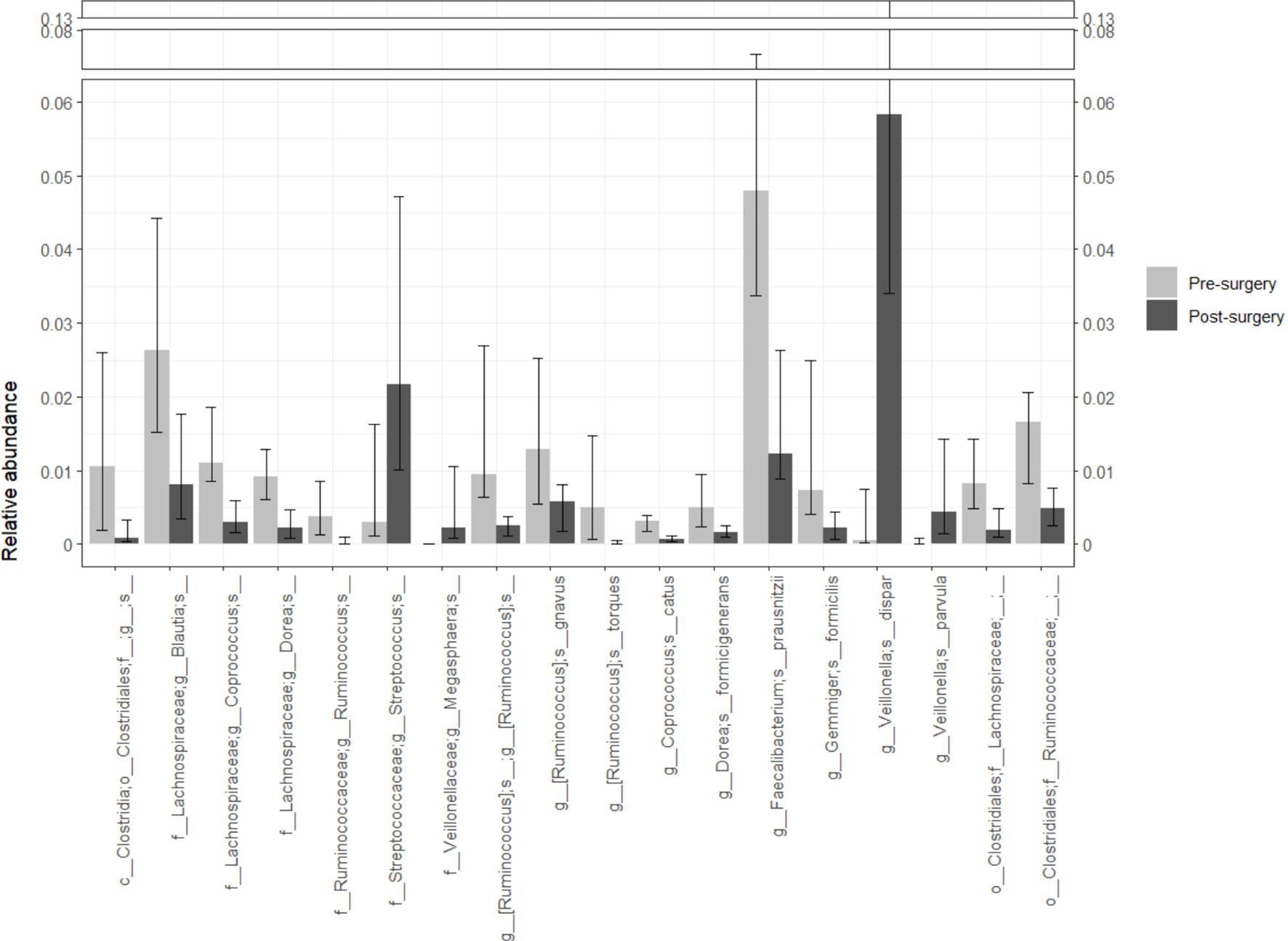
Figure 3. Bacterial abundance changes after patients underwent a malabsorptive bariatric surgery. Only bacteria with pFDR < 0.05 and a median sequence count of 50 in the rarefied abundance table are shown although the analysis included all bacteria identified by sequencing of the V4 variable region of the rDNA gene. A reduction in the abundance of most bacteria was observed. The genera Streptococcus, Megasphaera and the species Veillonella dispar and Veillonella parvula were more abundant in the post-surgery group. Statistical differences in bacterial abundance between groups were assessed using the paired Wilcoxon signed ranked-test. The p values were adjusted by FDR to control for false positives. Twenty-one individuals were included in each group. Analyses and plots were generated with R (version 4.3.0 [2023-04–21 ucrt]).
3.5 Enriched bacterial metabolic pathways
The enrichment of bacterial metabolic pathways is shown in Figure 4. In the pre-surgery group, the main enriched pathways were related to amino acid (lysine, ornithine, arginine, histidine, serine, glycine and isoleucine) and thiamin biosynthesis, sugars (sucrose, galactose, fucose, rhamnose, glucose, xylose and mannan) and starch degradation, acetate production and glycogen biosynthesis and degradation. In contrast, in the post-surgery group, pathways related to vitamin K synthesis were enriched.
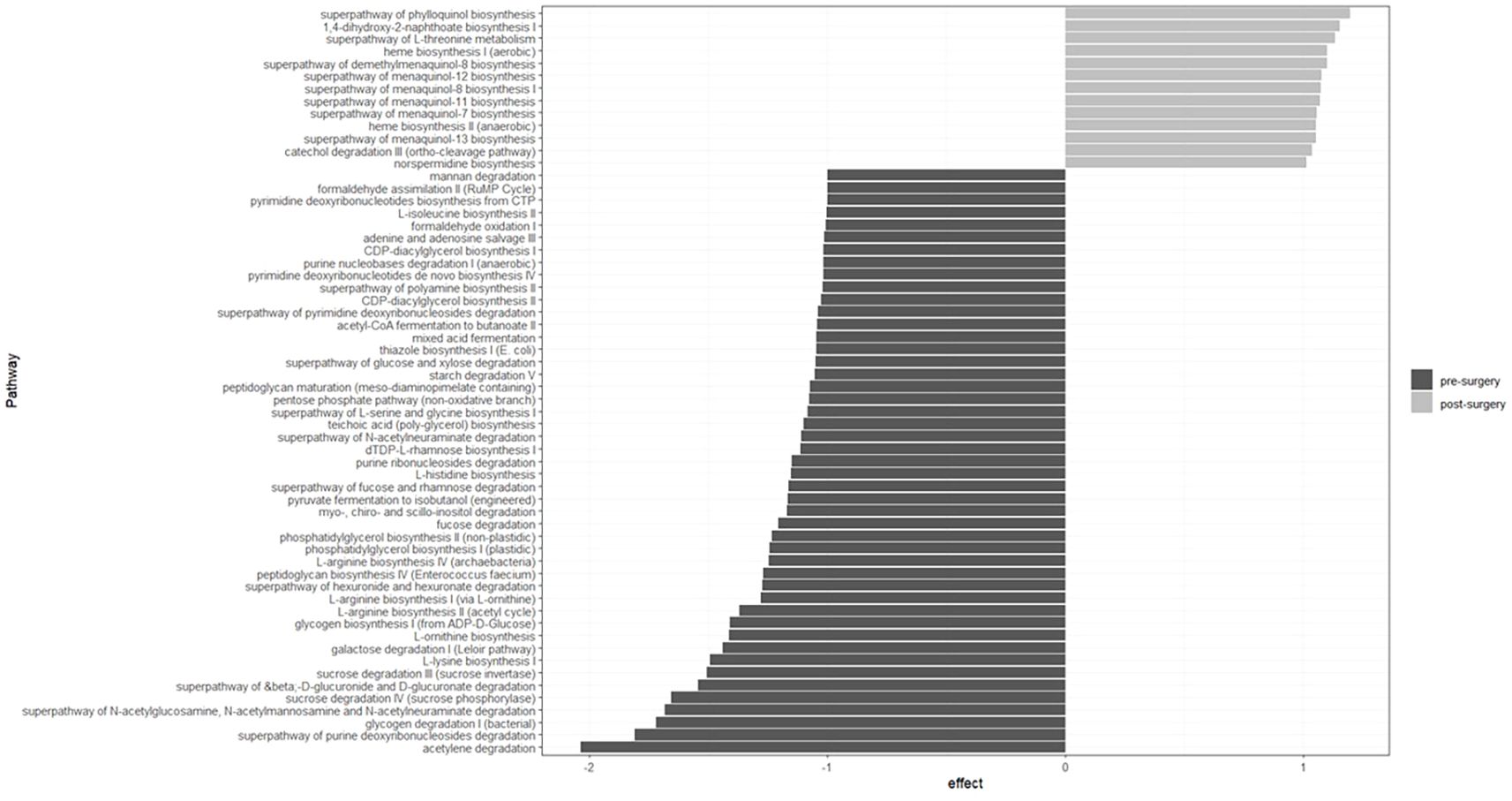
Figure 4. Bacterial metabolic pathways enriched in the pre- and post-surgery groups are related to amino acid, thiamin and K biosynthesis, sugar degradation and acetate production. Twenty-one individuals were included in each group. Metabolic pathways were predicted with Picrust2 and the effect size estimated using the ALDEx2 package in R (version 4.3.0 [2023-04–21 ucrt]).
4 Discussion
Bariatric surgery is a procedure that has been shown to substantially reduce weight and improve metabolic alterations associated with diabetes. Major remodeling of the digestive tract contributes to these changes by increasing the release of anorexigenic hormones and decreasing nutrient absorption but also by altering the gut microbiota composition (32). Indeed, our study revealed that bacterial diversity (alpha and beta) varied greatly after bariatric surgery with several species decreasing in abundance. The abundance of one of these bacteria (g_Coprococcus;s_ according to QIIME2 taxonomic assignation but referred as Coprococcus sp.) showed an opposite relationship with BMI before and after surgery and was positively correlated with magnesium and thiamin intake in the post-surgery group.
The results of this study are consistent with previously published findings showing that bariatric surgery improves patients´ health. Patients in the present study lost an average of 24% of their initial weight and they were reclassified from class III obesity to class I obesity. At baseline, 71.4% of the patients were taking some type of hypoglycemic medication, but this proportion was reduced to 14.3% after surgery. Biochemical variables such as fasting glucose, triglycerides, LDL and total cholesterol improved as well (Table 1).
Improvement in anthropometric and biochemical parameters was accompanied by a decrease in Shannon and Simpson diversity indices (Figure 1). This reduction in diversity is attributed to the extensive remodeling of the gut, which prevents the transit of food through the first portion of the small intestine, leading to incomplete digestion and overall changes in the gut microenvironment. Beta diversity estimated by Bray–Curtis, unweighted and weighted UniFrac indices showed clear differences between the compared groups (Figure 1). These results indicate that the bacterial composition is strongly modified after malabsorptive bariatric surgery. At the phylum level, the abundance of Firmicutes decreased, while that of Bacteroidetes and Fusobacteria increased (Supplementary Material S1). These findings are consistent with reports in the literature indicating that the microbiota of obese patients is enriched in Firmicutes (33). At lower taxonomic levels and after adjusting the p value by false discovery rate, we identified 18 taxa that varied in abundance (Figure 3). Most taxa decreased in abundance after bariatric surgery (14 out of 18). Taxa that decreased in abundance belonged to the order Clostridiales and the families Lachnospiraceae and Ruminococcaceae. The genomes of members of both families are known to contain ATP-binding cassette (ABC) genes and other genes encoding enzymes that participate starch and sugars degradation. Hydrolysis of these sugars results in the production of SCFAs (butyrate, acetate, and propionate) (34). Although increased SCFA production is generally considered beneficial for gut homeostasis, when combined with a greater bacterial capacity to extract energy from the diet, it may contribute to the obese phenotype. In line with this interpretation, we identified enrichment of several pathways involved in starch and sugar degradation (i.e. xylose, galactose, N-acetylmannosamine, N-acetylneuraminate, N-acetylglucosamine, mannan, glucose, rhamnose, fucose, and sucrose) in obese patients (Figure 4).
Another pathway that was enriched in the pre-surgery group was related to the acetylene degradation. The end product of this pathway is microbiota-derived acetate. Evidence indicating that acetate production by the microbiota due to high fructose consumption (which is relatively common in obese patients) leads to activation of hepatic ACSS2 enzyme which transforms acetate to lipogenic acetyl-CoA (35). This occurs in addition to the conversion of glucose and fructose to fatty acids in the liver (36). Thus, increased acetate and lipogenic acetyl-CoA production combined with high-caloric diets in obese pre-surgery patients may contribute to the maintenance of the obese phenotype.
Microbial glycogen degradation and glycogen biosynthesis pathways were enriched in the pre-surgery group as well. The enrichment of both energy-related pathways indicates that the microbiota of severely obese patients has the capacity to manage the energy surplus obtained due to the patients’ intake of high-caloric diets.
Previous reports indicate that approximately 27% of patients undergoing bariatric surgery have thiamin deficiency (37). This is clinically relevant because these patients require lifelong vitamin B1 (thiamin) supplementation. Thiamin is crucial and indirectly involved in the electron transport chain and extraction of energy from carbohydrates but also is required for bacterial growth (38). This essential vitamin is obtained from the diet and as a product of bacterial metabolism. However, not all bacteria have the genes necessary for its de novo synthesis (39). In this study, we found that the abundance of three bacteria (Faecalibacterium prausnitzii, Gemmigers formicilis and Ruminococcus) that require thiamine for their survival but cannot produce thiamine by themselves decreased (Figure 3). These bacteria belong to the Ruminococcaceae family (39). This abundance decrease corresponds to a reduction of the thiazole biosynthesis pathway after the surgery (Figure 4).
Although thiamin is required for normal physiological processes, our results suggest that fine tuning of thiamin intake is desirable in post-surgery patients. We observed that thiamin intake was positively correlated with Coprococcus sp. abundance which in turn was related to higher BMI after surgery (Figures 2b, c). We further observed that the Coprococcus sp. relative abundance in the post-surgery group was positively correlated with magnesium intake (Figure 2a) which is required for the proper activation of vitamin B1 (20, 40). The above-mentioned evidence suggest that thiamin and magnesium intake must be such that it does not have a detrimental effect on BMI mediated by Coprococcus sp. According to the recommended dietary allowance (RDA), 1.1 - 1.2 mg of thiamin and 320–420 mg of magnesium intake are required to maintain normal physiological processes (41, 42). These RDAs values fall near the midpoint of the range of micronutrient intakes quantified from the dietary recall questionnaires of post-surgery patients (Figures 3a, b). Thus, patients whose intake exceeds the RDA may benefit from reducing their daily thiamin and magnesium intake to levels that are closer to the RDA. Future well-controlled studies to explore the feasibility of diet-driven modulation of the Coprococcus sp. abundance in order to enhance weight loss and augment the metabolic benefits of bariatric surgery are needed. In addition, a better understanding of thiamin consumption and/or production by the Coprococcus sp. is necessary as a study by Soto-Martin et al. (43) reported that not all Coprococcus spp. have either the presence of the genetic repertoire for de novo thiamin biosynthesis or show the same growth rate in thiamin-depleted medium. Isolation of Coprococcus sp. from post-bariatric surgery samples followed by in vitro and in silico analysis of its capacity to produce or use thiamin as well as whole-genome sequencing are necessary to clarify the identified positive thiamin-Coprococcus sp.-BMI association.
Interestingly, a negative relationship between Coprococcus sp. abundance and BMI was observed in obese patients before the bariatric surgery (Figure 2d). This is in contrast to the positive relationship found in post-surgery patients. Lozano et al. (44) provided insight into a possible mechanism underlying this opposite relationship. They reported a positive association between high inflammatory index diets and ectopic fat accumulation, but only in patients with lower gut Coprococcus abundance. Obese individuals typically consume high-inflammatory-index diets with high caloric content. Thus, obese patients in this study with lower Coprococuss sp. abundance may be more prone to accumulate ectopic fat and to have a higher BMI. Post-surgery patients follow less inflammatory diets which indicates that this mechanism does not affect them. However, other mechanisms may play a role as determinants of weight loss. For instance, Coprococcuss spp. are SCFA producers with the capacity to extract energy from otherwise undigested diet components (43). Therefore, in the altered post-surgical gut environment, greater Coprococcus sp. abundance combined with a less diverse microbiota and high metabolic efficiency for extracting energy from limited substrates would make it more difficult for patients to lose the obese phenotype. No correlation between thiamin/magnesium intake and Coprococcus sp. abundance was found in severely obese individuals.
The role of Coprococcus in health remains a subject of debate. While a large portion of the scientific literature associates this genus with beneficial traits such as improved insulin sensitivity in metabolically healthy obese individuals (7–9), reduced inflammation, enhance anti-inflammatory molecules, mucin production and tight junctions restoration (10, 11), other studies linked it to with small intestine bacterial overgrowth (12), depressive symptoms (13), inflammatory bowel disease (14), liver tumorigenesis (15) and interstitial cystitis (16). The gut microbiota is a diverse and complex community in which bacterial growth is tightly linked to the available resources. Thus, divergent associations with health and disease may result from differences in microenvironments, diseases and bacteria-bacteria interactions. The findings of this study should be interpreted in light of certain limitations. First, although false discovery rate adjustment was applied in multiple comparison hypotheses testing to control for false positives, replication in larger, independent cohorts from diverse geographical locations is needed to strengthen the conclusions. Second, major intestinal rearrangement following bariatric surgery has been reported as the main driver of most of the changes in the gut microbiota profile (45). However, the contribution of other confounders not evaluated in this study such as medication use and lifestyle modifications should not be excluded and warrants further investigation.
5 Conclusion
Important changes in clinical variables and microbiota were observed after individuals with severe obesity underwent malabsorptive bariatric surgery. Only 20% of patients who used hypoglycemic medication before surgery continued doing so afterwards, while the remaining 80% did not longer required medication to control their glucose levels. This improvement was accompanied by changes in the gut microbiota alpha and beta diversity. After surgery, the gut microbiota was less diverse, with a decrease in the abundance of several bacterial species. These changes in the microbiota profile led to a reduction in the enrichment of metabolic pathways involved in amino acid, thiamin, sugar degradation and acetate production. The abundance of a species of the genus Coprococcus showed opposite relationships with BMI in pre-surgery and post-surgery individuals. Furthermore, its abundance correlated with magnesium and thiamin intake in individuals who underwent a malabsorptive bariatric surgery.
Data availability statement
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found below: https://www.ncbi.nlm.nih.gov/, PRJNA1181870.
Ethics statement
The studies involving humans were approved by Comite Nacional de Investigación Científica del Instituto Mexicano del Seguro Social, México (approval no. R-2017-785-063 and R-2020-785-016). The studies were conducted in accordance with the local legislation and institutional requirements. The participants provided their written informed consent to participate in this study.
Author contributions
FS-S: Formal Analysis, Conceptualization, Writing – review & editing, Supervision, Writing – original draft, Investigation. EP-R: Formal Analysis, Writing – review & editing. CR-S: Formal Analysis, Writing – review & editing. MM-A: Resources, Writing – review & editing. SR-G: Writing – review & editing, Methodology. LL-S: Writing – review & editing, Formal Analysis, Writing – original draft. LG-M: Methodology, Writing – review & editing. AR-G: Writing – review & editing, Resources. CM-O: Resources, Writing – review & editing. LM-T: Writing – review & editing, Methodology. MC-L: Writing – review & editing, Supervision, Resources.
Funding
The author(s) declare financial support was received for the research and/or publication of this article. This work was supported by the National Council of Science and Technology (CONACYT-México) with the grant SALUD-2017-C02-289961. CONACYT grants to Evelyn Pérez-Ruiz: 368994.
Acknowledgments
We thank Iliana Avila Soto and Sofia Barragan for their assistance during the application and nutrient calculations from the diet questionnaires as well as Carolina González and Javier Gaytán from the Sequecing Laborary, Centro de Instrumentos, División de Desarrollo de la Investigación, Instituto Mexicano del Seguro Social, for their technical assistance during the bacterial DNA sequencing.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fendo.2025.1613221/full#supplementary-material
References
1. Perdomo CM, Cohen RV, Sumithran P, Clément K, and Frühbeck G. Contemporary medical, device, and surgical therapies for obesity in adults. Lancet. (2023) 401:1116–30. doi: 10.1016/s0140-6736(22)02403-5
2. Purnell JQ. Definitions, classification, and epidemiology of obesity. In: Feingold KR, Anawalt B, Blackman MR, Boyce A, Chrousos G, et al, editors. Endotext. 2023 MDText.com, Inc, South Dartmouth (MA). Available online at: https://www.ncbi.nlm.nih.gov/books/NBK278943/.
3. Arterburn DE, Telem DA, Kushner RF, and Courcoulas AP. Benefits and risks of bariatric surgery in adults: A review. JAMA. (2020) 324:879–87. doi: 10.1001/jama.2020.12567
4. Robert M, Espalieu P, Pelascini E, Caiazzo R, Sterkers A, Khamphommala L, et al. Efficacy and safety of one anastomosis gastric bypass versus roux-en-Y gastric bypass for obesity (Yomega): A multicentre, randomised, open-label, non-inferiority trial. Lancet. (2019) 393:1299–309. doi: 10.1016/s0140-6736(19)30475-1
5. Fouladi F, Carroll IM, Sharpton TJ, Bulik-Sullivan E, Heinberg L, Steffen KJ, et al. A microbial signature following bariatric surgery is robustly consistent across multiple cohorts. Gut Microbes. (2021) 13:1930872. doi: 10.1080/19490976.2021.1930872
6. Cerdó T, García-Santos JA M, and Campoy C. The role of probiotics and prebiotics in the prevention and treatment of obesity. Nutrients. (2019) 11:635. doi: 10.3390/nu11030635
7. Cui J, Ramesh G, Wu M, Jensen ET, Crago O, Bertoni AG, et al. Butyrate-producing bacteria and insulin homeostasis: the microbiome and insulin longitudinal evaluation study (Miles). Diabetes. (2022) 71:2438–46. doi: 10.2337/db22-0168
8. Zhang M, Liu J, Li C, Gao J, Xu C, Wu X, et al. Functional fiber reduces mice obesity by regulating intestinal microbiota. Nutrients. (2022) 14:2676. doi: 10.3390/nu14132676
9. Olivares P, Pacheco ABF, Aranha LN, Oliveira BDS, Santos AA, Santos P, et al. Gut microbiota of adults with different metabolic phenotypes. Nutrition. (2021) 90:111293. doi: 10.1016/j.nut.2021.111293
10. Huang X, Qiu Y, Gao Y, Zhou R, Hu Q, He Z, et al. Gut microbiota mediate melatonin signalling in association with type 2 diabetes. Diabetologia. (2022) 65:1627–41. doi: 10.1007/s00125-022-05747-w
11. Yang R, Shan S, Shi J, Li H, An N, Li S, et al. Coprococcus eutactus, a potent probiotic, alleviates colitis via acetate-mediated iga response and microbiota restoration. J Agric Food Chem. (2023) 71:3273–84. doi: 10.1021/acs.jafc.2c06697
12. Guo H, Chen Y, Dong W, Lu S, Du Y, and Duan L. Fecal coprococcus, hidden behind abdominal symptoms in patients with small intestinal bacterial overgrowth. J Transl Med. (2024) 22:496. doi: 10.1186/s12967-024-05316-2
13. Radjabzadeh D, Bosch JA, Uitterlinden AG, Zwinderman AH, Ikram MA, van Meurs JBJ, et al. Gut microbiome-wide association study of depressive symptoms. Nat Commun. (2022) 13:7128. doi: 10.1038/s41467-022-34502-3
14. Liu B, Ye D, Yang H, Song J, Sun X, Mao Y, et al. Two-sample mendelian randomization analysis investigates causal associations between gut microbial genera and inflammatory bowel disease, and specificity causal associations in ulcerative colitis or crohn's disease. Front Immunol. (2022) 13:921546. doi: 10.3389/fimmu.2022.921546
15. Pedersen KB, Pulliam CF, Patel A, Del Piero F, Watanabe TTN, Wankhade UD, et al. Liver tumorigenesis is promoted by a high saturated fat diet specifically in male mice and is associated with hepatic expression of the proto-oncogene agap2 and enrichment of the intestinal microbiome with coprococcus. Carcinogenesis. (2019) 40:349–59. doi: 10.1093/carcin/bgy141
16. Fu C, Zhao Y, Zhou X, Lv J, Jin S, Zhou Y, et al. Gut microbiota and interstitial cystitis: exploring the gut-bladder axis through mendelian randomization, biological annotation and bulk rna sequencing. Front Immunol. (2024) 15:1395580. doi: 10.3389/fimmu.2024.1395580
17. Barra NG, Anhê FF, Cavallari JF, Singh AM, Chan DY, and Schertzer JD. Micronutrients impact the gut microbiota and blood glucose. J Endocrinol. (2021) 250:R1–R21. doi: 10.1530/joe-21-0081
18. Costliow ZA and Degnan PH. Thiamine acquisition strategies impact metabolism and competition in the gut microbe bacteroides thetaiotaomicron. mSystems. (2017) 2:e00116–17. doi: 10.1128/mSystems.00116-17
19. Ferenc K, Sokal-Dembowska A, Helma K, Motyka E, Jarmakiewicz-Czaja S, and Filip R. Modulation of the gut microbiota by nutrition and its relationship to epigenetics. Int J Mol Sci. (2024) 25:1228. doi: 10.3390/ijms25021228
20. Piuri G, Zocchi M, Della Porta M, Ficara V, Manoni M, Zuccotti GV, et al. Magnesium in obesity, metabolic syndrome, and type 2 diabetes. Nutrients. (2021) 13:320. doi: 10.3390/nu13020320
21. Shamah- Levy T V-HS and Rivera-Dommarco J. Manual de procedimientos para proyectos de nutrición. Cuernavaca México Instituto Nacional Salud Pública. (2006). Available online at: https://salud.gob.mx/unidades/cdi/documentos/proy_nutricion.pdf?utm_source=chatgpt.com.
22. Denova-Gutiérrez E, Ramírez-Silva I, Rodríguez-Ramírez S, Jiménez-Aguilar A, Shamah-Levy T, Rivera-Dommarco JA, et al. Validity of a food frequency questionnaire to assess food intake in Mexican adolescent and adult population. Salud Publica Mex. (2016) 58:617–28. doi: 10.21149/spm.v58i6.7862
23. Ramírez-Silva I, Jiménez-Aguilar A, Valenzuela-Bravo D, Martinez-Tapia B, Rodríguez-Ramírez S, Gaona-Pineda EB, et al. Methodology for estimating dietary data from the semi-quantitative food frequency questionnaire of the mexican national health and nutrition survey 2012. Salud Publica Mex. (2016) 58:629–38. doi: 10.21149/spm.v58i6.7974
24. Ramírez Silva I, Barragán-Vázquez S, Rodríguez Ramírez S, Rivera Dommarco JA, Mejía-Rodríguez F, Barquera Cervera S, et al. Base De Alimentos De México (Bam): Compilación de la Composición De Los Alimentos Frecuentemente Consumidos En El País. (2021) Version 18:1.1. Available online at: https://insp.mx/informacion-relevante/bam-bienvenida?utm_source=chatgpt.com
25. Bognar A. Tables of weight yield of food and retention factors of food constituents for the cal-culation of nutrient composition of cooked foods (Dishes). (2002). Karlsruhe: Bundesforschungsanstalt für Ernährung. (Berichte der Bundesforschungsanstalt für Ernährung, BFE-R-02-03). doi: 5445/IR/9642002
26. Nutrient Data LaboratoryTable of Nutrient Retention Factors. Release 6. U.S. Department of Agriculture, Agricultural Research Service. Beltsville, MD. (2007). Available online at: https://www.ars.usda.gov/ARSUserFiles/80400525/Data/retn/retn06.pdf.
27. Kozich JJ, Westcott SL, Baxter NT, Highlander SK, and Schloss PD. Development of a dual-index sequencing strategy and curation pipeline for analyzing amplicon sequence data on the miseq illumina sequencing platform. Appl Environ Microbiol. (2013) 79:5112–20. doi: 10.1128/aem.01043-13
28. Bolyen E, Rideout JR, Dillon MR, Bokulich NA, Abnet CC, Al-Ghalith GA, et al. Reproducible, interactive, scalable and extensible microbiome data science using qiime 2. Nat Biotechnol. (2019) 37:852–7. doi: 10.1038/s41587-019-0209-9
29. Callahan BJ, McMurdie PJ, Rosen MJ, Han AW, Johnson AJ, and Holmes SP. Dada2: high-resolution sample inference from illumina amplicon data. Nat Methods. (2016) 13:581–3. doi: 10.1038/nmeth.3869
30. McDonald D, Price MN, Goodrich J, Nawrocki EP, DeSantis TZ, Probst A, et al. An improved greengenes taxonomy with explicit ranks for ecological and evolutionary analyses of bacteria and archaea. Isme J. (2012) 6:610–8. doi: 10.1038/ismej.2011.139
31. Douglas GM, Maffei VJ, Zaneveld JR, Yurgel SN, Brown JR, Taylor CM, et al. Picrust2 for prediction of metagenome functions. Nat Biotechnol. (2020) 38:685–8. doi: 10.1038/s41587-020-0548-6
32. Madsbad S, Dirksen C, and Holst JJ. Mechanisms of changes in glucose metabolism and bodyweight after bariatric surgery. Lancet Diabetes Endocrinol. (2014) 2:152–64. doi: 10.1016/s2213-8587(13)70218-3
33. Indiani C, Rizzardi KF, Castelo PM, Ferraz LFC, Darrieux M, and Parisotto TM. Childhood obesity and firmicutes/bacteroidetes ratio in the gut microbiota: A systematic review. Child Obes. (2018) 14:501–9. doi: 10.1089/chi.2018.0040
34. Biddle A, Stewart L, Blanchard J, and Leschine S. Untangling the genetic basis of fibrolytic specialization by lachnospiraceae and ruminococcaceae in diverse gut communities. Diversity. (2013) 5:627–40. doi: 10.3390/d5030627
35. Zhao S, Jang C, Liu J, Uehara K, Gilbert M, Izzo L, et al. Dietary fructose feeds hepatic lipogenesis via microbiota-derived acetate. Nature. (2020) 579:586–91. doi: 10.1038/s41586-020-2101-7
36. Softic S, Gupta MK, Wang GX, Fujisaka S, O'Neill BT, Rao TN, et al. Divergent effects of glucose and fructose on hepatic lipogenesis and insulin signaling. J Clin Invest. (2017) 127:4059–74. doi: 10.1172/jci94585
37. Bahardoust M, Eghbali F, Shahmiri SS, Alijanpour A, Yarigholi F, Valizadeh R, et al. B1 vitamin deficiency after bariatric surgery, prevalence, and symptoms: A systematic review and meta-analysis. Obes Surg. (2022) 32:3104–12. doi: 10.1007/s11695-022-06178-7
38. Manzetti S, Zhang J, and van der Spoel D. Thiamin function, metabolism, uptake, and transport. Biochem (Mosc). (2014) 53:821–35. doi: 10.1021/bi401618y
39. Park J, Hosomi K, Kawashima H, Chen YA, Mohsen A, Ohno H, et al. Dietary vitamin B1 intake influences gut microbial community and the consequent production of short-chain fatty acids. Nutrients. (2022) 14. doi: 10.3390/nu14102078
40. LeBlanc JG, Chain F, Martín R, Bermúdez-Humarán LG, Courau S, and Langella P. Beneficial effects on host energy metabolism of short-chain fatty acids and vitamins produced by commensal and probiotic bacteria. Microbial Cell factories. (2017) 16:79. doi: 10.1186/s12934-017-0691-z
41. Institute of Medicine Standing Committee on the Scientific Evaluation of Dietary Reference I. The national academies collection: reports funded by national institutes of health. In: Dietary reference intakes for calcium, phosphorus, magnesium, vitamin D, and fluoride. (1997) (US, Washington DC): National Academies Press. doi: 10.17226/5776
42. Institute of Medicine (US) Standing Committee on the Scientific Evaluation of Dietary Reference Intakes and its Panel on Folate, Other B Vitamins, and Choline. In: Dietary Reference Intakes for Thiamin, Riboflavin, Niacin, Vitamin B6, Folate, Vitamin B12, Pantothenic Acid, Biotin, and Choline. (1998) (Washington DC): National Academies Press (US). doi: 10.17226/6015
43. Soto-Martin EC, Warnke I, Farquharson FM, Christodoulou M, Horgan G, Derrien M, et al. Vitamin biosynthesis by human gut butyrate-producing bacteria and cross-feeding in synthetic microbial communities. mBio. (2020) 11. doi: 10.1128/mBio.00886-20
44. Lozano CP, Wilkens LR, Shvetsov YB, Maskarinec G, Park SY, Shepherd JA, et al. Associations of the dietary inflammatory index with total adiposity and ectopic fat through the gut microbiota, lps, and C-reactive protein in the multiethnic cohort-adiposity phenotype study. Am J Clin Nutr. (2022) 115:1344–56. doi: 10.1093/ajcn/nqab398
45. Sánchez-Alcoholado L, Gutiérrez-Repiso C, Gómez-Pérez AM, García-Fuentes E, Tinahones FJ, and Moreno-Indias I. Gut microbiota adaptation after weight loss by roux-en-Y gastric bypass or sleeve gastrectomy bariatric surgeries. Surg Obes related diseases: Off J Am Soc Bariatric Surg. (2019) 15:1888–95. doi: 10.1016/j.soard.2019.08.551
Keywords: magnesium, thiamin, Coprococcus, bariatric surgery, acetate, microbiota
Citation: Suárez-Sánchez F, Pérez-Ruiz E, Ramírez-Silva CI, Molina-Ayala MA, Rivera-Gutiérrez S, León-Solís L, García-Morales L, Rodríguez-González A, Martínez-Ortiz C, Meneses-Tapia LA and Cruz-López M (2025) A species of Coprococcus is related to BMI in patients who underwent malabsorptive bariatric surgery and its abundance is modified by magnesium and thiamin intake. Front. Endocrinol. 16:1613221. doi: 10.3389/fendo.2025.1613221
Received: 16 April 2025; Accepted: 22 October 2025;
Published: 06 November 2025.
Edited by:
Lara Costantini, University of Tuscia, ItalyReviewed by:
Francesco Pizza, ASL Napoli 2 Nord, ItalyMatthew Vincent, Washington University in St. Louis, United States
Copyright © 2025 Suárez-Sánchez, Pérez-Ruiz, Ramírez-Silva, Molina-Ayala, Rivera-Gutiérrez, León-Solís, García-Morales, Rodríguez-González, Martínez-Ortiz, Meneses-Tapia and Cruz-López. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Fernando Suárez-Sánchez, ZnMuYmlvcS5pbXNzQGdtYWlsLmNvbQ==