 Luis M. Molinos-Albert
Luis M. Molinos-Albert Bonaventura Clotet1,2
Bonaventura Clotet1,2 Julià Blanco
Julià Blanco- 1IrsiCaixa AIDS Research Institute, Institut de Recerca Germans Trias i Pujol (IGTP), Germans Trias i Pujol University Hospital, Barcelona, Spain
- 2Universitat de Vic – Universitat Central de Catalunya, Barcelona, Spain
Broadly neutralizing antibodies (bNAbs) targeting conserved regions within the human immunodeficiency virus type-1 (HIV-1) envelope glycoprotein (Env) can be generated by the human immune system and their elicitation by vaccination will be a key point to protect against the wide range of viral diversity. The membrane proximal external region (MPER) is a highly conserved region within the Env gp41 subunit, plays a major role in membrane fusion and is targeted by naturally induced bNAbs. Therefore, the MPER is considered as an attractive vaccine target. However, despite many attempts to design MPER-based immunogens, further study is still needed to understand its structural complexity, its amphiphilic feature, and its limited accessibility by steric hindrance. These particular features compromise the development of MPER-specific neutralizing responses during natural infection and limit the number of bNAbs isolated against this region, as compared with other HIV-1 vulnerability sites, and represent additional hurdles for immunogen development. Nevertheless, the analysis of MPER humoral responses elicited during natural infection as well as the MPER bNAbs isolated to date highlight that the human immune system is capable of generating MPER protective antibodies. Here, we discuss the recent advances describing the immunologic and biochemical features that make the MPER a unique HIV-1 vulnerability site, the different strategies to generate MPER-neutralizing antibodies in immunization protocols and point the importance of extending our knowledge toward new MPER epitopes by the isolation of novel monoclonal antibodies. This will be crucial for the redesign of immunogens able to skip non-neutralizing MPER determinants.
Introduction
An Apparently Easy Vaccine Target
The human immunodeficiency virus type-1 (HIV-1) envelope glycoprotein (Env) is the sole viral antigen exposed on the virion surface. Env is synthetized as a precursor gp160 glycoprotein that will yield after cleavage a mature complex constituted by the non-covalent association of three gp120 (surface) and three gp41 (transmembrane) subunits, resulting in a heavily glycosylated trimer of heterodimers (1–5). Env determines the process of HIV-1 entry into the target cell that will lead to the fusion of the viral and host cell membranes (6). This process initiates with the high affinity interaction between gp120 and the CD4 molecule on the surface of target cells. This interaction promotes a series of conformational changes that transiently expose the gp120 coreceptor binding site allowing the gp120 attachment to the CCR5 or CXCR4 chemokine receptors (7–9). Coreceptor ligation triggers structural rearrangements in gp41 that permit the initiation of viral fusion. The gp41 fusion peptide (FP) inserts into the target cell membrane accounting for a short-life prehairpin fusion intermediate in which both cellular and viral membranes are connected by an extended conformation of gp41. Next, alpha-helical domains HR1 and HR2 of each gp41 monomer are folded back together to generate a 6-helix bundle conformation that brings both target cell and viral membranes closer to finally produce the membrane merge (10, 11). During this process both FP and the membrane proximal external region (MPER) play a crucial role in membrane destabilization (12).
Given its exposure on the virion surface and its role in viral infectivity, Env is the main target of HIV-1 protective humoral responses. The elicitation of Env broadly neutralizing antibodies (bNAbs), defined as those capable of neutralize the wide viral diversity, is one of the main goals for a successful HIV-1 vaccine (13). The notion that the human immune system is capable of producing HIV-1 bNAbs has been established by two pieces of evidence: (i) the identification of such immune responses in sera from HIV-1 infected individuals and (ii) the isolation of monoclonal bNAbs from these individuals (14, 15). These naturally induced bNAbs allowed the identification of conserved Env regions that helped researchers to delineate an HIV-1 Env vulnerability map. The study of bNAbs and the epitopes targeted by them are contributing enormously to our understanding of the HIV-1 humoral response as well as to the rational design of immunogen candidates (14, 16).
Whereas a big collection of bNAbs against gp120 Env subunit has been generated, a limited number has been also isolated against the less exposed gp41 subunit (17). Although neutralizing antibodies targeting the HR1 alpha-helical region have been described (18–20), the MPER is the major gp41 neutralizing determinant (21, 22). This highly conserved and unusual tryptophan-rich motif is located adjacent to the viral membrane, covering the last C-terminal residues of the gp41 ectodomain (aa 660–683, HXB2 numbering) and connects the extracellular portion of Env with the TM domain (23, 24). The importance of the MPER on Env functionality was highlighted by analysis of mutant viruses containing deletions, insertions or substitutions within this region (24–26). Substitution of the five MPER conserved tryptophan residues dramatically compromised the incorporation of gp41 into virions and, thus, blocked viral entry (24). Moreover, simple deletion of the W666-I682 spanning sequence completely abolished syncytium formation (27). These observations indicated that the MPER plays a major role in the HIV-1 Env-mediated fusion and viral infectivity, which is consistent with the high level of sequence conservation (23). The functional implications in viral infectivity, the high level of conservation and the lack of N-linked glycosylated residues, together with the discovery of potent and/or bNAbs targeting linear MPER sequences (2F5, 4E10, 10E8), all able to protect against viral challenge in non-human primates (NHP) (28–30), points that the elicitation of MPER-specific neutralizing responses by immunogen candidates is highly desirable (21, 22, 31). In addition, the MPER has a role in HIV-1 CD4-independent viral transcytosis at the epithelial barrier (32), where the conserved 662ELDKWA667 gp41 sequence interacts with galactosyl ceramide receptors (33). Secretory IgA from cervicovaginal secretions of HIV-1 infected individuals are capable of blocking viral transcytosis via 662ELDKWA667 sequence binding (34).
The MPER presents some immunological, physical, and structural, properties that impact directly on its immunogenicity, explaining the lower MPER neutralizing response of HIV-1 infected individuals comparing with other Env vulnerability regions (35, 36). Those include steric hindrance by gp120 and high hydrophobicity that makes the MPER to be partially embedded within the viral membrane (37). Structurally, the information regarding the native conformation of the MPER within the Env trimer is still limited (5), adding the challenge of developing an immunogen against a structurally ambiguous epitope. Finally, MPER-specific bNAbs show reactivity against self-antigens and host tolerance mechanisms have been suggested to influence the elicitation of MPER neutralizing responses (38).
Here, we discuss the properties that make the MPER both a unique as well as a challenging HIV-1 vaccine target; we review the MPER immune response during natural infection, the particular features of MPER bNAbs isolated and the different attempts to generate MPER-specific neutralizing antibodies by immunization within the last years. Although the results reflect a generalized failure, new insights into our knowledge have been achieved. The fact that other Env vulnerability sites have followed a similar path supports the notion that the MPER is still an HIV-1 vaccine target worth exploring (31).
Isolation of MPER Neutralizing Antibodies
The strongest evidence supporting that the human immune system can develop a potent neutralizing MPER-specific response results from the isolation of monoclonal antibodies from HIV-infected individuals. From the naturally induced 2F5, 4E10, 10E8, z13, m66.6, and CH12 antibodies identified, three of them (2F5, 4E10, and 10E8) display a broadly neutralizing activity (28, 39–46). 2F5 and 4E10 are among the first HIV-1 bNAbs discovered. They were generated by electrofusion of peripheral blood mononuclear cells mixtures from different HIV-1 infected individuals (47). 2F5 targets the linear sequence 662ELDKWA667 (39) within the N-terminal moiety of the MPER, where the central core 664DKW666 is essential for neutralization, as demonstrated by alanine-scanning mutagenesis assays (48). 2F5 has a relatively high potency and was found to neutralize 57–67% of the viral isolates tested with an IC50 below 50 µg/mL (42, 49). However, HIV-1 subtype C viruses are usually 2F5-resistant due to a mutation in the central core epitope (DSW instead of DKW) (49–51). 4E10 targets the distal conserved tryptophan rich moiety located C-terminal to the 2F5 epitope which includes the sequence 671NWFDIT676 and is extended toward C-terminal residues, where W672, F673, I675, T676, L679, and W680 have the most significant contacts with the antibody (43). Although presenting a moderate potency, 4E10 displays a remarkable breadth against 98–100% of the viral isolates, depending of the panel tested, with an IC50 below 50 µg/mL (49, 52). Further characterization of 2F5 and 4E10 antibodies has shown reduced potency of both antibodies, against transmitted-founder viruses (T/F IMC) or against replicating viruses obtained from primary lymphocytes when compared with pseudovirus obtained in 293 T cells (53–56). Despite these potential limitations, both 2F5 and 4E10 were shown to protect against viral challenge in NHP (28, 29) and their administration into human recipients showed no major clinical complications (57).
In order to delineate a complete map of HIV-1 neutralizing determinants, starting in 2009, a substantial effort has been made on the isolation of new bNAbs. The development of high-throughput analysis of single memory B cells and the use of fluorescently labeled Env-based protein probes to isolate antigen specific B cells (58–60) contributed enormously to the discovery of new HIV-1 neutralizing antibodies. In this context, the discovery in 2012 of the monoclonal antibody 10E8 recovered the interest toward the MPER region as a major vaccine target (42). 10E8 neutralized 98% of a panel of 181 pseudovirus with an IC50 below 50 µg/mL, showing a mean IC50 for sensitive viruses of 0.25 µg/mL, whereas mean IC50 values for 4E10 and 2F5 were 1.3 and 1.92 µg/mL, respectively (42). Interestingly, 72% of the panel was neutralized by 10E8 with an IC50 below 1 µg/mL, comparing with 37 and 16% for 4E10 and 2F5, respectively (42). Therefore, 10E8 could neutralize with a far greater potency and breadth than previously discovered anti-MPER bNAbs 2F5 and 4E10, and was comparable with some of the most potent HIV-1 bNAbs like VRC01 or PG9/PG16 (15). Notably, 10E8 was also reported to protect against viral challenge in vivo (30).
Interestingly, 2F5, 4E10, and 10E8 antibodies are IgG3 (42, 61); however, the role of this IgG subclass in the neutralizing properties of these antibodies, if any, remains elusive. Although IgG1 and IgG3 are the predominant antibodies elicited against viral antigens (62), both subclasses show important differences. IgG3 shows higher affinity for Fcγ receptors than IgG1, a shorter half-life and a long highly flexible hinge region which has been suggested to be crucial to facilitate the access of these antibodies to the MPER and mediate their neutralizing activity (63, 64). However, it is still unclear whether an IgG3 background is absolutely required, since anti-MPER neutralizing responses have been identified in the non-IgG3 fraction of some HIV-infected individuals (65), and a change to IgG1 did not affect the neutralizing activity of 2F5 and 4E10 antibodies (61, 66). In this context, anti-MPER bNAbs could have been specifically generated from germline precursors preferentially undergoing IgG3 class switching (67) and, in some cases, after affinity maturation and antigen selection by somatic hypermutation, switching to a more downstream IgG subclasses, such as IgG1, by sequential class switching recombination (68). Because IgG3 is one of the less represented IgG subclasses, with the shortest half-life in plasma and IgG3-dominant humoral responses are uncommon (63), elucidating whether this IgG subclass is required for the development of anti-MPER bNAbs, might be crucial to define immunization strategies aimed to generate effective long-lasting anti-MPER responses.
Independently of their origin, all these antibodies are the result of a long process of affinity maturation and are highly mutated with an unusually long and hydrophobic IgH complementary determining region 3 (CDR H3) (42, 69, 70). Notably, these antibodies share a common neutralization mechanism in which the interaction of the hydrophobic CDR H3 apex with the membrane seems to be essential (see next section) (71, 72). Accordingly, autoreactivity/polyreactivity are odd characteristics of 2F5 and 4E10 antibodies. Initially, 10E8 was reported to be non-polyreactive but subsequent studies suggested that 10E8 needs to bind membrane lipids, especially cholesterol, to mediate neutralization (42, 73, 74).
Depending on the bound antibody, the MPER can acquire a particular conformation. Crystal structures of 2F5 in complex with an MPER peptide showed that the core motif DKW forms a type 1 β-turn structure (75). Contrary, the MPER in complex with 4E10 was found to form an α-helical conformation from D674 to K683 (70, 76). Recently, the crystal structure of 10E8 bound to an scaffolded MPER construct revealed that the full epitope of 10E8 is composed of both MPER and lipids (74). Encouragingly, the frequency of 10E8-like antibodies in HIV-infected individuals seemed to be superior to 2F5 or 4E10 specificities in the cohort where 10E8 was isolated (42).
Very recently, a new lineage of distal MPER-specific bNAbs, designated as DH511, was isolated from memory B-cells and plasma of an HIV-infected donor (67). DH511 lineage presented long CDR H3 loops of 23 to 24 aminoacids, an VH and VL somatic mutation rate of 15–22 and 14–18%, respectively, and was derived from the same heavy chain germline gene family as 10E8 (VH 3–15). Similarly to 2F5, 4E10, and 10E8, DH511 clonal lineage presented an IgG3 isotype. Interestingly, the most potent mAb of this clonal lineage, DH511.2, neutralized 206 out of 208 pseudovirus of a geographically and genetically diverse panel with a median IC50 of 1 µg/mL, being slightly more broad but less potent than 10E8 (67).
Lipid Binding and the Concern of Polyreactivity
MPER and Lipids
Biophysical models suggest that the MPER acquires an alpha-helical conformation partially embedded into the viral membrane, constituted by two independent domains separated by a flexible hinge (37, 77). These two segments showed to present different membrane-interacting properties. The C-terminal domain remains embedded into the membrane, whereas the N-terminal domain is more exposed (37, 77–79). The high tryptophan content is likely responsible of the MPER potential to interact with and destabilize lipid membranes (80, 81). According to its amphiphilic characteristics, hydrophobic residues remain buried into the membrane whereas the most polar ones are solvent-exposed (37). Of note, the MPER topology depends on the membrane context where it is presented (82, 83) and membrane lipids such as cholesterol and sphingomyelin can modulate the capacity of the MPER to destabilize membranes (82, 83). MPER and cholesterol interactions are further supported by the existence of the sequence 679-LWYIK-683 located at the C-terminus which was identified as a cholesterol recognition amino acid consensus motif (84). This motif seems to play an important role during the incorporation of Env into the virion, stabilizing the trimer complex (22).
Neutralization Mechanisms and the Importance of Membrane Interaction
Antibody binding to a precise peptide sequence is necessary but not sufficient to achieve MPER-dependent antibody neutralization. Accordingly, z13e1 or 13H11 antibodies overlap the sequences bound by 4E10 and 2F5 respectively with similar affinities but displaying a far low neutralization potency (44, 85). MPER bNAbs show an enrichment of their long CDR H3 loops in hydrophobic residues that seem to be important for their neutralization capability (48, 86, 87). Whereas some residues of the CDRs are important for binding to the peptidic epitope, the most hydrophobic loops interact directly with membrane lipids (71, 72, 87). SPR-based studies demonstrated that whereas anti-MPER bNAbs bind to a peptide sequence following a Langmuir curve model, binding against peptide-membrane complexes follow a two steps (encounter-docking) model. First, the antibody attaches to the lipid membrane through its long hydrophobic CDR H3 and concentrates within the proximity of the MPER epitope to subsequently bind to the prehairpin intermediate of gp41, once the conformational change takes place (71, 72). This mechanism facilitates the accessibility of the antibody to its epitope, overcoming the poor exposure of the MPER and takes advantage of its close proximity to the viral membrane. Of note, upon binding, 2F5 or 4E10 promote an MPER conformational change, due to the extraction of the membrane-embedded epitope (37, 77).
Interestingly, the 2F5 antibody was predicted to bind lipids via CDRL1 and CDRH3 (88) and lipid binding sites were recently determined for 4E10 and 10E8 by X-ray crystallography (74, 89). 4E10 was shown to interact specifically with phosphatidic acid, phosphatidylglicerol and glycerol phosphate by using the CDR H1 and CDR H3 loops to bind polar head and hydrophobic tail groups respectively (89). In a second study, 10E8 lipid binding site was identified at the proximity of CDR L1 and CDR H3 loops (74). Therefore, the full epitope of MPER bNAbs is constituted by both peptide residues and membrane lipids. Notably, neutralizing activity of an anti-MPER single-chain bivalent llama antibody induced by immunization was also dependent of the hydrophobic CDR H3 apex without being involved in peptide recognition (87). Membrane interaction, thus, seems to play a major role in the neutralization mechanism of MPER bNAbs (26, 37, 72, 73, 77, 86).
The widely described importance of the membrane in MPER structure and functionality of the specific bNAbs suggest a role of lipids as a natural scaffold shaping the MPER structure. In this regard it is likely that lipids participate in the selection of germline precursors of bNAbs, pointing their relevance for immunogen design. Therefore, the generation of neutralizing anti-MPER responses may require its presentation within a membrane environment to properly present neutralizing determinants and to implement lipid cross-reactivity. The role of membrane lipids over MPER immunogenicity is, thus, a relevant issue currently being evaluated in immunization studies.
Binding to Self-Antigens: A Major Roadblock for MPER Neutralizing Antibodies?
Reactivity with self-antigens was suggested to explain the failure of generating MPER neutralizing antibodies by immunization as well as their low frequencies during natural infection (38, 90, 91). Gp41 antibodies generated during acute infection are usually derived from polyreactive antibodies whose precursors cross-react with antigens from intestinal microbiota (92–94). In 2005, polyspecific binding of 4E10 and 2F5 mAbs to cardiolipin and other anionic phospholipids was reported (90). Furthermore, conserved host antigens bound by 2F5, 4E10 and 10E8 have been also identified (95, 96). 2F5 binds to the enzyme kinureninase (KYNU), which contains the identical sequence (ELDKWA) of the 2F5 epitope, and is highly conserved between different mammal species. 4E10 binds to splicing factor-3b subunit-3 and type I inositol triphosphate (IP3R1) (95) and, although initially described as non-autoreactive, 10E8 recognize the FAM84A protein (96). Collectively, these findings suggested that immunological tolerance might be involved in HIV-1 evasion of immune responses since autoreactive B-cells that cross-react with MPER sequences might be impaired in the naive repertoire (91, 97).
This hypothesis was tested by monitoring B-cell development in knock-in (KI) mice models carrying the same V(D)J rearrangements as mature bNAbs 2F5 and 4E10. These models showed a normal early B cell development but exhibited a blockade in the transition of pre-B to immature IgM+ B cells, which is defined by the first tolerance checkpoint (98–101). B-cell central tolerance takes place in the bone marrow (BM) and abrogates the development of autoreactive B-cells by several mechanisms such as clonal deletion or receptor edition (102). After that, some autoreactive B-cells can still egress from BM as anergic cells, which show a hyporesponder status and a reduced lifespan. However, in special circumstances anergic B-cells can be activated and differentiate to antibody-producing cells (103). In accordance with this, immunization of 2F5 KI mice with MPER peptide-liposome immunogens could rescue anergic B-cells to produce specific neutralizing antibodies (104, 105). More recently, a 2F5 germ-line KI mouse model showed 2F5 precursors deletion while the remaining anergic B cells could be also activated by germ-line mimicking immunogens (106). These outcomes indicated that the generation of 2F5 and 4E10 antibodies is likely controlled by immunological tolerance mechanisms and launched the hypothesis that HIV-1 host mimicry is an evolutionary strategy of pathogens and not particularly restricted to HIV-1 (95, 96). However, it is important to highlight that HIV-1 epitope mimicry does not impair the functionality of the host enzyme kynureninase, bound by 2F5 (107), and infusion of 2F5 or 4E10 in human recipients showed no major clinical complications (57), supporting the safety of eliciting MPER protective antibodies by vaccination (57, 107).
The MPER Response during Natural Infection and Balance between Neutralizing and Non-Neutralizing Antibodies
The whole gp41 is mostly occluded by gp120 within the native viral spike, being the MPER transiently exposed during the fusion process (25). In consequence, B-cells accessibility to gp41 and native MPER may be compromised. Despite this, a strong antibody response is generated against the gp41 subunit in the course of HIV-1 infection probably due to gp120 shedding, non-functional forms of Env or transient epitope exposure during viral entry (108). Interestingly, the anti-gp41 humoral response can be detected two weeks after HIV-1 acquisition (108). This response, typically non-neutralizing and highly cross-reactive to gut commensal bacteria (92–94), is mainly focused against more exposed regions of gp41 such as the immunodominant disulfide loop, different from the MPER (108, 109).
Whereas MPER antibodies can be easily detected by ELISA, the analysis of their contribution to neutralizing activity of human plasma samples was found to be challenging. With this purpose chimeric SIV or HIV-2 viruses engrafted with HIV-1 MPER sequences or peptide-coated beads adsorption assays were developed (110–112). Accordingly, the presence of anti-MPER antibodies and the evaluation of their neutralizing capacity have been reported (35, 36, 65, 111, 113–116). The characterization of different cohorts in Europe, the United States, and South Africa indicated that MPER-specific neutralizing responses are less represented during natural infection comparing with other neutralizing specificities. For example, in a South African cohort of 156 HIV-1 infected individuals, only three showed higher titers of anti-MPER antibodies (65). Depletion of these antibodies resulted in loss of the neutralization breadth but the antibody specificities were found to be targeting a distinct epitope from those recognized by previously identified neutralizing epitopes (bound by 2F5 and 4E10 bNAbs), highlighting the existence of additional neutralizing specificities within the MPER (65). A recent study of the Protocol C cohort analyzed the neutralization profile of 439 plasma samples showing a far great less prevalence of MPER-specific antibodies when comparing with other specificities, mainly V3 N332-dependent glycan supersite (36). Remarkably, 27% of HIV-1 infected patients from an American cohort presented MPER-specific neutralizing activity (42). We previously showed that 66% of ART-naive chronically HIV-1 infected subjects presented MPER antibodies that were stable, at least for 1 year, and with an heterogeneous neutralizing capacity, highlighting the coexistence of neutralizing and non-neutralizing antibodies targeting the MPER (117). Moreover, anti-MPER antibodies correlate with the total anti-Env humoral response (117) and neutralization breadth (113, 118) and have been identified in HIV-infected individuals at different stages of the infection (119). Therefore, this landscape highlights that regardless of the cohort of study, anti-MPER antibodies (neutralizing and non-neutralizing) are present in HIV-1 infected subjects but their prevalence seems to be highly heterogeneous and probably strongly dependent on the methodology used (42, 65, 114, 117–119). Thus, the optimization of the current methodology for the quantification of MPER antibodies is highly desirable in order to establish their real prevalence. Human studies characterizing the MPER-specific neutralizing response are summarized in Table 1.
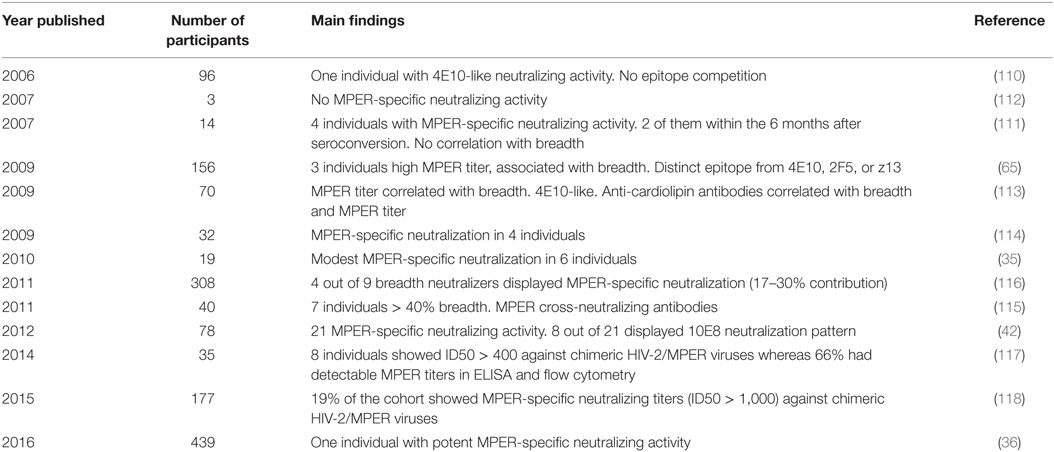
Table 1. Human studies detecting MPER-specific neutralizing responses.
The results obtained from these studies also point out that the MPER is sufficiently immunogenic to generate a humoral response and that no specific constraints limit antibody generation against this region. However, the relatively low prevalence of MPER-neutralizing responses identified to date indicates that some hurdles are involved in the generation of this type of antibodies. The low accessibility of this region, which may compromise the affinity maturation process, as well as other mechanisms such as lipid cross-reactivity, might be determinant for the establishment of a balance between neutralizing and non-neutralizing MPER antibodies. Therefore, this balance is a relevant issue with important implications for vaccine design, where immunogens exposing native MPER neutralizing determinants should be implemented.
Eliciting Anti-MPER Antibodies by Immunization
The particular features of the MPER described above, mainly low accessibility, close proximity to the membrane and subsequent hydrophobicity add additional hurdles for immunogen design against this vulnerability site. Moreover, the scarcity of MPER bNAbs isolated to date, comparing with other Env specificities does not contribute to enlarge our knowledge regarding the MPER complexity and the functional epitopes that should be targeted.
Initial approaches to induce 2F5 or 4E10-like antibodies attempted to introduce their corresponding binding sequences into chimeric viruses, fusion proteins or peptide-based vaccines (21). Although MPER-specific antibodies were elicited, neutralizing responses were not. Therefore, it became clear that additional variables beyond the recognition of specific peptidic sequences within the MPER should be considered. The common characteristics revealed later by MPER bNAbs, such as membrane cross-reactivity and binding to the gp41 prehairpin intermediate (72, 120), suggested that similar antibodies could be obtained by presenting MPER-based antigens in such precise conformational states in a membrane-like environment. In accordance, there are two major standpoints that are currently being addressed in MPER-based vaccinology: (i) what are the relevant structures that most likely mimic the native-bound form of MPER bNAbs and (ii) which is the role of membrane lipids over the MPER immunogenicity, including the precise lipid components and adjuvant systems. A summary of the most recent (since 2010) strategies followed in immunization protocols are listed in Table 2.
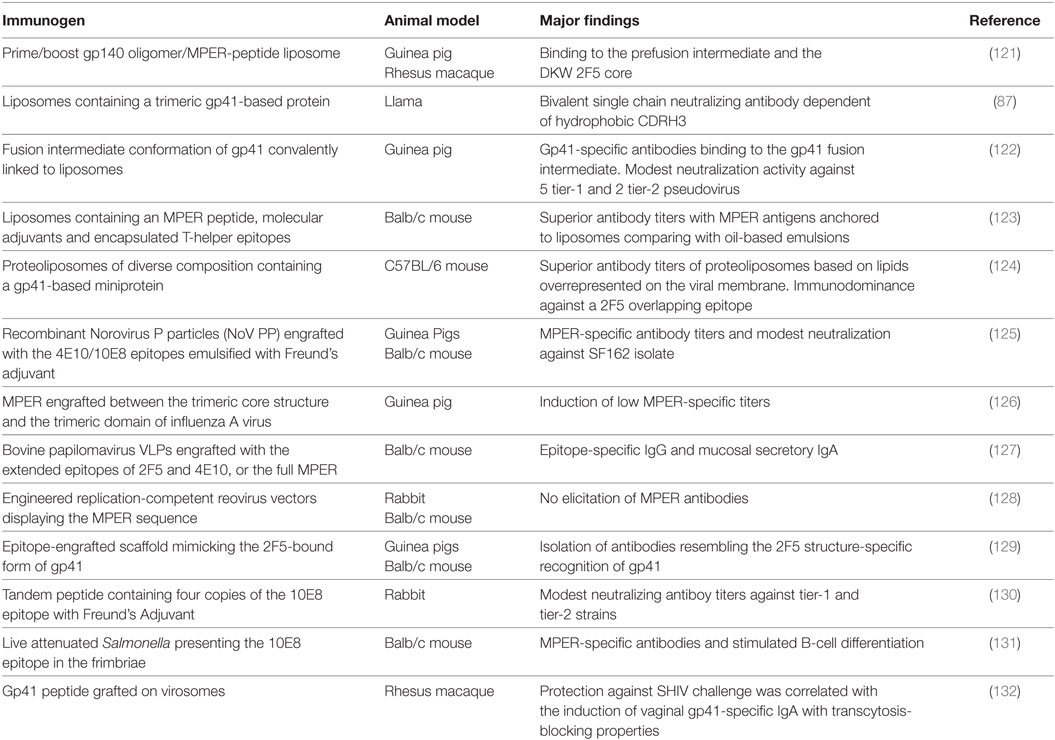
Table 2. Selection of recent immunization studies to elicit MPER neutralizing antibodies.
Conformational states bound by anti-MPER bNAbs have been approached (121, 122, 129, 133). The use of computational methods permitted the design of scaffolds consisting in unrelated protein structures selected from database but able to accommodate the neutralizing 2F5 binding sequence in a conformation close to the peptide-bound crystal structure. Such scaffolds induced polyclonal responses mimicking a 2F5-like binding profile in immunized animals (129). Crystallographic analysis confirmed that monoclonal antibodies isolated from immunized animals mimicked the conformation of 2F5 in a flexible gp41 peptide, high affinity to the same sequence and similar angle of epitope approach (129, 134). Same outcomes were obtained with scaffolds targeting the 4E10 (135) and z13e1 (136) binding motifs. In spite of such structural mimicry, neutralizing activity was not achieved, likely because additional features such as membrane binding were not addressed in the design of these scaffolds.
Due to the importance for neutralization and their implication in a substantial portion of the free energy of 2F5, 4E10, and 10E8 binding, lipid-containing immunogen are important platforms being explored (71, 87, 88). Given that the complete epitope of anti-MPER bNAbs includes membrane components (74, 89) and that lipid recognition by CDR H3 impacts into their functionality (69, 72, 73, 86, 87), their potential for contributing to MPER-specific neutralizing responses by immunization is worth exploring. In this regard, membrane-mimicking platforms including viral-like particles (VLP) (137, 138) or liposomes (122–124) have been approached. It has been shown that membrane lipids can modulate the MPER structure likely by promoting a native-like conformation and demonstrated to improve immunogenicity (123, 124). In particular, we previously demonstrated that those lipids overrepresented in the viral membrane such as cholesterol and sphingomyelin have the potential to induce stronger antibody titers comparing with simple POPC lipids (124). Interestingly, MPER-specific antibodies from long-lived Bone marrow plasma cells from mice immunized with antigen-coupled liposomes have been also reported. Those antibodies showed that were shaped under selective pressure promoted by the MPER in the context of lipids and did not display any polyreactive feature (139).
Whereas the implementation of lipid-based platforms achieved MPER-specific antibodies, modest neutralizing titers have been reported by a few studies. For example, liposome-peptide antigens in combination with MPLA molecular adjuvant led to the isolation of two MPER-specific IgM antibodies showing lipid cross-reactivity but limited neutralizing capacity (140). The use of an HA/gp41 fusion protein in viral like particles induced modest 4E10-like neutralizating titers (141). One study by Dennison and colleagues obtained MPER-specific antibodies in NHP which bound preferentially to the gp41 prehairpin fusion intermediate rather than a recombinant gp41 construct by using a gp140 oligomer prime boosted with liposomes exposing an MPER peptide regimen. Such preferential binding was thought to be primarily due to structural modifications induced by the liposomes where the antigen was presented (121). Furthermore, the response mapped specifically the 2F5 DKW neutralizing core (121). In spite of these promising results, neutralizing activity was not achieved. Mimicking the gp41 prehairpin intermediate has been also approached by the design of a gp41 immunogen formulated in proteoliposomes. Immunization of guinea pigs showed modest neutralizing titers against tier 1 viruses, although the specificities responsible for such neutralization were not delineated (122). Finally, the role of non-neutralizing antibodies in protection has been shown in some studies. The presence of vaginal IgA with ADCC and transcytosis-bocking properties induced by gp41-grafted virosomes was associated with protection of NHP against SHIV challenge (132). Such vaccine platform was also evaluated in a Phase I clinical trial in healthy women. Vaginal secretions of vaccinated subjects were found to present transcytosis-blocking properties in vitro (142).
Remark
In spite of the recent advances into the MPER physical and immunological properties, we still lack a full roadmap to generate a neutralizing response against this HIV-1 Env vulnerability site. The outcomes derived from MPER immunization studies clearly demonstrate that lipid cross-reactivity, binding to certain neutralizing epitopes or binding to gp41 native structures like the prehairpin intermediate are achievable. Although the implementation of these features will have a crucial role they will be likely insufficient to achieve the full properties of MPER-specific bNAbs in immunization protocols. In contrast, the selection of MPER non-neutralizing antibodies whose B-cell precursors may compete for the antigen presented cannot be excluded. While the knowledge gained from other Env vulnerability regions has advanced from the higher number of bNAbs isolated, to date only the potent 10E8 as well as 2F5 and 4E10 antibodies have been isolated. This fact highlights the need of the isolation of additional MPER bNAbs in order to bypass these gaps of our knowledge, improving immunogen design, while avoiding immunodominant non-neutralizing epitopes.
Author Contributions
LM-A drafted the manuscript, JC reviewed the manuscript and JB and BC made substantial, direct, and intellectual contribution to he work. All authors approved it for publication.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Funding
This work was supported by the HIVACAT Program, the CERCA Program (Generalitat de Catalunya), the Spanish AIDS network “Red Temática Cooperativa de Investigación en SIDA” (RD12/0017/0002), the Fondo de Investigaciones Sanitarias, and FEDER “Fondo Europeo de Desarrollo Regional” (grant number PI14/01307, to JB). JB is a researcher from Fundació Institut de Recerca en Ciències de la Salut Germans Trias i Pujol supported by the Health Department of the Catalan Government (Generalitat de Catalunya). LM-A was supported by an FI grant from Agència de Gestió d’Ajuts Universitaris i de Recerca from Generalitat de Catalunya and European Social Fund.
References
1. Wyatt R, Kwong PD, Desjardins E, Sweet RW, Robinson J, Hendrickson WA, et al. The antigenic structure of the HIV gp120 envelope glycoprotein. Nature (1998) 393:705–11. doi:10.1038/31514
2. Liu J, Bartesaghi A, Borgnia MJ, Sapiro G, Subramaniam S. Molecular architecture of native HIV-1 gp120 trimers. Nature (2008) 455:109–13. doi:10.1038/nature07159
3. Zanetti G, Briggs JAG, Grünewald K, Sattentau QJ, Fuller SD. Cryo-electron tomographic structure of an immunodeficiency virus envelope complex in situ. PLoS Pathog (2006) 2(8):e83. doi:10.1371/journal.ppat.0020083
4. Zhu P, Liu J, Bess J, Chertova E, Lifson JD, Grisé H, et al. Distribution and three-dimensional structure of AIDS virus envelope spikes. Nature (2006) 441:847–52. doi:10.1038/nature04817
5. Lee JH, Ozorowski G, Ward AB. Cryo-EM structure of a native, fully glycosylated, cleaved HIV-1 envelope trimer. Science (2016) 351:1043–8. doi:10.1126/science.aad2450
6. Chan DC, Kim PS. HIV entry and its inhibition. Cell (1998) 93:681–4. doi:10.1016/S0092-8674(00)81430-0
7. Maddon PJ, Dalgleish AG, McDougal JS, Clapham PR, Weiss RA, Axel R. The T4 gene encodes the AIDS virus receptor and is expressed in the immune system and the brain. Cell (1986) 47:333–48. doi:10.1016/0092-8674(86)90590-8
8. McDougal JS, Maddon PJ, Dalgleish AG, Clapham PR, Littman DR, Godfrey M, et al. The T4 glycoprotein is a cell-surface receptor for the AIDS virus. Cold Spring Harb Symp Quant Biol (1986) 51 Pt 2:703–11. doi:10.1101/SQB.1986.051.01.083
9. Rizzuto CD, Cgacgat G, Luc T-J, Rizzuto CD, Wyatt R, Herna N, et al. A conserved HIV gp120 glycoprotein structure involved in chemokine receptor binding. Science (1998) 280:1949–54. doi:10.1126/science.280.5371.1949
10. Blumenthal R, Durell S, Viard M. HIV entry and envelope glycoprotein-mediated fusion. J Biol Chem (2012) 287:40841–9. doi:10.1074/jbc.R112.406272
11. Klasse PJ. The molecular basis of HIV entry. Cell Microbiol (2012) 14:1183–92. doi:10.1111/j.1462-5822.2012.01812.x
12. Bellamy-McIntyre AK, Lay CS, Bär S, Maerz AL, Talbo GH, Drummer HE, et al. Functional links between the fusion peptide-proximal polar segment and membrane-proximal region of human immunodeficiency virus gp41 in distinct phases of membrane fusion. J Biol Chem (2007) 282:23104–16. doi:10.1074/jbc.M703485200
13. Haynes BF, Mascola JR. The quest for an antibody-based HIV vaccine. Immunol Rev (2017) 275:5–10. doi:10.1111/imr.12517
14. Mascola JR, Haynes BF. HIV-1 neutralizing antibodies: understanding nature’s pathways. Immunol Rev (2013) 254:225–44. doi:10.1111/imr.12075
15. West AP, Scharf L, Scheid JF, Klein F, Bjorkman PJ, Nussenzweig MC. Structural insights on the role of antibodies in HIV-1 vaccine and therapy. Cell (2014) 156:633–48. doi:10.1016/j.cell.2014.01.052
16. Kwong PD, Mascola JR, Nabel GJ. Broadly neutralizing antibodies and the search for an HIV-1 vaccine: the end of the beginning. Nat Rev Immunol (2013) 13:693–701. doi:10.1038/nri3516
17. McCoy LE, Burton DR. Identification and specificity of broadly neutralizing antibodies against HIV. Immunol Rev (2017) 275:11–20. doi:10.1111/imr.12484
18. Clerici MA, Barassi CB, Devito CD, Pastori CB, Piconi SA, Trabattoni DA, et al. Serum IgA of HIV-exposed uninfected individuals inhibit HIV through recognition of a region within the alpha-helix of gp41. AIDS (2002) 16:1731–41. doi:10.1097/00002030-200209060-00004
19. Pastori C, Tudor D, Diomede L, Drillet AS, Jegerlehner A, Röhn TA, et al. Virus like particle based strategy to elicit HIV-protective antibodies to the alpha-helic regions of gp41. Virology (2012) 431:1–11. doi:10.1016/j.virol.2012.05.005
20. Corti D, Langedijk JPM, Hinz A, Seaman MS, Vanzetta F, Fernandez-Rodriguez BM, et al. Analysis of memory B cell responses and isolation of novel monoclonal antibodies with neutralizing breadth from HIV-1-infected individuals. PLoS One (2010) 5:e8805. doi:10.1371/journal.pone.0008805
21. Montero M, van Houten NE, Wang X, Scott JK. The membrane-proximal external region of the human immunodeficiency virus type 1 envelope: dominant site of antibody neutralization and target for vaccine design. Microbiol Mol Biol Rev (2008) 72:54–84. doi:10.1128/MMBR.00020-07
22. Gach JS, Leaman DP, Zwick MB. Targeting HIV-1 gp41 in close proximity to the membrane using antibody and other molecules. Curr Top Med Chem (2011) 11:2997–3021. doi:10.2174/156802611798808505
23. Salzwedel K, West JT, Hunter E. A conserved tryptophan-rich motif in the membrane-proximal region of the human immunodeficiency virus type 1 gp41 ectodomain is important for Env-mediated fusion and virus infectivity. J Virol (1999) 73:2469–80.
24. Muñoz-Barroso I, Salzwedel K, Hunter E, Blumenthal R. Role of the membrane-proximal domain in the initial stages of human immunodeficiency virus type 1 envelope glycoprotein-mediated membrane fusion. J Virol (1999) 73:6089–92.
25. Dimitrov AS, Jacobs A, Finnegan CM, Stiegler G, Katinger H, Blumenthal R. Exposure of the membrane-proximal external region of HIV-1 gp41 in the course of HIV-1 envelope glycoprotein-mediated fusion. Biochemistry (2007) 46:1398–401. doi:10.1021/bi062245f
26. Vishwanathan SA, Hunter E. Importance of the membrane-perturbing properties of the membrane-proximal external region of human immunodeficiency virus type 1 gp41 to viral fusion. J Virol (2008) 82:5118–26. doi:10.1128/JVI.00305-08
27. Dimitrov AS, Rawat SS, Jiang S, Blumenthal R. Role of the fusion peptide and membrane-proximal domain in HIV-1 envelope glycoprotein-mediated membrane fusion. Biochemistry (2003) 42:14150–8. doi:10.1021/bi035154g
28. Hessell AJ, Rakasz EG, Tehrani DM, Huber M, Weisgrau KL, Landucci G, et al. Broadly neutralizing monoclonal antibodies 2F5 and 4E10 directed against the human immunodeficiency virus type 1 gp41 membrane-proximal external region protect against mucosal challenge by simian-human immunodeficiency virus SHIVBa-L. J Virol (2010) 84:1302–13. doi:10.1128/JVI.01272-09
29. Mascola JR, Stiegler G, VanCott TC, Katinger H, Carpenter CB, Hanson CE, et al. Protection of macaques against vaginal transmission of a pathogenic HIV-1/SIV chimeric virus by passive infusion of neutralizing antibodies. Nat Med (2000) 6:207–10. doi:10.1097/00002030-200102001-00018
30. Pegu A, Yang Z-Y, Boyington JC, Wu L, Ko S-Y, Schmidt SD, et al. Neutralizing antibodies to HIV-1 envelope protect more effectively in vivo than those to the CD4 receptor. Sci Transl Med (2014) 6:243ra88. doi:10.1126/scitranslmed.3008992
31. Zwick MB. The membrane-proximal external region of HIV-1 gp41: a vaccine target worth exploring. AIDS (2005) 19:1725–37. doi:10.1097/01.aids.0000189850.83322.41
32. Bomsel M. Transcytosis of infectious human immunodeficiency virus across a tight human epithelial cell line barrier. Nat Med (1997) 3:42–7. doi:10.1038/nm0197-42
33. Alfsen A, Bomsel M. HIV-1 gp41 envelope residues 650-685 exposed on native virus act as a lectin to bind epithelial cell galactosyl ceramide. J Biol Chem (2002) 277:25649–59. doi:10.1074/jbc.M200554200
34. Alfsen A, Iniguez P, Bouguyon E, Bomsel M. Secretory IgA specific for a conserved epitope on gp41 envelope glycoprotein inhibits epithelial transcytosis of HIV-1. J Immunol (2001) 166:6257–65. doi:10.4049/jimmunol.166.10.6257
35. Walker LM, Simek MD, Priddy F, Gach JS, Wagner D, Zwick MB, et al. A limited number of antibody specificities mediate broad and potent serum neutralization in selected HIV-1 infected individuals. PLoS Pathog (2010) 6:e1001028. doi:10.1371/journal.ppat.1001028
36. Landais E, Huang X, Havenar-Daughton C, Murrell B, Price MA, Wickramasinghe L, et al. Broadly neutralizing antibody responses in a large longitudinal sub-Saharan HIV primary infection cohort. PLoS Pathog (2016) 12:e1005369. doi:10.1371/journal.ppat.1005369
37. Sun ZYJ, Oh KJ, Kim M, Yu J, Brusic V, Song L, et al. HIV-1 broadly neutralizing antibody extracts its epitope from a kinked gp41 ectodomain region on the viral membrane. Immunity (2008) 28:52–63. doi:10.1016/j.immuni.2007.11.018
38. Kelsoe G, Haynes BF. Host controls of HIV broadly neutralizing antibody development. Immunol Rev (2017) 275:79–88. doi:10.1111/imr.12508
39. Muster T, Steindl F, Purtscher M, Trkola A, Klima A, Himmler G, et al. A conserved neutralizing epitope on gp41 human immunodeficiency virus type 1. J Virol (1993) 67:6642–7.
40. Muster T, Guinea R, Trkola A, Purtscher M, Klima A, Steindl F, et al. Cross-neutralizing activity against divergent human immunodeficiency virus type 1 isolates induced by the gp41 sequence ELDKWAS. J Virol (1994) 68:4031–4.
41. Stiegler G, Kunert R, Purtscher M, Wolbank S, Voglauer R, Steindl F, et al. A potent cross-clade neutralizing human monoclonal antibody against a novel epitope on gp41 of human immunodeficiency virus type 1. AIDS Res Hum Retroviruses (2001) 17:1757–65. doi:10.1089/08892220152741450
42. Huang J, Ofek G, Laub L, Louder MK, Doria-Rose NA, Longo NS, et al. Broad and potent neutralization of HIV-1 by a gp41-specific human antibody. Nature (2012) 491:406–12. doi:10.1038/nature11544
43. Zwick MB, Labrijn AF, Wang M, Spenlehauer C, Saphire EO, Binley JM, et al. Broadly neutralizing antibodies targeted to the membrane-proximal external region of human immunodeficiency virus type 1. J Virol (2001) 75:10892–905. doi:10.1128/JVI.75.22.10892
44. Nelson JD, Brunel FM, Jensen R, Crooks ET, Cardoso RMF, Wang M, et al. An affinity-enhanced neutralizing antibody against the membrane-proximal external region of human immunodeficiency virus type 1 gp41 recognizes an epitope between those of 2F5 and 4E10. J Virol (2007) 81:4033–43. doi:10.1128/JVI.02588-06
45. Ofek G, Zirkle B, Yang Y, Zhu Z, McKee K, Zhang B, et al. Structural basis for HIV-1 neutralization by 2F5-like antibodies m66 and m66.6. J Virol (2014) 88:2426–41. doi:10.1128/JVI.02837-13
46. Morris L, Chen X, Alam M, Tomaras G, Zhang R, Marshall DJ, et al. Isolation of a human anti-HIV gp41 membrane proximal region neutralizing antibody by antigen-specific single B cell sorting. PLoS One (2011) 6:e23532. doi:10.1371/journal.pone.0023532
47. Buchacher A, Predl R, Strutzenberger K, Steinfellner W, Trkola A, Purtscher M, et al. Generation of human monoclonal antibodies against HIV-1 proteins; electrofusion and Epstein-Barr virus transformation for peripheral blood lymphocyte immortalization. AIDS Res Hum Retroviruses (1994) 10:359–69. doi:10.1089/aid.1994.10.359
48. Zwick MB, Jensen R, Church S, Wang M, Stiegler G, Kunert R, et al. Anti-human immunodeficiency virus type 1 (HIV-1) antibodies 2F5 and 4E10 require surprisingly few crucial residues in the membrane-proximal external region of glycoprotein gp41 to neutralize HIV-1. J Virol (2005) 79:1252–61. doi:10.1128/JVI.79.2.1252-1261.2005
49. Binley JM, Wrin T, Korber B, Zwick MB, Wang M, Chappey C, et al. Comprehensive cross-clade neutralization analysis of a panel of anti-human immunodeficiency virus type 1 monoclonal antibodies comprehensive cross-clade neutralization analysis of a panel of anti-human immunodeficiency virus type 1 monoclonal antibodies. J Virol (2004) 78:13232–52. doi:10.1128/JVI.78.23.13232
50. Bures R, Morris L, Williamson C, Deers M, Fiscus SA, Abdool-karim S, et al. Regional clustering of shared neutralization determinants on primary isolates of clade C human immunodeficiency virus type 1 from South Africa. J Virol (2002) 76:2233–44. doi:10.1128/JVI.76.5.2233
51. Gray ES, Meyers T, Gray G, Montefiori DC, Morris L. Insensitivity of paediatric HIV-1 subtype C viruses to broadly neutralising monoclonal antibodies raised against subtype B. PLoS Med (2006) 3:1023–31. doi:10.1371/journal.pmed.0030255
52. Walker LM, Phogat SK, Chan-Hui P-Y, Wagner D, Phung P, Goss JL, et al. Broad and potent neutralizing antibodies from an African donor reveal a new HIV-1 vaccine target. Science (2009) 326:285–9. doi:10.1126/science.1178746
53. Louder MK, Sambor A, Chertova E, Hunte T, Barrett S, Ojong F, et al. HIV-1 envelope pseudotyped viral vectors and infectious molecular clones expressing the same envelope glycoprotein have a similar neutralization phenotype, but culture in peripheral blood mononuclear cells is associated with decreased neutralization sensi. Virology (2005) 339:226–38. doi:10.1016/j.virol.2005.06.003
54. Provine NM, Puryear WB, Wu X, Overbaugh J, Haigwood NL. The infectious molecular clone and pseudotyped virus models of human immunodeficiency virus type 1 exhibit significant differences in virion composition with only moderate differences in infectivity and inhibition sensitivity. J Virol (2009) 83:9002–7. doi:10.1128/JVI.00423-09
55. Provine NM, Cortez V, Chohan V, Overbaugh J. The neutralization sensitivity of viruses representing human immunodeficiency virus type 1 variants of diverse subtypes from early in infection is dependent on producer cell, as well as characteristics of the specific antibody and envelope variant. Virology (2012) 427:25–33. doi:10.1016/j.virol.2012.02.001
56. Miglietta R, Pastori C, Venuti A, Ochsenbauer C, Lopalco L. Synergy in monoclonal antibody neutralization of HIV-1 pseudoviruses and infectious molecular clones. J Transl Med (2014) 12:346. doi:10.1186/s12967-014-0346-3
57. Trkola A, Kuster H, Rusert P, Joos B, Fischer M, Leemann C, et al. Delay of HIV-1 rebound after cessation of antiretroviral therapy through passive transfer of human neutralizing antibodies. Nat Med (2005) 11:615–22. doi:10.1038/nm1244
58. Wu X, Yang Z-Y, Li Y, Hogerkorp C-M, Schief WR, Seaman MS, et al. Rational design of envelope identifies broadly neutralizing human monoclonal antibodies to HIV-1. Science (2010) 329:856–61. doi:10.1126/science.1187659
59. Doria-Rose NA, Klein RM, Manion MM, O’Dell S, Phogat A, Chakrabarti B, et al. Frequency and phenotype of human immunodeficiency virus envelope-specific B cells from patients with broadly cross-neutralizing antibodies. J Virol (2009) 83:188–99. doi:10.1128/JVI.01583-08
60. Scheid JF, Mouquet H, Feldhahn N, Seaman MS, Velinzon K, Pietzsch J, et al. Broad diversity of neutralizing antibodies isolated from memory B cells in HIV-infected individuals. Nature (2009) 458:636–40. doi:10.1038/nature07930
61. Kunert R, Steinfellner W, Purtscher M, Assadian A, Katinger H. Stable recombinant expression of the anti HIV-1 monoclonal antibody 2F5 after IgG3/IgG1 subclass switch in CHO cells. Biotechnol Bioeng (2000) 67:97–103. doi:10.1002/(SICI)1097-0290(20000105)67:1<97:AID-BIT11>3.0.CO;2-2
62. Ferrante A, Lorrain BJ, Feldman RG. IgG subclass distribution of antibodies to bacterial and viral antigens. Pediatr Infect Dis J (1990) 9:516–24. doi:10.1097/00006454-199008001-00004
63. Bruhns P, Iannascoli B, England P, Mancardi DA, Fernandez N, Jorieux S. Specificity and affinity of human Fcy receptors and their polymorphic variants for human IgG subclasses. Blood (2009) 113:3716–25. doi:10.1182/blood-2008-09-179754.The
64. Cavacini LA, Kuhrt D, Duval M, Mayer K, Posner MR. Binding and neutralization activity of human IgG1 and IgG3 from serum of HIV-infected individuals. AIDS Res Hum Retroviruses (2003) 19:785–92. doi:10.1089/088922203769232584
65. Gray ES, Madiga MC, Moore PL, Mlisana K, Abdool Karim SS, Binley JM, et al. Broad neutralization of human immunodeficiency virus type 1 mediated by plasma antibodies against the gp41 membrane proximal external region. J Virol (2009) 83:11265–74. doi:10.1128/JVI.01359-09
66. Kunert R, Wolbank S, Stiegler G, Weik R, Katinger H. Characterization of molecular features, antigen-binding, and in vitro properties of IgG and IgM variants of 4E10, an anti-HIV type 1 neutralizing monoclonal antibody. AIDS Res Hum Retroviruses (2004) 20:755–62. doi:10.1089/0889222041524571
67. Williams LD, Ofek G, Schätzle S, Mcdaniel JR, Lu X, Nicely NI, et al. Potent and broad HIV-neutralizing antibodies in memory B cells and plasma. Sci Immunol (2017) 2:eaal2200. doi:10.1126/sciimmunol.aal2200
68. Jackson KJL, Wang Y, Collins AM. Human immunoglobulin classes and subclasses show variability in VDJ gene mutation levels. Immunol Cell Biol (2014) 92:1–5. doi:10.1038/icb.2014.44
69. Zwick MB, Komori HK, Stanfield RL, Church S, Wang M, Parren PWHI, et al. The long third complementarity-determining region of the heavy chain is important in the activity of the broadly neutralizing anti-human immunodeficiency virus type 1 antibody 2F5. J Virol (2004) 78:3155–61. doi:10.1128/JVI.78.6.3155
70. Cardoso RMF, Zwick MB, Stanfield RL, Kunert R, Binley JM, Katinger H, et al. Broadly neutralizing anti-HIV antibody 4E10 recognizes a helical conformation of a highly conserved fusion-associated motif in gp41. Immunity (2005) 22:163–73. doi:10.1016/j.immuni.2004.12.011
71. Alam SM, McAdams M, Boren D, Rak M, Scearce RM, Gao F, et al. The role of antibody polyspecificity and lipid reactivity in binding of broadly neutralizing anti-HIV-1 envelope human monoclonal antibodies 2F5 and 4E10 to glycoprotein 41 membrane proximal envelope epitopes. J Immunol (2007) 178:4424–35. doi:10.4049/jimmunol.178.7.4424
72. Alam SM, Morelli M, Dennison SM, Liao H-X, Zhang R, Xia S-M, et al. Role of HIV membrane in neutralization by two broadly neutralizing antibodies. Proc Natl Acad Sci U S A (2009) 106:20234–9. doi:10.1073/pnas.0908713106
73. Chen J, Frey G, Peng H, Rits-Volloch S, Garrity J, Seaman MS, et al. Mechanism of HIV-1 neutralization by antibodies targeting a membrane-proximal region of gp41. J Virol (2014) 88:1249–58. doi:10.1128/JVI.02664-13
74. Irimia A, Serra AM, Sarkar A, Jacak R, Kalyuzhniy O, Sok D, et al. Lipid interactions and angle of approach to the HIV-1 viral membrane of broadly neutralizing antibody 10E8: insights for vaccine and therapeutic design. PLoS Pathog (2017) 13:e1006212. doi:10.1371/journal.ppat.1006212
75. Bryson S, Cunningham A, Ho J, Hynes R, Isenman D, Barber B, et al. Cross-neutralizing human monoclonal anti-HIV-1 antibody 2F5: preparation and crystallographic analysis of the free and epitope-complexed forms of its Fab fragment. Protein Pept Lett (2001) 8:413–8. doi:10.2174/0929866013409201
76. Cardoso RMF, Brunel FM, Ferguson S, Zwick M, Burton DR, Dawson PE, et al. Structural basis of enhanced binding of extended and helically constrained peptide epitopes of the broadly neutralizing HIV-1 antibody 4E10. J Mol Biol (2007) 365:1533–44. doi:10.1016/j.jmb.2006.10.088
77. Song L, Sun Z-YJ, Coleman KE, Zwick MB, Gach JS, Wang J, et al. Broadly neutralizing anti-HIV-1 antibodies disrupt a hinge-related function of gp41 at the membrane interface. Proc Natl Acad Sci U S A (2009) 106:9057–62. doi:10.1186/1742-4690-6-S3-O5
78. Huarte N, Lorizate M, Maeso R, Kunert R, Arranz R, Valpuesta JM, et al. The broadly neutralizing anti-human immunodeficiency virus type 1 4E10 monoclonal antibody is better adapted to membrane-bound epitope recognition and blocking than 2F5. J Virol (2008) 82:8986–96. doi:10.1128/JVI.00846-08
79. Kim M, Song L, Moon J, Sun Z-YJ, Bershteyn A, Hanson M, et al. Immunogenicity of membrane-bound HIV-1 gp41 membrane-proximal external region (MPER) segments is dominated by residue accessibility and modulated by stereochemistry. J Biol Chem (2013) 288:31888–901. doi:10.1074/jbc.M113.494609
80. Suárez T, Nir S, Goñi FM, Saéz-Cirión A, Nieva JL. The pre-transmembrane region of the human immunodeficiency virus type-1 glycoprotein: a novel fusogenic sequence. FEBS Lett (2000) 477:145–9. doi:10.1016/S0014-5793(00)01785-3
81. Stano P, Bufali S, Domazou AS, Luisi PL. Effect of tryptophan oligopeptides on the size distribution of POPC liposomes: a dynamic light scattering and turbidimetric study. J Liposome Res (2005) 15:29–47. doi:10.1081/LPR-64956
82. Huarte N, Lorizate M, Kunert R, Nieva JL. Lipid modulation of membrane-bound epitope recognition and blocking by HIV-1 neutralizing antibodies. FEBS Lett (2008) 582:3798–804. doi:10.1016/j.febslet.2008.10.012
83. Sáez-Cirión A, Nir S, Lorizate M, Agirre A, Cruz A, Pérez-Gil J, et al. Sphingomyelin and cholesterol promote HIV-1 gp41 pretransmembrane sequence surface aggregation and membrane restructuring. J Biol Chem (2002) 277:21776–85. doi:10.1074/jbc.M202255200
84. Vincent N, Genin C, Malvoisin E. Identification of a conserved domain of the HIV-1 transmembrane protein gp41 which interacts with cholesteryl groups. Biochim Biophys Acta (2002) 1567:157–64. doi:10.1016/S0005-2736(02)00611-9
85. Alam SM, Scearce RM, Parks RJ, Plonk K, Plonk SG, Sutherland LL, et al. Human immunodeficiency virus type 1 gp41 antibodies that mask membrane proximal region epitopes: antibody binding kinetics, induction, and potential for regulation in acute infection. J Virol (2008) 82:115–25. doi:10.1128/JVI.00927-07
86. Ofek G, Mckee K, Yang Y, Yang Z-Y, Skinner J, Guenaga FJ, et al. Relationship between antibody 2F5 neutralization of HIV-1 and hydrophobicity of its heavy chain third complementarity-determining region. J Virol (2010) 84:2955–62. doi:10.1128/JVI.02257-09
87. Lutje Hulsik D, Liu Y, Strokappe NM, Battella S, El Khattabi M, McCoy LE, et al. A gp41 MPER-specific llama VHH requires a hydrophobic CDR3 for neutralization but not for antigen recognition. PLoS Pathog (2013) 9:e1003202. doi:10.1371/journal.ppat.1003202
88. Julien JP, Bryson S, Nieva JL, Pai EF. Structural details of HIV-1 recognition by the broadly neutralizing monoclonal antibody 2F5: epitope conformation, antigen-recognition loop mobility, and anion-binding site. J Mol Biol (2008) 384:377–92. doi:10.1016/j.jmb.2008.09.024
89. Irimia A, Sarkar A, Stanfield RL, Wilson I. Crystallographic identification of lipid as an integral component of the epitope of HIV broadly neutralizing antibody 4E10. Immunity (2016) 44:1–11. doi:10.1016/j.immuni.2015.12.001
90. Haynes BF, Fleming J, St Clair EW, Katinger H, Stiegler G, Kunert R, et al. Cardiolipin polyspecific autoreactivity in two broadly neutralizing HIV-1 antibodies. Science (2005) 308:1906–8. doi:10.1126/science.1111781
91. Haynes BF, Moody MA, Verkoczy L, Kelsoe G, Alam SM. Antibody polyspecificity and neutralization of HIV-1: a hypothesis. Hum Antibodies (2005) 14:59–67.
92. Liao H-X, Chen X, Munshaw S, Zhang R, Marshall DJ, Vandergrift N, et al. Initial antibodies binding to HIV-1 gp41 in acutely infected subjects are polyreactive and highly mutated. J Exp Med (2011) 208:2237–49. doi:10.1084/jem.20110363
93. Trama AM, Moody MA, Alam SM, Jaeger FH, Lockwood B, Parks R, et al. HIV-1 envelope gp41 antibodies can originate from terminal ileum B cells that share cross-reactivity with commensal bacteria. Cell Host Microbe (2014) 16:215–26. doi:10.1016/j.chom.2014.07.003
94. Williams WB, Liao H-X, Moody MA, Kepler TB, Alam SM, Gao F, et al. Diversion of HIV-1 vaccine-induced immunity by gp41-microbiota cross-reactive antibodies. Science (2015) 349:aab1253. doi:10.1126/science.aab1253
95. Yang G, Holl TM, Liu Y, Li Y, Lu X, Nicely NI, et al. Identification of autoantigens recognized by the 2F5 and 4E10 broadly neutralizing HIV-1 antibodies. J Exp Med (2013) 210:241–56. doi:10.1084/jem.20121977
96. Liu M, Yang G, Wiehe K, Nicely NI, Vandergrift NA, Rountree W, et al. Polyreactivity and autoreactivity among HIV-1 antibodies. J Virol (2015) 89:784–98. doi:10.1128/JVI.02378-14
97. Verkoczy L, Kelsoe G, Haynes BF. HIV-1 envelope gp41 broadly neutralizing antibodies: hurdles for vaccine development. PLoS Pathog (2014) 10:3–6. doi:10.1371/journal.ppat.1004073
98. Verkoczy L, Diaz M. Autoreactivity in HIV-1 broadly neutralizing antibodies: implications for their function and induction by vaccination. Curr Opin HIV AIDS (2014) 9:224–34. doi:10.1097/COH.0000000000000049
99. Verkoczy L, Diaz M, Holl TM, Ouyang Y-B, Bouton-Verville H, Alam SM, et al. Autoreactivity in an HIV-1 broadly reactive neutralizing antibody variable region heavy chain induces immunologic tolerance. Proc Natl Acad Sci U S A (2010) 107:181–6. doi:10.1073/pnas.0912914107
100. Verkoczy L, Chen Y, Zhang J, Bouton-Verville H, Newman A, Lockwood B, et al. Induction of HIV-1 broad neutralizing antibodies in 2F5 knock-in mice: selection against membrane proximal external region-associated autoreactivity limits T-dependent responses. J Immunol (2013) 191:2538–50. doi:10.4049/jimmunol.1300971
101. Doyle-Cooper C, Hudson KE, Cooper AB, Ota T, Skog P, Dawson PE, et al. Immune tolerance negatively regulates B cells in knock-in mice expressing broadly neutralizing HIV antibody 4E10. J Immunol (2013) 191:3186–91. doi:10.4049/jimmunol.1301285
102. Nemazee D. Mechanisms of central tolerance for B cells. Nat Rev Immunol (2017) 17:281–94. doi:10.1038/nri.2017.19
103. von Boehmer H, Melchers F. Checkpoints in lymphocyte development and autoimmune disease. Nat Immunol (2010) 11:14–20. doi:10.1038/ni.1794
104. Dennison SM, Stewart SM, Stempel KC, Liao H-X, Haynes BF, Alam SM. Stable docking of neutralizing human immunodeficiency virus type 1 gp41 membrane-proximal external region monoclonal antibodies 2F5 and 4E10 is dependent on the membrane immersion depth of their epitope regions. J Virol (2009) 83:10211–23. doi:10.1128/JVI.00571-09
105. Verkoczy L, Chen Y, Bouton-Verville H, Zhang J, Diaz M, Hutchinson J, et al. Rescue of HIV-1 broad neutralizing antibody-expressing B cells in 2F5 VH x VL knockin mice reveals multiple tolerance controls. J Immunol (2011) 187:3785–97. doi:10.4049/jimmunol.1101633
106. Zhang R, Verkoczy L, Wiehe K, Alam SM, Nicely NI, Santra S, et al. Initiation of immune tolerance – controlled HIV gp41 neutralizing B cell lineages. Sci Transl Med (2016) 8:1–14. doi:10.1126/scitranslmed.aaf0618
107. Bradley T, Yang G, Ilkayeva O, Holl TM, Zhang R, Zhang J, et al. HIV-1 envelope mimicry of host enzyme kynureninase does not disrupt tryptophan metabolism. J Immunol (2016) 197(12):4663–73. doi:10.4049/jimmunol.1601484
108. Tomaras GD, Yates NL, Liu P, Qin L, Fouda GG, Chavez LL, et al. Initial B-cell responses to transmitted human immunodeficiency virus type 1: virion-binding immunoglobulin M (IgM) and IgG antibodies followed by plasma anti-gp41 antibodies with ineffective control of initial viremia. J Virol (2008) 82:12449–63. doi:10.1128/JVI.01708-08
109. Yates NL, Stacey AR, Nolen TL, Vandergrift NA, Moody MA, Montefiori DC, et al. HIV-1 gp41 envelope IgA is frequently elicited after transmission but has an initial short response half-life. Mucosal Immunol (2013) 6:692–703. doi:10.1038/mi.2012.107
110. Yuste E, Sanford HB, Carmody J, Little S, Zwick MB, Greenough T, et al. Simian immunodeficiency virus engrafted with human immunodeficiency virus type neutralization, and survey of HIV-1-positive plasma simian immunodeficiency virus engrafted with human immunodeficiency virus type 1 (HIV-1)-specific epitopes : replication. J Virol (2006) 80:3030–41. doi:10.1128/JVI.80.6.3030
111. Gray ES, Moore PL, Choge IA, Decker JM, Bibollet-Ruche F, Li H, et al. Neutralizing antibody responses in acute human immunodeficiency virus type 1 subtype C infection. J Virol (2007) 81:6187–96. doi:10.1128/JVI.00239-07
112. Dhillon AK, Donners H, Pantophlet R, Johnson WE, Decker JM, Shaw GM, et al. Dissecting the neutralizing antibody specificities of broadly neutralizing sera from human immunodeficiency virus type 1-infected donors. J Virol (2007) 81:6548–62. doi:10.1128/JVI.02749-06
113. Gray ES, Taylor N, Wycuff D, Moore PL, Tomaras GD, Wibmer CK, et al. Antibody specificities associated with neutralization breadth in plasma from human immunodeficiency virus type 1 subtype C-infected blood donors. J Virol (2009) 83:8925–37. doi:10.1128/JVI.00758-09
114. Li Y, Svehla K, Louder MK, Wycuff D, Phogat S, Tang M, et al. Analysis of neutralization specificities in polyclonal sera derived from human immunodeficiency virus type 1-infected individuals. J Virol (2009) 83:1045–59. doi:10.1128/JVI.01992-08
115. Gray ES, Madiga MC, Hermanus T, Moore PL, Wibmer CK, Tumba NL, et al. The neutralization breadth of HIV-1 develops incrementally over four years and is associated with CD4+ T cell decline and high viral load during acute infection. J Virol (2011) 85:4828–40. doi:10.1128/JVI.00198-11
116. Tomaras GD, Binley JM, Gray ES, Crooks ET, Osawa K, Moore PL, et al. Polyclonal B cell responses to conserved neutralization epitopes in a subset of HIV-1-infected individuals. J Virol (2011) 85:11502–19. doi:10.1128/JVI.05363-11
117. Molinos-Albert LM, Carrillo J, Curriu M, Rodriguez de la Concepción ML, Marfil S, García E, et al. Anti-MPER antibodies with heterogeneous neutralization capacity are detectable in most untreated HIV-1 infected individuals. Retrovirology (2014) 11:44. doi:10.1186/1742-4690-11-44
118. Jacob RA, Moyo T, Schomaker M, Abrahams F, Grau Pujol B, Dorfman JR. Anti-V3/glycan and anti-MPER neutralizing antibodies, but not anti-V2/glycan site antibodies, are strongly associated with greater anti-HIV-1 neutralization breadth and potency. J Virol (2015) 89:5264–75. doi:10.1128/JVI.00129-15
119. Brombin C, Diomede L, Tudor D, Drillet AS, Pastori C, Poli E, et al. A nonparametric procedure for defining a new humoral immunologic profile in a pilot study on HIV infected patients. PLoS One (2013) 8:e58768. doi:10.1371/journal.pone.0058768
120. Frey G, Peng H, Rits-Volloch S, Morelli M, Cheng Y, Chen B. A fusion-intermediate state of HIV-1 gp41 targeted by broadly neutralizing antibodies. Proc Natl Acad Sci U S A (2008) 105:3739–44. doi:10.1073/pnas.0800255105
121. Dennison SM, Sutherland LL, Jaeger FH, Anasti KM, Parks R, Stewart S, et al. Induction of antibodies in rhesus macaques that recognize a fusion-intermediate conformation of HIV-1 gp41. PLoS One (2011) 6:e27824. doi:10.1371/journal.pone.0027824
122. Lai RPJ, Hock M, Radzimanowski J, Tonks P, Hulsik DL, Effantin G, et al. A fusion intermediate gp41 immunogen elicits neutralizing antibodies to HIV-1. J Biol Chem (2014) 289:29912–26. doi:10.1074/jbc.M114.569566
123. Hanson MC, Abraham W, Crespo MP, Chen SH, Liu H, Szeto GL, et al. Liposomal vaccines incorporating molecular adjuvants and intrastructural T-cell help promote the immunogenicity of HIV membrane-proximal external region peptides. Vaccine (2015) 33:861–8. doi:10.1016/j.vaccine.2014.12.045
124. Molinos-albert LM, Bilbao E, Agulló L, Ma S, García E, Luisa M, et al. Proteoliposomal formulations of an HIV-1 gp41-based miniprotein elicit a lipid-dependent immunodominant response overlapping the 2F5 binding motif. Sci Rep (2017) 7:40800. doi:10.1038/srep40800
125. Yu Y, Fu L, Shi Y, Guan S, Yang L, Gong X, et al. Elicitation of HIV-1 neutralizing antibodies by presentation of 4E10 and 10E8 epitopes on Norovirus P particles. Immunol Lett (2015) 168:1–8. doi:10.1016/j.imlet.2015.10.003
126. Zang Y, Du D, Li N, Su W, Liu X, Zhang Y, et al. Eliciting neutralizing antibodies against the membrane proximal external region of HIV-1 Env by chimeric live attenuated influenza A virus vaccines. Vaccine (2015) 33:3859–64. doi:10.1016/j.vaccine.2015.06.072
127. Zhai Y, Zhong Z, Zariffard M, Spear GT, Qiao L. Bovine papillomavirus-like particles presenting conserved epitopes from membrane-proximal external region of HIV-1 gp41 induced mucosal and systemic antibodies. Vaccine (2013) 31:5422–9. doi:10.1016/j.vaccine.2013.09.003
128. Boehme K, Ikizler M, Iskarpatyoti J, Wetzel J, Willis J, Crowe J Jr, et al. Engineering recombinant reoviruses to display gp41 membrane-proximal external-region epitopes from HIV-1. mSphere (2016) 1:1–15. doi:10.1128/mSphere.00086-16
129. Ofek G, Guenaga FJ, Schief WR, Skinner J, Baker D, Wyatt R, et al. Elicitation of structure-specific antibodies by epitope scaffolds. Proc Natl Acad Sci U S A (2010) 107:17880–7. doi:10.1073/pnas.1004728107
130. Sun Z, Zhu Y, Wang Q, Ye L, Dai Y, Su S, et al. An immunogen containing four tandem 10E8 epitope repeats with exposed key residues induces antibodies that neutralize HIV-1 and activates an ADCC reporter gene. Emerg Microbes Infect (2016) 5:e65. doi:10.1038/emi.2016.86
131. Li Q-H, Jin G, Wang J-Y, Li H-N, Liu H, Chang X-Y, et al. Live attenuated Salmonella displaying HIV-1 10E8 epitope on fimbriae: systemic and mucosal immune responses in BALB/c mice by mucosal administration. Sci Rep (2016) 6:29556. doi:10.1038/srep29556
132. Bomsel M, Tudor D, Drillet AS, Alfsen A, Ganor Y, Roger MG, et al. Immunization with HIV-1 gp41 subunit virosomes induces mucosal antibodies protecting nonhuman primates against vaginal SHIV challenges. Immunity (2011) 34:269–80. doi:10.1016/j.immuni.2011.01.015
133. Vassell R, He Y, Vennakalanti P, Dey AK, Zhuang M, Wang W, et al. Immunogens modeling a fusion-intermediate conformation of gp41 elicit antibodies to the membrane proximal external region of the HIV envelope glycoprotein. PLoS One (2015) 10:e0128562. doi:10.1371/journal.pone.0128562
134. Guenaga J, Dosenovic P, Ofek G, Baker D, Schief WR, Kwong PD, et al. Heterologous epitope-scaffold prime:boosting immuno-focuses B cell responses to the HIV-1 gp41 2F5 neutralization determinant. PLoS One (2011) 6:e16074. doi:10.1371/journal.pone.0016074
135. Correia BE, Ban YEA, Holmes MA, Xu H, Ellingson K, Kraft Z, et al. Computational design of epitope-scaffolds allows induction of antibodies specific for a poorly immunogenic HIV vaccine epitope. Structure (2010) 18:1116–26. doi:10.1016/j.str.2010.06.010
136. Stanfield RL, Julien J-P, Pejchal R, Gach JS, Zwick MB, Wilson IA. Structure-based design of a protein immunogen that displays an HIV-1 gp41 neutralizing epitope. J Mol Biol (2011) 414:460–76. doi:10.1016/j.jmb.2011.10.014
137. Kamdem Toukam D, Tenbusch M, Stang A, Temchura V, Storcksdieck Genannt Bonsmann M, Grewe B, et al. Targeting antibody responses to the membrane proximal external region of the envelope glycoprotein of human immunodeficiency virus. PLoS One (2012) 7:1–10. doi:10.1371/journal.pone.0038068
138. Benen TD, Tonks P, Kliche A, Kapzan R, Heeney JL, Wagner R. Development and immunological assessment of VLP-based immunogens exposing the membrane-proximal region of the HIV-1 gp41 protein. J Biomed Sci (2014) 21:79. doi:10.1186/s12929-014-0079-x
139. Donius LR, Cheng Y, Choi J, Sun Z-YJ, Hanson M, Zhang M, et al. Generation of long-lived bone marrow plasma cells secreting antibodies specific for HIV-1 gp41 MPER in the absence of polyreactivity. J Virol (2016) 90:8875–90. doi:10.1128/JVI.01089-16
140. Matyas GR, Wieczorek L, Beck Z, Ochsenbauer-Jambor C, Kappes JC, Michael NL, et al. Neutralizing antibodies induced by liposomal HIV-1 glycoprotein 41 peptide simultaneously bind to both the 2F5 or 4E10 epitope and lipid epitopes. AIDS (2009) 23:2069–77. doi:10.1186/1742-4690-6-S3-P80
141. Ye L, Wen Z, Dong K, Wang X, Bu Z, Zhang H, et al. Induction of HIV neutralizing antibodies against the MPER of the HIV envelope protein by HA/gp41 chimeric protein-based DNA and VLP vaccines. PLoS One (2011) 6:e14813. doi:10.1371/journal.pone.0014813
142. Leroux-Roels G, Maes C, Clement F, van Engelenburg F, van den Dobbelsteen M, Adler M, et al. Randomized phase I: safety, immunogenicity and mucosal antiviral activity in young healthy women vaccinated with HIV-1 Gp41 P1 peptide on virosomes. PLoS One (2013) 8:e55438. doi:10.1371/journal.pone.0055438
Keywords: human immunodeficiency virus type-1, broadly neutralizing antibodies, membrane proximal external region, B-cells, polyreactivity, membrane interaction, immunization, immunogens
Citation: Molinos-Albert LM, Clotet B, Blanco J and Carrillo J (2017) Immunologic Insights on the Membrane Proximal External Region: A Major Human Immunodeficiency Virus Type-1 Vaccine Target. Front. Immunol. 8:1154. doi: 10.3389/fimmu.2017.01154
Received: 11 July 2017; Accepted: 31 August 2017;
Published: 19 September 2017
Edited by:
Francesca Chiodi, Karolinska Institute (KI), SwedenReviewed by:
Guido Ferrari, Duke University, United StatesLucia Lopalco, San Raffaele Hospital (IRCCS), Italy
Copyright: © 2017 Molinos-Albert, Clotet, Blanco and Carrillo. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Jorge Carrillo, amNhcnJpbGxvQGlyc2ljYWl4YS5lcw==
†Present address: Luis M. Molinos-Albert, Laboratory of Humoral Response to Pathogens, Department of Immunology, Institut Pasteur, Paris, France