 Günther Schönrich
Günther Schönrich Martin J. Raftery
Martin J. Raftery- Berlin Institute of Health, Institute of Virology, Charité—Universitätsmedizin Berlin, Humboldt-Universität zu Berlin, Berlin, Germany
Some of the clinically most important viruses persist in the human host after acute infection. In this situation, the host immune system and the viral pathogen attempt to establish an equilibrium. At best, overt disease is avoided. This attempt may fail, however, resulting in eventual loss of viral control or inadequate immune regulation. Consequently, direct virus-induced tissue damage or immunopathology may occur. The cluster of differentiation 1 (CD1) family of non-classical major histocompatibility complex class I molecules are known to present hydrophobic, primarily lipid antigens. There is ample evidence that both CD1-dependent and CD1-independent mechanisms activate CD1-restricted T cells during persistent virus infections. Sophisticated viral mechanisms subvert these immune responses and help the pathogens to avoid clearance from the host organism. CD1-restricted T cells are not only crucial for the antiviral host defense but may also contribute to tissue damage. This review highlights the two edged role of CD1-restricted T cells in persistent virus infections and summarizes the viral immune evasion mechanisms that target these fascinating immune cells.
Introduction
The majority of virus infections are self-limiting. Immunocompetent hosts often eliminate the invading pathogen without causing permanent tissue damage. Some viruses, however, can resist clearance, and persist in the host organism in the face of intact antiviral immune responses. Persisting viruses belong to different RNA and DNA virus families and represent a global threat to human health (1). As a strategy for persistence, they utilize chronic infection, latency, or both. Chronic infection is characterized by the continuous generation of infectious virus particles as observed during infection with hepatitis B virus (HBV) (2) or hepatitis C virus (HCV) (3). During latent infection, production of viral progeny is put on ice while the viral genome replicates with the host DNA. In the latent stage, viruses are nearly invisible for the host defense but can reinitiate production of infectious viral particles. Primarily human papillomavirus (HPV) (4) and the human herpesviruses (HHVs) (5) establish latency. Human immunodeficiency virus (HIV) establishes both chronic and latent infection (6). The immunological challenges associated with persisting virus infections are distinct from acute self-limiting infections (7).
Virus persistence is largely the result of an evolutionary arms race between the host immune system and viral immune evasion mechanisms (8). For example, persisting viruses have learned to subvert the attack of cytotoxic T cells, which recognize viral peptides presented by major histocompatibility complex (MHC) class I molecules (9). Cluster of differentiation 1 (CD1) molecules are structurally related to MHC class I heavy chain molecules and also associate with β2-microglobulin (10–12). They are expressed in mammals, birds and reptiles, although isoforms, antigen-binding sites, recycling motifs and genomic locations are not well conserved (13, 14). In striking contrast to MHC class I molecules, CD1 molecules are non-polymorphic and present hydrophobic, primarily lipid antigens to specialized T cells (15, 16). These CD1-restricted T cells are at the frontline of the human immune response against pathogenic microbes including viruses (17). In this review, we discuss lipid-driven T cell responses during persistent virus infections and the corresponding viral counter measures.
CD1-Restricted T Cells
In humans CD1 molecules are divided into group 1 (CD1a, CD1b, CD1c) and group 2 (CD1d) by sequence homology (18). Group 1 genes can be induced in a coordinated fashion primarily in professional antigen-presenting cells such as dendritic cells (DCs). In contrast, CD1d is constitutively expressed in a wider range of hematopoietic and non- hematopoietic cells (19). CD1-restricted T cells belong to the “unconventional” T cells that do not recognize MHC-bound peptides, and which often show rapid effector functions and orchestrate other immune cells (20–23).
Group 1 CD1-restricted T cells express diverse αβ T-cell receptors (TCRs). They can undergo clonal expansion in the periphery after recognition of stimulatory self-lipids or exogenous lipid antigens derived from bacteria such as Mycobacterium tuberculosis (24–26). CD1d-restricted T cells are referred to as natural killer T (NKT) cells because they usually—but not always—express NK1.1 (CD161), a NK cell activating C-type lectin. These cells are further divided into type 1 and type 2 NKT cells.
Type 1 NKT or invariant NKT (iNKT) cells express a semi-invariant αβ TCR, defined by expression of the Vα14-Jα18 TCR in mice and Vα24-Jα18 TCR in humans. They have been analyzed in great detail [recently reviewed in Ref. (17, 27–30)]. The iNKT cells are optimally stimulated by α-galactosylceramide (αGalCer), a glycolipid antigen derived from marine sponge (31). Physiological ligands include cellular and microbial lipids (32). It has been shown that iNKT cells contribute to antiviral responses although the relevant lipid antigens have not yet been defined [recently reviewed in Ref. (33–37)].
Type 2 NKT cells are not stimulated by αGalCer and show much more TCR sequence variability than iNKT cells (38–40). They are more prevalent in humans than iNKT cells, show features of both conventional T cells and iNKT cells and also influence the outcome of infections with persisting viruses (38–41).
Group 1 CD1-Restricted T Cells
Group 1 CD1-restricted T cells have been analyzed much less intensively than NKT cells. They can be CD4+, CD8+, or double negative (DN) (42, 43). In vitro-studies revealed that after recognition of self or foreign lipid antigens group 1 CD1-restricted T cells become cytoloytic and secrete large amounts of cytokines such as IFN-γ and TNF-α (44–46). These cytokines have a profound antiviral effect and play a pivotal role in controlling persistent virus infections (47). Self-reactive group 1 CD1-restricted T cells are common and the frequency is comparable to alloreactive T cells (estimated to be 1/10 to 1/300 of circulating T cells) (42, 48). They can induce TNF-α dependent DC maturation (49). Intriguingly, self-reactive CD1b-restricted T cells can acquire the phenotype of T helper 17 (Th17) cells and recruit neutrophils (50). The autoreactivity of group 1 CD1-resticted T cells is enhanced by stimulation through pattern recognition receptors (PRRs) (51). It is probable that viruses also trigger activation of self-reactive group 1 CD1-restricted T cells through virus-sensing PRRs and that this contributes to antiviral immunity or virus-induced immunopathogenesis. This notion is supported by the finding that HHV-5, also called human cytomegalovirus (HCMV) can interfere with localization of group 1 CD1 molecules to the cell surface especially CD1b (52).
CD1d-Restricted T Cells
iNKT Cells
As part of the innate immune system iNKT cells perform multiple effector functions rapidly after being activated. Human iNKT cells comprise phenotypically and functionally distinct subsets: CD4+, a small fraction of CD4−CD8αα+ and DN cells (53). The CD4+ subset can produce Th2-type cytokines whereas both the CD4+ and CD4− subsets can secrete Th1-type cytokines and cytotoxic molecules such as perforin or granzyme B (54–56). The iNKT cells can contribute to both protection from pathogenesis and enhancement of disease (57). In mice, functionally polarized subsets of iNKT cells are generated in the thymus. They secrete Th1-, Th2-, or Th17-like cytokines (NKT1, NKT2, and NKT17 subsets) similar to MHC class II-restricted CD4+ T cells and innate lymphoid cells (58–60). In fact, these subsets and the corresponding MHC-restricted T cell subsets share the same transcription factors that regulate their function (61). Further subsets of iNKT cells that are potentially relevant during viral infections have been defined: follicular helper-like iNKT cells (62), IL-9-producing iNKT cells (63), regulatory (Foxp3+) iNKT cells (64, 65), and IL-10-producing iNKT cells called NKT10 cells (66).
The iNKT cells can lyse CD1d+ target cells, mostly through the interaction between CD95 (Fas) and CD178 (FasL) (67, 68). Virus-infected cells are eliminated by iNKT cells in a CD1d-dependent manner (69). In line with this notion, iNKT cells limit the number of Epstein–Barr virus (EBV)-transformed human B cells in vitro by a mechanism requiring direct contact with EBV-infected cells (70). EBV, a HHV that infects more than 90% of the human population worldwide, is associated with tumors, such as Burkitt lymphoma, Hodgkin disease, and lymphomas, in immunosuppressed patients (71). The iNKT cell frequency in human tissue is very low (approximately 0.1% in peripheral blood and spleen) (72). Additionally, in contrast to mice iNKT cells are not enriched in the human liver (73, 74). This suggests that human iNKT cells may be more important in helping orchestrate the antiviral immune response rather than in killing virus-infected cells.
There is ample evidence that iNKT cells attract, stimulate, and regulate other innate cells with antiviral effector functions such as NK cells and neutrophils (21, 22). NK cells are found in most compartments of the human organism at a higher frequency than iNKT cells and are critically involved in protection from persisting viruses (75, 76). The iNKT cells transactivate NK cells through the release of IFN-γ in mice (77–80) or IL-2 in humans (81). Moreover, iNKT cells can directly or indirectly recruit and activate neutrophils. These innate cells contribute to both antiviral defense and virus-induced immunopathogenesis (82–85). NK1.1-negative iNKT lymphocytes can directly recruit neutrophils through preferential IL-17 secretion (86, 87). The iNKT cells can also indirectly promote neutrophil responses by interacting with monocyte-derived DCs resulting in prolonged Ca+ influx and release of inflammatory mediators such as PGE2 (88). In mice infected with murine cytomegalovirus (MCMV), a well-established model of persistent herpesvirus infection, IL-22 secreted by iNKT cells contributes to recruitment of antiviral neutrophils expressing TNF-related apoptosis-inducing ligand (TRAIL) (85). Moreover, iNKT cells can reverse the suppressive phenotype of neutrophils that is induced by acute-phase reactant serum amyloid A-1 (89). Vice versa, neutrophils may reduce inflammatory iNKT cell responses by a contact-dependent mechanism thereby contributing to reduction of inflammation (90). These examples illustrate how intensely iNKT cells and other innate immune cells cross-regulate each other.
Another important antiviral function of iNKT cells is the regulation of adaptive immunity (91). For example, iNKT cells help B cells to proliferate and produce high titers of antibodies (92, 93). B cells present bacterial glycolipids through CD1d to iNKT cells which in turn differentiate into follicular helper-like iNKT cells that provide cognate help to B cells (62, 94–96). Recent evidence suggests that iNKT cells are also critical for production of specific antibodies during viral infections despite the absence of exogenous lipid antigens (97, 98). The underlying mechanism, however, is neither based on follicular helper-like iNKT cell differentiation nor CD1d-mediated cognate B cell help by iNKT cells but on tightly regulated early cytokine secretion by iNKT cells at the interfollicular area (97). On the other hand, iNKT cells licensed by a specialized neutrophil subset restrict activation of autoreactive B cells during inflammasome-driven inflammation through FasL (67).
In addition to helping B cells, iNKT cells promote T cell responses directed against intracellular pathogens such as viruses (79, 99–103). This is accomplished by iNKT-mediated DC maturation, which is of central importance for an efficient antiviral host defense (99, 104–106). It has also been reported that activated CD4−CD8αα+ iNKT cells curb expansion of antigen-stimulated conventional T cells by CD1d-dependent killing of DCs (107). Thus, iNKT cell subsets regulate adaptive immune responses at different stages.
The importance of crosstalk between iNKT cells and monocyte-derived DCs is underlined by the observation that both cell types are programmed to migrate to inflamed peripheral tissue (55, 108–110). Many persistent viruses such as herpes simplex virus type 1 (HSV-1), HSV-2 and HCMV target monocyte-derived DCs to induce apoptosis or impair antigen presentation through MHC class I molecules (111–113). As a consequence antiviral CD8+ T cells have to be activated by uninfected bystander DCs that take up apoptotic debris containing viral antigens and cross-present it through MHC class I molecules (114, 115). This cross-presentation, also called cross-priming, requires pathogen-associated molecular patterns (PAMPs) and/or specialized subsets of Th cells that mature or “license” the cross-presenting DCs (116). It has been shown that uninfected DCs can be licensed for cross-priming by iNKT cells (117). On the other hand, iNKT cells suppress CD8+ T cell responses to viral proteins expressed in skin epithelial cells by blocking cross-priming in the draining lymph nodes (118). This suppressive iNKT cell effect may increase the persistence of viruses such as HPV and HSV-1 in the skin. The effect of iNKT cells on CD8+ T cell priming (activating versus inhibitory) may be dependent on the type of antigen and the requirement for CD4+ T cell helper epitopes (118).
A recent study has shown that iNKT cells release the brake on the adaptive immune response against influenza virus by interfering with myeloid-derived suppressor cells (MDSCs) (119). MDSCs are in fact a heterogeneous population of myeloid progenitor cells including immature DCs, immature macrophages, and granulocytes (120). The iNKT cells abolish the suppressive activity of MDSCs in a crosstalk that requires CD1d and CD40. The finding by De Santo et al. is important as MDSCs facilitate the development of persistent virus infections by impairing antiviral functions of T cells, NK cells, and APCs (121).
Type 2 NKT Cells
Evidence that type 2 NKT cells play a protective role in virus infections came from a study that compared virus-induced disease severity between wild-type mice, Jα18 KO mice (lacking only type 1 NKT cells) and CD1d KO mice (lacking both type 1 and type 2 NKT cells) (122). In comparison to iNKT cells little is known about the function of type 2 NKT cells due to the lack of suitable animal models and the difficulty in tracking this poorly defined cell type in vivo (41). However, there is evidence that type 2 NKT cell subsets with distinct functional profiles exist (41, 123).
Clinical Observations
A number of clinical observations suggest involvement of CD1-restricted T cells in either the control of persisting viruses or virus-induced immunopathogenesis.
Group 1 CD1-Restricted T Cells
Group 1 CD1-restricted T cells from patients with active tuberculosis expand after reexposure to cognate antigen similar to adaptive MHC-restricted T cells (124). In HIV-infected patients, CD1b-restricted T cells recognizing mycobacterial glycolipids are strongly reduced (125). The reduced frequency of mycobacteria-specific CD1b-restricted T cells may contribute to the increased incidence of tuberculosis in this group. The kinetics of group 1 CD1-restricted T cells stimulated by virus-induced self-lipids is not understood. Self-reactive group 1 CD1-restricted T cells show an adaptive-like population dynamics (42). It is possible but has not yet been proven that group 1 CD1-restricted T cell reacting to virus-induced stimulatory self-lipids expand in a similar way (Figure 1A). On the other hand, self-reactive CD1b-restricted T cells have been described that are more like innate T cells (126). These cells may contribute to early antiviral host defense similar to iNKT cells.
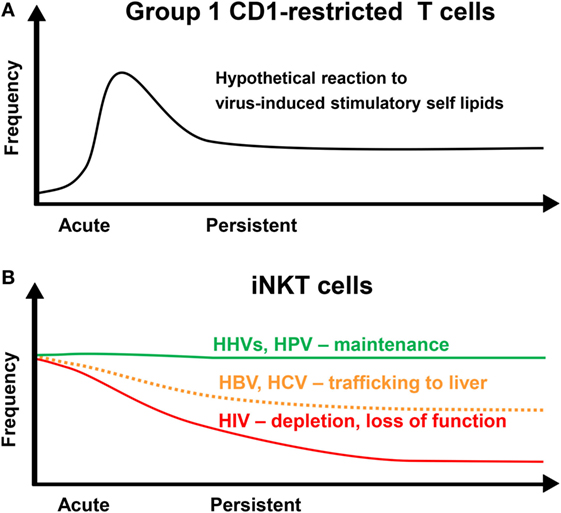
Figure 1. Dynamics of CD1-restricted T cells after persistent virus infections. (A) Group 1 CD1-restricted T cells are thought to undergo clonal expansion in response to an acute infection followed by a contraction phase leaving an increased population of memory cells. (B) The frequency of invariant NKT (iNKT) cells in the blood is maintained at a stable level over time in individuals infected with human herpesviruses (HHVs) or human papillomavirus (HPV). Human immunodeficiency virus (HIV) infection results in a loss of iNKT cells and a decrease in functionality. Hepatitis B virus (HBV) and hepatitis C virus (HCV) may result in a decrease in iNKT cell frequency due to redistribution into the liver.
CD1d-Restricted T Cells
Patients with defective iNKT cell responses suffer from severe infection with HHVs (127). For example, boys with X-linked lymphoproliferative disorder have severely impaired iNKT cell development and T cell function that results in life-threatening lymphoproliferation after primary infection with EBV or Kaposi’s sarcoma-associated herpesvirus (KSHV) (128–131). A similar disease was observed in girls with a homozygous mutation in the IL-2-inducible T cell kinase (ITK) gene on chromosome 5q31–5q32 (132). Moreover, patients with profound iNKT cell deficiency develop severe VZV-associated disease after vaccination with the live attenuated varicella vaccine (133, 134). Hermansky–Pudlak syndrome type 2, an autosomal recessive disease caused by mutations of the AP3B1 gene, encoding for the beta3A subunit of AP-3, also show reduced numbers of iNKT cells (135). Intriguingly, iNKT cell numbers decrease regardless of the HIV-status in patients with multicentric Castleman disease (MCD), a rare polyclonal lymphoproliferative disorder that is associated with KSHV (136). This observation suggests that a deficiency in iNKT cells contributes to the pathogenesis of KHSV-associated MCD. Although deficient iNKT cells can be regarded as the common denominator of these primary immundeficiencies it has to be kept in mind that other immune cells are affected as well.
In humans with an intact immune system, distinct dynamics of circulating iNKT cells are observed after infection with different persisting viruses. The frequency of peripheral blood iNKT cells after infection with persisting viruses that establish latent infection such as HHVs and HPV remain unaltered (Figure 1B) (98). In contrast, it has been reported that the frequencies of circulating iNKT cells are significantly decreased during chronic HBV and HCV infection and recover during antiviral therapy (Figure 1B) (137, 138). This decrease may be due to trafficking of iNKT cells to the liver as intrahepatic enrichment of iNKT cells is observed in patients with chronic viral hepatitis (137). In another study, however, iNKT cell frequencies between patients with chronic viral hepatitis and healthy individuals were not different (139). CD1d is strongly upregulated on hepatocytes and other liver cells during chronic viral hepatitis (137, 140). In fact, CD1d-restricted T cells contribute to virus-induced liver injury by killing hepatocytes and production of pro-inflammatory cytokines, which promote liver fibrosis and cirrhosis (74, 137, 141–144).
Several studies have shown that iNKT cells are depleted during HIV infection (145–152) (Figure 1B). The remaining iNKT cells display an exhausted phenotype with reduced functionality that may improve with early antiretroviral therapy (152–157). In fact, the level of iNKT cell activation in HIV-infected patients is associated with disease progression markers and the number and functionality of iNKT cells are preserved in non-progressive HIV infection (155, 158). Intriguingly, both CD4+ and CD4− iNKT cells are depleted in HIV-infected individuals (149) although CD4− iNKT cells are resistant to HIV infection (148, 150). This could be due to apoptosis of uninfected bystander cells. Triggering of both extrinsic and intrinsic death pathways in T cells due to systemic immune activation has been described in many studies (159).
CD1-Restricted T Cells in Animal Models of Persistent Virus Infections
Group 1 CD1-Restricted T Cells
The function of group 1 CD1-restricted T cells has proven difficult to address in vivo as laboratory mice do not express group 1 CD1 molecules (160–162). In contrast, mice with a humanized immune system express human group 1 CD1 molecules (163) and open up a new avenue for studying human immune responses to viral pathogens (164). Similarly, human group 1 CD1 transgenic mice have recently been developed (126, 165, 166). Future studies have to investigate a possible role of group 1 CD1-restricted T cells in persisting virus infections by using suitable animal models and tetramers.
CD1d-Restricted T Cells
The protective and pathogenic role of CD1d-restricted T cells has been investigated in several animal models of persistent virus infections (see Table 1).

Table 1. Studies of CD1-restricted T cells in animal models.
Herpesviral Infection
Studies in CD1d KO and iNKT cell-deficient (Jα18 KO) mice indicate that CD1d-restricted T cells have a protective role after infection with HSV-1 (98, 167, 168) or HSV-2 (169). In addition, intravaginal pretreatment of C57BL/6 mice with αGalCer lowers HSV-2 disease and inhibits viral replication (170). One report did not confirm an antiviral role for NKT cells, most likely because mice were infected with a less virulent HSV-1 strain (171).
In mice infected with MCMV, activation of iNKT cells by exogenous αGalCer reduces viral replication (172). Furthermore, it has been shown in the MCMV model that iNKT cells can protect from MCMV-induced myelosuppression (173). In C57BL/6 mice NK cells can compensate for iNKT deficiency (172, 174, 175) whereas in BALB/c mice NK cells are less well activated by MCMV and thus iNKT deficiency results in higher viral loads (176).
Chronic Viral Hepatitis
In contrast to HHVs, HBV, and HCV infect target cells without causing cytopathic effects. Injection of αGalCer into HBV transgenic mice activates intrahepatic resident iNKT cells and NK cells resulting in noncytopathic control of HBV replication through secretion of type I and II IFN (80, 177). Hepatocytes directly control iNKT cell homeostasis through modulating the balance between activating and non-activating lipids presented by CD1d molecules (178). Accordingly, iNKT cells act as an early warning system for HBV infection, which profoundly alters lipid metabolism (179). For example, type 2 NKT cells recognize CD1d-bound lysophospholipids, antigenic self-lipids that are generated in hepatocytes by a HBV-induced secretory phospholipase (180). The resulting activation of type 2 NKT cells also leads to IL-12 mediated transactivation of iNKT cells and plays a pivotal role in virus control (180). However, type 2 NKT cells can also cause liver injury in HBV transgenic mice (181, 182). It has to be kept in mind that there are major differences between human and murine liver NKT cells when extrapolating these results to humans (73, 74). Nevertheless, human NKT cell lines are stimulated in a CD1d-dependent manner by human hepatocytes (180). Taken together, these findings support clinical observations suggesting a role for CD1d-restricted T cells in antiviral immunity and virus-induced immunopathogenesis in human liver.
HIV Infection
In a humanized mouse model of HIV-1 infection activation of type 2 NKT cells inhibits viral replication and prevents virus-induced pancytopenia (183). In Asian macaques, which develop acquired immunodeficiency syndrome (AIDS) during persistent simian immunodeficiency virus (SIV) infection, NKT cell depletion inversely correlates with viral load (184). Comparative studies of Asian macaques and sooty mangabeys, which serve as a natural host and do not develop AIDS during persistent SIV infection, suggest that NKT cells protect from SIV-induced immune activation and immunodeficiency (185).
Activation of CD1-Restricted T Cells
In contrast to other microbes such as bacteria, viruses have not been investigated with regard to CD1 ligands, although viral infection can stimulate CD1-restricted antiviral T cells. Activation of CD1-restricted T cells during viral infection could be triggered by CD1 molecules presenting antigenic self-lipids. In addition, CD1-independent mechanisms that activate CD1-restricted T cells such as cytokine release have been described.
Group 1 CD1-Restricted T Cells
Autoreactive group 1 CD1-restricted T cells are stimulated by DCs that express group 1 CD1 molecules and present increased amounts of antigenic self-lipids upon activation by bacterial PAMPs through TLRs (186, 187). It is very likely but has not yet been shown that group 1 CD1-restricted autoreactive T cells are activated in a similar fashion during viral infections upon activation of DCs through virus-sensing PRRs.
CD1d-Restricted T Cells
CD1d-Dependent Activation
The limited repertoire of microbial and endogenous ligands presented by CD1d is expanding (188–190). However, the identity of self-lipids presented during viral infection is still enigmatic. Stimulatory self-lipids could be channeled into the CD1d-restricted antigen presentation pathway by several distinct mechanisms. First, there is ample evidence that TLR stimulation increases CD1d presentation of antigenic self-lipids resulting in iNKT cell activation (191–197). Besides modulating the repertoire of self-lipids, viruses also increase CD1 surface expression through triggering PRRs (198). Second, viral invasion and inflammation is associated with pathological hypoxia that activates hypoxia-inducible factor (HIF), a “master regulator” of adaptive immune responses (199, 200). HIF alters lipid metabolism in such a way that self-reactive NKT cells are activated (201, 202). Third, viruses profoundly rewire host lipid metabolism and remodel lipid distribution to boost in a coordinated fashion viral entry, replication, assembly, and egress (203–208). Disturbance of the normal lipid trafficking patterns within the cell due to enveloped virus production (HHVs, HBV, HCV) or even naked virus (HPV) will allow access of otherwise sequestered lipids or altered lipids to the relevant CD1 presenting compartment. Furthermore, after assembly and release from the infected host cell the envelope of the newly built viral particle may carry the ligand with it and deliver it during infection to another host cell. Taken together, several distinct mechanisms facilitate CD1d-dependent iNKT cell activation during different stages of the viral life cycle.
Of note, viral immune evasion mechanisms targeting other immune components may alert iNKT cells. For example, MHC class I downregulation by virus-encoded immunevasins induces CD1d upregulation and enhances NKT cell activation (209). This finding is supported by other observations. First, low pH stripping of MHC class I molecules augments CD1d surface expression and activates iNKT cells (210). Second, APCs from transporter associated with antigen presentation 1 (TAP1)-deficient mice are defective in MHC class I antigen presentation but show an increased capacity to stimulate iNKT cells (210, 211). This reverse correlation between CD1d and MHC class I surface expression may enable iNKT cells to detect the loss of MHC class I molecules, a situation called “missing self” (212). The underlying mechanism is not yet fully understood but may involve masking of CD1d by MHC class I molecules on the cell surface (210). Increased iNKT cell activation during persistent virus infections may also result from viral blockers inhibiting the autophagic machinery, which downregulates iNKT cell responses through CD1d internalization (213, 214).
CD1d-Independent Activation
CD1d-restricted T cells can also be stimulated independently of the TCR. First, there is ample evidence that pro-inflammatory cytokines (IL-12, IL-18, or type I IFN) released from APCs after stimulation through PRRs activate iNKT cells predominantly in a CD1d-independent manner (215, 216). It may be that an initial CD1d-mediated TCR signal is still required (217). CD1d-independent iNKT cell activation was observed in mice infected with MCMV, and was driven by type I IFN and IL-12 after DC stimulation (176, 218–220). The factors that determine whether pro-inflammatory cytokines are sufficient for activation of antiviral iNKT cells are unclear.
Second, engagement of NKG2D on NKT cells results in CD1d-independent activation and subsequent killing of ligand-expressing target cells (182, 221). NKG2D is an activating receptor expressed on CD4− iNKT cells and is triggered by stress ligands. The latter are upregulated during viral infections after stimulation of virus-sensing PRRs such as RIG-I and MDA-5 (222, 223). In addition, NKG2D can also co-stimulate TCR-mediated activation of CD4− iNKT cells (221).
Third, iNKT cells can be activated by apoptotic cells through TIM-1, a member of the T-cell immunoglobulin mucin (TIM) family of cell surface proteins (224). TIM-1 is constitutively expressed by NKT cells and serves as a receptor for phosphatidylserine, an important marker of cells undergoing apoptosis (225). This mode of iNKT cell activation may be relevant during persistent virus infections because viruses frequently drive their host cells into apoptosis (226).
Viral Evasion of CD1-Restricted T Cells
During the evolutionary arms race with the host immune system persisting viruses have developed multilayered defense strategies to interfere with antiviral CD1-restricted T cells (17, 33, 35, 36, 227). They target the CD1 antigen presentation machinery either on the transcriptional, posttranscriptional, or posttranslational level.
Group 1 CD1-Restricted T Cells
The group 1 CD1 molecules are expressed on professional APCs in response to certain cytokines such as granulocyte-macrophage colony-stimulating factor (228). The anti-inflammatory cytokine IL-10 prevents upregulation of group 1 CD1 molecules on professional APCs such as monocyte-derived DCs (229–231). In fact, the activity of IL-10 is crucial for establishing viral persistence (232). Large DNA viruses such as herpesviruses have acquired numerous genes from their host including viral homologs of IL-10 (vIL-10) to subvert the host immune response (233, 234). For example, vIL-10 from HCMV and EBV not only prevent upregulation of MHC class I/II molecules but also group 1 CD1 molecules (52, 235). Recent reports suggest that vIL-10 strongly induces the expression of its cellular counterpart in monocyte-derived cells thereby potentiating its immunomodulatory effect (236, 237). In addition, viral proteins with no homology to cellular proteins such as HCMV-encoded pUL11 or EBV-encoded latent membrane protein 1 highjack cellular signaling pathways to induce expression of cellular IL-10 (238, 239).
Group 1 CD1 molecules are downregulated from the cell surface during the early phase of HCMV infection with CD1b being especially sensitive to this effect (52). This block is not performed by known HCMV-encoded MHC class I-blocking molecules and results in the intracellular accumulation of group 1 CD1 molecules (52). The underlying mechanism has not yet been described. In addition, HIV-1 negative factor (Nef) downregulates CD1a after infection of immature DCs with HIV-1 and impairs stimulation of CD1a-restricted T cells (240, 241). In order to modulate host membrane trafficking pathways HIV-1 Nef has to interact with various host proteins through distinct motifs (242). Nef-mediated CD1a downregulation might require the interaction of Nef with hematopoietic cell kinase and p21-activated kinase 2, which are both expressed in immature DCs (241). Collectively, these findings support the idea that group 1 CD1 molecules contribute to lipid-driven antiviral immune responses.
CD1d-Restricted T Cells
Viral Downregulation of CD1d Gene Transcription
In contrast to group 1 CD1 molecules, CD1d is constitutively expressed not only in myeloid cells such as B cells, macrophages, and DCs and but also in epithelial tissue. Latent EBV infection that is associated with transformation of B cells results in complete shutdown of CD1d mRNA expression and the lack of iNKT cell activation (70). This is due to increased binding of lymphoid enhancer-binding factor 1 to the distal region of the CD1d promotor (Figure 2). Moreover, CD1d transcription is downregulated during severe primary HCMV infection (243).
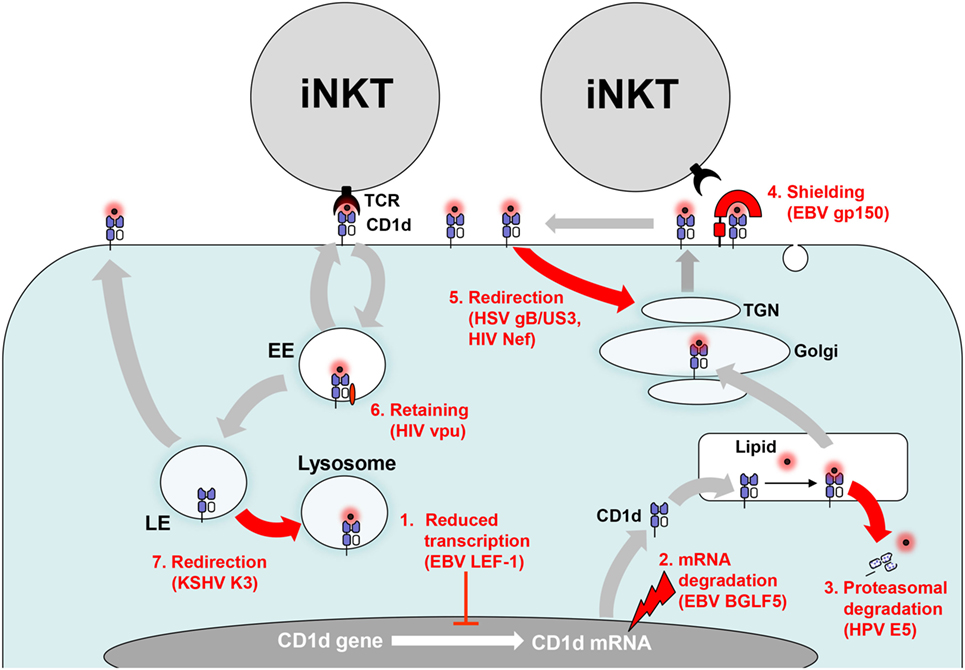
Figure 2. Evasion of CD1d antigen presentation. Persisting viruses evade invariant NKT (iNKT) cell activation by interfering with CD1d biosynthesis and CD1d trafficking. (1) CD1d gene transcription is downregulated by increased binding of Epstein–Barr virus (EBV)-encoded lymphoid enhancer-binding factor 1 (LEF-1) to the distal region of the CD1d promotor. (2) Viral host shutoff factors such as EBV BGLF5, an early lytic phase protein, inhibit protein synthesis by degrading mRNA. (3) Human papillomavirus (HPV) E5 translocates CD1d into the cytosol for proteasomal degradation. (4) EBV gp150 provides an abundantly sialylated glycan shield for CD1d on the cell surface preventing recognition by iNKT cells. (5) Herpes simplex virus type 1 (HSV-1) glycoprotein B/US3 and human immunodeficiency virus (HIV) negative factor (Nef) redirect CD1d to the trans-Golgi network (TGN). (6) In contrast, HIV Nef retains CD1d in the early endosome (EE). (7) Kaposi’s sarcoma-associated herpesvirus (KSHV) K3 redirects CD1d from the late endosome (LE) to the lysosome.
Virus-Induced Degradation of CD1d mRNA
Replicating α- and γ-herpesviruses induce global mRNA degradation to shut off host protein synthesis and reallocate cellular resources to their own need by different mechanisms (244). It has been shown that EBV-encoded shutoff protein BGLF5, a lytic phase protein, downregulates multiple immune components including CD1d (Figure 2) (245, 246).
Viral Interference With Trafficking of CD1d Molecules
After translation, CD1 heavy chains bind to β2-microglobulin and are loaded with self-lipids in the endoplasmic reticulum (ER). This trimeric complex travels then to the cell surface through the secretory pathway. Subsequently, a tyrosine motif within their cytoplasmic tail enables human CD1 molecules to bind the μ-subunit of adaptor protein complex (AP)-2 (CD1b-d) and AP-3 (CD1b and murine CD1d). In contrast, CD1a lacks AP-2 and AP-3 sorting motifs. Because of these differences CD1 molecules traffic through distinct intracellular compartments for acquiring lipids and present these lipids to CD1-restricted T cells (247–249). Persistent viruses utilize various posttranslational mechanisms to interfere with antigen presentation through CD1d molecules in different cellular compartments.
Virus-Induced Proteasomal Degradation of CD1d in the Cytosol
HPV subverts immune responses through expression of E5 (250). Clinical samples of HPV-infected cervical epithelium show decreased CD1d expression (251). Further analysis of transfected cell lines revealed that HPV E5 interacts with calnexin, a chaperone involved in CD1d folding in the ER, and finally targets CD1d to the cytosolic proteolytic pathway (Figure 2) (251).
Viral Shielding of Surface CD1d Molecules
A novel broadly acting mechanism of herpesviral subversion of antigen presentation has been discovered recently (252). EBV-encoded gp150, which is heavily glycosylated and expressed in the late phase of EBV replication, inhibits antigen presentation by MHC class I and II as well as CD1d molecules. The EBV-encoded gp150 does not interfere with recycling of CD1d molecules but prevents their detection by iNKT cells on the cell surface through an abundantly sialylated glycan shield (Figure 2) (252). Of interest, the Ebola virus glycoprotein masks MHC class I molecules by a similar mechanism but an effect on CD1d has not yet been tested (253). Further studies have to elucidate whether other herpesviruses have developed are similar strategy to mask antigen-presenting molecules on the cell surface.
Viral Modulation of CD1d Recycling
Persisting viruses can also subvert CD1 antigen presentation by interfering with CD1 recycling. For example, CD1d surface expression is downregulated during productive infection with HHVs such as HSV-1 (209, 254) and KSHV (255). This effect was dependent on the viral titer used for infection of CD1d-expressing cells (209).
Further attempts identified several viral proteins interfering with CD1d recycling. For example, KSHV-encoded modulator of immune recognition proteins 1 and 2 (also known as K3 and K5) function as membrane-bound E3-ubiquitin ligases and reduce expression of a number of immunologically important surface molecules including MHC class I and CD1d (256). In fact, K3 and K5 have been pilfered from the host genome and belong to the Membrane Associated RING-CH family of E3 ligase (MARCH) proteins. They reroute both MHC class I and CD1d to the lysosomal compartment by ubiquitinylating a unique lysine residue on the cytoplasmic tail (Figure 2) (255). In contrast to MHC class I, however, the CD1d complexes are resistant to lysosomal degradation resulting in reduced CD1d levels on KSHV-infected B cells although the total cellular amount of CD1d is not altered (255).
A recent study shows that CD1d molecules on the APC surface build nanoclusters and that these structures are important for iNKT cell activation (257). Intriguingly, the actin cytoskeleton prevents CD1d nanoclustering (257). This finding is in accordance with a previous study showing that disruption of actin filaments increases CD1d antigen presentation (258). It may also explain why HSV-1 lacking VP22 (UL49), a viral tegument protein that stabilizes microtubules, lacks the ability to inhibit CD1d recycling (259). Although VP22 is necessary for HSV-1 induced CD1d downregulation, other viral proteins are additionally required. The type II kinesin motor protein KIF3A, which transports proteins along the microtubule network, is necessary for CD1d surface expression (260). Of relevance, KIF3A is phosphorylated by US3, a HSV-1 encoded protein that induces downregulation of CD1d by suppressing its recycling (261). Another HSV-1 protein, glycoprotein B (gB), binds to CD1d within the ER and remains stably associated throughout CD1d trafficking (261). Both US3 and gB seem to be required for relocalization of CD1d to the trans-Golgi network (TGN) thereby reducing CD1d surface expression and iNKT cell activation (Figure 2) (261). In contrast, a previous study has reported that CD1d is trapped in lysome-like structures during HSV-1 infection (254). Similarly, another study reported that CD1d molecules are degraded in the lysosomes after HSV-1 induced phosphorylation of a dual residue motif (T322/S323) in the cytoplasmic tail of CD1d (262, 263). However, under normal conditions CD1d complexes are resistant to lysosomal degradation (255, 264).
HIV-1 encodes two proteins, Nef and viral protein U (Vpu) which target numerous immunologically important surface molecules including CD1d (265). In HIV-1 infected DCs, Vpu neither induces endocytosis nor rapid degradation but suppresses CD1d recycling by retaining CD1d in the early endosome (196, 266, 267). In contrast, Nef increases CD1d internalization and re-localizes CD1d molecules to the TGN (196, 268, 269). Thus, Vpu and Nef interfere with iNKT cell activation by complementary mechanisms (Figure 2). As a result, the capacity of HIV-1 infected DCs to stimulate iNKT cells is impaired (196).
The unique short (US) region of HCMV encodes several proteins including US2 and US11 that interfere with antigen presentation through MHC class I molecules (270). US2 and US11 induce rapid translocation of MHC class I heavy chains from the ER into the cytosol and subsequent degradation by the proteasome (271, 272). US2 and US11 also physically interact with CD1d but do not downregulate CD1d surface expression (273, 274). However, reduced activation of iNKT cells after stimulation with αGalCer-pulsed US2-expressing APCs has been reported but the underlying mechanism is unclear (274).
Viral Disruption of the iNKT Cell–DC Axis
The bidirectional interaction between DCs and iNKT cells is crucial for an efficient antiviral immune response (28, 275). During infection triggering of a combination of PRRs stimulates IL-12 release from DCs and increases the presentation of antigenic self-lipids by CD1d (191, 195, 197). In fact, signaling through PRRs inhibits degradation of glyosylceramides in the lysosome thereby increasing the amount of lipids that stimulate iNKT cells (193, 276). Once activated iNKT cells further enhance IL-12 production by DCs through CD40–CD40 ligand stimulation. This amplification loop then results in NK cell transactivation and increased responses of MHC-restricted CD4+ as well as CD8+ T cell responses. Persisting viruses such as HHVs use several different strategies to blunt DC function (277). For example, VZV interferes with TLR signaling in DCs thereby decreasing IL-12 production (278). Moreover, herpesviral IL-10 homologs can decrease the ability of uninfected DCs to secrete IL-12 (234). Virus-infected DCs that do not produce biologically active IL-12 fail to activate the antiviral functions of iNKT cells (279). Moreover, HCMV-infected DCs upregulate on the surface death ligands such as TRAIL that kill T cells (113). Taken together, it is likely that multilayered viral countermeasures severely impair activation and function of iNKT cells thereby disrupting the important iNKT cell–DC axis.
Viral Interference With CD1d-Independent Activation of CD1d-Restricted T Cells
Persistent viruses such as HHVs also interfere with NKG2D ligand upregulation on virus-infected cells thereby reducing the likelihood of NKG2D-mediated activation of antiviral iNKT and type 2 NKT cells (280, 281). Moreover, HSV-1-infected keratinocytes block cytokine-dependent activation of iNKT cells (282).
Virus-Induced Functional Impairment of CD1d-Restricted T Cells
Persisting viruses not only target CD1 antigen presentation and interfere with activation of CD1-restricted T cells but can also induce a state of unresponsiveness in CD1-restricted T cells. It has been demonstrated in vitro that HSV-1-infected keratinocytes impair TCR signaling in iNKT cells in a contact-dependent manner (282). In contrast to DCs, CD1d is not downregulated on keratinocytes after infection with HSV-1. This finding nicely illustrates that depending on the infected cell type one and the same virus use different strategies to evade CD1d-restricted T cell responses. Moreover, iNKT cells from patients with KSHV-associated MCD, a severe B-cell lymphoproliferative disorder, show impaired proliferation when stimulated with αGalCer (136). This hyporeactive state is indicative of virus-induced iNKT cell anergy or exhaustion. In accordance, induction of anergy in iNKT cells has been described in many reports and is required for prevention of uncontrolled inflammation and tissue destruction (283). On the other hand, it also allows viruses to replicate and spread more efficiently. The underlying mechanisms of virus-induced iNKT cell anergy are unknown but may involve viral modulation of costimulatory molecules and other signaling molecules on APCs (284).
Recently, a regulatory iNKT cell subset called NKT10 has been described that bear similarities to Tregs (66). Tregs suppress protective immune responses thereby supporting virus persistence and at the same time also reducing the inflammation-mediated tissue damage (285). The NKT10 cells occur naturally in mice and humans and expand after stimulation (66). It would be interesting to investigate whether persisting viruses increase the frequency and activity of NKT10 cells to curb antiviral immune responses.
Conclusion and Future Directions
The risk for the host in terms of severe disease from a persistent virus infection ranges from low in case of well-adapted viruses such as the HHVs to ineluctable in case of a recent émigré such as HIV-1. CD1-reactive cells play a significant role in this process. Although the role of self-reactive NKT cells in antiviral immunity and immunopathogenesis has been analyzed in the clinical setting and in experimental models, it is unclear how group 1 CD1-restricted T cells contribute to antiviral immunity. Future efforts should define in detail the antigenic self-lipids that are induced and loaded on CD1 molecules during viral infections and how they program antiviral immunity. This knowledge can then be harnessed to develop novel vaccines and adjuvants for protection from persistent virus infection.
Author Contributions
Both authors contributed to the conception, writing, and critical revising of this review.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Funding
The authors acknowledge support from the German Research Foundation (DFG) and the Open Access Publication Fund of Charité—Universitätsmedizin Berlin.
References
1. Boldogh I, Albrecht T, Porter DD. Persistent viral infections. In: Baron S, editor. Medical Microbiology. 4th ed. Galveston, TX: University of Texas Medical Branch at Galveston (1996).
2. Stanaway JD, Flaxman AD, Naghavi M, Fitzmaurice C, Vos T, Abubakar I, et al. The global burden of viral hepatitis from 1990 to 2013: findings from the Global Burden of Disease study 2013. Lancet (2016) 388(10049):1081–8. doi:10.1016/S0140-6736(16)30579-7
3. Mohd Hanafiah K, Groeger J, Flaxman AD, Wiersma ST. Global epidemiology of hepatitis C virus infection: new estimates of age-specific antibody to HCV seroprevalence. Hepatology (2013) 57(4):1333–42. doi:10.1002/hep.26141
4. zur Hausen H. Papillomaviruses and cancer: from basic studies to clinical application. Nat Rev Cancer (2002) 2(5):342–50. doi:10.1038/nrc798
5. Pellett PE, Roizman B. Herpesviridae. In: Knipe DM, Howley PM, Griffin DE, Lamb RA, Martin MA, Roizman B, editors. Fields Virology. 6th ed. Vol. 2. Philadelphia, PA: Lippincott Williams & Wilkins (2013). p. 1802–22.
6. Maartens G, Celum C, Lewin SR. HIV infection: epidemiology, pathogenesis, treatment, and prevention. Lancet (2014) 384(9939):258–71. doi:10.1016/S0140-6736(14)60164-1
7. Virgin HW, Wherry EJ, Ahmed R. Redefining chronic viral infection. Cell (2009) 138(1):30–50. doi:10.1016/j.cell.2009.06.036
8. Kane M, Golovkina T. Common threads in persistent viral infections. J Virol (2010) 84(9):4116–23. doi:10.1128/JVI.01905-09
9. van de Weijer ML, Luteijn RD, Wiertz EJ. Viral immune evasion: lessons in MHC class I antigen presentation. Semin Immunol (2015) 27(2):125–37. doi:10.1016/j.smim.2015.03.010
10. Calabi F, Milstein C. A novel family of human major histocompatibility complex-related genes not mapping to chromosome 6. Nature (1986) 323(6088):540–3. doi:10.1038/323540a0
11. Martin LH, Calabi F, Milstein C. Isolation of CD1 genes: a family of major histocompatibility complex-related differentiation antigens. Proc Natl Acad Sci U S A (1986) 83(23):9154–8. doi:10.1073/pnas.83.23.9154
12. McMichael AJ, Pilch JR, Galfre G, Mason DY, Fabre JW, Milstein C. A human thymocyte antigen defined by a hybrid myeloma monoclonal antibody. Eur J Immunol (1979) 9(3):205–10. doi:10.1002/eji.1830090307
13. Reinink P, Van Rhijn I. Mammalian CD1 and MR1 genes. Immunogenetics (2016) 68(8):515–23. doi:10.1007/s00251-016-0926-x
14. Rogers SL, Kaufman J. Location, location, location: the evolutionary history of CD1 genes and the NKR-P1/ligand systems. Immunogenetics (2016) 68(8):499–513. doi:10.1007/s00251-016-0938-6
15. Beckman EM, Porcelli SA, Morita CT, Behar SM, Furlong ST, Brenner MB. Recognition of a lipid antigen by CD1-restricted alpha beta+ T cells. Nature (1994) 372(6507):691–4. doi:10.1038/372691a0
16. Porcelli S, Morita CT, Brenner MB. CD1b restricts the response of human CD4-8- T lymphocytes to a microbial antigen. Nature (1992) 360(6404):593–7. doi:10.1038/360593a0
17. Crosby CM, Kronenberg M. Invariant natural killer T cells: front line fighters in the war against pathogenic microbes. Immunogenetics (2016) 68(8):639–48. doi:10.1007/s00251-016-0933-y
18. Calabi F, Jarvis JM, Martin L, Milstein C. Two classes of CD1 genes. Eur J Immunol (1989) 19(2):285–92. doi:10.1002/eji.1830190211
19. Dougan SK, Kaser A, Blumberg RS. CD1 expression on antigen-presenting cells. Curr Top Microbiol Immunol (2007) 314:113–41.
20. Godfrey DI, Uldrich AP, McCluskey J, Rossjohn J, Moody DB. The burgeoning family of unconventional T cells. Nat Immunol (2015) 16(11):1114–23. doi:10.1038/ni.3298
21. Mori L, Lepore M, De Libero G. The immunology of CD1- and MR1-restricted T cells. Annu Rev Immunol (2016) 34:479–510. doi:10.1146/annurev-immunol-032414-112008
22. Salio M, Silk JD, Jones EY, Cerundolo V. Biology of CD1- and MR1-restricted T cells. Annu Rev Immunol (2014) 32:323–66. doi:10.1146/annurev-immunol-032713-120243
23. Van Rhijn I, Godfrey DI, Rossjohn J, Moody DB. Lipid and small-molecule display by CD1 and MR1. Nat Rev Immunol (2015) 15(10):643–54. doi:10.1038/nri3889
24. Dellabona P, Consonni M, de Lalla C, Casorati G. Group 1 CD1-restricted T cells and the pathophysiological implications of self-lipid antigen recognition. Tissue Antigens (2015) 86(6):393–405. doi:10.1111/tan.12689
25. Siddiqui S, Visvabharathy L, Wang CR. Role of group 1 CD1-restricted T cells in infectious disease. Front Immunol (2015) 6:337. doi:10.3389/fimmu.2015.00337
26. Van Rhijn I, van Berlo T, Hilmenyuk T, Cheng TY, Wolf BJ, Tatituri RV, et al. Human autoreactive T cells recognize CD1b and phospholipids. Proc Natl Acad Sci U S A (2016) 113(2):380–5. doi:10.1073/pnas.1520947112
27. Berzins SP, Ritchie DS. Natural killer T cells: drivers or passengers in preventing human disease? Nat Rev Immunol (2014) 14(9):640–6. doi:10.1038/nri3725
28. Brennan PJ, Brigl M, Brenner MB. Invariant natural killer T cells: an innate activation scheme linked to diverse effector functions. Nat Rev Immunol (2013) 13(2):101–17. doi:10.1038/nri3369
29. Kohlgruber AC, Donado CA, LaMarche NM, Brenner MB, Brennan PJ. Activation strategies for invariant natural killer T cells. Immunogenetics (2016) 68(8):649–63. doi:10.1007/s00251-016-0944-8
30. Van Kaer L, Parekh VV, Wu L. Invariant natural killer T cells as sensors and managers of inflammation. Trends Immunol (2013) 34(2):50–8. doi:10.1016/j.it.2012.08.009
31. Kawano T, Cui J, Koezuka Y, Toura I, Kaneko Y, Motoki K, et al. CD1d-restricted and TCR-mediated activation of valpha14 NKT cells by glycosylceramides. Science (1997) 278(5343):1626–9. doi:10.1126/science.278.5343.1626
32. Rossjohn J, Pellicci DG, Patel O, Gapin L, Godfrey DI. Recognition of CD1d-restricted antigens by natural killer T cells. Nat Rev Immunol (2012) 12(12):845–57. doi:10.1038/nri3328
33. Chung BK, Priatel JJ, Tan R. CD1d expression and invariant NKT cell responses in herpesvirus infections. Front Immunol (2015) 6:312. doi:10.3389/fimmu.2015.00312
34. Diana J, Lehuen A. NKT cells: friend or foe during viral infections? Eur J Immunol (2009) 39(12):3283–91. doi:10.1002/eji.200939800
35. Juno JA, Keynan Y, Fowke KR. Invariant NKT cells: regulation and function during viral infection. PLoS Pathog (2012) 8(8):e1002838. doi:10.1371/journal.ppat.1002838
36. Opasawatchai A, Matangkasombut P. iNKT cells and their potential lipid ligands during viral infection. Front Immunol (2015) 6:378. doi:10.3389/fimmu.2015.00378
37. Tessmer MS, Fatima A, Paget C, Trottein F, Brossay L. NKT cell immune responses to viral infection. Expert Opin Ther Targets (2009) 13(2):153–62. doi:10.1517/14712590802653601
38. Dasgupta S, Kumar V. Type II NKT cells: a distinct CD1d-restricted immune regulatory NKT cell subset. Immunogenetics (2016) 68(8):665–76. doi:10.1007/s00251-016-0930-1
39. Dhodapkar MV, Kumar V. Type II NKT cells and their emerging role in health and disease. J Immunol (2017) 198(3):1015–21. doi:10.4049/jimmunol.1601399
40. Marrero I, Ware R, Kumar V. Type II NKT cells in inflammation, autoimmunity, microbial immunity, and cancer. Front Immunol (2015) 6:316. doi:10.3389/fimmu.2015.00316
41. Macho-Fernandez E, Brigl M. The extended family of CD1d-restricted NKT cells: sifting through a mixed bag of TCRs, antigens, and functions. Front Immunol (2015) 6:362. doi:10.3389/fimmu.2015.00362
42. de Lalla C, Lepore M, Piccolo FM, Rinaldi A, Scelfo A, Garavaglia C, et al. High-frequency and adaptive-like dynamics of human CD1 self-reactive T cells. Eur J Immunol (2011) 41(3):602–10. doi:10.1002/eji.201041211
43. Young MH, Gapin L. Group 1 CD1-restricted T cells take center stage. Eur J Immunol (2011) 41(3):592–4. doi:10.1002/eji.201141408
44. Kawashima T, Norose Y, Watanabe Y, Enomoto Y, Narazaki H, Watari E, et al. Cutting edge: major CD8 T cell response to live bacillus Calmette-Guerin is mediated by CD1 molecules. J Immunol (2003) 170(11):5345–8. doi:10.4049/jimmunol.170.11.5345
45. Ulrichs T, Moody DB, Grant E, Kaufmann SH, Porcelli SA. T-cell responses to CD1-presented lipid antigens in humans with Mycobacterium tuberculosis infection. Infect Immun (2003) 71(6):3076–87. doi:10.1128/IAI.71.6.3076-3087.2003
46. Vincent MS, Xiong X, Grant EP, Peng W, Brenner MB. CD1a-, b-, and c-restricted TCRs recognize both self and foreign antigens. J Immunol (2005) 175(10):6344–51. doi:10.4049/jimmunol.175.10.6344
47. Guidotti LG, Chisari FV. Noncytolytic control of viral infections by the innate and adaptive immune response. Annu Rev Immunol (2001) 19:65–91. doi:10.1146/annurev.immunol.19.1.65
48. de Jong A, Pena-Cruz V, Cheng TY, Clark RA, Van Rhijn I, Moody DB. CD1a-autoreactive T cells are a normal component of the human alphabeta T cell repertoire. Nat Immunol (2010) 11(12):1102–9. doi:10.1038/ni.1956
49. Vincent MS, Leslie DS, Gumperz JE, Xiong X, Grant EP, Brenner MB. CD1-dependent dendritic cell instruction. Nat Immunol (2002) 3(12):1163–8. doi:10.1038/ni851
50. Bagchi S, He Y, Zhang H, Cao L, Van Rhijn I, Moody DB, et al. CD1b-autoreactive T cells contribute to hyperlipidemia-induced skin inflammation in mice. J Clin Invest (2017) 127(6):2339–52. doi:10.1172/JCI92217
51. Bagchi S, Li S, Wang CR. CD1b-autoreactive T cells recognize phospholipid antigens and contribute to antitumor immunity against a CD1b(+) T cell lymphoma. Oncoimmunology (2016) 5(9):e1213932. doi:10.1080/2162402X.2016.1213932
52. Raftery MJ, Hitzler M, Winau F, Giese T, Plachter B, Kaufmann SH, et al. Inhibition of CD1 antigen presentation by human cytomegalovirus. J Virol (2008) 82(9):4308–19. doi:10.1128/JVI.01447-07
53. Takahashi T, Chiba S, Nieda M, Azuma T, Ishihara S, Shibata Y, et al. Cutting edge: analysis of human V alpha 24+CD8+ NK T cells activated by alpha-galactosylceramide-pulsed monocyte-derived dendritic cells. J Immunol (2002) 168(7):3140–4. doi:10.4049/jimmunol.168.7.3140
54. Gumperz JE, Miyake S, Yamamura T, Brenner MB. Functionally distinct subsets of CD1d-restricted natural killer T cells revealed by CD1d tetramer staining. J Exp Med (2002) 195(5):625–36. doi:10.1084/jem.20011786
55. Kim CH, Johnston B, Butcher EC. Trafficking machinery of NKT cells: shared and differential chemokine receptor expression among V alpha 24(+)V beta 11(+) NKT cell subsets with distinct cytokine-producing capacity. Blood (2002) 100(1):11–6. doi:10.1182/blood-2001-12-0196
56. Lee PT, Benlagha K, Teyton L, Bendelac A. Distinct functional lineages of human V(alpha)24 natural killer T cells. J Exp Med (2002) 195(5):637–41. doi:10.1084/jem.20011908
57. Godfrey DI, Kronenberg M. Going both ways: immune regulation via CD1d-dependent NKT cells. J Clin Invest (2004) 114(10):1379–88. doi:10.1172/JCI23594
58. Constantinides MG, Bendelac A. Transcriptional regulation of the NKT cell lineage. Curr Opin Immunol (2013) 25(2):161–7. doi:10.1016/j.coi.2013.01.003
59. Lee YJ, Holzapfel KL, Zhu J, Jameson SC, Hogquist KA. Steady-state production of IL-4 modulates immunity in mouse strains and is determined by lineage diversity of iNKT cells. Nat Immunol (2013) 14(11):1146–54. doi:10.1038/ni.2731
60. Watarai H, Sekine-Kondo E, Shigeura T, Motomura Y, Yasuda T, Satoh R, et al. Development and function of invariant natural killer T cells producing T(h)2- and T(h)17-cytokines. PLoS Biol (2012) 10(2):e1001255. doi:10.1371/journal.pbio.1001255
61. Engel I, Seumois G, Chavez L, Samaniego-Castruita D, White B, Chawla A, et al. Innate-like functions of natural killer T cell subsets result from highly divergent gene programs. Nat Immunol (2016) 17(6):728–39. doi:10.1038/ni.3437
62. King IL, Fortier A, Tighe M, Dibble J, Watts GF, Veerapen N, et al. Invariant natural killer T cells direct B cell responses to cognate lipid antigen in an IL-21-dependent manner. Nat Immunol (2011) 13(1):44–50. doi:10.1038/ni.2172
63. Monteiro M, Agua-Doce A, Almeida CF, Fonseca-Pereira D, Veiga-Fernandes H, Graca L. IL-9 expression by invariant NKT cells is not imprinted during thymic development. J Immunol (2015) 195(7):3463–71. doi:10.4049/jimmunol.1403170
64. Monteiro M, Almeida CF, Caridade M, Ribot JC, Duarte J, Agua-Doce A, et al. Identification of regulatory Foxp3+ invariant NKT cells induced by TGF-beta. J Immunol (2010) 185(4):2157–63. doi:10.4049/jimmunol.1000359
65. Moreira-Teixeira L, Resende M, Devergne O, Herbeuval JP, Hermine O, Schneider E, et al. Rapamycin combined with TGF-beta converts human invariant NKT cells into suppressive Foxp3+ regulatory cells. J Immunol (2012) 188(2):624–31. doi:10.4049/jimmunol.1102281
66. Sag D, Krause P, Hedrick CC, Kronenberg M, Wingender G. IL-10-producing NKT10 cells are a distinct regulatory invariant NKT cell subset. J Clin Invest (2014) 124(9):3725–40. doi:10.1172/JCI72308
67. Hagglof T, Sedimbi SK, Yates JL, Parsa R, Salas BH, Harris RA, et al. Neutrophils license iNKT cells to regulate self-reactive mouse B cell responses. Nat Immunol (2016) 17(12):1407–14. doi:10.1038/ni.3583
68. Wingender G, Krebs P, Beutler B, Kronenberg M. Antigen-specific cytotoxicity by invariant NKT cells in vivo is CD95/CD178-dependent and is correlated with antigenic potency. J Immunol (2010) 185(5):2721–9. doi:10.4049/jimmunol.1001018
69. Kok WL, Denney L, Benam K, Cole S, Clelland C, McMichael AJ, et al. Pivotal advance: invariant NKT cells reduce accumulation of inflammatory monocytes in the lungs and decrease immune-pathology during severe influenza A virus infection. J Leukoc Biol (2012) 91(3):357–68. doi:10.1189/jlb.0411184
70. Chung BK, Tsai K, Allan LL, Zheng DJ, Nie JC, Biggs CM, et al. Innate immune control of EBV-infected B cells by invariant natural killer T cells. Blood (2013) 122(15):2600–8. doi:10.1182/blood-2013-01-480665
71. Young LS, Rickinson AB. Epstein-Barr virus: 40 years on. Nat Rev Cancer (2004) 4(10):757–68. doi:10.1038/nrc1452
72. Chan AC, Leeansyah E, Cochrane A, d’Udekem d’Acoz Y, Mittag D, Harrison LC, et al. Ex-vivo analysis of human natural killer T cells demonstrates heterogeneity between tissues and within established CD4(+) and CD4(-) subsets. Clin Exp Immunol (2013) 172(1):129–37. doi:10.1111/cei.12045
73. Bandyopadhyay K, Marrero I, Kumar V. NKT cell subsets as key participants in liver physiology and pathology. Cell Mol Immunol (2016) 13(3):337–46. doi:10.1038/cmi.2015.115
74. Gao B, Radaeva S, Park O. Liver natural killer and natural killer T cells: immunobiology and emerging roles in liver diseases. J Leukoc Biol (2009) 86(3):513–28. doi:10.1189/JLB.0309135
75. Bjorkstrom NK, Ljunggren HG, Michaelsson J. Emerging insights into natural killer cells in human peripheral tissues. Nat Rev Immunol (2016) 16(5):310–20. doi:10.1038/nri.2016.34
76. Münz C, Chijioke O. Natural killer cells in herpesvirus infections [version 1; referees: 2 approved]. F1000Res (2017) 6(F1000 Faculty Rev):1231. doi:10.12688/f1000research.11197.1
77. Carnaud C, Lee D, Donnars O, Park SH, Beavis A, Koezuka Y, et al. Cutting edge: cross-talk between cells of the innate immune system: NKT cells rapidly activate NK cells. J Immunol (1999) 163(9):4647–50.
78. Eberl G, MacDonald HR. Selective induction of NK cell proliferation and cytotoxicity by activated NKT cells. Eur J Immunol (2000) 30(4):985–92. doi:10.1002/(SICI)1521-4141(200004)30:4<985::AID-IMMU985>3.0.CO;2-E
79. Ishikawa H, Tanaka K, Kutsukake E, Fukui T, Sasaki H, Hata A, et al. IFN-gamma production downstream of NKT cell activation in mice infected with influenza virus enhances the cytolytic activities of both NK cells and viral antigen-specific CD8+ T cells. Virology (2010) 407(2):325–32. doi:10.1016/j.virol.2010.08.030
80. Kakimi K, Guidotti LG, Koezuka Y, Chisari FV. Natural killer T cell activation inhibits hepatitis B virus replication in vivo. J Exp Med (2000) 192(7):921–30. doi:10.1084/jem.192.7.921
81. Metelitsa LS, Naidenko OV, Kant A, Wu HW, Loza MJ, Perussia B, et al. Human NKT cells mediate antitumor cytotoxicity directly by recognizing target cell CD1d with bound ligand or indirectly by producing IL-2 to activate NK cells. J Immunol (2001) 167(6):3114–22. doi:10.4049/jimmunol.167.6.3114
82. Galani IE, Andreakos E. Neutrophils in viral infections: current concepts and caveats. J Leukoc Biol (2015) 98(4):557–64. doi:10.1189/jlb.4VMR1114-555R
83. Raftery MJ, Lalwani P, Krautkrmer E, Peters T, Scharffetter-Kochanek K, Kruger R, et al. beta2 integrin mediates hantavirus-induced release of neutrophil extracellular traps. J Exp Med (2014) 211(7):1485–97. doi:10.1084/jem.20131092
84. Schonrich G, Raftery MJ. Neutrophil extracellular traps go viral. Front Immunol (2016) 7:366. doi:10.3389/fimmu.2016.00366
85. Stacey MA, Marsden M, Pham NT, Clare S, Dolton G, Stack G, et al. Neutrophils recruited by IL-22 in peripheral tissues function as TRAIL-dependent antiviral effectors against MCMV. Cell Host Microbe (2014) 15(4):471–83. doi:10.1016/j.chom.2014.03.003
86. Leite-de-Moraes MC, Lisbonne M, Arnould A, Machavoine F, Herbelin A, Dy M, et al. Ligand-activated natural killer T lymphocytes promptly produce IL-3 and GM-CSF in vivo: relevance to peripheral myeloid recruitment. Eur J Immunol (2002) 32(7):1897–904. doi:10.1002/1521-4141(200207)32:7<1897:AID-IMMU1897>3.0.CO;2-Y
87. Michel ML, Keller AC, Paget C, Fujio M, Trottein F, Savage PB, et al. Identification of an IL-17-producing NK1.1(neg) iNKT cell population involved in airway neutrophilia. J Exp Med (2007) 204(5):995–1001. doi:10.1084/jem.20061551
88. Xu X, Pocock GM, Sharma A, Peery SL, Fites JS, Felley L, et al. Human iNKT cells promote protective inflammation by inducing oscillating purinergic signaling in monocyte-derived DCs. Cell Rep (2016) 16(12):3273–85. doi:10.1016/j.celrep.2016.08.061
89. De Santo C, Arscott R, Booth S, Karydis I, Jones M, Asher R, et al. Invariant NKT cells modulate the suppressive activity of IL-10-secreting neutrophils differentiated with serum amyloid A. Nat Immunol (2010) 11(11):1039–46. doi:10.1038/ni.1942
90. Wingender G, Hiss M, Engel I, Peukert K, Ley K, Haller H, et al. Neutrophilic granulocytes modulate invariant NKT cell function in mice and humans. J Immunol (2012) 188(7):3000–8. doi:10.4049/jimmunol.1101273
91. Van Kaer L, Parekh VV, Wu L. Invariant natural killer T cells: bridging innate and adaptive immunity. Cell Tissue Res (2011) 343(1):43–55. doi:10.1007/s00441-010-1023-3
92. Dellabona P, Abrignani S, Casorati G. iNKT-cell help to B cells: a cooperative job between innate and adaptive immune responses. Eur J Immunol (2014) 44(8):2230–7. doi:10.1002/eji.201344399
93. Vomhof-DeKrey EE, Yates J, Leadbetter EA. Invariant NKT cells provide innate and adaptive help for B cells. Curr Opin Immunol (2014) 28:12–7. doi:10.1016/j.coi.2014.01.007
94. Barral P, Eckl-Dorna J, Harwood NE, De Santo C, Salio M, Illarionov P, et al. B cell receptor-mediated uptake of CD1d-restricted antigen augments antibody responses by recruiting invariant NKT cell help in vivo. Proc Natl Acad Sci U S A (2008) 105(24):8345–50. doi:10.1073/pnas.0802968105
95. Chang PP, Barral P, Fitch J, Pratama A, Ma CS, Kallies A, et al. Identification of Bcl-6-dependent follicular helper NKT cells that provide cognate help for B cell responses. Nat Immunol (2011) 13(1):35–43. doi:10.1038/ni.2166
96. Leadbetter EA, Brigl M, Illarionov P, Cohen N, Luteran MC, Pillai S, et al. NK T cells provide lipid antigen-specific cognate help for B cells. Proc Natl Acad Sci U S A (2008) 105(24):8339–44. doi:10.1073/pnas.0801375105
97. Gaya M, Barral P, Burbage M, Aggarwal S, Montaner B, Warren Navia A, et al. Initiation of antiviral B cell immunity relies on innate signals from spatially positioned NKT cells. Cell (2017) 172(3):517–33.e20. doi:10.1016/j.cell.2017.11.036
98. Raftery MJ, Wolter E, Fillatreau S, Meisel H, Kaufmann SH, Schonrich G. NKT cells determine titer and subtype profile of virus-specific IgG antibodies during herpes simplex virus infection. J Immunol (2014) 192(9):4294–302. doi:10.4049/jimmunol.1300148
99. Hermans IF, Silk JD, Gileadi U, Salio M, Mathew B, Ritter G, et al. NKT cells enhance CD4+ and CD8+ T cell responses to soluble antigen in vivo through direct interaction with dendritic cells. J Immunol (2003) 171(10):5140–7. doi:10.4049/jimmunol.171.10.5140
100. Ito H, Ando K, Ishikawa T, Nakayama T, Taniguchi M, Saito K, et al. Role of Valpha14+ NKT cells in the development of hepatitis B virus-specific CTL: activation of Valpha14+ NKT cells promotes the breakage of CTL tolerance. Int Immunol (2008) 20(7):869–79. doi:10.1093/intimm/dxn046
101. Johnson TR, Hong S, Van Kaer L, Koezuka Y, Graham BS. NK T cells contribute to expansion of CD8(+) T cells and amplification of antiviral immune responses to respiratory syncytial virus. J Virol (2002) 76(9):4294–303. doi:10.1128/JVI.76.9.4294-4303.2002
102. Joyee AG, Qiu H, Fan Y, Wang S, Yang X. Natural killer T cells are critical for dendritic cells to induce immunity in chlamydial pneumonia. Am J Respir Crit Care Med (2008) 178(7):745–56. doi:10.1164/rccm.200804-517OC
103. Taraban VY, Martin S, Attfield KE, Glennie MJ, Elliott T, Elewaut D, et al. Invariant NKT cells promote CD8+ cytotoxic T cell responses by inducing CD70 expression on dendritic cells. J Immunol (2008) 180(7):4615–20. doi:10.4049/jimmunol.180.7.4615
104. Fujii S, Shimizu K, Smith C, Bonifaz L, Steinman RM. Activation of natural killer T cells by alpha-galactosylceramide rapidly induces the full maturation of dendritic cells in vivo and thereby acts as an adjuvant for combined CD4 and CD8 T cell immunity to a coadministered protein. J Exp Med (2003) 198(2):267–79. doi:10.1084/jem.20030324
105. Munz C, Steinman RM, Fujii S. Dendritic cell maturation by innate lymphocytes: coordinated stimulation of innate and adaptive immunity. J Exp Med (2005) 202(2):203–7. doi:10.1084/jem.20050810
106. Nishimura T, Kitamura H, Iwakabe K, Yahata T, Ohta A, Sato M, et al. The interface between innate and acquired immunity: glycolipid antigen presentation by CD1d-expressing dendritic cells to NKT cells induces the differentiation of antigen-specific cytotoxic T lymphocytes. Int Immunol (2000) 12(7):987–94. doi:10.1093/intimm/12.7.987
107. Ho LP, Urban BC, Jones L, Ogg GS, McMichael AJ. CD4(-)CD8alphaalpha subset of CD1d-restricted NKT cells controls T cell expansion. J Immunol (2004) 172(12):7350–8. doi:10.4049/jimmunol.172.12.7350
108. Bobryshev YV, Lord RS. Co-accumulation of dendritic cells and natural killer T cells within rupture-prone regions in human atherosclerotic plaques. J Histochem Cytochem (2005) 53(6):781–5. doi:10.1369/jhc.4B6570.2005
109. Segura E, Touzot M, Bohineust A, Cappuccio A, Chiocchia G, Hosmalin A, et al. Human inflammatory dendritic cells induce Th17 cell differentiation. Immunity (2013) 38(2):336–48. doi:10.1016/j.immuni.2012.10.018
110. Thomas SY, Hou R, Boyson JE, Means TK, Hess C, Olson DP, et al. CD1d-restricted NKT cells express a chemokine receptor profile indicative of Th1-type inflammatory homing cells. J Immunol (2003) 171(5):2571–80. doi:10.4049/jimmunol.171.5.2571
111. Muller DB, Raftery MJ, Kather A, Giese T, Schonrich G. Frontline: induction of apoptosis and modulation of c-FLIPL and p53 in immature dendritic cells infected with herpes simplex virus. Eur J Immunol (2004) 34(4):941–51. doi:10.1002/eji.200324509
112. Pollara G, Speidel K, Samady L, Rajpopat M, McGrath Y, Ledermann J, et al. Herpes simplex virus infection of dendritic cells: balance among activation, inhibition, and immunity. J Infect Dis (2003) 187(2):165–78. doi:10.1086/367675
113. Raftery MJ, Schwab M, Eibert SM, Samstag Y, Walczak H, Schonrich G. Targeting the function of mature dendritic cells by human cytomegalovirus: a multilayered viral defense strategy. Immunity (2001) 15(6):997–1009. doi:10.1016/S1074-7613(01)00239-4
114. Busche A, Jirmo AC, Welten SP, Zischke J, Noack J, Constabel H, et al. Priming of CD8+ T cells against cytomegalovirus-encoded antigens is dominated by cross-presentation. J Immunol (2013) 190(6):2767–77. doi:10.4049/jimmunol.1200966
115. Nopora K, Bernhard CA, Ried C, Castello AA, Murphy KM, Marconi P, et al. MHC class I cross-presentation by dendritic cells counteracts viral immune evasion. Front Immunol (2012) 3:348. doi:10.3389/fimmu.2012.00348
116. Gottschalk C, Mettke E, Kurts C. The role of invariant natural killer T cells in dendritic cell licensing, cross-priming, and memory CD8(+) T cell generation. Front Immunol (2015) 6:379. doi:10.3389/fimmu.2015.00379
117. Semmling V, Lukacs-Kornek V, Thaiss CA, Quast T, Hochheiser K, Panzer U, et al. Alternative cross-priming through CCL17-CCR4-mediated attraction of CTLs toward NKT cell-licensed DCs. Nat Immunol (2010) 11(4):313–20. doi:10.1038/ni.1848
118. Mattarollo SR, Yong M, Gosmann C, Choyce A, Chan D, Leggatt GR, et al. NKT cells inhibit antigen-specific effector CD8 T cell induction to skin viral proteins. J Immunol (2011) 187(4):1601–8. doi:10.4049/jimmunol.1100756
119. De Santo C, Salio M, Masri SH, Lee LY, Dong T, Speak AO, et al. Invariant NKT cells reduce the immunosuppressive activity of influenza A virus-induced myeloid-derived suppressor cells in mice and humans. J Clin Invest (2008) 118(12):4036–48. doi:10.1172/JCI36264
120. Gabrilovich DI, Bronte V, Chen SH, Colombo MP, Ochoa A, Ostrand-Rosenberg S, et al. The terminology issue for myeloid-derived suppressor cells. Cancer Res (2007) 67(1):425; author reply 426. doi:10.1158/0008-5472.CAN-06-3037
121. Goh C, Narayanan S, Hahn YS. Myeloid-derived suppressor cells: the dark knight or the joker in viral infections? Immunol Rev (2013) 255(1):210–21. doi:10.1111/imr.12084
122. Exley MA, Bigley NJ, Cheng O, Tahir SM, Smiley ST, Carter QL, et al. CD1d-reactive T-cell activation leads to amelioration of disease caused by diabetogenic encephalomyocarditis virus. J Leukoc Biol (2001) 69(5):713–8.
123. Kumar V, Delovitch TL. Different subsets of natural killer T cells may vary in their roles in health and disease. Immunology (2014) 142(3):321–36. doi:10.1111/imm.12247
124. Montamat-Sicotte DJ, Millington KA, Willcox CR, Hingley-Wilson S, Hackforth S, Innes J, et al. A mycolic acid-specific CD1-restricted T cell population contributes to acute and memory immune responses in human tuberculosis infection. J Clin Invest (2011) 121(6):2493–503. doi:10.1172/JCI46216
125. Kasprowicz VO, Cheng TY, Ndung’u T, Sunpath H, Moody DB, Kasmar AG. HIV disrupts human T cells that target mycobacterial glycolipids. J Infect Dis (2016) 213(4):628–33. doi:10.1093/infdis/jiv455
126. Li S, Choi HJ, Felio K, Wang CR. Autoreactive CD1b-restricted T cells: a new innate-like T-cell population that contributes to immunity against infection. Blood (2011) 118(14):3870–8. doi:10.1182/blood-2011-03-341941
127. Zeissig S, Blumberg RS. Primary immunodeficiency associated with defects in CD1 and CD1-restricted T cells. Ann N Y Acad Sci (2012) 1250:14–24. doi:10.1111/j.1749-6632.2011.06380.x
128. Nichols KE, Hom J, Gong SY, Ganguly A, Ma CS, Cannons JL, et al. Regulation of NKT cell development by SAP, the protein defective in XLP. Nat Med (2005) 11(3):340–5. doi:10.1038/nm1189
129. Pasic S, Cupic M, Lazarevic I. HHV-8-related hemophagocytic lymphohistiocytosis in a boy with XLP phenotype. J Pediatr Hematol Oncol (2012) 34(6):467–71. doi:10.1097/MPH.0b013e3182375372
130. Pasquier B, Yin L, Fondaneche MC, Relouzat F, Bloch-Queyrat C, Lambert N, et al. Defective NKT cell development in mice and humans lacking the adapter SAP, the X-linked lymphoproliferative syndrome gene product. J Exp Med (2005) 201(5):695–701. doi:10.1084/jem.20042432
131. Rigaud S, Fondaneche MC, Lambert N, Pasquier B, Mateo V, Soulas P, et al. XIAP deficiency in humans causes an X-linked lymphoproliferative syndrome. Nature (2006) 444(7115):110–4. doi:10.1038/nature05257
132. Huck K, Feyen O, Niehues T, Ruschendorf F, Hubner N, Laws HJ, et al. Girls homozygous for an IL-2-inducible T cell kinase mutation that leads to protein deficiency develop fatal EBV-associated lymphoproliferation. J Clin Invest (2009) 119(5):1350–8. doi:10.1172/JCI37901
133. Banovic T, Yanilla M, Simmons R, Robertson I, Schroder WA, Raffelt NC, et al. Disseminated varicella infection caused by varicella vaccine strain in a child with low invariant natural killer T cells and diminished CD1d expression. J Infect Dis (2011) 204(12):1893–901. doi:10.1093/infdis/jir660
134. Levy O, Orange JS, Hibberd P, Steinberg S, LaRussa P, Weinberg A, et al. Disseminated varicella infection due to the vaccine strain of varicella-zoster virus, in a patient with a novel deficiency in natural killer T cells. J Infect Dis (2003) 188(7):948–53. doi:10.1086/378503
135. Jung J, Bohn G, Allroth A, Boztug K, Brandes G, Sandrock I, et al. Identification of a homozygous deletion in the AP3B1 gene causing Hermansky-Pudlak syndrome, type 2. Blood (2006) 108(1):362–9. doi:10.1182/blood-2005-11-4377
136. Sbihi Z, Dossier A, Boutboul D, Galicier L, Parizot C, Emarre A, et al. iNKT and memory B-cell alterations in HHV-8 multicentric Castleman disease. Blood (2017) 129(7):855–65. doi:10.1182/blood-2016-06-719716
137. de Lalla C, Galli G, Aldrighetti L, Romeo R, Mariani M, Monno A, et al. Production of profibrotic cytokines by invariant NKT cells characterizes cirrhosis progression in chronic viral hepatitis. J Immunol (2004) 173(2):1417–25. doi:10.4049/jimmunol.173.2.1417
138. Jiang X, Zhang M, Lai Q, Huang X, Li Y, Sun J, et al. Restored circulating invariant NKT cells are associated with viral control in patients with chronic hepatitis B. PLoS One (2011) 6(12):e28871. doi:10.1371/journal.pone.0028871
139. Inoue M, Kanto T, Miyatake H, Itose I, Miyazaki M, Yakushijin T, et al. Enhanced ability of peripheral invariant natural killer T cells to produce IL-13 in chronic hepatitis C virus infection. J Hepatol (2006) 45(2):190–6. doi:10.1016/j.jhep.2006.01.034
140. Durante-Mangoni E, Wang R, Shaulov A, He Q, Nasser I, Afdhal N, et al. Hepatic CD1d expression in hepatitis C virus infection and recognition by resident proinflammatory CD1d-reactive T cells. J Immunol (2004) 173(3):2159–66. doi:10.4049/jimmunol.173.3.2159
141. Exley MA, He Q, Cheng O, Wang RJ, Cheney CP, Balk SP, et al. Cutting edge: compartmentalization of Th1-like noninvariant CD1d-reactive T cells in hepatitis C virus-infected liver. J Immunol (2002) 168(4):1519–23. doi:10.4049/jimmunol.168.4.1519
142. Gao B, Radaeva S. Natural killer and natural killer T cells in liver fibrosis. Biochim Biophys Acta (2013) 1832(7):1061–9. doi:10.1016/j.bbadis.2012.09.008
143. Kumar V. NKT-cell subsets: promoters and protectors in inflammatory liver disease. J Hepatol (2013) 59(3):618–20. doi:10.1016/j.jhep.2013.02.032
144. Yanagisawa K, Yue S, van der Vliet HJ, Wang R, Alatrakchi N, Golden-Mason L, et al. Ex vivo analysis of resident hepatic pro-inflammatory CD1d-reactive T cells and hepatocyte surface CD1d expression in hepatitis C. J Viral Hepat (2013) 20(8):556–65. doi:10.1111/jvh.12081
145. Crowe NY, Godfrey DI, Baxter AG. Natural killer T cells are targets for human immunodeficiency virus infection. Immunology (2003) 108(1):1–2. doi:10.1046/j.1365-2567.2003.01580.x
146. Fernandez CS, Kelleher AD, Finlayson R, Godfrey DI, Kent SJ. NKT cell depletion in humans during early HIV infection. Immunol Cell Biol (2014) 92(7):578–90. doi:10.1038/icb.2014.25
147. Fleuridor R, Wilson B, Hou R, Landay A, Kessler H, Al-Harthi L. CD1d-restricted natural killer T cells are potent targets for human immunodeficiency virus infection. Immunology (2003) 108(1):3–9. doi:10.1046/j.1365-2567.2003.01560.x
148. Motsinger A, Haas DW, Stanic AK, Van Kaer L, Joyce S, Unutmaz D. CD1d-restricted human natural killer T cells are highly susceptible to human immunodeficiency virus 1 infection. J Exp Med (2002) 195(7):869–79. doi:10.1084/jem.20011712
149. Mureithi MW, Cohen K, Moodley R, Poole D, Mncube Z, Kasmar A, et al. Impairment of CD1d-restricted natural killer T cells in chronic HIV type 1 clade C infection. AIDS Res Hum Retroviruses (2011) 27(5):501–9. doi:10.1089/aid.2010.0237
150. Sandberg JK, Fast NM, Palacios EH, Fennelly G, Dobroszycki J, Palumbo P, et al. Selective loss of innate CD4(+) V alpha 24 natural killer T cells in human immunodeficiency virus infection. J Virol (2002) 76(15):7528–34. doi:10.1128/JVI.76.15.7528-7534.2002
151. van der Vliet HJ, von Blomberg BM, Hazenberg MD, Nishi N, Otto SA, van Benthem BH, et al. Selective decrease in circulating V alpha 24+V beta 11+ NKT cells during HIV type 1 infection. J Immunol (2002) 168(3):1490–5. doi:10.4049/jimmunol.168.3.1490
152. Vasan S, Poles MA, Horowitz A, Siladji EE, Markowitz M, Tsuji M. Function of NKT cells, potential anti-HIV effector cells, are improved by beginning HAART during acute HIV-1 infection. Int Immunol (2007) 19(8):943–51. doi:10.1093/intimm/dxm055
153. Moll M, Kuylenstierna C, Gonzalez VD, Andersson SK, Bosnjak L, Sonnerborg A, et al. Severe functional impairment and elevated PD-1 expression in CD1d-restricted NKT cells retained during chronic HIV-1 infection. Eur J Immunol (2009) 39(3):902–11. doi:10.1002/eji.200838780
154. Moll M, Snyder-Cappione J, Spotts G, Hecht FM, Sandberg JK, Nixon DF. Expansion of CD1d-restricted NKT cells in patients with primary HIV-1 infection treated with interleukin-2. Blood (2006) 107(8):3081–3. doi:10.1182/blood-2005-09-3636
155. Singh D, Ghate M, Godbole S, Kulkarni S, Thakar M. CD1d-restricted natural killer T cells are preserved in Indian long-term nonprogressors. J Acquir Immune Defic Syndr (2017) 75(4):e104–12. doi:10.1097/QAI.0000000000001322
156. Snyder-Cappione JE, Loo CP, Carvalho KI, Kuylenstierna C, Deeks SG, Hecht FM, et al. Lower cytokine secretion ex vivo by natural killer T cells in HIV-infected individuals is associated with higher CD161 expression. AIDS (2009) 23(15):1965–70. doi:10.1097/QAD.0b013e32832b5134
157. van der Vliet HJ, van Vonderen MG, Molling JW, Bontkes HJ, Reijm M, Reiss P, et al. Cutting edge: rapid recovery of NKT cells upon institution of highly active antiretroviral therapy for HIV-1 infection. J Immunol (2006) 177(9):5775–8. doi:10.4049/jimmunol.177.9.5775
158. Bachle SM, Malone DF, Buggert M, Karlsson AC, Isberg PE, Biague AJ, et al. Elevated levels of invariant natural killer T-cell and natural killer cell activation correlate with disease progression in HIV-1 and HIV-2 infections. AIDS (2016) 30(11):1713–22. doi:10.1097/QAD.0000000000001147
159. Gougeon ML, Piacentini M. New insights on the role of apoptosis and autophagy in HIV pathogenesis. Apoptosis (2009) 14(4):501–8. doi:10.1007/s10495-009-0314-1
160. Balk SP, Bleicher PA, Terhorst C. Isolation and expression of cDNA encoding the murine homologues of CD1. J Immunol (1991) 146(2):768–74.
161. Bradbury A, Belt KT, Neri TM, Milstein C, Calabi F. Mouse CD1 is distinct from and co-exists with TL in the same thymus. EMBO J (1988) 7(10):3081–6.
162. Bradbury A, Calabi F, Milstein C. Expression of CD1 in the mouse thymus. Eur J Immunol (1990) 20(8):1831–6. doi:10.1002/eji.1830200830
163. Lockridge JL, Chen X, Zhou Y, Rajesh D, Roenneburg DA, Hegde S, et al. Analysis of the CD1 antigen presenting system in humanized SCID mice. PLoS One (2011) 6(6):e21701. doi:10.1371/journal.pone.0021701
164. Schonrich G, Raftery MJ. Exploring the immunopathogenesis of viral hemorrhagic fever in mice with a humanized immune system. Front Immunol (2017) 8:1202. doi:10.3389/fimmu.2017.01202
165. Felio K, Nguyen H, Dascher CC, Choi HJ, Li S, Zimmer MI, et al. CD1-restricted adaptive immune responses to mycobacteria in human group 1 CD1 transgenic mice. J Exp Med (2009) 206(11):2497–509. doi:10.1084/jem.20090898
166. Zhao J, Siddiqui S, Shang S, Bian Y, Bagchi S, He Y, et al. Mycolic acid-specific T cells protect against Mycobacterium tuberculosis infection in a humanized transgenic mouse model. Elife (2015) 4:e08525. doi:10.7554/eLife.08525
167. Grubor-Bauk B, Arthur JL, Mayrhofer G. Importance of NKT cells in resistance to herpes simplex virus, fate of virus-infected neurons, and level of latency in mice. J Virol (2008) 82(22):11073–83. doi:10.1128/JVI.00205-08
168. Grubor-Bauk B, Simmons A, Mayrhofer G, Speck PG. Impaired clearance of herpes simplex virus type 1 from mice lacking CD1d or NKT cells expressing the semivariant V alpha 14-J alpha 281 TCR. J Immunol (2003) 170(3):1430–4. doi:10.4049/jimmunol.170.3.1430
169. Ashkar AA, Rosenthal KL. Interleukin-15 and natural killer and NKT cells play a critical role in innate protection against genital herpes simplex virus type 2 infection. J Virol (2003) 77(18):10168–71. doi:10.1128/JVI.77.18.10168-10171.2003
170. Iversen MB, Jensen SK, Hansen AL, Winther H, Issazadeh-Navikas S, Reinert LS, et al. NKT cell activation by local alpha-galactosylceramide administration decreases susceptibility to HSV-2 infection. Immunobiology (2015) 220(6):762–8. doi:10.1016/j.imbio.2014.12.019
171. Cornish AL, Keating R, Kyparissoudis K, Smyth MJ, Carbone FR, Godfrey DI. NKT cells are not critical for HSV-1 disease resolution. Immunol Cell Biol (2006) 84(1):13–9. doi:10.1111/j.1440-1711.2005.01396.x
172. van Dommelen SL, Tabarias HA, Smyth MJ, Degli-Esposti MA. Activation of natural killer (NK) T cells during murine cytomegalovirus infection enhances the antiviral response mediated by NK cells. J Virol (2003) 77(3):1877–84. doi:10.1128/JVI.77.3.1877-1884.2003
173. Broxmeyer HE, Dent A, Cooper S, Hangoc G, Wang ZY, Du W, et al. A role for natural killer T cells and CD1d molecules in counteracting suppression of hematopoiesis in mice induced by infection with murine cytomegalovirus. Exp Hematol (2007) 35(4 Suppl 1):87–93. doi:10.1016/j.exphem.2007.01.015
174. Arase H, Mocarski ES, Campbell AE, Hill AB, Lanier LL. Direct recognition of cytomegalovirus by activating and inhibitory NK cell receptors. Science (2002) 296(5571):1323–6. doi:10.1126/science.1070884
175. Brown MG, Dokun AO, Heusel JW, Smith HR, Beckman DL, Blattenberger EA, et al. Vital involvement of a natural killer cell activation receptor in resistance to viral infection. Science (2001) 292(5518):934–7. doi:10.1126/science.1060042
176. Tyznik AJ, Verma S, Wang Q, Kronenberg M, Benedict CA. Distinct requirements for activation of NKT and NK cells during viral infection. J Immunol (2014) 192(8):3676–85. doi:10.4049/jimmunol.1300837
177. Kakimi K, Lane TE, Chisari FV, Guidotti LG. Cutting edge: inhibition of hepatitis B virus replication by activated NK T cells does not require inflammatory cell recruitment to the liver. J Immunol (2001) 167(12):6701–5. doi:10.4049/jimmunol.167.12.6701
178. Zeissig S, Peuker K, Iyer S, Gensollen T, Dougan SK, Olszak T, et al. CD1d-restricted pathways in hepatocytes control local natural killer T cell homeostasis and hepatic inflammation. Proc Natl Acad Sci U S A (2017) 114(39):10449–54. doi:10.1073/pnas.1701428114
179. Godfrey DI, Uldrich AP, Baxter AG. NKT cells – an early warning system for HBV infection. Nat Med (2012) 18(7):1014–6. doi:10.1038/nm.2853
180. Zeissig S, Murata K, Sweet L, Publicover J, Hu Z, Kaser A, et al. Hepatitis B virus-induced lipid alterations contribute to natural killer T cell-dependent protective immunity. Nat Med (2012) 18(7):1060–8. doi:10.1038/nm.2811
181. Baron JL, Gardiner L, Nishimura S, Shinkai K, Locksley R, Ganem D. Activation of a nonclassical NKT cell subset in a transgenic mouse model of hepatitis B virus infection. Immunity (2002) 16(4):583–94. doi:10.1016/S1074-7613(02)00305-9
182. Vilarinho S, Ogasawara K, Nishimura S, Lanier LL, Baron JL. Blockade of NKG2D on NKT cells prevents hepatitis and the acute immune response to hepatitis B virus. Proc Natl Acad Sci U S A (2007) 104(46):18187–92. doi:10.1073/pnas.0708968104
183. Sundell IB, Halder R, Zhang M, Maricic I, Koka PS, Kumar V. Sulfatide administration leads to inhibition of HIV-1 replication and enhanced hematopoeisis. J Stem Cells (2010) 5(1):33–42.
184. Fernandez CS, Chan AC, Kyparissoudis K, De Rose R, Godfrey DI, Kent SJ. Peripheral NKT cells in simian immunodeficiency virus-infected macaques. J Virol (2009) 83(4):1617–24. doi:10.1128/JVI.02138-08
185. Rout N, Greene J, Yue S, O’Connor D, Johnson RP, Else JG, et al. Loss of effector and anti-inflammatory natural killer T lymphocyte function in pathogenic simian immunodeficiency virus infection. PLoS Pathog (2012) 8(9):e1002928. doi:10.1371/journal.ppat.1002928
186. De Libero G, Moran AP, Gober HJ, Rossy E, Shamshiev A, Chelnokova O, et al. Bacterial infections promote T cell recognition of self-glycolipids. Immunity (2005) 22(6):763–72. doi:10.1016/j.immuni.2005.04.013
187. De Libero G, Mori L. Recognition of lipid antigens by T cells. Nat Rev Immunol (2005) 5(6):485–96. doi:10.1038/nri1631
188. de Jong A. Activation of human T cells by CD1 and self-lipids. Immunol Rev (2015) 267(1):16–29. doi:10.1111/imr.12322
189. Gapin L, Godfrey DI, Rossjohn J. Natural killer T cell obsession with self-antigens. Curr Opin Immunol (2013) 25(2):168–73. doi:10.1016/j.coi.2013.01.002
190. McEwen-Smith RM, Salio M, Cerundolo V. CD1d-dependent endogenous and exogenous lipid antigen presentation. Curr Opin Immunol (2015) 34:116–25. doi:10.1016/j.coi.2015.03.004
191. Brennan PJ, Tatituri RV, Brigl M, Kim EY, Tuli A, Sanderson JP, et al. Invariant natural killer T cells recognize lipid self antigen induced by microbial danger signals. Nat Immunol (2011) 12(12):1202–11. doi:10.1038/ni.2143
192. Brigl M, Bry L, Kent SC, Gumperz JE, Brenner MB. Mechanism of CD1d-restricted natural killer T cell activation during microbial infection. Nat Immunol (2003) 4(12):1230–7. doi:10.1038/ni1002
193. Darmoise A, Teneberg S, Bouzonville L, Brady RO, Beck M, Kaufmann SH, et al. Lysosomal alpha-galactosidase controls the generation of self lipid antigens for natural killer T cells. Immunity (2010) 33(2):216–28. doi:10.1016/j.immuni.2010.08.003
194. Mattner J, Debord KL, Ismail N, Goff RD, Cantu C III, Zhou D, et al. Exogenous and endogenous glycolipid antigens activate NKT cells during microbial infections. Nature (2005) 434(7032):525–9. doi:10.1038/nature03408
195. Paget C, Mallevaey T, Speak AO, Torres D, Fontaine J, Sheehan KC, et al. Activation of invariant NKT cells by toll-like receptor 9-stimulated dendritic cells requires type I interferon and charged glycosphingolipids. Immunity (2007) 27(4):597–609. doi:10.1016/j.immuni.2007.08.017
196. Paquin-Proulx D, Gibbs A, Bachle SM, Checa A, Introini A, Leeansyah E, et al. Innate invariant NKT cell recognition of HIV-1-infected dendritic cells is an early detection mechanism targeted by viral immune evasion. J Immunol (2016) 197(5):1843–51. doi:10.4049/jimmunol.1600556
197. Salio M, Speak AO, Shepherd D, Polzella P, Illarionov PA, Veerapen N, et al. Modulation of human natural killer T cell ligands on TLR-mediated antigen-presenting cell activation. Proc Natl Acad Sci U S A (2007) 104(51):20490–5. doi:10.1073/pnas.0710145104
198. Raftery MJ, Winau F, Giese T, Kaufmann SH, Schaible UE, Schonrich G. Viral danger signals control CD1d de novo synthesis and NKT cell activation. Eur J Immunol (2008) 38(3):668–79. doi:10.1002/eji.200737233
199. Biddlestone J, Bandarra D, Rocha S. The role of hypoxia in inflammatory disease (review). Int J Mol Med (2015) 35(4):859–69. doi:10.3892/ijmm.2015.2079
200. Taylor CT, Colgan SP. Regulation of immunity and inflammation by hypoxia in immunological niches. Nat Rev Immunol (2017) 17(12):774–85. doi:10.1038/nri.2017.103
201. Fox LM, Cox DG, Lockridge JL, Wang X, Chen X, Scharf L, et al. Recognition of lyso-phospholipids by human natural killer T lymphocytes. PLoS Biol (2009) 7(10):e1000228. doi:10.1371/journal.pbio.1000228
202. Webb TJ, Carey GB, East JE, Sun W, Bollino DR, Kimball AS, et al. Alterations in cellular metabolism modulate CD1d-mediated NKT-cell responses. Pathog Dis (2016) 74(6):ftw055. doi:10.1093/femspd/ftw055
203. Altan-Bonnet N. Lipid tales of viral replication and transmission. Trends Cell Biol (2017) 27(3):201–13. doi:10.1016/j.tcb.2016.09.011
204. Chukkapalli V, Heaton NS, Randall G. Lipids at the interface of virus-host interactions. Curr Opin Microbiol (2012) 15(4):512–8. doi:10.1016/j.mib.2012.05.013
205. Fernandez de Castro I, Tenorio R, Risco C. Virus assembly factories in a lipid world. Curr Opin Virol (2016) 18:20–6. doi:10.1016/j.coviro.2016.02.009
206. Heaton NS, Randall G. Multifaceted roles for lipids in viral infection. Trends Microbiol (2011) 19(7):368–75. doi:10.1016/j.tim.2011.03.007
207. Mazzon M, Mercer J. Lipid interactions during virus entry and infection. Cell Microbiol (2014) 16(10):1493–502. doi:10.1111/cmi.12340
208. Strating JR, van Kuppeveld FJ. Viral rewiring of cellular lipid metabolism to create membranous replication compartments. Curr Opin Cell Biol (2017) 47:24–33. doi:10.1016/j.ceb.2017.02.005
209. Raftery MJ, Winau F, Kaufmann SH, Schaible UE, Schonrich G. CD1 antigen presentation by human dendritic cells as a target for herpes simplex virus immune evasion. J Immunol (2006) 177(9):6207–14. doi:10.4049/jimmunol.177.9.6207
210. Gourapura RJ, Khan MA, Gallo RM, Shaji D, Liu J, Brutkiewicz RR. Forming a complex with MHC class I molecules interferes with mouse CD1d functional expression. PLoS One (2013) 8(8):e72867. doi:10.1371/journal.pone.0072867
211. Lin Y, Roberts TJ, Spence PM, Brutkiewicz RR. Reduction in CD1d expression on dendritic cells and macrophages by an acute virus infection. J Leukoc Biol (2005) 77(2):151–8. doi:10.1189/jlb.0704399
212. Ljunggren HG, Karre K. In search of the ‘missing self’: MHC molecules and NK cell recognition. Immunol Today (1990) 11(7):237–44. doi:10.1016/0167-5699(90)90097-S
213. Keller CW, Loi M, Ewert S, Quast I, Theiler R, Gannage M, et al. The autophagy machinery restrains iNKT cell activation through CD1D1 internalization. Autophagy (2017) 13(6):1025–36. doi:10.1080/15548627.2017.1297907
214. Munz C. Beclin-1 targeting for viral immune escape. Viruses (2011) 3(7):1166–78. doi:10.3390/v3071166
215. Brigl M, Tatituri RV, Watts GF, Bhowruth V, Leadbetter EA, Barton N, et al. Innate and cytokine-driven signals, rather than microbial antigens, dominate in natural killer T cell activation during microbial infection. J Exp Med (2011) 208(6):1163–77. doi:10.1084/jem.20102555
216. Leite-De-Moraes MC, Hameg A, Arnould A, Machavoine F, Koezuka Y, Schneider E, et al. A distinct IL-18-induced pathway to fully activate NK T lymphocytes independently from TCR engagement. J Immunol (1999) 163(11):5871–6.
217. Wang X, Bishop KA, Hegde S, Rodenkirch LA, Pike JW, Gumperz JE. Human invariant natural killer T cells acquire transient innate responsiveness via histone H4 acetylation induced by weak TCR stimulation. J Exp Med (2012) 209(5):987–1000. doi:10.1084/jem.20111024
218. Holzapfel KL, Tyznik AJ, Kronenberg M, Hogquist KA. Antigen-dependent versus -independent activation of invariant NKT cells during infection. J Immunol (2014) 192(12):5490–8. doi:10.4049/jimmunol.1400722
219. Tyznik AJ, Tupin E, Nagarajan NA, Her MJ, Benedict CA, Kronenberg M. Cutting edge: the mechanism of invariant NKT cell responses to viral danger signals. J Immunol (2008) 181(7):4452–6. doi:10.4049/jimmunol.181.7.4452
220. Wesley JD, Tessmer MS, Chaukos D, Brossay L. NK cell-like behavior of Valpha14i NK T cells during MCMV infection. PLoS Pathog (2008) 4(7):e1000106. doi:10.1371/journal.ppat.1000106
221. Kuylenstierna C, Bjorkstrom NK, Andersson SK, Sahlstrom P, Bosnjak L, Paquin-Proulx D, et al. NKG2D performs two functions in invariant NKT cells: direct TCR-independent activation of NK-like cytolysis and co-stimulation of activation by CD1d. Eur J Immunol (2011) 41(7):1913–23. doi:10.1002/eji.200940278
222. Esteso G, Guerra S, Vales-Gomez M, Reyburn HT. Innate immune recognition of double-stranded RNA triggers increased expression of NKG2D ligands after virus infection. J Biol Chem (2017) 292(50):20472–80. doi:10.1074/jbc.M117.818393
223. Groh V, Rhinehart R, Randolph-Habecker J, Topp MS, Riddell SR, Spies T. Costimulation of CD8alphabeta T cells by NKG2D via engagement by MIC induced on virus-infected cells. Nat Immunol (2001) 2(3):255–60. doi:10.1038/85321
224. Lee HH, Meyer EH, Goya S, Pichavant M, Kim HY, Bu X, et al. Apoptotic cells activate NKT cells through T cell Ig-like mucin-like-1 resulting in airway hyperreactivity. J Immunol (2010) 185(9):5225–35. doi:10.4049/jimmunol.1001116
225. Rennert PD. Novel roles for TIM-1 in immunity and infection. Immunol Lett (2011) 141(1):28–35. doi:10.1016/j.imlet.2011.08.003
226. Danthi P. Viruses and the diversity of cell death. Annu Rev Virol (2016) 3(1):533–53. doi:10.1146/annurev-virology-110615-042435
227. Horst D, Geerdink RJ, Gram AM, Stoppelenburg AJ, Ressing ME. Hiding lipid presentation: viral interference with CD1d-restricted invariant natural killer T (iNKT) cell activation. Viruses (2012) 4(10):2379–99. doi:10.3390/v4102379
228. Kasinrerk W, Baumruker T, Majdic O, Knapp W, Stockinger H. CD1 molecule expression on human monocytes induced by granulocyte-macrophage colony-stimulating factor. J Immunol (1993) 150(2):579–84.
229. Fortsch D, Rollinghoff M, Stenger S. IL-10 converts human dendritic cells into macrophage-like cells with increased antibacterial activity against virulent Mycobacterium tuberculosis. J Immunol (2000) 165(2):978–87. doi:10.4049/jimmunol.165.2.978
230. Gerlini G, Tun-Kyi A, Dudli C, Burg G, Pimpinelli N, Nestle FO. Metastatic melanoma secreted IL-10 down-regulates CD1 molecules on dendritic cells in metastatic tumor lesions. Am J Pathol (2004) 165(6):1853–63. doi:10.1016/S0002-9440(10)63238-5
231. Thomssen H, Kahan M, Londei M. Differential effects of interleukin-10 on the expression of HLA class II and CD1 molecules induced by granulocyte/macrophage colony-stimulating factor/interleukin-4. Eur J Immunol (1995) 25(9):2465–70. doi:10.1002/eji.1830250909
232. Filippi CM, von Herrath MG. IL-10 and the resolution of infections. J Pathol (2008) 214(2):224–30. doi:10.1002/path.2272
233. Raftery M, Muller A, Schonrich G. Herpesvirus homologues of cellular genes. Virus Genes (2000) 21(1–2):65–75. doi:10.1023/A:1008184330127
234. Schonrich G, Abdelaziz MO, Raftery MJ. Herpesviral capture of immunomodulatory host genes. Virus Genes (2017) 53(6):762–73. doi:10.1007/s11262-017-1460-0
235. Raftery MJ, Wieland D, Gronewald S, Kraus AA, Giese T, Schonrich G. Shaping phenotype, function, and survival of dendritic cells by cytomegalovirus-encoded IL-10. J Immunol (2004) 173(5):3383–91. doi:10.4049/jimmunol.173.5.3383
236. Avdic S, McSharry BP, Steain M, Poole E, Sinclair J, Abendroth A, et al. Human cytomegalovirus-encoded human interleukin-10 (IL-10) homolog amplifies its immunomodulatory potential by upregulating human IL-10 in monocytes. J Virol (2016) 90(8):3819–27. doi:10.1128/JVI.03066-15
237. Chang WL, Baumgarth N, Yu D, Barry PA. Human cytomegalovirus-encoded interleukin-10 homolog inhibits maturation of dendritic cells and alters their functionality. J Virol (2004) 78(16):8720–31. doi:10.1128/JVI.78.16.8720-8731.2004
238. Lambert SL, Martinez OM. Latent membrane protein 1 of EBV activates phosphatidylinositol 3-kinase to induce production of IL-10. J Immunol (2007) 179(12):8225–34. doi:10.4049/jimmunol.179.12.8225
239. Zischke J, Mamareli P, Pokoyski C, Gabaev I, Buyny S, Jacobs R, et al. The human cytomegalovirus glycoprotein pUL11 acts via CD45 to induce T cell IL-10 secretion. PLoS Pathog (2017) 13(6):e1006454. doi:10.1371/journal.ppat.1006454
240. Shinya E, Owaki A, Shimizu M, Takeuchi J, Kawashima T, Hidaka C, et al. Endogenously expressed HIV-1 nef down-regulates antigen-presenting molecules, not only class I MHC but also CD1a, in immature dendritic cells. Virology (2004) 326(1):79–89. doi:10.1016/j.virol.2004.06.004
241. Shinya E, Shimizu M, Owaki A, Paoletti S, Mori L, De Libero G, et al. Hemopoietic cell kinase (Hck) and p21-activated kinase 2 (PAK2) are involved in the down-regulation of CD1a lipid antigen presentation by HIV-1 Nef in dendritic cells. Virology (2016) 487:285–95. doi:10.1016/j.virol.2015.10.023
242. Pereira EA, daSilva LL. HIV-1 Nef: taking control of protein trafficking. Traffic (2016) 17(9):976–96. doi:10.1111/tra.12412
243. Riou R, Bressollette-Bodin C, Boutoille D, Gagne K, Rodallec A, Lefebvre M, et al. Severe symptomatic primary human cytomegalovirus infection despite effective innate and adaptive immune responses. J Virol (2017) 91(5):e02245–16. doi:10.1128/JVI.02245-16
244. Gaglia MM, Covarrubias S, Wong W, Glaunsinger BA. A common strategy for host RNA degradation by divergent viruses. J Virol (2012) 86(17):9527–30. doi:10.1128/JVI.01230-12
245. Rowe M, Glaunsinger B, van Leeuwen D, Zuo J, Sweetman D, Ganem D, et al. Host shutoff during productive Epstein-Barr virus infection is mediated by BGLF5 and may contribute to immune evasion. Proc Natl Acad Sci U S A (2007) 104(9):3366–71. doi:10.1073/pnas.0611128104
246. van Gent M, Gram AM, Boer IG, Geerdink RJ, Lindenbergh MF, Lebbink RJ, et al. Silencing the shutoff protein of Epstein-Barr virus in productively infected B cells points to (innate) targets for immune evasion. J Gen Virol (2015) 96(Pt 4):858–65. doi:10.1099/jgv.0.000021
247. Moody DB, Cotton RN. Four pathways of CD1 antigen presentation to T cells. Curr Opin Immunol (2017) 46:127–33. doi:10.1016/j.coi.2017.07.013
248. Van Kaer L, Wu L, Joyce S. Mechanisms and consequences of antigen presentation by CD1. Trends Immunol (2016) 37(11):738–54. doi:10.1016/j.it.2016.08.011
249. Vartabedian VF, Savage PB, Teyton L. The processing and presentation of lipids and glycolipids to the immune system. Immunol Rev (2016) 272(1):109–19. doi:10.1111/imr.12431
250. de Freitas AC, de Oliveira THA, Barros MR Jr, Venuti A. hrHPV E5 oncoprotein: immune evasion and related immunotherapies. J Exp Clin Cancer Res (2017) 36(1):71. doi:10.1186/s13046-017-0541-1
251. Miura S, Kawana K, Schust DJ, Fujii T, Yokoyama T, Iwasawa Y, et al. CD1d, a sentinel molecule bridging innate and adaptive immunity, is downregulated by the human papillomavirus (HPV) E5 protein: a possible mechanism for immune evasion by HPV. J Virol (2010) 84(22):11614–23. doi:10.1128/JVI.01053-10
252. Gram AM, Oosenbrug T, Lindenbergh MF, Bull C, Comvalius A, Dickson KJ, et al. The Epstein-Barr virus glycoprotein gp150 forms an immune-evasive glycan shield at the surface of infected cells. PLoS Pathog (2016) 12(4):e1005550. doi:10.1371/journal.ppat.1005550
253. Francica JR, Varela-Rohena A, Medvec A, Plesa G, Riley JL, Bates P. Steric shielding of surface epitopes and impaired immune recognition induced by the ebola virus glycoprotein. PLoS Pathog (2010) 6(9):e1001098. doi:10.1371/journal.ppat.1001098
254. Yuan W, Dasgupta A, Cresswell P. Herpes simplex virus evades natural killer T cell recognition by suppressing CD1d recycling. Nat Immunol (2006) 7(8):835–42. doi:10.1038/ni1364
255. Sanchez DJ, Gumperz JE, Ganem D. Regulation of CD1d expression and function by a herpesvirus infection. J Clin Invest (2005) 115(5):1369–78. doi:10.1172/JCI24041
256. Boname JM, Lehner PJ. What has the study of the K3 and K5 viral ubiquitin E3 ligases taught us about ubiquitin-mediated receptor regulation? Viruses (2011) 3(2):118–31. doi:10.3390/v3020118
257. Torreno-Pina JA, Manzo C, Salio M, Aichinger MC, Oddone A, Lakadamyali M, et al. The actin cytoskeleton modulates the activation of iNKT cells by segregating CD1d nanoclusters on antigen-presenting cells. Proc Natl Acad Sci U S A (2016) 113(6):E772–81. doi:10.1073/pnas.1514530113
258. Gallo RM, Khan MA, Shi J, Kapur R, Wei L, Bailey JC, et al. Regulation of the actin cytoskeleton by Rho kinase controls antigen presentation by CD1d. J Immunol (2012) 189(4):1689–98. doi:10.4049/jimmunol.1101484
259. Liu J, Gallo RM, Duffy C, Brutkiewicz RR. A VP22-null HSV-1 is impaired in inhibiting CD1d-mediated antigen presentation. Viral Immunol (2016) 29(7):409–16. doi:10.1089/vim.2015.0145
260. Xiong R, Rao P, Kim S, Li M, Wen X, Yuan W. Herpes simplex virus 1 US3 phosphorylates cellular KIF3A to downregulate CD1d expression. J Virol (2015) 89(13):6646–55. doi:10.1128/JVI.00214-15
261. Rao P, Pham HT, Kulkarni A, Yang Y, Liu X, Knipe DM, et al. Herpes simplex virus 1 glycoprotein B and US3 collaborate to inhibit CD1d antigen presentation and NKT cell function. J Virol (2011) 85(16):8093–104. doi:10.1128/JVI.02689-10
262. Liu J, Glosson NL, Du W, Gervay-Hague J, Brutkiewicz RR. A Thr/Ser dual residue motif in the cytoplasmic tail of human CD1d is important for the down-regulation of antigen presentation following a herpes simplex virus 1 infection. Immunology (2013) 140(2):191–201. doi:10.1111/imm.12127
263. Liu J, Shaji D, Cho S, Du W, Gervay-Hague J, Brutkiewicz RR. A threonine-based targeting signal in the human CD1d cytoplasmic tail controls its functional expression. J Immunol (2010) 184(9):4973–81. doi:10.4049/jimmunol.0901448
264. Zhu Y, Zhang W, Veerapen N, Besra G, Cresswell P. Calreticulin controls the rate of assembly of CD1d molecules in the endoplasmic reticulum. J Biol Chem (2010) 285(49):38283–92. doi:10.1074/jbc.M110.170530
265. Sugden SM, Bego MG, Pham TN, Cohen EA. Remodeling of the host cell plasma membrane by HIV-1 Nef and Vpu: a strategy to ensure viral fitness and persistence. Viruses (2016) 8(3):67. doi:10.3390/v8030067
266. Bachle SM, Sauter D, Sibitz S, Sandberg JK, Kirchhoff F, Moll M. Involvement of a C-terminal motif in the interference of primate lentiviral Vpu proteins with CD1d-mediated antigen presentation. Sci Rep (2015) 5:9675. doi:10.1038/srep09675
267. Moll M, Andersson SK, Smed-Sorensen A, Sandberg JK. Inhibition of lipid antigen presentation in dendritic cells by HIV-1 Vpu interference with CD1d recycling from endosomal compartments. Blood (2010) 116(11):1876–84. doi:10.1182/blood-2009-09-243667
268. Chen N, McCarthy C, Drakesmith H, Li D, Cerundolo V, McMichael AJ, et al. HIV-1 down-regulates the expression of CD1d via Nef. Eur J Immunol (2006) 36(2):278–86. doi:10.1002/eji.200535487
269. Cho S, Knox KS, Kohli LM, He JJ, Exley MA, Wilson SB, et al. Impaired cell surface expression of human CD1d by the formation of an HIV-1 Nef/CD1d complex. Virology (2005) 337(2):242–52. doi:10.1016/j.virol.2005.04.020
270. Halenius A, Gerke C, Hengel H. Classical and non-classical MHC I molecule manipulation by human cytomegalovirus: so many targets-but how many arrows in the quiver? Cell Mol Immunol (2015) 12(2):139–53. doi:10.1038/cmi.2014.105
271. Wiertz EJ, Jones TR, Sun L, Bogyo M, Geuze HJ, Ploegh HL. The human cytomegalovirus US11 gene product dislocates MHC class I heavy chains from the endoplasmic reticulum to the cytosol. Cell (1996) 84(5):769–79. doi:10.1016/S0092-8674(00)81054-5
272. Wiertz EJ, Tortorella D, Bogyo M, Yu J, Mothes W, Jones TR, et al. Sec61-mediated transfer of a membrane protein from the endoplasmic reticulum to the proteasome for destruction. Nature (1996) 384(6608):432–8. doi:10.1038/384432a0
273. Cho S, Jun Y. Human CD1d molecules are resistant to human cytomegalovirus US2- and US11-mediated degradation. Biochem Biophys Res Commun (2011) 413(4):616–22. doi:10.1016/j.bbrc.2011.09.013
274. Han J, Rho SB, Lee JY, Bae J, Park SH, Lee SJ, et al. Human cytomegalovirus (HCMV) US2 protein interacts with human CD1d (hCD1d) and down-regulates invariant NKT (iNKT) cell activity. Mol Cells (2013) 36(5):455–64. doi:10.1007/s10059-013-0221-8
275. Keller CW, Freigang S, Lunemann JD. Reciprocal crosstalk between dendritic cells and natural killer T cells: mechanisms and therapeutic potential. Front Immunol (2017) 8:570. doi:10.3389/fimmu.2017.00570
276. Kain L, Webb B, Anderson BL, Deng S, Holt M, Costanzo A, et al. The identification of the endogenous ligands of natural killer T cells reveals the presence of mammalian alpha-linked glycosylceramides. Immunity (2014) 41(4):543–54. doi:10.1016/j.immuni.2014.08.017
277. Bosnjak L, Jones CA, Abendroth A, Cunningham AL. Dendritic cell biology in herpesvirus infections. Viral Immunol (2005) 18(3):419–33. doi:10.1089/vim.2005.18.419
278. Gutzeit C, Raftery MJ, Peiser M, Tischer KB, Ulrich M, Eberhardt M, et al. Identification of an important immunological difference between virulent varicella-zoster virus and its avirulent vaccine: viral disruption of dendritic cell instruction. J Immunol (2010) 185(1):488–97. doi:10.4049/jimmunol.0902817
279. Kovats S, Turner S, Simmons A, Powe T, Chakravarty E, Alberola-Ila J. West Nile virus-infected human dendritic cells fail to fully activate invariant natural killer T cells. Clin Exp Immunol (2016) 186(2):214–26. doi:10.1111/cei.12850
280. Lanier LL. NKG2D receptor and its ligands in host defense. Cancer Immunol Res (2015) 3(6):575–82. doi:10.1158/2326-6066.CIR-15-0098
281. Schmiedel D, Mandelboim O. Disarming cellular alarm systems-manipulation of stress-induced NKG2D ligands by human herpesviruses. Front Immunol (2017) 8:390. doi:10.3389/fimmu.2017.00390
282. Bosnjak L, Sahlstrom P, Paquin-Proulx D, Leeansyah E, Moll M, Sandberg JK. Contact-dependent interference with invariant NKT cell activation by herpes simplex virus-infected cells. J Immunol (2012) 188(12):6216–24. doi:10.4049/jimmunol.1100218
283. Singh AK, Gaur P, Das SN. Natural killer T cell anergy, co-stimulatory molecules and immunotherapeutic interventions. Hum Immunol (2014) 75(3):250–60. doi:10.1016/j.humimm.2013.12.004
284. Hu Z, Usherwood EJ. Immune escape of gamma-herpesviruses from adaptive immunity. Rev Med Virol (2014) 24(6):365–78. doi:10.1002/rmv.1791
Keywords: human CD1 molecules, antigen presentation, persisting viruses, viral immune evasion, NKT cells
Citation: Schönrich G and Raftery MJ (2018) CD1-Restricted T Cells During Persistent Virus Infections: “Sympathy for the Devil”. Front. Immunol. 9:545. doi: 10.3389/fimmu.2018.00545
Received: 18 January 2018; Accepted: 02 March 2018;
Published: 19 March 2018
Edited by:
Luc Van Kaer, Vanderbilt University, United StatesReviewed by:
Johan K. Sandberg, Karolinska Institute (KI), SwedenRandy Brutkiewicz, Indiana University Bloomington, United States
Copyright: © 2018 Schönrich and Raftery. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Günther Schönrich, Z3VlbnRoZXIuc2Nob2VucmljaEBjaGFyaXRlLmRl