 Pádraic J. Dunne1Christina O. Maher1
Pádraic J. Dunne1Christina O. Maher1 Michael Freeley2Katie Dunne3
Michael Freeley2Katie Dunne3 Andreea Petrasca1Judy Orikiiriza1
Andreea Petrasca1Judy Orikiiriza1 Margaret R. Dunne1Derval Reidy4Siobhan O’Dea4Aisling Loy4Jim Woo4
Margaret R. Dunne1Derval Reidy4Siobhan O’Dea4Aisling Loy4Jim Woo4 Aideen Long2
Aideen Long2 Thomas R. Rogers3Fiona Mulcahy4
Thomas R. Rogers3Fiona Mulcahy4 Derek G. Doherty1*
Derek G. Doherty1*
- 1Discipline of Immunology, School of Medicine, Trinity Translational Medicine Institute, Trinity College Dublin, Dublin, Ireland
- 2Discipline of Clinical Medicine, School of Medicine, Trinity Translational Medicine Institute, Trinity College Dublin, Dublin, Ireland
- 3Discipline of Clinical Microbiology, School of Medicine, Trinity Translational Medicine Institute, Trinity College Dublin, Dublin, Ireland
- 4Genitourinary Infectious Diseases Department, St. James’s Hospital, Dublin, Ireland
Human γδ T cells expressing the Vδ1 T cell receptor (TCR) recognize self and microbial antigens and stress-inducible molecules in a major histocompatibility complex-unrestricted manner and are an important source of innate interleukin (IL)-17. Vδ1 T cells are expanded in the circulation and intestines of patients with human immunodeficiency virus (HIV) infection. In this study, we show that patients with HIV have elevated frequencies, but not absolute numbers, of circulating Vδ1 T cells compared to control subjects. This increase was most striking in the patients with Candida albicans co-infection. Using flow cytometry and confocal microscopy, we identify two populations of Vδ1 T cells, based on low and high expression of the ε chain of the CD3 protein complex responsible for transducing TCR-mediated signals (denoted CD3εlo and CD3εhi Vδ1 T cells). Both Vδ1 T cell populations expressed the CD3 ζ-chain, also used for TCR signaling. Using lines of Vδ1 T cells generated from healthy donors, we show that CD3ε can be transiently downregulated by activation but that its expression is restored over time in culture in the presence of exogenous IL-2. Compared to CD3εhi Vδ1 T cells, CD3εlo Vδ1 T cells more frequently expressed terminally differentiated phenotypes and the negative regulator of T cell activation, programmed death-1 (PD-1), but not lymphocyte-activation gene 3, and upon stimulation in vitro, only the CD3εhi subset of Vδ1 T cells, produced IL-17. Thus, while HIV can infect and kill IL-17-producing CD4+ T cells, Vδ1 T cells are another source of IL-17, but many of them exist in a state of exhaustion, mediated either by the induction of PD-1 or by downregulation of CD3ε expression.
Introduction
T cells expressing the γδ T cell receptor (TCR) represent a minor population of lymphocytes that expands in blood and peripheral tissues upon exposure to bacteria (1, 2), fungi (3), yeast (4, 5), and viruses (6–8). γδ TCRs bind non-peptide antigens in a major histocompatibility complex (MHC) unrestricted manner, leading to phosphorylation of immunoreceptor tyrosine-based activation motifs (ITAM) on the CD3 γ, δ, ε, ζ, and sometimes FcRγ proteins (9, 10). They respond rapidly by killing target cells, releasing cytokines, and providing ligands that mediate the activation and differentiation of other cells of the immune system (11, 12, 13).
Human γδ T cells comprise three predominant cell populations (Vδ1, Vγ9Vδ2, and Vδ3) based upon differences in the δ chain of the TCR (14, 15). Vδ1 TCRs are diverse and can recognize the stress-inducible proteins MICA and MICB, which are expressed by some tumor and virus-infected cells (16), glycolipid antigens presented by CD1c (17) and CD1d (18, 19) and the algal protein phycoerythrin (20). In addition to the TCR, Vδ1 T cells can be activated via ligation of other stimulatory receptors, including NKG2C, NKG2D, NKp30, toll-like receptors, and the β-glucan receptor, dectin 1 (5, 21–24). Upon activation, Vδ1 T cells proliferate, release cytokines, such as interferon-γ (IFN-γ), tumor necrosis factor-α, and interleukin-17 (IL-17), chemokines, such as CCL3, CCL4, and CCL5, and they can kill CD4+ T cells in vitro (4, 21, 23, 25–27).
Vδ1 T cells are found at higher frequencies in the blood, intestinal mucosa, and bronchoalveolar fluid of patients with human immunodeficiency virus (HIV) compared with healthy subjects (28, 29, 30, 31, 32, 33). We have examined the frequencies, phenotypes, and functions of circulating Vδ1 T cells in a cohort of untreated and antiretroviral therapy (ART)-treated patients with HIV and healthy control subjects. We find that percentage frequencies, but not absolute numbers of Vδ1 T cell are higher in the untreated patients compared to ART-treated patients and control subjects. We also have identified two subsets of Vδ1 T cells based on low and high levels of expression of the CD3ε polypeptide, denoted CD3εlo and CD3εhi Vδ1 T cells. Both were expanded in patients with HIV and, in particular, in the patients with Candida albicans co-infection. Phenotypic and functional analysis of these Vδ1 T cell subsets indicated that the CD3εlo cells frequently express terminally differentiated (TD) and exhausted phenotypes and are unable to produce IL-17. These results suggest that HIV may induce a state of Vδ1 T cell inactivation.
Materials and Methods
Study Population
Venous blood was obtained from 36 patients with HIV infection (21 males and 15 females) attending the Genitourinary Infectious Diseases Department at St. James’s Hospital, Dublin. At the time of blood sample collection, 22 patients were receiving ART and 14 were not. The CD4+ T cell count ranged from 55 to 1,857 (median 529) cells/μl of blood in the treated patients and 261–1,115 (median 578) cell/μl in the untreated patients. The viral load ranged from <50 to 72,796 (median < 50) copies/ml in the treated patients and <50–51,000 (median 578) copies/ml in the untreated patients. Three patients were positive for hepatitis B virus and three were positive for hepatitis C. As controls, blood samples were obtained from 23 healthy age- and gender-matched control subjects. Ethical approval for this study was obtained from the Joint Research Ethics Committee of St. James’s Hospital and Tallaght Hospitals, Dublin, and all participants gave written, informed consent. Buffy coat packs from healthy blood donors were kindly provided by the Irish Blood Transfusion Service. Whole blood was used for enumerating T cells, as described below. Peripheral blood mononuclear cells (PBMCs) were prepared by density gradient centrifugation over Lymphoprep (Nycomed Pharma, Oslo, Norway) and used immediately in all procedures.
Antibodies and Flow Cytometry
Fluorochrome-conjugated monoclonal antibodies (mAbs) specific for the human Vδ1 TCR (clone TS-1), CD3ε (clones MEM-1 and HIT-3a), CD3ζ (clone 6B10.2), CD27 (clone 0323), CD45RA (clone HI100), programmed death-1 (PD-1) (clone EH12.1), lymphocyte-activation gene 3 (LAG-3) (clone 11C3C65), and CD31 (clone WM59) were obtained from Thermo Fisher Scientific (Dublin, Ireland), BioLegend (San Diego, CA, USA), and Beckman Coulter (High Wycombe, UK) and used according to the manufacturers’ recommendations. The CD3ε mAb (clone SP4) was kindly provided by Dr. Balbino Alcarón (Severo Ochoa Center for Molecular Biology, Madrid, Spain). Up to 106 PBMC, γδ T cell-enriched PBMC or expanded Vδ1 T cell lines were labeled with mAbs and analyzed using a CyAN ADP (Beckman Coulter) or FACSCanto (Becton Dickinson, Oxford, UK) flow cytometer. Data were analyzed with FlowJo v7.6 (Tree Star, Ashland, OR, USA) software. Single-stained OneComp Beads (Becton Dickinson) were used to set compensation parameters; fluorescence minus one (FMO) and isotype-matched Ab controls were used to set analysis gates. Fixable viability dye (eBioscience) was used to determine cell viability. The gating strategy for enumerating Vδ1 T cells is shown in Figure 1A. Total PBMC were analyzed for the enumeration of γδ T cell subsets. γδ T cell-enriched PBMC, prepared by negative selection using magnetic beads (Miltenyi Biotec, Bergische Gladbach, Germany), were used as a source of Vδ1 T cells for subsequent phenotypic and functional analysis.
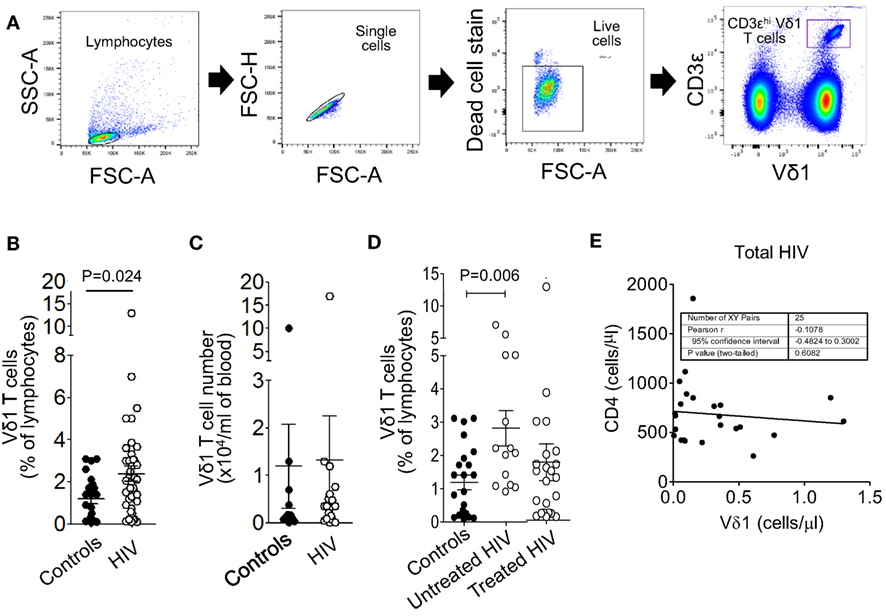
Figure 1. Circulating Vδ1 T cell frequencies but not numbers are higher in patients with untreated human immunodeficiency virus (HIV) infection. Peripheral blood mononuclear cells were prepared from blood samples of 36 patients with HIV infection and 23 healthy donors, stained with monoclonal antibodies specific for CD3 and the Vδ1 T cell receptor and analyzed by flow cytometry. (A) Gating strategy for the enumeration of Vδ1 T cells, showing sequential gates on the lymphocytes, single cells, live cells, and Vδ1 T cells. (B,C) Scatter plots showing circulating Vδ1 T cell frequencies (B) and absolute numbers (C). (D) Scatter plots showing Vδ1 T cell frequencies in control subjects (n = 23) and HIV patients divided into untreated (n = 14) and antiretroviral therapy-treated (n = 22) groups. Groups were compared using the Mann–Whitney U test. (E) Correlation between absolute counts of Vδ1 T cells and total CD4+ T cells in the blood of 25 patients with HIV infection.
Enumeration of Vδ1 T Cells
Absolute numbers of T cells per μl of blood were determined using Trucount tubes (BD Biosciences) according to the manufacturer’s protocol. The percentages of CD3+ cells that expressed Vδ1 TCRs, were determined by flow cytometry, as described above, allowing us to calculate the absolute counts of Vδ1 T cells (per μl of blood).
Vδ1 T Cell Sorting and Expansion
Lines of Vδ1 T cells were generated from healthy blood donors as described previously (5). Briefly, PBMC were prepared from buffy coat packs and monocytes were isolated by positive selection using CD14 Microbeads (Miltenyi Biotec, Gladbach Bergische, Germany). Monocytes were allowed to differentiate into immature dendritic cells (DCs) by culturing them for 6 days in the presence of granulocyte–monocyte colony-stimulating factor and IL-4 as described (34). Immature DC were plated at densities of 100,000 cells/ml and stimulated overnight with medium only, with heat- or ethanol-killed C. albicans (5 × 106 cells/ml) (5). C. albicans strain 10231 was obtained from the American Type Culture Collection and cultured for 24 h on malt extract agar. Fungi were cultured for 24 h, isolated, counted, and then inactivated by heating at 96°C for 60 min. Samples were then centrifuged at 5,000 × g for 10 min, the supernatants discarded, and the pellets washed with phosphate buffered saline (PBS). Inactivation was confirmed by plating an aliquot onto malt extract agar and incubating for 7 days to check for growth.
Total γδ T cells were enriched from PBMCs using human anti-TCR γ/δ Microbeads (Miltenyi Biotec). γδ-enriched cells (200,000 cells/ml) were cultured in the absence or presence of C. albicans- or curdlan-treated DCs at 2:1 ratios in complete serum-free AIM-V medium (AIM-V containing 0.05 mM l-glutamine, 100 U/ml penicillin, 100 U/ml streptomycin, 0.02 M HEPES, 55 µM β-mercaptoethanol, 1× essential amino acids, 1× nonessential amino acids, and 1 mM sodium pyruvate). Co-cultures were challenged with phytohemagglutinin (1 µg/ml; Sigma-Aldrich, Dublin, Ireland) and cultured with rIL-2 (40 U/ml; Miltenyi Biotec), which was added in fresh medium every 2–3 days. Cultures were restimulated every 2 weeks with activated DCs and phytohemagglutinin, which resulted in yields of >10 million Vδ1 T cells by day 28.
Confocal Microscopy
Expanded Vδ1 T cells were sorted into cells with high and low surface expression of CD3ε using a MoFlo XDP Cell Sorter (Beckman Coulter). The cell populations were subsequently incubated on poly l-lysine-coated 8-well Lab-Tek glass chamber slides (Nunc; Thermo Fisher Scientific) for 30 min at 37°C. The cells were fixed with an equal volume of 8% paraformaldehyde for 15 min at 37°C, permeabilized with 0.3% triton X-100 in PBS for 5 min at room temperature and then blocked with 3% bovine serum albumin in PBS for 30 min at room temperature. The samples were incubated with a fluorescein isothiocyanate (FITC)-conjugated mouse anti-human CD3ε antibody (clone SK7, BioLegend, 1/50 dilution in 3% BSA/PBS) and incubated overnight at 4°C. After two washes in PBS, the slides were counter-stained with Hoechst 33258 (Molecular Probes) for 30 min at room temperature to visualize the nuclei. The slides were then imaged under 63× oil immersion with a Zeiss laser scanning confocal 510 microscope (Carl Zeiss, Hertfordshire, UK). The mean fluorescence intensity (MFI) of CD3 staining and Hoechst staining in individual cells was quantified using Zen 2009 imaging software (Carl Zeiss). The MFI of Hoechst served as an internal reference control between the different populations.
Analysis of Intracellular Cytokine Production
Interleukin-17 expression by fresh, unexpanded Vδ1 T cells within γδ T cell-enriched PBMCs was examined by flow cytometry after stimulation of the cells for 6 h with medium alone or with 1 ng/ml phorbol myristate acetate (PMA) and 1 µg/ml ionomycin (PMA/I) in the presence of brefeldin A to prevent cytokine release from the cells (5, 34).
Statistical Analysis
Prism GraphPad software (San Diego, CA, USA) was used for data analysis. Cell frequencies and numbers determined by flow cytometry in subject groups and cytokine levels in treatment groups were compared using the Mann–Whitney U test. P values <0.05 were considered significant. Correlations were defined using Pearson’s correlation coefficient.
Results
Vδ1 T Cell Frequencies but Not Numbers Are Higher in Patients With Untreated HIV Infection
Peripheral blood mononuclear cells were prepared from blood samples of 36 patients with HIV infection and 23 healthy donors, stained with mAbs specific for CD3ε and the Vδ1 TCR and analyzed by flow cytometry (Figure 1A). Figure 1B shows that the frequencies, as percentages of lymphocytes, of Vδ1 T cells were significantly higher in the HIV patient samples. Absolute counts of Vδ1 T cells were not significantly different between patients and controls (Figure 1C), suggesting that the percentage increases in Vδ1 T cells are a result of the depletions of CD4+ T cells by HIV. When the patients were divided into untreated (n = 14) and ART-experienced (n = 22) groups, the frequencies of Vδ1 T cells were found to be higher only in the untreated patients (Figure 1D). Vδ1 T cell numbers did not correlate significantly with total CD4+ T cell counts (Figure 1E), suggesting that the increases in Vδ1 T cells in patients with HIV do not simply compensate for the depletions of CD4+ T cells. These data confirm and extend previous observations of altered Vδ1 T cell frequencies in patients with HIV.
Significant Numbers of Vδ1 T Cells Do Not Appear to Express CD3ε
A surprising observation made, while determining the frequencies of Vδ1 T cells in patients and control subjects, was that significant numbers of Vδ1 T cells do not appear to express CD3ε. CD3ε-negative Vδ1 T cells were detected in PBMC and in γδ T cell-enriched PBMC from both patients and control subjects using three different anti-CD3ε mAbs (clones MEM-1, SP4, and HIT-3a) after gating out dead cells, doublets and using FMO controls (Figure 2A). This allowed us to subdivide Vδ1 T cells into two groups on the basis of low and high expression of the TCR co-receptor, denoted CD3εlo and CD3εhi Vδ1 T cells, respectively. The levels of Vδ1 TCR expression were slightly higher in CD3εhi compared to CD3εlo Vδ1 T cells in both HIV patients and control subjects, although these differences did not reach statistical significance (Figure 2B). Further flow cytometric analysis revealed that both CD3εlo and CD3εhi Vδ1 T cells express the CD3ζ polypeptide (Figure 2C).
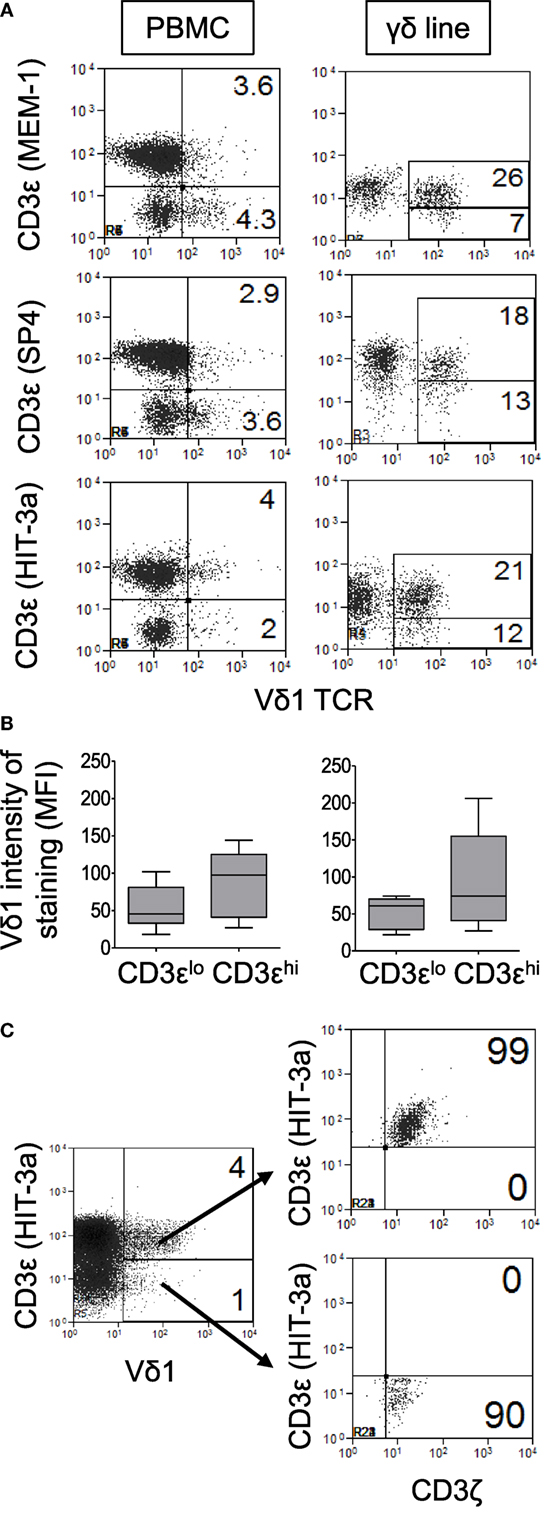
Figure 2. Significant numbers of Vδ1 T cells do not appear to express CD3ε. Peripheral blood mononuclear cell (PBMC) and γδ T cell-enriched PBMC were stained with antibodies specific for Vδ1 and CD3ζ and three different anti-CD3ε monoclonal antibodies (clones MEM-1, SP4, and HIT-3a) in separate tubes and analyzed by flow cytometry. (A) Representative flow cytometry dot plots of PBMC (left panels) and expanded Vδ1 T cells (right panels) from a patient with human immunodeficiency virus (HIV) infection showing the expression of CD3ε by Vδ1 T cells. (B) Box plots showing mean fluorescence intensities of staining for Vδ1 T cells in CD3εlo and CD3εhi Vδ1 T cells from six healthy donors (left) and nine HIV patients (right). (C) Representative flow cytometry dot plot showing CD3ζ expression by gated CD3εlo and CD3εhi Vδ1 T cells. Results are representative of PBMC or γδ T cell-enriched PBMC from four different donors.
CD3εlo Vδ1 T Cells Express Very Low Levels or No Intracellular CD3ε
The low levels of CD3ε expression by some Vδ1 T cells may be due to internalization of the CD3ε chain. To investigate if CD3εlo Vδ1 T cells express intracellular CD3ε, Vδ1 T cells were purified from two healthy donors and sorted by flow cytometry into cells with high and low surface expression of CD3ε (Figure 3A). The cell populations were then bound to slides, fixed, permeabilized, blocked with bovine serum albumin, and stained with a FITC-conjugated mouse anti-human CD3ε antibody and Hoechst 33258. Cells were imaged by confocal microscopy and MFIs were quantified. Figures 3B,C show that CD3εhi Vδ1 T cells express high levels of cell surface CD3ε and low levels of intracellular CD3ε. By contrast, CD3εlo Vδ1 T cells express very low levels of cell surface or intracellular CD3ε, indicating that the CD3εlo Vδ1 T cell phenotype is not the result of internalization CD3ε.
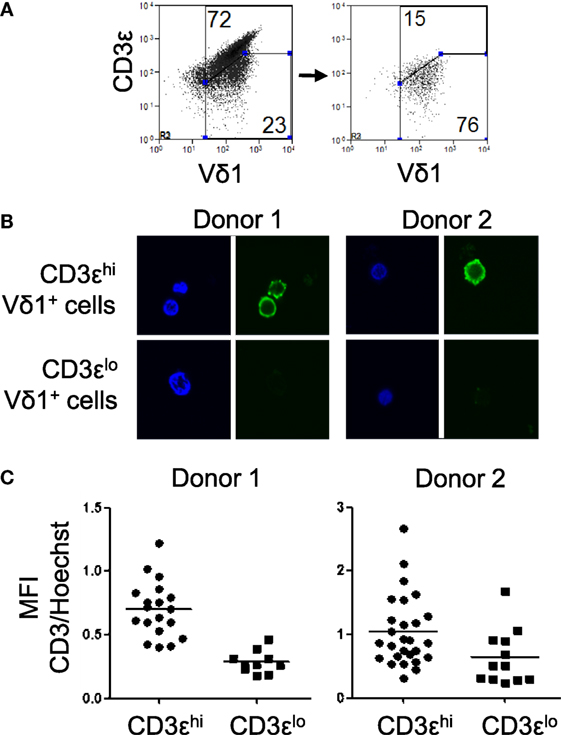
Figure 3. CD3εlo Vδ1 T cells express very low levels of intracellular CD3ε. Vδ1 T cells were purified from two independent blood donors and sorted using a flow cytometric cell sorter into cells with high and low surface expression of CD3ε. The cell populations were incubated on poly l-lysine-coated glass chamber slides, fixed, permeabilized, and blocked with BSA. The samples were then incubated overnight with a FITC-conjugated mouse anti-human CD3ε antibody, washed twice, and counter-stained with Hoechst 33258 to visualize the nuclei. The slides were then imaged under oil immersion with a laser scanning confocal microscope and mean fluorescence intensity (MFI) was quantified. (A) Flow cytometry dot plots showing CD3ε and Vδ1 T cell receptor expression by a line of expanded Vδ1 T cells (left) and sorted CD3εlo Vδ1 T cells (right). (B) Confocal micrographs showing CD3ε (green) and nuclei (blue) in sorted CD3εhi (top) and CD3εlo (bottom) Vδ1 T cells from two donors. (C) Scatter plots showing the ratios of MFI of CD3 to Hoechst staining in individual cells from the sorted populations of CD3εhi and CD3εlo Vδ1 T cells from two donors.
CD3εlo and CD3εhi Vδ1 T Cells Are Both Preserved in Patients With HIV and Especially in Patients With C. albicans Co-Infection
We next investigated if the percentage frequencies of CD3εlo and CD3εhi Vδ1 T cells correlated with the presence of HIV infection in untreated and ART-treated patients. PBMCs were prepared from 14 patients with untreated HIV infection, 22 patients receiving ART and 23 healthy donors, stained with mAbs specific for CD3ε and Vδ1 and analyzed by flow cytometry. Figure 4A shows that both subsets of Vδ1 T cells are expanded in the untreated patients, whereas CD3εhi Vδ1 T cells, only, are expanded in treated patients. There were no significant differences in the frequencies of CD3εlo and CD3εhi Vδ1 T cells in patients with HIV. We previously reported that Vδ1 T cells expand and release IL-17 in response to C. albicans, a common co-infection in patients with HIV (5). Figure 4B shows that the frequencies of both subsets of Vδ1 T cells were significantly higher in patients with Candida co-infection (n = 13) compared to patients with no evidence of fungal infection (n = 19), indicating that fungal infection makes a significant contribution to the increased frequencies of Vδ1 T cells reported in patients with HIV infection (28–33).
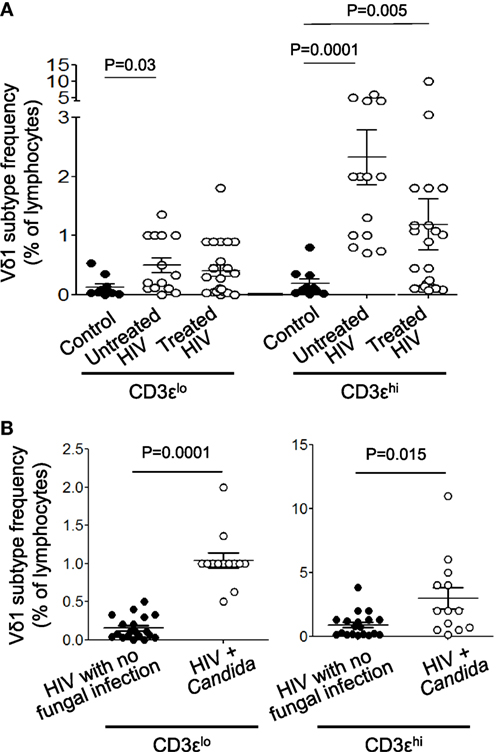
Figure 4. Both CD3εlo and CD3εhi Vδ1 T cells are expanded in untreated patients with human immunodeficiency virus (HIV) infection and especially in patients with Candida co-infection. Peripheral blood mononuclear cells were prepared from blood samples of 14 patients with untreated HIV infection, 22 patients receiving antiretroviral therapy and 23 healthy donors, stained with monoclonal antibodies specific for CD3ε and Vδ1 and analyzed by flow cytometry. (A) Scatter plots showing the frequencies of circulating CD3εlo and CD3εhi Vδ1 T cells in untreated and treated patients with HIV infection and healthy control subjects. (B) Scatter plots showing the frequencies of circulating CD3εlo and CD3εhi Vδ1 T cells in patients with HIV divided according to the absence (n = 19) or presence (n = 13) of Candida albicans co-infection. Groups were compared using the Mann–Whitney U test.
CD3ε Expression by Vδ1 T Cells Can Be Modulated by Activation
We next investigated if CD3ε expression by Vδ1 T cells is stable or if it can be modulated by activation. CD3εhi and CD3εlo Vδ1 T cells were sorted from lines of Vδ1 T cells that were expanded from three donors. Cells were restimulated with PMA/I (Figure 5A) or DC pulsed with heat-killed C. albicans and PHA (Figure 5B) and cultured in the presence of IL-2. The expression of CD3ε by gated Vδ1 T cells was examined at times 0, 1, 7, and 14 days by flow cytometry. CD3ε expression by sorted CD3εhi Vδ1 T cells was transiently downregulated by activation with PMA/I (Figure 5A). Figures 5B,C show that CD3ε expression by CD3εlo Vδ1 T cells can be upregulated over time post reactivation with antigen. Thus, CD3ε can be transiently downregulated in CD3εhi Vδ1 T cells and upregulated in the CD3εlo population following activation, indicating that CD3ε expression is not fixed and that its downregulation is reversible.
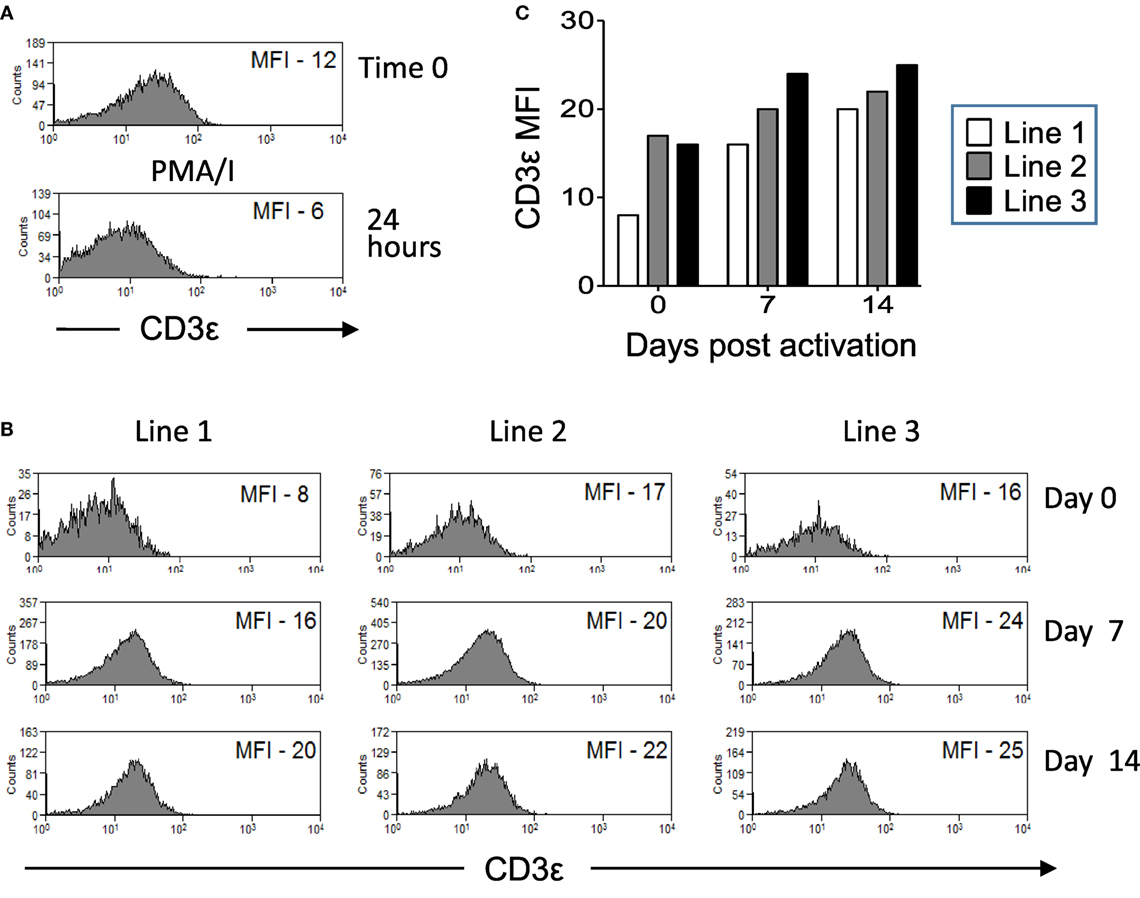
Figure 5. CD3ε expression by Vδ1 T cells is modulated over time post activation. CD3εhi and CD3εlo Vδ1 T cells were sorted from lines of expanded Vδ1 T cells from three donors. Cells were restimulated with phorbol myristate acetate with ionomycin (PMA/I) or heat-killed Candida albicans presented by monocyte-derived dendritic cell and PHA and cultured in the presence of interleukin (IL)-2. The expression of CD3ε by gated Vδ1 T cells was examined at times 0, 1, 7, and 14 days by flow cytometry. (A) Flow cytometry histograms showing downregulation of CD3ε by sorted CD3εhi Vδ1 T cells stimulated for 24 h with PMA/I. (B) Flow cytometry histograms showing the upregulation of CD3ε expression by sorted CD3εlo Vδ1 T cells after stimulation with C. albicans and culture with IL-2. (C) Mean fluorescence intensities (MFIs) of CD3ε expression by Vδ1 T cells from three donors over time after C. albicans stimulation.
CD3εlo Vδ1 T Cells More Frequently Have TD Phenotypes Than CD3εhi Vδ1 T Cells
The differentiation status of CD3εlo and CD3εhi Vδ1 T cells in 19 patients with HIV infection and 18 control subjects was examined by flow cytometric analysis of CD45RA and CD27 co-expression (34, 35). Figure 6A shows that significant proportions of total lymphocytes and gated CD3εhi Vδ1 T cells within γδ T cell-enriched PBMC from patients and controls expressed naïve (CD45RA+CD27+), central memory (CD45RA−CD27+), effector memory (CD45RA−CD27−), and TD (CD45RA+CD27−) phenotypes. By contrast, CD3εlo Vδ1 T cells from control subjects exhibited significantly higher frequencies of TD cells compared to CD3εhi Vδ1 T cells (Figures 6A,B). A similar increase in TD cells among CD3εlo Vδ1 T cells was found in the patients with HIV, with 90–100% of these cells being CD45RA+CD27+ in some patients, but this did not reach statistical significance. Interestingly, the proportions of CD3εhi Vδ1 T cells that expressed TD phenotypes were higher in the HIV patients compared to control subjects. When the HIV-infected patients were divided into untreated (n = 13) and ART-treated (n = 14) subjects, the proportions of CD3εlo Vδ1 T cells expressing TD phenotypes was only marginally higher than those of CD3εhi Vδ1 T cells (Figure 6B). These results show that significant proportions of CD3εlo Vδ1 T cells express TD phenotypes, suggesting that they are exhausted as a result of HIV infection.
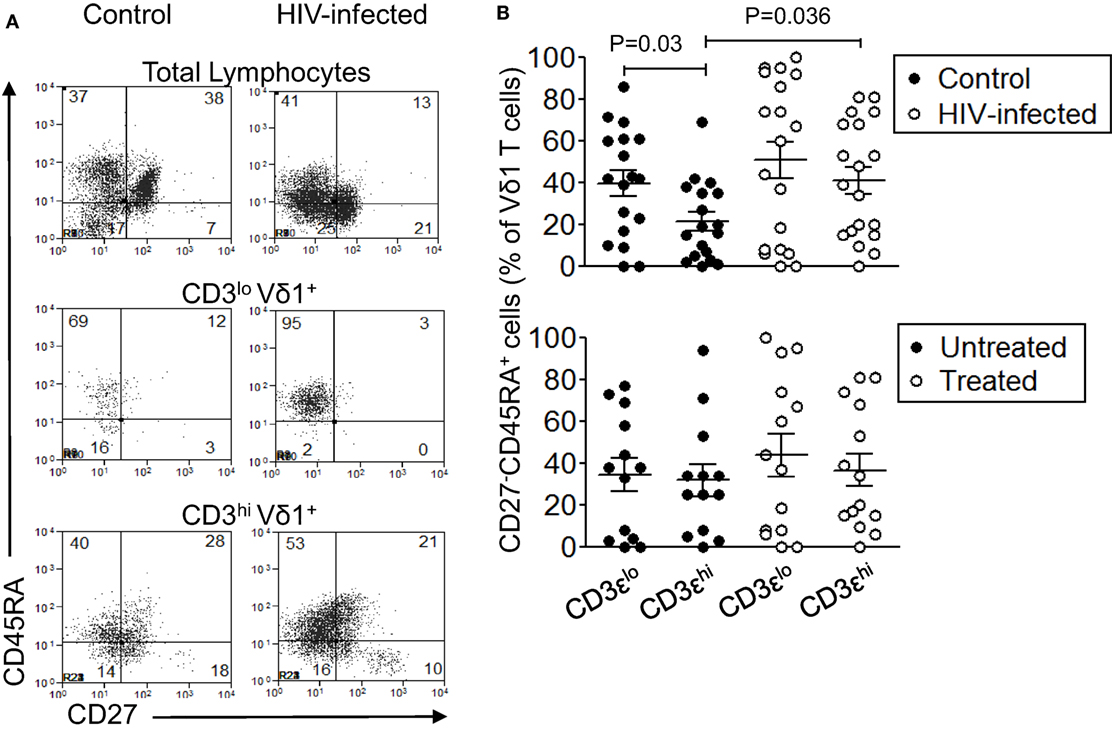
Figure 6. Significant proportions of CD3εlo Vδ1 T cells have terminally differentiated (TD) phenotypes. Peripheral blood mononuclear cell (PBMC) from 19 patients with human immunodeficiency virus (HIV) infection and 18 control subjects were enriched for γδ T cells using magnetic bead separation, stained with monoclonal antibodies specific for CD3ε, Vδ1, CD45RA, and CD27, and analyzed by flow cytometry. (A) Representative flow cytometry dot plots showing CD45RA and CD27 expression by total lymphocytes (upper panels), CD3εlo Vδ1 T cells (center panels), and CD3εhi Vδ1 T cells (bottom panels) in a control subject (left panels) and a patient with HIV infection (right panels). (B) Scatter plots showing the frequencies of CD3εlo and CD3εhi Vδ1 T cells from the patients and controls (upper graph) that expressed TD (CD45RA+CD27−) phenotypes. The lower graph shows the frequencies of CD3εlo and CD3εhi Vδ1 T cells from untreated (n = 13) and antiretroviral therapy-treated (n = 14) patients who expressed TD phenotypes. Groups were compared using the Mann–Whitney U test.
CD3εlo Vδ1 T Cells More Frequently Express PD-1, but Not LAG-3 or CD31, Than CD3εhi Vδ1 T Cells
Human immunodeficiency virus can induce the expression of the inhibitory receptors PD-1 and LAG-3 on HIV-specific T cells leading to their inactivation (36–42). Since Vδ1 T cells with TD phenotypes are preserved in patients with HIV infection, we investigated if CD3εlo and CD3εhi Vδ1 T cells from five untreated patients with HIV infection and eight control subjects express PD-1 or LAG-3. We also investigated if these cells express the naïve T cell marker CD31 (43). Figure 7 shows that PD-1 is expressed at higher levels on CD3εlo Vδ1 T cells compared to CD3εhi Vδ1 T cells from eight healthy donors. A similar trend, although not statistically significant was found in five untreated HIV patients (Figure 7B). PD-1 expression by CD3εlo and CD3εhi Vδ1 T cells was similar in patients and control subjects. By contrast, neither CD3εlo nor CD3εhi Vδ1 T cells from patients or controls expressed LAG-3. CD31 was expressed by variable proportions of CD3εlo and CD3εhi Vδ1 T cells and its expression was not altered in patients with HIV (Figure 7).
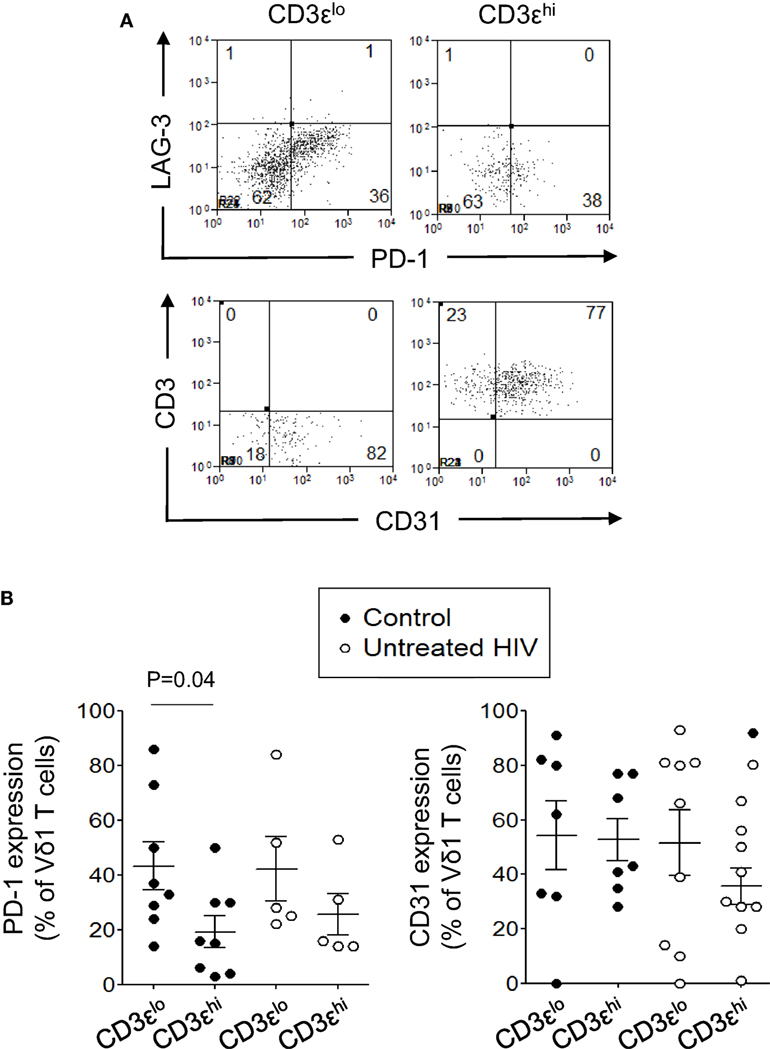
Figure 7. CD3εlo Vδ1 T cells more frequently express programmed death-1 (PD-1), but not lymphocyte-activation gene 3 (LAG-3) or CD31, than CD3εhi Vδ1 T cells. Peripheral blood mononuclear cell prepared from eight healthy donors and five untreated patients with human immunodeficiency virus (HIV) were enriched for γδ T cells using magnetic bead separation, stained with monoclonal antibodies specific for CD3ε, Vδ1, PD-1, LAG-3, and CD31 and analyzed by flow cytometry. (A) Flow cytometry dot plots showing PD-1, LAG-3, and CD31 expression by gated CD3εlo and CD3εhi Vδ1 T cells from a patient with HIV. (B) Scatter plots showing the frequencies of CD3εlo and CD3εhi Vδ1 T cells from the patients with HIV and control subjects that expressed PD-1 and CD31. Groups were compared using the Mann–Whitney U test.
CD3εlo Vδ1 T Cells Exhibit Impaired IL-17 Production
The increased expression of PD-1 and TD phenotypes of CD3εlo Vδ1 T cells suggest that these cells are in a state of exhaustion. We and others have shown that Vδ1 T cells are rapid and potent producers of IL-17 (4, 5). We investigated if CD3εlo and CD3εhi Vδ1 T cells from patients with HIV infection and control subjects differ in their ability to produce IL-17. γδ T cell-enriched PBMC from 13 healthy donors and 11 patients with HIV were stimulated for 6 h with PMA/I or incubated in medium alone and IL-17A expression by gated CD3εlo and CD3εhi Vδ1 T cells was examined by flow cytometry (Figure 8A). Figures 8B,C show that PMA/I treatment induced the production of IL-17 by significant numbers of CD3εhi Vδ1 T cells from both control subjects and HIV patients. However, stimulation of CD3εlo Vδ1 T cells with PMA/I did not lead to IL-17 production, suggesting that these cells are at least partially inactivated (Figure 8).
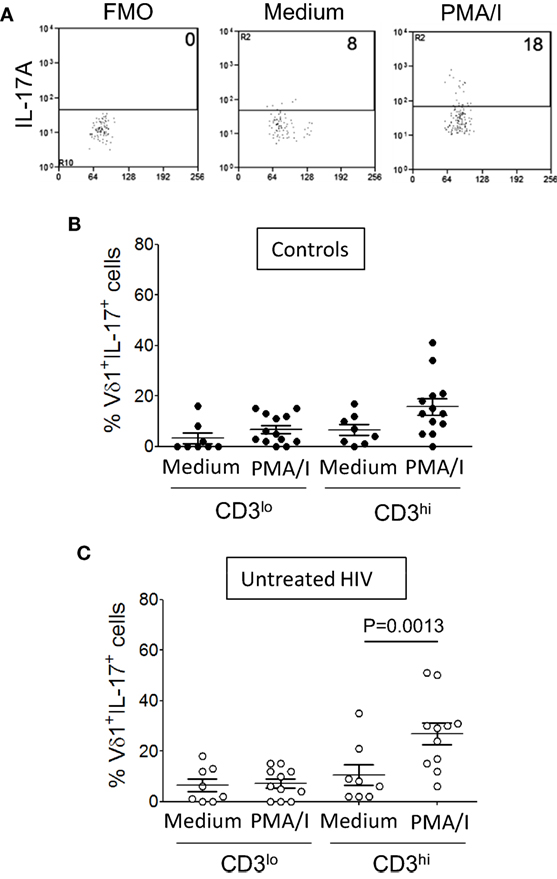
Figure 8. CD3εlo Vδ1 T cells exhibit impaired interleukin (IL)-17 production. Peripheral blood mononuclear cell (PBMC) from 13 healthy donors and 11 patients with untreated human immunodeficiency virus (HIV) infection were enriched for γδ T cells using magnetic bead separation. The cells were stimulated for 6 h with phorbol myristate acetate with ionomycin (PMA/I) or incubated in medium alone in the presence of brefeldin A. (A) Flow cytometry dot plots showing IL-17A expression by gated CD3εhi Vδ1 T cells within unstimulated PBMC (center panel) and PMA/ionomycin-stimulated PBMC (right panel) from a patient with HIV. The left panel shows a fluorescence minus one (FMO) control dot plot. (B,C) Scatter plots show the frequencies of CD3εlo and CD3εhi Vδ1 T cells from healthy donors (B) and HIV patients (C) that produced IL-17. Groups were compared using the Mann–Whitney U test.
Discussion
Numerous studies have shown that Vδ1 T cells are proportionally expanded in patients with HIV (28–33). Vδ1 T cells may contribute to immunity against HIV by killing infected CD4+ T cells (21, 25), releasing antiviral cytokines (4, 25, 27) and chemokines (23). They may also contribute to the immunodeficiency associated with HIV infection, by depleting CD4+ T cells (26). In this study, we have shown that Vδ1 T cells are not expanded in our patients with HIV infection, but their overall percentages are increased, suggesting that these cells are merely preserved in patients with HIV, while other cells are depleted. Since Vδ1 T cells are an important source of innate IL-17 (4, 5), it is also possible that their main role in patients with HIV is to stimulate immunity against co-infecting bacteria and fungi (1–5). Consistent with this hypothesis, we and others have found that Vδ1 T cells expand and produce IL-17 in response to C. albicans and that their frequencies are highest in HIV-positive patients with Candida co-infection (4, 5). Vδ1 T cells are also thought to be major producers of IL-17 in patients with colorectal cancer, in whom they have reduced IFN-γ production (44, 45). However, very few of these cells from healthy donors and patients with primary immunodeficiencies were reported to produce IL-17 (46), suggesting that IL-17 production by Vδ1 T cells is dependent on environmental factors, such as infection.
The TCR consists of a clonotypic αβ or a γδ glycoprotein heterodimer, generated by somatic recombination of germline gene segments, that recognizes antigens associated with antigen-presenting molecules, such as MHC, MR1, or CD1 (10). The TCR polypeptides associate with the CD3 complex, formed by the CD3 γ, δ (not to be confused with the TCR γ and δ polypeptides), ε and ζ subunits, which are invariable and mediate signal transduction. CD3ε can form heterodimers with CD3γ and CD3δ, while CD3ζ frequently exists as a homodimer, and CD3δε, CD3γε, and CD3ζζ are all capable of transducing activating signals in response to TCR ligation (9, 10). The CD3 γ, δ, ε, and ζ polypeptides all contain ITAMs in their cytoplasmic domains, which are required for intracellular assembly and surface expression of the TCR and signal transduction events that mediate thymocyte maturation and mature αβ T cell activation (47–50). Humans and mice lacking CD3ε have no αβ or γδ T cells (49, 51), indicating an absolute requirement for CD3ε in early T cell development. However, unlike in αβ T cells, γδ TCR rearrangement can occur in the absence of CD3ε (50) and some mature γδ T cells do not express CD3ε (52). γδ TCRs can also signal through FcRγ homodimers and CD3ζ-FcRγ heterodimers (52).
In this study, we have identified two populations of Vδ1 T cells, one of which expresses normal levels of CD3ε and the other which appears to express no or low levels of CD3ε, but normal levels of CD3ζ. CD3εlo and CD3εhi Vδ1 T cells were present in PBMC from patients with HIV and in control subjects and in expanded lines of Vδ1 T cells. Using confocal microscopy of sorted CD3εlo and CD3εhi Vδ1 T cells, we show that the absence of CD3ε is unlikely to be due to internalization of the polypeptide, since intracellular CD3ε was not detected. To investigate the stability of CD3ε expression, CD3εlo and CD3εhi Vδ1 T cells were sorted from lines of Vδ1 T cells and restimulated with C. albicans and cultured in the presence of IL-2. We found that CD3εhi Vδ1 T cells could downregulate CD3ε and CD3εlo Vδ1 T cells could upregulate CD3ε expression, suggesting that the expression of this component of CD3 can be modulated by activation and that its downregulation is reversible. CD3ε expression is required for progression of thymocyte maturation from the double positive CD4+CD8+ stage to the single positive CD4+ or CD8+ stage and for assembly of the pre-TCR (49, 50, 53, 54), but appears to be dispensible in mature T cells, where it may act to amplify weak signals from the TCR (55, 56). Thus, it is possible that CD3εlo Vδ1 T cells have a lower responsiveness to antigenic stimulation than CD3εhi Vδ1 T cells. Interestingly, Vδ1 TCR expression was slightly lower in CD3εlo compared to CD3εhi Vδ1 T cells in HIV patients and control subjects, adding further support to this idea. CD3ε contains endocytosis determinants that may contribute to the up- and downregulation of CD3ε on T cells (57) and recent studies have provided evidence that CD3ε expression can be downregulated by tumor-educated tolerogenic DC (58) and possibly by HIV (59, 60). We found that both CD3εlo and CD3εhi Vδ1 T cells are expanded in patients with untreated HIV infection compared to control subjects, but especially in patients with C. albicans co-infection. Thus, CD3εlo Vδ1 T cells accounted for 0.1% of lymphocytes in controls, compared to 0.5% in untreated HIV patients (P = 0.03) and >1% in patients with HIV and Candida infection (P = 0.0001). Likewise, CD3εhi Vδ1 T cells accounted for 0.2% of controls, compared to 2.3% of untreated HIV patients (P = 0.0001) and >3% of patients with HIV and Candida infection (P = 0.015). Future studies are required to identify the antigenic specificities of the Vδ1 TCR and to ascertain if Vδ1 T cell numbers or the ratios of CD3εhi to CD3εlo Vδ1 T cells can be used as a prognostic marker of Candida co-infection.
To determine if CD3εlo Vδ1 T cells display phenotypic or functional differences from CD3εhi Vδ1 T cells, PBMC freshly isolated from healthy donors were enriched for γδ T cells and further analyzed by flow cytometry. We found that CD3εlo Vδ1 T cells more frequently have TD phenotypes and express PD-1, but not LAG-3, compared to CD3εhi Vδ1 T cells, suggesting that they have previously been activated and exist in a state of inactivation. PD-1 and LAG-3 expression by HIV-specific CD4+ and CD8+ T cells is a feature of HIV infection, is associated with T-cell exhaustion and disease progression, and is thought to promote viral persistence (36–42). Our finding that Vδ1 T cells, and especially the CD3εlo subset of Vδ1 T cells, frequently express PD-1 indicates that this induction of exhaustion in HIV infection extends to γδ T cells and suggests that mAb blocking of PD-1 may benefit patients with HIV (61). Previous workers have reported a skewing of Vγ9Vδ2 T cells toward TD in patients with HIV (32, 62), which is associated with impaired IFN-γ production (63). We tested if CD3εlo Vδ1 T cells display properties of exhaustion by testing their ability to produce IL-17, a cardinal function of Vδ1 T cells (4, 5). We found that significant proportions of CD3εhi Vδ1 T cells, but not CD3εlo Vδ1 T cells, produced IL-17 in response to PMA/I stimulation ex vivo. Therefore, CD3εlo Vδ1 T cells may represent a population of inactive, TD T cells. Since IL-17 production is only one of multiple effector activities of Vδ1 T cells, future studies are required to determine if other activities, such as IFN-γ production, are deficient in CD3εlo Vδ1 T cells. Vδ1 and Vγ9Vδ2 T cells expressing low levels of CD3 and exhibiting impaired responses to stimulation have been reported to accumulate in sites of active Mycobacterium tuberculosis infection (64, 65) and Paget et al. (66) reported that murine Vγ6Vδ1+ T cells with low levels of CD3 predominantly produce IFN-γ whereas the same cells with high levels of CD3 produce IL-17. Thus, modulation of CD3ε expression may be a general mechanism for the regulation of γδ T cell activity.
The results of this study indicate that Vδ1 T cells persist in the blood of patients with untreated HIV infection, and especially in patients with Candida co-infection, while other T cells are depleted. Although it is not known if Vδ1 T cells can directly recognize HIV or HIV-infected cells, a recent study has shown that γδ TCR exposure to viruses can promote the expansion of virus-reactive T cells, providing strong evidence that γδ T cells mediate adaptive immune responses to viruses (67). Previous studies have demonstrated that Vδ1 T cells proliferate and release IL-17 in response to C. albicans, by a mechanism that requires IL-23 release from DC (4, 5). The preservation of Vδ1 T cells in patients whose IL-17-producing CD4+ T cells may be depleted by HIV, identifies Vδ1 T cells as an alternative potential source of IL-17. However, it appears that significant proportions of Vδ1 T cells in patients with HIV have been driven to a state of inactivation, expressing TD phenotypes and the inhibitory receptor PD-1 and failing to produce IL-17 upon stimulation. Downregulation of CD3ε, a signaling molecule known to augment TCR-mediated responses (55, 56), may represent another mechanism by which the effector functions of Vδ1 T cells can be inhibited.
Ethics Statement
Ethical approval for this study was obtained from the Joint Research Ethics Committee of St. James’s Hospital and Tallaght Hospitals, Dublin, and all participants gave written, informed consent.
Author Contributions
PD, TR, and DD: conceived the study. PD, CM, MF, KD, AP, JO, A Long, and MD: performed experiments, acquired and analyzed data. DR, SO, and FM: directed and coordinated sample collection. A Loy, JW, and FM: performed sample collection. DD: wrote the manuscript.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
This study was supported by a grant from the Health Research Board of Ireland. JO was supported by a training grant from Irish Aid (Combat Diseases of Poverty Consortium). We thank Dr. Balbino Alcarón for providing the anti-CD3ε mAb (clone SP4). We thank Conleth Feighery, Mark Little, Yasmeen Ghnewa, Vincent O’Reilly, Serena Arduini, Tanya Coulter, and Eilis Dockry for helpful discussions.
Abbreviations
ART, antiretroviral therapy; DC, dendritic cell; FITC, fluorescein isothiocyanate; FMO, fluorescence minus one; HIV, human immunodeficiency virus; IL-17, interleukin-17; IFN-γ, interferon-γ; mAb, monoclonal antibody; MFI, mean fluorescence intensity; MHC, major histocompatibility complex; PBMCs, peripheral blood mononuclear cells; PBS, phosphate buffered saline; PD-1, programmed death-1; PMA/I, phorbol myristate acetate with ionomycin; TCR, T cell receptor; TD, terminally differentiated.
References
1. Poccia F, Malkovsky M, Pollak A, Colizzi V, Sireci G, Salerno A, et al. In vivo γδ T cell priming to mycobacterial antigens by primary Mycobacterium tuberculosis infection and exposure to nonpeptidic ligands. Mol Med (1999) 5:471–6.
2. Matsuzaki G, Yamada H, Kishihara K, Yoshikai Y, Nomoto K. Mechanism of murine Vγ1+ γδ T cell-mediated innate immune response against Listeria monocytogenes infection. Eur J Immunol (2002) 32:928–35. doi:10.1002/1521-4141(200204)32:4<928::AID-IMMU928>3.0.CO;2-I
3. Hebart H, Bollinger C, Fisch P, Sarfati J, Meisner C, Baur M, et al. Analysis of T-cell responses to Aspergillus fumigatus antigens in healthy individuals and patients with hematologic malignancies. Blood (2002) 100:4521–8. doi:10.1182/blood-2002-01-0265
4. Fenoglio D, Poggi A, Catellani S, Battaglia F, Ferrera A, Setti M, et al. Vδ1 T lymphocytes producing IFN-γ and IL-17 are expanded in HIV-1-infected patients and respond to Candida albicans. Blood (2009) 113:6611–8. doi:10.1182/blood-2009-01-198028
5. Maher CO, Dunne K, Comerford R, O’Dea S, Loy A, Woo J, et al. Candida albicans stimulates IL-23 release by human dendritic cells and downstream IL-17 secretion by Vδ1 T cells. J Immunol (2015) 194:5953–60. doi:10.4049/jimmunol.1403066
6. Dechanet J, Merville P, Lim A, Retiere C, Pitard V, Lafarge X, et al. Implication of γδ T cells in the human immune response to cytomegalovirus. J Clin Invest (1999) 103:1437–49. doi:10.1172/JCI5409
7. Nishimura H, Yajima T, Kagimoto Y, Ohata M, Watase T, Kishihara K, et al. Intraepithelial γδ T cells may bridge a gap between innate immunity and acquired immunity to herpes simplex virus type 2. J Virol (2004) 78:4927–30. doi:10.1128/JVI.78.9.4927-4930.2004
8. Roux A, Mourin G, Larsen M, Fastenackels S, Urrutia A, Gorochov G, et al. Differential impact of age and cytomegalovirus infection on the γδ T cell compartment. J Immunol (2013) 191:1300–6. doi:10.4049/jimmunol.1202940
9. Kuhns MS, Badgandi HB. Piecing together the family portrait of TCR-CD3 complexes. Immunol Rev (2012) 250:120–43. doi:10.1111/imr.12000
10. Alcover A, Alarcón B, Di Bartolo V. Cell biology of T cell receptor expression and regulation. Annu Rev Immunol (2018) 36:103–25. doi:10.1146/annurev-immunol-042617-053429
11. Bonneville M, O’Brien RL, Born WK. γδ T cell effector functions: a blend of innate programming and acquired plasticity. Nat Rev Immunol (2010) 10:467–78. doi:10.1038/nri2781
12. Vantourout P, Hayday AC. Six-of-the-best: unique contributions of γδ T cells to immunology. Nat Rev Immunol (2013) 13:88–100. doi:10.1038/nri3384
13. Chien YH, Meyer C, Bonneville M. γδ T cells: first line of defense and beyond. Annu Rev Immunol (2014) 32:121–55. doi:10.1146/annurev-immunol-032713-120216
14. Morita CT, Jin C, Sarikonda G, Wang H. Nonpeptide antigens, presentation mechanisms, and immunological memory of human Vγ2Vδ2 T cells: discriminating friend from foe through the recognition of prenyl pyrophosphate antigens. Immunol Rev (2007) 215:59–76. doi:10.1111/j.1600-065X.2006.00479.x
15. Mangan BA, Dunne MR, O’Reilly VP, Dunne PJ, Exley MA, O’Shea D, et al. Cutting edge: CD1d restriction and Th1/Th2/Th17 cytokine secretion by human Vδ3 T cells. J Immunol (2013) 191:30–4. doi:10.4049/jimmunol.1300121
16. Groh V, Steinle A, Bauer S, Spies T. Recognition of stress-induced MHC molecules by intestinal epithelial γδ T cells. Science (1998) 279:1737–40. doi:10.1126/science.279.5357.1737
17. Spada FM, Grant EP, Peters PJ, Sugita M, Melián A, Leslie DS, et al. Self-recognition of CD1 by γδ T cells: implications for innate immunity. J Exp Med (2000) 191:937–48. doi:10.1084/jem.191.6.937
18. Bai L, Picard D, Anderson B, Chaudhary V, Luoma A, Jabri B, et al. The majority of CD1d-sulfatide-specific T cells in human blood use a semiinvariant Vδ1 TCR. Eur J Immunol (2012) 42:2505–10. doi:10.1002/eji.201242531
19. Uldrich AP, Le Nours J, Pellicci DG, Gherardin NA, McPherson KG, Lim RT, et al. CD1d-lipid antigen recognition by the γδ TCR. Nat Immunol (2013) 14:113711–45. doi:10.1038/ni.2713
20. Zeng X, Wei YL, Huang J, Newell EW, Yu H, Kidd BA, et al. γδ T cells recognize a microbial encoded B cell antigen to initiate a rapid antigen-specific interleukin-17 response. Immunity (2012) 37:524–34. doi:10.1016/j.immuni.2012.06.011
21. Fausther-Bovendo H, Wauquier N, Cherfils-Vicini J, Cremer I, Debré P, Vieillard V. NKG2C is a major triggering receptor involved in the Vδ1 T cell-mediated cytotoxicity against HIV-infected CD4 T cells. AIDS (2008) 22:217–26. doi:10.1097/QAD.0b013e3282f46e7c
22. Wesch D, Peters C, Oberg HH, Pietschmann K, Kabelitz D. Modulation of γδ T cell responses by TLR ligands. Cell Mol Life Sci (2011) 68:2357–70. doi:10.1007/s00018-011-0699-1
23. Hudspeth K, Fogli M, Correia DV, Mikulak J, Roberto A, Della Bella S, et al. Engagement of NKp30 on Vδ1 T cells induces the production of CCL3, CCL4, and CCL5 and suppresses HIV-1 replication. Blood (2012) 119:4013–6. doi:10.1182/blood-2011-11-390153
24. Kuroda H, Saito H, Ikeguchi M. Decreased number and reduced NKG2D expression of Vδ1 γδ T cells are involved in the impaired function of Vδ1 γδ T cells in the tissue of gastric cancer. Gastric Cancer (2012) 15:433–9. doi:10.1007/s10120-011-0138-x
25. Boullier S, Dadaglio G, Lafeuillade A, Debord T, Gougeon ML. Vδ1 T cells expanded in the blood throughout HIV infection display a cytotoxic activity and are primed for TNF-α and IFN-γ production but are not selected in lymph nodes. J Immunol (1997) 159:3629–37.
26. Sindhu ST, Ahmad R, Morisset R, Ahmad A, Menezes J. Peripheral blood cytotoxic γδ T lymphocytes from patients with human immunodeficiency virus type 1 infection and AIDS lyse uninfected CD4+ T cells, and their cytocidal potential correlates with viral load. J Virol (2003) 77:1848–55. doi:10.1128/JVI.77.3.1848-1855.2003
27. Dobmeyer TS, Dobmeyer R, Wesch D, Helm EB, Hoelzer D, Kabelitz D. Reciprocal alterations of Th1/Th2 function in γδ T-cell subsets of human immunodeficiency virus-1-infected patients. Br J Haematol (2002) 118:282–8. doi:10.1046/j.1365-2141.2002.03555.x
28. De Maria A, Ferrazin A, Ferrini S, Ciccone E, Terragna A, Moretta L. Selective increase of a subset of T cell receptor γδ T lymphocytes in the peripheral blood of patients with human immunodeficiency virus type 1 infection. J Infect Dis (1992) 165:917–9. doi:10.1093/infdis/165.5.917
29. Agostini C, Zambello R, Trentin L, Cerutti A, Bulian P, Crivellaro C, et al. γδ T cell receptor subsets in the lung of patients with HIV-1 infection. Cell Immunol (1994) 153:194–205. doi:10.1006/cimm.1994.1017
30. Poles MA, Barsoum S, Yu W, Yu J, Sun P, Daly J, et al. Human immunodeficiency virus type 1 induces persistent changes in mucosal and blood γδ T cells despite suppressive therapy. J Virol (2003) 77:10456–67. doi:10.1128/JVI.77.19.10456-10467.2003
31. Nilssen DE, Brandtzaeg P. Intraepithelial γδ T cells remain increased in the duodenum of AIDS patients despite antiretroviral treatment. PLoS One (2012) 7(1):e29066. doi:10.1371/journal.pone.0029066
32. Li Z, Li W, Li N, Jiao Y, Chen D, Cui L, et al. γδ T cells are involved in acute HIV infection and associated with AIDS progression. PLoS One (2014) 9:e106064. doi:10.1371/journal.pone.0106064
33. Cimini E, Agrati C, D’Offizi G, Vlassi C, Casetti R, Sacchi A, et al. Primary and chronic HIV infection differently modulates mucosal Vδ1 and Vδ2 T-cells differentiation profile and effector functions. PLoS One (2015) 10:e0129771. doi:10.1371/journal.pone.0129771
34. Dunne MR, Madrigal-Estebas L, Tobin LM, Doherty DG. (E)-4-hydroxy-3-methyl-but-2 enyl pyrophosphate-stimulated Vγ9Vδ2 T cells possess T helper type 1-promoting adjuvant activity for human monocyte-derived dendritic cells. Cancer Immunol Immunother (2010) 59:1109–20. doi:10.1007/s00262-010-0839-8
35. Dieli F, Poccia F, Lipp M, Sireci G, Caccamo N, Di Sano C, et al. Differentiation of effector/memory Vδ2 T cells and migratory routes in lymph nodes or inflammatory sites. J Exp Med (2003) 198:391–7. doi:10.1084/jem.20030235
36. Petrovas C, Casazza JP, Brenchley JM, Price DA, Gostick E, Adams WC, et al. PD-1 is a regulator of virus-specific CD8+ T cell survival in HIV infection. J Exp Med (2006) 203:2281–92. doi:10.1084/jem.20061496
37. Day CL, Kaufmann DE, Kiepiela P, Brown JA, Moodley ES, Reddy S, et al. PD-1 expression on HIV-specific T cells is associated with T-cell exhaustion and disease progression. Nature (2006) 443:350–4. doi:10.1038/nature05115
38. Trautmann L, Janbazian L, Chomont N, Said EA, Gimmig S, Bessette B, et al. Upregulation of PD-1 expression on HIV-specific CD8+ T cells leads to reversible immune dysfunction. Nat Med (2006) 12:1198–202. doi:10.1038/nm1106-1329b
39. Tendeiro R, Foxall RB, Baptista AP, Pinto F, Soares RS, Cavaleiro R, et al. PD-1 and its ligand PD-L1 are progressively up-regulated on CD4 and CD8 T-cells in HIV-2 infection irrespective of the presence of viremia. AIDS (2012) 26:1065–71. doi:10.1097/QAD.0b013e32835374db
40. Cockerham LR, Jain V, Sinclair E, Glidden DV, Hartogenesis W, Hatano H, et al. Programmed death-1 expression on CD4+ and CD8+ T cells in treated and untreated HIV disease. AIDS (2014) 28:1749–58. doi:10.1097/QAD.0000000000000314
41. Tian X, Zhang A, Qiu C, Wang W, Yang Y, Qiu C, et al. The upregulation of LAG-3 on T cells defines a subpopulation with functional exhaustion and correlates with disease progression in HIV-infected subjects. J Immunol (2015) 194:3873–82. doi:10.4049/jimmunol.1402176
42. Hurst J, Hoffmann M, Pace M, Williams JP, Thornhill J, Hamlyn E, et al. Immunological biomarkers predict HIV-1 viral rebound after treatment interruption. Nat Commun (2015) 6:8495. doi:10.1038/ncomms9495
43. Kimmig S, Przybylski GK, Schmidt CA, Laurisch K, Möwes B, Radbruch A, et al. Two subsets of naive T helper cells with distinct T cell receptor excision circle content in human adult peripheral blood. J Exp Med (2002) 195:789–94. doi:10.1084/jem.20011756
44. Wu P, Wu D, Ni C, Ye J, Chen W, Hu G, et al. γδT17 cells promote the accumulation and expansion of myeloid-derived suppressor cells in human colorectal cancer. Immunity (2014) 40:785–800. doi:10.1016/j.immuni.2014.03.013
45. Meraviglia S, Lo Presti E, Tosolini M, La Mendola C, Orlando V, Todaro M, et al. Distinctive features of tumor-infiltrating γδ T lymphocytes in human colorectal cancer. Oncoimmunology (2017) 6:e1347742. doi:10.1080/2162402X.2017.1347742
46. Dillen CA, Pinsker BL, Marusina AI, Merleev AA, Farber ON, Liu H, et al. Clonally expanded γδ T cells protect against Staphylococcus aureus skin reinfection. J Clin Invest (2018) 128:1026–42. doi:10.1172/JCI96481
47. Love PE, Shores EW, Johnson MD, Tremblay ML, Lee EJ, Grinberg A, et al. T cell development in mice that lack the ζ chain of the T cell antigen receptor complex. Science (1993) 261:918–21. doi:10.1126/science.7688481
48. Haks MC, Krimpenfort P, Borst J, Kruisbeek AM. The CD3γ chain is essential for development of both the TCRαβ and TCRγδ lineages. EMBO J (1998) 17:1871–82. doi:10.1093/emboj/17.7.1871
49. DeJarnette JB, Sommers CL, Huang K, Woodside KJ, Emmons R, Katz K, et al. Specific requirement for CD3ε in T cell development. Proc Natl Acad Sci U S A (1998) 95:14909–14. doi:10.1073/pnas.95.25.14909
50. Malissen M, Gillet A, Ardouin L, Bouvier G, Trucy J, Ferrier P, et al. Altered T cell development in mice with a targeted mutation of the CD3-ε gene. EMBO J (1995) 14:4641–53.
51. de Saint Basile G, Geissmann F, Flori E, Uring-Lambert B, Soudais C, Cavazzana-Calvo M, et al. Severe combined immunodeficiency caused by deficiency in either the δ or the ε subunit of CD3. J Clin Invest (2004) 114:1512–7. doi:10.1172/JCI200422588
52. Hayes SM, Love PE. Distinct structure and signaling potential of the γδ TCR complex. Immunity (2002) 16:827–38. doi:10.1016/S1074-7613(02)00320-5
53. Wang B, Wang N, Whitehurst CE, She J, Chen J, Terhorst C. T lymphocyte development in the absence of CD3 ε or CD3 γδεζ. J Immunol (1999) 162:88–94.
54. Brodeur JF, Li S, da Silva Martins M, Larose L, Dave VP. Critical and multiple roles for the CD3ε intracytoplasmic tail in double negative to double positive thymocyte differentiation. J Immunol (2009) 182:4844–53. doi:10.4049/jimmunol.0803679
55. Tailor P, Tsai S, Shameli A, Serra P, Wang J, Robbins S, et al. The proline-rich sequence of CD3ε as an amplifier of low-avidity TCR signaling. J Immunol (2008) 181:243–55. doi:10.4049/jimmunol.181.1.243
56. Mingueneau M, Sansoni A, Grégoire C, Roncagalli R, Aguado E, Weiss A, et al. The proline-rich sequence of CD3ε controls T cell antigen receptor expression on and signaling potency in preselection CD4+CD8+ thymocytes. Nat Immunol (2008) 9:522–32. doi:10.1038/ni.1608
57. Borroto A, Lama J, Niedergang F, Dautry-Varsat A, Alarcón B, Alcover A. The CD3 ε subunit of the TCR contains endocytosis signals. J Immunol (1999) 163:25–31.
58. Kuang DM, Zhao Q, Xu J, Yun JP, Wu C, Zheng L. Tumor-educated tolerogenic dendritic cells induce CD3ε down-regulation and apoptosis of T cells through oxygen-dependent pathways. J Immunol (2008) 181:3089–98. doi:10.4049/jimmunol.181.5.3089
59. Dorival C, Brizzi F, Lelièvre JD, Sol-Foulon N, Six E, Henry A, et al. HIV-1 Nef protein expression in human CD34+ progenitors impairs the differentiation of an early T/NK cell precursor. Virology (2008) 377:207–15. doi:10.1016/j.virol.2008.04.009
60. Rotem E, Reuven EM, Klug YA, Shai Y. The transmembrane domain of HIV-1 gp41 inhibits T-cell activation by targeting multiple T-cell receptor complex components through its GxxxG motif. Biochemistry (2016) 55:1049–57. doi:10.1021/acs.biochem.5b01307
61. Seung E, Dudek TE, Allen TM, Freeman GJ, Luster AD, Tager AM. PD-1 blockade in chronically HIV-1-infected humanized mice suppresses viral loads. PLoS One (2013) 8:e77780. doi:10.1371/journal.pone.0077780
62. Hartjen P, Meyer-Olson D, Lehmann C, Stellbrink HJ, van Lunzen J, Schulze zur Wiesch J. Vγ2Vδ2 T cells are skewed toward a terminal differentiation phenotype in untreated HIV infection. J Infect Dis (2013) 208:180–2. doi:10.1093/infdis/jit141
63. Martini F, Poccia F, Goletti D, Carrara S, Vincenti D, D’Offizi G, et al. Acute human immunodeficiency virus replication causes a rapid and persistent impairment of Vγ9Vδ2 T cells in chronically infected patients undergoing structured treatment interruption. J Infect Dis (2002) 186:847–50. doi:10.1086/342410
64. Yokobori N, Schierloh P, Geffner L, Balboa L, Romero M, Musella R, et al. CD3 expression distinguishes two gammadeltaT cell receptor subsets with different phenotype and effector function in tuberculous pleurisy. Clin Exp Immunol (2009) 157:385–94. doi:10.1111/j.1365-2249.2009.03974.x
65. El Daker S, Sacchi A, Montesano C, Altieri AM, Galluccio G, Colizzi V, et al. An abnormal phenotype of lung Vγ9Vδ2 T cells impairs their responsiveness in tuberculosis patients. Cell Immunol (2013) 282:106–12. doi:10.1016/j.cellimm.2013.05.001
66. Paget C, Chow MT, Gherardin NA, Beavis PA, Uldrich AP, Duret H, et al. CD3bright signals on γδ T cells identify IL-17A-producing Vγ6Vδ1+ T cells. Immunol Cell Biol (2015) 93:198–212. doi:10.1038/icb.2014.94
Keywords: human immunodeficiency virus, Vδ1 T cells, CD3ε, interleukin-17, programmed death-1, flow cytometry
Citation: Dunne PJ, Maher CO, Freeley M, Dunne K, Petrasca A, Orikiiriza J, Dunne MR, Reidy D, O’Dea S, Loy A, Woo J, Long A, Rogers TR, Mulcahy F and Doherty DG (2018) CD3ε Expression Defines Functionally Distinct Subsets of Vδ1 T Cells in Patients With Human Immunodeficiency Virus Infection. Front. Immunol. 9:940. doi: 10.3389/fimmu.2018.00940
Received: 31 January 2018; Accepted: 16 April 2018;
Published: 02 May 2018
Edited by:
Francesco Dieli, Università degli Studi di Palermo, ItalyReviewed by:
Andrea Knight, Masaryk University, CzechiaMartin S. Davey, University of Birmingham, United Kingdom
Copyright: © 2018 Dunne, Maher, Freeley, Dunne, Petrasca, Orikiiriza, Dunne, Reidy, O’Dea, Loy, Woo, Long, Rogers, Mulcahy and Doherty. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Derek G. Doherty, ZGVyZWsuZG9oZXJ0eUB0Y2QuaWU=