 Susanna Valanne1
Susanna Valanne1 Mika Rämet
Mika Rämet- 1Laboratory of Experimental Immunology, Faculty of Medicine and Life Sciences, BioMediTech Institute, University of Tampere, Tampere, Finland
- 2PEDEGO Research Unit, Medical Research Center Oulu, University of Oulu, Oulu, Finland
- 3Department of Children and Adolescents, Oulu University Hospital, Oulu, Finland
A commentary on
Phagocytosis of particles by cells is an ancient, evolutionarily highly conserved process. It is essential for normal development, tissue homeostasis, and immunity in a wide range of organisms from flies to man. In Drosophila melanogaster, plasmatocytes, the most abundant blood cell type, participate both in the elimination of apoptotic corpses during development (1, 2) and in the receptor-mediated phagocytosis of microbes, an indispensable process for immunity (3–5).
The ability of phagocytic cells to recognize and internalize particles is based on the expression of various phagocytic receptors. In D. melanogaster, the key receptors for recognition of both apoptotic corpses (Six-Microns-Under, Draper, Croquemort) and microbes (Eater, Scavenger receptor-CI, Nimrod) are rather well described (3, 6–11).
In the recent issue of Frontiers in Immunology, Shlyakhover and his co-workers (12) elegantly describe the central role of transcriptional regulator Serpent for the phagocytic ability of embryonic macrophages in D. melanogaster. Serpent is a GATA factor, which is shown by the authors to be both required and sufficient for the expression of phagocytic receptors needed for engulfment of apoptotic corpses in the embryonic macrophages. In the Serpent mutant embryos, phagocytosis of apoptotic corpses was severely impaired, and this was associated with a loss of Six-Microns-Under, Draper, and Croquemort expression. Furthermore, expression of any of these receptors partially rescued the phagocytosis deficiency in the Serpent mutants.
Thus, this recent study beautifully demonstrates the central role of Serpent as the master regulator of phagocytosis of apoptotic cells by controlling the expression of receptors required for recognition of these particles. However, it has been a long-standing observation that serpent expression—together with components of the transcriptional coactivator Mediator complex Med12–Med13—is also required for phagocytosis of microbes in macrophage-like, D. melanogaster embryo-derived S2 cells (13, 14) (Figure 1). Serpent was identified in the very first high-throughput RNA interference (RNAi) screen as a regulator of bacterial cell surface binding and phagocytosis (13). RNAi targeting serpent reduced phagocytosis of heat-killed, FITC-labeled Escherichia coli (phagocytic index 21 ± 11 of controls) and Staphylococcus aureus (phagocytic index 20 ± 6 of controls) (3). Therefore, Serpent appears to control the expression of cell surface proteins responsible for microbial binding and uptake in addition to receptors required for the recognition of apoptotic corpses (Figure 1). In fact, by performing a transcriptional analysis of S2 cells in which serpent was knocked down by RNAi (3, 14), 46 genes were identified with signal sequence and/or transmembrane domain whose expression was downregulated by more than twofold after serpent RNAi (3). These included known phagocytic receptors for microbes (Eater and Scavenger receptor-CI) indicating that Serpent is a master regulator of also microbial pattern recognition in phagocytosis. Of note, our analysis of Serpent-dependently expressed genes revealed also Six-Microns-Under (also called Nimrod C4 and CG16876) as one of the genes expressed in Serpent-dependent manner.
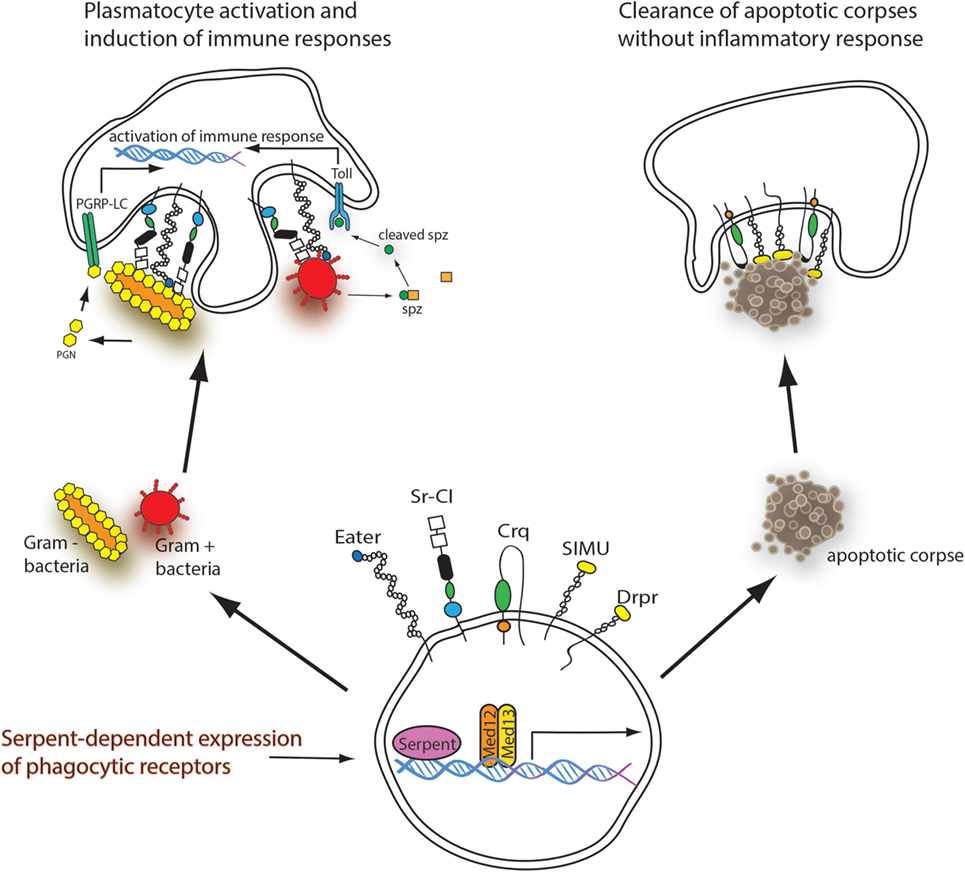
Figure 1. Serpent-dependent expression of phagocytic receptors. The naïve hemocyte expresses both apoptotic and immune-related phagocytosis receptors in a Serpent and Med12/Med13-dependent manner. Upon recognition of “eat me” signals produced by apoptotic corpses, the receptors Croquemort (Crq), Draper (Drpr), and Six-Microns-Under (SIMU) bind to the apoptotic corpse and initiate its phagocytosis and degradation without inflammatory response (right). When the hemocyte meets bacteria, the phagocytic receptors Eater and scavenger receptor CI (Sr-CI) bind to the bacteria initiating phagocytosis. Bacteria also induce systemic and cellular immune responses via the Imd and the toll pathways (left).
Altogether, these findings elaborate the importance of GATA factor Serpent in transcriptional control of the overall phagocytic competence of macrophage-like cells in D. melanogaster. As the same transcription factor controls the expression of genes necessary for clearance of particles without inflammatory response as well as immune response associated receptors, it seems that in Drosophila, professional phagocytic cells possess capability for both anti- and pro-inflammatory responses depending on the cargo they recognize. Upon immune activation, Drosophila hemocytes produce antimicrobial peptides (15) and change their morphology (16, 17). This resembles polarization of mammalian macrophages toward a “pro-inflammatory” M1 phenotype by exposure to lipopolysaccharide together with TH1 cytokine IFN-γ. As a consequence, M1 polarized macrophage is an effector cell in TH1 cellular immune responses, whereas the alternatively activated M2 macrophage has immunosuppressive properties, for example, in wound healing and tissue repair. In contrast to mammals, Drosophila lacks interferon-γ and adaptive immunity, so there must be alternative means to direct plasmatocyte polarization. With numerous advantageous genetic tools together with efforts to define hemocyte lineages using in vivo hemocyte reporters (16), D. melanogaster will continue to be an exciting model to study factors affecting differentiation of hemocyte lineages and mechanisms controlling phagocytic competence and activation of plasmatocytes.
Author Contributions
All authors wrote and approved the final version of the manuscript.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
1. Abrams JM, White K, Fessler LI, Steller H. Programmed cell death during Drosophila embryogenesis. Development (1993) 117(1):29–43.
2. Tepass U, Fessler LI, Aziz A, Hartenstein V. Embryonic origin of hemocytes and their relationship to cell death in Drosophila. Development (1994) 120(7):1829–37.
3. Kocks C, Cho JH, Nehme N, Ulvila J, Pearson AM, Meister M, et al. Eater, a transmembrane protein mediating phagocytosis of bacterial pathogens in Drosophila. Cell (2005) 123(2):335–46. doi:10.1016/j.cell.2005.08.034
4. Elrod-Erickson M, Mishra S, Schneider D. Interactions between the cellular and humoral immune responses in Drosophila. Curr Biol (2000) 10(13):781–4. doi:10.1016/S0960-9822(00)00569-8
5. Ulvila J, Vanha-aho LM, Kleino A, Vähä-Mäkilä M, Vuoksio M, Eskelinen S, et al. Cofilin regulator 14-3-3zeta is an evolutionarily conserved protein required for phagocytosis and microbial resistance. J Leukoc Biol (2011) 89(5):649–59. doi:10.1189/jlb.0410195
6. Franc NC, Dimarcq JL, Lagueux M, Hoffmann J, Ezekowitz RA. Croquemort, a novel Drosophila hemocyte/macrophage receptor that recognizes apoptotic cells. Immunity (1996) 4(5):431–43. doi:10.1016/S1074-7613(00)80410-0
7. Kurant E, Axelrod S, Leaman D, Gaul U. Six-microns-under acts upstream of draper in the glial phagocytosis of apoptotic neurons. Cell (2008) 133(3):498–509. doi:10.1016/j.cell.2008.02.052
8. Freeman MR, Delrow J, Kim J, Johnson E, Doe CQ. Unwrapping glial biology: GCM target genes regulating glial development, diversification, and function. Neuron (2003) 38(4):567–80. doi:10.1016/S0896-6273(03)00289-7
9. Rämet M, Pearson A, Manfruelli P, Li X, Koziel H, Göbel V, et al. Drosophila scavenger receptor CI is a pattern recognition receptor for bacteria. Immunity (2001) 15(6):1027–38. doi:10.1016/S1074-7613(01)00249-7
10. Kurucz E, Márkus R, Zsámboki J, Folkl-Medzihradszky K, Darula Z, Vilmos P, et al. Nimrod, a putative phagocytosis receptor with EGF repeats in Drosophila plasmatocytes. Curr Biol (2007) 17(7):649–54. doi:10.1016/j.cub.2007.02.041
11. Ulvila J, Vanha-aho LM, Rämet M. Drosophila phagocytosis – still many unknowns under the surface. APMIS (2011) 119(10):651–62. doi:10.1111/j.1600-0463.2011.02792.x
12. Shlyakhover E, Shklyar B, Hakim-Mishnaevski K, Levy-Adam F, Kurant E. Drosophila GATA factor serpent establishes phagocytic ability of embryonic macrophages. Front Immunol (2018) 9:266. doi:10.3389/fimmu.2018.00266
13. Rämet M, Manfruelli P, Pearson A, Mathey-Prevot B, Ezekowitz RA. Functional genomic analysis of phagocytosis and identification of a Drosophila receptor for E. coli. Nature (2002) 416(6881):644–8. doi:10.1038/nature735
14. Kuuluvainen E, Hakala H, Havula E, Sahal Estimé M, Rämet M, Hietakangas V, et al. Cyclin-dependent kinase 8 module expression profiling reveals requirement of mediator subunits 12 and 13 for transcription of serpent-dependent innate immunity genes in Drosophila. J Biol Chem (2014) 289(23):16252–61. doi:10.1074/jbc.M113.541904
15. Irving P, Ubeda JM, Doucet D, Troxler L, Lagueux M, Zachary D, et al. New insights into Drosophila larval haemocyte functions through genome-wide analysis. Cell Microbiol (2005) 7(3):335–50. doi:10.1111/j.1462-5822.2004.00462.x
16. Anderl I, Vesala L, Ihalainen TO, Vanha-aho LM, Andó I, Rämet M, et al. Transdifferentiation and proliferation in two distinct hemocyte lineages in Drosophila melanogaster larvae after wasp infection. PLoS Pathog (2016) 12(7):e1005746. doi:10.1371/journal.ppat.1005746
Keywords: phagocytosis, Drosophila melanogaster, apoptosis, innate immune response, plasmatocyte
Citation: Valanne S, Vesala L and Rämet M (2018) Commentary: Drosophila GATA Factor Serpent Establishes Phagocytic Ability of Embryonic Macrophages. Front. Immunol. 9:1582. doi: 10.3389/fimmu.2018.01582
Received: 09 April 2018; Accepted: 26 June 2018;
Published: 06 July 2018
Edited by:
Uday Kishore, Brunel University London, United KingdomReviewed by:
Raymond B. Birge, The State University of New Jersey, United StatesJonathan P. Rast, Emory University School of Medicine, United States
Copyright: © 2018 Valanne, Vesala and Rämet. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Mika Rämet, bWlrYS5yYW1ldEB1dGEuZmk=