 Julia Roider1,2,3,4
Julia Roider1,2,3,4 Abigail Ngoepe1
Abigail Ngoepe1 Maximilian Muenchhoff5,6
Maximilian Muenchhoff5,6 Emily Adland2Andreas Groll7Thumbi Ndung'u1,3,8,9,10
Emily Adland2Andreas Groll7Thumbi Ndung'u1,3,8,9,10 Henrik Kløverpris1,10,11
Henrik Kløverpris1,10,11 Philip Goulder2,3*†
Philip Goulder2,3*† Alasdair Leslie1,10*†
Alasdair Leslie1,10*†- 1Africa Health Research Institute, University of KwaZulu-Natal, Durban, South Africa
- 2Department of Paediatrics, Peter Medawar Building for Pathogen Research, Oxford University, Oxford, United Kingdom
- 3HIV Pathogenesis Programme, Doris Duke Medical Research Institute, University of KwaZulu-Natal, Durban, South Africa
- 4Department of Infectious Diseases, Medizinische Klinik IV, Ludwig-Maximilians-University Munich, Munich, Germany
- 5Department of Virology, Max von Pettenkofer Institute, Ludwig-Maximilians-University Munich, Munich, Germany
- 6German Center for Infection Research (DZIF), Partner Site Munich, Munich, Germany
- 7Faculty of Statistics, TU Dortmund University, Dortmund, Germany
- 8Ragon Institute of Massachusetts General Hospital, Massachusetts Institute of Technology and Harvard University, Cambridge, MA, United States
- 9Max Planck Institute for Infection Biology, Berlin, Germany
- 10Department of Infection and Immunity, University College London, London, United Kingdom
- 11Department of Immunology and Microbiology, University of Copenhagen, Copenhagen, Denmark
Pediatric slow progressors (PSP) are rare ART-naïve, HIV-infected children who maintain high CD4 T-cell counts and low immune activation despite persistently high viral loads. Using a well-defined cohort of PSP, we investigated the role of regulatory T-cells (TREG) and of IL-7 homeostatic signaling in maintaining normal-for-age CD4 counts in these individuals. Compared to children with progressive disease, PSP had greater absolute numbers of TREG, skewed toward functionally suppressive phenotypes. As with immune activation, overall T-cell proliferation was lower in PSP, but was uniquely higher in central memory TREG (CM TREG), indicating active engagement of this subset. Furthermore, PSP secreted higher levels of the immunosuppressive cytokine IL-10 than children who progressed. The frequency of suppressive TREG, CM TREG proliferation, and IL-10 production were all lower in PSP who go on to progress at a later time-point, supporting the importance of an active TREG response in preventing disease progression. In addition, we find that IL-7 homeostatic signaling is enhanced in PSP, both through preserved surface IL-7receptor (CD127) expression on central memory T-cells and increased plasma levels of soluble IL-7receptor, which enhances the bioactivity of IL-7. Combined analysis, using a LASSO modeling approach, indicates that both TREG activity and homeostatic T-cell signaling make independent contributions to the preservation of CD4 T-cells in HIV-infected children. Together, these data demonstrate that maintenance of normal-for-age CD4 counts in PSP is an active process, which requires both suppression of immune activation through functional TREG, and enhanced T-cell homeostatic signaling.
Introduction
In the absence of antiretroviral therapy (ART), HIV-infected children typically progress rapidly to AIDS (1). “Pediatric slow progressors” (PSP) are a subgroup of vertically HIV-infected children who maintain their CD4 counts despite being antiretroviral treatment-naive (2–7). These children share certain fundamental immunological characteristics with the natural hosts of nonpathogenic SIV infection such as sooty mangabeys, including low immune activation on CD4 T-cells in the face of persistent high viral loads (7–10). Here we investigate the role of immune regulation by regulatory T-cells (TREG) and of IL-7-mediated T-cell homeostasis, in maintaining this beneficial phenotype in a well-described cohort of PSP (7).
Persistent immune activation is a hallmark of HIV infection in humans and is an important driver of disease progression that is independent of viral load (11–13). Indeed, in pediatric HIV infection, it is CD4 T-cell activation and not viral load that is the strongest predictor of CD4 decline (7, 13, 14). In the non-human primate model of HIV infection, the rapid resolution of immune activation during acute infection also distinguishes non-pathogenic infection of SIV in sooty mangabeys from pathogenic infection in rhesus macaques (15). In sooty mangabeys, this resolution of immune activation is associated with an up-regulation of the canonical TREG markers CD25 and FoxP3 on CD4 and CD8 T-cells (16), and increased production of the key regulatory cytokine IL-10 (17). Moreover, TREG numbers in sooty mangabeys remain stable, whilst they are severely depleted in rhesus macaques, in correlation with disease state (18). The importance of TREG in pediatric non-progression has not been assessed to date, but the similarities between PSP and non-progressive SIV infection suggest a possible role for these cells.
TREG are required to prevent autoimmunity and to regulate over-reactive immunity in the setting of chronic infections, including HIV infection (19–21). They can exert their suppressive function through cell-cell contact (22, 23), disruption of metabolic pathways (24), or by releasing soluble suppressive factors like IL-10 or TGF-b (25, 26) [reviewed in Yamaguchi et al. (27)]. TREG make up ~1% of CD4 T cells in untreated HIV-infected adults, but are more frequent in early life, and can constitute up to 15% of total CD4 T-cells in the fetus (28). Their frequency is inversely correlated with HIV-driven immune activation (14, 29–34), and a decline in absolute TREG numbers coincides with disease progression (14, 33, 35–38). In addition, TREG have been shown to be comprised of phenotypically and functionally diverse subsets. Miyara et al. identified three phenotypically distinct subsets that differed in their suppressive capacity: CD45RA+FoxP3lo resting TREG cells and CD45RA−FoxP3hi activated TREG cells, both of which were suppressive in vitro, and a cytokine-secreting CD45RA−FoxP3lo subset that did not display suppressive activity in vitro. Terminally differentiated activated TREG cells rapidly died whereas resting TREG cells proliferated and converted into activated TREG cells in vitro and in vivo (39).
In addition to TREG activity, non-progression in sooty mangabeys has been linked to preservation of IL-7 signaling in T-cells (40). This pleiotropic cytokine is crucial for the development and homeostasis of T-cells, promotes antigen-specific expansion and memory formation (41–44), and can reverse T-cell exhaustion (45). Immune failure in both adult and pediatric HIV infection is associated with perturbations in IL-7 signaling (46, 47) and reduced responsiveness (48, 49). In adult long-term non-progressors, IL-7R (CD127) expression is preserved on central memory and effector memory CD4 T-cell compartments when compared to untreated progressors (50). Again, however, the importance of IL-7 to sustaining CD4 T-cell levels in PSP remains unknown.
In this study, we investigate two mechanisms by which PSP maintain their CD4 counts in the face of on-going viral replication: first, via strong regulatory T-cell responses that reduce immune activation; and, second, via intact IL-7 receptor signaling that preserves homeostatic proliferation. Both mechanisms are interlinked by the main driver of pathogenesis in HIV infection: chronic immune activation.
Materials and Methods
Study Participants
Peripheral blood mononuclear cells (PBMC) and plasma of vertically HIV-1 C clade-infected children and age matched healthy controls all from Southern Africa and predominantly of Zulu origin, were obtained from clinics in Durban, South Africa (Ithembalabantu Clinic and Prince Mshiyeni Hospital) (see Table 1 for cohort characteristics). In the present study, pediatric slow progressors (PSPN; n = 12) are defined as vertically HIV-infected, ART-naïve, CD4 count >450/mm3 at age >5 years. Pediatric future progressors (PFP; n = 11) were meeting inclusion criteria for PSPN at the time point used for the experiments but progressed in the longitudinal follow-up. Pediatric progressors (PP; n = 10) are defined here as vertically HIV-infected, ART-naïve, CD4 count <350/mm3 at age >5 years. This pediatric cohort has been followed up for over 5 years.
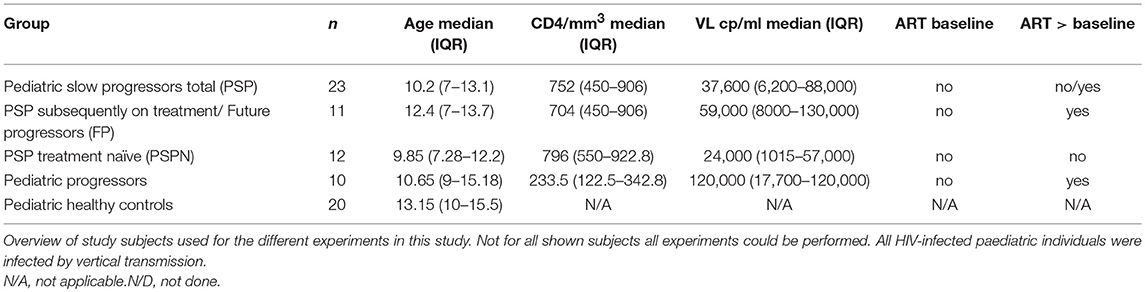
Table 1. Clinical characteristics of study cohort.
Viral load measurement was performed using the COBRA AmpliPrep/COBAS TaqMan HIV-1 Test version 2.0 by Roche (CAP/CTMv2.0) (range, 20–10 millioncopies/ml) except at the Ithembalabantu Clinic where the BioMérieux NucliSens Version 2.0 Easy Q/Easy Mag (NucliSens v2.0) assay (range, 20–10 million copies/ml) was used.
Written informed consent was obtained for underage children from their caregivers. Additionally, assent to participate in the study was given directly by children in the appropriate age groups. Studies were approved by the University of the Free State Ethics Committee, Bloemfontein; Biomedical Research Ethics Committee, University of KwaZulu-Natal, Durban; and Research Ethics Committee, University of Oxford.
Sample Processing—PBMC and Plasma
Plasma was separated by centrifugation and crypopreserved at −80°C. Peripheral blood mononuclear cells (PBMCs) were isolated by Ficoll density gradient centrifugation and stored in liquid nitrogen until use.
Flow Cytometry and ICS Assays
PBMCs were stained with fluorescent monoclonal antibodies against markers previously associated with TREG / immune activation /IL-7R (Supplementary Table 1). Briefly, cells were thawed and rested in R10 medium overnight at 37°C in 5% CO2. For phenotyping cells were washed in 15 ml of PBS, centrifuged at 500 g for 5 min and room temperature and the supernatant removed. Cells were resuspended in a volume of 25 μl with titrated concentrations of fluorochrome-conjugated monoclonal antibodies against cell surface markers and the L/D fixable blue dead stain (Thermo Fisher Scientific) as a viability marker (antibodies used are listed in Supplementary Table S1) and stained for 20 min in the dark. Cells were again washed x 2 in PBS and resuspended in Fix/Perm solution (ebioscience) for intracellular staining (Ki-67, FoxP3) and incubated for 20 min at 4°C, followed by washing and incubating 20 min in 20% Goat serum for Fc-receptor blockade. Afterwards, cells were incubated with intracellular antibodies for 20 min at room temperature in the dark, washed x 2 and the pellet was suspended in 2% paraformaldehyde in PBS. For intracellular cytokine staining assays cells were first stimulated with PMA/Ionomycin (at a final concentration of 4 × 10−5M) in the presence of anti-CD28 and anti-CD49 (1 mg/ml), Brefeldin A and Monensin (5 mg/ml) (BD biosciences), for 5 h, followed by surface staining and intracellular staining as above. Rainbow beads were run at every experiment and compensation adjusted to keep peaks identical to ensure longitudinal comparability of experiments. Flow cytometry acquisition was performed on a BD LSRFortessa within 5 h of staining and analyzed using FlowJo version 9.9.5.
IL-7 and sCD127 Plasma Assays
Plasma levels of IL-7 and sCD127 were quantified using commercially available enzyme-linked immunosorbent assay kits (R&D Systems and Neobiolab, respectively) neat in duplicate. Results are expressed in picograms per millilitre for IL-7 and nanograms per millilitre for sCD127.
Statistical Analyses
Most statistical analyses were undertaken using Prism GraphPad Software version 7.0. After confirming non-Gaussian distribution within the majority of the parameters analyzed, Mann–Whitney test was used to compare continuous factors between two groups and Kruskal-Wallis test to compare continuous factors between > two groups and Benjamini, Krieger and Yekutieli's procedure for false discovery rate to correct for multiple comparisons. For paired analyses, Wilcoxon test was used and for paired analyses at multiple time points 2-way ANOVA analyses. Bivariate correlation analyses were performed using the Spearman rank correlation method with exact permutation P-values calculated. All P-values are two-sided, and a P-value of <0.05 was considered significant. In scatterplots, median values, and interquartile range are indicated.
The influence of several predictor variables (n = 39) on the absolute number of CD4 T-cell counts was assessed by least absolute shrinkage and selection operator (LASSO) principle (51) on scaled covariates using the generalized linear model (GLM) glmnet package of the statistical software R as described before (7, 52). For this method, the optimal tuning parameter λ, which reflects the amount of penalization and, hence, controls variable selection, is determined via 5-fold cross-validation on the basis of the model's deviance. A final unregularized conventional GLM (post-LASSO) is fitted on the selected set of covariates using the R-function glm (Supplementary Figures 1A,B).
A correlation matrix was used to visualize Spearman's correlations between selected variables of the LASSO analysis, comprising absolute CD4 T-cell count; viral load; frequencies of HLA-DR, Ki-67, IL-7R expression on bulk CD4 and CD8 T-cells; HLA-DR and IL-7R expression on CM CD4 T-cells; Ki-67 expression on CM TREG; frequencies of TREG TEMRA, TREG EM, suppressive TREG, non-suppressive cytokine secreting TREG, CD25−CD127−CD4 T-cells; absolute count of TREG/μl; as well as IL-10 production of TREG and CD4 T-cells upon PMA/Ionomycin stimulation. A data set of n = 26 vertically HIV-infected, ART-naïve children above 5 years of age for whom all measurements were available from the same study visit was used for analysis. Variables are grouped on the basis of principal components analysis using the R package correlogram.
Results
Pediatric Slow Progressors Display Low General Immune Activation and Proliferation
We defined pediatric slow progressors (PSP) as ART-naïve children aged >5years whose absolute CD4 count is >450 cells/mm3. To examine the T-cell correlates of low immune activation observed among PSP, we divided subjects into four study groups: (i) Pediatric slow progressors, who maintained CD4 counts at >450/mm3 throughout the study period and remained ART-naïve (PSPN; n = 12); (ii) Pediatric future progressors (PFP; n = 11): children who met the criteria to be categorized as PSP at baseline, but who later experienced CD4 decline to <350 cells/mm3 and commenced ART (iii) Pediatric progressors (PP; n = 10), defined as being ART-naïve with CD4 <350/mm3 who commenced treatment after the blood draw; (iv) Age-matched HIV-uninfected children (n = 20). The absolute CD4 counts, CD4 percentage, CD4:CD8 ratios and viral loads are shown in Figures 1A–D and clinical features summarized in Table 1. As previously reported (7), CD4 T-cell activation, measured by HLA-DR expression, was lower in both PSPN and PFP compared to PP and correlated with disease progression by CD4% (Figures 1E,F). CD4 T-cell proliferation, measured by Ki-67, also correlated very strongly with CD4%, but unlike HLA-DR, was significantly higher on PSPN and PFP compared to healthy controls (Figures 1G,H). The pattern of HLA-DR expression is similar on CD8 T-cells, but Ki-67 expression does not distinguish between progressors non-progressors (Figures 1I–L). These data in a subset of the pediatric cohorts previously described (7) confirm the previous findings showing low immune activation and, additionally, low proliferative activity on CD4 T-cells of PSP, both of which are strongly linked to CD4 count in ART-naïve children.
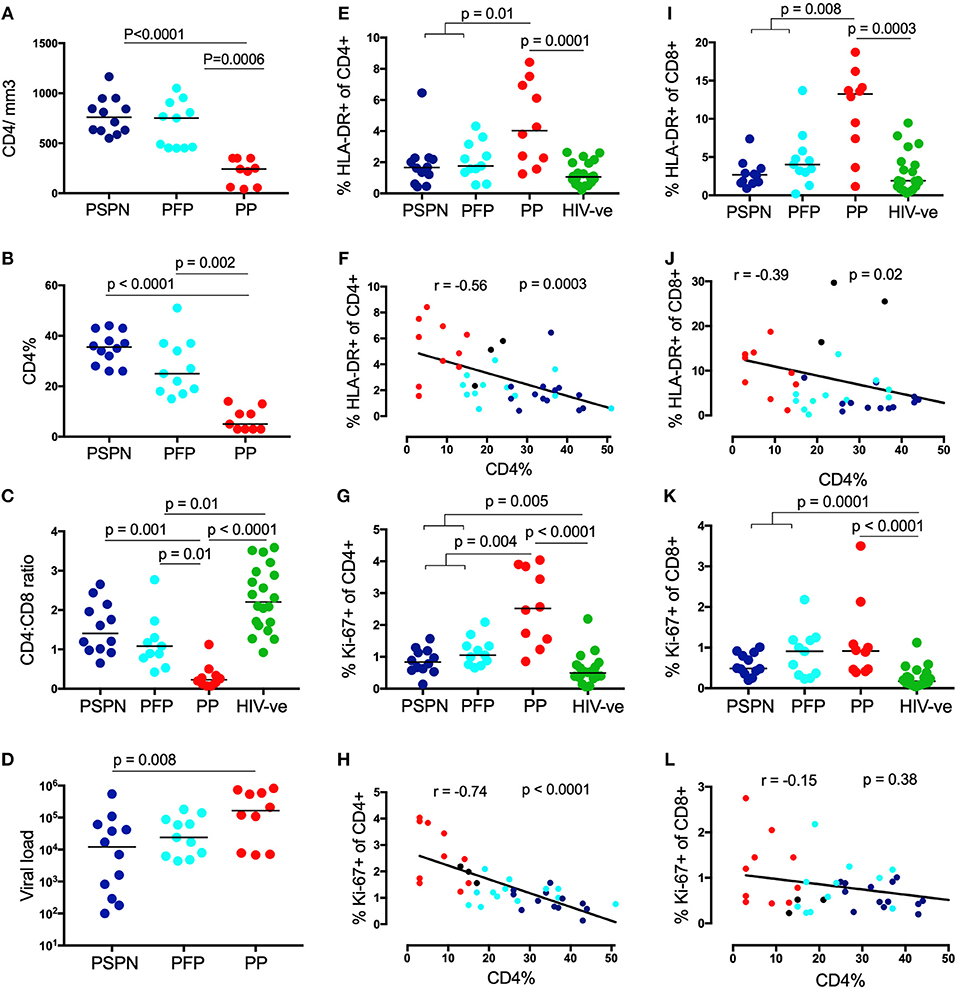
Figure 1. Pediatric slow progressors show low immune activation preferentially on CD4 T-cells. CD4 count absolute (A), CD4 percentage (B), viral load (C) in three pediatric groups: Pediatric slow progressors (PSP; n = 23) were defined here as treatment naïve, aged >5 years with CD4 counts >450/mm3 at the time point the experiments were performed. According to the longitudinal follow up we separated “pediatric future progressors” (PFP; n = 11; light blue) who would go onto treatment from true pediatric slow progressors, who stayed treatment naïve during the follow-up (PSPN; n = 12; dark blue). Pediatric progressors (PP; n = 10; red) were defined as treatment naïve, aged >5 years with CD4 counts <350/mm3. For CD4:CD8 ratio (D) age-matched uninfected controls (n = 20; green) were included as control group. (E) Frequency of HLA expression and its correlation with CD4 percentage (F) on CD4 T-cells within the 4 pediatric groups. (G,H), same as (E,F) but for Ki-67 expression. (I–L) same as (E–H) but of CD8 T-cells. For scatterplots, median, and interquartile range are shown. Kruskal-Wallis test was performed and corrected for multiple comparisons. Only significant p-values (<0.05) are given. For correlations, Spearman ranks test was used. For correlations with CD4 percentage, all HIV-infected infants were used including those not fulfilling criteria for PSP nor PP (black dots; n = 3; CD4 count >350 and <450/mm3).
Suppressive Regulatory T-Cells Are Increased in Pediatric Slow Progressors
To determine whether TREG play a role in controlling immune activation in these children, we studied CD4 T-cells co-expressing CD25 and the canonical transcription factor FoxP3 (39, 53) in the pediatric study groups described. The absolute number of TREG was significantly elevated in both PSPN and PFP (Figure 2A), and this correlated negatively with HLA-DR and Ki-67 expression on CD4 T-cells (Figures 2B,C), consistent with a role for TREG in preserving low immune activation in PSP. A similar trend was observed for TREG frequency, although statistically not significant (Supplementary Figure 2).
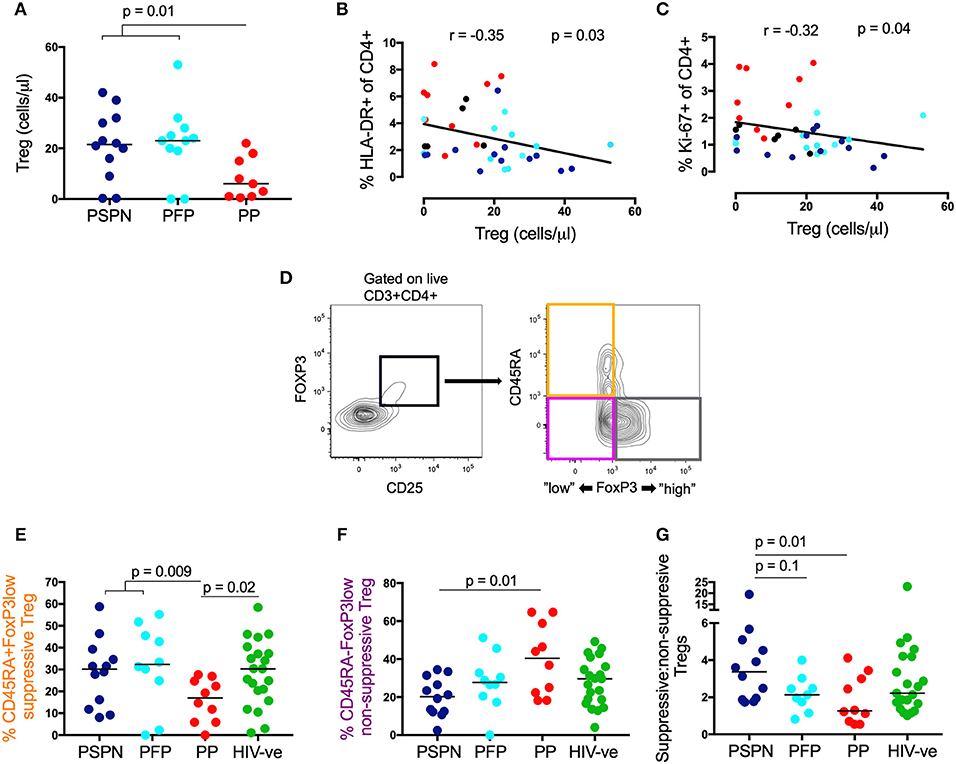
Figure 2. Suppressive regulatory T-cells are increased in pediatric slow progressors. (A) Absolute TREG count/μl (gated on CD25+FoxP3+ CD4 T-cells) within the different pediatric groups: PSPN (dark blue; n = 12), PFP (light blue; n = 11), PP (red; n = 10) and (B) its correlation with HLA-DR and Ki-67 (C) on CD4 T-cells. For correlations all HIV-infected children were used including those not fulfilling criteria for PSP nor PP (black dots; n = 3; CD4 count >350 and <450/mm3). (D) Gating strategy for three functional TREG (gated of CD25+FoxP3+ CD4 T-cells) memory subsets (39): (i) “activated”-suppressive TREG (CD45RA−FoxP3high; gray); (ii) “resting”-suppressive TREG (CD45RA+FoxP3low; orange) and cytokine-producing non-suppressive TREG (CD45RA−FoxP3low; pink). (E) Same groups as (A) but including uninfected controls (green; n = 20) and showing the frequency of “resting”- suppressive TREG (CD45RA+FoxP3low) and cytokine-producing non-suppressive TREG (CD45RA−FoxP3low; F). (G) Same as (E,F) but showing the ratio of all suppressive TREG (CD45RA+FoxP3low and CD45RA−FoxP3high; orange and gray population, respectively) to non-suppressive TREG (CD45RA−FoxP3low; pink population) within the pediatric groups. For scatterplots, median, and interquartile range are shown. Kruskal-Wallis test was performed and corrected for multiple comparisons. For correlations, Spearman ranks test was used.
To investigate whether these differences in TREG numbers between the pediatric study groups might correspond with functional differences, we examined TREG phenotype using expression levels of CD45RA and FoxP3 to identify three previously defined distinct functional TREG subsets: (1) CD45RA+ve FoxP3lo resting TREG (Figure 2D; orange population), (2) CD45RA-ve FoxP3hi activated TREG cells (Figure 2D; gray population), both of which are suppressive in vitro; and (3) CD45RA-ve FoxP3lo TREG that secrete cytokines but have no direct immunosuppressive activity (Figure 2D; pink population) (39). No significant differences between the study groups were observed in the frequency of CD45RA-ve FoxP3hi activated TREG (data not shown). However, CD45RA+veFoxP3lo resting TREG (a functionally suppressive subset) were significantly higher in PSP compared to PP (Figure 2E), whilst non-suppressive, CD45-ve FoxP3lo TREG, were highest in PP (Figure 2F). Together, the overall ratio of suppressive (combining the resting CD45+veFoxP3lo and the activated CD45RA-ve FoxP3hi subset): non-suppressive TREG is higher in PSPN compared to both PP and PFP (Figure 2G). Overall these data suggest TREG are both expanded and more functional in PSP compared to PP.
Memory TREG Proliferation and IL-10 Secretion Are Increased in Pediatric Slow Progressors
To further investigate TREG activity, we next measured proliferation and production of the key anti-inflammatory cytokine, IL-10 (27). Much higher levels of ex-vivo proliferation, by Ki-67, were observed in the central memory TREG subset of PSPN, compared to both PP and PFP (Figure 3A, see Supplementary Figure 3A for exemplary gating plots). Importantly, this was highly specific to central memory TREG, and not observed on central memory CD4 T-cells, on total TREG, or on total CD4s (Supplementary Figure 3B, Figure 1G). These data are consistent with the hypothesis that the expanded and phenotypically functional TREG observed in PSP, are maintained through active, on-going proliferation in these subjects, and that this activity is required to reduce immune activation and prevent CD4 T-cell decline.
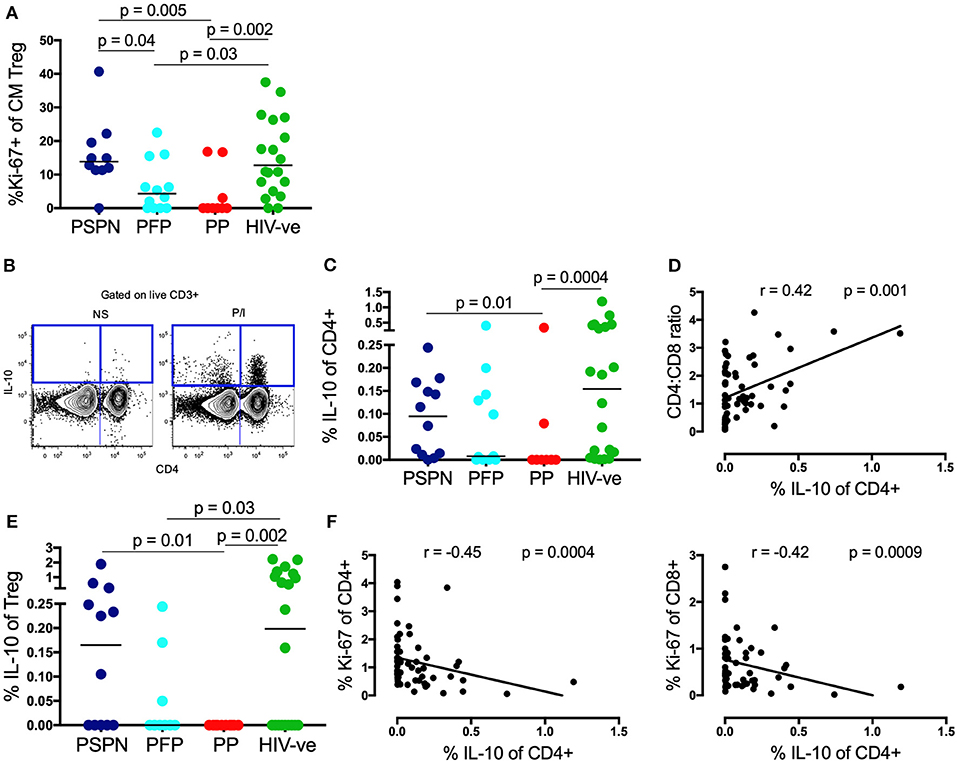
Figure 3. Memory TREG proliferation and IL-10 secretion are increased in pediatric slow progressors. (A) Frequency of Ki-67+ve “central memory” TREG (gated on CD4+CD25+FoxP3+CD45RA−CCR7+) within the pediatric groups: PSPN (dark blue; n = 12), PFP (light blue; n = 11), PP (red; n = 10), and uninfected controls (green; n = 20). (B) Exemplary FACS plot of one PSPN study subject showing CD4 (x-axis) vs. IL-10 (y-axis) staining (left: unstimulated control, right: stimulated with PMA/Ionomycin). (C) Same groups as (A) but showing IL-10 production of CD4 T-cells as measured by intracellular staining assay after 5 h stimulation with PMA/ Ionomycin and (D) its correlation with CD4:CD8 ratio within the whole pediatric cohort. (E) Same as (C) but of TREG. (F) Same as (D) but correlating IL-10 production of CD4 T-cells with Ki-67 expression on CD4 (left) and CD8 (right) T-cells. For correlations, all data available from pediatric subjects was used, including those not fitting criteria for PSP nor PP. For scatterplots, median, and interquartile range are shown. For correlations, Spearman ranks test was used. Kruskal-Wallis test was performed and corrected for multiple comparisons.
Evidence of this on-going immune regulation in PSP was also observed when we measured IL-10 production (Figure 3B). In response to non-specific stimulation with PMA/ionomycin, IL-10 production was severely impaired in PP, but preserved in PSPN (p = 0.01, Figure 3C). Again, PFP display an intermediate phenotype, and overall IL-10 production correlated with immune health, as measured by CD4:CD8 ratio (Figure 3D). As previously reported (54) IL-10 production by TREG themselves was low, but remained elevated in PSPN compared to PP (Figure 3E). Overall, total IL-10 production by CD4 T-cells within all pediatric study subjects correlated inversely with Ki-67 expression in both bulk CD4 and CD8 T-cells (Figure 3F), consistent with a direct role for this cytokine in limiting immune activation. Taken together, these data suggest that the maintenance of CD4 T-cells in PSP is an active process, impairment of which, as in PFP, may precipitate disease progression.
Increased IL-7/ sIL-7 Receptor in Pediatric Slow Progressors
Having established a role for immune regulation in PSP, we next examined the importance of homeostatic IL-7 signaling in these individuals. CD4 T-cell depletion in adult HIV infection is associated with a loss of the IL-7 receptor, CD127, whilst both non-progressing adults, and SIV-infected sooty mangabeys maintain high CD127 expression (40, 55, 56). Consistent with these data, we find CD127 is reduced on CD4 T-cells from PP compared to uninfected controls (p = 0.001) but preserved in both PSP groups (p = 0.01; Figure 4A). This is primarily a result of reduced expression on central memory T-cells (Figure 4B) and not in other memory subsets (Supplementary Figure 4). Interestingly, CD127 expression on CD8 T-cells is significantly lower in all HIV-infected individuals compared to healthy controls, irrespective of disease progression (Figure 4C). Furthermore, with respect to homeostatic T-cell proliferation mediated by IL-2 signaling via the high affinity IL-2 receptor CD25, we observed that CD25 CD127 double negative CD4 T-cells are expanded in PP compared to healthy controls, but not in PSP (Figure 4D). This finding, among HIV-infected children, is also consistent with the association in adults described between an expansion of CD4 T-cells lacking both CD25 and CD127 and progressive disease (55).
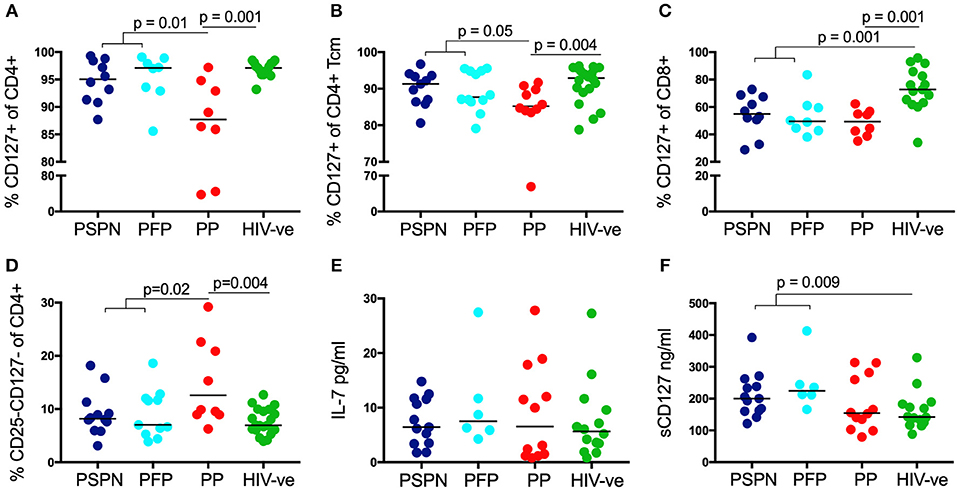
Figure 4. Increased IL-7/sIL-7 receptor in pediatric slow progressors. Frequency of IL-7R (CD127) expression on CD4 (A) and on central memory CD4 (B) T-cells in PSPN (dark blue; n = 10), PFP (light blue; n = 8), PP (red; n = 8), and uninfected pediatric controls (green; n = 15). (C) Same groups as before but showing frequency of IL-7R expression on CD8 T-cells and (D) frequency of CD4 T-cells double-negative for IL-2R (CD25) and IL-7R (CD127). (E) Ex-vivo plasma levels of IL-7 in pg/ml in PSPN (n = 13, dark blue), PFP (n = 6, light blue), PP (n = 12; red), and pediatric uninfected controls (n = 14; green). (F) Same as in (E) but showing plasma levels of IL-7R (sCD127) in ng/ml in the different groups. For scatterplots, median, and interquartile range are shown. Kruskal-Wallis test was performed and corrected for multiple comparisons.
As CD4 T-cells are a main consumer of IL-7 in vivo, loss of these cells in progressive adult HIV infection has been linked with an increase in plasma levels of IL-7 (43). Thus, IL-7 itself is considered unlikely to be a limiting resource in HIV infection. However, recent data demonstrate that the bio-activity of IL-7 is directly enhanced by the presence of soluble CD127 (sCD127) (57). Indeed, the positive benefit of IL-7 therapy in SIV infected rhesus macaques was dependent on high levels of sCD127 (58). We therefore measured plasma levels of IL-7 and sCD127 in HIV-infected children. As expected, plasma IL-7 was not limiting in any of our designated groups, and indeed no significant differences were observed (Figure 4E). However, sCD127 levels were markedly higher in PSP (median 212 ng/ml) compared to both PP (154 ng/ml), and uninfected controls (142 ng/ml; p = 0.009). As reported elsewhere, plasma level of sIL-7R are far greater than that of IL7 itself, being in the 100–400 ng/ml range compared to 1–30 pg/ml for IL-7 [Figure 4F; (57, 59)]. Importantly, the fact they have significantly higher levels of sCD127 than healthy controls, again suggest that this is an active process in PSP, rather than a bystander effect of non-progression. Together with preserved expression of the surface receptor CD127, IL-7 signaling in PSP is likely to be enhanced by high levels of sIL-7R in plasma, to potentiate the homeostatic activity of IL-7, and preserve CD4 counts.
Preservation of IL-7Ra and IL-2Ra Expression on CD4 T-Cells and TREG Count Are Independent From Immune Activation and Strong Associates for Slow Progression in Pediatric HIV Infection
To determine the independent contribution of the multiple, often correlated parameters (see correlogram, Supplementary Figure 5) described to the maintenance of normal-for-age CD4 T-cell counts, we assessed the influence of each using a generalized linear model (GLM), applying the least absolute shrinkage and selection operator (LASSO) principle on scaled covariates as described before (Table 2) (7). In agreement with this previous study, a primary driver of slow progression in this current analysis is low immune activation on CD4 T-cells. However, Ki-67 expression on CD4 T-cells, which was not included in the previous analysis, supersedes HLA-DR expression on CM CD4 T-cells, previously identified as the strongest predictor of CD4 count. In addition to these markers of immune activation, the LASSO model, also identifies IL-7R (CD127) and IL-2R (CD25) expression on CD4 T-cells, IL7-R of CD8 T-cells, as well as absolute TREG count and phenotype, as making independent contributions on CD4 count. Taken together, these data strongly support an important role for an active TREG response and T-cell homeostatic signaling in maintaining normal-for-age CD4 counts in PSP.
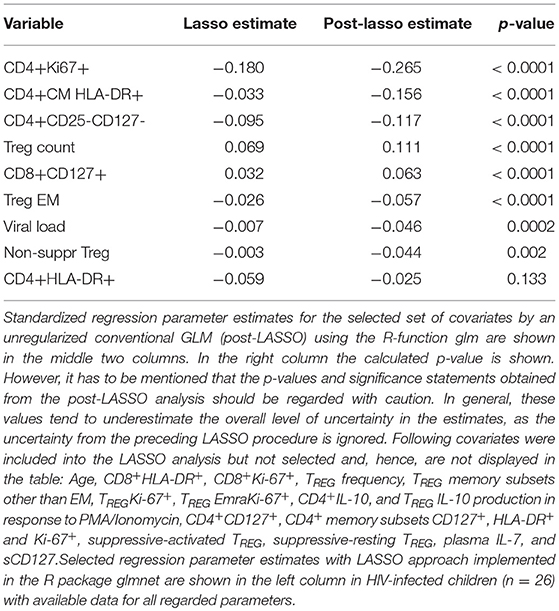
Table 2. Association of immunological parameters with CD4 T-cell count.
Discussion
In this study, we investigate the underlying mechanisms of CD4 preservation in a well-defined cohort of HIV-infected pediatric slow progressors (PSP). These children share fundamental characteristics with the natural hosts of non-pathogenic SIV infection, most importantly the preservation of circulating CD4 T-cells, and the lack of immune activation, despite on-going viral replication. Here we find strong evidence that the maintenance of this non-progressive state is an active process, requiring both immune regulation via TREG activity and IL-10 production, and enhancement of homeostatic IL-7 signaling. Notably, we also observe differences in phenotype and TREG activity between PSP who remain stable through the course of study (PSPN) and those who go onto progress at a later time-point (PFP). This suggests that TREG activity is particularly important in preventing progression, and that loss of this key function will precipitate development of disease. Moreover, this observation strengthens the argument that active involvement of TREG is required to maintain non-progression in PSP, rather being a consequence of a preserved CD4 T-cell compartment.
The role of regulatory T-cells in HIV infection has been controversial, with some studies suggesting they are detrimental as they inhibit antiviral T-cell responses (60–62), whilst others find a beneficial role through the reduction of immune activation (63–66) [reviewed in Phetsouphanh et al. (67)]. However, in general, preservation of TREG has been associated with positive outcomes; including non-progression in HIV elite controllers (68, 69), adult viremic slow progressors (70) and natural hosts of SIV infection (71). Conversely, a loss of circulating TREG in adult progressors inversely correlates with the activation and apoptosis of CD8 T-cells (72), and experimental depletion in SIV-infected natural hosts increases immune activation and viral replication (73). The importance of measuring absolute counts has been emphasized previously (14, 35), in studies finding that the relative frequency of TREG was increased in all HIV-infected individuals, compared to controls, but that the absolute number was greatly decreased, with the exception of elite controllers. Taken together, these studies strongly suggest TREG can play a beneficial role in limiting HIV disease progression.
Our hypothesis that activation of TREG in PSP is an important indicator of their functional involvement in immune regulation is supported by previous work showing adult HIV-infected elite controllers (EC) and viremic long term non-progressors (LTNPs) have more activated TREG than healthy controls (74). Moreover, similar to our PSP, these EC have preserved IL-10 production in contrast to the controls. IL-10 is an anti-inflammatory cytokine that is critically important in preventing inflammatory and autoimmune diseases and is produced by multiple cells of the adaptive immune system including CD4 T-cells and TREG as well as by innate cells [reviewed in Kwon and Kaufmann (54)]. Not all studies have shown a positive effect of IL-10 in adult HIV infection (75, 76) due to its negative impact on HIV-specific T-cell proliferation and effector function. It is possible that IL-10 has different effects in adult and pediatric HIV infection, as cytotoxic T-cells are not thought to be important in PSP (77). However, other studies have shown a correlation between IL-10 production, low immune activation, and slow disease progression in adult HIV infection (78) and genetic polymorphisms in the IL-10 promoter that are associated with higher IL-10 production have been shown to attenuate CD4 T-cell loss in HIV-infected individuals (79–81).
In addition to immune regulation, a key observation of our study is the apparent preservation and augmentation of IL-7 signaling in PSP. Firstly, and in contrast to progressing children, IL-7 receptor expression (CD127) is preserved in PSP, especially within the CD4 T-cell compartment. This correlates with parameters of disease progression and is in accordance to various studies of adult and pediatric HIV infection (50, 82–86). Notably, differences in CD127 expression are most prominent within the central memory compartment, which has recently been shown to be particularly dependent on IL-7 signaling (87). Similar preservation of CD127 expression in the central memory compartment has been described in adult elite controllers and LTNP (50, 88). In these studies, CD127 expression was directly linked to viral load, which cannot be true in the case of PSP, who are all viremic, again highlighting the likely differences in mechanism underlying HIV non-progression in adults and children.
Second, and most unexpectedly, we found PSP had increased plasma levels of sCD127 compared to PP and healthy controls. The precise details of how sCD127 influences IL-7 signaling and T-cell homeostasis are unclear. Progressive adult HIV infection was linked to increased plasma levels of sCD127, hypothesized to inhibit IL-7 availability (59). However, lower levels of sCD127 have been observed by others in HIV-infected patients compared to healthy controls (89). Moreover, higher levels of sCD127 in plasma were associated with improved immune reconstitution in SIV infected rhesus macaques treated with recombinant IL-7 (58). The potentiating effect of sCD127 on IL-7 signaling is supported by mechanistic studies showing it enhances the bioactivity of IL-7 when the cytokine is limiting, as it is presumed to be the situation in-vivo (57, 59). Therefore, we hypothesize that increased plasma sCD127 leads to enhanced bioactivity of IL-7 in PSP and, along with preserved surface CD127 expression, improves maintenance of CD4 T-cells through IL-7 homeostatic signaling. The fact that sCD127 levels are higher in PSP than healthy controls, suggests this is an important mechanism in preserving CD4 T-cell levels in these rare PSP individuals.
One potential limitation of this study is the use of surface CD25 as a marker of TREG, as this molecule is also an activation marker, whose expression may be affected by HIV-driven immune activation (90, 91). However, we have previously examined CD25 in HIV-infected and uninfected children and did not observe a significant impact of HIV on overall CD25 expression levels on CD4 T-cells (92). Furthermore, as the PSP group displayed very low immune activation, it is unlikely to have caused elevated CD25 expression within this group. An additional limitation is the lack of in vitro data to directly determine TREG activity (39). Unfortunately, we did not have sufficient samples remaining from this historical cohort to perform such assays. However, the fact that we have identified multiple independent measures of enhanced TREG activity in PSP, including increased TREG frequency and proliferation, TREG skewing toward more suppressive phenotypes, and enhanced IL-10 production, strongly supports the existence of enhanced TREG suppressive capacity in the unique subset of PSP.
In this study, we observed various mechanisms contributing to CD4 T-cell preservation shared between PSP and the natural hosts of SIV infection. Firstly, as observed in natural hosts (8, 10, 16), PSP had low T-cell immune activation and proliferation as measured by HLA-DR and Ki-67 expression despite viremia and this correlated inversely with parameters of disease progression. Secondly, PSP showed increased absolute TREG numbers and this was strongly inversely associated with immune activation (18). Thirdly, both CD4 T-cells and TREG of PSP secreted more IL-10 than those of progressors which fits with the observation that in African green monkeys, but not in rhesus macaques, SIV infection leads to expression of an anti-inflammatory profile including IL-10, and TGF-β secretion and an increase in TREG (17). And lastly, both PSP and natural hosts have a low frequency of CD4 T-cells lacking both IL-7R and IL-2R (CD127 and CD25) (55). These CD127-veCD25-ve CD4 T-cells show features of activated effectors and could therefore be key promoters of the overall level of immune activation and disease progression (55). LASSO modeling suggests that, although some of these parameters maybe linked through immune activation, both TREG activity and T-cell homeostasis are independently involved in maintaining CD4 counts in PSP.
Over all, our findings support a model where PSP preserve their CD4 T-cells through actively maintaining low immune activation, requiring both immune regulation via TREG activity and IL-10 production, and enhancement of T-cell homeostatic IL-7 signaling.
Author Contributions
JR designed the study, conducted experimental work within the study, analyzed the data, and wrote the paper. AN, MM, and EA conducted experimental work within the study. AG performed statistical analyses within the study. TN supervised experimental work within the study. HK, PG, and AL supervised experimental work within the study, analyzed data, and wrote the paper. PG established research cohorts.
Funding
AL: WT and HHMI; PG: WT (Grant 104748MA) and NIH (RO1 AI133673); HK: WT (Grant 202485/Z/16/Z).
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We thank all children and their care givers for their time and commitment to this study. We also thank all staff at the Ithembalabantu Clinic in Umlazi, Durban, for their essential support and all core staff members at the HIV Pathogenesis Programme who helped with patient recruitment and sample processing.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fimmu.2019.00213/full#supplementary-material
Supplementary Figure 1. (A) LASSO coefficient paths vs. tuning parameter λ and (B) deviance of the model via 5-fold cross-validation.
Supplementary Figure 2. Frequency of TREG (%CD25+FoxP3+ of CD4 T-cells) in PSPN (dark blue; n = 12), PFP (light blue; n = 11), PP (red; n = 10) and uninfected pediatric controls (green; n = 20).
Supplementary Figure 3. (A) Exemplary gating strategy for Ki-67 expression of “central memory” TREG (CD4+CD25+FoxP3+CD45RA−CCR7+) in one PSPN (blue; top row) and one PP child (red; bottom row). (B) Frequency of Ki-67 expression on all TREG (gated on CD4+CD25+FoxP3+; left), of all central memory CD4 T-cells (CD45RA-CCR7+; CM; middle) and of Ki-67 expression on all CM T-cells (CD45RA−CCR7+; right) within the different pediatric groups: PSPN (dark blue; n = 12), PFP (light blue; n = 11), PP (red; n = 10) and uninfected pediatric controls (green; n = 20). Kruskal-Wallis test was performed and corrected for multiple comparisons.
Supplementary Figure 4. Frequency of IL-7R expression on naïve (TN; left), effector memory (EM; middle) and terminally differentiated (Temra; right) CD4 T-cells in PSPN (dark blue; n = 12), PFP (light blue; n = 11), PP (red; n = 10) and uninfected pediatric controls (green; n = 20). For scatterplots, median and interquartile range are shown. Kruskal-Wallis test was performed and corrected for multiple comparisons.
Supplementary Figure 5. Correlation matrix in ART-naïve HIV-infected children (n = 25) with available data for all regarded parameters. Positive correlations are colored in blue and inverse correlations in red with deeper color shading and bigger circle size reflecting stronger r-values. Correlations with p > 0.05 are left blank.
Supplementary Table 1. List of antibodies for flow cytometry.
References
1. Goulder PJ, Lewin SR, Leitman EM. Paediatric HIV infection: the potential for cure. Nat Rev Immunol. (2016) 16:259–71. doi: 10.1038/nri.2016.19
2. Blanche S, Newell ML, Mayaux MJ, Dunn DT, Teglas JP, Rouzioux C, et al. Morbidity and mortality in European children vertically infected by HIV-1. The french pediatric HIV infection study group and european collaborative study. J Acquir Immune Defic Syndr Hum Retrovirol. (1997) 14:442–50
3. Paul ME, Mao C, Charurat M, Serchuck L, Foca M, Hayani K, et al. Predictors of immunologic long-term nonprogression in HIV-infected children: implications for initiating therapy. J Allergy Clin Immunol. (2005) 115:848–55. doi: 10.1016/j.jaci.2004.11.054
4. Ssewanyana I, Elrefaei M, Dorsey G, Ruel T, Jones NG, Gasasira A, et al. Profile of T cell immune responses in HIV-infected children from Uganda. J Infect Dis. (2007) 196:1667–70. doi: 10.1086/522013
5. Ananworanich J, Apornpong T, Kosalaraksa P, Jaimulwong T, Hansudewechakul R, Pancharoen C, et al. Characteristics of lymphocyte subsets in HIV-infected, long-term nonprogressor, and healthy Asian children through 12 years of age. J Allergy Clin Immunol. (2010) 126:1294–1301 e1210. doi: 10.1016/j.jaci.2010.09.038
6. Hainaut M, Verscheure V, Ducarme M, Schandene L, Levy J, Mascart F. Cellular immune responses in human immunodeficiency virus (HIV)-1-infected children: is immune restoration by highly active anti-retroviral therapy comparable to non-progression? Clin Exp Immunol. (2011) 165:77–84. doi: 10.1111/j.1365-2249.2011.04403.x
7. Muenchhoff M, Adland E, Karimanzira O, Crowther C, Pace M, Csala A, et al. Nonprogressing HIV-infected children share fundamental immunological features of nonpathogenic SIV infection. Sci Transl Med. (2016) 8:358ra125. doi: 10.1126/scitranslmed.aag1048
8. Silvestri G, Sodora DL, Koup RA, Paiardini M, O'Neil SP, McClure HM, et al. Nonpathogenic SIV infection of sooty mangabeys is characterized by limited bystander immunopathology despite chronic high-level viremia. Immunity (2003) 18:441–52. doi: 10.1016/S1074-7613(03)00060-8
9. Brenchley JM, Paiardini M, Knox KS, Asher AI, Cervasi B, Asher TE, et al. Differential Th17 CD4 T-cell depletion in pathogenic and nonpathogenic lentiviral infections. Blood (2008) 112:2826–35. doi: 10.1182/blood-2008-05-159301
10. Chahroudi A, Bosinger SE, Vanderford TH, Paiardini M, Silvestri G. Natural SIV hosts: showing AIDS the door. Science (2012) 335:1188–93. doi: 10.1126/science.1217550
11. Giorgi JV, Hultin LE, McKeating JA, Johnson TD, Owens B, Jacobson LP, et al. Shorter survival in advanced human immunodeficiency virus type 1 infection is more closely associated with T lymphocyte activation than with plasma virus burden or virus chemokine coreceptor usage. J Infect Dis. (1999) 179:859–70. doi: 10.1086/314660
12. Deeks SG, Kitchen CM, Liu L, Guo H, Gascon R, Narvaez AB, et al. Immune activation set point during early HIV infection predicts subsequent CD4+ T-cell changes independent of viral load. Blood (2004) 104:942–7. doi: 10.1182/blood-2003-09-3333
13. Roider JM, Muenchhoff M, Goulder PJ. Immune activation and paediatric HIV-1 disease outcome. Curr Opin HIV AIDS (2016) 11:146–55. doi: 10.1097/COH.0000000000000231
14. Prendergast A, O'Callaghan M, Menson E, Hamadache D, Walters S, Klein N, et al. Factors influencing T cell activation and programmed death 1 expression in HIV-infected children. AIDS Res Hum Retroviruses (2012) 28:465–8. doi: 10.1089/AID.2011.0113
15. Brenchley JM, Paiardini M. Immunodeficiency lentiviral infections in natural and non-natural hosts. Blood (2011) 118:847–54. doi: 10.1182/blood-2010-12-325936
16. Favre D, Lederer S, Kanwar B, Ma ZM, Proll S, Kasakow Z, et al. Critical loss of the balance between Th17 and T regulatory cell populations in pathogenic SIV infection. PLoS Pathog. (2009) 5:e1000295. doi: 10.1371/journal.ppat.1000295
17. Kornfeld C, Ploquin MJ, Pandrea I, Faye A, Onanga R, Apetrei C, et al. Antiinflammatory profiles during primary SIV infection in African green monkeys are associated with protection against AIDS. J Clin Invest. (2005) 115:1082–91. doi: 10.1172/JCI23006
18. Pereira LE, Villinger F, Onlamoon N, Bryan P, Cardona A, Pattanapanysat K, et al. Simian immunodeficiency virus (SIV) infection influences the level and function of regulatory T cells in SIV-infected rhesus macaques but not SIV-infected sooty mangabeys. J Virol. (2007) 81:4445–56. doi: 10.1128/JVI.00026-07
19. Boettler T, Spangenberg HC, Neumann-Haefelin C, Panther E, Urbani S, Ferrari C, et al. T cells with a CD4+CD25+ regulatory phenotype suppress in vitro proliferation of virus-specific CD8+ T cells during chronic hepatitis C virus infection. J Virol. (2005) 79:7860–7. doi: 10.1128/JVI.79.12.7860-7867.2005
20. Rouse BT, Sarangi PP, Suvas S. Regulatory T cells in virus infections. Immunol Rev. (2006) 212:272–86. doi: 10.1111/j.0105-2896.2006.00412.x
22. Huang CT, Workman CJ, Flies D, Pan X, Marson AL, Zhou G, et al. Role of LAG-3 in regulatory T cells. Immunity (2004) 21:503–13. doi: 10.1016/j.immuni.2004.08.010
23. Wing K, Onishi Y, Prieto-Martin P, Yamaguchi T, Miyara M, Fehervari Z, et al. CTLA-4 control over Foxp3+ regulatory T cell function. Science (2008) 322:271–5. doi: 10.1126/science.1160062
24. Borsellino G, Kleinewietfeld M, Di Mitri D, Sternjak A, Diamantini A, Giometto R, et al. Expression of ectonucleotidase CD39 by Foxp3+ Treg cells: hydrolysis of extracellular ATP and immune suppression. Blood (2007) 110:1225–32. doi: 10.1182/blood-2006-12-064527
25. Duhen T, Duhen R, Lanzavecchia A, Sallusto F, Campbell DJ. Functionally distinct subsets of human FOXP3+ Treg cells that phenotypically mirror effector Th cells. Blood (2012) 119:4430–40. doi: 10.1182/blood-2011-11-392324
26. Kindlund B, Sjoling A, Yakkala C, Adamsson J, Janzon A, Hansson LE, et al. CD4(+) regulatory T cells in gastric cancer mucosa are proliferating and express high levels of IL-10 but little TGF-beta. Gastr Cancer (2017) 20:116–25. doi: 10.1007/s10120-015-0591-z
27. Yamaguchi T, Wing JB, Sakaguchi S. Two modes of immune suppression by Foxp3(+) regulatory T cells under inflammatory or non-inflammatory conditions. Semin Immunol. (2011) 23:424–30. doi: 10.1016/j.smim.2011.10.002
28. Prendergast AJ, Klenerman P, Goulder PJ. The impact of differential antiviral immunity in children and adults. Nat Rev Immunol. (2012b) 12:636–48. doi: 10.1038/nri3277
29. Ndhlovu LC, Loo CP, Spotts G, Nixon DF, Hecht FM. FOXP3 expressing CD127lo CD4+ T cells inversely correlate with CD38+ CD8+ T cell activation levels in primary HIV-1 infection. J Leukoc Biol. (2008) 83:254–62. doi: 10.1189/jlb.0507281
30. Cao W, Jamieson BD, Hultin LE, Hultin PM, Detels R. Regulatory T cell expansion and immune activation during untreated HIV type 1 infection are associated with disease progression. AIDS Res Hum Retroviruses (2009) 25:183–91. doi: 10.1089/aid.2008.0140
31. Freguja R, Gianesin K, Mosconi I, Zanchetta M, Carmona F, Rampon O, et al. Regulatory T cells and chronic immune activation in human immunodeficiency virus 1 (HIV-1)-infected children. Clin Exp Immunol. (2011) 164:373–80. doi: 10.1111/j.1365-2249.2011.04383.x
32. Mendez-Lagares G, Pozo-Balado MM, Genebat M, Garcia Perganeda A, Leal M, Pacheco YM. Severe immune dysregulation affects CD4(+)CD25(hi)FoxP3(+) regulatory T cells in HIV-infected patients with low-level CD4 T-cell repopulation despite suppressive highly active antiretroviral therapy. J Infect Dis. (2012) 205:1501–9. doi: 10.1093/infdis/jis230
33. Degaffe G, Zakhour R, Zhang W, Contreras GA, Bell CS, Rodriguez G, et al. Forkhead box protein 3(+) regulatory T cells and Helios(+) subset in perinatally acquired HIV. Clin Exp Immunol. (2015) 180:108–17. doi: 10.1111/cei.12560
34. Khaitan A, Kravietz A, Mwamzuka M, Marshed F, Ilmet T, Said S, et al. FOXP3+Helios+ regulatory T cells, immune activation, and advancing disease in HIV-infected children. J Acquir Immune Defic Syndr. (2016) 72:474–84. doi: 10.1097/QAI.0000000000001000
35. Schulze Zur Wiesch J, Thomssen A, Hartjen P, Toth I, Lehmann C, Meyer-Olson D, et al. Comprehensive analysis of frequency and phenotype of T regulatory cells in HIV infection: CD39 expression of FoxP3+ T regulatory cells correlates with progressive disease. J Virol. (2011) 85:1287–97. doi: 10.1128/JVI.01758-10
36. Shaw JM, Hunt PW, Critchfield JW, McConnell DH, Garcia JC, Pollard RB, et al. Increased frequency of regulatory T cells accompanies increased immune activation in rectal mucosae of HIV-positive noncontrollers. J Virol. (2011) 85:11422–34. doi: 10.1128/JVI.05608-11
37. Angin M, Kwon DS, Streeck H, Wen F, King M, Rezai A, et al. Preserved function of regulatory T cells in chronic HIV-1 infection despite decreased numbers in blood and tissue. J Infect Dis. (2012) 205:1495–500. doi: 10.1093/infdis/jis236
38. Arguello RJ, Balbaryski J, Barboni G, Candi M, Gaddi E, Laucella S. Altered frequency and phenotype of CD4+ forkhead box protein 3+ T cells and its association with autoantibody production in human immunodeficiency virus-infected paediatric patients. Clin Exp Immunol. (2012) 168:224–33. doi: 10.1111/j.1365-2249.2012.04569.x
39. Miyara M, Yoshioka Y, Kitoh A, Shima T, Wing K, Niwa A, et al. Functional delineation and differentiation dynamics of human CD4+ T cells expressing the FoxP3 transcription factor. Immunity (2009) 30:899–911. doi: 10.1016/j.immuni.2009.03.019
40. Sumpter B, Dunham R, Gordon S, Engram J, Hennessy M, Kinter A, et al. Correlates of preserved CD4(+) T cell homeostasis during natural, nonpathogenic simian immunodeficiency virus infection of sooty mangabeys: implications for AIDS pathogenesis. J Immunol. (2007) 178:1680–91. doi: 10.4049/jimmunol.178.3.1680
41. Schluns KS, Kieper WC, Jameson SC, Lefrancois L. Interleukin-7 mediates the homeostasis of naive and memory CD8 T cells in vivo. Nat Immunol. (2000) 1:426–32. doi: 10.1038/80868
42. Kaech SM, Tan JT, Wherry EJ, Konieczny BT, Surh CD, Ahmed R. Selective expression of the interleukin 7 receptor identifies effector CD8 T cells that give rise to long-lived memory cells. Nat Immunol. (2003) 4:1191–8. doi: 10.1038/ni1009
43. Rethi B, Fluur C, Atlas A, Krzyzowska M, Mowafi F, Grutzmeier S, et al. Loss of IL-7Ralpha is associated with CD4 T-cell depletion, high interleukin-7 levels and CD28 down-regulation in HIV infected patients. AIDS (2005) 19:2077–86. doi: 10.1097/01.aids.0000189848.75699.0f
44. Sprent J, Surh CD. Interleukin 7, maestro of the immune system. Semin Immunol. (2012) 24:149–50. doi: 10.1016/j.smim.2012.04.011
45. Pellegrini M, Calzascia T, Toe JG, Preston SP, Lin AE, Elford AR, et al. IL-7 engages multiple mechanisms to overcome chronic viral infection and limit organ pathology. Cell (2011) 144:601–13. doi: 10.1016/j.cell.2011.01.011
46. Shive CL, Clagett B, McCausland MR, Mudd JC, Funderburg NT, Freeman ML, et al. Inflammation perturbs the IL-7 axis, promoting senescence and exhaustion that broadly characterize immune failure in treated HIV infection. J Acquir Immune Defic Syndr. (2016) 71:483–92. doi: 10.1097/QAI.0000000000000913
47. Hartling HJ, Jespersen S, Gaardbo JC, Sambleben C, Thorsteinsson K, Gerstoft J, et al. Reduced IL-7R T cell expression and increased plasma sCD127 in late presenting HIV-infected individuals. J Acquir Immune Defic Syndr. (2017) 74:81–90. doi: 10.1097/QAI.0000000000001153
48. Alfonzo MA, Diaz A, Siciliano L, Lopez MG, Hung A, Garcia JF. Functional state of CD4+ and CD8+ T lymphocytes and their role in the slow progression of HIV infection in pediatric patients. J Pediatr. (2012) 88:161–8. doi: 10.2223/JPED.2183
49. Nguyen TP, Shukla S, Asaad R, Freeman ML, Lederman MM, Harding CV, et al. Responsiveness to IL-7 but not to IFN-alpha is diminished in CD4+ T cells from treated HIV infected patients who experience poor CD4+ T-cell recovery. AIDS (2016) 30:2033–42. doi: 10.1097/QAD.0000000000001161
50. Mercier F, Boulassel MR, Yassine-Diab B, Tremblay C, Bernard NF, Sekaly RP, et al. Persistent human immunodeficiency virus-1 antigenaemia affects the expression of interleukin-7Ralpha on central and effector memory CD4+ and CD8+ T cell subsets. Clin Exp Immunol. (2008) 152:72–80. doi: 10.1111/j.1365-2249.2008.03610.x
51. Tibshirani R. Regression shrinkage and selection via the lasso. J R Stat Soc Ser B (1996) 58, 267–88
52. Friedman J, Hastie T, Tibshirani R. Regularization paths for generalized linear models via coordinate descent. J Stat Softw. (2010) 33:1–22
53. Sakaguchi S, Yamaguchi T, Nomura T, Ono M. Regulatory T cells and immune tolerance. Cell (2008) 133:775–87. doi: 10.1016/j.cell.2008.05.009
54. Kwon DS, Kaufmann DE. Protective and detrimental roles of IL-10 in HIV pathogenesis. Eur Cytokine Netw. (2010) 21:208–14. doi: 10.1684/ecn.2010.0201
55. Dunham RM, Cervasi B, Brenchley JM, Albrecht H, Weintrob A, Sumpter B, et al. CD127 and CD25 expression defines CD4+ T cell subsets that are differentially depleted during HIV infection. J Immunol. (2008) 180:5582–92
56. Gaardbo JC, Hartling HJ, Gerstoft J, Nielsen SD. Thirty years with HIV infection-nonprogression is still puzzling: lessons to be learned from controllers and long-term nonprogressors. AIDS Res Treat. (2012) 2012:161584. doi: 10.1155/2012/161584
57. Lundstrom W, Highfill S, Walsh ST, Beq S, Morse E, Kockum I, et al. Soluble IL7Ralpha potentiates IL-7 bioactivity and promotes autoimmunity. Proc Natl Acad Sci USA. (2013) 110:E1761–70. doi: 10.1073/pnas.1222303110
58. Steele AK, Carrasco-Medina L, Sodora DL, Crawley AM. Increased soluble IL-7 receptor concentrations associate with improved IL-7 therapy outcomes in SIV-infected ART-treated Rhesus macaques. PLoS ONE (2017) 12:e0188427. doi: 10.1371/journal.pone.0188427
59. Crawley AM, Faucher S, Angel JB. Soluble IL-7R alpha (sCD127) inhibits IL-7 activity and is increased in HIV infection. J Immunol. (2010) 184:4679–87. doi: 10.4049/jimmunol.0903758
60. Aandahl EM, Michaelsson J, Moretto WJ, Hecht FM, Nixon DF. Human CD4+ CD25+ regulatory T cells control T-cell responses to human immunodeficiency virus and cytomegalovirus antigens. J Virol. (2004) 78:2454–9. doi: 10.1128/JVI.78.5.2454-2459.2004
61. Weiss L, Donkova-Petrini V, Caccavelli L, Balbo M, Carbonneil C, Levy Y. Human immunodeficiency virus-driven expansion of CD4+CD25+ regulatory T cells, which suppress HIV-specific CD4 T-cell responses in HIV-infected patients. Blood (2004) 104:3249–56. doi: 10.1182/blood-2004-01-0365
62. Kinter A, McNally J, Riggin L, Jackson R, Roby G, Fauci AS. Suppression of HIV-specific T cell activity by lymph node CD25+ regulatory T cells from HIV-infected individuals. Proc Natl Acad Sci USA. (2007) 104:3390–5. doi: 10.1073/pnas.0611423104
63. Eggena MP, Barugahare B, Jones N, Okello M, Mutalya S, Kityo C, et al. Depletion of regulatory T cells in HIV infection is associated with immune activation. J Immunol. (2005) 174:4407–14. doi: 10.4049/jimmunol.174.7.4407
64. Fazekas de St Groth B, Landay AL. Regulatory T cells in HIV infection: pathogenic or protective participants in the immune response? AIDS (2008) 22:671–83. doi: 10.1097/QAD.0b013e3282f466da
65. Moreno-Fernandez ME, Rueda CM, Rusie LK, Chougnet CA. Regulatory T cells control HIV replication in activated T cells through a cAMP-dependent mechanism. Blood (2011) 117:5372–80. doi: 10.1182/blood-2010-12-323162
66. Okeke EB, Okwor I, Uzonna JE. Regulatory T cells restrain CD4+ T cells from causing unregulated immune activation and hypersensitivity to lipopolysaccharide challenge. J Immunol. (2014) 193:655–62. doi: 10.4049/jimmunol.1303064
67. Phetsouphanh C, Xu Y, Zaunders J. CD4 T cells mediate both positive and negative regulation of the immune response to HIV infection: complex role of T follicular helper cells and regulatory T cells in pathogenesis. Front Immunol. (2014) 5:681. doi: 10.3389/fimmu.2014.00681
68. Chase AJ, Yang HC, Zhang H, Blankson JN, Siliciano RF. Preservation of FoxP3+ regulatory T cells in the peripheral blood of human immunodeficiency virus type 1-infected elite suppressors correlates with low CD4+ T-cell activation. J Virol. (2008) 82:8307–15. doi: 10.1128/JVI.00520-08
69. Owen RE, Heitman JW, Hirschkorn DF, Lanteri MC, Biswas HH, Martin JN, et al. HIV+ elite controllers have low HIV-specific T-cell activation yet maintain strong, polyfunctional T-cell responses. AIDS (2010) 24:1095–105. doi: 10.1097/QAD.0b013e3283377a1e
70. Choudhary SK, Vrisekoop N, Jansen CA, Otto SA, Schuitemaker H, Miedema F, et al. Low immune activation despite high levels of pathogenic human immunodeficiency virus type 1 results in long-term asymptomatic disease. J Virol. (2007) 81:8838–42. doi: 10.1128/JVI.02663-06
71. Chan ML, Petravic J, Ortiz AM, Engram J, Paiardini M, Cromer D, et al. Limited CD4+ T cell proliferation leads to preservation of CD4+ T cell counts in SIV-infected sooty mangabeys. Proc Biol Sci. (2010) 277:3773–81. doi: 10.1098/rspb.2010.0972
72. Jiao Y, Fu J, Xing S, Fu B, Zhang Z, Shi M, et al. The decrease of regulatory T cells correlates with excessive activation and apoptosis of CD8+ T cells in HIV-1-infected typical progressors, but not in long-term non-progressors. Immunology (2009) 128(1 Suppl.):e366–75. doi: 10.1111/j.1365-2567.2008.02978.x
73. Pandrea I, Gaufin T, Brenchley JM, Gautam R, Monjure C, Gautam A, et al. Cutting edge: experimentally induced immune activation in natural hosts of simian immunodeficiency virus induces significant increases in viral replication and CD4+ T cell depletion. J Immunol. (2008) 181:6687–91. doi: 10.4049/jimmunol.181.10.6687
74. Gaardbo JC, Ronit A, Hartling HJ, Gjerdrum LM, Springborg K, Ralfkiaer E, et al. Immunoregulatory T cells may be involved in preserving CD4 T cell counts in HIV-infected long-term nonprogressors and controllers. J Acquir Immune Defic Syndr. (2014) 65:10–8. doi: 10.1097/QAI.0b013e3182a7c932
75. Clerici M, Balotta C, Salvaggio A, Riva C, Trabattoni D, Papagno L, et al. Human immunodeficiency virus (HIV) phenotype and interleukin-2/ interleukin-10 ratio are associated markers of protection and progression in HIV infection. Blood (1996) 88:574–9
76. Brockman MA, Kwon DS, Tighe DP, Pavlik DF, Rosato PC, Sela J, et al. IL-10 is up-regulated in multiple cell types during viremic HIV infection and reversibly inhibits virus-specific T cells. Blood (2009) 114:346–56. doi: 10.1182/blood-2008-12-191296
77. Adland E, Paioni P, Thobakgale C, Laker L, Mori L, Muenchhoff M, et al. Discordant impact of HLA on viral replicative capacity and disease progression in pediatric and adult HIV infection. PLoS Pathog. (2015) 11:e1004954. doi: 10.1371/journal.ppat.1004954
78. Chevalier MF, Didier C, Petitjean G, Karmochkine M, Girard PM, Barre-Sinoussi F, et al. Phenotype alterations in regulatory T-cell subsets in primary HIV infection and identification of Tr1-like cells as the main interleukin 10-producing CD4+ T cells. J Infect Dis. (2015) 211:769–79. doi: 10.1093/infdis/jiu549
79. Shin HD, Winkler C, Stephens JC, Bream J, Young H, Goedert JJ, et al. Genetic restriction of HIV-1 pathogenesis to AIDS by promoter alleles of IL10. Proc Natl Acad Sci USA. (2000) 97:14467–72. doi: 10.1073/pnas.97.26.14467
80. Erikstrup C, Kallestrup P, Zinyama-Gutsire RB, Gomo E, Butterworth AE, Pedersen BK, et al. Reduced mortality and CD4 cell loss among carriers of the interleukin-10−1082G allele in a Zimbabwean cohort of HIV-1-infected adults. AIDS (2007) 21:2283–91. doi: 10.1097/QAD.0b013e3282f153ed
81. Naicker DD, Werner L, Kormuth E, Passmore JA, Mlisana K, Karim SA, et al. Interleukin-10 promoter polymorphisms influence HIV-1 susceptibility and primary HIV-1 pathogenesis. J Infect Dis. (2009) 200:448–52. doi: 10.1086/600072
82. Koesters SA, Alimonti JB, Wachihi C, Matu L, Anzala O, Kimani J, et al. IL-7Ralpha expression on CD4+ T lymphocytes decreases with HIV disease progression and inversely correlates with immune activation. Eur J Immunol. (2006) 36:336–44. doi: 10.1002/eji.200535111
83. Benito JM, Lopez M, Lozano S, Gonzalez-Lahoz J, Soriano V. Down-regulation of interleukin-7 receptor (CD127) in HIV infection is associated with T cell activation and is a main factor influencing restoration of CD4(+) cells after antiretroviral therapy. J Infect Dis. (2008) 198:1466–73. doi: 10.1086/592716
84. Kiazyk SA, Fowke KR. Loss of CD127 expression links immune activation and CD4(+) T cell loss in HIV infection. Trends Microbiol. (2008) 16:567–73. doi: 10.1016/j.tim.2008.08.011
85. Hasley RB, Hong C, Li W, Friesen T, Nakamura Y, Kim GY, et al. HIV immune activation drives increased Eomes expression in memory CD8 T cells in association with transcriptional downregulation of CD127. AIDS (2013) 27:1867–77. doi: 10.1097/QAD.0b013e3283618487
86. Tanaskovic S, Fernandez S, Price P, French MA. Interleukin-7 signalling defects in naive CD4+ T cells of HIV patients with CD4+ T-cell deficiency on antiretroviral therapy are associated with T-cell activation and senescence. AIDS (2014) 28:821–30. doi: 10.1097/QAD.0000000000000213
87. Okoye AA, Rohankhedkar M, Konfe AL, Abana CO, Reyes MD, Clock JA, et al. Effect of IL-7 therapy on naive and memory T cell homeostasis in aged rhesus macaques. J Immunol. (2015) 195:4292–305. doi: 10.4049/jimmunol.1500609
88. Potter SJ, Lacabaratz C, Lambotte O, Perez-Patrigeon S, Vingert B, Sinet M, et al. Preserved central memory and activated effector memory CD4+ T-cell subsets in human immunodeficiency virus controllers: an ANRS EP36 study. J Virol. (2007) 81:13904–15. doi: 10.1128/JVI.01401-07
89. Rose T, Lambotte O, Pallier C, Delfraissy JF, Colle JH. Identification and biochemical characterization of human plasma soluble IL-7R: lower concentrations in HIV-1-infected patients. J Immunol. (2009) 182:7389–97. doi: 10.4049/jimmunol.0900190
90. Biancotto A, Iglehart SJ, Vanpouille C, Condack CE, Lisco A, Ruecker E, et al. HIV-1 induced activation of CD4+ T cells creates new targets for HIV-1 infection in human lymphoid tissue ex vivo. Blood (2008) 111:699–704. doi: 10.1182/blood-2007-05-088435
91. Angerami MT, Suarez GV, Vecchione MB, Laufer N, Ameri D, Ben G, et al. Expansion of CD25-negative forkhead box P3-positive T cells during HIV and Mycobacterium tuberculosis infection. Front Immunol. (2017) 8:528. doi: 10.3389/fimmu.2017.00528
Keywords: pediatric HIV-infection, regulatory T cells (Treg), immune activation (IA), IL-7, homeostatic signaling, pediatric slow progression, IL-10, immune regulation
Citation: Roider J, Ngoepe A, Muenchhoff M, Adland E, Groll A, Ndung'u T, Kløverpris H, Goulder P and Leslie A (2019) Increased Regulatory T-Cell Activity and Enhanced T-Cell Homeostatic Signaling in Slow Progressing HIV-infected Children. Front. Immunol. 10:213. doi: 10.3389/fimmu.2019.00213
Received: 10 October 2018; Accepted: 24 January 2019;
Published: 12 February 2019.
Edited by:
Claire Anne Chougnet, Cincinnati Children's Research Foundation, United StatesReviewed by:
Keith R. Fowke, University of Manitoba, CanadaAlka Khaitan, New York University, United States
Copyright © 2019 Roider, Ngoepe, Muenchhoff, Adland, Groll, Ndung'u, Kløverpris, Goulder and Leslie. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Philip Goulder, cGhpbGlwLmdvdWxkZXJAcGFlZGlhdHJpY3Mub3guYWMudWs=
Alasdair Leslie, YWwubGVzbGllQGFocmkub3Jn
†These authors have contributed equally to this work