 Yun Shan Goh
Yun Shan Goh Daniel McGuire
Daniel McGuire Laurent Rénia
Laurent Rénia- 1Singapore Immunology Network (SIgN), Agency for Science, Technology and Research (A*STAR), Biopolis, Singapore, Singapore
- 2School of Biological Sciences, Nanyang Technological University, Singapore, Singapore
- 3Department of Microbiology and Immunology, Yong Loo Lin School of Medicine, National University of Singapore, Singapore, Singapore
Despite continuous efforts, the century-old goal of eradicating malaria still remains. Multiple control interventions need to be in place simultaneously to achieve this goal. In addition to effective control measures, drug therapies and insecticides, vaccines are critical to reduce mortality and morbidity. Hence, there are numerous studies investigating various malaria vaccine candidates. Most of the malaria vaccine candidates are subunit vaccines. However, they have shown limited efficacy in Phase II and III studies. To date, only whole parasite formulations have been shown to induce sterile immunity in human. In this article, we review and discuss the recent developments in vaccination with sporozoites and the mechanisms of protection involved.
Introduction
Malaria is one of the deadliest diseases, causing a major public health problem with high mortality and morbidity. In 2017, the World Health Organization reported 219 million clinical cases and 435,000 deaths (1). The use of different control interventions such as insecticide-treated bed nets, combination drug therapies and early diagnostics has greatly reduced malaria mortality worldwide (2). However, with increasing drug resistance and insecticide resistance, these efforts are insufficient to eradicate malaria globally (3, 4). It has become increasingly clear that there is no control intervention that can singly eradicate malaria. Multiple control interventions need to be in place simultaneously and a malaria vaccine is integral to global malaria eradication (5).
Parasite Life Cycle
Plasmodium parasites have a complex life cycle, infecting two hosts, the human and the mosquito. In the human host, the Plasmodium life cycle consists of two stages, the liver stage and the blood stage. Infected female Anopheles mosquitoes inject sporozoites into the dermis of their mammalian host upon feeding. Sporozoites then enter the bloodstream and migrate to the liver, where the liver stage begins. The sporozoites multiply in hepatocytes, eventually forming merozoites in vesicles called merosomes. These vesicles rupture and release the merozoites into the bloodstream, commencing the blood stage by infecting red blood cells (RBCs). It is the continual cycling of malaria parasites within the RBCs, and the immune responses directed against this stage of the parasite, that causes most of the pathologies observed in malaria infections. The malaria parasites are then transmitted back to the mosquito following blood feeding by a female mosquito. The sexual forms of the blood stage parasites, gametocytes, develop into male and female gametes which fertilize each other, eventually forming oocysts in the mosquito's midgut wall. The oocysts then lyse to release sporozoites, which migrate to the mosquito's salivary glands. When the Anopheles mosquito takes a blood meal on another human, the injected sporozoites migrate from the dermis to the liver, thereby beginning a new cycle of infection.
Vaccines Against Malaria
The development of vaccines for malaria has been met with many difficulties. Despite decades of research efforts, there is still no available vaccine for human use. This has led to the development of a wide range of approaches, in the search for an efficacious malaria vaccine. These approaches can be broadly divided into three main categories: (1) whole parasite-based vaccines, (2) subunit vaccines, and (3) viral, bacterial and parasite vectors as delivery vectors.
Whole Parasite-Based Vaccines
Whole parasite-based vaccines have had considerably more success than other vaccines. Whole parasite-based vaccines contain all parasitic antigens. This approach allows the development of different types of immune responses. Whole parasites used for the vaccines are obtained by dissecting sporozoites from mosquitoes or harvesting asexual blood stages from culture. There are many technical, logistical, and regulatory hurdles associated with large scale production and delivery of whole parasite vaccines in the field. However, recent sporozoite vaccine trials have shown considerable progress in overcoming these hurdles (6, 7).
History
The development of malaria vaccines began with whole parasite-based vaccines more than a 100 years ago when the Sergent brothers used heat-inactivated P. relictum sporozoites to immunize canaries and obtained partial protection (8). This was followed by the work of Russell and Mohan where both cellular and humoral responses against malaria were induced in immunized domestic fowls (9). In 1946, Jules Freund invented the Freund adjuvant and formulated the vaccine by combining the adjuvant with formalin-inactivated-blood infected with P. lophurae, an avian malaria parasite, or P. knowlesi, a monkey malaria parasite (10, 11). The formulations showed promising efficacy. However, the toxic side effects of the Freund adjuvant have prevented its use in humans. The first attempt in humans was done by Heidelberger et al., using formalin-inactivated P. vivax-infected blood to immunize volunteers, however no protection was induced (12). These initial studies, though suboptimal, have paved the way for future whole parasite-based vaccine development.
Vaccination With Sporozoites
Among the whole parasite-based vaccine candidates, there is considerable research on the pre-erythrocytic parasites. The pre-erythrocytic stage is an asymptomatic phase. Very few sporozoites are injected and subsequently developed in the hepatocytes during natural infection in human volunteers (13). The idea of inducing an immune response that can neutralize sporozoites in the skin and circulation and prevent the penetration of a low number of sporozoites into hepatocytes or destroy a low number of infected hepatocytes during the asymptomatic phase make pre-erythrocytic stage vaccines attractive. By preventing the pre-erythrocytic stage development, the vaccines would prevent blood stage infection, hence preventing pathology. In addition, the pre-erythrocytic stage vaccines have had more success than the other stages, which provides more support for their development.
Whole sporozoite-based vaccines developed thus far in human include (1) irradiated parasites (6, 14), (2) genetically-attenuated parasites (15, 16), and (3) drug-infection-treatment vaccination (17, 18).
Vaccination with irradiated sporozoites
Irradiated sporozoite vaccine is the most clinically developed whole parasite-based vaccine, and also the most clinically developed pre-erythrocytic vaccine (17). The first few studies that showed definitive protective immunity with irradiated sporozoites were done in chicks and subsequently in mice (18, 19). This was later demonstrated in humans, where vaccination with irradiated sporozoites, via infective mosquito bites, protected 92% of the volunteers from infection (20–22). However, >1,000 mosquito bites are required to introduce sufficient irradiated sporozoites to induce the high level of efficacy. This prevented the development of this approach for mass vaccination.
More recently, delivery of cyropreserved irradiated sporozoites into the host by direct venous inoculation via needle and syringe, has been tested in humans and showed promising efficacy data (6, 17, 23). While four doses only protected 33% of the individuals (6), five doses protected 100% of the individuals (6). More studies to perfect the vaccination regimes would allow direct venous inoculation via needle and syringe to replace mosquito bites as a delivery system. Another hurdle with irradiated sporozoite vaccines is the need for a high dose of irradiated parasites. Vaccine dosage, vaccination regimen, and route of administration have been investigated in malaria-naive adults (24). In the study, administration of higher doses may further enhance protection— four intravenous immunizations with a higher dose of 2.7 × 105 irradiated sporozoites was found to be the most optimal, where 55% of vaccinated subjects remained uninfected following controlled human malaria infections (CHMI) 21 weeks after immunization. The timing of the CHMI following vaccination has also been found to be important, with vaccine efficacy being higher when CHMI was performed 3 weeks after immunization, instead of 21 weeks. While vaccination with irradiated sporozoites led to sterile protection in 100% (6/6) of vaccinated malaria naïve volunteers (6), irradiated sporozoites vaccination in malaria-endemic Mali yielded a lower protection (14). There are fundamental differences between the two studies, such as the first study examines protection against homologous challenge and the latter study examines protection against heterologous challenge. Naturally transmitted parasites are often different from the vaccine strain. Twenty-four weeks after final immunization regimen, five doses of 2.7 × 105 irradiated sporozoites protected 7 of 10 against homologous CHMI, but only 1 of 10 against heterologous CHMI (25), showing that the vaccine efficacy against heterologous infection is markedly reduced. In addition, the findings also suggest that pre-exposure to the malaria parasites may prevent the establishment of a protective immune response since blood stage infection is able to induce immune suppression (26). This also further highlights the need for optimization of the required dose and regime in the target population to achieve robust and sterile protection.
The irradiation of parasites is a delicate process that requires the sporozoites to retain a certain degree of viability. Similar to heat-inactivated and frozen-thawed sporozoites, over-irradiated sporozoites do not induce protection (27, 28). Irradiating the sporozoites leads to DNA damage in the sporozoites with no or limited reduction on hepatocyte infectivity (29, 30). Although irradiation results in an inhibition of parasite DNA replication, ultrastructure modification and alteration in gene expression (31, 32), eventually leading to developmental arrest of the liver stage within hepatocytes (33, 34), it still allows for parasite antigen presentation and priming of immune responses.
Vaccination with genetically-attenuated sporozoites
Non-irradiated sporozoites have been postulated to be more efficacious as whole parasite-based vaccines because, as compared to irradiated sporozoites, they are able to progress to a later stage of pre-erythrocytic development, (35). By doing so, the host immune system is exposed to a wide repertoire of malarial antigens and thus able to target more of the pre-erythrocytic stage. To this end, research efforts have focused on developing ways, other than irradiation, to attenuate the growth of the parasite. Recent advances in Plasmodium research such as genetic manipulation have brought forward a new approach to attenuate parasites. Genetically-attenuated parasites are modified by deleting key essential genes that result in developmental arrest of the liver stage after hepatocyte infection, but do not affect parasite viability, mosquito infectivity, and sporozoite production in animal models (36). Inactivation of UIS3, UIS4, or P36p prevented the attenuated parasites from developing beyond the early pre-erythrocytic stage in mice (37–39). Vaccination of these live genetically-attenuated parasites offered sterile protection against a challenge with a wild type isolate. While promising, one of the major concerns for the development of genetically-attenuated parasites as vaccines is the possible occurrence of breakthrough infections. The first clinical trial using live genetically-attenuated parasites that lack the two genes, p52 and p36, led to breakthrough infections (15). Breakthrough infections in mice have also been observed for UIS4- and P36p-deficient parasites (37, 39). However, great progress has been made recently. Live genetically-attenuated parasites lacking three genes (p52–/p36–/sap1–; “PfGAP3KO”) arrest early in liver-stage development, and were safe with no observed breakthrough infection following administration into human subjects by infective mosquito bites (16). Compared to irradiated sporozoite vaccine and early liver stage-arresting genetically-attenuated parasites, second generation genetically-attenuated parasites that arrest late liver stages have shown to demonstrate superior anti-malarial immunity following vaccination in mice by having a greater antigen repertoire (40, 41). These candidate vaccines could also have greater efficacy in humans, but this remains to be demonstrated.
Drug-infection-treatment vaccination
Drug-infection-treatment vaccination is the last approach. It involves vaccination with live wild-type parasites under drug prophylaxis, where the drug targets and eliminates the blood stage parasites. This approach allows full liver development and a limited initial blood stage development, thereby focusing immunity toward the liver stages. Pioneer mouse studies have, indeed, shown greater efficacy when the mice were vaccinated with live P. berghei or P. yoelii sporozoites under drug prophylaxis than when the mice were vaccinated with irradiated sporozoites—fewer inoculations and less sporozoites were required to induce sterile protection (42–44). Vaccination with live sporozoites under chloroquine prophylaxis is the most investigated vaccine formulation under this approach, and it has shown very promising efficacy data. The first study in humans examined the efficacy of the vaccine where the live sporozoites were introduced into the volunteers via infective mosquito bites (45). The study utilized CHMI and demonstrated sterile protection, where 100% of the volunteers were protected from infection following a wild-type sporozoite challenge. A subsequent CHMI study investigated the efficacy of the vaccine where the live sporozoites were intravenously inoculated via a needle and syringe. In this study, a dose-dependent protection was observed, where only three doses of 5.12 × 104 sporozoites were sufficient to protect all volunteers from the challenge (46). In addition to chloroquine, vaccination with live wild-type parasites under prophylaxis of other antimalarials has also been investigated. Primaquine (47), mefloquine (48) and artemisinin derivatives such as artesunate (49) have been used in place of chloroquine and vaccination with live wild-type parasites under prophylaxis of these antimalarials has demonstrated protective immunity against a homologous sporozoite challenge in mice and in humans.
However, there are various challenges with this approach such as the technical and logistical issues associated with generating sporozoites at large scale and field deployment. There are also concerns that the sporozoite injections might not be properly followed with antimalarial treatment, which could lead to sickness. In addition, while sterile immunity can be achieved against a homologous sporozoite challenge, protection was suboptimal when immunized volunteers were challenged with a heterologous strain (50, 51), suggesting that the protective immune mechanisms target polymorphic antigens. Nevertheless, the current published findings have demonstrated very encouraging efficacy data and suggest that vaccination with live sporozoites under chloroquine prophylaxis, following vaccination regime optimization, could potentially be the most efficacious sporozoite vaccine until date.
Subunit Vaccines
Subunit vaccines allow rational design of the vaccine to induce the desired immune effectors against the parasite. In addition, subunit vaccines are safe and generally easy to produce at large scale and to administer in the field. Hence, efficacious subunit vaccines that offer long term protection are the preferred vaccines of choice.
Peptides and Recombinant Proteins
Subunit vaccines have been developed either as peptides, multi-peptide constructs or recombinant proteins. They can be based on a single parasite antigen or a combination of multiple parasite antigens, and often in a formulation that includes adjuvants. Most constructs have been designed for the circumsporozoite protein (CSP), a major sporozoite surface protein (52), as it is the first cloned malaria antigen (53, 54). Both peptides and multi-peptide constructs containing either the B epitope alone or both B and T epitopes induced protection in mouse models (55–58). However, when tested in humans, the peptide constructs did not induce significant protection (59). The main reason for the failure in humans is that the immune response to these constructs was genetically restricted by major histocompatibility gene (60) and thus could not induce an efficient immune response in most volunteers.
To ameliorate immunogenicity and protective efficacy, peptides and proteins have been designed to contain T cell epitopes from the parasites or an unrelated proteins recognized by many MHC genes (61), coupled to diverse chemical backbones, or fused with other proteins to create particle vaccines to be used with or without various adjuvants (62–67). All these new constructs demonstrated a high efficacy in mouse models but have yet to be validated in humans.
Of all the subunit vaccines, RTS,S, a CSP-based subunit vaccine, is the current most clinically-advanced malaria vaccine, being the only malaria vaccine to have progressed to the pivotal Phase III clinical trials (68). Enormous resources have been spent on rationally improving RTS,S efficacy, which include developing novel adjuvant systems. Multiple studies have demonstrated a need for RTS,S be formulated with adjuvants such as monophosphoryl lipid A and QS21, to achieve immunogenicity (69–71). This was followed up by a series of clinical trials, where significant progress has been made to improve immunogenicity and efficacy (72, 73). The RTS,S vaccine has been designed to inhibit the liver stage and prevent blood stage infection. RTS,S is, ultimately, formulated with a chimeric molecule based on CSP, fused to the S antigen of the hepatitis B virus, together with a potent adjuvant, AS01. The first Phase IIb trials performed in adult volunteers in the USA showed ~50% protection against clinical malaria (69, 70, 74).
When tested in the endemic regions, RTS,S's efficacy against infection was less impressive and was of short-duration (<3 months) (75, 76). What is encouraging is that RTS,S/AS01 vaccination has been found to induce a significant reduction (~60%) in the incidence of clinical infections in children in the same study (76). This finding encouraged GlaxoSmithKline and the Malaria Vaccine initiative, with financial support from the Bill and Melinda Gates Foundation, to further develop this vaccine for infants and young children in Africa. However, RTS,S efficacy against clinical malaria was later found to be suboptimal in malaria-endemic populations, with a vaccine efficacy against clinical infection of 36.3% in young children and 25.9% in infants (77–81). One possible explanation is that the CSP used in the vaccine contains several T cell epitopes, which are all highly polymorphic in parasite population in the field. Neafsey et al. elegantly demonstrated that the overall vaccine efficacy was very low in field settings with minimal matching of the CSP alleles in the field with the CSP allele in RTS,S (80).
Another explanation for the limited efficacy of the CSP-based vaccines is that it may not be the best antigen to induce protection. It is likely that other antigens may be better vaccine candidates, alone or in combination with the CSP. This hypothesis was supported by various studies, which showed that the sterile protection against a sporozoite challenge obtained in mice immunized with irradiated sporozoites was independent of the immune response against the CSP (82–85).
With the limited success of subunit vaccines developed thus far, there have been many efforts to identify new pre-erythrocytic (liver) targets for vaccine development using various approaches (86–90). These new antigens have shown encouraging efficacy data in animal models either alone (91, 92) or in combinations (93, 94), however efficacy in humans has yet to be demonstrated (95).
DNA Vaccines
DNA has been identified as a vaccine delivery system in the 1990s (96). This approach was quickly taken up, and DNA vaccines against CSP were developed and tested in human (97–99). Although the CSP-based constructs induced high levels of protection in mice (100), they had poor immunogenicity in humans (101). DNA constructs encoding multiple genes (102, 103) or epitopes were also developed (104). However, none of these constructs induced high level of protection against a sporozoite challenge (104, 105). To enhance the immune responses, malaria DNA vaccines were developed in combination with DNA constructs encoding for cytokines such as GM-CSF. Although some of these constructs had increased immunogenicity and efficacy in murine models (106), they did not induce protection against sporozoite challenge in human (105).
Viral, Bacterial, and Parasite Vectors as Delivery Vectors
Viral, bacterial and parasite vectors have been developed as delivery vectors for malaria vaccines. As these vectors are based on whole organisms, they usually do not need to be adjuvanted to stimulate the innate immune system which is necessary for the development of an optimal adaptive immune response (107). However, in some studies, various adjuvants have also been used to increase vector constructs immunogenicity (108).
The use of viral vectors as delivery vectors for malaria vaccines is the most common (109). Recombinant influenza viruses (110), pox viruses such as vaccinia virus (110, 111), Sindbis virus (112), yellow fever virus (113, 114), adenovirus (115), human cytomegalovirus (116) as delivery vectors have shown promising efficacy in animal models. Bacterial vectors such as Salmonella (117, 118), Bacille-Calmette Guerin (BCG) (119), Shigella flexneri 2A strain (120) as delivery vectors have also shown good immunogenicity and efficacy against sporozoite challenge in mice. Currently, only Salmonella vectors as delivery vectors have been examined in humans, showing good safety profile and immunogenicity (121). Parasites such as Leishmania (122) and Toxoplasma (123, 124) as delivery vectors were also able to induce partial protection in mice. However, it remains to be seen if these vectors can induce protection in human.
Prime-Boost Combinations
To enhance humoral and T cell responses, various prime-boost strategies have been developed using combinations of different vaccine approaches. Vaccination with subunits or DNA constructs, followed by viral vectors or combination of viral vectors encoding one or multiple malaria antigens has been examined and efficacy has been demonstrated in mouse models (125–129). However, of all these combinations, only a few have shown significant efficacy in humans (104, 109, 130–133). Recent strategies, prime-and-target (134) and prime-and-trap (135), have shown that the best combination, that can induce high level of protection in mice, depends on the capacity to induce and maintain tissue-resident memory cells in the liver.
Model Systems
The use of experimental models has an important role in the development of vaccines. It is essential for the first assessments of safety, immunogenicity and potential protective efficacy of vaccine candidates. The early studies of malaria candidate vaccines utilized avian models, despite being a poor alternative to study pathogens with mammalian hosts. The eventual establishment of other malaria models has brought new insights and greatly facilitated malaria vaccine research. Current models used include: (1) mouse, (2) non-human primates (NHP), (3) humanized mice, and (4) human volunteers (CHMI).
Mouse Models
Until date, the traditional mouse model still remains the most commonly used model as it is less costly and more easily available. P. berghei and P. yoelii are two of the more commonly used rodent malaria species for in vivo and in vitro studies. It is often the starting ground for in vivo studies examining the development of pre-erythrocytic stage and whole sporozoite vaccine candidates. Human parasites such as P. falciparum and P. vivax, which contribute to the majority of the malaria global disease burden, display highly restricted host-cell tropism—they cannot establish an infection and develop the pre-erythrocytic stages effectively in vitro easily. A reproducible model of full development of the pre-erythrocytic stage has only been described in primary human hepatocytes (136). The mouse model allows the examination of the parasite infection in the liver in vivo, and the effectiveness of the whole sporozoite vaccine candidates to protect the host from infection. While vaccine efficacy in the mice does not necessarily predict vaccine efficacy in humans, there have not been any examples where an absence of vaccine efficacy in mice was contradicted by vaccine efficacy in humans. Quite a number of whole sporozoite vaccine candidates that are first identified to be protective in mouse models (19, 42, 44) have went on to be validated in humans (22, 45). The first study demonstrating that vaccination with a whole sporozoite vaccine candidate, irradiated sporozoites, can induce sterile protection from infective sporozoite challenge was in a P. berghei mouse model (19). This has been established as the gold standard as human volunteers vaccinated with irradiated P. falciparum sporozoites were found to develop protective immunity (22). Similarly, vaccination with live sporozoites under drug prophylaxis was also first identified to have promising efficacy in the P. berghei and P. yoelii mouse models (42, 44), before demonstrating sterile protection in all vaccinated human volunteers (45). A new approach to chemically attenuate sporozoites has been identified using the P. berghei model. This was performed by treating sporozoites with centanamycin, a DNA alkylating agent (137–139). This may also offer protection in humans, however further studies to validate its efficacy are still pending.
With the development of transgenic rodent malaria parasites, knock-in (KI) parasites expressing P. falciparum (140) or P. vivax (141) genes have been generated. These KI parasites allow the examination of the efficacy of immunogens (142–144) or antibodies against human malaria pre-erythrocytic antigens in vivo (145, 146).
While the mouse models have greatly contributed to the development of malaria vaccines, there are limitations. It is still largely unknown how relevant the mouse models are for the human parasite. The ability to interpret and draw conclusion from the mouse and translate it to the human remains unclear. There are major fundamental differences, both at genetic and proteomic levels, between the rodent and human malaria species, with the rodent parasite genomes missing orthologs for more than 730 P. falciparum genes (147, 148). In addition, there are differences in the both the liver and blood stages of infection. While the mouse parasites, P. berghei and P. yoelii, emerge from the liver after 2–3 days of infection, the human parasites, P. falciparum and P. vivax, require 7–10 days of pre-erythrocytic stage development (149). The formation of dormant pre-erythrocytic stages in P. vivax infections, which is a hallmark of P. vivax infections (150), is also not present in P. berghei and P. yoelii infections in mice, although liver forms of P. yoelii have been observed in the liver of their natural host, Thamnomys gazellae, at least 8 months post-sporozoite infection (151). Lastly, most murine studies are performed with genetically homogenous inbred mice with a limited MHC gene repertoire, which do not mimic the large genetic diversity of the human population. Laboratory mice are maintained in clean specific-pathogen—free facilities, hence the absence of the effect of environment (e.g., microbiome) on the mouse immune system may bias infection and vaccine studies (152). Taken together, while it is a powerful experiment tool, the traditional mouse model is not an ideal model, especially for studying pre-erythrocytic malaria vaccines.
Non-human Primates
Due to the limitations of the traditional mouse models, there have been substantial efforts to develop alternative animal models that are able to generate adequate parallels in an in vivo approach of the human immune system. Historically, non-human primates (NHP) have been used as the alternative model. Compared to the mouse, NHP share a lot more similarities with the human. Simian Plasmodium species can infect various NHP species. In particular, human parasites can be adapted to NHP and some NHP can even support direct infection with P. falciparum and P. vivax (153). The Aotus monkeys have served as a valuable model. They can be infected by P. falciparum and P. vivax (154, 155). The NHP models are particularly important for assessments of P. vivax pre-erythrocytic stage vaccines (156), due to the formation of dormant pre-erythrocytic stages in P. vivax infections, which cannot be recapitulated in the mouse models. Simian malaria parasites, such as P knowlesi or P. cynomolgi in macaques, have also been used for immune and vaccine studies (157, 158). Vaccination with live sporozoites under chloroquine prophylaxis has shown promising efficacy data in Toque monkeys immunized with P. cynomolgi (159). However, the lack of availability, the high costs to maintain a colony and the restriction of utilization due to ethical issues limit the use of NHP, especially where large numbers are required.
Humanized Mouse
In the more recent years, the development of a humanized mouse as an animal model (160, 161) has greatly facilitated the study of human malaria research. These models mainly arise from the xenotransplantation of human hepatopoietic cells and/or tissues, allowing the long-term establishment of components of human immunity in permissive immunodeficient mice.
Using a human liver chimeric SCID/Alb-uPA mouse, studies on the pre-erythrocytic stages can be performed (162, 163). While it has been shown to be a viable model to study P. falciparum pre-erythrocytic stage development, the study of human malaria in this model is limited to the pre-erythrocytic stage as P. falciparum cannot transit from the pre-erythrocytic stages to the blood stages in this model (164). Other drawbacks include infertility of the mice due to the SCID/Alb-uPA immunodeficient background (165), and hepatotoxicity and high neonatal mortality due to the uPA transgene expression (166). These have made the generation of large number of the mice extremely costly and difficult.
Due to these drawbacks, an alternative, a human liver chimeric FAH−/−Rag2−/−IL2Rγnull (FRG) mouse, has been developed. These mice can be bred relatively easily and do not suffer from hepatotoxicity. In addition, this model has been shown to support robust pre-erythrocytic stage infection and development (149). When human RBCs were transplanted into these mice, the new model supported the transition from a pre-erythrocytic infection to a blood stage infection (167). The NOD mice deficient for the IL2Rγ gene and transgenic for the thymidine kinase gene (TK-NOG) is another model that has also been developed. These mice do not suffer from liver failure. A transient injection of the drug gancyclovir induces a controlled ablation of the mouse hepatocytes. Treated mice are easily repopulated with human hepatocytes (168). These mice can also be doubly engrafted with human red blood cells and this allows the full development of P. falciparum in the liver and the transition to the blood stage. Interestingly, this model also supported the liver stage development of another human parasite, P. ovale (169).These findings are extremely encouraging as this raises the possibility of using these models to study a liver stage infection and also a combined liver and blood stage infection. While the use of the humanized mice offers many new possibilities to study human malaria biology in a non-human model in vivo, it is worth noting that these mice are immuno-compromised, which makes them unsuitable for vaccine immunogenicity and efficacy studies. Nonetheless, they have shown to be useful in passive transfer experiments to test antibody efficacy (170–172).
Studies on the development of humanized mice with a fully reconstituted immune system are underway (173). In fact, humanized mice that possess the human immune system (HIS) have been established for malaria research, using recombinant adeno-associated virus (AAV)-based gene transfer technologies (174). With functional human CD4 T cells and B cells (HIS-CD4/B mice), these HIS mice were able to produce a significant level of human IgG against P. falciparum CSP upon immunization (175). The HIS-CD4/B mice were also protected against infection from an in vivo challenge with transgenic P. berghei sporozoites expressing the PfCSP protein following immunization. While these models are essential pre-clinical models to understand immune responses against human malaria, it is worth noting that these HIS mice still retain mouse myeloid compartments that are likely to influence antigen presentation and immune cell residency, and in vivo vaccine efficacy can only be examined using transgenic rodent malaria parasites expressing selected P. falciparum proteins. New iterations of humanized mice that possess the humanization of the liver, bone marrow, lymphoid compartments, and human erythrocytes would be the ideal mouse model and would greatly help to understand human malaria parasite infection and immunology. It would be an essential tool in providing a more accurate initial assessment of the safety profile and vaccine efficacy of malaria vaccine candidates before moving onto human studies.
Human Volunteers
The most relevant model is the human host itself. The establishment of the CHMI model has greatly helped malaria research. The CHMI model involves exposing healthy human volunteers to the parasite via infective mosquito bites, monitoring the volunteers closely for signs and symptoms of malaria infection, and treating the volunteers with drug upon detection of fever and/or detection of parasites (45, 176).
The CHMI model uses the most relevant host-parasite pair. While CHMI has been performed via other routes such as intravenously and intramuscularly, it is more commonly done via infective mosquito bites. The use of infective mosquito bites in the model mimics the natural route of infection, offering advantages in the prediction of the potential efficacy of vaccine candidate against natural infections. However, it also has its limitations. The CHMI model is often performed with one parasite strain, while there are many antigenically diverse heterologous parasites in the field. Infection in the CHMI model is controlled and the parasite load is administered at one single time, whereas high parasite load at one single time is uncommon in natural field setting. Despite the limitations, CHMI studies with no observed efficacy could halt the pursuit of large and expensive clinical trials in malaria-endemic areas in time. CHMI studies with partial efficacy could provide insights on how protective efficacy could be improved by alterations in vaccination regimes such as the number of doses and number of immunizations. In particular, through a series of CHMI studies, the company Sanaria was able to optimize their PfSPZ vaccine, which is composed of radiation-attenuated, aseptic, purified, cryopreserved P. falciparum sporozoites, to induce sterile protection against homologous challenge for at least 59 weeks (24) and heterologous challenge for at least 33 weeks (51) in malaria-naïve individuals. The vaccination also prevented naturally transmitted heterogeneous P. falciparum in malaria-endemic adults in Mali for at least 24 weeks (vaccine efficacy of 29%) (14). Further CHMI studies to optimize dosage and vaccination regimes could improve the vaccine efficacy.
Immunity and Correlates of Protection Against the Pre-erythrocytic Stage
The malaria parasite has a complex life cycle. Depending on the stage of development in its mammalian host, the parasite can be extracellular or intracellular. They can also infect different cell types. Hence, various innate and adaptive immune mechanisms are needed for parasite control and elimination. In order to develop an efficacious pre-erythrocytic stage vaccine, it is important to know the protective immune mechanisms to induce (Table 1).
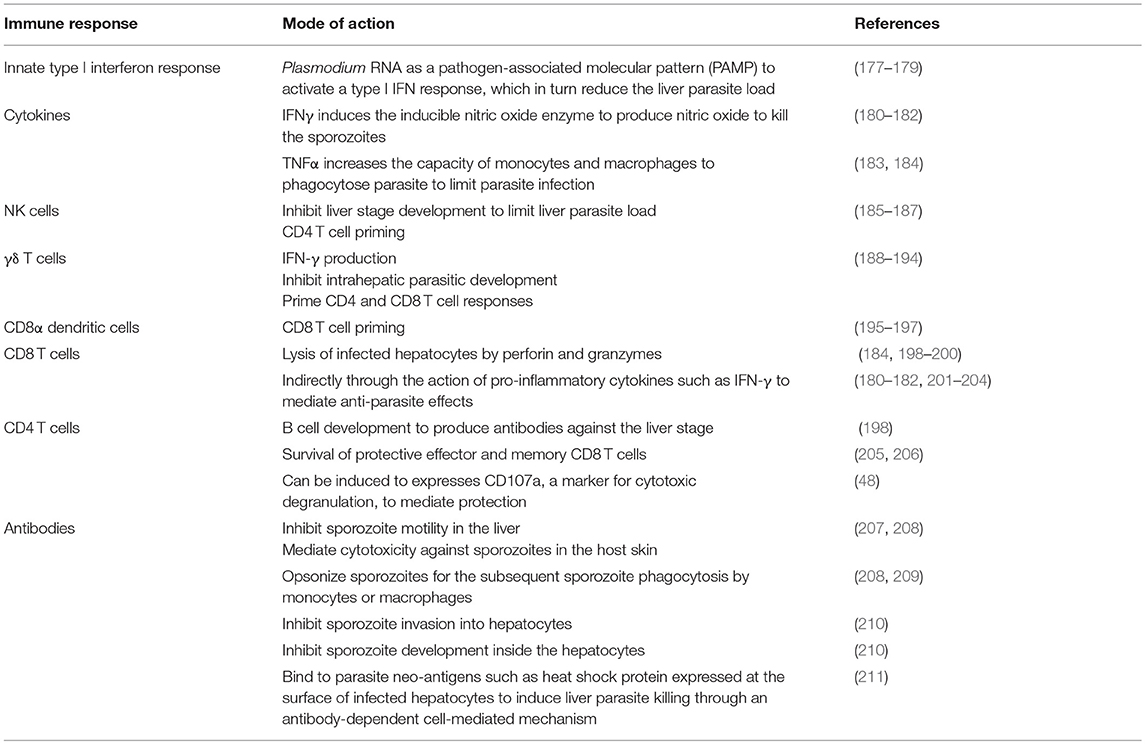
Table 1. Immunity against pre-erythrocytic stage parasites.
In addition, through better understanding of the mechanisms involved in the protection, we could potentially identify correlates of protection. The identification of correlates of protection is particularly important to the vaccine development as it helps to assess vaccine efficacy and design better immunogens. Through various animal models, we are beginning to tease out the potential correlates of protective immunity.
Innate Immunity
Upon infection, the innate immunity is triggered by the malaria parasites. Immune responses initiated by the innate immune system in response to parasites play key roles in protective immunity development. Early pro-inflammatory responses regulate anti-parasitic Th1 development and promote effector cell function for efficiently clearing infections. The use of a proper adjuvant is necessary to trigger the adequate innate pathway.
Cytokines
Cytokines play an important role in the protection against malaria. Upon infection, the RNA of the parasites is recognized by the cytosolic pathogen-recognition receptors of mouse hepatocytes. As a result, type I interferon pathway is induced, which can inhibit late stage parasites. Type I interferon leads to the recruitment of leukocytes that inhibit late liver forms through IFNγ (179, 212). IFNγ can inhibit the development of P. yoelii and P. berghei in vitro and in vivo in mice (177, 178), P. falciparum in human hepatocytes in vitro (213) and P. vivax infected chimpanzees in vivo (214). The effect of IFNγ is through the induction of the inducible nitric oxide synthase enzyme in hepatocyte which generates high of toxic nitric oxide (180–182).
In addition to IFNγ, IL6, and TNFα have also been implicated in protection. TNFα is able to inhibit parasite liver stage indirectly through induction of yet-to-be-identified mediators secreted by hepatocytes (215) or through IL-6 on non-parenchymal cells (216). IL-6 inhibits liver stage development through the induction of iNOS (217, 218).
NK and NKT Cells
NK and NKT cells are abundant in the liver, and interact with the parasites to initiate liver-stage cell-mediated immunity. Following the activation of the type I interferon pathway, the hepatocytes produce chemokines to recruit macrophages, neutrophils, lymphocytes, NK and NKT cells to the site of infection in mice (179, 219). This eventually leads to the killing of late liver stage parasites by NKT cells. NK cells have also been shown to inhibit the development of the liver stages in the hepatocytes, limiting the infection and/or reinfection in mice (185). NK cells also play an important role in CD4 T cell priming during murine malaria infections (186, 187), bridging between the innate and adaptive immunity.
γδ T Cells
Similar to NK T cells, γδ T cells are innate-like T cells that have been postulated to bridge the gap between innate and adaptive immunity (220). Early production of IFNγ by γδ T cells was detected following in vitro exposure of P. falciparum-infected RBCs to PBMCs from malaria-naïve donors (188). In mice, γδ T cells induced by whole sporozoites vaccination can inhibit intrahepatic parasitic development (190). γδ T cells are also important for the induction of early immunity against malaria (191). γδ T cell-deficient mice immunized with irradiated sporozoites were more susceptible to liver stage infection 42 h following a sporozoite challenge (191). γδ T cells can also directly prime CD4 and CD8 T cell responses in vitro (192, 193).
Sterile protection in mice following vaccination with irradiated sporozoites requires γδ T cells (194). Without γδ T cells, protective CD8 T cell responses were impaired (194). γδ T cells have been postulated to act either as effector cells that operate in the absence of αβ T cells, or as accessory cells for appropriate protective responses from other cells (194). In humans, γδ T cells have also been shown to recognize malaria antigens and proliferate, conferring immunity against clinical malaria in children from Uganda (221). In addition to influencing the protective CD8 T cell response, γδ T cells can also influence cytokine production. Higher frequencies and higher cytokine production by γδ T cells correlate with protection against subsequent infection in children living in endemic settings (189, 222). Recent studies on irradiated sporozoite vaccines have shown that γδ T cells expanded in a dose-dependent manner in immunized malaria-naïve subjects (6, 24), and were associated with protection (24). Hence, γδ T cells could be a potential correlate of protection, and further studies to better define a most appropriate outcome to represent a measurable positive correlation of γδ T cells with protection would be advantageous for vaccine development.
CD8α Dendritic Cells
Early adaptive immunity is triggered as early as a few hours post an infective mosquito bite, with T cell activation being observed in the skin draining lymph nodes in the murine model (196). After dermal inoculation, a fraction of sporozoites actively migrates to the draining lymph nodes (195). There is a direct uptake of the parasites by lymph-node resident CD8α dendritic cells followed by CD8 T cell-dendritic cell cluster formation in the draining lymph nodes (223). CD8α dendritic cells are also shown to be essential for the development of the protective immunity induced by intravenous injection of irradiated sporozoites since mice depleted of these subsets are not protected against a sporozoite challenge (224, 225). A subsequent study showed that splenic but not liver CD8α dendritic cells are the main cells involved in effector parasite specific-T cell priming (226). It was also shown recently that monocyte-derived CD11c cells infiltrated the liver after infection, acquired parasite-derived antigens and primed protective CD8 T cells (227). The role and functions of other dendritic subsets is controversial and remains to be determined (199, 228, 229).
Adaptive Immunity
As with any vaccination, the focus has been to trigger the adaptive immunity to induce an efficacious and long-lasting immunity. Various arms of the adaptive immunity are required to act in concert to provide protection against malaria.
CD8 T Cells
CD8 T cells have been implicated as the principal effector cells, central to protection against malaria. The importance of CD8 T cells in protective immunity was first demonstrated in mice vaccinated with irradiated sporozoites (177). The sterile immunity induced by the vaccination was abolished when CD8 T cells were depleted (177, 178). CD8 T cells can kill the parasites in mice (200) either directly through lysis of infected hepatocytes by perforin and granzymes (184, 230) or indirectly through IFNγ-mediated protection (180–182, 201–204). It must be stressed that while leukocytes and, in particular, CD8 T cells can kill liver parasites by these mechanisms, they differ depending of the host/parasite combinations (231).
Given the central role of CD8 T cells in protection, it is one potential correlate of protective immunity. In humans, CD8 T cells have been shown to be associated with protection from severe malaria (232), and a few of the identified CD8 T cell responses are directed against pre-erythrocytic stage antigens such as LSA1 and CSP (233, 234). However, vaccination studies, where human volunteers were immunized with irradiated sporozoites, showed that, while CD8 T cell response were also detected against various pre-erythrocytic stage antigens, the responses were not found to be associated with protection (235). More recently, a human trial where human volunteers were immunized with irradiated sporozoites showed seemingly contradicting data, where sterile protection correlated with the numbers of IFNγ-producing CD8 T cells in isolated PBMCs (6). This is also evident in animal studies on irradiated sporozoite vaccines, where high frequency of parasite-specific CD8 T cells was observed in the liver of non-human primates and mice, and was associated with protection in mice (236). The differences are likely due to the differences in the vaccination regimes and the methods to detect/quantify T cells. Indeed, in humans, T cell activity is measured in peripheral blood, whereas, in mice, T cell activity is usually measured in spleen or liver.
Central memory T cells
Memory T cells provide long-term protection. Upon re-infection, these cells rapidly gain effector functions including cytokine production and lytic activity. There are three subsets of memory T cells: (1) central memory T cells, which predominantly reside in lymphoid tissues, (2) effector memory T cells, which reside in the spleen and peripheral tissues, and (3) tissue-resident memory T cells, which reside in the tissues and do not recirculate. The role of central memory T cells in protection against malaria is limited. While central memory T cells can produce IFN-γ after in vitro stimulation (201), their presence has not been associated with protection. Despite having a large proportion of central memory T cell, mice that were vaccinated with modified vaccinia ANKARA expressing the multiple epitope string and thrombospondin-related adhesion protein (ME-TRAP) were not protected from malaria challenge (237).
CD8 effector memory T cells
In contrast to the central memory T cells, the presence of effector memory T cells has been associated with sterile protection in the murine model, although large numbers are required for protection against malaria (238). Long-term sterile protection was only observed in mice when the parasite-specific CD8 T cells made up >1% of the total peripheral blood CD8 T cell population (238). Degree of protection in mice correlated with the frequencies of CD8 effector memory T cells present in liver, and failure to achieve the protective threshold frequency of these cells might make the host susceptible to infection (203).
CD8 tissue-resident memory T cells
More recently, a new subset of memory T cells, with a distinct gene expression profile, has been characterized (202). The liver tissue-resident memory T cells develop naturally during the course of an immune response following TCR stimulation, with rapidly expanding population due to the liver infection or inflammation in mice (239). These cells are found to be patrolling within the liver sinusoids, a process dependent on LFA-1–ICAM-1 interactions (240). Tissue-resident memory T cells were essential for sterile protection against sporozoite infection in mice following immunization with irradiated sporozoites (241).
CD4 T Cells
In contrast to CD8 T cells, the role of CD4 T cells in protection against malaria is not well understood. Despite this, it is clear that the development and maturation of an effective CD8T cell response is dependent on CD4 T cells help. CD4 T cells are activated to amplify the anti-pathogen response by driving B cell germinal responses and supporting CD8 T cell activation. Mouse hepatocytes express MHC Class I and Class II molecules that can be loaded with parasite antigen-derived epitopes following the TAP or the endosomal pathways (242–245). CD4 T cells are required to prime effective immunity. CD4 T cells recognizing CSP have been shown to protect against a P. yoelii sporozoite challenge in mice (243, 246). Pre-immunization but not pre-challenge depletion of CD4 T cells also resulted in a loss of protection in mice immunized with sporozoites, suggesting that CD4 T cells might provide signals for efficient maturation of effector CD8 T cells (247). In mice, CD4 T cells were essential to ensure survival of protective effector and memory CD8 T cell induced by irradiated sporozoites (205, 206). In humans, many studies have described a CSP-specific CD4 T cell response that is associated with protection against natural infection and disease (248), and is able to inhibit pre-erythrocytic stage development (249). CD4 T cells have also been shown to correlate with sterile protection in humans following immunization with live sporozoites under chloroquine prophylaxis. CSP-specific CD4 T cells were induced to express CD107a, a marker for cytotoxic degranulation, after immunizations with live sporozoites under chloroquine prophylaxis in humans, and these cytotoxic markers has been shown to be associated with sterile protection against the pre-erythrocytic stages (48). In addition to being crucial for B cell development to produce antibodies, CD4 T cells are also important for CD8 T cell responses.
While it is clear that CD4 T cells are involved in protective immunity against malaria, its use as a potential correlate of protection needs further validation. Together with CD8 T cells, the definitive role of CD4 T cells in protection requires more unraveling and the information will be critical to the development of a validated T cell-based correlate of protection for vaccine efficacy assessment.
Antibodies
Lastly, in addition to inducing an effective CD8 T cell response, the development of many malaria candidate vaccines also aims at being able to induce an effective antibody response. Antibodies are often the first host immune response being studied. Antibodies against the pre-erythrocytic stage can mediate protection by limiting pre-erythrocytic stage infection and development. More specifically, the antibodies do so by (1) inhibiting sporozoite motility in the dermis and liver (207), (2) mediating cytotoxicity against sporozoites in the host skin (208, 250), (3) opsonizing the sporozoites and subsequently facilitating sporozoite phagocytosis by monocytes or macrophages in the spleen or the liver (208, 209), (4) inhibiting sporozoite invasion into hepatocytes (210), (5) inhibiting sporozoite development inside the hepatocytes (210), and (6) binding to parasite neo-antigens such as heat shock protein expressed at the surface of infected hepatocytes and eventually inducing liver parasite killing through an antibody-dependent cell-mediated mechanism that is likely to involve Kupffer cells or NK cells (211).
Antibodies are potential correlates of protection. Passive transfer of RTS,S-induced human anti-CSP monoclonal antibodies into humanized mice at concentrations within the range observed in human, protected the mice against P. falciparum challenge (170). Immunization with genetically-attenuated sporozoites that arrest late in the liver stage development elicited protection against both a sporozoite challenge and a direct blood stage challenge by inducing the production of stage-transcending protective antibodies in mice (251). Sporozoite-specific antibodies induced by vaccination with irradiated sporozoites (252) or genetically-attenuated sporozoites (253) have also been shown to inhibit sporozoite invasion into human hepatocytes in vitro and correlate with protection in human individuals (254–256). However, a recent human study examining protection following vaccination with irradiated sporozoites in malaria-naïve individuals has found no significant correlation of antibody response with protection (46). In addition, there is no distinct antibody profile that allows differentiation of protected individuals from the susceptible individuals (257) following RTS,S vaccination. There is increasing awareness that, in addition of high level of antibodies, the quality of the antibodies is also important. In vitro assays to examine the functionality of the induced antibodies following vaccination of pre-erythrocytic stage vaccine have been developed. These assays include gliding motility assays (258), sporozoite traversal and invasion inhibition assays (6, 253, 259), and pre-erythrocytic stage development inhibition assays (136, 260). Recently, human monoclonal antibodies have been derived from volunteers immunized with irradiated sporozoites. These antibodies recognized an important epitope at the junction of the N terminal part and the repeat regions of the CSP. This can lead to the design of better CSP-based vaccines (146, 261). Further studies to draw parallels between the readouts of these assays and the protection in the field are necessary to develop a validated antibody-based correlate of protection for vaccine efficacy assessment.
Concluding Remarks
In contrast to the limited efficacy of RTS,S and other subunit vaccines, vaccination with sporozoites has had more success. In addition to promising efficacy data, a series of recent clinical trials on sporozoite-based vaccines has demonstrated formidable advances in overcoming issues in vaccine manufacturing and delivery (6, 14, 23, 46). Furthermore, there are now data showing that sporozoite vaccines are safe and tolerated in malaria-endemic areas (14, 23). While the vaccine efficacy is markedly reduced against heterologous CHMI (as compared with homologous CHMI), it is encouraging that it offers some protection against heterologous CHMI (25). Further studies to optimize the immunization regimen could potentially improve the vaccine efficacy. Here, we reviewed the various types of vaccination strategies with sporozoites and the different animal models being used for the vaccination studies. We also discussed the mechanisms of protection against the pre-erythrocytic parasites. While the mechanisms of protection are slowly being unraveled, the establishment of validated correlates of protection for assessment of vaccine efficacy has proved to be challenging. Half of the world population is at risk of a malaria infection. The target population is highly diverse, with individuals from different age groups (infants, adults and elderly), different exposed status (endemic and non-endemic), and different immunological background (immunocompromised and pregnant). The presence of co-infections in some populations in malaria-endemic regions adds further complexity. In addition, the complexity of the parasite and the diversity of its genome also makes it difficult to definitively establish correlates of protection. Depending on which part of the parasite life cycle the malaria vaccine candidates target, different forms of immunity are induced. As it is still unclear if the ultimate goal of a malaria vaccine should be to protect against infection or simply to protect against disease, different clinical endpoints have been used to measure vaccine efficacy. Vaccine-induced immune responses that correlate with protection against one endpoint may not necessarily correlate with protection against a different endpoint. Hence, until date, there is no validated correlate of protection. A concerted effort to develop/refine relevant animal models, investigate the definitive mechanisms of protection and identify validated correlates of protection would greatly help to inform critical decisions in human vaccine clinical trial, which will accelerate future progress in the development of an efficacious malaria vaccine.
Author Contributions
All authors listed have made a substantial, direct and intellectual contribution to the work, and approved it for publication.
Funding
This work was supported by the Agency for Science Technology and Research (A*STAR).
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
1. WHO. Malaria Fact Sheet. [online]. Available online at: http://www.who.int/mediacentre/factsheets/fs094/en/ (accessed January, 2019) (Online) (2018).
2. Bhatt S, Weiss DJ, Cameron E, Bisanzio D, Mappin B, Dalrymple U, et al. The effect of malaria control on Plasmodium falciparum in Africa between 2000 and 2015. Nature. (2015) 526:207–11. doi: 10.1038/nature15535
3. Hemingway J, Ranson H, Magill A, Kolaczinski J, Fornadel C, Gimnig J, et al. Averting a malaria disaster: will insecticide resistance derail malaria control? Lancet. (2016) 387:1785–8. doi: 10.1016/S0140-6736(15)00417-1
4. Wiebe A, Longbottom J, Gleave K, Shearer FM, Sinka ME, Massey NC, et al. Geographical distributions of African malaria vector sibling species and evidence for insecticide resistance. Malar J. (2017) 16:85. doi: 10.1186/s12936-017-1734-y
5. Healer J, Cowman AF, Kaslow DC, Birkett AJ. Vaccines to accelerate malaria elimination and eventual eradication. Cold Spring Harb Perspect Med. (2017) 7:a025627. doi: 10.1101/cshperspect.a025627
6. Seder RA, Chang LJ, Enama ME, Zephir KL, Sarwar UN, Gordon IJ, et al. Protection against malaria by intravenous immunization with a nonreplicating sporozoite vaccine. Science. (2013) 341:1359–65. doi: 10.1126/science.1241800
7. Jongo SA, Shekalaghe SA, Church LWP, Ruben AJ, Schindler T, Zenklusen I, et al. Safety, immunogenicity, and protective efficacy against controlled human malaria infection of Plasmodium falciparum sporozoite vaccine in Tanzanian adults. Am J Trop Med Hyg. (2018) 99:338–49. doi: 10.4269/ajtmh.17-1014
8. Sergent E, Sergent E. Sur l'immunite sdans le paludismedes oiseaux. Conservation in vitro des sporozoites de plasmodium de Plasmodium relictum immmunite relative obtenue par inoculations de ces sporozoites. C R Acad Sci. (1910) 151:407–9.
9. Russell PF, Mohan BN. The immunization of fowls against mosquito-borne Plasmodium gallinaceum by injections of serum and of inactivated homologous sporozoites. J Exp Med. (1942) 76:477–95. doi: 10.1084/jem.76.5.477
10. Freund J, Sommer HE, Walter AW. Immunization against malaria: vaccination of ducks with killed parasites incorporated with adjuvants. Science. (1945) 102:200–2. doi: 10.1126/science.102.2643.200
11. Freund J, Thomson KJ, Sommer HE, Walter AW, Schenkein EL. Immunization of rhesus monkeys against malarial infection (P. Knowlesi) with killed parasites and adjuvants. Science. (1945) 102:202–4. doi: 10.1126/science.102.2643.202
12. Heidelberger M, Mayer MM, Demarest CR. Studies in human malaria; the preparation of vaccines and suspensions containing plasmodia. J Immunol. (1946) 52:325–30.
13. Rosenberg R. Ejection of malaria sporozoites by feeding mosquitoes. Trans R Soc Trop Med Hyg. (1992) 86:109. doi: 10.1016/0035-9203(92)90466-P
14. Sissoko MS, Healy SA, Katile A, Omaswa F, Zaidi I, Gabriel EE, et al. Safety and efficacy of PfSPZ vaccine against Plasmodium falciparum via direct venous inoculation in healthy malaria-exposed adults in Mali: a randomised, double-blind phase 1 trial. Lancet Infect Dis. (2017) 17:498–509. doi: 10.1016/S1473-3099(17)30104-4
15. Spring M, Murphy J, Nielsen R, Dowler M, Bennett JW, Zarling S, et al. First-in-human evaluation of genetically attenuated Plasmodium falciparum sporozoites administered by bite of anopheles mosquitoes to adult volunteers. Vaccine. (2013) 31:4975–83. doi: 10.1016/j.vaccine.2013.08.007
16. Kublin JG, Mikolajczak SA, Sack BK, Fishbaugher ME, Seilie A, Shelton L, et al. Complete attenuation of genetically engineered Plasmodium falciparum sporozoites in human subjects. Sci Transl Med. (2017) 9:eaad9099. doi: 10.1126/scitranslmed.aad9099
17. Richie TL, Billingsley PF, Sim BK, James ER, Chakravarty S, Epstein JE, et al. Progress with Plasmodium falciparum sporozoite (PfSPZ)-based malaria vaccines. Vaccine. (2015) 33:7452–61. doi: 10.1016/j.vaccine.2015.09.096
18. Richards WH. Active immunization of chicks against Plasmodium gallinaceum by inactivated homologous sporozoites and erythrocytic parasites. Nature. (1966) 212:1492–4. doi: 10.1038/2121492a0
19. Nussenzweig RS, Vanderberg J, Most H, Orton C. Protective immunity produced by the injection of x-irradiated sporozoites of Plasmodium berghei. Nature. (1967) 216:160–2. doi: 10.1038/216160a0
20. Clyde DF, Mccarthy VC, Miller RM, Hornick RB. Specificity of protection of man immunized against sporozoite-induced Falciparum malaria. Am J Med Sci. (1973) 266:398–403. doi: 10.1097/00000441-197312000-00001
21. Rieckmann KH, Carson PE, Beaudoin RL, Cassells JS, Sell KW. Letter: sporozoite induced immunity in man against an Ethiopian strain of Plasmodium falciparum. Trans R Soc Trop Med Hyg. (1974) 68:258–9. doi: 10.1016/0035-9203(74)90129-1
22. Hoffman SL, Goh LM, Luke TC, Schneider I, Le TP, Doolan DL, et al. Protection of humans against malaria by immunization with radiation-attenuated Plasmodium falciparum sporozoites. J Infect Dis. (2002) 185:1155–64. doi: 10.1086/339409
23. Hickey BW, Lumsden JM, Reyes S, Sedegah M, Hollingdale MR, Freilich DA, et al. Mosquito bite immunization with radiation-attenuated Plasmodium falciparum sporozoites: safety, tolerability, protective efficacy and humoral immunogenicity. Malar J. (2016) 15:377. doi: 10.1186/s12936-016-1435-y
24. Ishizuka AS, Lyke KE, Dezure A, Berry AA, Richie TL, Mendoza FH, et al. Protection against malaria at 1 year and immune correlates following PfSPZ vaccination. Nat Med. (2016) 22:614–23. doi: 10.1038/nm.4110
25. Epstein JE, Paolino KM, Richie TL, Sedegah M, Singer A, Ruben AJ, et al. Protection against Plasmodium falciparum malaria by PfSPZ vaccine. JCI Insight. (2017) 2:e89154. doi: 10.1172/jci.insight.89154
26. Renia L, Goh YS. Malaria parasites: the great escape. Front Immunol. (2016) 7:463. doi: 10.3389/fimmu.2016.00463
27. Vanderberg JP, Nussenzweig RS, Most H, Orton CG. Protective immunity produced by the injection of x-irradiated sporozoites of Plasmodium berghei. II Effects of radiation on sporozoites. J Parasitol. (1968) 54:1175–80. doi: 10.2307/3276987
28. Spitalny GL, Nussenzweig RS. Effect of various routes of immunization and methods of parasite attenuation on development of protection against sporozoite-induced rodent malaria. Proc Helminthol Soc Wash. (1972) 39:506–14.
29. Nussler A, Follezou JY, Miltgen F, Mazier D. Effect of irradiation on Plasmodium sporozoites depends on the species of hepatocyte infected. Trop Med Parasitol. (1989) 40:468–9.
30. Mellouk S, Lunel F, Sedegah M, Beaudoin RL, Druilhe P. Protection against malaria induced by irradiated sporozoites. Lancet. (1990) 335:721. doi: 10.1016/0140-6736(90)90832-P
31. Hoffman BU, Chattopadhyay R. Plasmodium falciparum: effect of radiation on levels of gene transcripts in sporozoites. Exp Parasitol. (2008) 118:247–52. doi: 10.1016/j.exppara.2007.08.014
32. Oakley MS, Verma N, Zheng H, Anantharaman V, Takeda K, Gao Y, et al. Molecular markers of radiation induced attenuation in intrahepatic Plasmodium falciparum parasites. PLoS ONE. (2016) 11:e0166814. doi: 10.1371/journal.pone.0166814
33. Suhrbier A, Winger LA, Castellano E, Sinden RE. Survival and antigenic profile of irradiated malarial sporozoites in infected liver cells. Infect Immun. (1990) 58:2834–9.
34. Silvie O, Semblat JP, Franetich JF, Hannoun L, Eling W, Mazier D. Effects of irradiation on Plasmodium falciparum sporozoite hepatic development: implications for the design of pre-erythrocytic malaria vaccines. Parasite Immunol. (2002) 24:221–3. doi: 10.1046/j.1365-3024.2002.00450.x
35. Renia L, Gruner AC, Mauduit M, Snounou G. Vaccination against malaria with live parasites. Expert Rev Vaccines. (2006) 5:473–81. doi: 10.1586/14760584.5.4.473
36. Vaughan AM, Wang R, Kappe SH. Genetically engineered, attenuated whole-cell vaccine approaches for malaria. Hum Vaccin. (2010) 6:107–13. doi: 10.4161/hv.6.1.9654
37. Mueller AK, Camargo N, Kaiser K, Andorfer C, Frevert U, Matuschewski K, et al. Plasmodium liver stage developmental arrest by depletion of a protein at the parasite-host interface. Proc Natl Acad Sci USA. (2005) 102:3022–7. doi: 10.1073/pnas.0408442102
38. Mueller AK, Labaied M, Kappe SH, Matuschewski K. Genetically modified Plasmodium parasites as a protective experimental malaria vaccine. Nature. (2005) 433:164–7. doi: 10.1038/nature03188
39. van Dijk MR, Douradinha B, Franke-Fayard B, Heussler V, Van Dooren MW, Van Schaijk B, et al. Genetically attenuated, P36p-deficient malarial sporozoites induce protective immunity and apoptosis of infected liver cells. Proc Natl Acad Sci USA. (2005) 102:12194–9. doi: 10.1073/pnas.0500925102
40. Butler NS, Schmidt NW, Vaughan AM, Aly AS, Kappe SH, Harty JT. Superior antimalarial immunity after vaccination with late liver stage-arresting genetically attenuated parasites. Cell Host Microbe. (2011) 9:451–62. doi: 10.1016/j.chom.2011.05.008
41. Vaughan AM, Sack BK, Dankwa D, Minkah N, Nguyen T, Cardamone H, et al. A plasmodium parasite with complete late liver stage arrest protects against preerythrocytic and erythrocytic stage infection in mice. Infect Immun. (2018) 86:e00088–18. doi: 10.1128/IAI.00088-18
42. Beaudoin RL, Strome CP, Mitchell F, Tubergen TA. Plasmodium berghei: immunization of mice against the ANKA strain using the unaltered sporozoite as an antigen. Exp Parasitol. (1977) 42:1–5. doi: 10.1016/0014-4894(77)90054-6
43. Orjih AU, Cochrane AH, Nussenzweig RS. Comparative studies on the immunogenicity of infective and attenuated sporozoites of Plasmodium berghei. Trans R Soc Trop Med Hyg. (1982) 76:57–61. doi: 10.1016/0035-9203(82)90019-0
44. Belnoue E, Costa FT, Frankenberg T, Vigario AM, Voza T, Leroy N, et al. Protective T cell immunity against malaria liver stage after vaccination with live sporozoites under chloroquine treatment. J Immunol. (2004) 172:2487–95. doi: 10.4049/jimmunol.172.4.2487
45. Roestenberg M, Mccall M, Hopman J, Wiersma J, Luty AJ, Van Gemert GJ, et al. Protection against a malaria challenge by sporozoite inoculation. N Engl J Med. (2009) 361:468–77. doi: 10.1056/NEJMoa0805832
46. Mordmuller B, Surat G, Lagler H, Chakravarty S, Ishizuka AS, Lalremruata A, et al. Sterile protection against human malaria by chemoattenuated PfSPZ vaccine. Nature. (2017) 542:445–9. doi: 10.1038/nature21060
47. Friesen J, Silvie O, Putrianti ED, Hafalla JC, Matuschewski K, Borrmann S. Natural immunization against malaria: causal prophylaxis with antibiotics. Sci Transl Med. (2010) 2:40ra49. doi: 10.1126/scitranslmed.3001058
48. Bijker EM, Teirlinck AC, Schats R, Van Gemert GJ, Van De Vegte-Bolmer M, Van Lieshout L, et al. Cytotoxic markers associate with protection against malaria in human volunteers immunized with Plasmodium falciparum sporozoites. J Infect Dis. (2014) 210:1605–15. doi: 10.1093/infdis/jiu293
49. Peng X, Keitany GJ, Vignali M, Chen L, Gibson C, Choi K, et al. Artesunate versus chloroquine infection-treatment-vaccination defines stage-specific immune responses associated with prolonged sterile protection against both pre-erythrocytic and erythrocytic Plasmodium yoelii infection. J Immunol. (2014) 193:1268–77. doi: 10.4049/jimmunol.1400296
50. Schats R, Bijker EM, Van Gemert GJ, Graumans W, Van De Vegte-Bolmer M, Van Lieshout L, et al. Heterologous protection against malaria after immunization with Plasmodium falciparum sporozoites. PLoS ONE. (2015) 10:e0124243. doi: 10.1371/journal.pone.0124243
51. Lyke KE, Ishizuka AS, Berry AA, Chakravarty S, Dezure A, Enama ME, et al. Attenuated PfSPZ vaccine induces strain-transcending T cells and durable protection against heterologous controlled human malaria infection. Proc Natl Acad Sci USA. (2017) 114:2711–6. doi: 10.1073/pnas.1615324114
52. Yoshida N, Nussenzweig RS, Potocnjak P, Nussenzweig V, Aikawa M. Hybridoma produces protective antibodies directed against the sporozoite stage of malaria parasite. Science. (1980) 207:71–3. doi: 10.1126/science.6985745
53. Dame JB, Williams JL, Mccutchan TF, Weber JL, Wirtz RA, Hockmeyer WT, et al. Structure of the gene encoding the immunodominant surface antigen on the sporozoite of the human malaria parasite Plasmodium falciparum. Science. (1984) 225:593–9. doi: 10.1126/science.6204383
54. Enea V, Ellis J, Zavala F, Arnot DE, Asavanich A, Masuda A, et al. DNA cloning of Plasmodium falciparum circumsporozoite gene: amino acid sequence of repetitive epitope. Science. (1984) 225:628–30. doi: 10.1126/science.6204384
55. Egan JE, Weber JL, Ballou WR, Hollingdale MR, Majarian WR, Gordon DM, et al. Efficacy of murine malaria sporozoite vaccines: implications for human vaccine development. Science. (1987) 236:453–6. doi: 10.1126/science.3551073
56. Zavala F, Tam JP, Barr PJ, Romero PJ, Ley V, Nussenzweig RS, et al. Synthetic peptide vaccine confers protection against murine malaria. J Exp Med. (1987) 166:1591–6. doi: 10.1084/jem.166.5.1591
57. Tam JP, Clavijo P, Lu YA, Nussenzweig V, Nussenzweig R, Zavala F. Incorporation of T and B epitopes of the circumsporozoite protein in a chemically defined synthetic vaccine against malaria. J Exp Med. (1990) 171:299–306. doi: 10.1084/jem.171.1.299
58. Marussig M, Renia L, Motard A, Miltgen F, Petour P, Chauhan V, et al. Linear and multiple antigen peptides containing defined T and B epitopes of the Plasmodium yoelii circumsporozoite protein: antibody-mediated protection and boosting by sporozoite infection. Int Immunol. (1997) 9:1817–24. doi: 10.1093/intimm/9.12.1817
59. Herrington DA, Clyde DF, Losonsky G, Cortesia M, Murphy JR, Davis J, et al. Safety and immunogenicity in man of a synthetic peptide malaria vaccine against Plasmodium falciparum sporozoites. Nature. (1987) 328:257–9. doi: 10.1038/328257a0
60. Good MF, Berzofsky JA, Maloy WL, Hayashi Y, Fujii N, Hockmeyer WT, et al. Genetic control of the immune response in mice to a Plasmodium falciparum sporozoite vaccine. Widespread nonresponsiveness to single malaria T epitope in highly repetitive vaccine. J Exp Med. (1986) 164:655–60. doi: 10.1084/jem.164.2.655
61. Sinigaglia F, Guttinger M, Kilgus J, Doran DM, Matile H, Etlinger H, et al. A malaria T-cell epitope recognized in association with most mouse and human MHC class II molecules. Nature. (1988) 336:778–80. doi: 10.1038/336778a0
62. Schodel F, Wirtz R, Peterson D, Hughes J, Warren R, Sadoff J, et al. Immunity to malaria elicited by hybrid hepatitis B virus core particles carrying circumsporozoite protein epitopes. J Exp Med. (1994) 180:1037–46. doi: 10.1084/jem.180.3.1037
63. Kubler-Kielb J, Majadly F, Biesova Z, Mocca CP, Guo C, Nussenzweig R, et al. A bicomponent Plasmodium falciparum investigational vaccine composed of protein-peptide conjugates. Proc Natl Acad Sci USA. (2010) 107:1172–7. doi: 10.1073/pnas.0913374107
64. Powell TJ, Tang J, Derome ME, Mitchell RA, Jacobs A, Deng Y, et al. Plasmodium falciparum synthetic LbL microparticle vaccine elicits protective neutralizing antibody and parasite-specific cellular immune responses. Vaccine. (2013) 31:1898–904. doi: 10.1016/j.vaccine.2013.02.027
65. Collins KA, Snaith R, Cottingham MG, Gilbert SC, Hill AVS. Enhancing protective immunity to malaria with a highly immunogenic virus-like particle vaccine. Sci Rep. (2017) 7:46621. doi: 10.1038/srep46621
66. Espinosa DA, Christensen D, Munoz C, Singh S, Locke E, Andersen P, et al. Robust antibody and CD8(+) T-cell responses induced by P. falciparum CSP adsorbed to cationic liposomal adjuvant CAF09 confer sterilizing immunity against experimental rodent malaria infection. NPJ Vaccines. (2017) 2:10. doi: 10.1038/s41541-017-0011-y
67. Kaba SA, Karch CP, Seth L, Ferlez KMB, Storme CK, Pesavento DM, et al. Self-assembling protein nanoparticles with built-in flagellin domains increases protective efficacy of a Plasmodium falciparum based vaccine. Vaccine. (2018) 36:906–14. doi: 10.1016/j.vaccine.2017.12.001
68. Cohen J, Nussenzweig V, Nussenzweig R, Vekemans J, Leach A. From the circumsporozoite protein to the RTS, S/AS candidate vaccine. Hum Vaccin. (2010) 6:90–6. doi: 10.4161/hv.6.1.9677
69. Gordon DM, Mcgovern TW, Krzych U, Cohen JC, Schneider I, Lachance R, et al. Safety, immunogenicity, and efficacy of a recombinantly produced Plasmodium falciparum circumsporozoite protein-hepatitis B surface antigen subunit vaccine. J Infect Dis. (1995) 171:1576–85. doi: 10.1093/infdis/171.6.1576
70. Stoute JA, Slaoui M, Heppner DG, Momin P, Kester KE, Desmons P, et al. A preliminary evaluation of a recombinant circumsporozoite protein vaccine against Plasmodium falciparum malaria. RTS,S malaria vaccine evaluation group. N Engl J Med. (1997) 336:86–91. doi: 10.1056/NEJM199701093360202
71. Kester KE, Mckinney DA, Tornieporth N, Ockenhouse CF, Heppner DG, Hall T, et al. Efficacy of recombinant circumsporozoite protein vaccine regimens against experimental Plasmodium falciparum malaria. J Infect Dis. (2001) 183:640–7. doi: 10.1086/318534
72. Garcon N, Chomez P, Van Mechelen M. GlaxoSmithKline adjuvant systems in vaccines: concepts, achievements and perspectives. Expert Rev Vaccines. (2007) 6:723–39. doi: 10.1586/14760584.6.5.723
73. Kester KE, Cummings JF, Ofori-Anyinam O, Ockenhouse CF, Krzych U, Moris P, et al. Randomized, double-blind, phase 2a trial of falciparum malaria vaccines RTS,S/AS01B and RTS,S/AS02A in malaria-naive adults: safety, efficacy, and immunologic associates of protection. J Infect Dis. (2009) 200:337–46. doi: 10.1086/600120
74. Heppner DG Jr, Walsh DS, Uthaimongkol N, Tang DB, Tulyayon S, Permpanich B, et al. Randomized, controlled, double-blind trial of daily oral azithromycin in adults for the prophylaxis of Plasmodium vivax malaria in Western Thailand. Am J Trop Med Hyg. (2005) 73:842–9. doi: 10.4269/ajtmh.2005.73.842
75. Bojang KA, Milligan PJ, Pinder M, Vigneron L, Alloueche A, Kester KE, et al. Efficacy of RTS,S/AS02 malaria vaccine against Plasmodium falciparum infection in semi-immune adult men in the Gambia: a randomised trial. Lancet. (2001) 358:1927–34. doi: 10.1016/S0140-6736(01)06957-4
76. Alonso PL, Sacarlal J, Aponte JJ, Leach A, Macete E, Milman J, et al. Efficacy of the RTS,S/AS02A vaccine against Plasmodium falciparum infection and disease in young African children: randomised controlled trial. Lancet. (2004) 364:1411–20. doi: 10.1016/S0140-6736(04)17223-1
77. Olotu A, Moris P, Mwacharo J, Vekemans J, Kimani D, Janssens M, et al. Circumsporozoite-specific T cell responses in children vaccinated with RTS,S/AS01E and protection against P falciparum clinical malaria. PLoS ONE. (2011) 6:e25786. doi: 10.1371/journal.pone.0025786
78. Rts SCTP, Agnandji ST, Lell B, Fernandes JF, Abossolo BP, Methogo BG, et al. A phase 3 trial of RTS,S/AS01 malaria vaccine in African infants. N Engl J Med. (2012) 367:2284–95. doi: 10.1056/NEJMoa1208394
79. Kaslow DC, Biernaux S. RTS,S: toward a first landmark on the malaria vaccine technology roadmap. Vaccine. (2015) 33:7425–32. doi: 10.1016/j.vaccine.2015.09.061
80. Neafsey DE, Juraska M, Bedford T, Benkeser D, Valim C, Griggs A, et al. Genetic diversity and protective efficacy of the RTS,S/AS01 malaria vaccine. N Engl J Med. (2015) 373:2025–37. doi: 10.1056/NEJMoa1505819
81. Rts SCTP. Efficacy and safety of RTS,S/AS01 malaria vaccine with or without a booster dose in infants and children in Africa: final results of a phase 3, individually randomised, controlled trial. Lancet. (2015) 386:31–45. doi: 10.1016/S0140-6736(15)60721-8
82. Kumar KA, Sano G, Boscardin S, Nussenzweig RS, Nussenzweig MC, Zavala F, et al. The circumsporozoite protein is an immunodominant protective antigen in irradiated sporozoites. Nature. (2006) 444:937–40. doi: 10.1038/nature05361
83. Gruner AC, Mauduit M, Tewari R, Romero JF, Depinay N, Kayibanda M, et al. Sterile protection against malaria is independent of immune responses to the circumsporozoite protein. PLoS ONE. (2007) 2:e1371. doi: 10.1371/journal.pone.0001371
84. Mauduit M, Gruner AC, Tewari R, Depinay N, Kayibanda M, Chavatte JM, et al. A role for immune responses against non-CS components in the cross-species protection induced by immunization with irradiated malaria sporozoites. PLoS ONE. (2009) 4:e7717. doi: 10.1371/journal.pone.0007717
85. Mauduit M, Tewari R, Depinay N, Kayibanda M, Lallemand E, Chavatte JM, et al. Minimal role for the circumsporozoite protein in the induction of sterile immunity by vaccination with live rodent malaria sporozoites. Infect Immun. (2010) 78:2182–8. doi: 10.1128/IAI.01415-09
86. Tarun AS, Peng X, Dumpit RF, Ogata Y, Silva-Rivera H, Camargo N, et al. A combined transcriptome and proteome survey of malaria parasite liver stages. Proc Natl Acad Sci USA. (2008) 105:305–10. doi: 10.1073/pnas.0710780104
87. Trieu A, Kayala MA, Burk C, Molina DM, Freilich DA, Richie TL, et al. Sterile protective immunity to malaria is associated with a panel of novel P. falciparum antigens. Mol Cell Proteomics. (2011) 10:M111.007948. doi: 10.1074/mcp.M111.007948
88. Duffy PE, Sahu T, Akue A, Milman N, Anderson C. Pre-erythrocytic malaria vaccines: identifying the targets. Expert Rev Vaccines. (2012) 11:1261–80. doi: 10.1586/erv.12.92
89. Peng K, Goh YS, Siau A, Franetich JF, Chia WN, Ong AS, et al. Breadth of humoral response and antigenic targets of sporozoite-inhibitory antibodies associated with sterile protection induced by controlled human malaria infection. Cell Microbiol. (2016) 18:1739–50. doi: 10.1111/cmi.12608
90. Schussek S, Trieu A, Apte SH, Sidney J, Sette A, Doolan DL. Novel Plasmodium antigens identified via genome-based antibody screen induce protection associated with polyfunctional T cell responses. Sci Rep. (2017) 7:15053. doi: 10.1038/s41598-017-15354-0
91. Wang R, Charoenvit Y, Corradin G, De La Vega P, Franke ED, Hoffman SL. Protection against malaria by Plasmodium yoelii sporozoite surface protein 2 linear peptide induction of CD4+ T cell- and IFN-gamma-dependent elimination of infected hepatocytes. J Immunol. (1996) 157:4061–7.
92. Daubersies P, Thomas AW, Millet P, Brahimi K, Langermans JA, Ollomo B, et al. Protection against Plasmodium falciparum malaria in chimpanzees by immunization with the conserved pre-erythrocytic liver-stage antigen 3. Nat Med. (2000) 6:1258–63. doi: 10.1038/81366
93. Speake C, Pichugin A, Sahu T, Malkov V, Morrison R, Pei Y, et al. Identification of novel pre-erythrocytic malaria antigen candidates for combination vaccines with circumsporozoite protein. PLoS ONE. (2016) 11:e0159449. doi: 10.1371/journal.pone.0159449
94. Bruder JT, Chen P, Ekberg G, Smith EC, Lazarski CA, Myers BA, et al. Profiling the targets of protective CD8(+) T cell responses to infection. Mol Ther Methods Clin Dev. (2017) 7:20–31. doi: 10.1016/j.omtm.2017.08.003
95. Cummings JF, Spring MD, Schwenk RJ, Ockenhouse CF, Kester KE, Polhemus ME, et al. Recombinant liver stage antigen-1 (LSA-1) formulated with AS01 or AS02 is safe, elicits high titer antibody and induces IFN-gamma/IL-2 CD4+ T cells but does not protect against experimental Plasmodium falciparum infection. Vaccine. (2010) 28:5135–44. doi: 10.1016/j.vaccine.2009.08.046
96. Ulmer JB, Donnelly JJ, Parker SE, Rhodes GH, Felgner PL, Dwarki VJ, et al. Heterologous protection against influenza by injection of DNA encoding a viral protein. Science. (1993) 259:1745–9. doi: 10.1126/science.8456302
97. Wang R, Doolan DL, Le TP, Hedstrom RC, Coonan KM, Charoenvit Y, et al. Induction of antigen-specific cytotoxic T lymphocytes in humans by a malaria DNA vaccine. Science. (1998) 282:476–80. doi: 10.1126/science.282.5388.476
98. Le TP, Coonan KM, Hedstrom RC, Charoenvit Y, Sedegah M, Epstein JE, et al. Safety, tolerability and humoral immune responses after intramuscular administration of a malaria DNA vaccine to healthy adult volunteers. Vaccine. (2000) 18:1893–901. doi: 10.1016/S0264-410X(99)00407-7
99. Wang R, Epstein J, Baraceros FM, Gorak EJ, Charoenvit Y, Carucci DJ, et al. Induction of CD4(+) T cell-dependent CD8(+) type 1 responses in humans by a malaria DNA vaccine. Proc Natl Acad Sci USA. (2001) 98:10817–22. doi: 10.1073/pnas.181123498
100. Hedstrom RC, Sedegah M, Hoffman SL. Prospects and strategies for development of DNA vaccines against malaria. Res Immunol. (1994) 145:476–83. doi: 10.1016/S0923-2494(94)80179-7
101. Vuola JM, Keating S, Webster DP, Berthoud T, Dunachie S, Gilbert SC, et al. Differential immunogenicity of various heterologous prime-boost vaccine regimens using DNA and viral vectors in healthy volunteers. J Immunol. (2005) 174:449–55. doi: 10.4049/jimmunol.174.1.449
102. Wang R, Doolan DL, Charoenvit Y, Hedstrom RC, Gardner MJ, Hobart P, et al. Simultaneous induction of multiple antigen-specific cytotoxic T lymphocytes in nonhuman primates by immunization with a mixture of four Plasmodium falciparum DNA plasmids. Infect Immun. (1998) 66:4193–202.
103. Haddad D, Bilcikova E, Witney AA, Carlton JM, White CE, Blair PL, et al. Novel antigen identification method for discovery of protective malaria antigens by rapid testing of DNA vaccines encoding exons from the parasite genome. Infect Immun. (2004) 72:1594–602. doi: 10.1128/IAI.72.3.1594-1602.2004
104. McConkey SJ, Reece WH, Moorthy VS, Webster D, Dunachie S, Butcher G, et al. Enhanced T-cell immunogenicity of plasmid DNA vaccines boosted by recombinant modified vaccinia virus Ankara in humans. Nat Med. (2003) 9:729–35. doi: 10.1038/nm881
105. Richie TL, Charoenvit Y, Wang R, Epstein JE, Hedstrom RC, Kumar S, et al. Clinical trial in healthy malaria-naive adults to evaluate the safety, tolerability, immunogenicity and efficacy of MuStDO5, a five-gene, sporozoite/hepatic stage Plasmodium falciparum DNA vaccine combined with escalating dose human GM-CSF DNA. Hum Vaccin Immunother. (2012) 8:1564–84. doi: 10.4161/hv.22129
106. Weiss WR, Ishii KJ, Hedstrom RC, Sedegah M, Ichino M, Barnhart K, et al. A plasmid encoding murine granulocyte-macrophage colony-stimulating factor increases protection conferred by a malaria DNA vaccine. J Immunol. (1998) 161:2325–32.
107. Iwasaki A, Medzhitov R. Control of adaptive immunity by the innate immune system. Nat Immunol. (2015) 16:343–53. doi: 10.1038/ni.3123
108. Cabral-Miranda G, Heath MD, Gomes AC, Mohsen MO, Montoya-Diaz E, Salman AM, et al. Microcrystalline tyrosine (MCT®): a depot adjuvant in licensed allergy immunotherapy offers new opportunities in malaria. Vaccines. (2017) 5:E32. doi: 10.3390/vaccines5040032
109. Ewer KJ, Sierra-Davidson K, Salman AM, Illingworth JJ, Draper SJ, Biswas S, et al. Progress with viral vectored malaria vaccines: a multi-stage approach involving “unnatural immunity”. Vaccine. (2015) 33:7444–51. doi: 10.1016/j.vaccine.2015.09.094
110. Rodrigues M, Li S, Murata K, Rodriguez D, Rodriguez JR, Bacik I, et al. Influenza and vaccinia viruses expressing malaria CD8+ T and B cell epitopes. Comparison of their immunogenicity and capacity to induce protective immunity. J Immunol. (1994) 153:4636–48.
111. Tine JA, Lanar DE, Smith DM, Wellde BT, Schultheiss P, Ware LA, et al. NYVAC-Pf7: a poxvirus-vectored, multiantigen, multistage vaccine candidate for Plasmodium falciparum malaria. Infect Immun. (1996) 64:3833–44.
112. Tsuji M, Bergmann CC, Takita-Sonoda Y, Murata K, Rodrigues EG, Nussenzweig RS, et al. Recombinant Sindbis viruses expressing a cytotoxic T-lymphocyte epitope of a malaria parasite or of influenza virus elicit protection against the corresponding pathogen in mice. J Virol. (1998) 72:6907–10.
113. Bonaldo MC, Garratt RC, Caufour PS, Freire MS, Rodrigues MM, Nussenzweig RS, et al. Surface expression of an immunodominant malaria protein B cell epitope by yellow fever virus. J Mol Biol. (2002) 315:873–85. doi: 10.1006/jmbi.2001.5258
114. Stoyanov CT, Boscardin SB, Deroubaix S, Barba-Spaeth G, Franco D, Nussenzweig RS, et al. Immunogenicity and protective efficacy of a recombinant yellow fever vaccine against the murine malarial parasite Plasmodium yoelii. Vaccine. (2010) 28:4644–52. doi: 10.1016/j.vaccine.2010.04.071
115. Hollingdale MR, Sedegah M, Limbach K. Development of replication-deficient adenovirus malaria vaccines. Expert Rev Vaccines. (2017) 16:261–71. doi: 10.1080/14760584.2016.1228454
116. Hansen SG, Womack J, Scholz I, Renner A, Edgel KA, Xu G, et al. Cytomegalovirus vectors expressing Plasmodium knowlesi antigens induce immune responses that delay parasitemia upon sporozoite challenge. PLoS ONE. (2019) 14:e0210252. doi: 10.1371/journal.pone.0210252
117. Sadoff JC, Ballou WR, Baron LS, Majarian WR, Brey RN, Hockmeyer WT, et al. Oral Salmonella typhimurium vaccine expressing circumsporozoite protein protects against malaria. Science. (1988) 240:336–8. doi: 10.1126/science.3281260
118. Aggarwal A, Kumar S, Jaffe R, Hone D, Gross M, Sadoff J. Oral Salmonella: malaria circumsporozoite recombinants induce specific CD8+ cytotoxic T cells. J Exp Med. (1990) 172:1083–90. doi: 10.1084/jem.172.4.1083
119. Arama C, Assefaw-Redda Y, Rodriguez A, Fernandez C, Corradin G, Kaufmann SH, et al. Heterologous prime-boost regimen adenovector 35-circumsporozoite protein vaccine/recombinant bacillus calmette-guerin expressing the Plasmodium falciparum circumsporozoite induces enhanced long-term memory immunity in BALB/c mice. Vaccine. (2012) 30:4040–5. doi: 10.1016/j.vaccine.2012.04.029
120. Bergmann-Leitner ES, Hosie H, Trichilo J, Deriso E, Ranallo RT, Alefantis T, et al. Self-adjuvanting bacterial vectors expressing pre-erythrocytic antigens induce sterile protection against malaria. Front Immunol. (2013) 4:176. doi: 10.3389/fimmu.2013.00176
121. Gonzalez C, Hone D, Noriega FR, Tacket CO, Davis JR, Losonsky G, et al. Salmonella typhi vaccine strain CVD 908 expressing the circumsporozoite protein of Plasmodium falciparum: strain construction and safety and immunogenicity in humans. J Infect Dis. (1994) 169:927–31. doi: 10.1093/infdis/169.4.927
122. Wang HH, Rogers WO, Kang YH, Sedegah M, Hoffman SL. Partial protection against malaria by immunization with Leishmania enriettii expressing the Plasmodium yoelii circumsporozoite protein. Mol Biochem Parasitol. (1995) 69:139–48. doi: 10.1016/0166-6851(94)00159-K
123. Di Cristina M, Ghouze F, Kocken CH, Naitza S, Cellini P, Soldati D, et al. Transformed Toxoplasma gondii tachyzoites expressing the circumsporozoite protein of Plasmodium knowlesi elicit a specific immune response in rhesus monkeys. Infect Immun. (1999) 67:1677–82.
124. Charest H, Sedegah M, Yap GS, Gazzinelli RT, Caspar P, Hoffman SL, et al. Recombinant attenuated Toxoplasma gondii expressing the Plasmodium yoelii circumsporozoite protein provides highly effective priming for CD8+ T cell-dependent protective immunity against malaria. J Immunol. (2000) 165:2084–92. doi: 10.4049/jimmunol.165.4.2084
125. Sedegah M, Jones TR, Kaur M, Hedstrom R, Hobart P, Tine JA, et al. Boosting with recombinant vaccinia increases immunogenicity and protective efficacy of malaria DNA vaccine. Proc Natl Acad Sci USA. (1998) 95:7648–53. doi: 10.1073/pnas.95.13.7648
126. Bruna-Romero O, Hafalla JC, Gonzalez-Aseguinolaza G, Sano G, Tsuji M, Zavala F. Detection of malaria liver-stages in mice infected through the bite of a single Anopheles mosquito using a highly sensitive real-time PCR. Int J Parasitol. (2001) 31:1499–502. doi: 10.1016/S0020-7519(01)00265-X
127. Hill AV, Reyes-Sandoval A, O'hara G, Ewer K, Lawrie A, Goodman A, et al. Prime-boost vectored malaria vaccines: progress and prospects. Hum Vaccin. (2010) 6:78–83. doi: 10.4161/hv.6.1.10116
128. Bliss CM, Bowyer G, Anagnostou NA, Havelock T, Snudden CM, Davies H, et al. Assessment of novel vaccination regimens using viral vectored liver stage malaria vaccines encoding ME-TRAP. Sci Rep. (2018) 8:3390. doi: 10.1038/s41598-018-21630-4
129. de Camargo TM, De Freitas EO, Gimenez AM, Lima LC, De Almeida Caramico K, Francoso KS, et al. Prime-boost vaccination with recombinant protein and adenovirus-vector expressing Plasmodium vivax circumsporozoite protein (CSP) partially protects mice against Pb/Pv sporozoite challenge. Sci Rep. (2018) 8:1118. doi: 10.1038/s41598-017-19063-6
130. Webster DP, Dunachie S, Vuola JM, Berthoud T, Keating S, Laidlaw SM, et al. Enhanced T cell-mediated protection against malaria in human challenges by using the recombinant poxviruses FP9 and modified vaccinia virus Ankara. Proc Natl Acad Sci USA. (2005) 102:4836–41. doi: 10.1073/pnas.0406381102
131. Hodgson SH, Juma E, Salim A, Magiri C, Njenga D, Molyneux S, et al. Lessons learnt from the first controlled human malaria infection study conducted in Nairobi, Kenya. Malar J. (2015) 14:182. doi: 10.1186/s12936-015-0671-x
132. Ogwang C, Kimani D, Edwards NJ, Roberts R, Mwacharo J, Bowyer G, et al. Prime-boost vaccination with chimpanzee adenovirus and modified vaccinia Ankara encoding TRAP provides partial protection against Plasmodium falciparum infection in Kenyan adults. Sci Transl Med. (2015) 7:286re285. doi: 10.1126/scitranslmed.aaa2373
133. Rampling T, Ewer KJ, Bowyer G, Bliss CM, Edwards NJ, Wright D, et al. Safety and high level efficacy of the combination malaria vaccine regimen of RTS,S/AS01B with chimpanzee adenovirus 63 and modified vaccinia ankara vectored vaccines expressing ME-TRAP. J Infect Dis. (2016) 214:772–81. doi: 10.1093/infdis/jiw244
134. Gola A, Silman D, Walters AA, Sridhar S, Uderhardt S, Salman AM, et al. Prime and target immunization protects against liver-stage malaria in mice. Sci Transl Med. (2018) 10:eaap9128. doi: 10.1126/scitranslmed.aap9128
135. Olsen TM, Stone BC, Chuenchob V, Murphy SC. Prime-and-trap malaria vaccination to generate protective CD8(+) liver-resident memory T cells. J Immunol. (2018) 201:1984–93. doi: 10.4049/jimmunol.1800740
136. Mazier D, Beaudoin RL, Mellouk S, Druilhe P, Texier B, Trosper J, et al. Complete development of hepatic stages of Plasmodium falciparum in vitro. Science. (1985) 227:440–2. doi: 10.1126/science.3880923
137. Purcell LA, Wong KA, Yanow SK, Lee M, Spithill TW, Rodriguez A. Chemically attenuated Plasmodium sporozoites induce specific immune responses, sterile immunity and cross-protection against heterologous challenge. Vaccine. (2008) 26:4880–4. doi: 10.1016/j.vaccine.2008.07.017
138. Purcell LA, Yanow SK, Lee M, Spithill TW, Rodriguez A. Chemical attenuation of Plasmodium berghei sporozoites induces sterile immunity in mice. Infect Immun. (2008) 76:1193–9. doi: 10.1128/IAI.01399-07
139. Raja AI, Stanisic DI, Good MF. Chemical attenuation in the development of a whole-organism malaria vaccine. Infect Immun. (2017) 85:e00062–17. doi: 10.1128/IAI.00062-17
140. Tewari R, Spaccapelo R, Bistoni F, Holder AA, Crisanti A. Function of region I and II adhesive motifs of Plasmodium falciparum circumsporozoite protein in sporozoite motility and infectivity. J Biol Chem. (2002) 277:47613–8. doi: 10.1074/jbc.M208453200
141. Espinosa DA, Yadava A, Angov E, Maurizio PL, Ockenhouse CF, Zavala F. Development of a chimeric Plasmodium berghei strain expressing the repeat region of the P. vivax circumsporozoite protein for in vivo evaluation of vaccine efficacy. Infect Immun. (2013) 81:2882–7. doi: 10.1128/IAI.00461-13
142. Kastenmuller K, Espinosa DA, Trager L, Stoyanov C, Salazar AM, Pokalwar S, et al. Full-length Plasmodium falciparum circumsporozoite protein administered with long-chain poly(I.C) or the Toll-like receptor 4 agonist glucopyranosyl lipid adjuvant-stable emulsion elicits potent antibody and CD4+ T cell immunity and protection in mice. Infect Immun. (2013) 81:789–800. doi: 10.1128/IAI.01108-12
143. Bauza K, Malinauskas T, Pfander C, Anar B, Jones EY, Billker O, et al. Efficacy of a Plasmodium vivax malaria vaccine using ChAd63 and modified vaccinia Ankara expressing thrombospondin-related anonymous protein as assessed with transgenic Plasmodium berghei parasites. Infect Immun. (2014) 82:1277–86. doi: 10.1128/IAI.01187-13
144. Deal C, Balazs AB, Espinosa DA, Zavala F, Baltimore D, Ketner G. Vectored antibody gene delivery protects against Plasmodium falciparum sporozoite challenge in mice. Proc Natl Acad Sci USA. (2014) 111:12528–32. doi: 10.1073/pnas.1407362111
145. Espinosa DA, Gutierrez GM, Rojas-Lopez M, Noe AR, Shi L, Tse SW, et al. Proteolytic cleavage of the Plasmodium falciparum circumsporozoite protein is a target of protective antibodies. J Infect Dis. (2015) 212:1111–9. doi: 10.1093/infdis/jiv154
146. Kisalu NK, Idris AH, Weidle C, Flores-Garcia Y, Flynn BJ, Sack BK, et al. A human monoclonal antibody prevents malaria infection by targeting a new site of vulnerability on the parasite. Nat Med. (2018) 24:408–16. doi: 10.1038/nm.4512
147. Carlton JM, Angiuoli SV, Suh BB, Kooij TW, Pertea M, Silva JC, et al. Genome sequence and comparative analysis of the model rodent malaria parasite Plasmodium yoelii yoelii. Nature. (2002) 419:512–9. doi: 10.1038/nature01099
148. Hall N, Karras M, Raine JD, Carlton JM, Kooij TW, Berriman M, et al. A comprehensive survey of the Plasmodium life cycle by genomic, transcriptomic, and proteomic analyses. Science. (2005) 307:82–6. doi: 10.1126/science.1103717
149. Vaughan AM, Mikolajczak SA, Wilson EM, Grompe M, Kaushansky A, Camargo N, et al. Complete Plasmodium falciparum liver-stage development in liver-chimeric mice. J Clin Invest. (2012) 122:3618–28. doi: 10.1172/JCI62684
150. Krotoski WA, Collins WE, Bray RS, Garnham PC, Cogswell FB, Gwadz RW, et al. Demonstration of hypnozoites in sporozoite-transmitted Plasmodium vivax infection. Am J Trop Med Hyg. (1982) 31:1291–3. doi: 10.4269/ajtmh.1982.31.1291
151. Landau I, Chabaud AG, Adam JP, Michel JC, Boulard Y. Morphologie et evolution des schizontes hépatiques secondaires dans le paludisme des rongeurs de la Maboké. Cahiers de la Maboké. (1968) 6:63–72.
152. Beura LK, Hamilton SE, Bi K, Schenkel JM, Odumade OA, Casey KA, et al. Normalizing the environment recapitulates adult human immune traits in laboratory mice. Nature. (2016) 532:512–6. doi: 10.1038/nature17655
153. Beignon AS, Le Grand R, Chapon C. In vivo imaging in NHP models of malaria: challenges, progress and outlooks. Parasitol Int. (2014) 63:206–15. doi: 10.1016/j.parint.2013.09.001
154. Herrera S, Perlaza BL, Bonelo A, Arevalo-Herrera M. Aotus monkeys: their great value for anti-malaria vaccines and drug testing. Int J Parasitol. (2002) 32:1625–35. doi: 10.1016/S0020-7519(02)00191-1
155. Collins WE, Sullivan JS, Williams A, Nace D, Williams T, Galland GG, et al. Aotus nancymaae as a potential model for the testing of anti-sporozoite and liver stage vaccines against Plasmodium falciparum. Am J Trop Med Hyg. (2006) 74:422–4. doi: 10.4269/ajtmh.2006.74.422
156. Joyner C, Barnwell JW, Galinski MR. No more monkeying around: primate malaria model systems are key to understanding Plasmodium vivax liver-stage biology, hypnozoites, and relapses. Front Microbiol. (2015) 6:145. doi: 10.3389/fmicb.2015.00145
157. Deans JA. Protective antigens of bloodstage Plasmodium knowlesi parasites. Philos Trans R Soc Lond B Biol Sci. (1984) 307:159–69. doi: 10.1098/rstb.1984.0116
158. Wijayalath WA, Cheesman S, Rajakaruna J, Handunnetti SM, Carter R, Pathirana PP. Evidence for strain-specific protective immunity against blood-stage parasites of Plasmodium cynomolgi in toque monkey. Parasite Immunol. (2008) 30:630–6. doi: 10.1111/j.1365-3024.2008.01065.x
159. Wijayalath W, Cheesman S, Tanabe K, Handunnetti S, Carter R, Pathirana S. Strain-specific protective effect of the immunity induced by live malarial sporozoites under chloroquine cover. PLoS ONE. (2012) 7:e45861. doi: 10.1371/journal.pone.0045861
160. Good MF, Hawkes MT, Yanow SK. Humanized mouse models to study cell-mediated immune responses to liver-stage malaria vaccines. Trends Parasitol. (2015) 31:583–94. doi: 10.1016/j.pt.2015.06.008
161. Minkah NK, Schafer C, Kappe SHI. Humanized mouse models for the study of human malaria parasite biology, pathogenesis, and immunity. Front Immunol. (2018) 9:807. doi: 10.3389/fimmu.2018.00807
162. Morosan S, Hez-Deroubaix S, Lunel F, Renia L, Giannini C, Van Rooijen N, et al. Liver-stage development of Plasmodium falciparum, in a humanized mouse model. J Infect Dis. (2006) 193:996–1004. doi: 10.1086/500840
163. Sacci JB Jr, Alam U, Douglas D, Lewis J, Tyrrell DL, Azad AF, et al. Plasmodium falciparum infection and exoerythrocytic development in mice with chimeric human livers. Int J Parasitol. (2006) 36:353–60. doi: 10.1016/j.ijpara.2005.10.014
164. Mikolajczak SA, Sacci JB Jr, de La Vega P, Camargo N, Vanbuskirk K, Krzych U, et al. Disruption of the Plasmodium falciparum liver-stage antigen-1 locus causes a differentiation defect in late liver-stage parasites. Cell Microbiol. (2011) 13:1250–60. doi: 10.1111/j.1462-5822.2011.01617.x
165. Brezillon NM, Dasilva L, L'hote D, Bernex F, Piquet J, Binart N, et al. Rescue of fertility in homozygous mice for the urokinase plasminogen activator transgene by the transplantation of mouse hepatocytes. Cell Transplant. (2008) 17:803–12. doi: 10.3727/096368908786516800
166. Heckel JL, Sandgren EP, Degen JL, Palmiter RD, Brinster RL. Neonatal bleeding in transgenic mice expressing urokinase-type plasminogen activator. Cell. (1990) 62:447–56. doi: 10.1016/0092-8674(90)90010-C
167. Mikolajczak SA, Lakshmanan V, Fishbaugher M, Camargo N, Harupa A, Kaushansky A, et al. A next-generation genetically attenuated Plasmodium falciparum parasite created by triple gene deletion. Mol Ther. (2014) 22:1707–15. doi: 10.1038/mt.2014.85
168. Hasegawa M, Kawai K, Mitsui T, Taniguchi K, Monnai M, Wakui M, et al. The reconstituted 'humanized liver' in TK-NOG mice is mature and functional. Biochem Biophys Res Commun. (2011) 405:405–10. doi: 10.1016/j.bbrc.2011.01.042
169. Soulard V, Bosson-Vanga H, Lorthiois A, Roucher C, Franetich JF, Zanghi G, et al. Plasmodium falciparum full life cycle and Plasmodium ovale liver stages in humanized mice. Nat Commun. (2015) 6:7690. doi: 10.1038/ncomms8690
170. Foquet L, Hermsen CC, Van Gemert GJ, Van Braeckel E, Weening KE, Sauerwein R, et al. Vaccine-induced monoclonal antibodies targeting circumsporozoite protein prevent Plasmodium falciparum infection. J Clin Invest. (2014) 124:140–4. doi: 10.1172/JCI70349
171. Sack B, Kappe SH, Sather DN. Towards functional antibody-based vaccines to prevent pre-erythrocytic malaria infection. Expert Rev Vaccines. (2017) 16:403–14. doi: 10.1080/14760584.2017.1295853
172. Foquet L, Schafer C, Minkah NK, Alanine DGW, Flannery EL, Steel RWJ, et al. Plasmodium falciparum liver stage infection and transition to stable blood stage infection in liver-humanized and blood-humanized FRGN KO mice enables testing of blood stage inhibitory antibodies (reticulocyte-binding protein homolog 5) in vivo. Front Immunol. (2018) 9:524. doi: 10.3389/fimmu.2018.00524
173. Walsh NC, Kenney LL, Jangalwe S, Aryee KE, Greiner DL, Brehm MA, et al. Humanized mouse models of clinical disease. Annu Rev Pathol. (2017) 12:187–215. doi: 10.1146/annurev-pathol-052016-100332
174. Huang J, Li X, Coelho-Dos-Reis JG, Wilson JM, Tsuji M. An AAV vector-mediated gene delivery approach facilitates reconstitution of functional human CD8+ T cells in mice. PLoS ONE. (2014) 9:e88205. doi: 10.1371/journal.pone.0088205
175. Huang J, Li X, Coelho-Dos-Reis JG, Zhang M, Mitchell R, Nogueira RT, et al. Human immune system mice immunized with Plasmodium falciparum circumsporozoite protein induce protective human humoral immunity against malaria. J Immunol Methods. (2015) 427:42–50. doi: 10.1016/j.jim.2015.09.005
176. Dunachie SJ, Walther M, Vuola JM, Webster DP, Keating SM, Berthoud T, et al. A clinical trial of prime-boost immunisation with the candidate malaria vaccines RTS,S/AS02A and MVA-CS. Vaccine. (2006) 24:2850–9. doi: 10.1016/j.vaccine.2005.12.041
177. Schofield L, Villaquiran J, Ferreira A, Schellekens H, Nussenzweig R, Nussenzweig V. Gamma interferon, CD8+ T cells and antibodies required for immunity to malaria sporozoites. Nature. (1987) 330:664–6. doi: 10.1038/330664a0
178. Weiss WR, Sedegah M, Beaudoin RL, Miller LH, Good MF. CD8+ T cells (cytotoxic/suppressors) are required for protection in mice immunized with malaria sporozoites. Proc Natl Acad Sci USA. (1988) 85:573–6. doi: 10.1073/pnas.85.2.573
179. Liehl P, Zuzarte-Luis V, Chan J, Zillinger T, Baptista F, Carapau D, et al. Host-cell sensors for Plasmodium activate innate immunity against liver-stage infection. Nat Med. (2014) 20:47–53. doi: 10.1038/nm.3424
180. Nussler AK, Renia L, Pasquetto V, Miltgen F, Matile H, Mazier D. In vivo induction of the nitric oxide pathway in hepatocytes after injection with irradiated malaria sporozoites, malaria blood parasites or adjuvants. Eur J Immunol. (1993) 23:882–7. doi: 10.1002/eji.1830230417
181. Mellouk S, Hoffman SL, Liu ZZ, De La Vega P, Billiar TR, Nussler AK. Nitric oxide-mediated antiplasmodial activity in human and murine hepatocytes induced by gamma interferon and the parasite itself: enhancement by exogenous tetrahydrobiopterin. Infect Immun. (1994) 62:4043–6.
182. Seguin MC, Klotz FW, Schneider I, Weir JP, Goodbary M, Slayter M, et al. Induction of nitric oxide synthase protects against malaria in mice exposed to irradiated Plasmodium berghei infected mosquitoes: involvement of interferon gamma and CD8+ T cells. J Exp Med. (1994) 180:353–8. doi: 10.1084/jem.180.1.353
183. Muniz-Junqueira MI, Dos Santos-Neto LL, Tosta CE. Influence of tumor necrosis factor-alpha on the ability of monocytes and lymphocytes to destroy intraerythrocytic Plasmodium falciparum in vitro. Cell Immunol. (2001) 208:73–9. doi: 10.1006/cimm.2001.1770
184. Butler NS, Schmidt NW, Harty JT. Differential effector pathways regulate memory CD8 T cell immunity against Plasmodium berghei versus P. yoelii sporozoites. J Immunol. (2010) 184:2528–38. doi: 10.4049/jimmunol.0903529
185. Roland J, Soulard V, Sellier C, Drapier AM, Di Santo JP, Cazenave PA, et al. NK cell responses to Plasmodium infection and control of intrahepatic parasite development. J Immunol. (2006) 177:1229–39. doi: 10.4049/jimmunol.177.2.1229
186. Ing R, Stevenson MM. Dendritic cell and NK cell reciprocal cross talk promotes gamma interferon-dependent immunity to blood-stage Plasmodium chabaudi AS infection in mice. Infect Immun. (2009) 77:770–82. doi: 10.1128/IAI.00994-08
187. Ryg-Cornejo V, Nie CQ, Bernard NJ, Lundie RJ, Evans KJ, Crabb BS, et al. NK cells and conventional dendritic cells engage in reciprocal activation for the induction of inflammatory responses during Plasmodium berghei ANKA infection. Immunobiology. (2013) 218:263–71. doi: 10.1016/j.imbio.2012.05.018
188. D'Ombrain MC, Hansen DS, Simpson KM, Schofield L. Gammadelta-T cells expressing NK receptors predominate over NK cells and conventional T cells in the innate IFN-gamma response to Plasmodium falciparum malaria. Eur J Immunol. (2007) 37:1864–73. doi: 10.1002/eji.200636889
189. D'Ombrain MC, Robinson LJ, Stanisic DI, Taraika J, Bernard N, Michon P, et al. Association of early interferon-gamma production with immunity to clinical malaria: a longitudinal study among Papua New Guinean children. Clin Infect Dis. (2008) 47:1380–7. doi: 10.1086/592971
190. Tsuji M, Mombaerts P, Lefrancois L, Nussenzweig RS, Zavala F, Tonegawa S. Gamma delta T cells contribute to immunity against the liver stages of malaria in alpha beta T-cell-deficient mice. Proc Natl Acad Sci USA. (1994) 91:345–9. doi: 10.1073/pnas.91.1.345
191. McKenna KC, Tsuji M, Sarzotti M, Sacci JB Jr, Witney AA, Azad AF. Gammadelta T cells are a component of early immunity against preerythrocytic malaria parasites. Infect Immun. (2000) 68:2224–30. doi: 10.1128/IAI.68.4.2224-2230.2000
192. Brandes M, Willimann K, Moser B. Professional antigen-presentation function by human gammadelta T Cells. Science. (2005) 309:264–8. doi: 10.1126/science.1110267
193. Brandes M, Willimann K, Bioley G, Levy N, Eberl M, Luo M, et al. Cross-presenting human gammadelta T cells induce robust CD8+ alphabeta T cell responses. Proc Natl Acad Sci USA. (2009) 106:2307–12. doi: 10.1073/pnas.0810059106
194. Zaidi I, Diallo H, Conteh S, Robbins Y, Kolasny J, Orr-Gonzalez S, et al. Gammadelta T cells are required for the induction of sterile immunity during irradiated sporozoite vaccinations. J Immunol. (2017) 199:3781–8. doi: 10.4049/jimmunol.1700314
195. Amino R, Thiberge S, Martin B, Celli S, Shorte S, Frischknecht F, et al. Quantitative imaging of Plasmodium transmission from mosquito to mammal. Nat Med. (2006) 12:220–4. doi: 10.1038/nm1350
196. Chakravarty S, Cockburn IA, Kuk S, Overstreet MG, Sacci JB, Zavala F. CD8+ T lymphocytes protective against malaria liver stages are primed in skin-draining lymph nodes. Nat Med. (2007) 13:1035–41. doi: 10.1038/nm1628
197. Obeid M, Franetich JF, Lorthiois A, Gego A, Gruner AC, Tefit M, et al. Skin-draining lymph node priming is sufficient to induce sterile immunity against pre-erythrocytic malaria. EMBO Mol Med. (2013) 5:250–63. doi: 10.1002/emmm.201201677
198. Janeway CAJ, Travers P, Walport M, Shlomcik MJ. Immunobiology: The Immune System in Health and Disease. New York, NY: Garland Science (2001).
199. Renia L, Maranon C, Hosmalin A, Gruner AC, Silvie O, Snounou G. Do apoptotic Plasmodium-infected hepatocytes initiate protective immune responses? J Infect Dis. (2006) 193:163–4; author reply 164–5. doi: 10.1086/498536
200. Cockburn IA, Amino R, Kelemen RK, Kuo SC, Tse SW, Radtke A, et al. In vivo imaging of CD8+ T cell-mediated elimination of malaria liver stages. Proc Natl Acad Sci USA. (2013) 110:9090–5. doi: 10.1073/pnas.1303858110
201. Berenzon D, Schwenk RJ, Letellier L, Guebre-Xabier M, Williams J, Krzych U. Protracted protection to Plasmodium berghei malaria is linked to functionally and phenotypically heterogeneous liver memory CD8+ T cells. J Immunol. (2003) 171:2024–34. doi: 10.4049/jimmunol.171.4.2024
202. Tse SW, Cockburn IA, Zhang H, Scott AL, Zavala F. Unique transcriptional profile of liver-resident memory CD8+ T cells induced by immunization with malaria sporozoites. Genes Immun. (2013) 14:302–9. doi: 10.1038/gene.2013.20
203. Patel H, Yadav N, Parmar R, Patel S, Singh AP, Shrivastava N, et al. Frequent inoculations with radiation attenuated sporozoite is essential for inducing sterile protection that correlates with a threshold level of Plasmodia liver-stage specific CD8(+) T cells. Cell Immunol. (2017) 317:48–54. doi: 10.1016/j.cellimm.2017.05.001
204. Sacci JBJr, Hollingdale MR, Sedegah M. Cellular immune response to DNA and vaccinia prime-boost immunization kills Plasmodium yoelii-infected hepatocytes in vitro. Pathog Dis. (2017) 75. doi: 10.1093/femspd/ftx051
205. Carvalho LH, Sano G, Hafalla JC, Morrot A, Curotto De Lafaille MA, Zavala F. IL-4-secreting CD4+ T cells are crucial to the development of CD8+ T-cell responses against malaria liver stages. Nat Med. (2002) 8:166–70. doi: 10.1038/nm0202-166
206. Overstreet MG, Chen YC, Cockburn IA, Tse SW, Zavala F. CD4+ T cells modulate expansion and survival but not functional properties of effector and memory CD8+ T cells induced by malaria sporozoites. PLoS ONE. (2011) 6:e15948. doi: 10.1371/journal.pone.0015948
207. Vanderberg JP, Frevert U. Intravital microscopy demonstrating antibody-mediated immobilisation of Plasmodium berghei sporozoites injected into skin by mosquitoes. Int J Parasitol. (2004) 34:991–6. doi: 10.1016/j.ijpara.2004.05.005
208. Aliprandini E, Tavares J, Panatieri RH, Thiberge S, Yamamoto MM, Silvie O, et al. Cytotoxic anti-circumsporozoite antibodies target malaria sporozoites in the host skin. Nat Microbiol. (2018) 3:1224–33. doi: 10.1038/s41564-018-0254-z
209. Schwenk R, Lumsden JM, Rein LE, Juompan L, Kester KE, Heppner DG, et al. Immunization with the RTS,S/AS malaria vaccine induces IFN-gamma(+)CD4 T cells that recognize only discrete regions of the circumsporozoite protein and these specificities are maintained following booster immunizations and challenge. Vaccine. (2011) 29:8847–54. doi: 10.1016/j.vaccine.2011.09.098
210. Nudelman S, Renia L, Charoenvit Y, Yuan L, Miltgen F, Beaudoin RL, et al. Dual action of anti-sporozoite antibodies in vitro. J Immunol. (1989) 143:996–1000.
211. Renia L, Mattei D, Goma J, Pied S, Dubois P, Miltgen F, et al. A malaria heat-shock-like determinant expressed on the infected hepatocyte surface is the target of antibody-dependent cell-mediated cytotoxic mechanisms by nonparenchymal liver cells. Eur J Immunol. (1990) 20:1445–9. doi: 10.1002/eji.1830200706
212. Miller JL, Sack BK, Baldwin M, Vaughan AM, Kappe SHI. Interferon-mediated innate immune responses against malaria parasite liver stages. Cell Rep. (2014) 7:436–47. doi: 10.1016/j.celrep.2014.03.018
213. Mellouk S, Maheshwari RK, Rhodes-Feuillette A, Beaudoin RL, Berbiguier N, Matile H, et al. Inhibitory activity of interferons and interleukin 1 on the development of Plasmodium falciparum in human hepatocyte cultures. J Immunol. (1987) 139:4192–5.
214. Ferreira A, Schofield L, Enea V, Schellekens H, Van Der Meide P, Collins WE, et al. Inhibition of development of exoerythrocytic forms of malaria parasites by gamma-interferon. Science. (1986) 232:881–4. doi: 10.1126/science.3085218
215. Depinay N, Franetich JF, Gruner AC, Mauduit M, Chavatte JM, Luty AJ, et al. Inhibitory effect of TNF-alpha on malaria pre-erythrocytic stage development: influence of host hepatocyte/parasite combinations. PLoS ONE. (2011) 6:e17464. doi: 10.1371/journal.pone.0017464
216. Nussler A, Pied S, Goma J, Renia L, Miltgen F, Grau GE, et al. TNF inhibits malaria hepatic stages in vitro via synthesis of IL-6. Int Immunol. (1991) 3:317–21. doi: 10.1093/intimm/3.4.317
217. Nussler A, Drapier JC, Renia L, Pied S, Miltgen F, Gentilini M, et al. L-arginine-dependent destruction of intrahepatic malaria parasites in response to tumor necrosis factor and/or interleukin 6 stimulation. Eur J Immunol. (1991) 21:227–30. doi: 10.1002/eji.1830210134
218. Pied S, Renia L, Nussler A, Miltgen F, Mazier D. Inhibitory activity of IL-6 on malaria hepatic stages. Parasite Immunol. (1991) 13:211–7. doi: 10.1111/j.1365-3024.1991.tb00276.x
219. Liehl P, Meireles P, Albuquerque IS, Pinkevych M, Baptista F, Mota MM, et al. Innate immunity induced by Plasmodium liver infection inhibits malaria reinfections. Infect Immun. (2015) 83:1172–80. doi: 10.1128/IAI.02796-14
220. Uldrich AP, Le Nours J, Pellicci DG, Gherardin NA, Mcpherson KG, Lim RT, et al. CD1d-lipid antigen recognition by the gammadelta TCR. Nat Immunol. (2013) 14:1137–45. doi: 10.1038/ni.2713
221. Jagannathan P, Kim CC, Greenhouse B, Nankya F, Bowen K, Eccles-James I, et al. Loss and dysfunction of Vdelta2(+) gammadelta T cells are associated with clinical tolerance to malaria. Sci Transl Med. (2014) 6:251ra117. doi: 10.1126/scitranslmed.3009793
222. Jagannathan P, Lutwama F, Boyle MJ, Nankya F, Farrington LA, Mcintyre TI, et al. Vdelta2+ T cell response to malaria correlates with protection from infection but is attenuated with repeated exposure. Sci Rep. (2017) 7:11487. doi: 10.1038/s41598-017-10624-3
223. Radtke AJ, Kastenmuller W, Espinosa DA, Gerner MY, Tse SW, Sinnis P, et al. Lymph-node resident CD8alpha+ dendritic cells capture antigens from migratory malaria sporozoites and induce CD8+ T cell responses. PLoS Pathog. (2015) 11:e1004637. doi: 10.1371/journal.ppat.1004637
224. Jung S, Unutmaz D, Wong P, Sano G, De Los Santos K, Sparwasser T, et al. In vivo depletion of CD11c+ dendritic cells abrogates priming of CD8+ T cells by exogenous cell-associated antigens. Immunity. (2002) 17:211–20. doi: 10.1016/S1074-7613(02)00365-5
225. Jobe O, Donofrio G, Sun G, Liepinsh D, Schwenk R, Krzych U. Immunization with radiation-attenuated Plasmodium berghei sporozoites induces liver cCD8alpha+DC that activate CD8+T cells against liver-stage malaria. PLoS ONE. (2009) 4:e5075. doi: 10.1371/journal.pone.0005075
226. Lau LS, Fernandez-Ruiz D, Mollard V, Sturm A, Neller MA, Cozijnsen A, et al. CD8+ T cells from a novel T cell receptor transgenic mouse induce liver-stage immunity that can be boosted by blood-stage infection in rodent malaria. PLoS Pathog. (2014) 10:e1004135. doi: 10.1371/journal.ppat.1004135
227. Kurup SP, Anthony SM, Hancox LS, Vijay R, Pewe LL, Moioffer SJ, et al. Monocyte-derived CD11c(+) cells acquire plasmodium from hepatocytes to prime CD8 T cell immunity to liver-stage malaria. Cell Host Microbe. (2019) 25:565–77.e6. doi: 10.1016/j.chom.2019.02.014
228. Leiriao P, Mota MM, Rodriguez A. Apoptotic plasmodium-infected hepatocytes provide antigens to liver dendritic cells. J Infect Dis. (2005) 191:1576–81. doi: 10.1086/429635
229. Mauduit M, See P, Peng K, Renia L, Ginhoux F. Dendritic cells and the malaria pre-erythrocytic stage. Immunol Res. (2012) 53:115–26. doi: 10.1007/s12026-012-8269-7
230. Trimnell A, Takagi A, Gupta M, Richie TL, Kappe SH, Wang R. Genetically attenuated parasite vaccines induce contact-dependent CD8+ T cell killing of Plasmodium yoelii liver stage-infected hepatocytes. J Immunol. (2009) 183:5870–8. doi: 10.4049/jimmunol.0900302
231. Chakravarty S, Baldeviano GC, Overstreet MG, Zavala F. Effector CD8+ T lymphocytes against liver stages of Plasmodium yoelii do not require gamma interferon for antiparasite activity. Infect Immun. (2008) 76:3628–31. doi: 10.1128/IAI.00471-08
232. Hill AV, Elvin J, Willis AC, Aidoo M, Allsopp CE, Gotch FM, et al. Molecular analysis of the association of HLA-B53 and resistance to severe malaria. Nature. (1992) 360:434–9. doi: 10.1038/360434a0
233. Aidoo M, Lalvani A, Gilbert SC, Hu JT, Daubersies P, Hurt N, et al. Cytotoxic T-lymphocyte epitopes for HLA-B53 and other HLA types in the malaria vaccine candidate liver-stage antigen 3. Infect Immun. (2000) 68:227–32. doi: 10.1128/IAI.68.1.227-232.2000
234. Bonelo A, Valmori D, Triponez F, Tiercy JM, Mentha G, Oberholzer J, et al. Generation and characterization of malaria-specific human CD8(+) lymphocyte clones: effect of natural polymorphism on T cell recognition and endogenous cognate antigen presentation by liver cells. Eur J Immunol. (2000) 30:3079–88. doi: 10.1002/1521-4141(200011)30:11<3079::AID-IMMU3079>3.0.CO;2-7
235. Doolan DL, Southwood S, Freilich DA, Sidney J, Graber NL, Shatney L, et al. Identification of Plasmodium falciparum antigens by antigenic analysis of genomic and proteomic data. Proc Natl Acad Sci USA. (2003) 100:9952–7. doi: 10.1073/pnas.1633254100
236. Epstein JE, Tewari K, Lyke KE, Sim BK, Billingsley PF, Laurens MB, et al. Live attenuated malaria vaccine designed to protect through hepatic CD8(+) T cell immunity. Science. (2011) 334:475–80. doi: 10.1126/science.1211548
237. Reyes-Sandoval A, Wyllie DH, Bauza K, Milicic A, Forbes EK, Rollier CS, et al. CD8+ T effector memory cells protect against liver-stage malaria. J Immunol. (2011) 187:1347–57. doi: 10.4049/jimmunol.1100302
238. Schmidt NW, Podyminogin RL, Butler NS, Badovinac VP, Tucker BJ, Bahjat KS, et al. Memory CD8 T cell responses exceeding a large but definable threshold provide long-term immunity to malaria. Proc Natl Acad Sci USA. (2008) 105:14017–22. doi: 10.1073/pnas.0805452105
239. Holz LE, Prier JE, Freestone D, Steiner TM, English K, Johnson DN, et al. CD8(+) T cell activation leads to constitutive formation of liver tissue-resident memory T cells that seed a large and flexible niche in the liver. Cell Rep. (2018) 25:68–79 e64. doi: 10.1016/j.celrep.2018.08.094
240. McNamara HA, Cai Y, Wagle MV, Sontani Y, Roots CM, Miosge LA, et al. Up-regulation of LFA-1 allows liver-resident memory T cells to patrol and remain in the hepatic sinusoids. Sci Immunol. (2017) 2:eaaj1996. doi: 10.1126/sciimmunol.aaj1996
241. Fernandez-Ruiz D, Ng WY, Holz LE, Ma JZ, Zaid A, Wong YC, et al. Liver-resident memory CD8(+) T cells form a front-line defense against malaria liver-stage infection. Immunity. (2016) 45:889–902. doi: 10.1016/j.immuni.2016.08.011
242. Renia L, Marussig MS, Grillot D, Pied S, Corradin G, Miltgen F, et al. In vitro activity of CD4+ and CD8+ T lymphocytes from mice immunized with a synthetic malaria peptide. Proc Natl Acad Sci USA. (1991) 88:7963–7. doi: 10.1073/pnas.88.18.7963
243. Renia L, Grillot D, Marussig M, Corradin G, Miltgen F, Lambert PH, et al. Effector functions of circumsporozoite peptide-primed CD4+ T cell clones against Plasmodium yoelii liver stages. J Immunol. (1993) 150:1471–8.
244. Bongfen SE, Torgler R, Romero JF, Renia L, Corradin G. Plasmodium berghei-infected primary hepatocytes process and present the circumsporozoite protein to specific CD8+ T cells in vitro. J Immunol. (2007) 178:7054–63. doi: 10.4049/jimmunol.178.11.7054
245. Pichugin A, Steers N, De La Vega P, Zarling S, Chalom I, Krzych U. TAP-mediated processing of exoerythrocytic antigens is essential for protection induced with radiation-attenuated Plasmodium sporozoites. Eur J Immunol. (2016) 46:885–96. doi: 10.1002/eji.201545748
246. Takita-Sonoda Y, Tsuji M, Kamboj K, Nussenzweig RS, Clavijo P, Zavala F. Plasmodium yoelii: peptide immunization induces protective CD4+ T cells against a previously unrecognized cryptic epitope of the circumsporozoite protein. Exp Parasitol. (1996) 84:223–30. doi: 10.1006/expr.1996.0108
247. Weiss WR, Sedegah M, Berzofsky JA, Hoffman SL. The role of CD4+ T cells in immunity to malaria sporozoites. J Immunol. (1993) 151:2690–8.
248. Reece WH, Pinder M, Gothard PK, Milligan P, Bojang K, Doherty T, et al. A CD4(+) T-cell immune response to a conserved epitope in the circumsporozoite protein correlates with protection from natural Plasmodium falciparum infection and disease. Nat Med. (2004) 10:406–10. doi: 10.1038/nm1009
249. Schwenk R, Asher LV, Chalom I, Lanar D, Sun P, White K, et al. Opsonization by antigen-specific antibodies as a mechanism of protective immunity induced by Plasmodium falciparum circumsporozoite protein-based vaccine. Parasite Immunol. (2003) 25:17–25. doi: 10.1046/j.1365-3024.2003.00495.x
250. Flores-Garcia Y, Nasir G, Hopp CS, Munoz C, Balaban AE, Zavala F, et al. Antibody-mediated protection against Plasmodium sporozoites begins at the dermal inoculation site. MBio. (2018) 9:e02194–18. doi: 10.1128/mBio.02194-18
251. Sack BK, Keitany GJ, Vaughan AM, Miller JL, Wang R, Kappe SH. Mechanisms of stage-transcending protection following immunization of mice with late liver stage-arresting genetically attenuated malaria parasites. PLoS Pathog. (2015) 11:e1004855. doi: 10.1371/journal.ppat.1004855
252. Zenklusen I, Jongo S, Abdulla S, Ramadhani K, Lee Sim BK, Cardamone H, et al. Immunization of malaria-preexposed volunteers with PfSPZ vaccine elicits long-lived IgM invasion-inhibitory and complement-fixing antibodies. J Infect Dis. (2018) 217:1569–78. doi: 10.1093/infdis/jiy080
253. Finney OC, Keitany GJ, Smithers H, Kaushansky A, Kappe S, Wang R. Immunization with genetically attenuated P. falciparum parasites induces long-lived antibodies that efficiently block hepatocyte invasion by sporozoites. Vaccine. (2014) 32:2135–8. doi: 10.1016/j.vaccine.2014.02.055
254. Herrington D, Davis J, Nardin E, Beier M, Cortese J, Eddy H, et al. Successful immunization of humans with irradiated malaria sporozoites: humoral and cellular responses of the protected individuals. Am J Trop Med Hyg. (1991) 45:539–47. doi: 10.4269/ajtmh.1991.45.539
255. Edelman R, Hoffman SL, Davis JR, Beier M, Sztein MB, Losonsky G, et al. Long-term persistence of sterile immunity in a volunteer immunized with X-irradiated Plasmodium falciparum sporozoites. J Infect Dis. (1993) 168:1066–70. doi: 10.1093/infdis/168.4.1066
256. Egan JE, Hoffman SL, Haynes JD, Sadoff JC, Schneider I, Grau GE, et al. Humoral immune responses in volunteers immunized with irradiated Plasmodium falciparum sporozoites. Am J Trop Med Hyg. (1993) 49:166–73. doi: 10.4269/ajtmh.1993.49.166
257. Campo JJ, Aponte JJ, Skinner J, Nakajima R, Molina DM, Liang L, et al. RTS,S vaccination is associated with serologic evidence of decreased exposure to Plasmodium falciparum liver- and blood-stage parasites. Mol Cell Proteomics. (2015) 14:519–31. doi: 10.1074/mcp.M114.044677
258. Behet MC, Foquet L, Van Gemert GJ, Bijker EM, Meuleman P, Leroux-Roels G, et al. Sporozoite immunization of human volunteers under chemoprophylaxis induces functional antibodies against pre-erythrocytic stages of Plasmodium falciparum. Malar J. (2014) 13:136. doi: 10.1186/1475-2875-13-136
259. Kaushansky A, Rezakhani N, Mann H, Kappe SH. Development of a quantitative flow cytometry-based assay to assess infection by Plasmodium falciparum sporozoites. Mol Biochem Parasitol. (2012) 183:100–3. doi: 10.1016/j.molbiopara.2012.01.006
260. Zou X, House BL, Zyzak MD, Richie TL, Gerbasi VR. Towards an optimized inhibition of liver stage development assay (ILSDA) for Plasmodium falciparum. Malar J. (2013) 12:394. doi: 10.1186/1475-2875-12-394
Keywords: Plasmodium falciparum, malaria, sporozoites, protection, vaccine, models, correlates
Citation: Goh YS, McGuire D and Rénia L (2019) Vaccination With Sporozoites: Models and Correlates of Protection. Front. Immunol. 10:1227. doi: 10.3389/fimmu.2019.01227
Received: 29 October 2018; Accepted: 14 May 2019;
Published: 05 June 2019.
Edited by:
Ian Marriott, University of North Carolina at Charlotte, United StatesReviewed by:
Stefan Kappe, Seattle Children's Research Institute, United StatesJulius Clemence Hafalla, London School of Hygiene and Tropical Medicine (LSHTM), United Kingdom
Copyright © 2019 Goh, McGuire and Rénia. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Yun Shan Goh, Z29oX3l1bl9zaGFuQGltbXVub2wuYS1zdGFyLmVkdS5zZw==