 Lucia Stranavova1†Ondrej Pelak2†
Lucia Stranavova1†Ondrej Pelak2† Michael Svaton2
Michael Svaton2 Petra Hruba1
Petra Hruba1 Eva Fronkova2Antonij Slavcev3Klara Osickova4Jana Maluskova5Petr Hubacek6Jiri Fronek7
Eva Fronkova2Antonij Slavcev3Klara Osickova4Jana Maluskova5Petr Hubacek6Jiri Fronek7 Petra Reinke8
Petra Reinke8 Hans-Dieter Volk8
Hans-Dieter Volk8 Tomas Kalina2‡
Tomas Kalina2‡ Ondrej Viklicky1,4*‡
Ondrej Viklicky1,4*‡- 1Transplant Laboratory, Institute for Clinical and Experimental Medicine, Prague, Czechia
- 2CLIP – Childhood Leukaemia Investigation Prague, Department of Paediatric Haematology and Oncology, 2nd Faculty of Medicine, Charles University Prague and University Hospital Motol, Prague, Czechia
- 3Department of Immunogenetics, Institute for Clinical and Experimental Medicine, Prague, Czechia
- 4Department of Nephrology, Transplant Centre, Institute for Clinical and Experimental Medicine, Prague, Czechia
- 5Department of Transplant Pathology, Transplant Centre, Institute for Clinical and Experimental Medicine, Prague, Czechia
- 6Department of Paediatric Haematology and Oncology, 2nd Faculty of Medicine and Motol University Hospital, Charles University, Prague, Czechia
- 7Department of Transplant Surgery, Institute for Clinical and Experimental Medicine, Prague, Czechia
- 8BIH Centre for Regenerative Therapies, Berlin Centre for Advanced Therapies, Charité University Medicine Berlin, Berlin, Germany
Cytomegalovirus (CMV) infection is associated with allograft rejection but the mechanisms behind are poorly defined yet. Although cross-reactivity of T cells to alloantigen and CMV has been hypothesized, direct evidence in patients is lacking. In this observational cohort study, we tested the pre-transplant effector/memory T cell response to CMV peptide pools and alloantigen in 78 living donor/recipient pairs using the interferon-gamma Enzyme-Linked ImmunoSpot (ELISPOT) assay. To prove the hypothesis of cross-reactivity, we analyzed by applying next-generation sequencing the T cell receptor ß (TCR- ß) repertoire of CMV- and alloantigen-reactive T cells enriched from peripheral pre-transplant blood of 11 CMV-seropositive and HLA class I mismatched patients. Moreover, the TCR-repertoire was also analyzed in the allograft biopsies of those patients. There was a significant association between the presence of pre-transplant CMV immediate-early protein 1 (IE-1)-specific effector/memory T cells and acute renal allograft rejection and function (p = 0.01). Most importantly, we revealed shared TCR-ß sequences between CMV-IE1 and donor alloantigen-reactive T cells in all pre-transplant peripheral blood samples analyzed in CMV-seropositive patients who received HLA class I mismatched grafts. Identical TCR sequences were also found in particular in post-transplant allograft biopsies of patients with concomitant CMV infection and rejection. Our data show the presence of functional, cross-reactive T cells and their clonotypes in peripheral blood and in kidney allograft tissue. It is therefore likely that CMV-donor cross-reactivity as well as CMV specific T cell elicited inflammation is involved in the processes that affect allograft outcomes.
Introduction
Although the human cytomegalovirus (CMV) infection establishes a broad immunity which controls infection, rendering it asymptomatic in majority of immunocompetent hosts even if the CMV reactivates repeatedly following different stressors during life-time (1, 2). However, it might be associated with life-threatening complications in organ transplant recipients (3). Interestingly, it is widely acknowledged that, in addition to its direct pathogenic effects, CMV infection in organ transplant recipients is associated with more frequent acute and chronic rejection (4, 5). Although several possibilities have been proposed for those indirect negative effects, the mechanisms behind are poorly understood so far and direct proofs are missing. Persistent CMV infection elicits strong and lifelong T cell immunity that controls CMV reactivation/reinfection and prevents CMV disease by permanent dynamic interaction between the virus and the CMV-reactive T cell clones. However, CMV-reactive T cells can cause tissue damage by several mechanisms: (i) direct cytotoxic effect on CMV infected (allograft) cells, (ii) indirect bystander activation and proinflammatory milieu formation, and (iii) heterologous (cross-reactive) allorecognition (6).
The cross-reactivity of CMV-reactive effector T cells to HLA class I antigens has been discussed (7) and those cross-reactive cells were transiently found in the peripheral blood of kidney transplant recipients (8). T cell receptor (TCR) cross-reactivity has been suggested as primary means of increasing the effective size of T cell compartment, while cross-reactive memory cells have been shown to expand and activate more rapidly (9). Several mechanisms have been proposed for TCR cross-reactivity, including molecular mimicry and the ability of TCR to recognize different peptide-MHC complexes (10). Several other studies have shown the presence of cross-reactive virus-specific memory T cells and donor HLA molecules (7, 8, 11, 12). However, direct evidence of the role of heterologous TCR immunity in renal allograft rejection has not been shown so far.
Herein, we demonstrate that (i) the presence of CMV-reactive T cells pre-transplant predicts risk of acute allograft rejection, (ii) heterologous CMV- and donor-reactive cross-reactivity TCR-ß identical T cells pre-exists in patients prior to kidney transplantation, and (iii) identical cross-reactive T cell clones are detectable in renal allograft biopsies post-transplantation. Our data support the impact of heterologous immunity to CMV-IE1 and alloantigen among pre-transplant memory T cells on allograft outcome and indicate the need of adequate control not only by immunosuppression but also efficient antiviral strategies.
Materials and Methods
Patient Characteristics
In this observational cohort study, we evaluated the role of pre-transplant CMV-specific T cell immunity in acute rejection using an ELISPOT cohort consisting of 78 living donor kidney transplant recipients and their respective donors, all of whom underwent transplant surgery in the Institute for Clinical and Experimental Medicine in Prague between the years 2014 and 2017. The demographic data of the patients are summarized in Table 1. The peripheral blood of both donors and recipients was drawn pre-operatively to isolate PBMCs. All patients received tacrolimus, mycophenolate mofetil, and steroids as maintenance immunosuppression, initiated 48 h before the scheduled surgery. Patients at low immunological risk [panel-reactive antibody (PRA) <20%] received the anti-CD25 monoclonal antibody basiliximab (Simulect, Novartis), while patients at higher risk received rabbit polyclonal anti-thymocyte globulin (rATG, Thymoglobulin®, Genzyme Corporation) as induction immunosuppression. CMV prophylaxis with valganciclovir (Valcyte®, Roche) was given to CMV-seronegative recipients who had received grafts from seropositive donors or to CMV-seropositive recipients who had undergone rATG induction. Fourteen out of 78 patients experienced acute rejection episodes during the 1st year after transplantation and were treated as previously reported (13). For a detailed description of the histological findings, see Table S1.
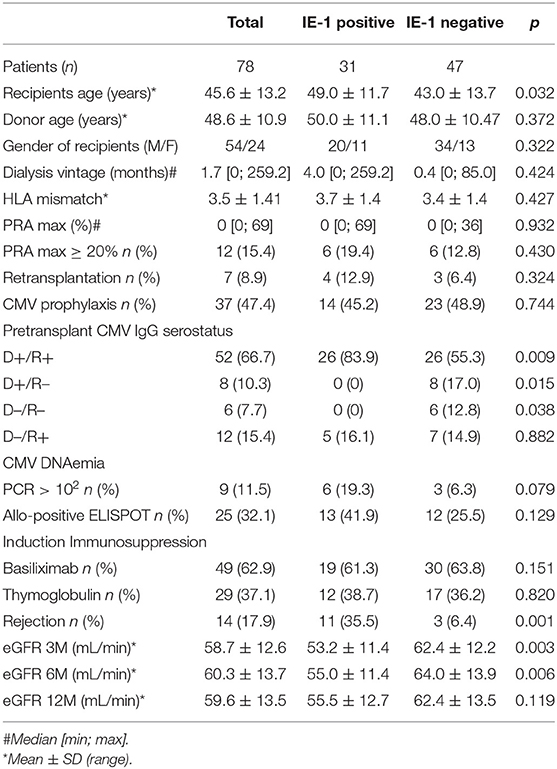
Table 1. Demographics of the “ELISPOT” patient cohort.
For the analysis of the TCR repertoire of CMV- and donor alloantigen-reactive T cells (the “cross-reactive” cohort) 11 donor/recipient pairs were selected with primary low risk and pre-transplant CMV-seropositive living donor renal allograft recipients from the years 2014 to 2016. For a summary of the demographic data of these patients, see Table 2. All patients received tacrolimus, mycophenolate mofetil, and steroids as maintenance immunosuppression (initiated 48 h before the scheduled surgery) and basiliximab as induction immunosuppression. All patients underwent a 3-month protocol kidney graft biopsy according to the centre's standard practice, while case biopsies were performed to histologically verify clinically suspected acute rejection. In 5 out of 11 patients, histological proven acute rejection episodes occurred within 3 months after the operation. Histological findings are given in Table S2. In the case of this patient cohort and their respective living donors, peripheral blood was drawn prior to transplantation in order to isolate PBMCs and the allograft biopsy was performed, with 2–3 mm of the tissue samples stored in Ambion RNAlater® Stabilization Solution (Thermo Fisher Scientific) for future molecular evaluation.
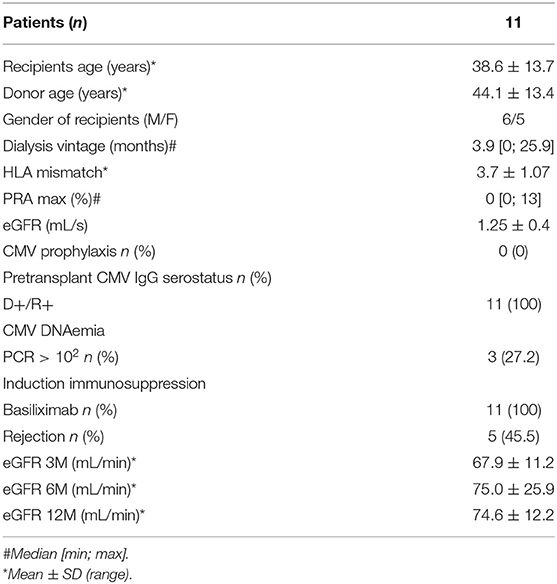
Table 2. Demographics of the “cross-reactive” cohort.
All patients from the ELISPOT and cross-reactive cohorts as well as their respective donors gave their written informed consent to participate in the study. The local ethics committee approved the study protocol under No. G14-08-38.
IFN-γ ELISPOT Assay
In the “ELISPOT” cohort, allo- and CMV-specific T cells were assessed using the IFN-γ ELISPOT method according to recently described protocols (14, 15). Peripheral blood mononuclear cells (PBMCs) were isolated from heparinized peripheral blood samples of donors and recipients taken prior to transplantation (using standard density gradient centrifugation) and cryopreserved in liquid nitrogen as described previously (16). After thawing, PBMCs were re-suspended with complete media [RPMI 1640 supplemented with 10% heat-inactivated fetal calf serum (FCS), penicillin + streptomycin (50 U/mL) and 1.7 mM sodium glutamate] and left for 24 h at 37°C in a CO2 incubator. Next, 3 × 105 recipient PBMCs were stimulated with CD3-depleted donor cells to detect allospecific T cells, and with CMV antigens [whole protein-spanning overlapping peptide pools of immediate-early protein 1 (IE-1) and phosphoprotein 65 (pp65), length of each is 15 amino acids with 11 amino acid overlap] (Miltenyi Biotec) to detect CMV-reactive T cells [using pokeweed mitogen from Autoimmune Diagnostika (AID), GmbH as a positive control]. CMV peptide pools were used in the concentration of 1 μg/mL of pp65 or IE-1. PBMCs were seeded in an ELISPOT plate and incubated for 24 h at 37°C in a CO2 incubator. The ELISPOT kit used to detect IFN-γ-producing cells was obtained from AID. After 24 h incubation at 37°C in the CO2 incubator, cells were removed and the ELISPOT plate processed according to the manufacturer's protocol. The resulting numbers of spots were measured semi-automatically using an ELISPOT reader (AID iSpot FluoroSpot Reader System ELR07 IFL).
Antigen Specific T Cells by Flow Cytometry and FACS Sorting
Antigen specific T cells (virus specific or donor reactive) were detected as proliferating CD8+ T cells by dye dilution technique. Eleven patients from the “cross-reactive” cohort were selected for flow cytometry sorting. Cryopreserved PBMCs from recipients and donors were thawed, suspended with complete media and left for 24 h at 37°C in a CO2 incubator. After resting, donor PBMCs were inactivated with Mitomycin C (50 μg/1 mL) (Sigma-Aldrich) for 2 h at 37°C in a CO2 incubator and washed twice with complete media (210 RCF/10 min). Afterwards, both recipient and donor PBMCs were labeled with the dilution dyes CellTrace™ Violet and Far Red Cell Proliferation kits (Thermo Fisher Scientific), respectively, according to the manufacturer's instructions. Labeled recipient PBMCs were aliquoted by 0.5 × 106 in a 96-well plate (2 mL, V-bottom, Greiner Bio-One, GmbH) with a culture medium [RPMI 1640 supplemented with 10% heat-inactivated FCS, penicillin+ streptomycin (50 U/mL), 1.7 mM sodium glutamate, 0.00036% (v/v) β-mercaptoethanol, and 10 U/mL IL-2]. CellTrace™ Violet dilution dye labeled PBMC were stimulated with the following CMV antigens: 1 μg/mL of pp65, 1 μg/mL of IE-1 (Miltenyi Biotec), or whole CMV lysate (Vidia) for 6 days. To detect alloreactive and cross-reactive T cells, inactivated donor cells (ratio 1:1) were used as a stimulus (Far Red dye labeled). Additional controls consisting of unstimulated recipient cells and recipient cells with additional IL-2 (50 U/mL) (Sigma-Aldrich) were used to eliminate bystander cell proliferation (data not shown). After 6 days of stimulation, the cells were harvested in 5 ml tubes and washed once with PBS containing 2 mM EDTA. Antigen specific cells proliferate in response to antigen and loose their dilution dye (CellTrace Violet low cells in Figure 2). Washed cells were stained with CMV peptide-pentamers (Pro5® MHC Pentamers, Proimmune) matched to the patient's HLA allele (Table S4) for 15 min at 4°C. Cells were washed once with PBS containing 2 mM EDTA and stained with antibodies against CD3 (anti-CD3 PC7, Beckman Coulter), CD4 (anti-CD4 ECD, Immunotech), CD8 (anti-CD8 APC-H7, BD Biosciences), and CD45 (anti-CD45 PerCP, Exbio) for 20 min at 4°C. As a subsequent step, cells were washed once with PBS and analyzed using the FACSAria™ III Cell sorter (BD Biosciences). Proliferating CellTrace™ Violet low cells were FACS sorted into a 50 μl elution buffer (Qiagen) (sort gate position is denoted as R1 gate in Figure 2). Sorted cells were then incubated in a thermo block heater at 99°C and at 500 rpm for 60 min to isolate DNA; the samples were subsequently stored at −20°C for next-generation sequencing (NGS) library preparation. FlowJo software (FlowJo, LLC) was used for flow cytometry data analysis.
To limit the possibility that cells stimulating the Allo responses contain CMV-infected cells, CMV seropositive donors' blood was examined for the presence of CMV genome by real-time quantitative PCR. DNA was extracted from whole blood using MagNA Pure Compact Nucleic Acid Isolation Kit (Roche, Basel, Switzerland), and the viral loads were normalized to 10,000 human genomic equivalents (17, 18). No CMV genome was found in those samples (Table S3).
Kidney Biopsies and DNA Isolation
Kidney biopsy tissue samples taken from the 3-month protocol biopsies or from case biopsies due to deterioration of kidney graft function were available in 7 out of 11 patients from the “cross-reactive” patient cohort. All biopsy tissue samples were stored in Ambion RNAlater® Stabilization Solution (Thermo Fisher Scientific) at −80°C. The tissue samples were thawed and genomic DNA isolated using the QIAamp DNA Micro Kit (Qiagen) according to the manufacturer's protocol. Isolated DNA were then stored at −20°C for NGS sequencing library preparation.
TCR-β Repertoire Sequencing Using a Next Generation Sequencing Approach
Genomic DNA isolated from FACS-sorted cells and kidney biopsies were used for sequencing library preparation. Two-step PCR was used to detect the majority of V and J segments in complementarity-determining region 3 (CDR3) of the TCR-β sequence, as described previously (19–21). The established libraries were sequenced on the IonTorrent PGM using Hi-Q 400 bp chemistry (Thermo Fisher) and data were analyzed using the Vidjil application (VIDJIL web platform) (22). Samples with analyzed reads below 1,000 or reads in which the CDR3 sequence could not be identified were discarded from further analysis. Clones from the analyzed samples with reads lower than 10 were considered a source of possible cross-contamination between barcoded samples and were thus disregarded from the analysis. Non-productive rearrangements presumably originating from the second allele were retained in the analysis as additional markers of clonotypes.
Statistical Analysis
Data were analyzed using GraphPad InStat 3 (GraphPad Software) and IBM SPSS 22 software. The normality of data distribution was tested using the Kolmogorov-Smirnov test. Since all data were shown not to correspond with standard normal distribution, only non-parametric statistical methods were used. The Mann-Whitney U and Chi-Square tests were used to compare the patient groups. Cox proportional hazards model were used to identify risk variables for rejection. The association of recipient, donor, and transplant parameters and immunological factors were first entered to univariate regression analyses (Table 3). All significant variables (p < 0.05) were included into final multivariate Cox model adjusted for induction treatment presenting the extent of immunological risk. Kaplan-Meier survival curves and the log-rank test were used to project rejection-free intervals and to compare groups. The Wilcoxon matched-pair signed-rank test was used to evaluate ELISPOT differences. To evaluate prediction of rejection risk on the basis of pretransplant pp65/IE-1/Allo ELISPOT receiver operating characteristic (ROC) curves and the calculation of the area under the curve (AUC) were used. Spearman's rank correlation coefficient was used to calculate correlations of pretransplant IE-1 ELISPOT and eGFR in 3, 6, and 12 months. All results with a P-value of <0.05 were considered statistically significant.
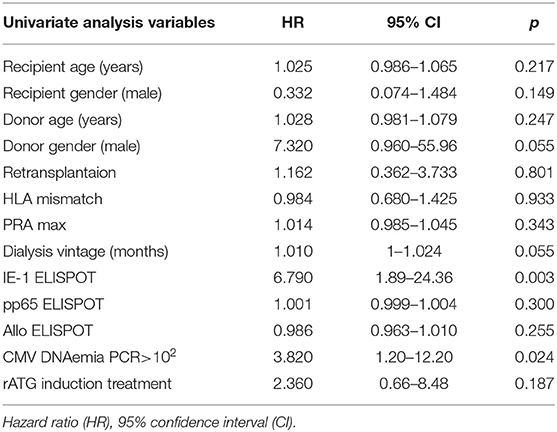
Table 3. Risk factors associated with graft rejection in univariate Cox regression.
Results
Pre-transplant Presence of CMV-Specific T Cells Predicts Acute Rejection
To evaluate the relation of pre-existing CMV-reactive and donor-alloreactive T cells and acute rejection of renal allografts, ELISPOT assays were performed in 78 consecutive patients. The design of the IFNγ-ELISPOT allows the detection only of memory/effector CD4+ and CD8+ T cells preformed in vivo. As expected, we observed significantly higher frequencies of pre-existing CMV-reactive than alloreactive memory/effector T cells (Figure 1A). Interestingly, the pre-transplant presence of a CMV-reactive response, both to IE-1 and pp65 whole protein overlapping peptide pools, had a stronger predictive power of acute rejection [IE-1 and pp65: AUC = 0.70, cut-off = 122.5 at 69.8% sensitivity and 80% specificity, 95% confidence interval (CI): 0.54–0.87; p = 0.014 and AUC = 0.59, cut-off = 332 at 63.5% sensitivity; and 53.3% specificity, 95% CI: 0.44–0.74, p = 0.27, respectively] than the donor-alloreactive ELISPOT (AUC = 0.40, cut-off = 25 at 66.7% sensitivity and 25.0% specificity, 95% CI: 0.27–0.59, p = 0.39, Figure 1B). Moreover, a shorter rejection-free interval was observed in patients with a positive pre-transplant IE-1 ELISPOT (Figure 1C).
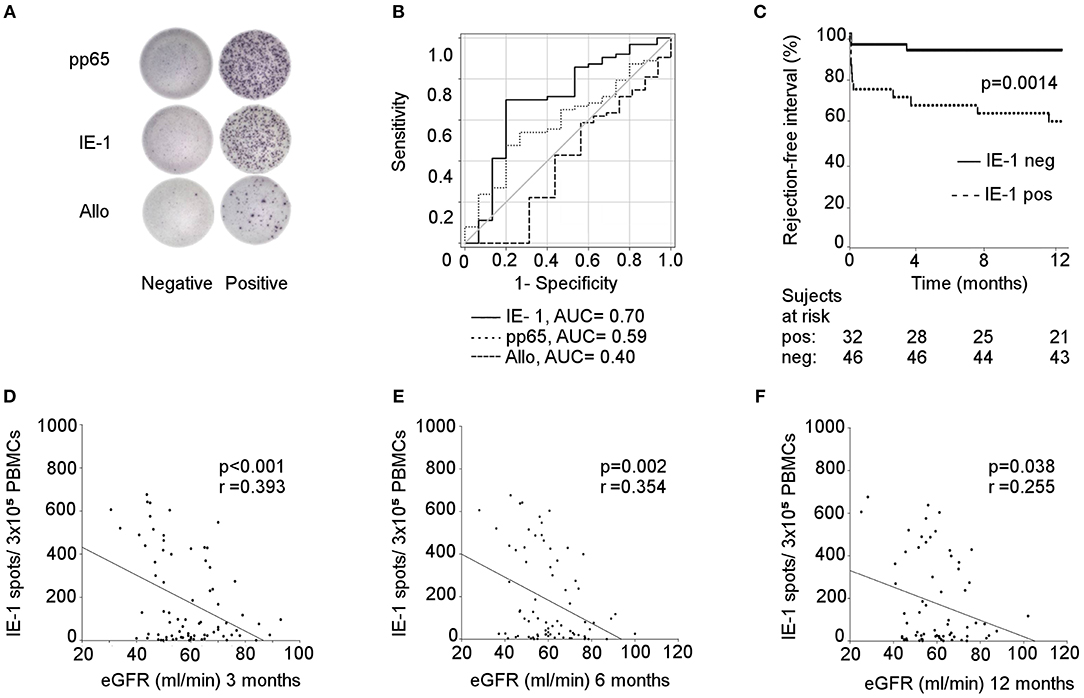
Figure 1. CMV-specific (but not allospecific) ELISPOT for predicting rejection and kidney allograft function. (A) visualization of IFN-γ spots after stimulation with pp65/IE-1/alloantigens in positive and negative recipients; (B) prediction of rejection risk based on the pre-transplant pp65/IE-1/allo ELISPOT; The operating characteristic (ROC) curves and the calculation of the area under the curve (AUC) were used for this analysis; IE-1: 95% confidence interval (CI): 0.54–0.87; p = 0.014; pp65: 95% CI: 0.44–0.74, p = 0.27; Allo: 95% CI: 0.27–0.59, p = 0.39; (C) rejection-free intervals of patients using IE-1-positive and -negative ELISPOTs expressed as Kaplan-Meier survival curves; p = 0.0014. Correlation between a pre-transplant IE-1 ELISPOT and eGFR at 3 (D), 6 (E), and 12 (F) months were established by Spearman's rank correlation coefficient; 3M: p < 0.001; 6M: p = 0.002; 12M: p = 0.038.
Univariate Cox regression analysis revealed as significant risk factors of acute rejection only pretransplant positive IE-1 ELISPOT [Hazard ratio (HR) = 6.8, 95% CI: 1.89–24.36, p = 0.003] and post transplant positive CMV viral load by PCR>102 (HR = 3.8, 95% CI: 1.2–12.2, p = 0.024) (Table 3). A multivariate Cox regression analysis adjusted for ATG induction treatment and CMV PCR > 102 revealed only IE-1 positive ELISPOT (HR = 6.2, 95% CI: 1.67–22.3, p = 0.006) to be independent risk factor of acute rejection.
Interestingly, significant correlations were also found between pre-transplant IE-1 ELISPOTs and kidney graft function (estimated glomerular filtration rate (eGFR) using the Chronic Kidney Disease Epidemiology Collaboration (CKD-EPI) equation) at 3, 6, and 12 months (p < 0.001, p = 0.002, and p = 0.038, respectively) (Figures 1D–F). The demographic characteristics of patients with positive and negative IE-1 ELISPOTs are summarized in Table 1.
Taken together, CMV-reactive cellular immunity predicts acute rejection and short-term outcome of renal allografts.
CMV- and Alloreactive T Cells Express Shared TCR Sequences
The strong association between the pre-transplant presence of CMV-reactive T cells and rejection prompted us to investigate the possible cross-reactivity of CMV-specific T cells to donor alloantigens by search for shared TCR sequences. First, we combined the donor alloantigen MLR with CMV-peptide pentamer staining to evaluate cross-reactivity at single cell level in pre-transplant peripheral blood mononuclear cells (PBMCs) (8). In contrast to previously published studies, we detected no T cells cross-reactive to the immunodominant pp65 CMV peptide and donor cells, as detected by the co-staining for CMV-pp65 pentamer and cell tracking dilution following proliferation to alloantigen stimulation (Figure 2A). However, the response to dominant epitopes (pentamer staining) is lower compared to the proliferative response to the whole CMV peptide pool as demonstrated in 10 out of 11 CMV-seropositive patients (Figures 2B–D and summarized in Figure S1). In parallel, donor alloreactive T cells were present in all patient samples (including CMV pentamer-negative ones) prior to transplantation.
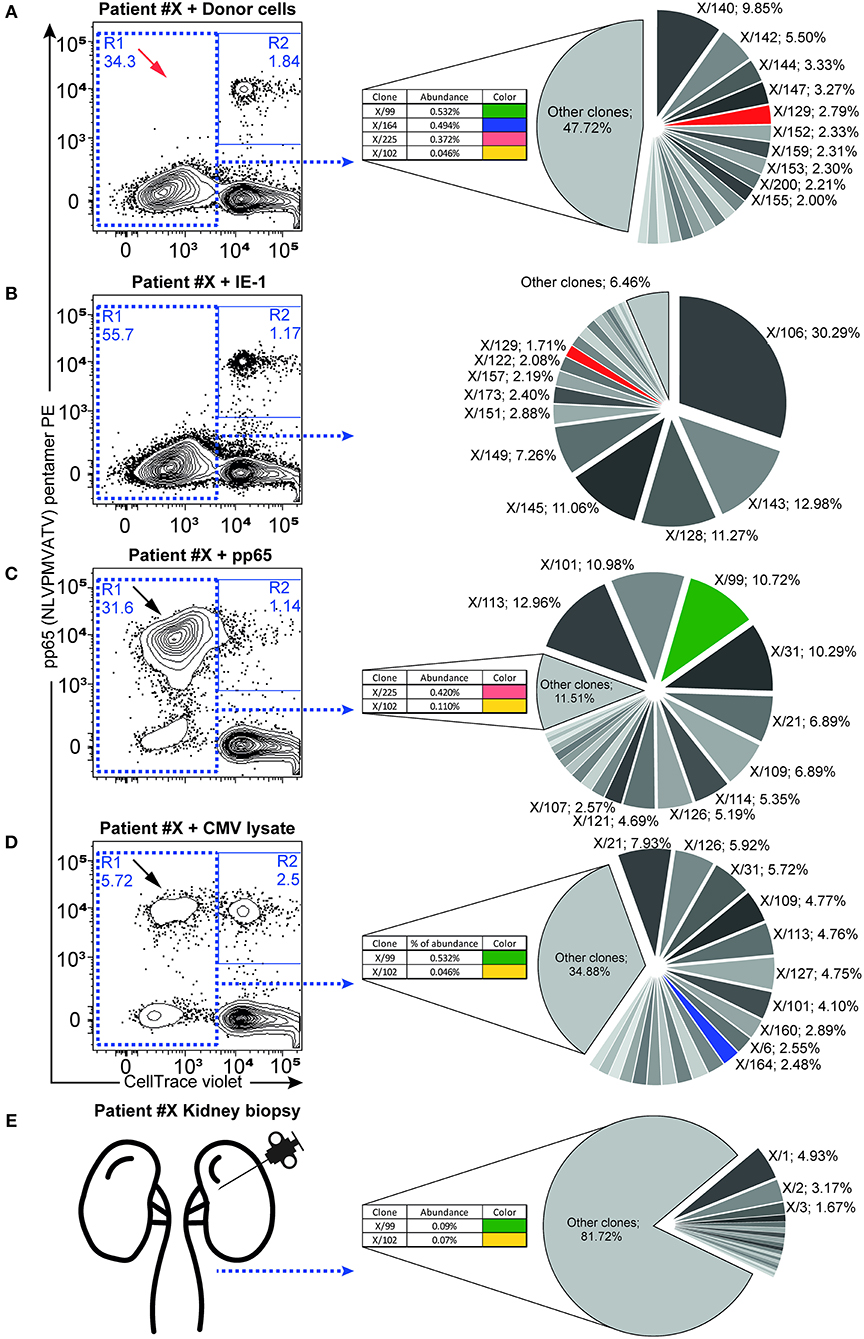
Figure 2. Antigen-responding CD8+ T cells containing cross-reactive TCR-β sequences. Antigen-specific responses were assessed as proliferating recipient CD8+ T cells (CellTrace low cells, plots show gated CD3+CD8+ lymphocytes) after 6-days of ex-vivo culture in combination with pentamer staining for the immunodominant (pp65: NLVPMVATV-specific) TCR receptor. Flow cytometry dot plots show the proliferation response of CD8+ T cells to donor cells (A), IE-1 (B), pp65 (C), and whole CMV lysate (D). Proliferating cells in R1 were FACS-sorted and used for subsequent NGS TCR-β repertoire analysis. Twenty of the most abundant TCR-β sequences are represented in the pie chart graph (right panels), while additional minor cross-reactive clones are shown in inlets. Color codes highlight the same TCR sequence clones found in the respective antigen-responding cells or in the kidney (E). Black arrows highlight antigen-specific proliferating T cells recognizing the immunodominant pp65 peptide, with red arrows indicating their absence from the donor cell-elicited response. Relative clone abundance is shown next to the clone name in 10 of the most abundant clones or in the inlets. One representative patient (#X) is shown, with a complete list of responding cell fractions. The amount of sorted cells and available NGS reads are given in Table 4, while the cross-reactive clones found are listed in Table S5.
To investigate whether cross-reactivity would be present among the total pool of CMV-reactive T cells, we isolated antigen-reactive T cells (either reactive to CMV peptide pool or donor PBMCs) by FACS sorting (Figures 2A–D) and performed NGS of TCR-β sequences (Table 4). In 10 out of 11 patients, we acquired a sufficient amount of reads for analysis. We hypothesize that while pentamer staining only reveals single immunodominant CD8+ T cell clones, cross-reactivity may be caused by other less-dominant TCR clones. We were able to identify hundreds of distinct TCR-ß sequences from sorted antigen-reactive T cells from all patients (median 392 [241; 491], Table S5). Indeed, multiple clones sharing the same unique TCR-ß sequences were found in both CMV- and donor-reactive samples (Figure 2, right panels) from all patients, regardless of occurrence of rejection (Table S6).
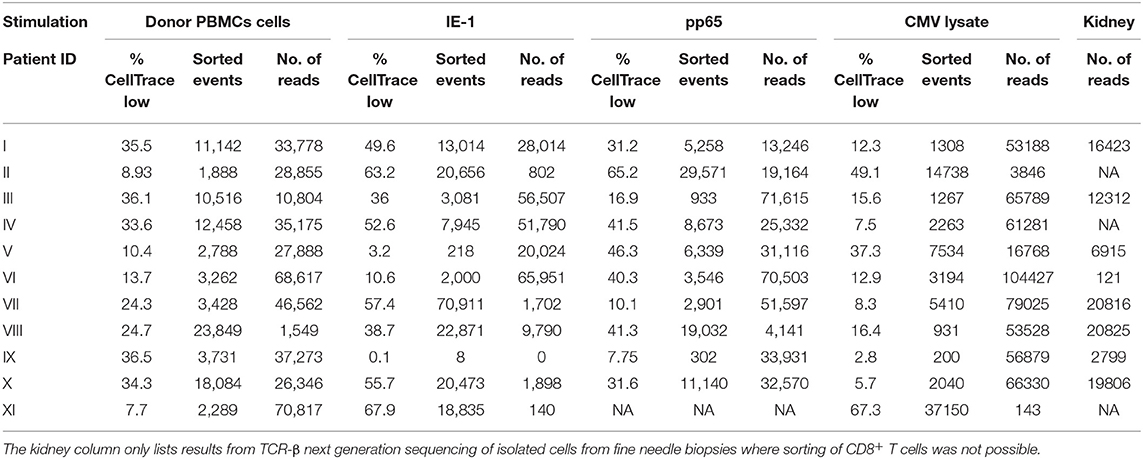
Table 4. Percentages of proliferating CD8+ T cells (% CellTrace low), the number of sorted proliferating CD8+ T cells (sorted events), and the number of reads obtained after TCR-β next generation sequencing of sorted proliferating CD8+ T cells (No. of reads) in response to different stimulations.
Our results also provide evidence that both donor cells and CMV antigens can trigger identical T cell clones for proliferation showing functional responsiveness.
Shared Cross-Reactive TCR-ß Clonotypes Are Detectable in Renal Allograft Biopsies
Next, we sought to investigate whether cross-reactive TCR-ß clonotypes would be detectable in the allografts. Allograft biopsy samples were made available for 7 patients investigated for alloreactive and CMV-specific clonotypes (see “cross-reactive” cohort described above). In the kidney biopsy samples of 6 out of 7 patients, we were able to find identical TCR-β CDR3 sequences as in the alloreactive T lymphocytes pre-transplant. For the remaining patient, only 16 clones could be analyzed from the sequencing results of the MLR tube. Therefore, cross-reactive clones could have been missed due to the lower coverage of this sequencing library. In parallel, CMV-reactive TCR-ß clonotypes were found in the biopsy samples even at higher frequencies in the same patients, with a median of 3 clones per patient and a maximum of 11 (Table 5). The CMV-reactive clonotypes in the kidney covered 0.5–6.4% of all TCR-β sequences (Table 5) found in the kidney biopsies (see individual clones in Table S6). Finally, in 3 out of 7 biopsy samples, we detected CMV/donor alloantigen cross-reactive clonotypes identified pre-transplant in peripheral blood (Table 5).

Table 5. CMV-, Allo-, and Cross-reactive clones identified from blood pre-transplant are found in the kidney.
Remarkably, in agreement with the acknowledged capacity of CMV-reactive T cell clones to expand upon CMV reactivation, cumulative abundance of CMV-reactive TCR-ß sequences was the highest (6.4 and 5.1%) in the two kidney tissue samples obtained from patients (No. VIII and X) suffering from significant CMV reactivation (viral load: 935 copies/ml and 1,020 copies/ml of plasma, respectively) concomitantly with biopsy-proven cellular rejection.
In summary, CMV-reactive and alloreactive clonotypes were found in all allograft biopsies patients analyzed, and in 3 out of 7 patients, we detected cross-reactive clonotypes as defined by pre-transplant analyses. Importantly, the highest number of these cross-reactive clones was observed in the two patients with CMV reactivation and concomitant cellular rejection.
Discussion
The immunity in response to previous virus infections can modify the immune response to other antigens. Although heterologous immunity can be beneficial by boosting protective responses, it can also result in severe immunopathologies (6). Here, for the first time, we provide evidence that heterologous immunity can be detected in blood and biopsies of renal allograft recipients. Firstly, we found that high frequencies of CMV-reactive effector/memory (but not of alloreactive) T cells detected pre-transplant were associated with subsequent occurrence of T cell-mediated rejection. Secondly, multiple cross-reactive T cell clones (shared TCR-ß sequences) were found in both CMV- and donor-reactive T cells enriched from pre-transplant peripheral blood samples. Finally, TCR-ß sequences of alloreactive [CMV-reactive, and cross-reactive (CMV & MLR)] clonotypes were found in renal allografts; the latter particularly in association with CMV-associated T cell mediated acute rejection. Therefore, our data demonstrate that identical clonotypes of T cells can react in response to alloantigens as well as CMV antigens. This observation might explain how CMV reactivation, especially in the case of high viral load during uncontrolled replication, boosts directly not only the CMV but also the alloimmune T cell response.
Interestingly, in contrast to the association between high levels of pre-transplant CMV memory/effector response, in our study, we found no associations between pre-transplant donor-reactive memory/effector T cell response and acute rejection. Given the standardized, validated and robust ELISPOT method applied (previously used in a large European multicentre clinical trial; see www.biodrim.eu), it is unlikely that any methodological bias occurred. An earlier study reported that higher pre-transplant T cell alloresponse was associated with acute allograft rejection in a study where patients received non-lymphocyte-depleting induction immunosuppression (23). Contrary, in our study some 40% of patients had received rATG T cell depletive induction immunosuppression. Similarly, pre-transplant allo-T cell responses have also been shown to correlate with lower post-transplant eGFR in patients with non-depleting induction (24). Apart from the association between CMV-specific memory/effector T cells and acute rejection, we found significant correlation with lower post-transplant eGFR at three different time-points during the first post-Tx year, rendering our observations more robust. Therefore, it is likely that the T cell-depletion strategy used in about half of our patients effectively reduced the available clonal size of alloreactive memory/effector T cells to a level that could be further controlled by maintenance immunosuppression.
Interestingly, there was weaker association of CMV-pp65- vs. CMV-IE-1-reactive T cells with acute rejection in our study. This phenomenon might be explained by higher CD8+ T cell response to IE-1 than to pp65 antigens (25).
In fact, subclinical CMV reactivation is frequently detected in over 30% of kidney transplant recipients despite CMV prophylaxis (26–28). We speculate that subtle localized CMV reactivations are even more frequent and, while undetected, provide antigen stimulation to CMV-specific T cells. This is in line with observations made by authors of a previous prospective randomized trial. They found that late-onset of CMV viremia, which developed in more than half of patients despite CMV prophylaxis, is associated with poorer outcomes (29).
Although it was reported that at least 151 of the 213 predicted CMV proteins, elicited T cell responses in at least one out of 33 donors (30), we and others could show that the T cell responses to IE-1 and pp65 CMV-proteins are the most dominant ones. Therefore, we concentrated in this study on the two immunodominant CMV proteins.
Applying the previously described method for detecting cross-reactive T cells based on MLR-reactivity combined with peptide-pentamer staining, was not effective in our scenario to detect cross-reactive T cells (7, 8, 31). One reason for this might be the use of unbiased PBMC samples with a scarcity of cross-reactive cells. In fact, as our access to patient material was limited by ethical reasons, we used only 5 × 105 T lymphocytes for functional stimulation, resulting after sort in limited yield of antigen-reactive T cells ranging from 3,846 to 104,427 and 1,888 to 23,849 CMV- and allospecific-proliferating T cells, respectively, for TCR repertoire analysis. Moreover, our recent data show that immunodominant epitopes for one particular HLA-type, as detected and enriched by peptide/dextramer staining, do not reflect the whole response to a particular CMV protein. Therefore, we developed recently the method of T cell stimulation by whole protein-spanning overlapping peptide pools covering almost all epitopes in a HLA-independent matter (32). Applying this method here, we could detect all three categories of CMV-, donor alloantigen-, and cross-reactive T cells in all patients with sufficient yield after sorting derived from pre-transplant blood samples despite limited amounts of reactive T cells (and resulting reads in NGS). These results show the potency of recipients' memory/effector T cell pool to react in case of CMV reactivation post-transplantation with both a protective CMV-specific and a putatively harmful CMV/allo-cross reactive response. In other words, CMV reactivation because of breakthrough through or after weaning of antiviral prophylaxis that might be amplified by TNF-release following ATG application can trigger putatively harmful alloresponse by crossreactivity (33). In line with this, we could detect cross-reactive TCR-ß clonotypes in the kidney biopsies of 3 out of 6 patients with sufficient yield for analysis. Whether the absence of detectable shared cross-reactive TCR-ß sequences in the remaining three biopsy samples is due to sensitivity problems or missing triggering by CMV is not clear, but the high abundancy in the samples just of the two patients suffering from enhanced CMV viral load and concomitant acute rejection supports their pathogenic role in CMV-associated graft injury.
In summary, our data show that within the large peripheral population of CMV-specific memory T cells there is a pool of cross-reactive T cell clonotypes that can produce effector T cells capable of migrating into kidney allografts. Moreover, these T cell clonotypes (when in the presence of chronic antigenic stimuli, such as CMV) may be susceptible to enhanced proliferation and allograft rejection. This phenomenon seems to be universal and corresponds with previous hypotheses about the cross-reactive virus-alloimmune response (12, 34). Specific allo-HLA cross-reactivity has been reported for EBV, CMV, varicella-zoster virus (VZV), and influenza A virus-specific T cells at clonal level, while cross-reactivity has been shown to be mediated by the same TCRs (35, 36). However, our data demonstrate for the first time their occurrence in the unbiased bulk T cell pool from peripheral blood and intragraft.
The limitations of this study must also be acknowledged. The analysis was confined by the limited number of patients and TCR-β chains; furthermore, TCR-α rearrangements were not examined. The configuration of TCR-β chains (including D segments) building in particular the CDR3 region ensures much greater variability of rearranged sequences than TCR-α. Therefore, TCR-β is considered more informative than TCR-α and has been widely used in similar studies. Aside of T cells several other cells (e.g., NK cells) may produce IFNγ after stimulation. Therefore, we phenotypized IFNγ-producing cells stimulated by CMV antigens by flow cytometry. However, the majority of IFNγ-producing cells were T lymphocytes (55%), while NK cells accounted for 5% of IFNγ-producing cells only. The “cross-reactive cohort” subjected TCR- β NGS comprised only by CMV seropositive donor-recipient pairs. Among CMV seropositive-donors cells the CMV-infected cells might be present (37). To minimize the risk of potential activation by CMV infected donor cells, we evaluated those donors for the presence of CMV in their peripheral blood and found none CMV genome.
Our data show that CMV-specific cellular response pre-transplant predicts rejection and document that surprisingly large proportion of patients harbors CMV and donor cross-reactive clones. CMV and donor cross-reactive T cells might thus directly damage the donor cells, being expanded by CMV antigenic stimulation during CMV reactivation. This effect might be supported by CMV specific response that builds inflammatory environment in the kidney. We recommend the approaches aimed at preventing CMV reactivation to be employed more aggressively; not only to prevent CMV disease but also to limit cross-reactivity-induced graft rejection.
In conclusion, we report that in our patient cohort the presence of cytomegalovirus IE-1-specific memory/effector IFN-gamma secreting T cells predict kidney transplant rejection and poorer 1 year graft function. Since we established the presence of functional, cross-reactive T cells and their clonotypes in peripheral blood, tracking the clonotypes directly in the kidney tissue, it is therefore likely that CMV-donor cross-reactivity as well as CMV specific T cell elicited inflammation is involved in the processes that affect allograft outcomes. Future studies should be carried out to determine whether more aggressive prevention and treatment of CMV reactivation might possibly limit alloimmune injury boosted by cross-reactive T cells.
Data Availability Statement
All datasets generated for this study are included in the article/Supplementary Material.
Ethics Statement
The study protocol was approved by the Ethics Commintee of the Institute for Clinical and Experimental Medicine and Thomayer Hospital under number: G14-08-38. All subjects gave written informed consent in accordance with the Declaration of Helsinki.
Author Contributions
OV and TK share senior authorship designed and supervised the research and wrote the manuscript. LS, OP, MS, PHr, AS, and EF performed the research, participated in the data analysis, and manuscript writing. JF, JM, PHu, and KO helped with carrying out the research. PR and H-DV helped in establishment of the ELISPOT technology and significantly contributed to writing the manuscript.
Funding
This study was supported by the Ministry of Health of the Czech Republic (Grant No. 15-26865A) and in part by Bio-DrIM (EU FP7 program). EF and MS were supported by PRIMUS/17/MED11. TK and OP were supported by the Ministry of Education, Youth and Sports of the Czech Republic (NPU I project LO1604).
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We are grateful to Petr Kolar from the Information Technology Division at IKEM for his clinical database support, Maik Stein from BIH Centre for Regenerative Therapies for his help with establishing the ELISPOT method, Janka Slatinska and Vladimir Hanzal from the Department of Nephrology at IKEM for their help with patient management, Renata Zamecnikova and Tereza Neradova for their help with living donor management, Michaela Cerna, Bohumira Kronosova, and Romana Polackova for the sample collection and, finally, Michael Fitzgerald for English correction.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fimmu.2019.02549/full#supplementary-material
Abbreviations
TCR, T cell receptor; ELISPOT, Interferon-gamma Enzyme-Linked ImmunoSpot; IE-1, CMV immediate-early protein 1; pp65, Phosphoprotein 65; CI, Confidence interval; eGFR, Estimated glomerular filtration rate; CKD-EPI, Chronic Kidney Disease Epidemiology Collaboration equation; MLR, Mixed lymphocyte reaction; PBMCs, Peripheral blood mononuclear cells; NGS, Next-generation sequencing; VZV, Varicella-zoster virus; rATG, Rabbit polyclonal antithymocyte globulin; PRA, Panel-reactive antibody; CDR3, Complementarity-determining region 3; HR, Hazard ratio; FCS, Fetal calf serum; ROC, Receiver operating characteristic curve; AUC, Area under the curve.
References
1. Reinke P, Prosch S, Kern F, Volk HD. Mechanisms of human cytomegalovirus (HCMV) (re)activation and its impact on organ transplant patients. Transpl Infect Dis. (1999) 1:157–64. doi: 10.1034/j.1399-3062.1999.010304.x
2. Rafailidis PI, Mourtzoukou EG, Varbobitis IC, Falagas ME. Severe cytomegalovirus infection in apparently immunocompetent patients: a systematic review. Virol J. (2008) 5:47. doi: 10.1186/1743-422X-5-47
3. Klenerman P, Oxenius A. T cell responses to cytomegalovirus. Nat Rev Immunol. (2016) 16:367–77. doi: 10.1038/nri.2016.38
4. Reinke P, Fietze E, Ode-Hakim S, Prosch S, Lippert J, Ewert R, et al. Late-acute renal allograft rejection and symptomless cytomegalovirus infection. Lancet. (1994) 344:1737–8. doi: 10.1016/S0140-6736(94)92887-8
5. Cainelli F, Vento S. Infections and solid organ transplant rejection: a cause-and-effect relationship? Lancet Infect Dis. (2002) 2:539–49. doi: 10.1016/S1473-3099(02)00370-5
6. Sharma S, Thomas PG. The two faces of heterologous immunity: protection or immunopathology. J Leukoc Biol. (2014) 95:405–16. doi: 10.1189/jlb.0713386
7. Gamadia LE, Remmerswaal EB, Surachno S, Lardy NM, Wertheim-van Dillen PM, van Lier RA, et al. Cross-reactivity of cytomegalovirus-specific CD8+ T cells to allo-major histocompatibility complex class I molecules. Transplantation. (2004) 77:1879–85. doi: 10.1097/01.TP.0000131158.81346.64
8. Heutinck KM, Yong SL, Tonneijck L, van den Heuvel H, van der Weerd NC, van der Pant KA, et al. Virus-specific CD8(+) T cells cross-reactive to donor-alloantigen are transiently present in the circulation of kidney transplant recipients infected with CMV and/or EBV. Am J Transplant. (2016) 16:1480–91. doi: 10.1111/ajt.13618
9. Kim SK, Brehm MA, Welsh RM, Selin LK. Dynamics of memory T cell proliferation under conditions of heterologous immunity and bystander stimulation. J Immunol. (2002) 169:90–8. doi: 10.4049/jimmunol.169.1.90
10. Yin Y, Mariuzza RA. The multiple mechanisms of T cell receptor cross-reactivity. Immunity. (2009) 31:849–51. doi: 10.1016/j.immuni.2009.12.002
11. van den Heuvel H, Heutinck KM, van der Meer-Prins EP, Yong SL, Claas FH, Ten Berge IJ. Detection of virus-specific CD8+ T cells with cross-reactivity against alloantigens: potency and flaws of present experimental methods. Transplant Direct. (2015) 1:e40. doi: 10.1097/TXD.0000000000000550
12. D'Orsogna L, van den Heuvel H, van Kooten C, Heidt S, Claas FHJ. Infectious pathogens may trigger specific allo-HLA reactivity via multiple mechanisms. Immunogenetics. (2017) 69:631–41. doi: 10.1007/s00251-017-0989-3
13. Viklicky O, Hribova P, Volk HD, Slatinska J, Petrasek J, Bandur S, et al. Molecular phenotypes of acute rejection predict kidney graft prognosis. J Am Soc Nephrol. (2010) 21:173–80. doi: 10.1681/ASN.2008121268
14. Nickel P, Bold G, Presber F, Biti D, Babel N, Kreutzer S, et al. High levels of CMV-IE-1-specific memory T cells are associated with less alloimmunity and improved renal allograft function. Transpl Immunol. (2009) 20:238–42. doi: 10.1016/j.trim.2008.11.002
15. Bestard O, Lucia M, Crespo E, Van Liempt B, Palacio D, Melilli E, et al. Pretransplant immediately early-1-specific T cell responses provide protection for CMV infection after kidney transplantation. Am J Transplant. (2013) 13:1793–805. doi: 10.1111/ajt.12256
16. Gebauer BS, Hricik DE, Atallah A, Bryan K, Riley J, Tary-Lehmann M, et al. Evolution of the enzyme-linked immunosorbent spot assay for post-transplant alloreactivity as a potentially useful immune monitoring tool. Am J Transplant. (2002) 2:857–66. doi: 10.1034/j.1600-6143.2002.20908.x
17. Tanaka N, Kimura H, Iida K, Saito Y, Tsuge I, Yoshimi A, et al. Quantitative analysis of cytomegalovirus load using a real-time PCR assay. J Med Virol. (2000) 60:455–62. doi: 10.1002/(SICI)1096-9071(200004)60:4<455::AID-JMV14>3.0.CO;2-Q
18. Hubacek P, Virgili A, Ward KN, Pohlreich D, Keslova P, Goldova B, et al. HHV-6 DNA throughout the tissues of two stem cell transplant patients with chromosomally integrated HHV-6 and fatal CMV pneumonitis. Br J Haematol. (2009) 145:394–8. doi: 10.1111/j.1365-2141.2009.07622.x
19. Kotrova M, Muzikova K, Mejstrikova E, Novakova M, Bakardjieva-Mihaylova V, Fiser K, et al. The predictive strength of next-generation sequencing MRD detection for relapse compared with current methods in childhood ALL. Blood. (2015) 126:1045–7. doi: 10.1182/blood-2015-07-655159
20. Theunissen P, Mejstrikova E, Sedek L, van der Sluijs-Gelling AJ, Gaipa G, Bartels M, et al. Standardized flow cytometry for highly sensitive MRD measurements in B-cell acute lymphoblastic leukemia. Blood. (2017) 129:347–57. doi: 10.1182/blood-2016-07-726307
21. van Dongen JJ, Langerak AW, Bruggemann M, Evans PA, Hummel M, Lavender FL, et al. Design and standardization of PCR primers and protocols for detection of clonal immunoglobulin and T-cell receptor gene recombinations in suspect lymphoproliferations: report of the BIOMED-2 concerted action BMH4-CT98–3936. Leukemia. (2003) 17:2257–317. doi: 10.1038/sj.leu.2403202
22. Duez M, Giraud M, Herbert R, Rocher T, Salson M, Thonier F. Vidjil: A web platform for analysis of high-throughput repertoire sequencing. PLoS ONE. (2016) 11:e0166126. doi: 10.1371/journal.pone.0166126
23. Crespo E, Lucia M, Cruzado JM, Luque S, Melilli E, Manonelles A, et al. Pre-transplant donor-specific T-cell alloreactivity is strongly associated with early acute cellular rejection in kidney transplant recipients not receiving T-cell depleting induction therapy. PLoS ONE. (2015) 10:e0117618. doi: 10.1371/journal.pone.0117618
24. Hricik DE. Transplant immunology and immunosuppression: core curriculum 2015. Am J Kidney Dis. (2015) 65:956–66. doi: 10.1053/j.ajkd.2015.01.026
25. Slezak SL, Bettinotti M, Selleri S, Adams S, Marincola FM, Stroncek DF. CMV pp65 and IE-1 T cell epitopes recognized by healthy subjects. J Transl Med. (2007) 5:17. doi: 10.1186/1479-5876-5-17
26. Selvey LA, Lim WH, Boan P, Swaminathan R, Slimings C, Harrison AE, et al. Cytomegalovirus viraemia and mortality in renal transplant recipients in the era of antiviral prophylaxis. Lessons from the western Australian experience. BMC Infect Dis. (2017) 17:501. doi: 10.1186/s12879-017-2599-y
27. Fisher RA. Cytomegalovirus infection and disease in the new era of immunosuppression following solid organ transplantation. Transpl Infect Dis. (2009) 11:195–202. doi: 10.1111/j.1399-3062.2009.00372.x
28. Humar A, Lebranchu Y, Vincenti F, Blumberg EA, Punch JD, Limaye AP, et al. The efficacy and safety of 200 days valganciclovir cytomegalovirus prophylaxis in high-risk kidney transplant recipients. Am J Transplant. (2010) 10:1228–37. doi: 10.1111/j.1600-6143.2010.03074.x
29. Reischig T, Hribova P, Jindra P, Hes O, Bouda M, Treska V, et al. Long-term outcomes of pre-emptive valganciclovir compared with valacyclovir prophylaxis for prevention of cytomegalovirus in renal transplantation. J Am Soc Nephrol. (2012) 23:1588–97. doi: 10.1681/ASN.2012010100
30. Sylwester AW, Mitchell BL, Edgar JB, Taormina C, Pelte C, Ruchti F, et al. Broadly targeted human cytomegalovirus-specific CD4+ and CD8+ T cells dominate the memory compartments of exposed subjects. J Exp Med. (2005) 202:673–85. doi: 10.1084/jem.20050882
31. Ford ML. Virally-induced heterologous immunity in renal transplant recipients: important or inconsequential? Am J Transplant. (2016) 16:1348–9. doi: 10.1111/ajt.13657
32. Kern F, Faulhaber N, Frommel C, Khatamzas E, Prosch S, Schonemann C, et al. Analysis of CD8 T cell reactivity to cytomegalovirus using protein-spanning pools of overlapping pentadecapeptides. Eur J Immunol. (2000) 30:1676–82. doi: 10.1002/1521-4141(200006)30:6<1676::AID-IMMU1676>3.0.CO;2-V
33. Prosch S, Staak K, Stein J, Liebenthal C, Stamminger T, Volk HD, et al. Stimulation of the human cytomegalovirus IE enhancer/promoter in HL-60 cells by TNFalpha is mediated via induction of NF-kappaB. Virology. (1995) 208:197–206. doi: 10.1006/viro.1995.1143
34. Zeng G, Huang Y, Huang Y, Lyu Z, Lesniak D, Randhawa P. Antigen-specificity of T cell infiltrates in biopsies with T cell-mediated rejection and BK polyomavirus viremia: analysis by next generation sequencing. Am J Transplant. (2016) 16:3131–8. doi: 10.1111/ajt.13911
35. Amir AL, D'Orsogna LJ, Roelen DL, van Loenen MM, Hagedoorn RS, de Boer R, et al. Allo-HLA reactivity of virus-specific memory T cells is common. Blood. (2010) 115:3146–57. doi: 10.1182/blood-2009-07-234906
36. D'Orsogna LJ, Roelen DL, van der Meer-Prins EM, van der Pol P, Franke-van Dijk ME, Eikmans M, et al. Tissue specificity of cross-reactive allogeneic responses by EBV EBNA3A-specific memory T cells. Transplantation. (2011) 91:494–500. doi: 10.1097/TP.0b013e318207944c
37. Jackson SE, Sedikides GX, Okecha G, Poole EL, Sinclair JH, Wills MR. Latent Cytomegalovirus (CMV) infection does not detrimentally alter T cell responses in the healthy old, but increased latent CMV carriage is related to expanded CMV-specific T cells. Front Immunol. (2017) 8:733. doi: 10.3389/fimmu.2017.00733
Keywords: kidney transplantation, cytomegalovirus, ELISPOT, cross-reactivity, rejection, heterologous immunity, TCR repertoire
Citation: Stranavova L, Pelak O, Svaton M, Hruba P, Fronkova E, Slavcev A, Osickova K, Maluskova J, Hubacek P, Fronek J, Reinke P, Volk H-D, Kalina T and Viklicky O (2019) Heterologous Cytomegalovirus and Allo-Reactivity by Shared T Cell Receptor Repertoire in Kidney Transplantation. Front. Immunol. 10:2549. doi: 10.3389/fimmu.2019.02549
Received: 08 April 2019; Accepted: 14 October 2019;
Published: 31 October 2019.
Edited by:
Hermann Einsele, Julius Maximilian University of Würzburg, GermanyReviewed by:
Fred Falkenburg, Leiden University, NetherlandsMirko Trilling, University of Duisburg-Essen, Germany
Copyright © 2019 Stranavova, Pelak, Svaton, Hruba, Fronkova, Slavcev, Osickova, Maluskova, Hubacek, Fronek, Reinke, Volk, Kalina and Viklicky. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Ondrej Viklicky, b25kcmVqLnZpa2xpY2t5QGlrZW0uY3o=
†Shared first authorship
‡Shared senior authorship