 Imen Ayadi1†Saidou Balam2,3
Imen Ayadi1†Saidou Balam2,3 Régine Audran4
Régine Audran4 Jean-Pierre Bikorimana4†Issa Nebie5
Jean-Pierre Bikorimana4†Issa Nebie5 Mahamadou Diakité2Ingrid Felger6Marcel Tanner6
Mahamadou Diakité2Ingrid Felger6Marcel Tanner6 François Spertini4
François Spertini4 Giampietro Corradin1
Giampietro Corradin1 Myriam Arevalo7,8
Myriam Arevalo7,8 Socrates Herrera8
Socrates Herrera8 Valentina Agnolon4*
Valentina Agnolon4*- 1Biochemistry Department, University of Lausanne, Epalinges, Switzerland
- 2University Clinical Research Center (UCRC), University of Sciences, Techniques, and Technologies of Bamako (USTTB), Bamako, Mali
- 3Department of Internal Medicine II—Nephrology, University Hospital Regensburg, Regensburg, Germany
- 4Division of Immunology and Allergy, Centre Hospitalier Universitaire Vaudois (CHUV), Lausanne, Switzerland
- 5Centre National de Recherche et de Formation sur le Paludisme, Ouagadougou, Burkina Faso
- 6Department of Medical Parasitology and Infection Biology, Swiss Tropical and Public Health Institute, Basel, Switzerland
- 7Malaria Vaccine and Drug Development Center, Cali, Colombia
- 8Caucaseco Scientific Research Center, Cali, Colombia
Over the last four decades, significant efforts have been invested to develop vaccines against malaria. Although most efforts are focused on the development of P. falciparum vaccines, the current availability of the parasite genomes, bioinformatics tools, and high throughput systems for both recombinant and synthetic antigen production have helped to accelerate vaccine development against the P. vivax parasite. We have previously in silico identified several P. falciparum and P. vivax proteins containing α-helical coiled-coil motifs that represent novel putative antigens for vaccine development since they are highly immunogenic and have been associated with protection in many in vitro functional assays. Here, we selected five pairs of P. falciparum and P. vivax orthologous peptides to assess their sero-reactivity using plasma samples collected in P. falciparum- endemic African countries. Pf-Pv cross-reactivity was also investigated. The pairs Pf27/Pv27, Pf43/Pv43, and Pf45/Pv45 resulted to be the most promising candidates for a cross-protective vaccine because they showed a high degree of recognition in direct and competition ELISA assays and cross-reactivity with their respective ortholog. The recognition of P. vivax peptides by plasma of P. falciparum infected individuals indicates the existence of a high degree of cross-reactivity between these two Plasmodium species. The design of longer polypeptides combining these epitopes will allow the assessment of their immunogenicity and protective efficacy in animal models.
Introduction
Malaria disease was globally estimated at 228 million of cases in 2018 (1). Plasmodium falciparum and P. vivax are the two most important parasites in terms of infection prevalence and global distribution, and are responsible for more than 98% of global malaria clinical cases. In particular, P. falciparum is the most prevalent malaria parasite in Africa, while P. vivax is predominant in Asia, Oceania, and America (1).
Although a steady reduction in malaria transmission rates and deaths had been observed during the last two decades as a results of intensified control activities (1), an increase in malaria has been recently reported (2). Additionally, resistance to artemisinin, the main current anti-malarial treatment for P. falciparum, has been identified in several countries (3–6), and a reduction of the sensitivity of P. vivax parasites to antimalarials like chloroquine and primaquine has been reported (7–10). This represents an enormous challenge to malaria eradication and indicates the need for new control and elimination strategies. In particular, vaccination is considered as a potential cost-effective complement tool for malaria control and elimination. RTS,S/AS01 is the most advanced malaria vaccine candidate to date. It is based on P. falciparum circumsporozoite (Pf-CS) protein (11) and has been successfully tested in several Phase III trials in several African countries (12). Other P. falciparum candidates based on different approaches have undergone clinical development, such as the PfSPZ vaccine made of irradiated whole sporozoites (13, 14). On the other hand, only a few P. vivax vaccine candidates have advanced to Phase I and II vaccine trials (15–18) or are currently under preclinical development (Pvs48) (19). Despite this significant progress, a greater effort has to be invested on the development of malaria vaccines that could target the different parasite phases and species. Considering P. falciparum and P. vivax global distribution and infection rates, a vaccine providing cross-species protection would strategically reduce the vast majority of malaria clinical cases. The availability of the Plasmodium genome and proteome as well as of bioinformatics tools have allowed the identification of parasite proteins containing specific domains with functional importance such as α-helical coiled-coil motifs. Typically, these motifs are short conformationally-stable fragments (around 30/40 residues) composed of two to six α-helices wrapped around each other to form a left-handed supercoil. Each motif is characterized by heptad repeated regions denoted (abcdefg)n, where “a” and “d” are hydrophobic amino acids whereas the rest are usually hydrophilic residues (20–23). While the hydrophobic residues (a and d) are crucial for interhelical interactions, the hydrophilic residues are exposed on the surface of the coiled-coil motif and are likely important for interaction with other proteins. Alpha-helical coiled-coil domains can be rapidly produced by chemical synthesis and they auto-fold into their native structure. These peptides are perceived as novel putative antigens for vaccine development (22, 24) and have been investigated for protection against several diseases such as HIV (25), meningitis (26), and influenza (27). In the field of malaria, α-helical coiled-coil peptides were shown to be highly immunogenic in mice (22, 28, 29), with a strong association between antibody levels and clinical immunity (24, 30). Importantly, antibodies specifically directed against these motifs were tested in ex vivo biological assays and were shown to induce parasite growth inhibition (22, 29). In detail, α-helical coiled-coil peptides expressed in erythrocytic asexual-stage (22, 23, 28, 29) were in silico identified from P. falciparum (n = 166) and P. vivax (n = 50) genomes. The corresponding domains were synthesized and tested for their reactivity with sera of individuals from malaria-endemic areas (21, 28, 31).
In the present study, we investigate five α-helical coiled-coil domains that are orthologous between P. vivax and P. falciparum and share high sequence homology. The five ortholog pairs were selected because they were highly recognized by plasma samples from African malaria-endemic countries. The same panel of orthologs have been previously identified by Cespedes et al. as highly reactive with sera from malaria-endemic areas of Colombia and Papua New Guinea, thus being recognized as potential vaccine candidates (20). These peptides in fact were shown to be highly immunogenic in mice and induce antibodies able to recognize native proteins on P. vivax asexual blood stages. We expanded the investigation on the ability of α-helical coiled-coil domains to elicit antibodies with serological cross-reactivity, with the aim of selecting a set of antigenic peptides to be combined in a cross-protective polypeptide antigen for protection against both parasite species.
Materials and Methods
Peptides
Alpha-helical coiled-coil P. falciparum (n = 166) and P. vivax (n = 50) peptides had been formerly selected from the corresponding malaria proteomes (22, 23, 28). Peptides were chemically synthesized, HPLC purified, and characterized as previously described (22, 28). Based on the data published by Cespedes et al. on the recognition of 50 α-helical coiled coil peptides by human plasma samples, 38 orthologues peptide pairs that showed highest reactivity with plasma from Colombia and Papua New Guinea (PNG) were selected for the present study (28).
Human Plasma Samples
Plasma samples from Mali were collected in Kenieroba, Bozokin, and Fourda villages located in the Bancoumana town at 75 km from Bamako and in Dangassa village in the Kourouba town at 80 km from Bamako. Plasma were collected from 2009 to 2011 from 35 donors, among which seven were aged from three to 13 years old. Research and ethical clearance for the study was obtained from the Faculty of medicine, pharmacology and odonto-stomatology (FMPOS) of Bamako University (N°0840/FMPOS).
Tanzanian plasma was collected from 37 adult donors of a large-scale community based study undertaken in Ifakara village, Kilombero District, Morogoro Region from 1982 to 1984. Blood samples from adults were taken by finger prick and the serum was kept at −70°C until use. Research and ethical clearance for the study was obtained by the Tanzanian Commission for Science & Technology.
Plasma of eight adult donors from Burkina Faso were collected in the capital city of Ouagadougou. Ethical clearance was obtained from the Ministry of Health, Burkina Faso. After obtaining informed consent from parents and caretakers, heparinized venous blood samples were collected during a cross sectional survey during the malaria low transmission season 1998.
All samples were anonymized and kept at −80°C until use. Samples from Tanzania and Burkina Faso were also used in our previous studies (22, 29, 32). In all studies written informed consent (IC) was obtained from all adult study participants, and an informed assent (IA) was obtained for children through an IC from their parents and legal caretakers. Samples from Burkina Faso were collected during the malaria low transmission season and from donors living in the capital city of Ouagadougou, whereas Tanzanian and Malian samples were collected from residents of the countryside, where people are potentially more exposed to malaria-infected mosquitoes. Plasma samples used as negative controls were collected from Swiss naïve donors who had no history of malaria and no previous travel to malaria-endemic areas.
ELISA
ELISA was performed using Maxisorp 96-well plates (Thermo scientific, Ref 442404). Plates were coated overnight at 4°C with 5 μg/mL of each peptide. Plates were blocked for 1 h at room temperature (RT) with PBS 1X-milk 3% before being incubated with primary antibodies. Human plasma from Burkina Faso, Tanzania, and Mali donors were tested at the dilution 1:50 or 1:100 in titration two fold dilution series and incubated on plate for 2 h at RT. Goat anti-human IgG conjugated to horseradish peroxidase (HRP) was used as secondary antibody at dilution 1:2,000 (Life technologies, Ref H10307) and 1:1,000 (Invitrogen, Cat No 62-8420) for 1 h at RT. Signal was revealed using TMB substrate reagent (BD OptEIA, cat 555214) for 20 min in the dark at RT, and the reaction was blocked using 1 M sulphuric acid (Merck, 1.00731.1000). Optical density (OD) was measured at 450 nm and 630 nm using a TECAN NanoQuant Infinit M200 PRO spectrophotometer.
ELISA Competition Assays
In principle, P. falciparum or P. vivax peptides were adsorbed on the plate as previously described, and then plasma antibodies were incubated, at the dilution giving 50% of the maximal binding to each coated peptide, with a serial dilution of the respective antigen or its orthologue (from 500 μg/mL, 10 fold titration), and without peptide as control (no competition, ODmax). The resulting inhibited samples were then dispensed onto the coated wells (50 μL/well), and assay was carried on as previously described. Percentage of inhibition was calculated as: 100-(OD−ODmin)/(ODmax−ODmin) ×100, where ODmin is the signal in well without serum and ODmax is the signal resulting from samples incubated without competitor protein.
Data Analysis
The percentages of positive responses of serum samples from Mali, Tanzania, and Burkina Faso were evaluated as OD values higher than the mean negative control plus three times the standard deviation at serum dilution 1:200. Orthologue pairs were analyzed in the same experiment on the same day, thus allowing for direct comparison inside each pair. In order to allow for a direct and more informative comparison among peptide pairs, we transformed the OD values in antibody endpoint titers by using an internal positive control. First, OD values were converted to arbitrary units (AU) using the same positive control for each pair of orthologous peptides. The positive control for each couples was a plasma sample giving a similar good response to both orthologues, it was serially diluted and present on each ELISA plate. The 35 plasma Malian samples were individually tested in a twofold dilution series to establish a correlation curve between antibody titers and AU. At this point, antibody titers for all the other field samples were extrapolated from the correlation curve log (titer) = f [log(AU)], with a coefficient of variability between expected and extrapolated titers below 25%. For both P. falciparum and P. vivax peptides, titers were extrapolated from the same correlation curve in order to directly compare both orthologues. Friedman test followed by Dunn's multiple comparison test were performed for a collective analysis of titers from the three countries against all peptides.
In cross-reactivity experiments, samples were considered cross-reactive if positive to both orthologues. The percentage of cross-reactivity was calculated over the total of samples positive for at least one of the two orthologues.
Results and Discussion
P. falciparum and P. vivax α-Helical Coiled-Coil Peptides Are Similarly Recognized by Plasma From P. falciparum Endemic Regions
Previous work published by Cespedes et al. reported the recognition of 50 α-helical coiled coil peptides by human plasma samples from Colombia and Papua New Guinea (PNG), identifying 38 peptides that showed the highest sero-reactivity (28). In the present study we expanded the analysis of the 38 peptides and their orthologous by testing them with three panels of plasma samples obtained from African donors living in Mali, Tanzania or Burkina Faso (Supplementary Table 1). Plasma samples were collected during the ‘90s and the first decade of the current century, when these countries were endemic for P. falciparum with no P. vivax detection (33, 34). Indeed, low prevalence of P. vivax was only recently reported in Mali (35, 36). From the panel of 38 P. falciparum and P. vivax orthologues, five were highly recognized by plasma samples from African malaria-endemic countries and were selected for further studies (Table 1). Notably, these five pairs coincided with the P. vivax peptides identified by Cespedes et al. (28) as potential vaccine candidates. The P. falciparum orthologues Pf 27, Pf 43, Pf 45, Pf 82.02, and Pf 96.03 were the only peptides recognized by at least 90% of the tested samples, with their P. vivax orthologues being recognized by 69 to 98% of the samples (Supplementary Table 1).
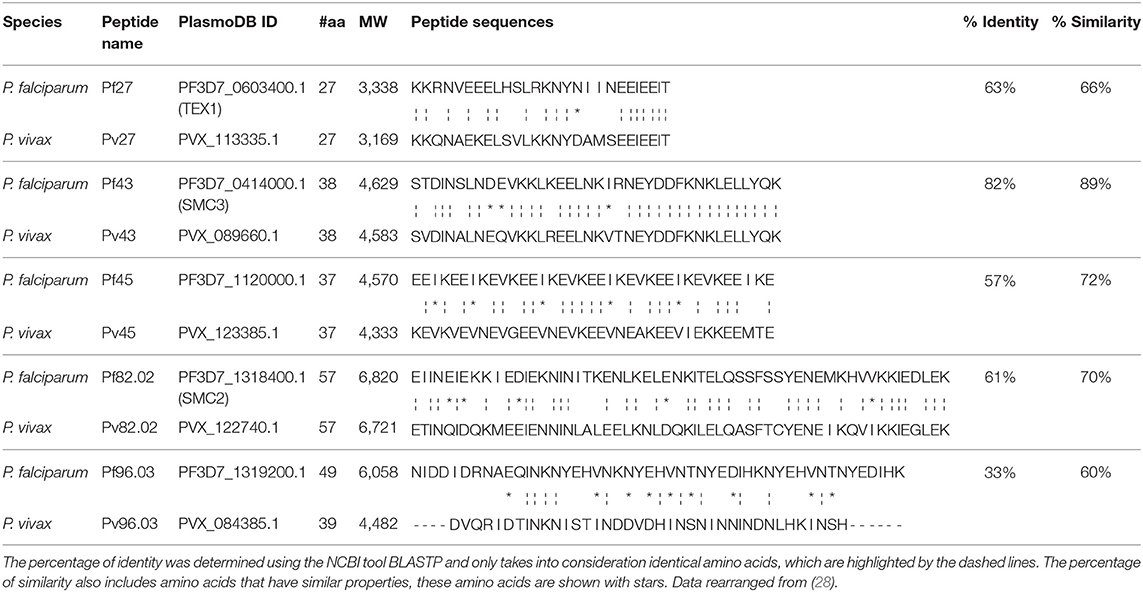
Table 1. Homology between P. falciparum and P. vivax peptides.
The selected five pairs of orthologous peptides have lengths between 27 and 59 amino acids and display sequence identities ranging from 33 to 82% and a similarity higher than 60% (Table 1). Orthologues Pf 27/Pv27, Pf 43/Pv43, Pf 45/Pv45, and Pf 82.02/Pv82.02 share more than 50% identity (82, 63, 61, and 57%, respectively), while the couple Pf 96.03/Pv96.03 have the lowest identity percentage (33%). However, this low Pf/Pv identity appears to be balanced by a 60% similarity that results in 100% recognition of the P. falciparum and 98% of the P. vivax orthologues by the African sample panel (Supplementary Table 1), underlining the importance of charged amino acids for antigen-antibody binding.
The antigenicity of the orthologous coiled-coil peptides was analyzed with the panel of plasma from African donors, and median OD values for each peptide together with percentages of positive responses are summarized in Table 2. No significant difference was observed among the median OD of P. falciparum peptides and their P. vivax orthologue. A complete descriptive analysis showing direct comparison between the recognition of P. falciparum vs. P. vivax coiled-coil peptides and each plasma in the African donors panel is provided in Supplementary Figure 1. Globally, from Table 2 and Supplementary Figure 1 we can recognize the orthologues pair Pf 45/Pv45 as the one resulting in the highest OD values. As an additional observation, the recognition of P. vivax peptides by plasma samples from P. falciparum endemic regions is an indication of cross-reactivity between the two Plasmodium species that could be exploited for the generation of a cross-protective vaccine. This type of analysis could now be expanded to P. ovale and P. malariae that are prevalent in most of the regions where malaria in endemic. However, their genomes have only been available in the last 2 years, but not at the time of peptide selection and synthesis (37).
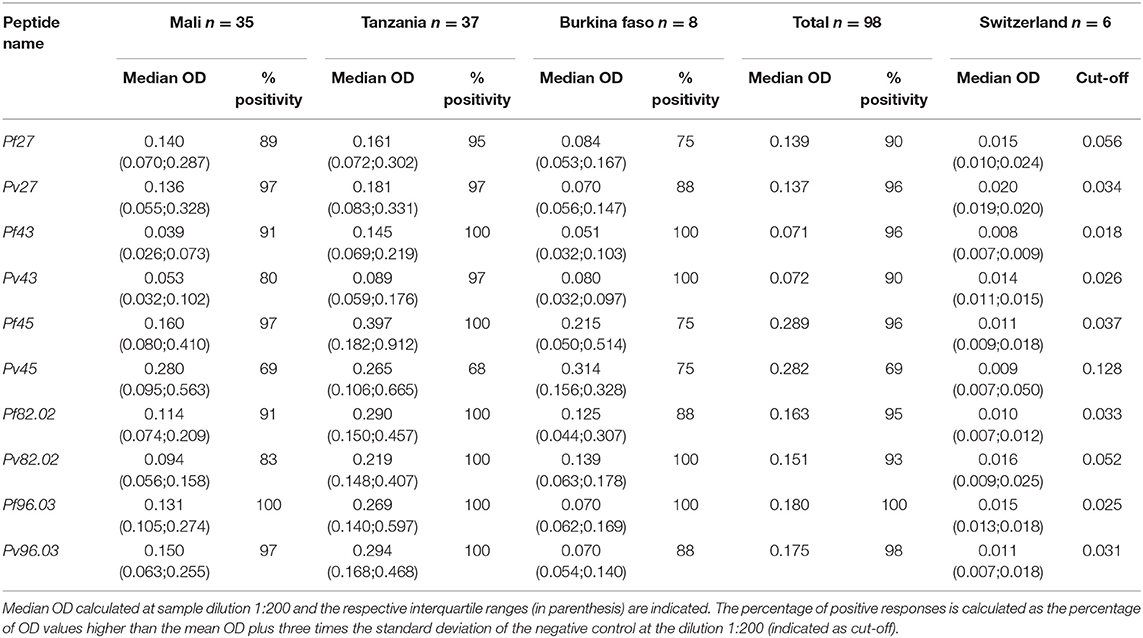
Table 2. Pf and Pv orthologous peptides are both recognized by African samples.
Antibody response to the five pairs of orthologous peptides (Figure 1) showed no statistical difference between P. falciparum vs. P. vivax titers when data from each individual country were analyzed. However, we could identify a particular trend in plasma from Burkina Faso showing the lowest titers probably because they were collected during the low transmission season and from donors living in the capital that are potentially less exposed to Plasmodium-infected mosquitoes. Similarly, plasma from Mali showed generally lower titers than plasma from Tanzania. Indeed, the former included children and adolescents among the donors panel, which have statistically had fewer contacts with Plasmodium-infected mosquitos. No significant difference was observed between Pf and Pv titers when all ELISA results from the three countries were combined. These results strongly suggest a cross-reactivity between P. falciparum and P. vivax coiled-coil peptides. As described by Olugbile et al. (29), the synthesis of a candidate vaccine consisting of two or more polypeptides will permit to maximize the proportion of the population that will respond to such a candidate while conserving the individual functional capacities of each constituent peptide. With reference to our data, we found titers of peptide 82.02 from both Plasmodium species to be significantly lower than those of the other peptides (Figure 1), identifying this pair as not appealing to be included in the combined polypeptide vaccine. In addition, the peptides pair 82.02 showed the biggest difference between the titers of the P. falciparum or P. vivax peptide (Figure 2). Concerning the other pairs, antibody titers resulted to be similar between each ortholog, and the ratio between P. falciparum and P. vivax peptide titers was close to one (Figure 2), suggesting a good cross-reactivity that might contribute to cross-species clinical protection.
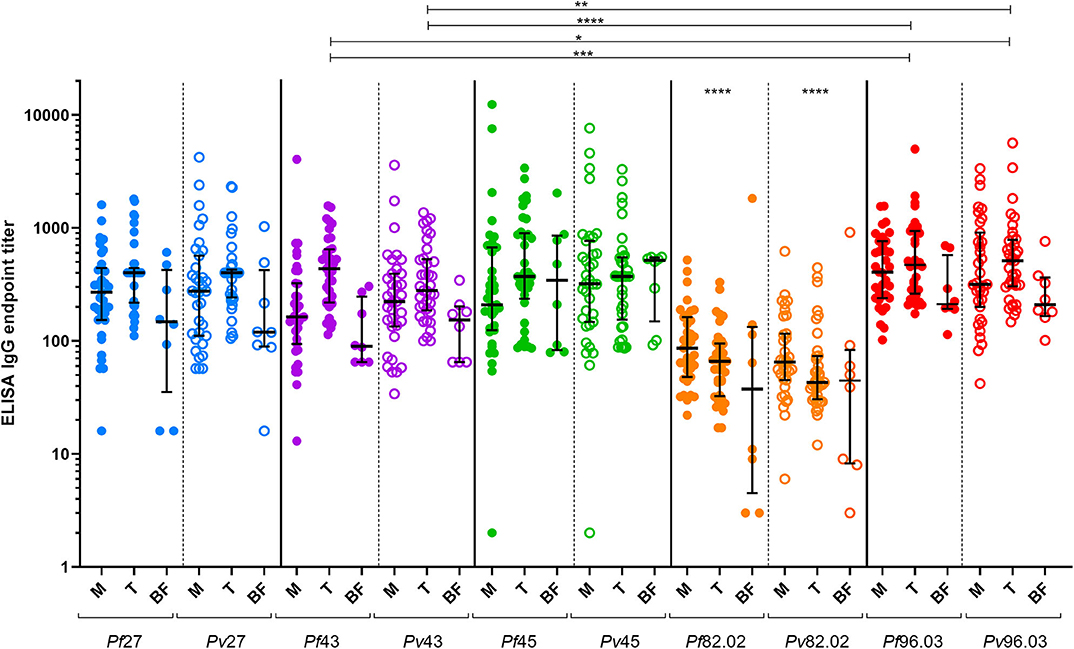
Figure 1. Antibody response to P. falciparum and P. vivax orthologous peptides. The dot-plot graph shows IgG endpoint titers to the different pairs of orthologous peptides tested with samples from Mali (M, n = 35), Tanzania (T, n = 37), and Burkina Faso (BF, n = 8). Median and interquartile ranges (25th and 75th percentiles) are shown. Median titers by peptide considering the three countries collectively: Pf27 = 335; Pv27 = 354; Pf43 = 237; Pv43 = 246; Pf45 = 315; Pv45 = 371; Pf82.02 = 68; Pv82.02 = 54; Pf96.03 = 429; Pv96.03 = 371. Friedman test followed by Dunn's multiple comparison test were performed. Statistical significance represented in the graph is referred to a collective analysis of titers from the three countries.
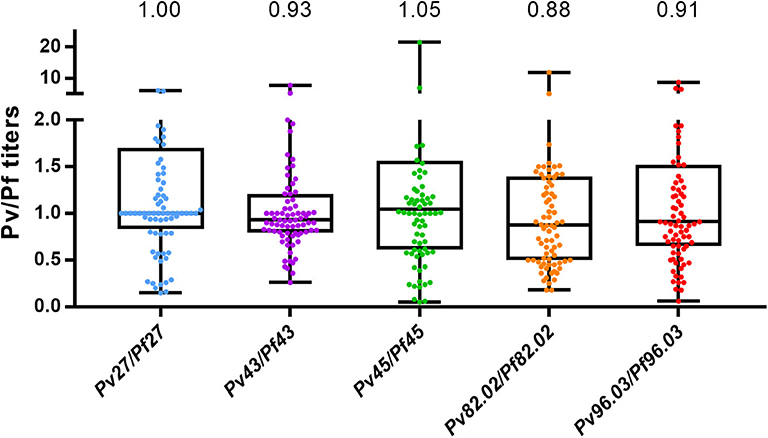
Figure 2. P. falciparum and P. vivax orthologous peptides are similarly recognized. The graph shows the titer ratio between Pf and Pv orthologues. Peptides were tested with samples from Mali (n = 35), Tanzania (n = 37), and Burkina Faso (n = 8) and results of the three countries are combined. Median and interquartile range (25th and 75th percentiles) are shown. Values the median. One dot corresponds to one sample. Kruskal Wallis followed by Dunn's multiple comparison test: no significant differences.
Cross-Reactivity Between P. falciparum and P. vivax α-Helical Coiled-Coil Peptides
To assess the frequency of plasma samples recognizing both orthologous peptides, we introduced a definition of positivity stricter than that applied before and only plasma samples that were positive for at least one of the two orthologues and had a titer higher than 200 were considered. Pairs Pf 27/Pv27, Pf 43/Pv43, and Pv96.03/Pv96.03 were recognized in more than 60% of the samples (62, 71, and 78%, respectively) (Table 3). Surprisingly, recognition of both peptides of pairs Pf 45/Pv45 and Pf 82.02/Pv82.02 was <50%, even though the similarity percentage between these two pairs of orthologues is around 70% and thus higher than that of the more cross-reactive pairs Pf 27/Pv27 and Pv96.03/Pv96.03, as shown in Table 1.
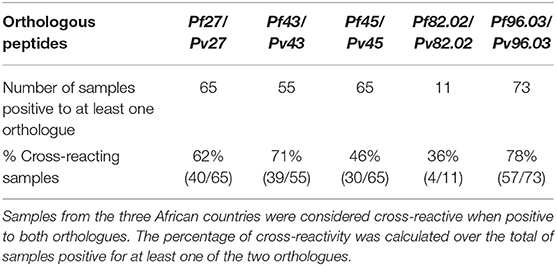
Table 3. Frequency of donors recognizing both Pf and Pv orthologous peptides.
To experimentally analyse cross-reactivity between P. falciparum and P. vivax orthologous peptides, competition ELISA were performed on each peptide. Competitors were the peptide itself or its ortholog. In Tables 4A,B (left panels), we reported the results of competition ELISA for a small panel of plasma for each peptide as representative of all the others. We identified few cases of cross-reactive inhibition, for example plasma A005 and KA006 were almost completely inhibited by Pf 27 and Pv27, resulting in a percentage of inhibition higher than 80%, the coated peptide being the same as the competitor or its ortholog. Overall, the results of competition ELISA were diverse, ranging from complete to zero inhibition, irrespective if the peptide used as inhibitor was the same as the coated one or its orthologue. We initially interpreted this as an effect of the lower antibody-antigen affinity in solution compared to the avidity of antibodies for an antigen adsorbed to the plate surface, which, together with their three-dimensional coiled-coil structure, is favored by the improved proximity among them. To investigate this possibility, we tested if the recognition among the panel of African plasma and the peptides was really specific. When testing each plasma sample without coating or in presence of the coiled coil peptides, some plasma gave a background signal equal or higher than the actual antigen-antibody reaction (Tables 4A,B, right panels).
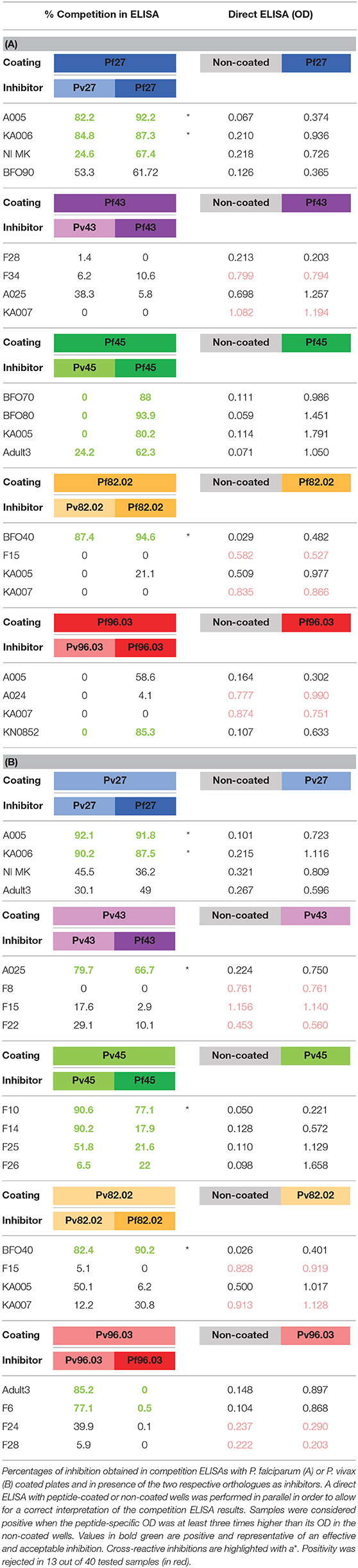
Table 4. Percentages of inhibition obtained in competition ELISAs.
This phenomenon has been already reported in the literature (38–41) and the term serum-specific background noise (SSBN) has been coined for this observation known to affect 4–32% of sera, especially when tested individuals had recent or ongoing bacterial infections at the time of blood sampling. In presence of SSBN, subtraction of the aspecific signal from the antigen-specific signal is suggested before data analysis, in order to increase assay sensitivity (38, 40). In the present investigation we decided to apply a more strict rule. Each plasma sample was tested without coating or in the presence of the coiled coil peptides, and was considered positive when its peptide-specific OD value was at least three times its OD value in the non-coated wells (Tables 4A,B, right panels). As a result, positivity was rejected in 13 out of 40 tested samples (highlighted in red in Table 4), corresponding to a 32.5% of samples with aspecific reactivity. Frequently, samples with background noise were associated to absence of inhibition with both orthologues. This is for example the case for plasma KA007, which gave similar high background with different peptides, confirming this effect is indeed sample-specific rather than due to the coating. Given these results, we wish to highlight their importance in order to raise awareness of SSBN in the scientific community. This issue should be taken into consideration—as in the present study—for proper interpretation of ELISA results. Indeed, SSBN analysis led us to identify real inhibition in almost half (19/40) of the samples tested (highlighted in bold and green in Table 4).
Particularly, Pf27/Pv27 and Pf45/Pv45 showed clear cross reactivity with the potential to be selected as candidates. Interestingly, these peptides have been previously tested for their antigenicity in sera obtained from donors from both P. falciparum and P. vivax endemic countries, and purified specific IgG antibodies showed capacity to recognize native proteins in IFAT and functional activity in ADCI assays (22, 28, 29). In addition, association of antibody responses with protection against infection from both parasites was observed (24, 29). Such results support the selection of Pf27/Pv27 and Pf45/Pv45 as promising candidates to be combined in a polypeptide vaccine conferring cross-protection against P. falciparum and P. vivax. The combination of two or eventually more antigens as a single product is cost-effective in terms of vaccine manufacturing and easy to scale-up.
Conclusions
We showed the recognition of P. vivax peptides by plasma of P. falciparum infected people living in Mali, Tanzania, and Burkina Faso, thus establishing the existence of a high degree of cross-reactivity between the ortholog coiled-coil domains of the two Plasmodium species. Current studies are focused on the development of hybrid nanoparticles, as recently described by Karch et al. (42), containing P. falciparum and P. vivax cross-reactive coiled-coil domains capable of eliciting a protective response toward the two parasites in experimental animals.
Data Availability Statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Ethics Statement
The studies involving human participants were reviewed and approved by Faculty of medicine, pharmacology and odonto-stomatology (FMPOS) of Bamako University (N°0840/FMPOS); Tanzanian Commission for Science & Technology; Ministry of Health, Burkina Faso. Written informed consent to participate in this study was provided by the participants' legal guardian/next of kin.
Author Contributions
GC, MA, and SH: conceived the study. IA, J-PB, and VA: performed the experiments. IA, RA, and VA: analyzed the data. SB, IN, MD, IF, and MT: contributed with reagents and material. IA: writing-original draft. VA, RA, IF, FS, and GC: writing-review and editing. All authors discussed the results, contributed to the revision of the manuscript, and approved the final version.
Funding
Studies were supported by the NIAID-NIH Grant No. R01AI148843.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The reviewer TT declared a past co-authorship with one of the author MD to the handling editor.
Acknowledgments
We are grateful for the participation of the community from malaria-endemic countries of Mali, Tanzania and Burkina Faso, as well as Swiss volunteers and Dr. Howard Hengers for critical reading of the manuscript.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fimmu.2020.574330/full#supplementary-material
References
2. Dhiman S. Are malaria elimination efforts on right track? An analysis of gains achieved and challenges ahead. Infect Dis Poverty. (2019) 8:14. doi: 10.1186/s40249-019-0527-7
3. WHO. Status Report on Artemisinin Resistance and ACT Efficacy. Geneva (2018). (accessed August 10, 2018).
4. Ikeda M, Kaneko M, Tachibana S-I, Balikagala B, Sakurai-Yatsushiro M, Yatsushiro S, et al. Artemisinin-resistant Plasmodium falciparum with high survival rates, Uganda, 2014–2016. Emerg Infect Dis. (2018) 24:718. doi: 10.3201/eid2404.170141
5. Oboh MA, Ndiaye D, Antony HA, Badiane AS, Singh US, Ali NA, et al. Status of artemisinin resistance in Malaria parasite Plasmodium falciparum from molecular analyses of the Kelch13 gene in Southwestern Nigeria. BioMed Res Int. (2018) 2018:2305062. doi: 10.1155/2018/2305062
7. Commons RJ, Simpson JA, Thriemer K, Humphreys GS, Abreha T, Alemu SG, et al. The effect of chloroquine dose and primaquine on Plasmodium vivax recurrence: a worldwide antimalarial resistance network systematic review and individual patient pooled meta-analysis. Lancet Infect Dis. (2018) 18:1025–34. doi: 10.1016/S1473-3099(18)30348-7
8. Ladeia-Andrade S, Menezes MJ, de Sousa TN, Silvino ACR, de Carvalho JF, Salla LC, et al. Monitoring the efficacy of chloroquine-primaquine therapy for uncomplicated Plasmodium vivax malaria in the main transmission hot spot of Brazil. Antimicrob Agents Chemother. (2019) 63:e01965–18. doi: 10.1128/AAC.01965-18
9. Silva SR, Almeida ACG, da Silva GAV, Ramasawmy R, Lopes SCP, Siqueira AM, et al. Chloroquine resistance is associated to multi-copy pvcrt-o gene in Plasmodium vivax malaria in the Brazilian Amazon. Malaria J. (2018) 17:267. doi: 10.1186/s12936-018-2411-5
10. Rieckmann K, Davis D, Hutton D. Plasmodium vivax resistance to chloroquine? Lancet. (1989) 334:1183–4. doi: 10.1016/S0140-6736(89)91792-3
11. Cohen J, Nussenzweig V, Vekemans J, Leach A. From the circumsporozoite protein to the RTS, S/AS candidate vaccine. Hum Vacc. (2010) 6:90–6. doi: 10.4161/hv.6.1.9677
12. Moncunill G, de Rosa SC, Ayestaran A, Nhabomba AJ, Mpina M, Cohen KW, et al. S/AS01E malaria vaccine induces memory and polyfunctional T cell responses in a pediatric African phase III trial. Front Immunol. (2017) 8:1008. doi: 10.3389/fimmu.2017.01008
13. Seder R, Chang L, Enama M, Zephir K, Sarwar U, Gordon I, et al. VRC 312 study team 2013 protection against malaria by intravenous immunization with a nonreplicating sporozoite vaccine. Science. (2013) 341:1359–65. doi: 10.1126/science.1241800
14. Mordmüller B, Surat G, Lagler H, Chakravarty S, Ishizuka AS, Lalremruata A, et al. Sterile protection against human malaria by chemoattenuated PfSPZ vaccine. Nature. (2017) 542:445–9. doi: 10.1038/nature21060
15. Mueller I, Shakri AR, Chitnis CE. Development of vaccines for Plasmodium vivax malaria. Vaccine. (2015) 33:7489–95. doi: 10.1016/j.vaccine.2015.09.060
16. Herrera S, Bonelo A, Perlaza BL, Fernandez OL, Victoria L, Lenis AM, et al. Safety and elicitation of humoral and cellular responses in Colombian malaria-naive volunteers by a Plasmodium vivax circumsporozoite protein–derived synthetic vaccine. Am J Trop Med Hygiene. (2005) 73:3–9. doi: 10.4269/ajtmh.2005.73.3
17. Herrera S, Bonelo A, Perlaza BL, Valencia AZ, Cifuentes C, Hurtado S, et al. Use of long synthetic peptides to study the antigenicity and immunogenicity of the Plasmodium vivax circumsporozoite protein. Int J Parasitol. (2004) 34:1535–46. doi: 10.1016/j.ijpara.2004.10.009
18. Yadava A, Sattabongkot J, Washington MA, Ware LA, Majam V, Zheng H, et al. A novel chimeric Plasmodium vivax circumsporozoite protein induces biologically functional antibodies that recognize both VK210 and VK247 sporozoites. Infect Immun. (2007) 75:1177–85. doi: 10.1128/IAI.01667-06
19. Arévalo-Herrera M, Vallejo AF, Rubiano K, Solarte Y, Marin C, Castellanos A, et al. Recombinant Pvs48/45 antigen expressed in E. coli generates antibodies that block malaria transmission in Anopheles albimanus mosquitoes. PLoS ONE. (2015) 10:e0119335. doi: 10.1371/journal.pone.0119335
20. Tripet B, Wagschal K, Lavigne P, Mant CT, Hodges RS. Effects of side-chain characteristics on stability and oligomerization state of a de Novo-designed model coiled-coil: 20 amino acid substitutions in position “d” 1. J Mol Biol. (2000) 300:377–402. doi: 10.1006/jmbi.2000.3866
21. Corradin G, Villard V, Kajava AV. Protein structure based strategies for antigen discovery and vaccine development against malaria and other pathogens. Endocr Metab Immune Disord Drug Targets. (2007) 7:259–65. doi: 10.2174/187153007782794371
22. Villard V, Agak GW, Frank G, Jafarshad A, Servis C, Nébié I, et al. Rapid identification of malaria vaccine candidates based on α-helical coiled coil protein motif. PLoS ONE. (2007) 2:e645. doi: 10.1371/journal.pone.0000645
23. Kulangara C, Kajava AV, Corradin G, Felger I. Sequence conservation in Plasmodium falciparum α-helical coiled coil domains proposed for vaccine development. PLoS ONE. (2009) 4:e5419. doi: 10.1371/journal.pone.0005419
24. Céspedes N C, Suen SLW, Koepfli C, França CT, Felger I, Nebie I, et al. Natural immune response to Plasmodium vivax alpha-helical coiled coil protein motifs and its association with the risk of P. vivax malaria. PLoS ONE. (2017) 12:e0179863. doi: 10.1371/journal.pone.0179863
25. Tong P, Lu Z, Chen X, Wang Q, Yu F, Zou P, et al. An engineered HIV-1 gp41 trimeric coiled coil with increased stability and anti-HIV-1 activity: implication for developing anti-HIV microbicides. J Antimicrob Chemother. (2013) 68:2533–44. doi: 10.1093/jac/dkt230
26. Liguori A, Iacono LD, Maruggi G, Benucci B, Merola M, Surdo PL, et al. NadA3 structures reveal undecad coiled coils and LOX1 binding regions competed by meningococcus B vaccine-elicited human antibodies. mBio. (2018) 9:e01914–18. doi: 10.1128/mBio.01914-18
27. Jiang Z, Gera L, Mant CT, Hirsch B, Yan Z, Shortt JA, et al. Platform technology to generate broadly cross-reactive antibodies to α-helical epitopes in hemagglutinin proteins from influenza A viruses. Peptide Sci. (2016) 106:144–59. doi: 10.1002/bip.22808
28. Cespedes N, Habel C, Lopez-Perez M, Castellanos A, Kajava AV, Servis C, et al. Plasmodium vivax antigen discovery based on alpha-helical coiled coil protein motif. PLoS ONE. (2014) 9:e100440. doi: 10.1371/journal.pone.0100440
29. Olugbile S, Villard V, Bertholet S, Jafarshad A, Kulangara C, Roussilhon C, et al. Malaria vaccine candidate: design of a multivalent subunit α-helical coiled coil poly-epitope. Vaccine. (2011) 29:7090–9. doi: 10.1016/j.vaccine.2011.06.122
30. Agak GW, Bejon P, Fegan G, Gicheru N, Villard V, Kajava AV, et al. Longitudinal analyses of immune responses to Plasmodium falciparum derived peptides corresponding to novel blood stage antigens in coastal Kenya. Vaccine. (2008) 26:1963–71. doi: 10.1016/j.vaccine.2008.02.020
31. Hviid L. Clinical disease, immunity and protection against Plasmodium falciparum malaria in populations living in endemic areas. Expert Rev Mol Med. (1998) 1998:1–10. doi: 10.1017/S1462399498000179
32. Olugbile S, Kulangara C, Bang G, Bertholet S, Suzarte E, Villard V, et al. Vaccine potentials of an intrinsically unstructured fragment derived from the blood stage-associated Plasmodium falciparum protein PFF0165c. Infect Immun. (2009) 77:5701–9. doi: 10.1128/IAI.00652-09
34. Kathrine PMA, Tan R, Stefanie Steele F. Yellow Fever & Malaria Information, by Country, CDC Yellow Book 2018: Health Information for International Travel. Oxford: Oxford University Press (2017).
35. Gunalan K, Niangaly A, Thera MA, Doumbo OK, Miller LH. Plasmodium vivax infections of duffy-negative erythrocytes: historically undetected or a recent adaptation? Trends Parasitol. (2018) 35:420–9. doi: 10.1016/j.pt.2018.02.006
36. Niangaly A, Gunalan K, Ouattara A, Coulibaly D, Sa JM, Adams M, et al. Plasmodium vivax infections over 3 years in duffy blood group negative Malians in Bandiagara, Mali. Am J Trop Med Hygiene. (2017) 97:744–52. doi: 10.4269/ajtmh.17-0254
37. Rutledge GG, Böhme U, Sanders M, Reid AJ, Cotton JA, Maiga-Ascofare O, et al. Plasmodium malariae and P. ovale genomes provide insights into malaria parasite evolution. Nature. (2017) 542:101–4. doi: 10.1038/nature21038
38. Güven E, Duus K, Lydolph MC, Jørgensen CS, Laursen I, Houen G. Non-specific binding in solid phase immunoassays for autoantibodies correlates with inflammation markers. J Immunol Methods. (2014) 403:26–36. doi: 10.1016/j.jim.2013.11.014
39. Terato K, Do CT, Cutler D, Waritani T, Shionoya H. Preventing intense false positive and negative reactions attributed to the principle of ELISA to re-investigate antibody studies in autoimmune diseases. J Immunol Methods. (2014) 407:15–25. doi: 10.1016/j.jim.2014.03.013
40. Moritz CP, Tholance Y, Lassablière F, Camdessanché J-P, Antoine J-C. Reducing the risk of misdiagnosis of indirect ELISA by normalizing serum-specific background noise: the example of detecting anti-FGFR3 autoantibodies. J Immunol Methods. (2019) 466:52–6. doi: 10.1016/j.jim.2019.01.004
41. Haberland A, Müller J, Wallukat G, Wenzel K. Antigen-free control wells in an ELISA set-up for the determination of autoantibodies against G protein-coupled receptors—a requisite for correct data evaluation. Anal Bioanal Chem. (2018) 410:5101–5. doi: 10.1007/s00216-018-1172-x
Keywords: malaria, plasmodium falciparum, plasmodium vivax, vaccine, coiled-coil peptides, immune response
Citation: Ayadi I, Balam S, Audran R, Bikorimana J-P, Nebie I, Diakité M, Felger I, Tanner M, Spertini F, Corradin G, Arevalo M, Herrera S and Agnolon V (2020) P. falciparum and P. vivax Orthologous Coiled-Coil Candidates for a Potential Cross-Protective Vaccine. Front. Immunol. 11:574330. doi: 10.3389/fimmu.2020.574330
Received: 19 June 2020; Accepted: 11 September 2020;
Published: 21 October 2020.
Edited by:
Urszula Krzych, Walter Reed Army Institute of Research, United StatesReviewed by:
Takafumi Tsuboi, Ehime University, JapanLilian Rose Pratt-Riccio, Oswaldo Cruz Foundation (Fiocruz), Brazil
Sean C. Murphy, University of Washington, United States
Copyright © 2020 Ayadi, Balam, Audran, Bikorimana, Nebie, Diakité, Felger, Tanner, Spertini, Corradin, Arevalo, Herrera and Agnolon. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Valentina Agnolon, dmFsZW50aW5hLmFnbm9sb25AY2h1di5jaA==
†Present address: Imen Ayadi, Department of Cell Physiology and Metabolism, University of Geneva, Geneva, Switzerland
Jean-Pierre Bikorimana, Department of Pharmacology and Physiology, University of Montreal, Montreal, QC, Canada