 Hector Teran-Navarro1†
Hector Teran-Navarro1† David Salcines-Cuevas1,2†
David Salcines-Cuevas1,2† Ricardo Calderon-Gonzalez1,2‡
Ricardo Calderon-Gonzalez1,2‡ Raquel Tobes3Jorge Calvo-Montes4Inmaculada Concepción Pérez-Del Molino Bernal4Sonsoles Yañez-Diaz5,6Manuel Fresno7,8
Raquel Tobes3Jorge Calvo-Montes4Inmaculada Concepción Pérez-Del Molino Bernal4Sonsoles Yañez-Diaz5,6Manuel Fresno7,8 Carmen Alvarez-Dominguez9*
Carmen Alvarez-Dominguez9*- 1Instituto de Investigación Marqués de Valdecilla (IDIVAL), Santander, Spain
- 2Grupo de Oncología y Nanovacunas, Santander, Spain
- 3Alamo Blanco, Granada, Spain
- 4Microbiology Department, Hospital Universitario Marqués de Valdecilla, Santander, Spain
- 5Dermatology Department, Hospital Universitario Marqués de Valdecilla, Santander, Spain
- 6Facultad de Medicina, Universidad de Cantabria, Santander, Spain
- 7DIOMUNE S.L., Parque Científico de Madrid, Madrid, Spain
- 8Centro de Biología Molecular Severo Ochoa, Universidad Autónoma de Madrid, Madrid, Spain
- 9Facultad de Educación y Facultad de Ciencias de la Salud, Universidad Internacional de La Rioja, Logroño, Spain
Cross-reactive vaccines recognize common molecular patterns in pathogens and are able to confer broad spectrum protection against different infections. Antigens common to pathogenic bacteria that induce broad immune responses, such as the glyceraldehyde-3-phosphate dehydrogenase (GAPDH) of the genera Listeria, Mycobacterium, or Streptococcus, whose sequences present more than 95% homology at the N-terminal GAPDH1−22 peptide, are putative candidates for universal vaccines. Here, we explore vaccine formulations based on dendritic cells (DC) loaded with two molecular forms of Listeria monocytogenes GAPDH (LM-GAPDH), such as mRNA carriers or recombinant proteins, and compare them with the same molecular forms of three other antigens used in experimental vaccines, listeriolysin O of Listeria monocytogeness, Ag85A of Mycobacterium marinum, and pneumolysin of Streptococcus pneumoniae. DC loaded with LM-GAPDH recombinant proteins proved to be the safest and most immunogenic vaccine vectors, followed by mRNA encoding LM-GAPDH conjugated to lipid carriers. In addition, macrophages lacked sufficient safety as vaccines for all LM-GAPDH molecular forms. The ability of DC loaded with LM-GAPDH recombinant proteins to induce non-specific DC activation explains their adjuvant potency and their capacity to trigger strong CD4+ and CD8+ T cell responses explains their high immunogenicity. Moreover, their capacity to confer protection in vaccinated mice against challenges with L. monocytogenes, M. marinum, or S. pneumoniae validated their efficiency as cross-reactive vaccines. Cross-protection appears to involve the induction of high percentages of GAPDH1−22 specific CD4+ and CD8+ T cells stained for intracellular IFN-γ, and significant levels of peptide-specific antibodies in vaccinated mice. We concluded that DC vaccines loaded with L. monocytogenes GAPDH recombinant proteins are cross-reactive vaccines that seem to be valuable tools in adult vaccination against Listeria, Mycobacterium, and Streptococcus taxonomic groups.
Introduction
Vaccines for adults is one of the biggest challenges of current vaccinology and several methodologies have been proposed for this purpose such as reverse vaccinology, a genome-based approach to vaccine development (1), or immune algorithm approaches (2–4). One of the main issues regarding vaccines for adults is the possibility to prepare bacterial vaccines that induce cross-protection against infections caused by different pathogens that provide cellular specific immunity, involving both T and B cells, known as cross-reactive vaccines (CRV). However, cross-protection against infections can also be achieved if innate immune cells acquire long functional states such as in trained immunity-based vaccines (TIbV) (5). Dendritic cells (DC) are pivotal cells for conventional, CRV, or TIbV vaccines and serve as efficient vaccine platforms. In this regard, DC based vaccines can recognize non-specific patterns in pathogens and can induce specific immunity (5–7), allowing cross-protection against infections. In fact, the COVID-19 pandemic has highlighted the possibility that vaccines designed for unrelated pathogens such as Mycobacterium bovis Bacillus Calmette-Guérin (BCG), could also confer some protection for a coronavirus (8, 9).
Bacterial pathogens such as Mycobacterium tuberculosis, Listeria monocytogenes, or Streptococcus pneumonia can cause severe meningitis both in the elderly and in adults with immunocompromising conditions, such as cancer patients, in all cases that require long-term antibiotic treatment (10). Opportunistic skin diseases, mild or severe, caused in adults by Mycobacterium marinum, Mycobacterium chelonae, Mycobacterium fortuitum, Listeria monocytogenes, or Streptococcus pyogenes also require long-term treatment with antibiotics that might contribute to the development of antibiotic resistance (11–13). On the other hand, there are no vaccines available for meningitis or severe skin diseases in the elderly (14). Preparing DC based vaccines that can cross-protect against bacterial genera of Listeria, Mycobacterium, or Streptococcus might therefore provide relevant tools for adult vaccination.
Poly-bacterial preparations such as MV130 (Bactek®) are composed of heat-inactivated bacteria with 90% gram-positive bacteria (Streptococcus pneumoniae, Staphylococcus aureus, Staphylococcus epidermidis) and 10% gram-negative bacteria (Klebsiella pneumoniae, Haemophilus influenza, and Moraxella catarrhalis) (15). MV130 acts as adjuvant and improves recurrent respiratory tract infections by inducing a specific T cell immunity against bacteria present in the preparation, but also with T cell responses to other different antigens (16, 17). The ability of MV130 to immunomodulate DC, implies the triggering of Toll-like (TLR) and Nod-like receptors (NLR) with the ability to stimulate Th1 and Th17 immune responses and increases the levels of IL-10 (18). Other bacterial adjuvants such as DIO-1, a lipopolysaccharide of Ochrobactrum intermedium that acts as a TLR-2/4 agonist, is also able to immunomodulate DC, inducing Th1 immune responses and conferring protection against experimental listeriosis in different vaccine formulations (19–21).
Bacterial ADP-ribosylating enterotoxins such as the heat-labile enterobacterial toxin subunit of Escherichia coli (LT), or the cholera toxin (CT) are also used as adjuvants as they promote multifaced antigen-specific responses inducing Th1, Th2, and Th17 patterns. The availability of LT and CT mutants lacking toxicity have allowed these bacterial toxins to be included in vaccine designs, as they retain their adjuvant capacities (22). Other bacterial enzymes with ADP-ribosylating abilities are the glyceraldehyde-3-phosphage dehydrogenases (GAPDH) of gram-positive bacteria, also proposed as universal vaccines against different Streptococcus serotypes, since they induce broad spectrum immune responses (23). Our group also described that the GAPDH of L. monocytogenes (GAPDH-LM, Lmo 2459), which also presents ADP-ribosylating abilities (24), showed two interesting abilities for vaccine designs—a 22 amino acid peptide at the N-terminal that presented 95–98% sequence homology to GAPDH of Mycobacterium and Streptococcus and the ability of anti-Listeria GAPDH antibodies to recognize Mycobacterium or Streptococcus spp (25–28).
Messenger RNA (mRNA) is a promising vehicle for vaccination (29), however, naked mRNA suffers a quick degradation by RNases activity and is consequently not internalized efficiently. Several delivery carriers for mRNA vaccines have been developed, mostly based on lipid particulate complexes. Typical examples are the COVID-19 vaccines by Moderna and Pfizer-BioNTech and others such as nanoparticles (30–33). In this regard, cationic lipids commercially available, such as lipofectamine (Invitrogen), can also serve as protective capsules to incorporate nucleic acids into eukaryotic cells. In fact, this is a classical procedure to transfect cDNA or antisense oligonucleotides into cells as well as showing antimicrobial abilities (34–36). In this study, we compare the immune response capacities of mRNA encoding GAPDH encapsulated in lipofectamine (mRNA-GAPDH-LIPO) and GAPDH recombinant proteins with antigens involved in experimental vaccines such as listeriolysin O (LLO) of L. monocytogenes (LM), Ag85A antigen of M. marinum (MM), or pneumolysin (PLY) of S. pneumoniae (SP) (37–42) and explore their potential as CRV vaccines to confer antigen cross-protection immunity.
Materials and Methods
Bacteria, Adjuvants, Cells, Reagents, and Cell Medium
We used L. monocytogenes wild type 10403S strain (LMWT) and LLO L. monocytogenes deficient mutant (LMΔLLO) derived from the 10403S strain (Prof. D.A. Portnoy, University of California, Berkley, CA, USA). The Mycobacterium smegmatis strain was donated by F.J. Sangari and A. Seoane (IBBTEC-University of Cantabria, Santander, Spain) and the S. pneumoniae non-pathogenic vaccine strain, 49619-19F, was obtained commercially from ATCC. Listeria monocytogenes (LM), Mycobacterium marinum (MM), M. chelonae (MC), Mycobacterium avium (MA), Mycobacterium tuberculosis (MTB), Streptococcus pneumoniae (SP) (all of them serotype 5), Streptococcus pyogenes (SPY), and Streptococcus agalactiae (SA) were all clinical isolates of the Microbiology Department at our institution (Hospital Universitario Marqués de Valdecilla, Santander, Spain). DIO-1 is a TLR2/4 targeted molecule that we used as an adjuvant (19–21). Bone-marrow-derived macrophages (DM) or bone-marrow-derived dendritic cells (DCs) were obtained from femurs of 8–12-week-old female mice. DMs or DCs were cultured at 2 × 106 cells/mL in six-well-plates in Dulbecco's Modified Eagle's Medium (DMEM) supplemented with 20% fetal calf serum (FCS), 1 mM glutamine, 1 mM non-essential amino acids, 50 μg/mL gentamicin, and 30 μg/mL vancomycin (DMEM complete medium) and 20 ng/mL granulocyte–macrophage colony-stimulating factor (GM-CSF) for DC, was added to the complete medium to obtain differentiated immune cells. On Day 7, the cells were harvested and analyzed by fluorescence-activated cell sorting (FACS) to evaluate cell surface markers and appropriate differentiation of DCs using the following markers: CD11b–fluorescein isothiocyanate (FITC), CD11c–phycoerythrin (PE), IAb–allophycocyanin (APC), F4/80–PE, CD80–FITC, and CD86–V450 (BD Biosciences, Palo Alto, CA). Cells were collected using cell scrapers to detach adherent cells. In certain samples we also used, after detachment, anti-mouse CD11c-coated magnetic beads and MACSTM separation columns (Miltenyi Biotech Inc., Auburn, CA) on day 7 for positive selection, as previously described (34). Lipofectamine was obtained from Invitrogen.
Mice
We used C57BL/6 mice from our animal facilities at the University of Cantabria at 20–24 weeks old, an age that mimics human beings that are 50 years old and older. LD50 of the L. monocytogenes strain 10403S in C57BL/6 mice is 2 × 105 CFU/mice (2, 39, 43). LD50 of LM (HUMV-01) was 2-fold higher 4 × 105 CFU/mice. LD50 of M. marinum (HUMV-MM01) is 2 × 104 CFU/mice in C57BL/6 mice and LD50 of S. pneumoniae (HUMV-SP01) is 5 × 104/mice in C57BL/6 mice. LD50 were evaluated in groups of mice (n = 10) i.v infected with 2 × 104 CFU/mice, 5 × 104 CFU/mice or 105 CFU/mice. Mice were examined for death every 12 h and checked for clinical parameters of illness every 24 h.
Bioinformatics Analyses
GAPDH of L. monocytogenes (GAPDH-LM) similarity searches were done online using FASTA (available at http://www.ebi.ac.uk/fasta33/) and BLAST (available at http://www.ebi.ac.uk/blast2/ and (http://www.ncbi.nlm.nih.gov/sutils/genom_table.cgi). The analysis of protein domains was based on the Pfam database (available at: https://www.ebi.ac.uk/interpro/) (44). Theoretical 3D predictive models for L. monocytogenes GAPDH (GAPDH-LM), M. tuberculosis GAPDH (GAPDH-MTB), and S. pyogenes GAPDH (GAPDH-SP) were obtained using the Automated Comparative Protein Modeling Server SWISSMODEL (available at https://swissmodel.expasy.org/). Multiple alignment and phylogenetic trees of GAPDH from L. monocytogenes, M. tuberculosis, M. marinum, M. chelonae, S. agalactiae, S. pneumoniae, and S. pyogenes were carried out using Clustal Omega, a multiple sequence alignment program that uses seeded guide trees and HMM profile-profile techniques (available at https://www.ebi.ac.uk/Tools/msa/clustalo/). The aligned regions correspond to the InterPro domain IPR020828 that all the proteins have at the beginning of their sequence. The InterPro domain IRP020828 corresponds to the glyceraldehyde 3-phosphate dehydrogenase, NAD(P) binding domain: https//www.ebi.ac.uk/interpro/entry/InterPro/IPR020828/. The consensus symbols of the alignments were taken from https://www.ebi.ac.uk/seqdb/confluence/display/JDSAT/Clustal+Omega+FAQ#ClustalOmegaFAQ-Whatdotheconsensussymbolsmeaninthealignment? Their meaning is the following: an * (asterisk) indicates positions which have a single, fully conserved residue; a: (colon) indicates conservation between groups of strongly similar properties as below—roughly equivalent to scoring >0.5 in the Gonnet PAM 250 matrix: (STA, NEQK, NHQK, NDEQ, QHRK, MILV, MILF, HY, FYW); a. (period) indicates conservation between groups of weakly similar properties as below—roughly equivalent to scoring = <0.5 and >0 in the Gonnet PAM 250 matrix (CSA, ATV, SAG, STNK, STPA, SGND, SNDEQK, NDEQHK, NEQHRK, FVLIM, HFY). Note that TV is included in the weaker scoring groups despite scoring 0–0 in the PAM 250 matrix, this is because it is a fairly common substitution as they are both beta-branched in fully buried residues, at the cost of a hydrogen bond. In fact, this substitution has been used in the past to make TS mutants (Information courtesy of Toby Gibson).
cDNA Plasmids, in vitro Transcription and Recombinant Proteins
cDNA plasmid clones of antigens from L. monocytogenes serovar 1/2 (listeriolysin O, LLO, and glyceraldehyde-3-phosphate-dehydrogenase, GAPDH), Ag85A antigen of M. marinum, or pneumolysin from S. pneumoniae were obtained from Bioclone Inc. Plasmids were first linearized to prepare mRNA by in vitro transcription (Qiagen in vitro transcription kit) and mRNA transcripts purified with spin columns that contain a silica-based membrane. Purity and concentrations were measured by Nanodrop and further quantification of purity and the size of transcripts was verified by electrophoresis. Escherichia coli strain BL21 bearing plasmids to express large quantities of His-fusion recombinant full-length proteins of LLO (LM-LLOrec or LLOrec) and GAPDH of L. monocytogenes (LM-GAPDHrec or GAPDHrec), pneumolysin O (PLYrec) of S. pneumoniae, and Ag85A of M. marinum (Ag85Arec) were obtained from Bioclone Inc. The expression of large quantities His-fusion proteins was induced with 1 mM IPTG for 5 h at 37°C. His-tagged recombinant proteins were purified with TALON resin, according to the manufacturer's instructions (Clontech). Purification of recombinant proteins was evaluated after SDS-PAGE gels loading 3 μg of protein per lane and Coomasie staining (Figure 2A, labeled as His-protein expression in E. coli) as previously reported by our group (39). Verification of protein purification was evaluated after cutting the bands from gels, TCA precipitation, and proteomic identification at the Centro Nacional of Biotechnology (Madrid). Protein purification was passed through the ToxinEraserTM kit (Genescript, catalog number L0038) to eliminate traces of endotoxin recombinant purified proteins and traces of endotoxin verified with the Genescript ToxiSensorTM chromogenic Limulus Amebocyte lysate kit (catalog number L0035C). The endotoxin elimination kit consists of columns composed by an affinity matrix of modified polymyxin B. Endotoxin levels in protein purifications were lower than 0.1 EU/mL, according to the manufacturer. All reagents to be incubated with DC were tested for endotoxin traces and confirmed to have <0.1 EU/mL of endotoxin.
Preparation of mRNA Encoding Antigens Conjugated to Lipid Carriers (mRNA-Antigen-LIPO)
We prepared the lipid carriers using lipofectamine 2000 (Invitrogen) (5 μl) which was added to mRNA encoding antigens (GAPDH, LLO, PLY and Ag85A) prepared in the previous section before (100 pmol), in a total volume of 100 μL of Opti-MEM. mRNA encoding antigens and lipofectamine mixtures (mRNA-antigens-LIPO) were incubated for 1 h at RT to allow conjugation to mRNA, followed by 5 min of incubation in a water-bath sonicator to allow for the forming of liposome-like carriers. DC prepared in 6-well-plates (1 × 106/well) were incubated with mRNA encoding antigens-LIPO mixtures in Opti-MEM medium without serum for 4 h. Supernatants were removed and cells were incubated for 12 h in DMEM-1% FCS. Efficiencies of mRNA uptake by DC are shown in Figure 2A (DC lysates Coomasie gel of immunoprecipates). Briefly, DC were loaded with 50 μg/mL of mRNA encoded PLY, Ag85A, LLO, or GAPDH conjugated to the lipid carrier, lipofectamine for 16 h. Next, DC were lysed and immunoprecipitated with rabbit anti-Mycobacterium antibody (Colorado University), rabbit anti-PLY (a gift of JR de los Toyos, Oviedo, Spain), and rabbit anti-Listeria monocytogenes GAPDH1-22 antibody (performed by C. Alvarez-Dominguez and M. Fresno at CBMSO facilities using GAPDH1-22 peptide and incomplete Freund's adjuvant) as previously reported (24). Immunoprecipates were stained with Coomasie blue.
Preparation of Murine DC Vaccines and Assays for DC Activation
Bone-marrow derived DC cells obtained from mice femurs were differentiated with GM-CSF (20 ng/mL) for 7 days. Differentiated DC presented a phenotype of 98% CD11c+MHC−II+CD11b−/+CD40−CD86− cells. These DC were used in vivo for T cell responses or vaccination protocols. For DC activation assays, differentiated DC were treated with different reagents for 16 h: 5 μg/mL of recombinant proteins LM-GAPDHrec or LM-LLOrec or 50 μg/mL of mRNA-LIPO complexes: mRNA-LLO-LIPO and mRNA-GAPDH-LIPO. Two adjuvants were also included as reference controls: LPS (10 ng/mL) and the Th1 adjuvant DIO-1 (10 ng/mL). Cell surface markers of DC activation were explored by flow cytometry. Activated DC presented a phenotype of 90% CD11c+IAb+CD40+CD86+ positive cells. Activation was also measured in DC supernatants after filtration and storage at −80°C to measure cytokine production using a multiparametric CBA kit of BD Biosciences (see Cytokine Measurement section).
Cell Toxicity and Apoptosis Assays on Macrophages and DC Vaccines
Bone-marrow derived macrophages (BM-DM) were obtained, as described above, from mice femurs and differentiated with M-CSF (20 ng/mL) for 7 days. BM-DM and activated DC were treated, or not, with the different recombinant proteins or mRNA encoded antigens conjugated to lipid carriers (50 μg/mL) for 16 h in culture medium, washed, and analyzed for cell toxicity or apoptosis. Cell toxicity was examined with Trypan-blue staining by light microscopy as well as by hemolysis of sheep red blood cells. Apoptosis was examined by flow cytometry using two reported products, annexin-V conjugated to allophycocyanin (APC) fluorochrome and 7-AAD (7-aminoactinomycin D) (BD Biosciences, San Jose, CA, USA). Staining of cells with 7-ADD corresponded to necrotic cell death, whereas staining of cells with annexin-V alone corresponded to apoptotic programmed cell death (mean ± SD). Results are expressed as the % of cell toxicity or as the percentages of apoptotic cells ± SD of triplicate samples, respectively (P < 0.05).
Virulence of Bacterial Clinical Isolates in vitro and in vivo
DC vaccines prepared in mice (1 × 106 cells/mL) were infected at a MOI of 10:1 (bacteria: cells) to evaluate the in vitro replication of invasive clinical isolates of LM (HUMV-LM01), MM (HUMV-MM01), and SP (HUMV-SP01) which were calculated as replication indexes (RI) as previously reported (2, 27, 39). RI are calculated by the CFU at 16 h post-infection, divided into CFU at 1 h post-infection. This parameter is considered an indicator of bacterial growth in DC and is comparable to in vivo virulence in spleens 72 h post-infection, as we have previously reported for listeriosis (27). We included the following bacteria as controls: LM 10403S strain (LMWT) as the LM basal control, LLO deficient strain, LMΔLLO as non-pathogenic LM, Mycobacterium smegmatis as non-pathogenic mycobacteria control and the vaccine strain 49619-19F of S. pneumoniae as the non-pathogenic SP control (Figure 1C). Similarly, to evaluate virulence in vivo, we inoculated intravenously 104 CFU of each clinical isolate to be tested. 104 CFU/mice corresponded to a bacterial dose lower than LD50 (see section Mice for LD50 calculations). Spleen homogenates were plated in agar plates to count CFU and results are expressed as CFU/mL. Bacterial controls were the same as those used for in vitro virulence assays.
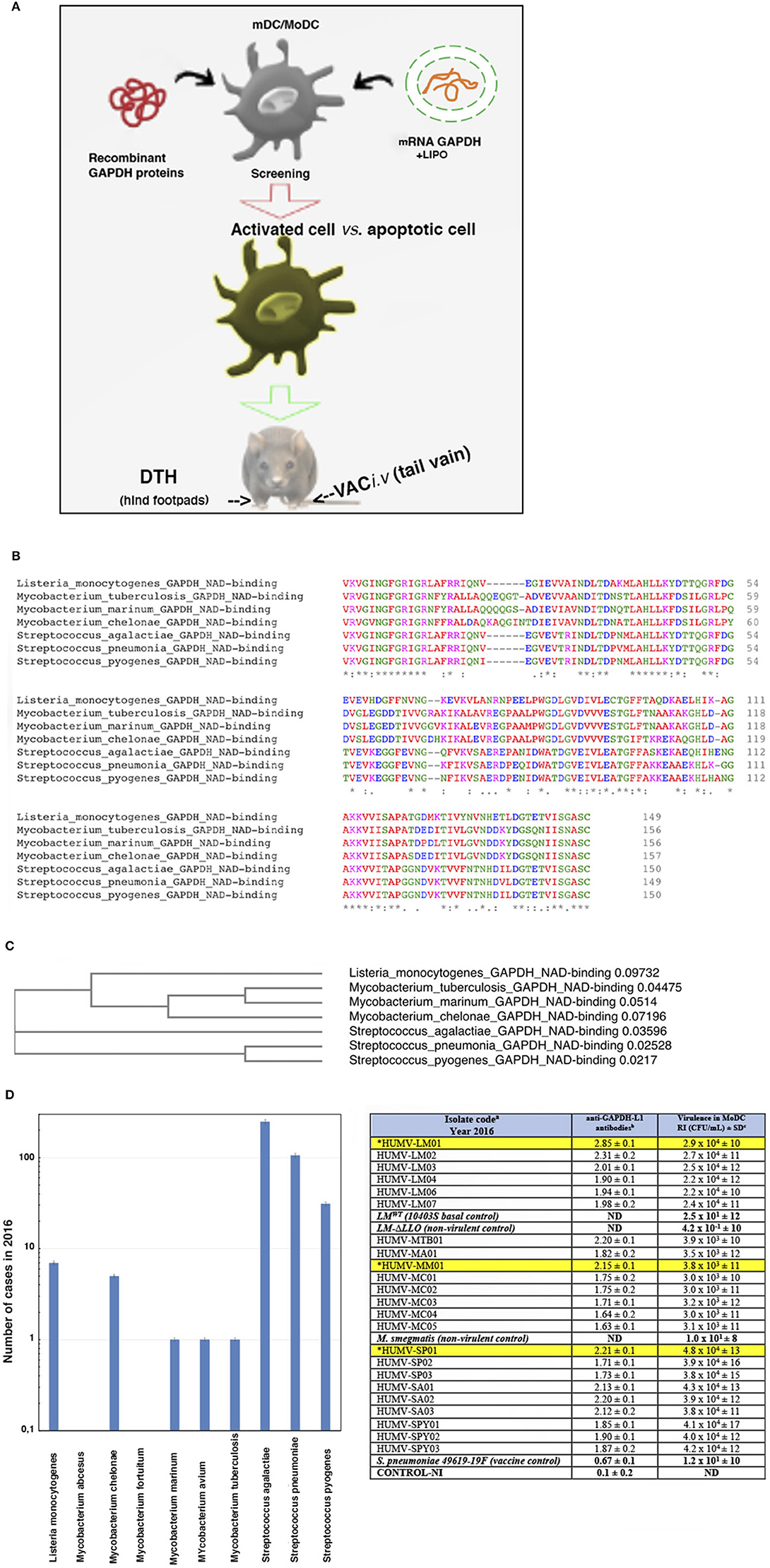
Figure 1. Scheme of the study and selection of the bacterial antigens for the vaccine vectors. (A) Scheme explaining our strategy in this study. First DC are incubated with the different antigen forms: recombinant L. monocytogenes GAPDH proteins or mRNA-LIPO-GAPDH carriers for screening of the suitable ones, causing DC activation with minimal apoptosis induction. DC vaccines loaded with the different antigens are tested for DTH responses as a measurement of T cell responses. DC vaccines with the maximal DTH responses are tested for vaccination experiments followed by bacterial challenge. (B) Multiple alignment of GAPDH sequences of NAD-binding domains of the following bacteria detected at our Health Institution showing more than 95% homology: Listeria monocytogenes (A0A0B8RGN3_LISM), M. tuberculosis (A0A045ITJ4_MYCTX), M. chelonae (A0A0E3TR96_MYCCH) M. marinum (A0A2Z5YDP2_MYCMR), S. agalactiae (Q9ALW2_STRAG), S. pyogenes (G3P_STRPY), and S. pneumoniae (I6L8L9_STREE) protein sequences using CLUSTAL O (1.2.4) multiple sequence alignment. The aligned regions correspond to the InterPro domain IPR020828 that all the proteins have at the beginning of their sequence. The InterPro domain IPR020828 corresponds to the Glyceraldehyde 3-phosphate dehydrogenase, NAD(P) binding domain: https://www.ebi.ac.uk/interpro/entry/InterPro/IPR020828/. The consensus symbols are taken from https://www.ebi.ac.uk/seqdb/confluence/display/JDSAT/Clustal+Omega+FAQ#ClustalOmegaFAQ-Whatdotheconsensussymbolsmeaninthealignment? The symbols meaning is explained in Materials and Methods section Bioinformatics Analyses. Colors on protein alignments correspond to residues according to their physicochemical properties: RED corresponds to Small (small + hydrophobic-including aromatic-Y), BLUE corresponds to acidic, MAGENTA corresponds to basic—H, GREEN corresponds to hydroxyl + sulfhydryl + amine + G and GRAY corresponds to unusual amino/imino acids (see Supplementary Material for complete residues description). (C) Phylogenetic tree of the seven bacteria species compared in this study. The tree data are the following: (Listeria_monocytogenes_GAPDH_NAD-binding:0.09732, ((Mycobacterium_tuberculosis_GAPDH_NAD-binding:0.04475, Mycobacterium_marinum_GAPDH_NAD-binding:0.05140):0.03060, Mycobacterium_chelonae_GAPDH_NAD-binding:0.07196):0.31226):0.14525, Streptococcus_agalactiae_GAPDH_NAD-binding:0.03596, (Streptococcus_pneumonia_GAPDH_NAD-binding:0.02528, Streptococcus_pyogenes_GAPDH_NAD-binding:0.02170):0.01751); (D) Analyses of the clinical cases of bacteria species after the bioinformatic analysis of GAPDH sequences showing 95% homologies in B and detected in the year 2016 at the Hospital U. Marqués de Valdecilla (Microbiology Dpt) from a complete study from 2014 to 2019 (graphic on the left). In the Table (on the right), we show sera from patients (HUMV codes) infected with the bacterial strains of B and examined for anti-LM-GAPDH1−22 antibodies using a peptide ELISA. Sera were collected from patients and storage at −80°C. In the table, we present the antibody titers of patients from a representative 2016 year and with anti-GAPDH-L1 titers higher than 2.0 OD after performing a peptide-specific ELISA. Results are presented as the mean ± SD of OD units in triplicate experiments (P < 0.05). Asterisks and highlighted in yellow correspond to the selected clinical isolates for our further study. Virulence of these clinical isolates into human MoDC (2 × 105/mL) from healthy donors is evaluated in vitro after MoDC infection with 2 × 106 CFU/sample of the clinical isolates detailed in the Table. MoDC were lysed at two different times, at 1 h and at 16 h infection and lysates cultured in agar plates to count CFU. In vitro virulence is expressed as a replication index (RI) of the ration of CFU at 16 h to CFU at 1-h post-infection. Results expressed as RI numbers ± SD of three different experiments. ELISA and virulence assays in vitro using MoDC are performed in triplicate. A Student t-Test is applied for statistical analysis (P ≤ 0.05).
Delayed Type Hypersensitivity (DTH) Reactions Elicited by DC-Vaccines
C57BL/6 mice were immunized i.p with LM (HUMV-LM01), MM (HUMV-MM01), or SP (HUMV-SP01) (5 × 103 CFU). Seven days later, mice were inoculated in the left hind footpads using DC vaccines (106 cells/mice) pre-loaded with the following reagents: the recombinant proteins of L. monocytogenes LLOrec, and GAPDHrec, M. marinum Ag85Arec or S. pneumoniae PLYrec, or the mRNA-Ag-LIPO complexes: mRNA-LLO-LIPO, mRNA-GAPDH-LIPO, mRNA-Ag85A-LIPO, or mRNA-PLY-LIPO. DC vaccines were formulated in the presence of DIO-1 (2 μg/mL) (2). The negative controls were the right hind footpads, since they were not inoculated. After 48 h, we measured the footpad thickness with a caliper; results are expressed in millimeters as the mean of three different experiments. To explore T cell responses in detail, we collected and homogenized the popliteal lymph nodes of mice analyzed for DTH reactions and cell homogenates were passed through cell strainers to analyze CD4+ and CD8+ T cells by flow cytometry. Results are expressed as the percentage of positive cells ± SD.
Vaccination Experiments With DC Vaccines Loaded With Listeria Recombinant Proteins or mRNA-LIPO
C57BL/6 female mice were vaccinated (n = 5/vaccine), or not (n = 5), via the lateral tail vain (i.v), with one dose of DC-vaccines (106 cells/mice) pre-loaded with recombinant Listeria proteins as GAPDHrec and LLOrec; mRNA-LIPO complexes such as mRNA-LIPO-LLO and mRNA- LIPO-GAPD-; or empty DC-LIPO. Seven days post-vaccination, mice were challenged i.v with 100 μL bacterial suspension of LM (HUMV-LM01), MM (HUMV-MM01), or SP (HUMV-SP01) in saline (1 × 104 CFU/mL). All animals were examined daily and 14 days after the bacterial challenge, the mice were bled and sacrificed to quantify viable CFU/mL in the spleens and the cytokines in mice serum. Results are expressed as the percentages of protection ± SD of CFU/mL of vaccinated vs. non-vaccinated animals, using two controls: empty DC and saline. CFU of non-vaccinated mice are as follows: saline LM (HUMV-LM01) 2.75 × 105 CFU/mL, DC-CONT LM (HUMV-LM01) 2.60 × 105 CFU/mL, saline MM (HUMV-MM01) 1 × 105 CFU/mL, DC-CONT MM (HUMV-MM01) 0.9 × 105 CFU/mL, saline SP (HUMV-SP01) 2.5 × 105 CFU/mL, and DC-CONT SP (HUMV-SP01) 2.49 × 105 CFU/mL.
Intracellular IFN-γ Staining
Spleen cells of vaccinated and non-vaccinated mice were cultured in 96-well plates (5 × 106 cells/mL) and stimulated with L. monocytogenes GAPDH1−22 peptide (50 μM) for 5 h in the presence of brefeldin A. Cells were surface labeled for CD4 or CD8, fixed, and permeabilized with a cytofix/cytoperm kit to measure IFN-γ (BD Biosciences). After sample acquisition by flow cytometry, data were gated for CD4+ or CD8+ events, and the percentages of these cells expressing IFN-γ were determined. Results were corrected according to the percentages of total CD4+ or CD8+ positive cells. Data were analyzed using FlowJo software (Treestar, Ashland, OR, USA).
Peptide-ELISA Assay to Measure Listeria monocytogenes GAPDH1−22 Antibody Titers
Ninety-six –well-plates were coated with L. monocytogenes GAPDH1−22 peptide (50 μg/mL) and coated to 96-well-plates in carbonate buffer (pH 8.0) overnight at 4°C. Plates were washed and incubated with 1 mg/mL of BSA (fraction V) to saturate all sites in the plates. Sera of patients infected with LM, MM, or SP or sera of vaccinated or non-vaccinated mice were 1/10 diluted and peptide coated plates were incubated with diluted sera for 2 h at RT, as previously described (2, 24). Reactions were developed with goat anti-human IgG or goat anti-mouse IgG and absorbances were analyzed at 450 nm. Results are presented as optical density measurements (OD) from mean values ± SD, of triplicate experiments.
Isolation of MoDC From Healthy Donors and in vitro Virulence With Clinical Isolates
Leukocytes from whole blood cells were isolated as the interphase of a Ficoll gradient. Leukocytes were incubated with microbeads conjugated to a mouse IgG2a monoclonal anti-CD14 antibody, and passed through MACS™ columns (Miltenyi, Bergisch Gladbach, Germany) to select monocytes (Mo) as CD14+ positive cells. Mo cells were differentiated to monocyte derived DC (MoDC) using standard procedures previously reported (27). In brief, Mo (1 × 106 of cells/mL) are cultured into 6-well-plates (Falcon™) over 7 days using GM-CSF (50 ng/mL) and IL-4 (20 ng/mL) in RPMI-20% FCS medium. Differentiated cells were 98% CD45+HLA-DR±CD86−CD14− positive cells and were used for the in vitro virulence analysis.
Adjuvant Effects of Vaccine Vectors on to MoDC From Healthy Donors
MoDC (2 × 106 cells/mL) were incubated with different recombinant proteins (5 μg/mL) or adjuvants (20 ng/mL), LLOrec, GAPDHrec, LPS, or DIO-1. After 16 h, supernatants were collected, filtered, and stored at −80°C until use for the cytokine analysis. Cell surface markers were analyzed by flow cytometry to evaluate the percentages of CD45, MHC-II, CD86, and CD14 positive cells to determine an activation phenotype of 99% CD45+HLA-DR++CD40++CD86++ positive cells.
Cytokine Measurement
Cytokines in mice sera, DC, or MoDC supernatants were quantified using multiparametric CBA kits, either for mice or for human samples (BD Biosciences, San Jose, CA, USA). The human Th1/Th2/Th17 CBA kit (catalog number 560484) was used to measure human cytokines in MoDC supernatants, and the mouse Th1/Th2/Th17 CBA kit (catalog number 560485) was used to measure cytokines in mice sera and DC supernatants. Cytokine concentrations were expressed as the average of three replicates in pg/mL ± SD. ANOVA was applied to these samples according to the manufacturer's instructions. Data were analyzed using the FlowJo software.
FACS Analysis
Cell surface markers of human MoDC, murine DC, or murine spleens were analyzed by FACS using the following antibodies: anti-HLA-DR-FITC, anti-CD45-PerCP, anti-CD14-PE, and anti-CD86-V450 (clone 2331) for human MoDC. For cell surface markers of murine DC, we used, biotin anti-IAb (clone AF6-120-1), anti-CD11c-PE (clone HL3), anti-CD40-APC (monoclonal 3/23 from BD Pharmingen), and anti-CD86-V450 (clone GL-1) and for murine spleens we also used anti-CD4-FITC (clone RPA-T4) and anti-CD8-PE (clone RPA-T8) (BD Biosciences). Data were analyzed using the FlowJo software. ANOVA was applied to these samples according to the manufacturer's instructions.
Statistical Analysis
For statistical analysis, the Student's t-test was applied to mice assays infected with bacterial pathogens. For statistical purposes, each group included five mice for all assays reported (P < 0.5 was considered significant). ANOVA analysis was applied to the cytokine measurements and flow cytometry analysis as per the manufacturer's recommendations (P ≤ 0.05 was considered significant). For statistical purposes, each flow cytometry sample was performed in triplicate. GraphPad software was used for generation of all the graphs presented.
Ethics Statement
This study was carried out in accordance with the Guide for the Care and Use of Laboratory Animals of the Spanish Ministry of Science, Research and Innovation. The Committee on the Ethics of Animal Experiments of the University of Cantabria approved the protocol (Permit Number: PI-01-17) that follows the Spanish legislation (RD 1201/2005). All surgeries were performed by cervical dislocation, and all efforts were made to minimize suffering. Similarly, for the use of human data of bacteria clinical isolates, we have an approved project from the Committee of Clinical Ethics of Cantabria (CEm) entitled: “Clinical Development of Listeria based vaccines” which includes Informed Consent and General Project Information documents of patients (Permit Acta Number: 29/2014, internal code: 2014.228).
Results and Discussion
We initiated this study with the hypothesis that bacterial vaccines for adults can benefit from the discovery of antigens that are able to immunodulate DC and drive a wide spectrum immunity that cross-protects against bacterial infectious diseases of the genera Listeria, Mycobacterium, and Streptococcus. Vaccines inducing cross-protection immunity has recently been suggested for these taxonomic groups as multivalent vaccines (28). They are differentiated from conventional vaccines as they have the capacity of cross-reactive immune responses. For this reason, here, we refer to them as CRV vaccines to differentiate them from other type of vaccines, such as trained-immunity based vaccines (TIbV). CRV and TIbV might share two features: (i) stimulation of non-specific protection against several pathogens that involves innate immune cells and (ii) induction of specific immune responses to the vaccine antigens (5, 6). DC are innate immune cells responsible for antigen presentation and is relevant in all types of vaccines, conventional, CRV, or TIbV. DC are explored here as vaccine platforms to evaluate any bacterial antigen as a candidate for cross-protection vaccination, if the antigens induced minimal DC apoptosis, along with maximal expansion of T cells. In this context, two types of vaccine carriers are explored: an mRNA-encoded antigen conjugated with lipid carriers and recombinant proteins (see scheme of our procedure in Figure 1A). The bacterial antigens we use in this study are those reported in experimental vaccines for the above-mentioned bacteria genera: L. monocytogenes GAPDH and LLO, M. marinum Ag85A and S. pneumoniae PLY (37–42). DC and macrophages were selected as vaccine vectors since they are innate immune cells that participate actively in cross-protection immunity (5, 6). We focused our study to L. monocytogenes GAPDH antigen (Lmo2459) since it presents similar ADP-ribosylating abilities, immunogenic domains, and cross-immune responses in three bacterial genera of our study, Listeria, Mycobacterium, and Streptococcus (24–26, 28). These features prompted us to hypothesize that L. monocytogenes GAPDH was a candidate for CRV vaccines.
Selection of Antigens and Antigen Forms
We performed two approaches to select the bacterial pathogens for our study: first a bioinformatic analysis we previously reported (28) to search for homologies higher than 80% among GAPDH of most common pathogenic bacteria communicated annually at our Health institution and virulence analysis of clinical isolates. From a 5-year study from 2014 to 2018, we chose year 2016 as representative and detected several bacterial genera with GAPDH homologies higher than 80%, such as Hemophilus, Klebsiella, Listeria, Mycobacterium, Pseudomonas, Staphylococcus, and Streptococcus (Table 1). However, if we quoted GAPDH homologies to 95% or higher, only clinical isolates of the bacterial taxonomic groups of L. monocytogenes, Mycobacterium, or Streptococcus fitted this category. The highest GAPDH homologies corresponded to L. monocytogenes (LM), M. chelonae (MC), M. tuberculosis (MTB), M. marinum (MM), S. agalactiae (SA), S. pneumoniae (SP), and S. pyogenes (SPY). Next, we focused on the NAD-binding domains of GAPDH from these taxonomic groups, using CLUSTAL 0 (1.24) multiple sequence alignment (Figure 1B), and, observed that protein sequences covering amino acids 3–25 displayed the highest identities (asterisks corresponds to 100% identity, colon symbol to 90%, and period symbol to 80% (detailed analysis is described in Material and Methods, section Bioinformatics Analyses). Amino acids are shown in a colored codes to distinguish homologies (Figure 1B) and color code explanations are provided in the Figure legend and Supplementary Material (Supplementary Table 1). These alignments might explain that the peptide-specific anti-Listeria monocytogenes GAPDH1−22 antibody prepared in rabbits, with the 1-22 amino acid sequence of LM, can also detect MTB, MM, and SP bacterial extracts and surface shapes of the bacteria as previously described by our group (24, 28), suggesting that LM, MTB, MM, and SP shared immunogenic domains, in addition to enzymatic abilities and enzymatic domains. This is especially relevant as the phylogenetic tree relates NAD-binding domains of LM with MTB, MM, and MC. Another branch of the phylogenetic tree relates the NAD-binding domains of LM and SA and a third branch relates NAD-binding domains of LM with SP and SPY (Figure 1C), suggesting that GAPDH could be a common virulence factor. To further investigate this issue, we collected sera from all patients reported with infections caused by these eight bacterial species, detected at year 2016 (graph plot in Figure 1D), and explored for the presence of antibodies recognizing the LM-GAPDH1−22 peptide, using a peptide-ELISA previously described (27). Several patients with infections caused by LM (HUMV-LM01, HUMV-LM02, and HUMV-LM03), MM (HUMV-MM01), MTB (HUMV-MTB01), SP(HUMV-SP01), SA (HUMV-SA01, HUMV-SA02, HUMV-SA03), and SPY (HUMV-SPY02) presented very high levels of antibodies recognizing the LM-GAPDH1−22 epitope with O.D. ≥ 2.0, (right table in Figure 1D, column labeled as anti-GAPDH-L1 antibodies), while the remaining patients presented high levels of antibodies with O.D. ≥ 1.5. We concluded that immune responses generated by Listeria, Mycobacterium, and Streptococcus taxonomic groups are mainly targeted to a common GAPDH1−22 epitope, strongly suggesting that GAPDH might be a common virulence factor to these pathogens. Evaluation of the in vitro virulence of their clinical isolates also supports our hypothesis. In vitro virulence was performed, infecting monocyte derived dendritic cells (MoDC) from healthy donors with the clinical isolates at a MOI of 10:1 and examining the bacteria replication indexes (RI). RI are defined as the ratio of CFU/mL at 16 h post-infection to CFU/mL at 1 h (27). We detected that those patients with the highest titers of antibodies recognizing the LM-GAPDH1−22 epitope, also presented the highest virulent strains of LM, MTB, MM, SP, or SPY, showing at least 100-fold higher replication indexes (RI) than non-virulent strains (right table in Figure 1D, column labeled as virulence in MoDC). This methodology was confirmed by the virulence of the clinical isolates in vivo using C57BL/6 mice and bacterial doses lower than LD50 [see Material and Methods section Mice and (41–44)], that reported similar results as in vitro virulence using MoDC (Supplementary Table 2). In brief, these data strongly suggests that GAPDH is a common virulence antigen of Listeria, Mycobacterium, and Streptococcus that needs to be explored as a candidate for CRV vaccines.

Table 1. Main bacteria detected at Hospital Universitario Marqués de Valdecilla yearly with GAPDH showing more than 80% sequence homology at N-terminus.
The second approach was to decipher the best antigen form to prepare a T-cell based vaccine vector from using DC loaded with the antigens and inoculation of mice hind footpads to examine a classical delayed-type hypersensitivity assay (DTH), a valid measure of T cell immunity (2). The antigens included in this strategy are commercially available as cDNAs (Bioclone Inc): Ag85A of MM (Ag85-MM), pneumolysin (PLY) of SP (PLY-SP), and GAPDH (GAPDH-LM) and listeriolysin O (LLO) of LM (LLO-LM). We prepared and compared two types of antigen forms, recombinant proteins and mRNA-lipid carrier complexes (mRNA-LIPO) because they can load different antigen processing compartments on DC. While recombinant proteins load the endo-lysosomal compartments relevant for MHC-class II antigen presentation, mRNA-lipid carrier complexes (LIPO) load the cross-presentation compartments relevant for MHC-class I antigen presentation (30, 45, 46). To prepare mRNA-lipid carrier complexes, commercially available DNA plasmids were first linearized (left upper cDNA gel in Figure 2A showed cDNA plasmids of each antigen) and mRNA samples were obtained by in vitro transcription (right upper mRNA gel in Figure 2A). Next, we added a CAP site at the 5′ end and a poly A tail at the 3′ end, following the manufacture's recommendation (see details in Materials and Methods, section cDNA Plasmids, in vitro Transcription, and Recombinant Proteins) (concentration and purity of transcripts are shown in Supplementary Figure 1A). Next, mRNA samples (100 pmol) were incubated with the lipid carrier, lipofectamine (5 μL), to obtain mRNA-Antigen-lipid carrier complexes (labeled here as mRNA-antigen-LIPO) and offered to DC to evaluate maximal uptake by antigen presenting cells (right lower Coomasie stained gel in Figure 2A). To prepare recombinant proteins, commercially available DNA plasmids were expressed in large quantities as His-fusion proteins in E. coli strain BL21 to obtain LLOrec, Ag85Arec, PLYrec, or GAPDHrec (left lower Coomasie stained gel in Figure 2A). Toxicities of mRNA-antigen-LIPO complexes and recombinant proteins were examined by hemolysis of sheep red blood cells in macrophages (BM-DM) and DC (Supplementary Figure 1B), as well as by Trypan blue staining in DC which reflects cell viabilities (Supplementary Figure 1C). Both methods of analyzing toxicities—hemolysis and Trypan blue—are relevant when using cytolysins (LLO or PLY) that are able to lyse red blood cells as LLO or PLY, while not causing significant reductions on cell viabilities. In fact, the high hemolysis detected with both cytolysins in macrophages, either as recombinant proteins or mRNA-antigen-LIPO complexes (Supplementary Figure 1B), drove us not to use macrophages for vaccine platforms. None of the antigen forms we used with DC caused hemolysis (Supplementary Figure 1B) or reduction of cell viability (Supplementary Figure 1C), therefore, we concluded that DC were the most suitable vaccine platform.
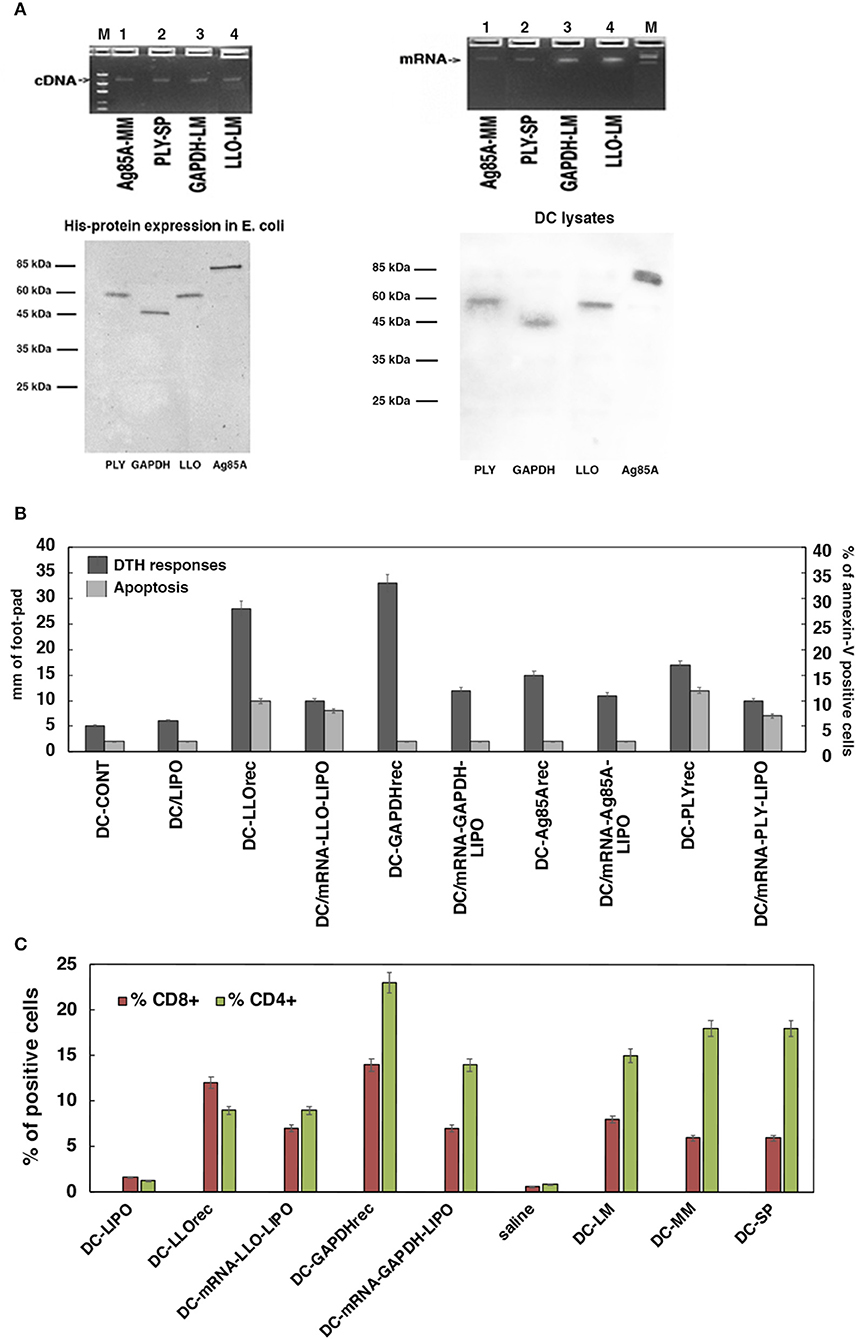
Figure 2. Selection of the antigen forms for the vaccines. (A) Upper geles correspond to preparation of the mRNA antigens from cDNA plasmids encoding for Ag85A antigen of M. marinum (MM), PLY of S. pneumoniae (SP), and GAPDH or LLO of L. monocytogenes (LM) after in vitro transcription. Gels show the linear plasmids (upper left) and mRNA transcripts (upper right). Concentration of the mRNA preparations and qualities are shown in Supplementary Figure 1A. Lower gels correspond to Coomasie stained gels of purification of His-recombinant proteins (lower left) and DC uptake of prepared mRNA from all antigens (PLY of SP, Ag85A of MM, LLO, or GAPDH of LM) conjugated to the lipid carrier, lipofectamine, and after 16 h DC cells are lysed. Lysates were immunoprecipitated with rabbit anti-LLO antibody (DIATEVA), rabbit anti-Mycobacterium antibody (Colorado University), rabbit anti-PLY (a gift from JR de los Toyos, Oviedo, Spain), and rabbit anti-LM-GAPDH1-22 antibody (C. A-D and M.F obtained at CBMSO) (24). All immunoprecitpates were stained with Coomasie blue. (B) DC apoptosis (light gray bars) and DTH responses measured as the footpad swelling (dark gray bars) evaluated after incubation of DC with different antigens: empty DC (DC-CONT), lipofectamine (DC-LIPO), recombinant proteins as PLYrec from SP, LLOrec, and GAPDHrec from LM and antigen Ag85A from MM or mRNA-LIPO complexes of PLY, LLO, GAPDH, or Ag85A. Apoptosis is measured in vitro by flow cytometry and results are expressed as the percentages of annexin-V positive cells ± SD of three different experiments. ANOVA test was applied for flow cytometry results (P ≤ 0.05). DTH responses are measured in vivo after inoculation of right hind footpads of C57BL/6 mice with the different DC vaccines (n = 5 per DC vaccine). Forty-eight hours after inoculation of DC vaccines, DTH responses are evaluated by the swelling of the hind footpads measured with a caliper. Results are expressed as millimeters ± SD of each group of 5 mice. Student t-Test was applied for statistical analysis (P ≤ 0.05). (C) C57BL/6 mice were immunized i.v with 5 × 103 CFU/mice (HUMV-LM01, HUMV-MM01, or HUMV-SP01) and 7 days later, left hind footpads were inoculated with 1 × 106 DC vaccines (pre-loaded with 5 μg/mL of LLOrec, GAPDHrec, or 50 g/mL of mRNA-LIPO-GAPDH or mRNA-LIPO-LLO, or 1 × 106 CFU of LM, MM or SP, LIPO incubated DC, or saline incubated DC). Popliteal lymph nodes are isolated from mice legs and after homogenization, T cells sub-populations are analyzed by flow cytometry. Percentages of CD4+ (green bars) or CD8+ T cells (red bars) are shown. Results are expressed as the percentages of positive cells ± SD of three different experiments. Student t-Test was applied for statistical analysis (P < 0.05).
Selection of Most Immunogenic Antigens
Once antigen forms were prepared, we examined the DTH responses in C57BL/6 mice previously challenged intravenously (i.v) with the pathogens LM (HUMV-LM01), MM (HUMV-MM01), or SP (HUMV-SP01). Seven days post-infection we inoculated the left hind footpads with 106 DC pre-loaded with the different bacterial antigens, either recombinant proteins (5 μg/mL) or mRNA-LIPO complexes (50 μg/mL), in solutions with DIO-1 adjuvant to amplify the immune response, per mouse. The DTH response was measured as the swelling on the left hind footpad of each mouse 48 h post-inoculation, compared to the right hind footpad, which acts as the negative control. DC loaded with recombinant LM-GAPDHrec presented the highest DTH responses, followed by DC loaded with recombinant LM-LLOrec, next were mRNA-LM-GAPDH-LIPO and mRNA-SP-PLY-LIPO complexes. DC loaded with MM-Ag85Arec, mRNA- MM-Ag85A-LIPO complexes (dark gray bars in Figure 2B) induced significant DTH responses but lower than LM-GAPDH or LM-LLO antigen forms. DC loaded with SP-PLYrec and mRNA-SP-PLY-LIPO show half the footpad swelling than GAPDH antigen forms, therefore they induce only partial DTH responses. We also explored the abilities of these antigens to induce apoptosis in DC as a measure of the undesired inactivation of DC (≥10% apoptosis) (see Material and Methods in section Cell Toxicity and Apoptosis Assays on Macrophages and DC Vaccines). Whole pathogens, LM, MM, or SP (HUMV-LM01, HUMV-MM01, or HUMV-SP01, respectively) induced high levels of apoptosis (12–17%) as well as recombinant cytolysins like SP-PLYrec and LM-LLOrec (11–18%) or mRNA-LIPO complexes of these cytolysins (10–13%) (light gray bars in Figure 2B). All the other molecular forms tested (mRNA-LIPO complexes of LM-GAPDH or MM-Ag85A, and their recombinant proteins) presented apoptosis below 5% and similar to controls: DC loaded with lipofectamine (DC-LIPO) or incubated with saline (DC-CONT) (Figure 2B). Therefore, we concluded that the highest immunogenic and less apoptotic antigen forms corresponded to recombinant LM-GAPDHrec. mRNA-LIPO complexes of LM-GAPDH show half the lower immunogenic DTH responses than LM-GAPDHrec, although we inoculated a 10-fold concentration of mRNA-LIPO complexes compared to recombinant proteins. In brief, we do not consider this antigen form, mRNA-LIPO complexes, as suitable for exploring CRV vaccines. Next, we collected the popliteal lymph nodes of mice with the highest DTH immune responses (LM-GAPDHrec, LM-LLOrec, mRNA-LIPO complexes of LM-GAPDH or LM-LLO) and cultured them in vitro with 1 μg/mL of each antigen for 72 h, and examined the percentages of T cell populations, both CD4+ or CD8+ T cells by flow cytometry. We detected the highest percentages of CD4+ (23%) and CD8+ (14%) T cells in mice inoculated with LM-GAPDHrec (Figure 2C). mRNA-LIPO complexes of GAPDH presented significant percentages of CD4+ (15%) T cells, but low percentages of CD8+ (7%) T cells. The molecular forms of LLO, presented low percentages of CD4+ (9%) T cells but significant percentages of CD8+ (12%) T cells. However, mRNA-LIPO complexes of LLO induced low percentages of CD4+ (9%) and CD8+ (7%) T cells. No significant T cell responses were observed in the controls, DC, or in saline. When we compared these results with the DTH responses, we confirmed a correlation between the highest DTH responses (dark gray bars in Figure 2B) and the highest percentages of CD4+ and CD8+ T cells induced in the popliteal lymph nodes (Figure 2C). We argue that antigens in vaccine platforms that induced high DTH responses reflect the high expansion of T cell responses they induced and explains their high immunogenicity; both features are specific of the antigen.
Adjuvant Abilities of Vaccine Vectors
There is another possible explanation for DC-LM-GAPDHrec vaccines generating high DTH immune responses with induction of CD8+ and CD4+ T cells that is not related to the antigen immunogenicity. Some antigens can also induce DC activation, such as adjuvants or cell-walls of dead bacteria (16–19) and are interesting compounds for vaccine platforms. Here, we tested the possibility that LM-GAPDHrec, LM-LLOrec, or mRNA-LIPO complexes of LM-GAPDH or LM-LLO serve as non-specific DC activators. We evaluated two characteristics of activated DC, the cell surface expression of activation markers and the production of cytokines. We treated DC with different reagents, LM (HUMV-LM01), MM (HUMV-MM01), SP (HUMV-SP01), LM-LLOrec, LM-GAPDHrec, mRNA-LIPO complexes of LM-GAPDH or LM-LLO for 16 h, to examine activation. Two different adjuvants were also included in the assay, LPS and DIO-1 (14). Classical cell surface activation markers of DC are CD11c, MHC-II, CD40, or CD86, while CD11b is a macrophage-DC marker that, upon DC activation, reduces its surface expression and GR1 is a classical polymorphonuclear leukocyte (PMN) marker. LM (HUMV-LM01), MM (HUMV-MM01), and SP (HUMV-SP01) bacteria clearly induce DC activation, reflected by high percentages of CD11c, MHC-II, CD40, and CD86 positive cells (Figure 3A). mRNA-LIPO complexes of LM-GAPDH or LM-LLO did not induce DC activation, as the percentages of positive cells for MHC-II, CD40, or CD86 were similar to non-infected controls (NI). Recombinant LM-GAPDHrec protein was the only antigen form that clearly increased the percentages of all DC activation markers, CD11c, MHC-II, CD40, and CD86. However, LM-GAPDHrec effect was different than the activation pattern induced with LPS that increased only the percentages of the MHC-II activation marker (violet bars in Figure 3A) and was also different to the activation pattern induced by the DIO-1 adjuvant that increased the percentages of two activation markers, MHC-II and CD40. Neither LPS (dark blue bars), nor DIO-1 (garnet bars) caused significant effects in the percentages of CD86 positive cells. We conclude that LM-GAPDHrec activation of DC affected the expression of all classical markers of DC activation (light blue bars), suggesting a broader activation pattern. Next, we explored other features of DC activation, after collection of DC supernatants and analysis of cytokines using a Th1-Th2 parametric flow cytometry assay (BD Biosciences). As shown in Figure 3B, DC stimulation with adjuvants as LPS released high levels of Th1 (MCP-1, TNF-α, or IFN-α and Th2 (IL-6 and IL-10) cytokines; while stimulation with adjuvants like DIO-1 produced Th1 (MCP-1, TNF- α, or IFN-α), but not Th2 cytokines. DC stimulation with mRNA-LIPO complexes of GAPDH or LLO produced no cytokine at all (undetectable levels) and LLOrec only showed low levels of Th1 cytokines (1–5 pg/mL). DC stimulated with recombinant LM-GAPDHrec released high levels of Th1 cytokines such as MCP-1, TNF-α, IFN-α, and IL-12, while no significant levels of Th2 cytokines such as IL-6 or IL-10 were observed. IL-12 production is associated with the ability to stimulate CD8+ T cells and might explain the effect of DC loaded with recombinant LM-GAPDHrec to promote DTH responses (red bars in Figure 2C) after DC activation. We also confirmed that GAPDHrec was able to activate monocyte derived DC (MoDC) from healthy donors, as they induced Th1 cytokines with high levels of IL-12 and very low levels of IL-6 and IL-10 (Supplementary Table 3). We conclude that LM-GAPDHrec is a classical pro-inflammatory adjuvant that is able to activate DC in a stronger and broader manner.
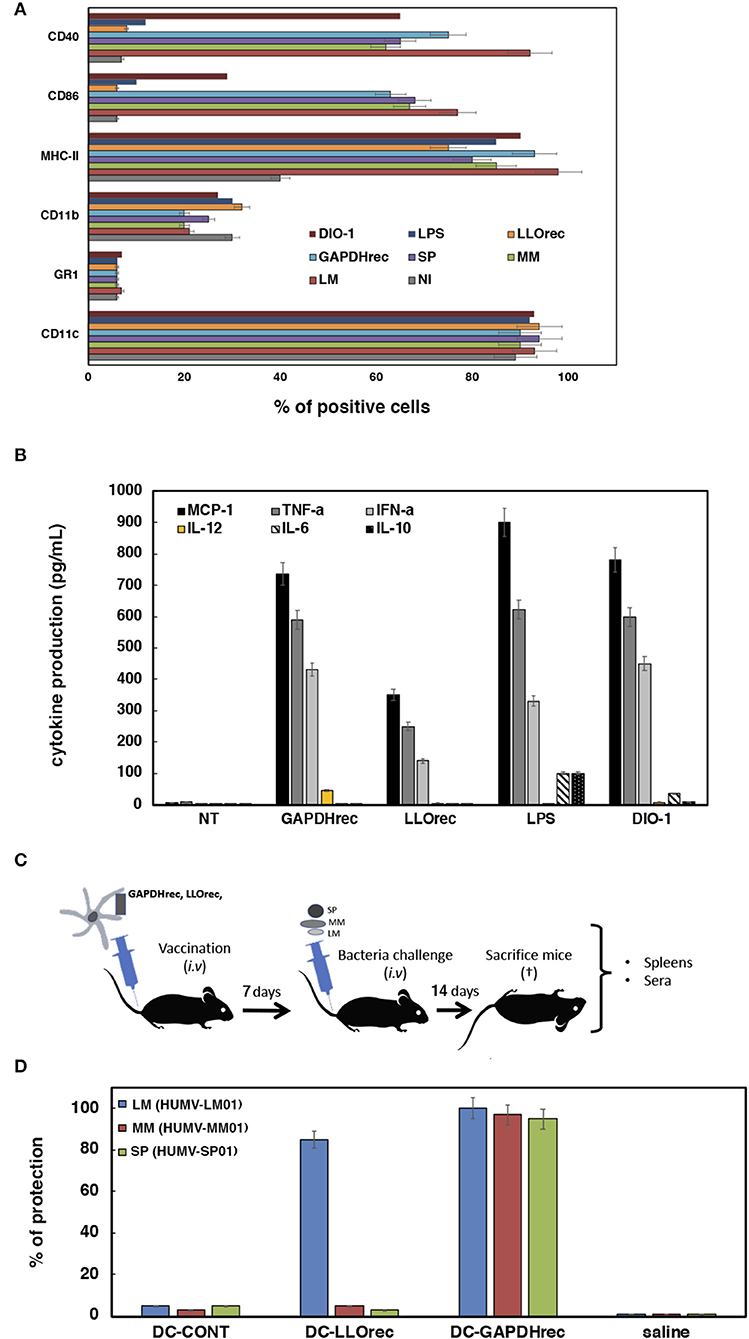
Figure 3. Adjuvant and vaccine abilities of DC vaccine vectors loaded with recombinant or mRNA-LIPO antigens. (A) Flow cytometry analysis of DC surface markers after incubation with recombinant proteins LLOrec or GAPDHrec or mRNA-LIPO complexes: mRNA-LIPO-LLO, mRNA-LIPO-GAPDH, bacteria MM (HUMV-MM01), SP (HUMV-SP01), or LM (HUMV-LM01) or two adjuvants, LPS or DIO-1. Results show the percentages of CD11c+, MHC-II+, CD40+, or CD86+ positive cells. Results are the mean of three different experiments ± SD. Student t-Test was applied for statistical analysis (P ≤ 0.05). (B) Cytokine levels released to the supernatants of DC and measured with a multiparametric CBA kit (BD Biosciences). Results are expressed as pg/mL of each cytokine ± SD of triplicate samples. ANOVA test was applied to the cytokine's concentrations according to the manufacturer recommendation (P ≤ 0.05). (C) Scheme of vaccination model and sample collection to analyze immune responses and protection: spleens and sera. (D) Vaccination of C57BL/6 mice with a single dose of DC vaccines. Seven days later, each group of vaccinated mice are divided in 3 sets and challenged i.v with 104 CFU/mice of hypervirulent strains of HUMV-LM01, HUMV-MM01, or HUMV-SP01. Next, after 14 days mice are bled, sacrificed and spleens collected. Vaccination results expressed percentages of protection as the mean ± SD of triplicates. Percentages are calculated as the number of CFU/mL counted in spleen homogenates of NV mice (saline) divided by CFU/mL of each set of vaccinated mice. Results are expressed as the mean ± SD of triplicates. Student t-Test was applied for statistical analysis (P ≤ 0.05). CFU of non-vaccinated mice are the following: saline LM (HUMV-LM01) 2.75 × 105 CFU/mL, DC-CONT LM (HUMV-LM01) 2.60 × 105 CFU/mL, saline MM (HUMV-MM01) 1 × 105 CFU/mL, DC-CONT MM (HUMV-MM01) 0.9 × 105 CFU/mL, saline SP (HUMV-SP01) 2.5 × 105 CFU/mL, DC-CONT SP (HUMV-SP01) 2.49 × 105 CFU/mL.
Validation of DC-GAPDHrec as CRV Vaccines for Listeria monocytogenes, Mycobacterium marinum, and Streptococcus pneumoniae Infections
Specific DC activation with production of IL-12 have been linked to vaccine efficiency (47), therefore, we tested the vaccine efficiency of DC loaded with the highest immunogenic antigen forms, recombinant proteins LM-GAPDHrec and LM-LLOrec (see Figure 3C for vaccination scheme). Five mice per group were inoculated i.v with a single dose of DC vaccines (106 cells/mice) pre-loaded with 5 μg/mL of LM-LLOrec or LM-GAPDHrec (DC-LM-LLOrec or DC-LM-GAPDHrec) for 7 days and was then challenged i.v with either LM (HUMV-LM01), MM (HUMV-MM01), or SP (HUMV-SP01) for 14 days. Next, mice were sacrificed and their sera and spleens were collected. CFU were examined in spleens by plating in specific agar plates and results were expressed as the percentages of protection (see Material and Methods, section Vaccination Experiments With DC Vaccines Loaded With Listeria Recombinant Proteins or mRNA-LIPO for the detailed procedure). Only DC vaccines pre-loaded with LM-GAPDHrec conferred good protection against a challenge with LM (HUMV-LM01), MM (HUMV-MM01), or SP (HUMV-SP01) (blue, red and green bars in Figure 3D), while DC-LM-LLOrec protected only for LM (HUMV-LM01) infection. Empty DC showed no protection at all against any of the pathogens (bars labeled as DC-CONT in Figure 3D).
We also checked specific humoral and cellular immune parameters in vaccinated and non-vaccinated mice reported in experimental listeriosis vaccines (38, 48), such as the presence of antibodies recognizing the LM-GAPDH1−22 peptide in sera and the percentages of CD4+ or CD8+ cells specific for LM-GAPDH1−22 peptide-specific and IFN-γ producers, as well as verification of high frequencies of CD8+ T cells specific for GAPDH1−22 peptide using H2-Kb:Ig dimers (Table 2, see procedures in Materials and Methods, section Intracellular IFN-γ Staining). We detected high titers of antibodies recognizing LM-GAPDH1−22 epitope, and high percentages of GAPDH1−22 specific CD4+ and CD8+ and IFN-γ producers after vaccination with DC-LM-GAPDHrec and being challenged with LM (HUMV-LM01), MM (HUMV-MM01), or SP (HUMV-SP01) infections. Moreover, these vaccinated mice presented very high frequencies of CD8+ T cells specific for the GAPDH1−22 peptide, while non-vaccinated mice challenged with LM, MM, or SP presented undetectable frequencies. We concluded that DC-LM-GAPDHrec vaccines caused mainly antigen specific DC immune stimulation that confer cross-protection against LM, MM, and SP and induced GAPDH specific immune responses, both in T and B cells. However, we cannot discard non-specific broader DC immune stimulation.
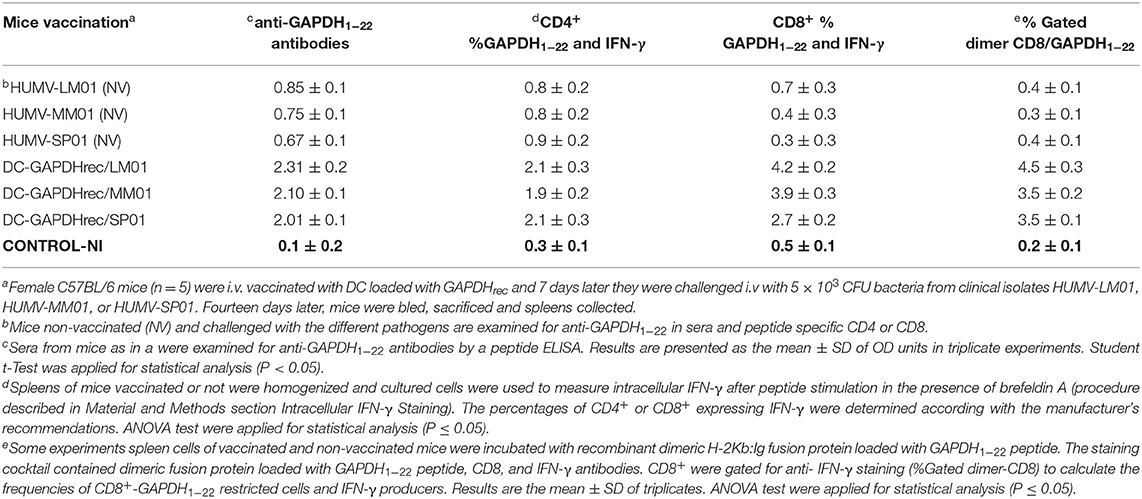
Table 2. Specific immune responses elicited after vaccination of mice with DC-LM-GAPDHrec and challenge with LM (HUMV-LM01), MM (HUMV-MM01), or SP (HUMV-SP01).
Conclusion
Listeria monocytogenes GAPDH in two forms, either as a recombinant protein or as an mRNA-GNP complex, appears to be a safe bacterial antigen that induce significant T cell mediated immune responses when used in DC vaccine vectors. However, only the Listeria GAPDH recombinant protein activates DC in a specific and non-specific but broader form, different than adjuvant activation, as it induces all relevant activation markers and high production of Th1 cytokines, including IL-12. Therefore, not only is stimulation of T cell immune responses required for an antigen form to be considered a good candidate for vaccines, but specific DC activation also seems necessary to induce cross-protection against Listeria, Mycobacterium, and Streptococcus infections. DC vaccines loaded with recombinant LM-GAPDH can be considered not only as CRV vaccines with cross-protection abilities, but also as TIbV vaccines, since they present broad-spectrum protection for the common GAPDH virulence factor of Listeria, Mycobacterium, and Streptococcus and induces specific GAPDH immune responses. In fact, cross-protection abilities of these vaccines correlate with high levels of antibodies and high percentages of specific CD4+, CD8+ T cells, and IFN-γ producers, recognizing the N-terminal GAPDH1−22 peptide that has 98% homology in Listeria, Mycobacterium, and Streptococcus. The ability of mRNA-lipid carrier complexes to induce DC activation and strong T cell responses should be improved to include them in vaccine formulations for multivalent vaccines.
We speculate that experimental multivalent vaccines that can protect against Listeria, Mycobacterium, Streptococcus, bacterial genera responsible for severe meningitis, and long-lasting cutaneous infections in adults and the elderly, are promising tools for the new generation of human vaccines that are based on cross-reactive immunity, either as multivalent or as trained immunity-based vaccines.
Patents
This study is protected by patent number WO200802108A1. https://patents.google.com/patent/WO2008020108A1/fi#patentCitations (accessed March 27, 2021), entitled to Fundación Marqués de Valdecilla and patent number WO2019243647. https://patentscope.wipo.int/search/es/detail.jsf?docId=WO2019243647 (accessed March 27, 2021), entitled to Fundación Instituto de Investigación Marqués de Valdecilla.
Data Availability Statement
The original contributions presented in the study are included in the article/Supplementary Material, further inquiries can be directed to the corresponding author/s.
Ethics Statement
The studies involving human participants were reviewed and approved by Committee of Clinical Ethics of Cantabria. The patients/participants provided their written informed consent to participate in this study. The animal study was reviewed and approved by Universidad de Cantabria Animal Ethical Committee.
Author Contributions
CA-D designed the experiments and directed the coordination of the experiments. DS-C and HT-N performed all the experiments and contributed equally to this study. RC-G helped in the performance of the vaccination experiments. RT performed the bioinformatic analyses. JC-M and IP-D provided all the clinical isolates, collected sera from patients, collected the Informed Consents of the patients, and have the custody of them. SY-D provided clinical isolates and clinical histories of patients infected with M. marinum, collected the Informed Consents of the patients and have their custody and helped with the mice models of infection. All authors contributed to the article and approved the submitted version.
Funding
This research was funded by the CICYT program of the Ministry of Science and Innovation, grant number SAF2012-34203, co-funded in part with the European FEDER funds A new way of making Europe, the COST European action ENOVA, the Instituto de Salud Carlos III (ISCIII) grant numbers, DTS18-00022 and PI19-01580 to CA-D, the Bio-Health Research Program of Cantabria Government, gran number INNVAL19/26 to SY-D, IP-D, and CA-D and INNVAL20/01 to CA-D; SAF2016-75988-R Comunidad de Madrid (S2017/BMD-3671. INFLAMUNE-CM) and the Fondo de Investigaciones Sanitarias (BIOIMID) to MF and a Predoctoral contract to DS-C by Bio-Health research program of Cantabria government.
Conflict of Interest
MF was employed by company DIOMUNE S.L.
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We acknowledge Carmen Punzon (Diomune, S. L, Centro de Biología Molecular Severo Ochoa, Universidad Autónoma de Madrid, Madrid, Spain) that kindly synthesize DIO-1 adjuvant. D.A. Portnoy (University of Berkley, UCLA, USA) for the LMWT and LMΔLLO strains, F. J. Sangari and A. Seoane (IBBTEC-UC, Santander, Spain) for M. smegmatis strain, J. R de los Toyos (U. Oviedo, Asturias, Spain) for the anti-PLY antibody, A. Paradela (CNB, CSIC, Madrid) for proteomic analysis of purified proteins and A. Criado and E. Lanagrán (Department of Health Processes, Education Faculty, at Universidad Internacional de La Rioja, Logroño, La Rioja, Spain) for personal support. We thank I. Garcia (CIC-biomaGUNE, San Sebastian-Donostia, Spain) for helping to design the image of Figure 1A.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fimmu.2021.632304/full#supplementary-material
References
1. Mora M, Telford JL. Genome-based approaches to vaccine development. J Mol Med. (2010) 88:143–7. doi: 10.1007/s00109-009-0574-9
2. Calderon-Gonzalez R, Tobes R, Pareja E, Frande-Cabanes E, Alaez-Alvarez L, Petrovsky N, et al. Identification and characterisation of T-cell epitopes for incorporation into dendritic cell-delivered Listeria vaccines. J Immunol Methods. (2015) 424:111–9. doi: 10.1016/j.jim.2015.05.009
3. Stephenson KE, Neubauer GH, Reimer U, Pawlowski N, Knaute T, Zerweck J, et al. Quantification of epitope diversity of HIV-1-specific binding antibodies by peptidemicroarrays for global HIV-1 vaccine development. J Immunol Methods. (2015) 416:105–23. doi: 10.1016/j.jim.2014.11.006
4. Martínez L, Milanič M, Malaina I, Álvarez C, Pérez MB, M de la Fuente I. Weighted lambda superstrings applied to vaccine design. PLoS ONE. (2019) 14:e0211714. doi: 10.1371/journal.pone.0211714
5. Sanchez-Ramon S, Conejero L, Netea MG, Sancho D, Palomares O, Subiza JL. Trained immunity based-vaccines: a new paradigm for the development of broad-spectrum anti-infectious formulations. Front Immunol. (2018) 9:2936. doi: 10.3389/fimmu.2018.02936
6. Netea MG, Joosten LA, Latz E, Mills KH, Natoli G, Stunnenberg HG, et al. Trained immunity: a program of innate immune memory in health and disease. Science. (2016) 352:aaf1098. doi: 10.1126/science.aaf1098
7. Sanchez-Cerrillo I, Landete P, Aldave B, Sanchez-Alonso S, Sanchez-Azogra A, Marcos-Jimenez A, et al. Differential redistribution of activated monocytes and dendritic cell subsets to the lung associates with severity of Covid-19. medRxiv [Preprint]. (2020). doi: 10.1101/2020.05.13.20100925
8. Miyasaka M. Is BCG vaccination causally related to reduced COVID-19 mortality? EMBO Mol Med. (2020) 7:e12661. doi: 10.15252/emmm.202012661
9. O'Neil LAJ, Netea MG. BCG-induced trained immunity: can it offer protection against COVID-19? Nat Rev Immunol. (2020) 20:335–7. doi: 10.1038/s41577-020-0337-y
10. McGill F, Heyderman RS, Panagiotou S, Tunkel AR, Solomon T. Acute bacterial meningitis in adults. Lancet. (2016) 388:3036–47. doi: 10.1016/S0140-6736(16)30654-7
11. Gonzalez-Santiago TM, Drage LA. Nontuberculous mycobacteria. skin and soft tissue infections. Dermatol Clin. (2015) 33:563–77. doi: 10.1016/j.det.2015.03.017
12. Godshall CE, Suh G, Lorber B. Cutaneous listeriosis. J Clin Microbiol. (2013) 51:3591–6. doi: 10.1128/JCM.01974-13
13. Johansson L, Thulin P, Low DE, Norrby-Teglund A. Getting under the skin: the immunopathogenesis of Streptococcus pyogenes deep tissue infections. Clin Infect Dis. (2010) 51:58–65. doi: 10.1086/653116
14. Weinberger B. Vaccines for the elderly: current use and future challenges. Immun Ageing. (2018) 215:3. doi: 10.1186/s12979-017-0107-2
15. Alecsandru D, Valor L, Sanchez-Ramon S, Gil J, Carbone J, Navarro J, et al. Sublingual therapeutic immunization with a polyvalent bacterial preparation in patients with recurrent respiratory infections: immunomodulatory effect on antigen-specific memory CD4+ T cells and impact on clinical outcome. Clin Exp Immunol. (2011) 164:100–7. doi: 10.1111/j.1365-2249.2011.04320.x
16. Sanchez-Ramon S, de Diego RP, Dieli-Crimi R, Subiza JL. Extending the clinical horizons of mucosal bacterial vaccines: current evidence and future prospects. Curr Drug Targets. (2014) 15:1132–43. doi: 10.2174/1389450115666141020160705
17. Tejera-Alhambra M, Palomares O, Perez de Diego R, Diaz-Lezcano I, Sanchez-Ramon S. New biological insights in the immunomodulatory effects of mucosal polybacterial vaccines in clinical practice. Curr Pharm Des. (2016) 22:6283–93. doi: 10.2174/1381612822666160829143129
18. Cirauqui C, Benito-Villalvilla C, Sanchez-Ramon S, Sirvent S, Diez-Rivero CM, Conejero L, et al. Human dendritic cells activated with MV130 induce Th1, Th17 and IL-10 responses via RIPK2 and MyD88 signalling pathways. Eur J Immunol. (2018) 48:180–93. doi: 10.1002/eji.201747024
19. Ovejero-Guisasola JI, Fresno-Escudero M. Lipopolysaccharide of Ochrobactrum Intermedium and Their Use as Immunostimulant of Mamalians. Patent Number. WO 2010/39352. Global Catalogue of Microorganisms (2010). Available online at: https://patents.google.com/patent/WO2010139352A1/en (accessed March 27, 2021).
20. Calderon-Gonzalez R, Frande-Cabanes E, Teran-Navarro H, Marimon JM, Freire J, Salcines-Cuevas D, et al. GNP-GAPDH1−22 nanovaccines prevent neonatal listeriosis by blocking microglia apoptosis and bacterial dissemination. Oncotarget. (2017) 8:53916–34. doi: 10.18632/oncotarget.19405
21. Calderon-Gonzalez R, Teran-Navarro H, Frande-Cabanes E, Fernandez-Ferrandez E, Freire J, Penades S, et al. Pregnancy vaccination with gold glyco-nanoparticles carrying Listeria monocytogenes peptides protects against listeriosis and brain- and cutaneous-associated morbidities. Nanomaterials. (2016) 6:pii: E151. doi: 10.3390/nano6080151
22. Clements JD, Norton EB. The mucosal vaccine adjuvant LT(R192G/L211A) or dmLT. mSphere. (2018) 3:e00215–18. doi: 10.1128/mSphere.00215-18
23. Alves J, Madureira P, Baltazar MT, Barros L, Oliveira K, Dinis-Oliveira RJ, et al. A safe and stable neonatal vaccine targeting GAPDH confers protection against Group B Streptococcus infections in adult susceptible mice. PLoS ONE. (2015) 10:e0144196. doi: 10.1371/journal.pone.0144196
24. Alvarez-Dominguez C, Madrazo-Toca F, Fernandez-Prieto L, Vandeckerhove J, Pareja E, Tobes R, et al. Characterization of a Listeria monocytogenes protein interfering with Rab5a. Traffic. (2008) 9:325–37. doi: 10.1111/j.1600-0854.2007.00683.x
25. Alvarez-Dominguez C, Carrasco-Marin E. Peptides Which are Immunogenic in Relation to the Genuses Listeria and Mycobacterium, Antibodies and Uses of these. Patent Number. W02008020108A1 (2006). Available online at: https://patents.google.com/patent/WO2008020108A1/fi#patentCitations (accessed March 27, 2021).
26. Alvarez-Dominguez C, Calderon-Gonzalez R, Teran-Navarro H, Salcines-Cuevas D, Frande-Cabanes E, Garcia I, et al. Multivalent Vaccine for the Treatment and Prevention of Tuberculosis, Listeriosis and Pneumonia. Patent Number. WO2019243647 (2019). Available online at: https://patentscope.wipo.int/search/es/detail.jsf?docId=WO2019243647 (accessed March 27, 2021).
27. Calderon-Gonzalez R, Teran-Navarro H, Marimon JM, Gonzalez-Rico C, Calvo-Montes J, Frande-Cabanes E, et al. Biomarkers tools to design clinical vaccines determined from a study of annual listeriosis incidence in northern Spain. Front Immunol. (2016) 7:541. doi: 10.3389/fimmu.2016.00541
28. Alvarez-Dominguez C, Salcines-Cuevas D, Teran-Navarro H, Calderon-Gonzlez R, Tobes R, Garcia I, et al. Epitopes for multivalent vaccines against Listeria, Mycobacterium and Streptococcus spp: a novel role for glyceraldehyde-3-phosphate dehydrogenase. Front Cell Infect Microbiol. (2020) 10:573348. doi: 10.3389/fcimb.2020.573348
29. Pardi N, Hogan MJ, Porter FW, Weissman D. “mRNA vaccines - a new era in vaccinology”. Nat Rev Drug Discov. (2018) 17:261–79. doi: 10.1038/nrd.2017.243
30. Midoux P, Pichon C. Lipid-based mRNA vaccine delivery systems. Expert Rev Vaccines. (2015) 14:221–34 doi: 10.1586/14760584.2015.986104
31. Flanagan K, Best E, Crawford NW, Giles M, Koirala A, Macartney K, et al. Progress and pitfalls in the quest for effective SARS-CoV-2 (COVID-19) vaccines. Front Immunol. (2020) 11:570250. doi: 10.3389/fimmu.2020.579250
32. Calderon-Gonzalez R, Terán-Navarro H, García I, Marradi M, Salcines-Cuevas D, Yañez-Diaz S, et al. Gold glyconanoparticles coupled to listeriolysin O 91-99 peptide serve as adjuvant therapy for solid tumours. Nanoscale. (2017) 9:10721–31. doi: 10.1039/C7NR02494K
33. Ryou SM, Kim JM, Yeom JH, Hyun S, Kim Sw, Bae J, et al. Gold nanoparticle-assisted delivery of small, highly structured RNA into the nuclei of human cells. Biochem Biophys Res Commun. (2011) 416:178–83. doi: 10.1016/j.bbrc.2011.11.020
34. Alvarez-Dominguez C, Stahl PD. Increased expression of Rab5a correlates directly with accelerated maturation of Listeria monocytogenes phagosomes. J Biol Chem. (1999) 274:17:11459–62. doi: 10.1074/jbc.274.17.11459
35. Singh R, Jha D, Gautam HK, Kumar P. Supramolecular self-assemblies of engineered polyethylenimines as multifunctional nanostructures for DNA transportation with excellent antimicrobial activity. Bioorg Chem. (2020) 6:104463. doi: 10.1016/j.bioorg.2020.104463
36. Calegari LP, Dias RS, de Oliveira MD, Pessoa CR, de Oliveira AS, Oliveira AF, et al. Multi-walled carbon nanotubes increase antibody-producing B cells in mice immunized with a tetravalent vaccine candidate for dengue virus. J Nanobiotechnology. (2016) 14:61. doi: 10.1186/s12951-016-0196-7
37. Calderon-Gonzalez R, Garcia I, Marradi M, Petrovsky N, Alvarez-Dominguez C. Novel nanoparticles vaccines. Human Vaccin Immunother. (2015) 11:2501–3. doi: 10.1080/21645515.2015.1063756
38. Kim JH, Jang HH, Ryou SM, Kim S, Bae J, Lee K, et al. A functionalized gold nanoparticles-assisted universal carrier for antisense DNA. Chem Commun. (2010) 46:4151–3. doi: 10.1039/c0cc00103a
39. Calderon-Gonzalez R, Frande-Cabanes E, Bronchalo-Vicente L, Lecea-Cuello MJ, Bosch-Martinez A, Fanarraga ML, et al. Cellular vaccines in listeriosis: role of the Listeria antigen GAPDH. Front Cell Infect Microbiol. (2014) 4:22. doi: 10.3389/fcimb.2014.00022
40. Kono M, Nakamura Y, Suda T, Uchijima M, Tsujimara K, Nagata T, et al. Enhancement of protective immunity against intracellular bacteria using type-1 polarized dendritic cell (DC) vaccine. Vaccine. (2012) 30:2633–9. doi: 10.1016/j.vaccine.2012.02.026
41. Myllymäki H, Niskanen M, Oksanen KE, Sheerwood E, Ahava M, Parikka M, et al. Identification of novel antigen candidates for a tuberculosis vaccine in the adult zebrafish (Danio rerio). PLoS ONE. (2017) 12:e0181942. doi: 10.1371/journal.pone.0181942
42. Kwon BE, Ahn JH, Min S, Kim H, Seo J, Yeo S, et al. Development of new preventive and therapeutic vaccines for tuberculosis. Immune Netw. (2018) 18:e17. doi: 10.4110/in.2018.18.e17
43. D'Orazio SEF, Halme DG, Ploegh HL, Starnbach M. Class Ia MHC-deficient BALB/c mice generate CD8+ T cell-mediated protective immunity against Listeria monocytogenes infection. J Immunol. (2003) 171:291–8. doi: 10.4049/jimmunol.171.1.291
44. Blum M, Chang H, Chuguransky S, Grego T, Kandasaamy S, Mitchell A, et al. The InterPro protein families and domains database: 20 years on. Nucleic Acids Res. (2021) 49:D344–54. doi: 10.1093/nar/gkaa977
45. Peters B, Sette A. Generating quantitative models describing the sequence specificity of biological processes with the stabilized matrix method. BMC Bioinformatics. (2005) 6:132. doi: 10.1186/1471-2105-6-132
46. Stier EM, Mandal M, Lee KD. Differential cytosolic delivery and presentation of antigen by listeriolysin O-liposomes to macrophages and dendritic cells. Mol Pharm. (2005) 2:74–82. doi: 10.1021/mp049896v
47. Buchanan RM, Briles DE, Arulanandam BP, Westerink MA, Raeder RH, Metzger DW. IL-12 mediated increases in protection elicited by pneumococcal and meningococcal conjugate vaccine. Vaccine. (2001) 19:2020–8. doi: 10.1016/S0264-410X(00)00421-7
48. Rodriguez-Del Rio E, Marradi M, Calderon-Gonzalez R, Frande-Cabanes E, Penadés S, Petrovsky N, et al. A gold glyco-nanoparticle carrying a Listeriolysin O peptide and formulated with Advax™ delta inulin adjuvant induces robust T-cell protection against listeria infection. Vaccine. (2015) 33:1465–73. doi: 10.1016/j.vaccine.2015.01.062
Keywords: glyceraldehyde-3-phosphate-dehydrogenase, listeriosis, pneumonia, tuberculosis, cross-reactive vaccines, innate immunity
Citation: Teran-Navarro H, Salcines-Cuevas D, Calderon-Gonzalez R, Tobes R, Calvo-Montes J, Pérez-Del Molino Bernal IC, Yañez-Diaz S, Fresno M and Alvarez-Dominguez C (2021) A Comparison Between Recombinant Listeria GAPDH Proteins and GAPDH Encoding mRNA Conjugated to Lipids as Cross-Reactive Vaccines for Listeria, Mycobacterium, and Streptococcus. Front. Immunol. 12:632304. doi: 10.3389/fimmu.2021.632304
Received: 23 November 2020; Accepted: 15 February 2021;
Published: 19 April 2021.
Edited by:
Jose Luis Subiza, Inmunotek SL, SpainReviewed by:
Nicholas M. Provine, University of Oxford, United KingdomHaizhen Wu, East China University of Science and Technology, China
Copyright © 2021 Teran-Navarro, Salcines-Cuevas, Calderon-Gonzalez, Tobes, Calvo-Montes, Pérez-Del Molino Bernal, Yañez-Diaz, Fresno and Alvarez-Dominguez. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Carmen Alvarez-Dominguez, Y2FybWVuLmFsdmFyZXpAdW5pci5uZXQ= orcid.org/0000-0002-4585-6959
†These authors have contributed equally to this work
‡Present address: Ricardo Calderon-Gonzalez, Wellcome-Wolfson Institute for Experimental Medicine-Queen's University Belfast, Belfast, United Kingdom