 Mohd Rehan1,2*†
Mohd Rehan1,2*† Firoz Ahmed3,4*†
Firoz Ahmed3,4*† Saad M. Howladar5
Saad M. Howladar5 Mohammed Y. Refai3,4
Mohammed Y. Refai3,4 Hanadi M. Baeissa3Torki A. Zughaibi1,2Khalid Mohammed Kedwa6Mohammad Sarwar Jamal7
Hanadi M. Baeissa3Torki A. Zughaibi1,2Khalid Mohammed Kedwa6Mohammad Sarwar Jamal7- 1King Fahd Medical Research Center, King Abdulaziz University, Jeddah, Saudi Arabia
- 2Department of Medical Laboratory Technology, Faculty of Applied Medical Sciences, King Abdulaziz University, Jeddah, Saudi Arabia
- 3Department of Biochemistry, College of Science, University of Jeddah, Jeddah, Saudi Arabia
- 4University of Jeddah Center for Research and Product Development, University of Jeddah, Jeddah, Saudi Arabia
- 5Department of Biology, College of Science, University of Jeddah, Jeddah, Saudi Arabia
- 6Hematology Department, Regional Lab Makkah, Haddah, Saudi Arabia
- 7Integrative Biosciences Center, Wayne State University, Detroit, MI, United States
Background: The newly identified betacoronavirus SARS-CoV-2 is the causative pathogen of the coronavirus disease of 2019 (COVID-19) that killed more than 3.5 million people till now. The cytokine storm induced in severe COVID-19 patients causes hyper-inflammation, is the primary reason for respiratory and multi-organ failure and fatality. This work uses a rational computational strategy to identify the existing drug molecules to target host pathways to reduce the cytokine storm.
Results: We used a “host response signature network” consist of 36 genes induced by SARS-CoV-2 infection and associated with cytokine storm. In order to attenuate the cytokine storm, potential drug molecules were searched against “host response signature network”. Our study identified that drug molecule andrographolide, naturally present in a medicinal plant Andrographis paniculata, has the potential to bind with crucial proteins to block the TNF-induced NFkB1 signaling pathway responsible for cytokine storm in COVID-19 patients. The molecular docking method showed the binding of andrographolide with TNF and covalent binding with NFkB1 proteins of the TNF signaling pathway.
Conclusion: We used a rational computational approach to repurpose existing drugs targeting host immunomodulating pathways. Our study suggests that andrographolide could bind with TNF and NFkB1 proteins, block TNF-induced cytokine storm in COVID-19 patients, and warrant further experimental validation.
Introduction
The ongoing pandemic by severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) infected more than 168 million people worldwide, causing more than 3.5 million deaths as of 26 May 2021 (https://www.worldometers.info/coronavirus/). SARS-CoV-2 infects the lower respiratory tract resulting in a severe respiratory disease called coronavirus disease of 2019 (COVID-19) (1). The majority of COVID-19 patients have mild symptoms, including cough, fever, body pain, which recover in a few days. However, a significant number of patients developed severe cases with difficulty in breath, respiratory and lung failure, organ damage, and even death (2–4). Accumulating studies found that an excessively high level of pro-inflammatory cytokines released, called cytokine storm, in response to SARS-CoV-2 infection triggers acute respiratory distress syndrome (ARDS) and multi-organ failure in COVID-19 patients (2, 4). The most prominent cytokines elevated in the blood plasma of severe COVID-19 patients include TNF, IL-6, IL-8, IL-10, and IL-2 (5, 6).
SARS-CoV-2 is classified as betacoronavirus closely related to two previously identified human pathogenic SARS-CoV and MERS-CoV, which caused the epidemics in 2002 and 2012, respectively (7). The SARS-CoV-2 infects the host cell with its spike protein that binds to the ACE2 receptor on the host cell surface and facilitates viral entry. SARS-CoV-2 has a plus-strand RNA genome of about 29,900 nucleotides that work as mRNA to encode viral proteins using host machinery. The genome also contains several cis-acting RNA elements that could be involved in viral infection and replications (8). Studies indicated that SARS-CoV-2 has evolutionary closely related to bat-derived SARS-CoV (8, 9).
Till now, no safe and effective therapy has been approved to cure COVID-19. However, life support treatment is recommended for the management and treatment of COVID-19 patients based on the symptoms and severity of the cases.
Based upon multiple clinical trial data on mild-to-severe COVID-19 patients, the US Food and Drug Administration (FDA) has approved only a single antiviral drug Veklury (remdesivir), for the treatment of COVID-19 on 22 October 2020. Furthermore, the FDA authorized eight drugs for emergency use to treat severe COVID-19 patients, and numerous other therapeutics are currently being under clinical trials (https://www.fda.gov/media/136832/download). The FDA is working with different organizations to facilitate the development of safe and effective drug molecules in combating COVID-19.
The elevated level of cytokines is the primary cause of severity in COVID-19 patients, and hence, attenuating the level of these cytokines would be a better approach to manage and treat the COVID-19 patients. Therefore, it is urgently required to identify the drug molecules for targeting the critical pathways in the human that could minimize cytokine storm in COVID-19. However, developing a new medicine is a scientifically challenging process that requires a very long time and massive money with the risk of failure. With the high rate of SARS-CoV-2 infection and the rise of new variant strains, drug repurposing became an extremely important process to identify the potential candidate drugs for treating the severe symptoms of COVID-19. Drug repurposing refers to an approach to identifying new therapeutic uses for approved or investigational drugs other than the scope of the original medical indication (10, 11). Since the repurpose drugs already passed the early-stage trial, including safety assessment, efficacy, toxicity, pharmacokinetic, pharmacodynamic, and preclinical testing, this approach has several advantages: drugs are sufficiently safe, low risk of failure, less investment, and shorten the timeline, could instantly start the tested trials in patients (11). Several drugs were repurposed against numerous diseases (12, 13). Furthermore, various computational methods, including molecular docking and dynamic simulations, have increasingly being used for structural insights into the action mechanism of existing drugs and for novel drug designing (14–18).
This study used a rational computational approach to repurpose drug molecules to minimize the cytokine storm associated with COVID-19 patients. Our previous study identified the SARS-CoV-2-mediated activation of the “host response signature network” responsible for cytokine storm (5). In this work, we identified andrographolide to target the “host response signature network” through a virtual drug screening method. Finally, we employed a structure-based molecular docking method to demonstrate that the andrographolide could bind with TNF and NFkB1, potentially blocking the TNF-mediated NFkB1 pathway underpinning cytokine storm in severe patients with COVID-19.
Materials and Methods
Host Regulatory Network and Biological Pathway Analysis
This study used the “host response signature network” identified in the human cells infected with SARS-CoV-2 in our previous study (5). Briefly, transcriptome data of SARS-CoV-2 infected normal human bronchial epithelial (NHBE) cells and controlled NHBE cells were used to identify the differentially expressed host genes (DEHGs) (19). A network was created by integrating DEHGs with human protein-protein interaction data. Furthermore, analysis of the network revealed an important sub-network of highly inter-connected 31 proteins (IL6, TNF, CXCL8, CXCL3, CXCL5, IRF9, SAA1, OAS3, CSF2, IFI6, OAS2, CSF3, IRF7, ICAM1, CXCL2, MX1, OAS1, MMP9, IL1A, IL1B, C3, TLR2, IFI27, CXCL6, CXCL1, CCL20, XAF1, IFI44L, MX2, BST2, IFITM1) predominantly involved in cytokine storm and under the regulation of transcription factors STAT1, STAT2, STAT3, POU2F2, and NFkB1. The sub-network of 31 proteins and their five master regulators are called as “host response signature network”. In order to understand the biological pathways associated with the “host response signature network”, KEGG (Kyoto Encyclopedia of Genes and Genomes) pathway enrichment analysis was performed using DAVID 6.8 (https://david.ncifcrf.gov/home.jsp).
Potential Drugs Against “Host Response Signature Network”
The “host response signature network” was analyzed to find potential drug molecules using the “Gene Association” module of Network Data Exchange (NDEx version 2.4.5) (https://ndexbio.org/) (20, 21). NDEx is an open-source platform for biological network analysis and knowledge discovery. The output of “Gene Association” analysis gives networks of query gene/protein and its direct or indirect interaction with chemical and drug molecules. All output networks of drug-target interaction were merged to make a non-redundant network and visualized with Cytoscape software version 3.7. Subsequently, we analyzed the potential drug molecules targeting the “host response signature network”. For further study, we selected a drug molecule, andrographolide, naturally present in a medicinal plant used to treat common cold and inflammation (22).
Data Retrieval
The three-dimensional coordinates of andrographolide were retrieved from the PubChem database with PubChem CID, 5318517. The crystal structure of human NFkB1, p50 subunit, with bound native DNA was selected and retrieved from the PDB database with PDB Id, 2V2T. The crystal structure of human TNF as a dimer with bound native inhibitor was selected and retrieved from the PDB database with PDB Id, 5MU8.
Predicting the Drug‐Likeness and Pharmacokinetic Properties
The drug‐likeness and pharmacokinetic properties, including absorption, distribution, metabolism, excretion, and toxicity, were analyzed using the online tool “pkCSM‐pharmacokinetics” (http://biosig.unimelb.edu.au/pkcsm/). This machine learning-based method uses the various graph-based signature of chemical compounds having sets of distance patterns between atoms (23).
Covalent Docking of Andrographolide to NFkB1
It was reported that andrographolide is a covalent inhibitor of NFkB1 and makes a covalent bond with Cys-62 of NFkB1 (24, 25). Autodock Vina (26) was used for covalent docking of andrographolide to NFkB1. To perform covalent docking, the reported atom of the andrographolide was linked to the sulfide atom of Cys-62 through a covalent bond (25). The adduct Cys-andrographolide was made flexible for rotation through various bonds using Autodock Tools (27). Then the docking of a very small molecule (e.g., water molecule) was performed, leading to the relaxed conformation of the flexible adduct Cys-andrographolide. Finally, this relaxed conformation of the adduct Cys-andrographolide was reported as the final covalent docking of andrographolide. Autodock Tools was used to prepare the protein, the ligand, and the grid box required for docking by Autodock Vina.
Molecular Docking and Protein-Ligand Complex Analysis
For normal molecular docking, we used Dock version 6.5 (28). For TNF, the homodimer (Chains A and B) was used for molecular docking, and the native inhibitor bound within the binding site formed by both monomers (Chains A and B) was used as a probe for the TNF homodimer binding site. The protein and ligand preparation required for molecular docking and visualization at different stages of docking was performed using Chimera version 1.6.2 (29). The illustrations for binding poses were generated using Pymol version 2.4.0 (Schrödinger, LLC) and the protein-ligand interaction plots were prepared using Ligplot+ version 2.1 (30). The binding energy and dissociation constant scores were predicted using XScore version 1.2.11 (31).
Results and Discussion
The integration and analysis of high-throughput biological data are extensively used to identify altered regulatory networks and the potential target molecules of complex diseases (5, 32). Furthermore, virtual drug screening and molecular docking were used to find therapeutic drug molecules against target molecules to treat the disease (17, 18). Therapeutic molecules against SARS-CoV-2 infection could be developed using two major approaches: (i) design the therapeutic molecules against the crucial targets of the virus (33–36); (ii) identify the disease-related gene regulatory network and then design the therapeutic molecules against it. Our previous work identified the “host response signature network” associated with the influx of cytokine storm through the TNF-induced NFkB1 signaling pathway (5). Our study also suggested vitamin D’s role in reducing the cytokine storm and viruses (5). In this work, we employed the later drug development strategy in which the “host response signature network” induced in the SARS-CoV-2 infection was used as targets to repurpose drug molecules to reduce the cytokine storm responsible for severe COVID-19.
Biological Pathways of “Host Response Signature Network”
The “host response signature network” was analyzed with the KEGG pathway, which identified several significantly associated biological pathways. We found that TNF signaling pathway is significantly associated with IL6, CSF2, CCL20, IL1B, CXCL1, CXCL3, TNF, CXCL2, MMP9, NFkB1, and ICAM1 of the “host response signature network” and responsible for the induction of cytokine storm (Figure 1). The complete list of all KEGG pathways is provided in the Supplementary Table S1.
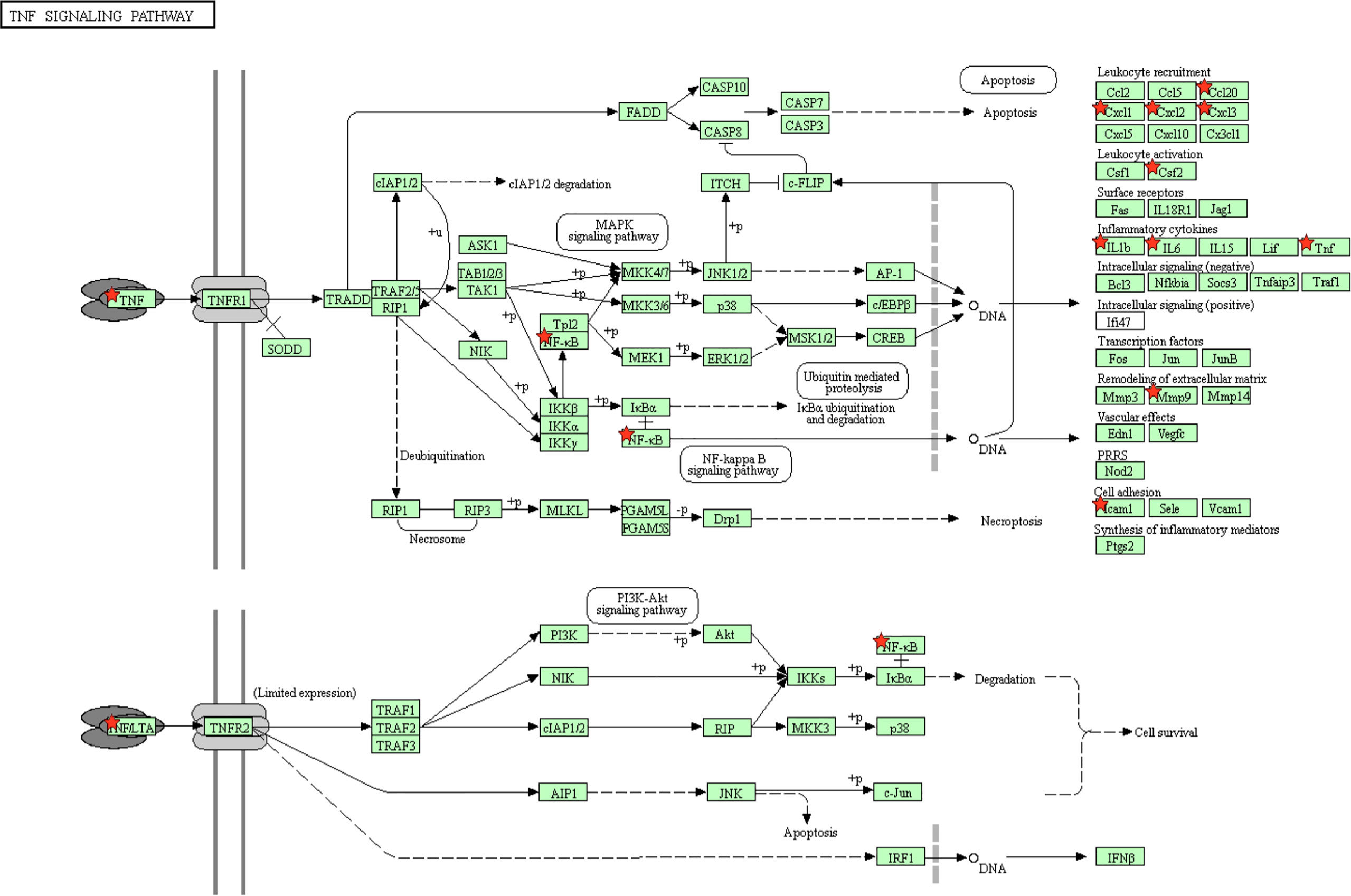
Figure 1 The KEGG pathway enrichment analysis showed that the “host response signature network” is associated with the TNF signaling pathway. The proteins IL6, CSF2, CCL20, IL1B, CXCL1, CXCL3, TNF, CXCL2, MMP9, NFKB1, and ICAM1 of the “host response signature network” are represented as red stars.
Potential Drug Molecules Targeting “Host Response Signature Network”
Analysis of potential drug molecules for “host response signature network” with the “Gene Association” module of online tool NDEx returns a list of drug-target interactions networks (Table 1). The list of drug-target interactions from different networks was merged to create a non-redundant drug-target network (Figure 2). The detailed information of drug molecules is provided in the Supplementary Table S2. The data was further investigated to find a potential drug for better management of the cytokine storm in severe COVID-19 patients. We have selected andrographolide for further study based upon the following criteria from the list of repurposed drugs.
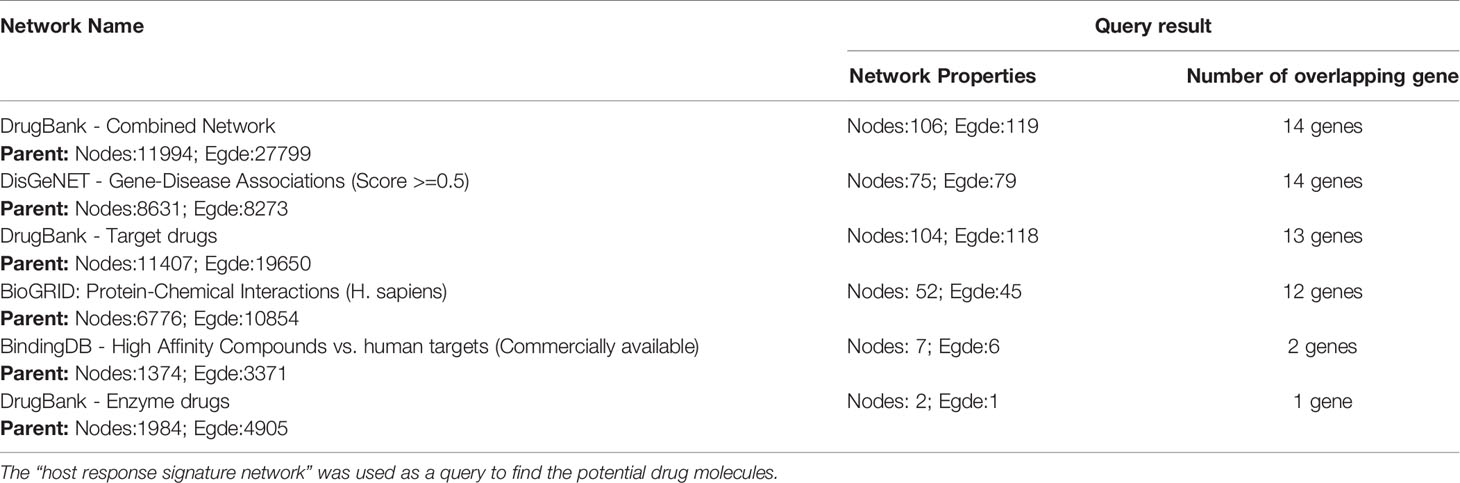
Table 1 Drug-target networks were retrieved using the “Gene Association” module of NDEx v2.4.5.
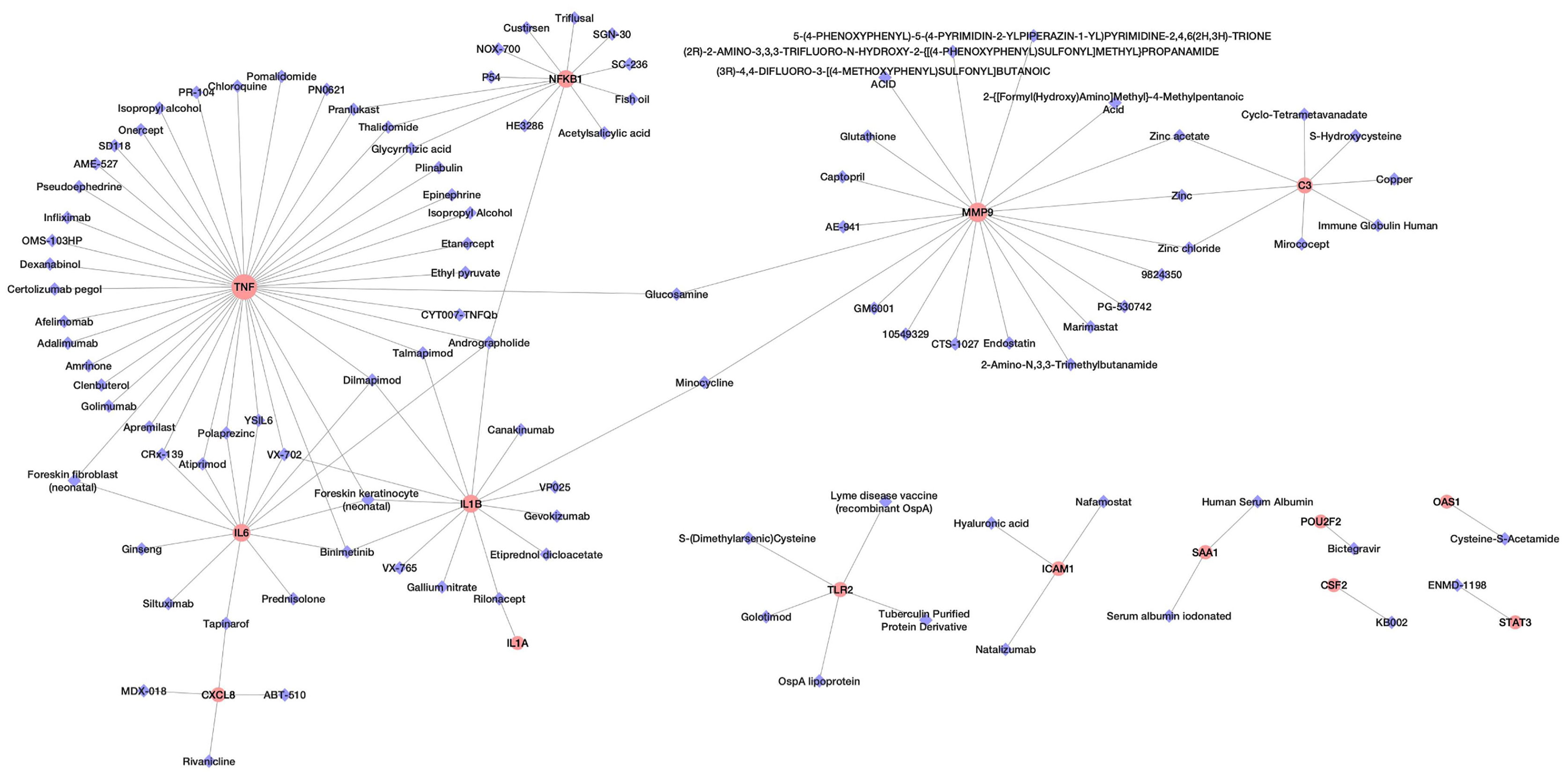
Figure 2 The network of drug molecules targeting the “host response signature network”.
(a) The drug must target/suppress the highest number of proteins of the “host response signature network”; (b) targeted proteins associate with the same biological pathway; (c) targeted proteins play a crucial role in the cytokine storm; and (d) a medicinal plant could be a source of the drug.
We found that andrographolide can target four different proteins IL1B, NFkB1, TNF, and IL6 (Table S2A) of the TNF signaling pathway (Figure 1), which might result in blockage of the cytokine storm in COVID-19. Andrographolide is a labdane diterpenoid isolated from Andrographis paniculata, also known as the “king of bitters”, a medicinal plant that mainly grows in Asian countries (Figure 3). Andrographis paniculata traditionally used in Unani, Ayurvedic, and Chinese herbal medicines with a broad range of therapeutic applications, including treating a common cold, upper respiratory tract infections, and inflammation (22).
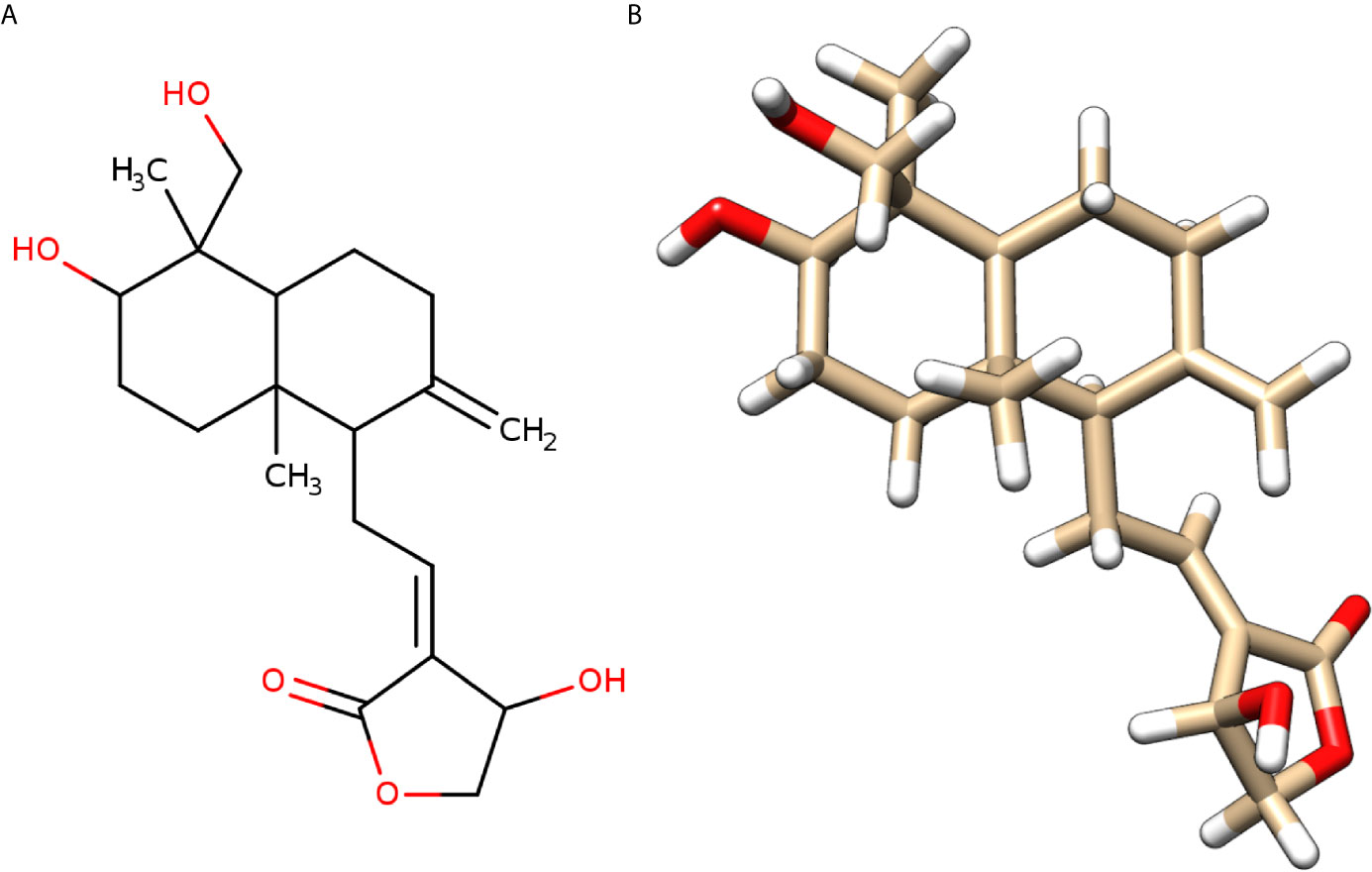
Figure 3 Molecular structure of andrographolide. (A) Two-dimensional sketch of andrographolide with O-atoms and -OH groups in red color. (B) Three-dimensional diagram of andrographolide with O-atoms in red and H-atoms in white colors.
Drug-Likeness and Pharmacokinetic Properties
As shown in Table 2, the andrographolide was having a low molecular weight of 350.46 (< 500) and the lipophilicity (LogP) value of 1.96 (<5), H-bond donors 3 (<5) and acceptors 5 (<10), and three rotatable bonds (<10). The values for all five conditions (Lipinski’s Rule of Five) for the andrographolide were well within the desired range of a drug molecule. While most of the pharmacokinetic properties, including absorption, distribution, metabolism, excretion, and toxicity, were acceptable for andrographolide (Table 3). Therefore, this suggests that andrographolide is a potentially safe drug candidate for use in humans.
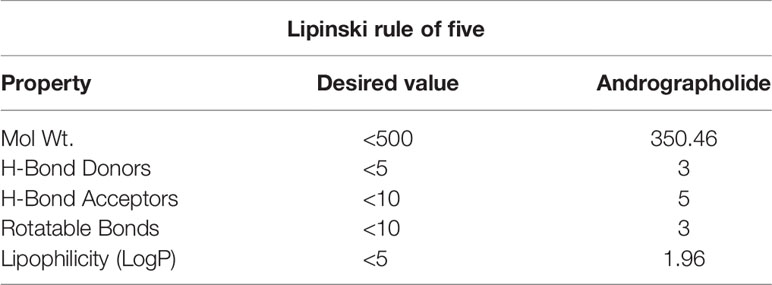
Table 2 Drug-likeness (Lipinski rule of five) for andrographolide.
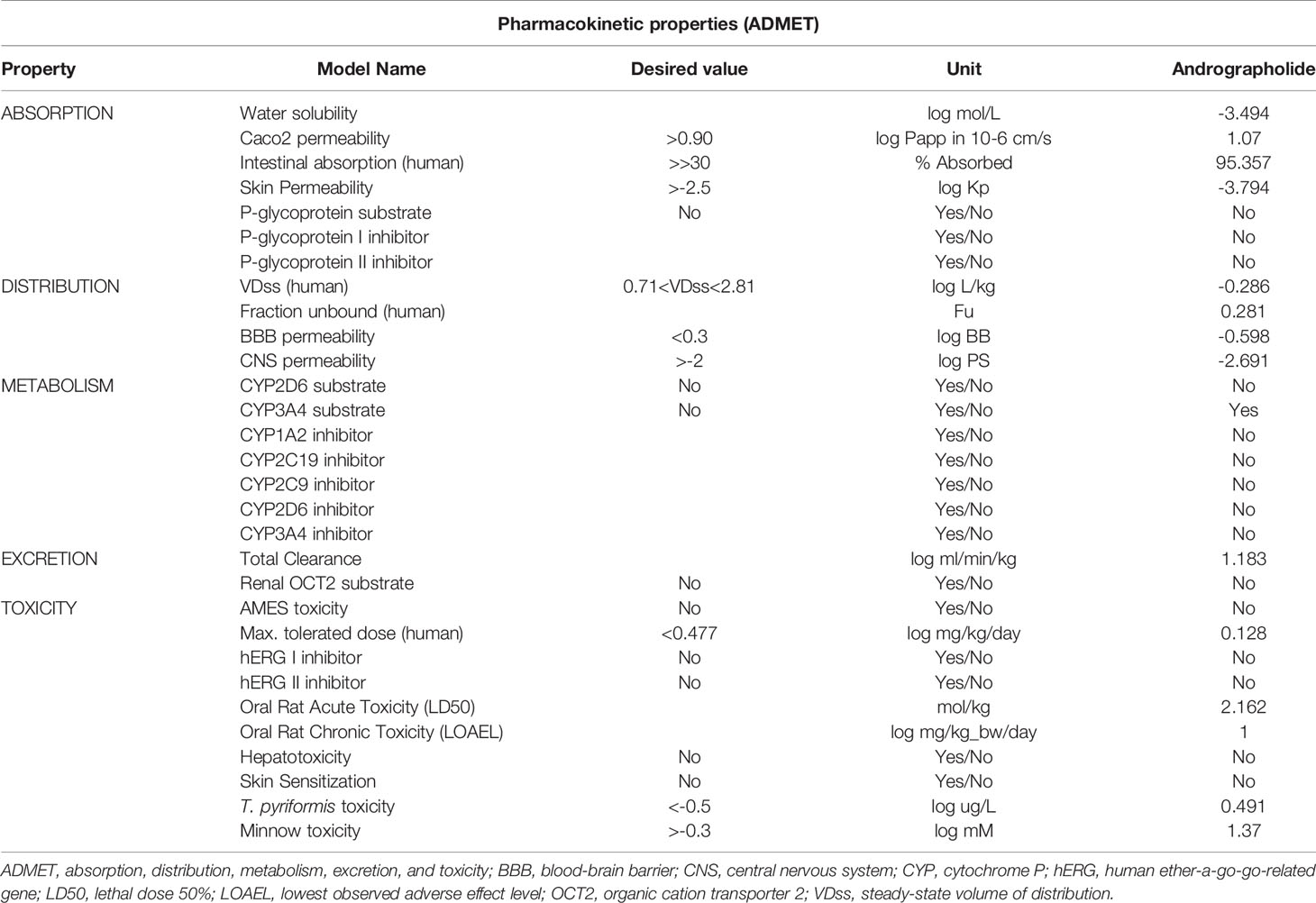
Table 3 Pharmacokinetic properties (ADMET) prediction for andrographolide.
Covalent Docking Analysis of Andrographolide With NFkB1
The PDB structures of IL1B and IL6 are available as a complex with their receptors (for example, PDB Ids: 3O4O and 1P9M, respectively) and with the antibody (for example, PDB Ids: 4G6J and 4CNI, respectively). The information for the binding site of ILIB and IL6 for a chemical compound was not obvious. So, we proceeded with the docking of andrographolide to NFkB1 and TNF only.
The experimental laboratories showed that andrographolide reacted with the sulfhydryl group (-SH) of Cys-62 of NFkB1 and formed a covalent bond (24, 25). One of the studies mentioned above further proceed to a normal docking of andrographolide to NFkB1, and they reported Cys-62 and Arg-57 as the interacting residues forming hydrogen bonds (25). We tried normal docking considering Cys-62 as a clue for the binding site and failed to see a binding site cavity close to Cys-62 residue where this compound may fit. Therefore, we proceeded with the covalent docking of andrographolide to NFkB1. The docking results showed that the andrographolide formed a covalent bond and a hydrogen bond (2.86 Å) with the sulfide atom of Cys-62 (Figure 4). The Cys-62 was also involved in 13 non-bonding contacts with andrographolide (Table 4). Another interacting residue Glu-63 was involved in another 13 non-bonding contacts. Thus, the andrographolide formed an adduct with Cys-62 and became part of the protein. In addition, it formed a hydrogen bond and 26 non-bonding interactions with the protein (Figure 4 and Table 4). Thus, our study suggests andrographolide could block the DNA binding site, interfering with the binding of DNA, and thus the transcription factor NFkB1 would not be able to carry out its function. This is in agreement with previous findings, which revealed that andrographolide binds with NFkB1, which blocks the binding of NFkB1 to DNA and prevents transcriptional activity (24, 25).
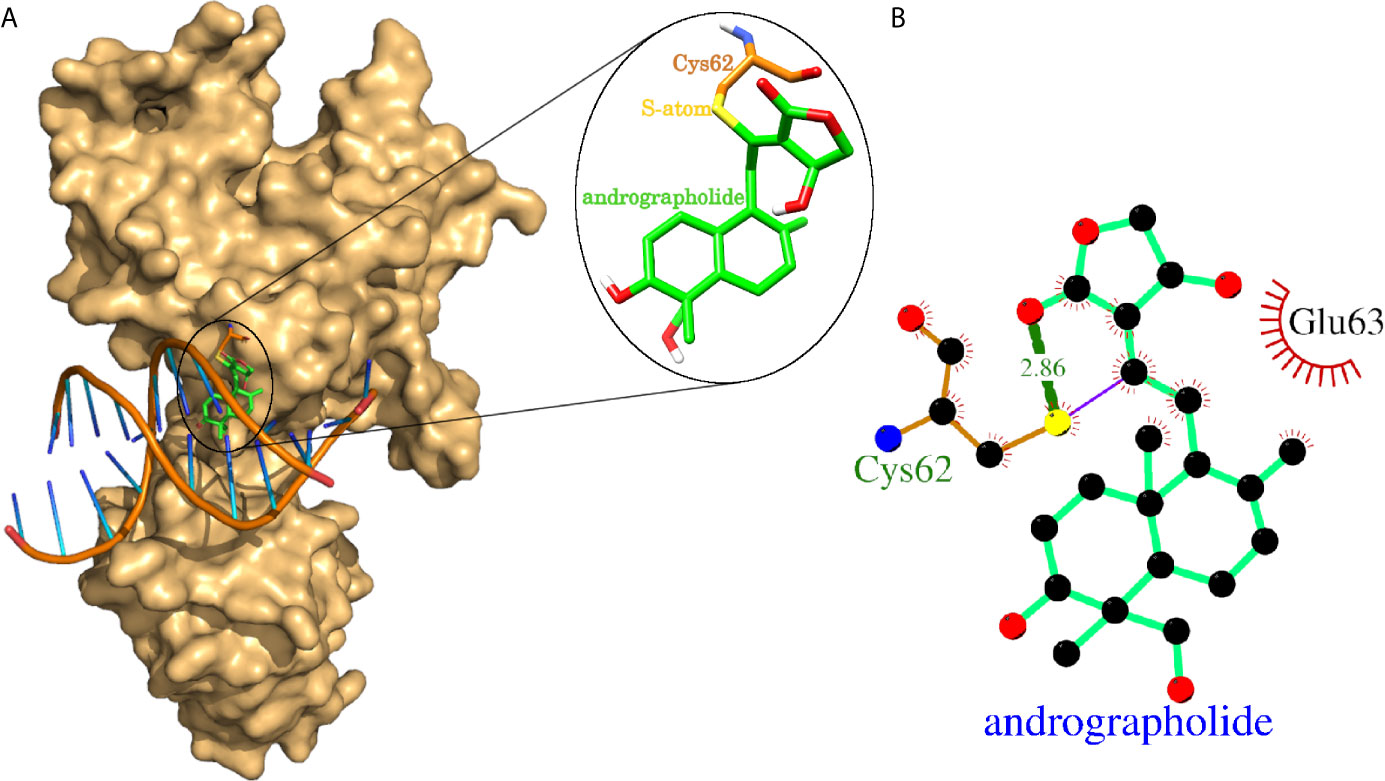
Figure 4 Covalent docking of andrographolide to human NFkB1. (A) NFkB1 is shown in surface representation colored light orange with bound native double-helical DNA and docked andrographolide. The docked andrographolide forming Cys-andrographolide adduct was shown as a close-up in the inset. The andrographolide, part of the adduct, was shown in sticks representation having carbon backbone in green color with heteroatoms oxygen, nitrogen, sulfur, and hydrogen atoms in red, blue, yellow, and white colors respectively. The Cys-62 part of the adduct forming the covalent bond through sulfur atom was also shown in sticks representation with the backbone in orange color. (B) Protein-ligand interaction plot of andrographolide with NFkB1. The andrographolide and hydrogen bonding residues are shown in ball and sticks representation with balls representing atoms and sticks representing the bond between two atoms. The color of balls distinguishes among atom types as C-atom in black, O-atoms in red, N-atoms in blue, and S-atom in yellow colors. The non-bonded interactions labeled with interacting residue are shown as red arcs with bristles, covalent bonds as purple line, and hydrogen bonds as thick green lines labeled with bond length in Å.

Table 4 The NFkB1 residues interacting with andrographolide.
Molecular Docking Analysis of Andrographolide With TNF Homodimer
The andrographolide docked well within the binding site formed by two monomeric units of TNF (Figure 5A) and stabilized by 20 non-bonded contacts and one hydrogen bond through nine interacting residues (Figure 5B and Table 5). The absolute values of binding scores, including dock score (-32.67), binding energy (-7.52 Kcal/mol), and dissociation constant measure pKd (5.51), were also reasonably high required for the stable protein-ligand complex. The nine interacting residues include four residues from chain A (Tyr-59, Tyr-119, Leu-120, and Gly-121) and five residues from chain B (Leu-94, Tyr-119, Leu-120, Gly-121, and Gly-122). Interestingly, the three identical residues Tyr-119, Leu-120, and Gly-121 from both the monomers, played a role in binding. Of nine interacting residues, Gly-121(B) formed hydrogen bonding interaction with the andrographolide with bond length 3.06 Å and also formed eight (maximum) non-bonded contacts. Therefore, the Gly-121(B) is proposed as the key residue playing a role in binding. While comparing the binding of andrographolide to that of the native inhibitor JNJ525 (Figures 5B, C), it was found that the andrographolide was also binding to the same site and sharing common six interacting residues Tyr-59 (A), Tyr-119 (A), Gly-121 (A), Tyr-119 (B), Gly-121 (B), and Gly-122 (B). Thus, the andrographolide was binding in the site where the TNF inhibitor is binding and inhibiting TNF activity.
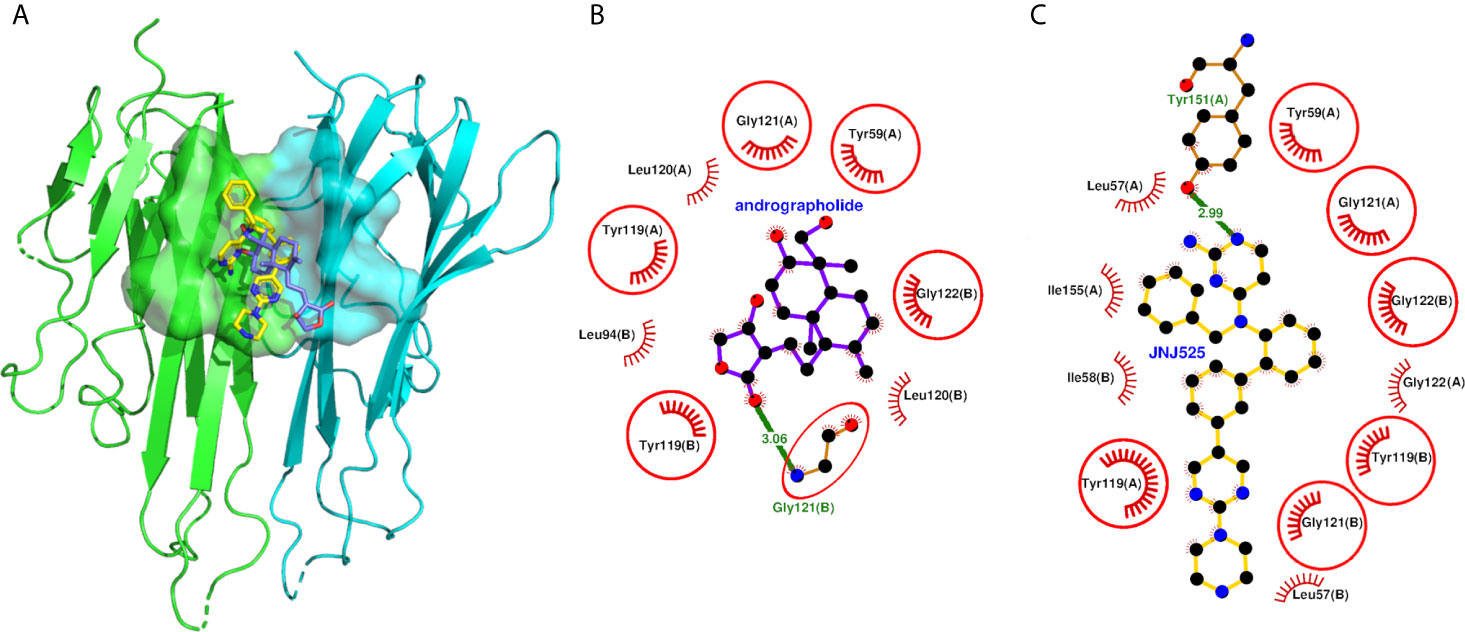
Figure 5 Molecular docking of andrographolide with human TNF homodimer. (A) The pose of andrographolide within the binding site of TNF homodimer. The NFkB1 is shown in cartoon representation colored green (Chain A) and cyan (Chain B) with the binding site in surface representation. The andrographolide and the native inhibitor are shown as sticks colored blue and yellow, respectively, with red oxygen and blue nitrogen atoms. (B, C) Protein-ligand interaction plots of andrographolide and the native inhibitor JNJ525 with TNF homodimer. The andrographolide and hydrogen bonding residue are shown in ball and sticks representation, with balls representing atoms and sticks representing the bond between two atoms. The color of balls distinguishes among atom types as C-atom in black, O-atoms in red, and N-atoms in blue colors. The non-bonded interactions labeled with interacting residue are shown as red arcs with bristles, while the hydrogen bond is shown as a green line labeled with bond length in Å. The residues common among interacting residues of both the native inhibitor and the andrographolide are encircled.

Table 5 The residues of TNF homodimer interacting with andrographolide.
Previous studies showed that andrographolide has a potent inhibitor of wide verities of viruses, including influenza virus (37, 38), hepatitis C virus (39), herpes simplex virus type 1 (40), Chikungunya virus (41). It also has the potential to inhibit the proliferation of lung carcinoma (42) and nasopharyngeal carcinoma (43), and tumor metastasis (44). Another report demonstrated that mice treated with andrographolide reduce the inflammation and fibrosis of damaged liver (45). It also prevents liver neutrophil infiltration and reduced TNF and COX-2 signaling (45). A recent study with the molecular docking method suggested andrographolide as a potential inhibitor of SARS-CoV-2 main protease Mpro (47). Another computational-based study found that andrographolide and its derivative, 14-deoxy-11,12-didehydroandrographolide, have a high binding affinity with three target proteins of SARS-CoV-2, i.e., main protease, papain-like protease (PLpro), and spike protein (48). In addition, the same study used a DIGEP-Pred tool and predicted that both compounds induce the level of several proteins involved in regulating immune systems, including the NFkB signaling pathway (48).
The NFkB1 is a family of transcription factors, which regulate the expression of various genes of pro-inflammatory cytokines, including IL6 and IL8 (CXCL8), mainly responsible for cytokine storm in COVID-19 (Figure 1) (5, 49). NFkB1 inhibitory protein (IkB) binds with NFkB1 dimer in an unstimulated cell, which prevents movement of NFkB1 from the cytoplasm to the nucleus. In response to SARS-CoV-2 infection, the TNF signaling pathway is activated, which causes degradation of IkB, resulting in the release and translocation of NFkB1 to the nucleus for transcription of various cytokine genes (5, 49). Our study found that andrographolide bound well with TNF and NFkB1 and provided structural insights into their binding. This binding of andrographolide blocks the TNF signaling pathways at these two critical steps, thus prevents the expression of various cytokine genes responsible for the influx of cytokine in COVID-19.
In addition, our study identified Dilmapimod as potential compounds to reduce the cytokine storm through targeting/suppressing IL1B, TNF, and IL6 (Table S2A). Activation of the p38 MAPK signaling pathway is associated with lung inflammation in asthma and chronic obstructive pulmonary disease (COPD) (50). The p38 MAPK regulated NFkB dependent transcription of multiple inflammatory cytokines (51). Clinical studies showed that Dilmapimod, a novel inhibitor of p38 MAPK, decreases the TNF and IL1B level in the blood of COPD patients (52, 53). Another study also suggested the potential use of Dilmapimod for COVID-19 management (54). Furthermore, our analysis identified that Prednisolone, a corticosteroid, has the potential to suppress the level of IL6 (Table S2A); thus, supported previous work indicating that Prednisolone could be useful in treating severe COVID-19 (55).
Previous studies showed the anti-inflammatory and anticancer effects of Dehydroxymethylepoxyquinomicin (DHMEQ), a new inhibitor of NFkB (56, 57). Another study found that DHMEQ reduces allergic airway inflammation and cytokines in a mice model of asthma (58). An investigation conducted in Jurkat T-lymphoblastic leukemia cells found that DHMEQ prevents the TNF-α-induced nuclear translocation of NFkB (46). These findings also raised the potential use of DHMEQ in inhibiting cytokine storms and thus need to be studied in more detail in treating COVID-19.
Our study used a rational computational approach to identify the potential drug molecule for COVID-19. Though it is associated with the following limitations: (i) we used the “host response signature network” identified in the NHBE cells infected with SARS-CoV-2 (5). The work included a limited sample size of transcriptomic data with three infected and three controlled groups (19). (ii) This study lacks in vitro drug screening data on human lung cells infected with SARS-CoV-2. Therefore, it is difficult to draw the effect of andrographolide on the “host response signature network” in SRAS-CoV-2 infected cells. (iii) Furthermore, our study lacks the experimental evaluation of the dose-dependent effect of the drug and its associated risk.
Conclusion
The SARS-CoV-2 infection resulted in the COVID-19 pandemic killed millions of people around the world. Therefore, it is urgently needed to develop safe and effective drug molecules in a limited time to combat COVID-19. The influx of cytokines in COVID-19 patients is the prominent reason for organ damage and death. Thus, our study used the information of altered regulatory network inducing cytokine storm in COVID-19 and then repurposed andrographolide as a potential drug molecule to reduce the cytokine storm. Molecular docking analysis showed that andrographolide could inhibit NFkB1 and TNF, and thus block the pathways responsible for cytokine storm. This naturally occurring compound possesses drug-like properties and could be a promising drug for further biochemical and cell-based experimental validation for combating severe COVID-19.
Data Availability Statement
The original contributions presented in the study are included in the article/Supplementary Material. Further inquiries can be directed to the corresponding authors.
Author Contributions
FA conceived the idea, generated the data, analyzed and interpreted the results, wrote and revised the manuscript, and designed and guided the project. MR improved the idea, generated the data, analyzed and interpreted the results, wrote, and revised the manuscript. SH and MYR verified the generated data and wrote and revised the manuscript. HB and TZ verified the computational methods, wrote and revised the manuscript. KK and MJ wrote and revised the manuscript. All authors contributed to the article and approved the submitted version.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
The authors would like to acknowledge the technical support provided by the King Fahd Medical Research Center, King Abdulaziz University; Department of Biochemistry, Department of Biology, University of Jeddah; and University of Jeddah Center for Research and Product Development (UJCRPD), University of Jeddah.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fimmu.2021.648250/full#supplementary-material
References
1. Ren LL, Wang YM, Wu ZQ, Xiang ZC, Guo L, Xu T, et al. Identification of a Novel Coronavirus Causing Severe Pneumonia in Human: A Descriptive Study. Med J (Engl) (2020) 133:1015–24. doi: 10.1097/CM9.0000000000000722
2. Moore BJB, June CH. Cytokine Release Syndrome in Severe COVID-19. Science (2020) 368:eabb8925. doi: 10.1126/science.abb8925
3. Yang X, Yu Y, Xu J, Shu H, Xia J, Liu H, et al. Clinical Course and Outcomes of Critically Ill Patients With SARS-Cov-2 Pneumonia in Wuhan, China: A Single-Centered, Retrospective, Observational Study. Lancet Respir Med (2020) 8:475–81. doi: 10.1016/S2213-2600(20)30079-5
4. Wang T, Du Z, Zhu F, Cao Z, An Y, Gao Y, et al. Comorbidities and Multi-Organ Injuries in the Treatment of COVID-19. Lancet (2020) 395(10228):e52. doi: 10.1016/S0140-6736(20)30558-4
5. Ahmed F. A Network-Based Analysis Reveals the Mechanism Underlying Vitamin D in Suppressing Cytokine Storm and Virus in SARS-Cov-2 Infection. Front Immunol (2020) 11(3084):590459. doi: 10.3389/fimmu.2020.590459
6. Zhang C, Wu Z, Li JW, Zhao H, Wang GQ. Cytokine Release Syndrome in Severe COVID-19: Interleukin-6 Receptor Antagonist Tocilizumab May Be the Key to Reduce Mortality. Int J Antimicrob Agents (2020) 55(5):105954. doi: 10.1016/j.ijantimicag.2020.105954
7. Choudhry H, Bakhrebah MA, Abdulaal WH, Zamzami MA, Baothman OA, Hassan MA, et al. Middle East Respiratory Syndrome: Pathogenesis and Therapeutic Developments. Future Virol (2019) 14(4):237–46. doi: 10.2217/fvl-2018-0201
8. Ahmed F, Sharma M, Al-Ghamdi AA, Al-Yami SM, Al-Salami AM, Refai MY, et al. A Comprehensive Analysis of Cis-Acting RNA Elements in the SARS-Cov-2 Genome by a Bioinformatics Approach. Front Genet (2020) 11(1385):1385. doi: 10.3389/fgene.2020.572702
9. Tang X, Wu C, Li X, Song Y, Yao X, Wu X, et al. On the Origin and Continuing Evolution of SARS-Cov-2. Natl Sci Rev (2020) 7(6):1012–23. doi: 10.1093/nsr/nwaa036
10. Ashburn TT, Thor KB. Drug Repositioning: Identifying and Developing New Uses for Existing Drugs. Nat Rev Drug Discovery (2004) 3(8):673–83. doi: 10.1038/nrd1468
11. Pushpakom S, Iorio F, Eyers PA, Escott KJ, Hopper S, Wells A, et al. Drug Repurposing: Progress, Challenges and Recommendations. Nat Rev Drug Discov (2019) 18(1):41–58. doi: 10.1038/nrd.2018.168
12. Kale VP, Habib H, Chitren R, Patel M, Pramanik KC, Jonnalagadda SC, et al. Old Drugs, New Uses: Drug Repurposing in Hematological Malignancies. Semin Cancer Biol (2020) 68:242–8. doi: 10.1016/j.semcancer.2020.03.005
13. Kumar R, Harilal S, Gupta SV, Jose J, Thomas Parambi DG, Uddin MS, et al. Exploring the New Horizons of Drug Repurposing: A Vital Tool for Turning Hard Work Into Smart Work. Eur J Med Chem (2019) 182:111602. doi: 10.1016/j.ejmech.2019.111602
14. Ferreira LG, Dos Santos RN, Oliva G, Andricopulo AD. Molecular Docking and Structure-Based Drug Design Strategies. Molecules (2015) 20(7):13384–421. doi: 10.3390/molecules200713384
15. Rehan M, Mahmoud MM, Tabrez S, Hassan HMA, Ashraf GM. Exploring Flavonoids for Potential Inhibitors of a Cancer Signaling Protein PI3Kγ Kinase Using Computational Methods. Anticancer Res (2020) 40(8):4547–56. doi: 10.21873/anticanres.14460
16. Rehan M. Anticancer Compound XL765 as PI3K/mTOR Dual Inhibitor: A Structural Insight Into the Inhibitory Mechanism Using Computational Approaches. PloS One (2019) 14(6):e0219180. doi: 10.1371/journal.pone.0219180
17. Rehan M, Mostafa M. Virtual Screening of 1,4-Naphthoquinone Derivatives for Inhibition of a Key Cancer Signaling Protein, AKT1 Kinase. Anticancer Res (2019) 39(7):3823–33. doi: 10.21873/anticanres.13532
18. Rehan M. A Structural Insight Into the Inhibitory Mechanism of an Orally Active PI3K/Mtor Dual Inhibitor, PKI-179 Using Computational Approaches. J Mol Graph Model (2015) 62:226–34. doi: 10.1016/j.jmgm.2015.10.005
19. Blanco-Melo D, Nilsson-Payant BE, Liu WC, Uhl S, Hoagland D, Møller R, et al. Imbalanced Host Response to SARS-Cov-2 Drives Development of COVID-19. Cell (2020) 181(5):1036–45.e9. doi: 10.1016/j.cell.2020.04.026
20. Pratt D, Chen J, Welker D, Rivas R, Pillich R, Rynkov V, et al. Ndex, the Network Data Exchange. Cell Syst (2015) 1(4):302–5. doi: 10.1016/j.cels.2015.10.001
21. Pratt D, Chen J, Pillich R, Rynkov V, Gary A, Demchak B, et al. Ndex 2.0: A Clearinghouse for Research on Cancer Pathways. Cancer Res (2017) 77(21):e58–61. doi: 10.1158/0008-5472.CAN-17-0606
22. Akbar S. Andrographis Paniculata: A Review of Pharmacological Activities and Clinical Effects. Altern Med Rev (2011) 16(1):66–77.
23. Pires DE, Blundell TL, Ascher DB. Pkcsm: Predicting Small-Molecule Pharmacokinetic and Toxicity Properties Using Graph-Based Signatures. J Med Chem (2015) 58(9):4066–72. doi: 10.1021/acs.jmedchem.5b00104
24. Xia YF, Ye BQ, Li YD, Wang JG, He XJ, Lin X, et al. Andrographolide Attenuates Inflammation by Inhibition of NF-Kappa B Activation Through Covalent Modification of Reduced Cysteine 62 of P50. J Immunol (2004) 173(6):4207–17. doi: 10.4049/jimmunol.173.6.4207
25. Wang J, Tan XF, Nguyen VS, Yang P, Zhou J, Gao M, et al. A Quantitative Chemical Proteomics Approach to Profile the Specific Cellular Targets of Andrographolide, a Promising Anticancer Agent That Suppresses Tumor Metastasis. Mol Cell Proteomics (2014) 13(3):876–86. doi: 10.1074/mcp.M113.029793
26. Trott O, Olson AJ. Autodock Vina: Improving the Speed and Accuracy of Docking With a New Scoring Function, Efficient Optimization, and Multithreading. J Comput Chem (2010) 31(2):455–61. doi: 10.1002/jcc.21334
27. Morris GM, Huey R, Lindstrom W, Sanner MF, Belew RK, Goodsell DS, et al. Autodock4 and Autodocktools4: Automated Docking With Selective Receptor Flexibility. J Comput Chem (2009) 30(16):2785–91. doi: 10.1002/jcc.21256
28. Ewing TJ, Makino S, Skillman AG, Kuntz ID. DOCK 4.0: Search Strategies for Automated Molecular Docking of Flexible Molecule Databases. J Comput Aided Mol Des (2001) 15(5):411–28. doi: 10.1023/a:1011115820450
29. Pettersen EF, Goddard TD, Huang CC, Couch GS, Greenblatt DM, Meng EC, et al. UCSF Chimera–A Visualization System for Exploratory Research and Analysis. J Comput Chem (2004) 25(13):1605–12. doi: 10.1002/jcc.20084
30. Laskowski RA, Swindells MB. Ligplot+: Multiple Ligand-Protein Interaction Diagrams for Drug Discovery. J Chem Inf Model (2011) 51(10):2778–86. doi: 10.1021/ci200227u
31. Wang R, Lai L, Wang S. Further Development and Validation of Empirical Scoring Functions for Structure-Based Binding Affinity Prediction. J Comput Aided Mol Des (2002) 16(1):11–26. doi: 10.1023/a:1016357811882
32. Ahmed F. Integrated Network Analysis Reveals FOXM1 and MYBL2 as Key Regulators of Cell Proliferation in Non-Small Cell Lung Cancer. Front Oncol (2019) 9:1011. doi: 10.3389/fonc.2019.01011
33. Wu C, Liu Y, Yang Y, Zhang P, Zhong W, Wang Y, et al. Analysis of Therapeutic Targets for SARS-Cov-2 and Discovery of Potential Drugs by Computational Methods. Acta Pharm Sin B (2020) 10(5):766–88. doi: 10.1016/j.apsb.2020.02.008
34. Gupta AK, Khan MS, Choudhury S, Mukhopadhyay A, Sakshi, Rastogi A, et al. Coronavr: A Computational Resource and Analysis of Epitopes and Therapeutics for Severe Acute Respiratory Syndrome Coronavirus-2. Front Microbiol (2020) 11:1858. doi: 10.3389/fmicb.2020.01858
35. Ahmed F, Raghava GP. Designing of Highly Effective Complementary and Mismatch SiRNAs for Silencing a Gene. PloS One (2011) 6(8):e23443. doi: 10.1371/journal.pone.0023443
36. Tyagi A, Ahmed F, Thakur N, Sharma A, Raghava GP, Kumar M. Hivsirdb: A Database of HIV Inhibiting SiRNAs. PloS One (2011) 6(10):e25917. doi: 10.1371/journal.pone.0025917
37. Chen JX, Xue HJ, Ye WC, Fang BH, Liu YH, Yuan SH, et al. Activity of Andrographolide and Its Derivatives Against Influenza Virus In Vivo and In Vitro. Biol Pharm Bull (2009) 32(8):1385–91. doi: 10.1248/bpb.32.1385
38. Yu B, Dai CQ, Jiang ZY, Li EQ, Chen C, Wu XL, et al. Andrographolide as an Anti-H1N1 Drug and the Mechanism Related to Retinoic Acid-Inducible Gene-I-Like Receptors Signaling Pathway. Chin J Integr Med (2014) 20(7):540–5. doi: 10.1007/s11655-014-1860-0
39. Lee JC, Tseng CK, Young KC, Sun HY, Wang SW, Chen WC, et al. Andrographolide Exerts Anti-Hepatitis C Virus Activity by Up-Regulating Haeme Oxygenase-1 via the P38 MAPK/Nrf2 Pathway in Human Hepatoma Cells. Br J Pharmacol (2014) 171(1):237–52. doi: 10.1111/bph.12440
40. Seubsasana S, Pientong C, Ekalaksananan T, Thongchai S, Aromdee C. A Potential Andrographolide Analogue Against the Replication of Herpes Simplex Virus Type 1 in Vero Cells. Med Chem (2011) 7(3):237–44. doi: 10.2174/157340611795564268
41. Wintachai P, Kaur P, Lee RC, Ramphan S, Kuadkitkan A, Wikan N, et al. Activity of Andrographolide Against Chikungunya Virus Infection. Sci Rep (2015) 5:14179. doi: 10.1038/srep14179
42. Luo X, Luo W, Lin C, Zhang L, Li Y. Andrographolide Inhibits Proliferation of Human Lung Cancer Cells and the Related Mechanisms. Int J Clin Exp Med (2014) 7(11):4220–5.
43. Peng T, Hu M, Wu TT, Zhang C, Chen Z, Huang S, et al. Andrographolide Suppresses Proliferation of Nasopharyngeal Carcinoma Cells via Attenuating NF-κB Pathway. BioMed Res Int 2015 (2015) p:735056. doi: 10.1155/2015/735056
44. Yu CC, Chen CA, Fu SL, Lin HY, Lee MS, Chiou WY, et al. Andrographolide Enhances the Anti-Metastatic Effect of Radiation in RAS-Transformed Cells via Suppression of ERK-Mediated MMP-2 Activity. PloS One (2018) 13(10):e0205666. doi: 10.1371/journal.pone.0205666
45. Lee TY, Chang HH, Wen CK, Huang TH, Chang YS. Modulation of Thioacetamide-Induced Hepatic Inflammations, Angiogenesis and Fibrosis by Andrographolide in Mice. J Ethnopharmacol (2014) 158 Pt A:423–30. doi: 10.1016/j.jep.2014.10.056
46. Ariga A, Namekawa J, Matsumoto N, Inoue J, Umezawa K. Inhibition of Tumor Necrosis Factor-Alpha -Induced Nuclear Translocation and Activation of NF-Kappa B by Dehydroxymethylepoxyquinomicin. J Biol Chem (2002) 277(27):24625–30. doi: 10.1074/jbc.M112063200
47. Enmozhi SK, Raja K, Sebastine I, Joseph J. Andrographolide as a Potential Inhibitor of SARS-CoV-2 Main Protease: An In Silico Approach. J Biol Chem (2021) 39(9):3092–8. doi: 10.1080/07391102.2020.1760136
48. Khanal P, Dey YN, Patil R, Chikhale R, Wanjari MM, Gurav SS, et al. Combination of System Biology to Probe the Anti-Viral Activity of Andrographolide and Its Derivative Against COVID-19. RSC Adv (2021) 11(9):5065–79. doi: 10.1039/D0RA10529E
49. Liu T, Zhang L, Joo D, Sun SC. Nf-κb Signaling in Inflammation. Signal Transduct Target Ther (2017) 2:e17023. doi: 10.1038/sigtrans.2017.23
50. Pelaia C, Vatrella A, Gallelli L, Lombardo N, Sciacqua A, Savino R, et al. Role of P38 Mitogen-Activated Protein Kinase in Asthma and COPD: Pathogenic Aspects and Potential Targeted Therapies. Drug Des Devel Ther (2021) 15:1275–84. doi: 10.2147/DDDT.S300988
51. Carter AB, Knudtson KL, Monick MM, Hunninghake GW. The P38 Mitogen-Activated Protein Kinase Is Required for NF-Kappab-Dependent Gene Expression. The Role of TATA-Binding Protein (TBP). J Biol Chem (1999) 274(43):30858–63. doi: 10.1074/jbc.274.43.30858
52. Singh D, Smyth L, Borrill Z, Sweeney L, Tal-Singer R. A Randomized, Placebo-Controlled Study of the Effects of the P38 MAPK Inhibitor SB-681323 on Blood Biomarkers of Inflammation in COPD Patients. J Clin Pharmacol (2010) 50(1):94–100. doi: 10.1177/0091270009347873
53. Betts JC, Mayer RJ, Tal-Singer R, Warnock L, Clayton C, Bates S, et al. Gene Expression Changes Caused by the P38 MAPK Inhibitor Dilmapimod in COPD Patients: Analysis of Blood and Sputum Samples From a Randomized, Placebo-Controlled Clinical Trial. Pharmacol Res Perspect (2015) 3(1):e00094. doi: 10.1002/prp2.94
54. Patra T, Ray R. IL-6 Induction and Signaling: Horizons of COVID-19-Related Pathogenesis. DNA Cell Biol (2021) 40(5):639–42. doi: 10.1089/dna.2021.0152
55. Gopalaswamy R, Subbian S. Corticosteroids for COVID-19 Therapy: Potential Implications on Tuberculosis. Int J Mol Sci (2021) 22(7):3773. doi: 10.3390/ijms22073773
56. Nishioka C, Ikezoe T, Jing Y, Umezawa K, Yokoyama A. DHMEQ, a Novel Nuclear Factor-Kappab Inhibitor, Induces Selective Depletion of Alloreactive or Phytohaemagglutinin-Stimulated Peripheral Blood Mononuclear Cells, Decreases Production of T Helper Type 1 Cytokines, and Blocks Maturation of Dendritic Cells. Immunology (2008) 124(2):198–205. doi: 10.1111/j.1365-2567.2007.02755.x
57. Lin Y, Ukaji T, Koide N, Umezawa K. Inhibition of Late and Early Phases of Cancer Metastasis by the NF-κb Inhibitor DHMEQ Derived From Microbial Bioactive Metabolite Epoxyquinomicin: A Review. Int J Mol Sci (2018) 19(3):729. doi: 10.3390/ijms19030729
Keywords: SARS-CoV-2, COVID-19, cytokine storm, systems bioinformatics, drug-repurposing, andrographolide, TNF signaling pathway, molecular docking
Citation: Rehan M, Ahmed F, Howladar SM, Refai MY, Baeissa HM, Zughaibi TA, Kedwa KM and Jamal MS (2021) A Computational Approach Identified Andrographolide as a Potential Drug for Suppressing COVID-19-Induced Cytokine Storm. Front. Immunol. 12:648250. doi: 10.3389/fimmu.2021.648250
Received: 31 December 2020; Accepted: 31 May 2021;
Published: 24 June 2021.
Edited by:
Ioannis Zabetakis, University of Limerick, IrelandReviewed by:
Shima Shahjouei, Geisinger Health System, United StatesHifzur Rahman Ansari, King Abdullah International Medical Research Center (KAIMRC), Saudi Arabia
Naoshad Muhammad, Washington University in St. Louis, United States
Copyright © 2021 Rehan, Ahmed, Howladar, Refai, Baeissa, Zughaibi, Kedwa and Jamal. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Mohd Rehan, bXJlaGFuNzg2QGdtYWlsLmNvbQ==; Firoz Ahmed, ZmFobWVkMUB1ai5lZHUuc2E=
†These authors have contributed equally to this work