 Pierre Frange1,2,3
Pierre Frange1,2,3 Thomas Montange4,5Jérôme Le Chenadec6Damien Batalie4,5Ingrid Fert4,5Catherine Dollfus7
Thomas Montange4,5Jérôme Le Chenadec6Damien Batalie4,5Ingrid Fert4,5Catherine Dollfus7 Albert Faye8Stéphane Blanche1Anne Chacé9Corine Fourcade10Isabelle Hau11Martine Levine12
Albert Faye8Stéphane Blanche1Anne Chacé9Corine Fourcade10Isabelle Hau11Martine Levine12 Nizar Mahlaoui1Valérie Marcou13Marie-Dominique Tabone7Florence Veber1
Nizar Mahlaoui1Valérie Marcou13Marie-Dominique Tabone7Florence Veber1 Alexandre Hoctin6Thierry Wack6
Alexandre Hoctin6Thierry Wack6 Véronique Avettand-Fenoël2,14Josiane Warszawski6,15
Véronique Avettand-Fenoël2,14Josiane Warszawski6,15 Florence Buseyne4,5* on behalf of ANRS-EP59-Study Group
Florence Buseyne4,5* on behalf of ANRS-EP59-Study Group- 1Immunologie, hématologie et rhumatologie pédiatrique, hôpital Necker–Enfants malades, AP–HP- Centre – Université de Paris, Paris, France
- 2Laboratoire de microbiologie clinique, hôpital Necker–Enfants malades, AP–HP-Centre – Université de Paris, Paris, France
- 3EHU 7328 PACT, Institut Imagine, Université de Paris, Paris, France
- 4Unité Epidémiologie et Physiopathologie des Virus Oncogènes, Institut Pasteur, Paris, France
- 5Département de Virologie, UMR CNRS 3569 Institut Pasteur, Paris, France
- 6Départment d’épidémiologie, Centre de Recherche en Épidémiologie et Santé des Populations, INSERM U1018, Le Kremlin-Bicêtre, Villejuif, France
- 7Hémato-oncologie pédiatrique, Hôpital Trousseau, AP-HP, Paris, France
- 8Pédiatrie Générale, Hôpital Robert Debré, AP-HP, Paris, France
- 9Pédiatrie et néonatologie, Centre hospitalier intercommunal de Villeuneuve-Saint-Georges, Villeuneuve-Saint-Georges, France
- 10Pédiatrie Générale, Hôpital Bicêtre, AP-HP, Paris, France
- 11Pédiatrie Générale, Centre hospitalier intercommunal de Créteil, Créteil, France
- 12Immuno-hématologie pédiatrique, Hôpital Robert Debré, AP-HP, Paris, France
- 13Médecine et réanimation néonatale, Hôpital Cochin, AP-HP-Centre – Université de Paris, Paris, France
- 14CNRS 8104/INSERM U1016, Institut Cochin, Université Paris Descartes, Paris, France
- 15INED, Université Paris Sud, Le Kremlin-Bicêtre, Orsay, France
Background: The early initiation of antiretroviral therapy (ART) in HIV-1-infected infants reduces mortality and prevents early CD4 T-cell loss. However, the impact of early ART on the immune system has not been thoroughly investigated in children over five years of age or adolescents. Here, we describe the levels of naive CD4 and CD8 T lymphocytes (CD4/CD8TN), reflecting the quality of immune reconstitution, as a function of the timing of ART initiation (early (<6 months) versus late (≥24 months of age)).
Methods: The ANRS-EP59-CLEAC study enrolled 27 children (5-12 years of age) and nine adolescents (13-17 years of age) in the early-treatment group, and 19 children (L-Ch) and 21 adolescents (L-Ado) in the late-treatment group. T lymphocytes were analyzed by flow cytometry and plasma markers were analyzed by ELISA. Linear regression analysis was performed with univariate and multivariate models.
Results: At the time of evaluation, all patients were on ART and had a good immunovirological status: 83% had HIV RNA loads below 50 copies/mL and the median CD4 T-cell count was 856 cells/µL (interquartile range: 685-1236 cells/µL). In children, early ART was associated with higher CD8TN percentages (medians: 48.7% vs. 31.0%, P = 0.001), and a marginally higher CD4TN (61.2% vs. 53.1%, P = 0.33). In adolescents, early ART was associated with low CD4TN percentages and less differentiated memory CD8 T cells. CD4TN and CD8TN levels were inversely related to cellular activation and gut permeability.
Conclusion: In children and adolescents, the benefits of early ART for CD8TN were clear after long-term ART. The impact of early ART on CD4TN appears to be modest, because pediatric patients treated late respond to HIV-driven CD4 T-lymphocyte loss by the de novo production of TN cells in the thymus. Our data also suggest that current immune activation and/or gut permeability has a negative impact on TN levels.
Clinical Trial Registration: ClinicalTrials.gov, identifier NCT02674867.
Introduction
HIV-1-infected children receiving antiretroviral treatment (ART) face a lifetime of chronic disease due to this virus (1–3). Early ART initiation has an immediate beneficial impact, by blocking clinical progression (4), decreasing the cell-associated viral reservoir in infants (5) and favoring faster CD4 T-lymphocyte recovery (1, 6, 7). However, the benefit of early ART for CD4 T-cell levels may decrease after longer periods on ART. Early ART initiation may preserve naive CD8 T lymphocytes (CD8TN) (8) and CD4/CD8 ratio (9). Early ART reduces the frequency of HIV-specific CD4 and CD8 T cells in children, but preserves the capacity of these cells to produce several cytokines (10–12). Overall, findings concerning the impact of early ART on CD4 and CD8 T cells in pediatric patients remain limited.
Several gaps in our knowledge of the long-term impact of early versus late ART remain. Few studies have investigated the immunological benefits of early ART initiation in children over the age of five years and those without sustained viral suppression. The benefit of early ART for CD4 T-cell levels may persist or decrease over longer periods of long-term effective ART. Finally, unplanned treatment interruptions frequently occur during early childhood and adolescence, and it remains unclear whether the deleterious impact of viral rebound on the benefits of early ART is transient or irreversible (13–15).
The ANRS-EP59-CLEAC study investigated the immunological and virological characteristics of HIV-1-infected children over five years of age and adolescents, as a function of age at ART initiation (< 6 months vs. ≥ 24 months of age). We included participants with an initial period of viral suppression but without any criteria on the persistence of viral control (16). Here, we focused on the proportions of CD4TN and CD8TN, the major indicators of qualitative immune reconstitution (17), and their relationships with current and past HIV disease parameters and current immune activation (3).
Patients and Methods
Patients
The ANRS-EP59-CLEAC study was conducted in accordance with the Helsinki Declaration and the protocol was approved by the “Comité de protection des personnes île-de-France V”. Agreement to participate was obtained from the participants, if they were old enough to give an opinion, and written informed consent was obtained from at least one parent. The main inclusion criteria were (1) HIV-1 infection after vertical transmission, (2) participant aged 5 to 17 years at the time of the study, (3) ART initiation for therapeutic purposes before six months of age or after two years of age, (4) with initial virological success (HIV-1 RNA < 400 copies/mL achieved within 24 months of treatment initiation), regardless of the subsequent course of viremia. Between 2016 and 2019, two 15 mL blood samples were collected for biological evaluations, at two consecutive routine clinical follow-up visits.
Biological Evaluations
Total blood cell-associated HIV-1 DNA was quantified by ultrasensitive real-time PCR (18). The CD4 and CD8 T-lymphocyte subsets were quantified in fresh blood by flow cytometry with combinations of antibodies targeting the CD3, CD4, CD8α, CD45RA, CCR7, CD27, CD28, CD31, CD95, HLA-DR, and CD38 molecules(Supplementary Material). Data were collected on a Gallios cytometer (Beckman Coulter) and analyzed with Kaluza software (Beckman Coulter). The gating strategy involved the sequential definition of lymphocytes, CD4 and CD8 T lymphocytes, and their subsets according to CD45RA and CCR7 expression. The subsets were further characterized by using CD27 and CD28 expression to define naive (TN), central memory (TCM), transitional memory (TTM), effector memory (TEM) and effector (TE) cells; CD31 and CD95 expression was used to define recent thymic emigrants (TRTE) and stem cell memory cells (TSCM); and HLA-DR and CD38 expression was used to define activated T cells (Supplementary Material).
C-reactive protein (CRP), interleukin-6 (IL-6), CXCL10, soluble CD14 (sCD14), soluble CD163 (sCD163), and intestinal fatty acid-binding protein (iFABP) were quantified by ELISA (Supplementary Table 1). The Liaison XL CMV IgG (Diasorin) chemiluminescence immunoassay and the CMV R-Gene (Biomérieux) PCR assay were used to quantify cytomegalovirus (CMV) antibodies and DNA in plasma samples. The frequency of interferon (IFN)-γ-producing cells was quantified in an Elispot assay with anti-IFN-γ capture/detection antibodies (Diaclone 869.060.010), streptavidin alkaline phosphatase conjugate (GE Healthcare RPN4402) and BCIP/NBT color development substrate (S3771, Promega), according to the kit manufacturers’ instructions. Fresh peripheral blood mononuclear cells (PBMCs) were used to seed High Protein-Binding Immobilon-P Membrane P96 plates (MSIPS4510, Merk Millipore) at a density of 2 x 105 and 5 x 104 cells/well in RMPI supplemented with 10% fetal calf serum, with the addition of pp65 CMV peptides (#11549, NIH AIDS Reagent Program, 1 µg/mL of each peptide) or peptide diluent (DMSO). The cells were then cultured for 24 h. Spot-forming cells (SFCs) were counted with an ImmunoSpot S6 UV Image Analyzer (CTL, Bonn, Germany).
HIV History Variables
The patients were either included at birth in The Agence Nationale de Recherche sur le SIDA et les Hépatites Virales (ANRS) EPF/ANRS CO10 national prospective multicenter cohort or were managed at the centers of the ANRS/CO10 cohort and diagnosed before the age of 13 years (1). Clinical and biological data were therefore collected for HIV-infected children via the completion of standardized questionnaires at six-month intervals, or were collected retrospectively after inclusion in the CLEAC study. All ART regimens initiated consisted of highly active antiretroviral treatment (HAART; i.e., any combination of at least three different antiretroviral drugs or any combination including one protease inhibitor, or one non-nucleoside reverse transcriptase inhibitor or one integrase inhibitor). During the first few weeks of life, 30 children received prophylactic ART consisting of zidovudine (n=21), zidovudine+lamivudine (n=5), zidovudine+lamivudine+lopinavir (n=2) or zidovudine+lamivudine+nevirapine (n=2). For the last four of these children, the date of first HAART (ART1) initiation was the date of prophylactic ART initiation, as treatment was continued without interruption after diagnosis. At the time of the study, all but one of the children were on HAART, the remaining child being on two nucleoside reverse transcriptase inhibitors and having an undetectable viral load.
CD4 and CD8 T cells and plasma HIV RNA levels were quantified at each clinical site. The threshold values used in HIV RNA assays depended on study site, and a value of 50 copies/mL was selected as the common cutoff for current HIV RNA detection. A cutoff of 400 copies/mL was used for the assessment of virological history, because of changes in HIV RNA quantification assays and cutoffs over time. Cumulative viremia was defined as the area under the HIV RNA curve over time (19). Past immunological parameters were defined using CD4 T-cell percentages, because the age-related variations of this parameter are smaller than those for CD4 T-cell counts. The duration of viral suppression and cumulative viremia were assessed from ART1 initiation. These parameters were normalized by dividing their values by the time since ART1 initiation.
Statistical Analysis
Analyses were stratified according to age at evaluation (children: 5-12 years of age; adolescents: 13-17 years of age). Wilcoxon-Mann-Whitney, Kruskal-Wallis and the Fisher’s exact tests were used to compare the characteristics of children and adolescents between the early and late treatment initiation groups. The corrplot package of R was used to display Spearman’s rank correlation coefficients. Univariate and multivariate analyses were performed by linear regression, with CD4/CD8TN percentages as the continuous dependent variables. Multivariate models included early ART as the main independent variable, together with noncollinear variables with P values <.2 in univariate analysis. Most models were built with a single biological variable, to prevent over-adjustment. Observations with high leverage values or studentized residuals were excluded to test the robustness of the models. Analyses were conducted with SAS statistical software. P values <.05 were considered statistically significant.
Results
Patient Characteristics
We prospectively enrolled 27 children (E-Ch) and nine adolescents (E-Ado) in the early ART (E) group, and 19 children (L-Ch) and 21 adolescents (L-Ado) in the late ART (L) group (Table 1). Just over half (54%) of the patients were girls. Most patients (74%) were born to mothers originating from Sub-Saharan Africa. However, 60% of patients were born in mainland France, and ART was initiated early more frequently in these children than in those born abroad. At the time of the study, children were more frequently aviremic (89% vs. 73%, P = .07), and had higher CD4 T-cell counts than adolescents (median ([IQR]: 954 [745;1320] vs. 766 [622;1092] cells/µL, P = .04). Immunovirological status was good in most patients at the time of the study.
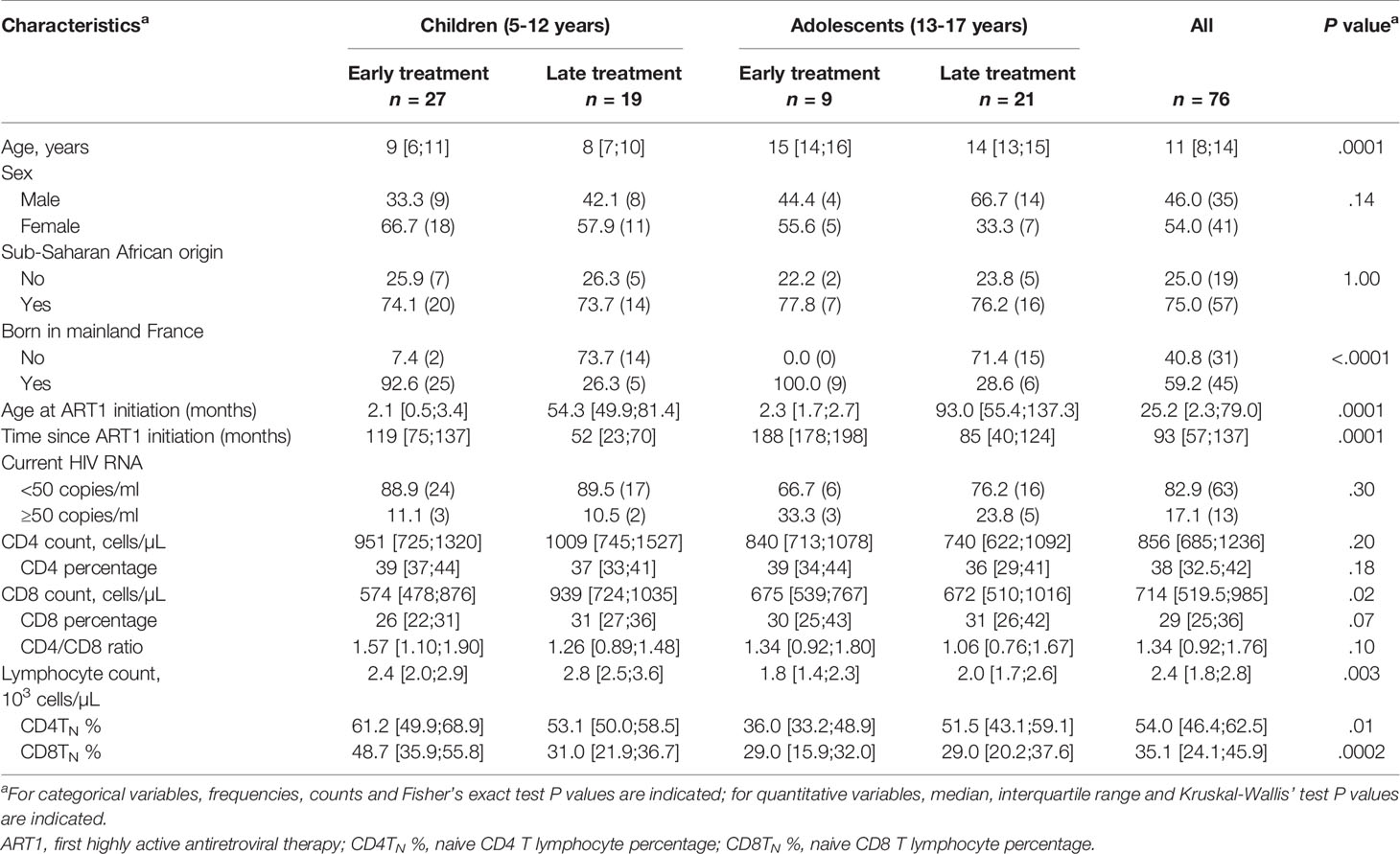
Table 1 Patient’s characteristics.
Early ART Was Not Significantly Associated With Higher CD4TN Percentages in Children, and Was Associated With Lower CD4TN Percentages In Adolescents
Blood CD4TN percentages were not significantly higher in E-Ch than in L-Ch (medians: 61.2% vs. 53.1%, P = .33). E-Ado had significantly lower CD4TN than L-Ado (36.0% vs. 51.5%, P = 0.02). The interaction between age and treatment group was significant (P = .008) and we performed analyses separately for children and adolescents.
High interindividual variation in the distribution of CD4 T-cell subsets was observed in all groups (Figures 1A, B). No significant differences were observed between the E-Ch and L-Ch groups (Figure 1A). E-Ado had significantly lower levels of recent thymic emigrants (TRTE, 27.2% vs. 41.4%, P = .03) and CD31negTN (4.0% vs. 6.5%, P = .01), and higher levels of transitional memory (TTM) cells (26.6% vs. 15.7%, P = .04) than L-Ado. CD4 T-cell counts were positively correlated with CD4TN and CD4TRTE, and negatively correlated with the levels of most memory subsets (Figure 1C).
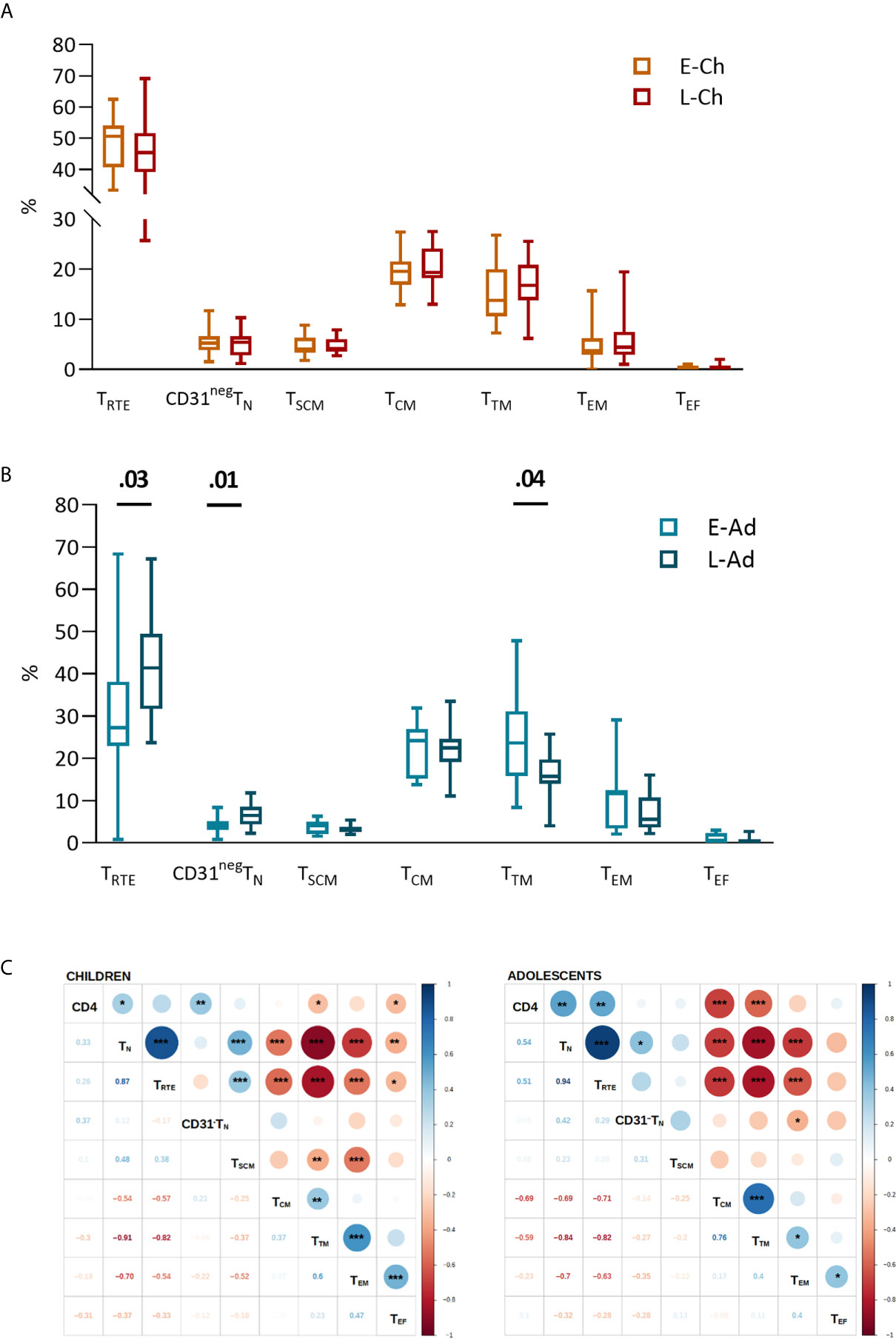
Figure 1 CD4 T-cell subsets in patients treated early and late. T-cell phenotypes were assessed by flow cytometry on fresh whole blood (Supplementary Material). The percentage of each subset among total CD4 T lymphocytes is presented for children (A) and adolescents (B), on box and whiskers plots showing the minimum and maximum values. The early and late treatment groups were compared in Mann-Whitney tests; when significant, P values are shown on the graph. Subsets are presented from the least to the most differentiated, and were defined as naive TN (CD45RA+CCR7+); recent thymic emigrants, TRTE (CD45RA+CCR7+CD31+); CD31negTN (CD45RA+CCR7+CD31-); central memory, TCM (CD45RA-CCR7+); transitional memory TTM (CD45RA-CCR7-CD27+); effector memory TEM (CD45RA-CCR7-CD27-) and effector TEF (CD45RA+CCR7-CD27-CD28-) cells. (C) The correlograms present Spearman’s rank correlation coefficients as symbols (upper quadrants) and values. Significant associations are indicated by symbols (*p < .05; **p < .01; ***p < .001). CD4 T-cell counts were used for correlation analyses.
We then searched for factors related to CD4TN levels, including demographic factors, immunovirological status at the time of the study and during the time since ART1 initiation. Biomarkers of immune activation (activated CD4/CD8TM, sCD14, sCD163, and CXCL10), inflammation (CRP and IL-6) and gut permeability (iFABP) (Figure 2 and Supplementary Table 2) were assessed in the samples used for TN quantification.
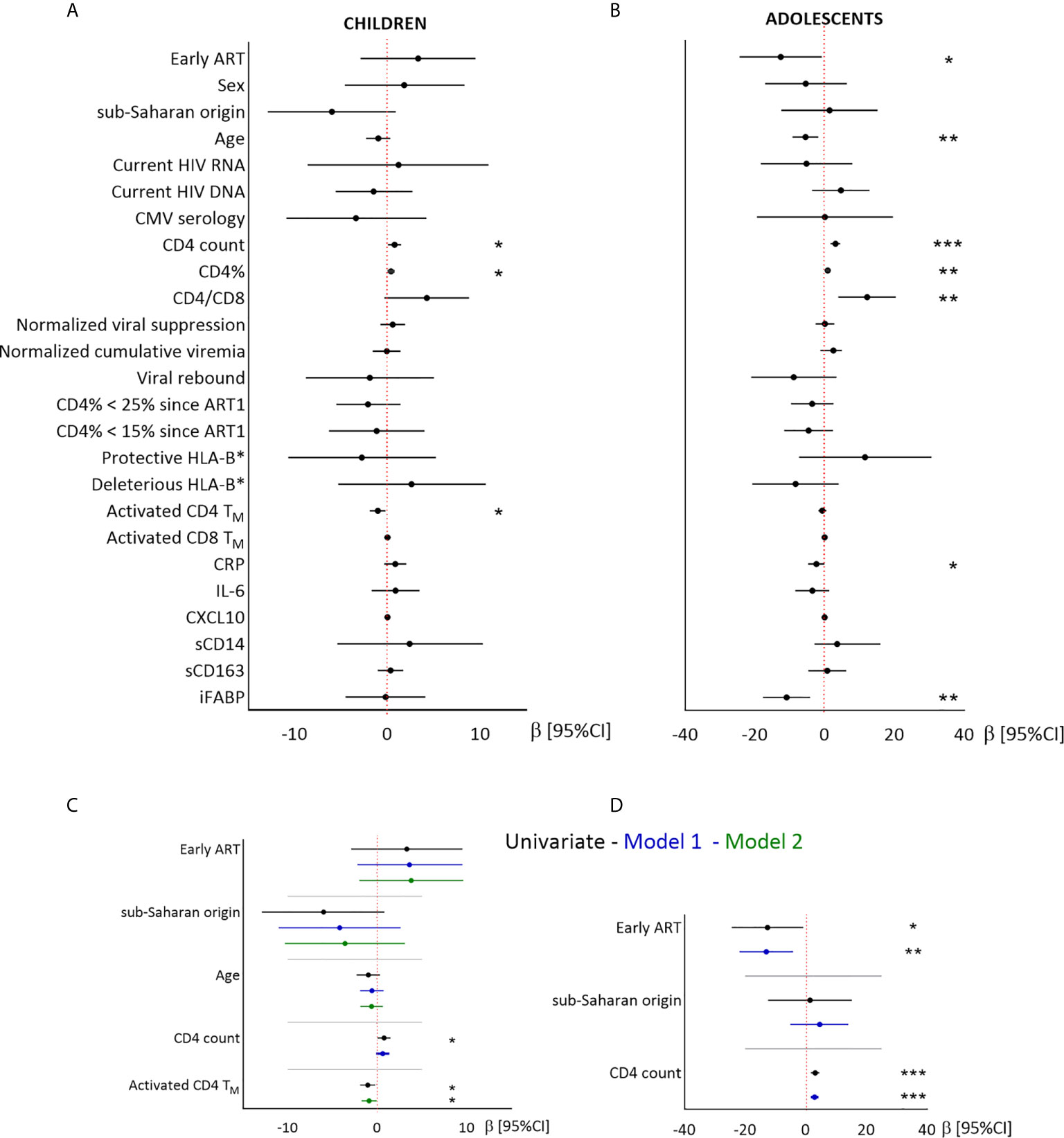
Figure 2 Linear regression analysis of the associations between CD4TN and demographic, virological and immunological factors in children and adolescents. Results from univariate (A, B) and multivariate (C, D) linear regressions are presented as estimates (β) and 95% confidence intervals. Significant associations are indicated by symbols (*p < .05; **p < .01; ***p < .001). Multivariate analysis included the covariables indicated on the plot. A and C: children; B and D: adolescents. Estimates are given per year, 100 CD4 T cells, and 1% activated CD4TM.
In children, higher CD4TN percentages were associated with higher CD4 T-cell counts and lower activated CD4TM percentages (Figure 2). Sex and geographic origin were not significantly associated with CD4TN percentages, but children of sub-Saharan origin tended to have lower CD4TN percentages than other children (56.5% vs 62.5%, P = 0.08, Figure 2 and Supplementary Table 2). After adjustment for treatment group, geographic origin and age, CD4TN percentage was not associated with CD4 T-cell counts (model 1), but was inversely correlated with activated CD4TM percentage (model 2).
In adolescents, higher CD4TN percentages were associated with late treatment, younger age, higher CD4 T-cell counts, and lower plasma CRP and iFABP levels, but not with either sex or geographic origin (Figure 2 and Supplementary Table 2). In multivariate analysis, higher CD4TN percentages were associated with late treatment and higher CD4 T-cell counts (model 1). Multivariate models including other variables were not robust to outliers (data not shown).
In conclusion, early ART was not significantly associated with higher CD4TN percentages in children, and was associated with lower CD4TN percentages in adolescents. In both groups, CD4TN percentages were associated with CD4 T-cell count and levels of immune activation and/or intestinal permeability.
Early ART Is Associated With Higher Naive CD8TN Percentages in Children but Not in Adolescents
E-Ch had significantly higher CD8TN percentages than L-Ch (48.7% vs. 31.0%, P = .001), and lower percentages of the most differentiated effector memory cells (CD27-CD28-CD8TEM, 6.1 vs. 14.6%, P = .009) (Figure 3A). E-Ado had significantly higher percentages of the least differentiated CD27+CD28+CD8TEM than L-ado (24.6% vs. 17.9%, P = .02), and similar levels of CD8TN (29.0% vs. 29.0%, P = .50) (Figure 3B). CD4 T-cell counts were correlated with CD8 T-cell subset levels (Figure 3C). Overall, early ART was associated with the least differentiated CD8 T lymphocytes.
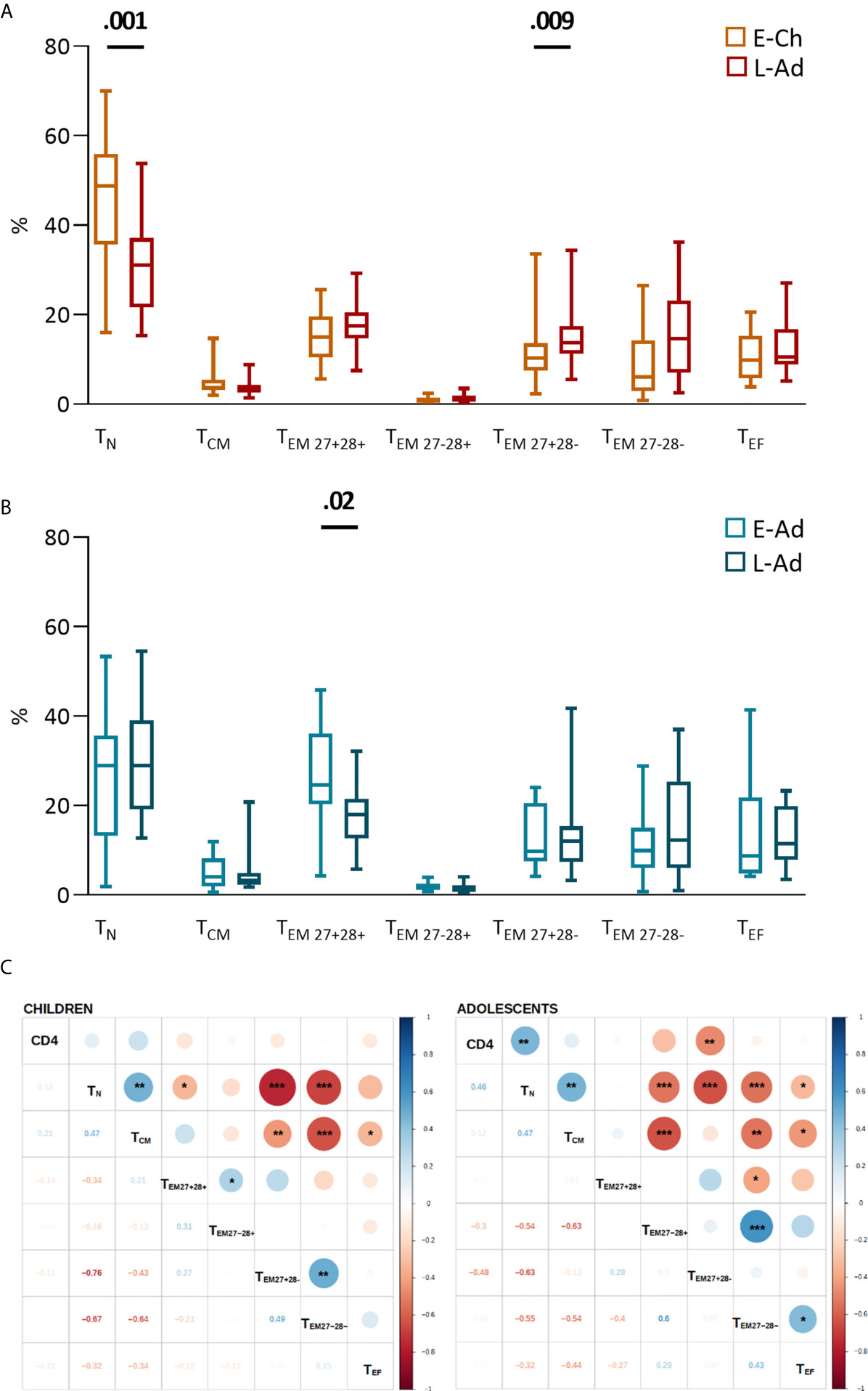
Figure 3 CD8 T-cell differentiation subsets in patients with early and late treatment initiation. T-cell phenotypes were assessed by flow cytometry on fresh whole blood (Supplementary Material). The percentage of each subset among total CD8 T lymphocytes is presented for children (A) and adolescents (B) on box and whiskers plots showing the minimum and maximum values. Early and late treatment groups were compared in Mann-Whitney tests; when significant, P values are shown on the graph. Subsets are presented from the least to the most differentiated, and were defined as naive TN (CD45RA+CCR7+); central memory TCM (CD45RA-CCR7+); effector memory TEM27+28+ (CD45RA-CCR7-CD27+CD28+); TEM27-28+ (CD45RA-CCR7- CD27-CD28+); TEM27+28- (CD45RA-CCR7- CD27+CD28-); TEM27-28- (CD45RA-CCR7- CD27-CD28-) and effector TEF (CD45RA+CCR7-CD27-CD28-) cells. (C) The correlograms present Spearman’s rank correlation coefficients as symbols (upper quadrants) and values. Significant associations are indicated by symbols (*p < .05; **p < .01; ***p < .001). CD4 T-cell counts were used to calculate correlations.
In children, higher CD8TN percentages were associated with early ART, a longer normalized duration of viral suppression, lower blood total HIV DNA levels, higher CD4/CD8 ratio, and lower sCD163 levels, but not with either sex or geographic origin (Figure 4 and Supplementary Table 3). Higher CD8TN percentages were independently associated with early ART after adjustment for the duration of viral suppression (model 1), and with higher CD4/CD8 ratio and lower HIV DNA levels (model 2). The associations with CMV serostatus and sCD163 were not significant in multivariate models (data not shown).
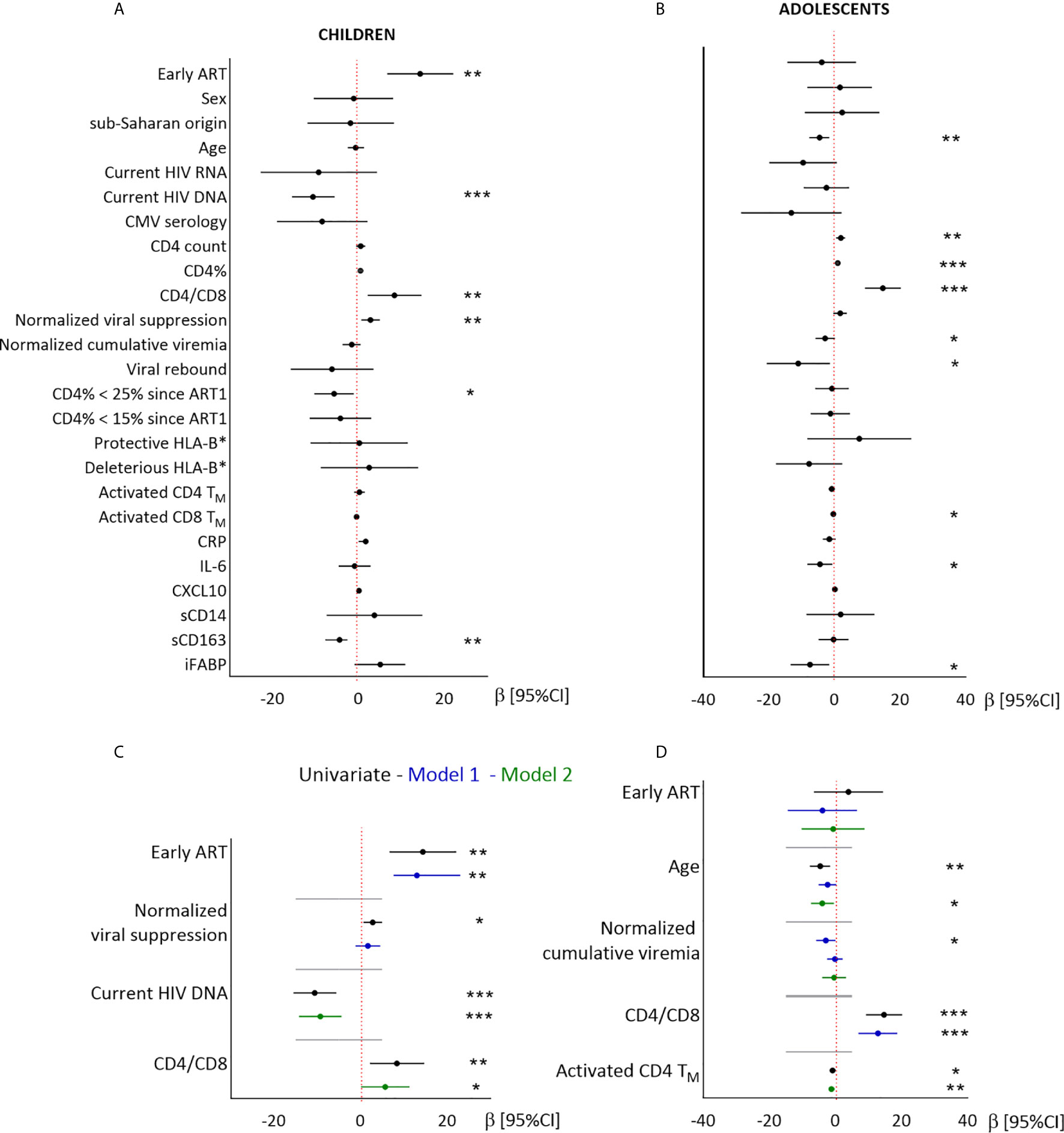
Figure 4 Linear regression analysis of the associations between CD8TN and demographic, virological and immunological factors in children and adolescents. Results from univariate (A, B) and multivariate (C, D) linear regressions are presented as estimates (β) and 95% confidence intervals. Significant associations are indicated by symbols (*p < .05; **p < .001; ***p < .0001). Multivariate analysis included the covariables indicated on the plot. Estimates are given per year, per 0.1 units of normalized viral suppression, per unit of normalized cumulative viremia, per point of CD4/CD8 ratio, and per 1% of activated CD4TM.
In adolescents, higher CD8TN percentages were not associated with early ART, sex or geographic origin, but were significantly associated with younger age, higher CD4/CD8 ratio, lower normalized cumulative viremia, and lower levels of activated CD4 and CD8TM cells, IL-6 and iFABP (Figure 4 and Supplementary Table 3). In multivariate models, higher CD8TN percentages were associated with younger age and higher CD4/CD8 ratio (model 1), or with younger age and lower activated CD4TM cell levels (model 2).
In conclusion, early ART was significantly associated with higher CD8TN percentages in children, but not in adolescents. In both groups, higher CD8TN percentages were associated with higher CD4/CD8 ratio and stronger immune activation.
Low CD4TN Levels in Adolescents Treated Early may Reflect Limited HIV-Driven TN Production by the Thymus
The lower CD4TN level in E-Ado than in other groups was unexpected. One patient had <5% CD4TN and this very low value was confirmed on the second blood sample. Relative to the reference values for Spain, a second patient had a CD4TN percentage below the 10th percentile, two had values above the 90th percentile, and five had CD4TN percentages between the 10th and 90th percentiles (20).
The E-Ado were born between 1999 and 2003, before early ART was recommended for all newborns. We hypothesized that they might have received early ART because of early diagnosis, reflecting in utero infection, or clinical symptoms observed from birth or developing during the first six months of life. We compared E-Ado to E-Ch, who were born between 2004 and 2012, when early ART was the standard of care (Table 2). HIV DNA and/or RNA tests were performed during the first week of life for 29 of the participants receiving early treatment: five of six E-Ado and 13 of 23 E-Ch tested positive, indicating prenatal HIV contamination (Fisher’s exact test, P = .36). Two patients in each group were diagnosed on the basis of suggestive symptoms (P = .25). Two of nine E-Ado and five of 27 E-Ch had at least one CDC stage C event (P = .99); these events occurred before the age of six months in both groups. Graphical analyses of CD4TN values as a function of early clinical events and early HIV diagnosis revealed no obvious relationship (Supplementary Figure 1). HIV RNA and CD4 levels before, at, and after ART1 initiation were not significantly different between E-Ch and E-Ado (Table 2). In conclusion, we found no statistically significant prescription bias (earlier ART related to early and/or severe infection) in E-Ado relative to E-Ch.
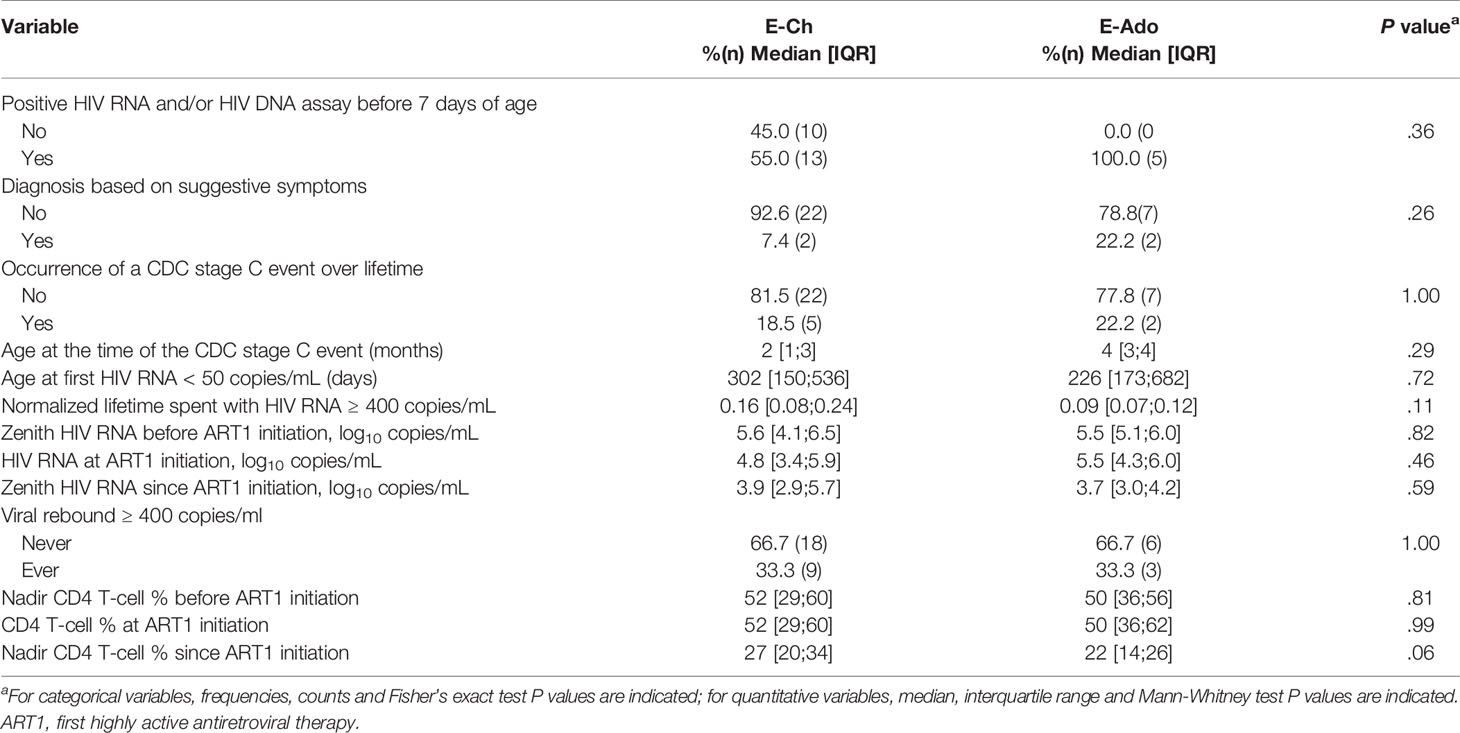
Table 2 HIV history variables in E-Ch and E-Ado.
HIV replication leads to a depletion of CD4 T cells and drives the de novo generation of TN by the thymus in young viremic patients (21–23). We compared E-Ado to L-Ado, whose median percentages of lifetime with HIV RNA levels > 400 copies/mL were 9% and 60%, respectively. The percentage TRTE among CD4TN — an indicator of thymic output —was lower in E-Ado than in L-Ado, but this difference was not significant (Table 3). The absolute duration of HIV RNA levels < 400 copies/mL since ART1 was inversely correlated with both CD4TN and CD4TRTE levels in adolescents (Spearman’s rho = -0.402, P = .02 and rho=-0.452, P = .01). Thus, the low CD4TN levels in E-Ado may reflect limited HIV-driven TN production by the thymus.
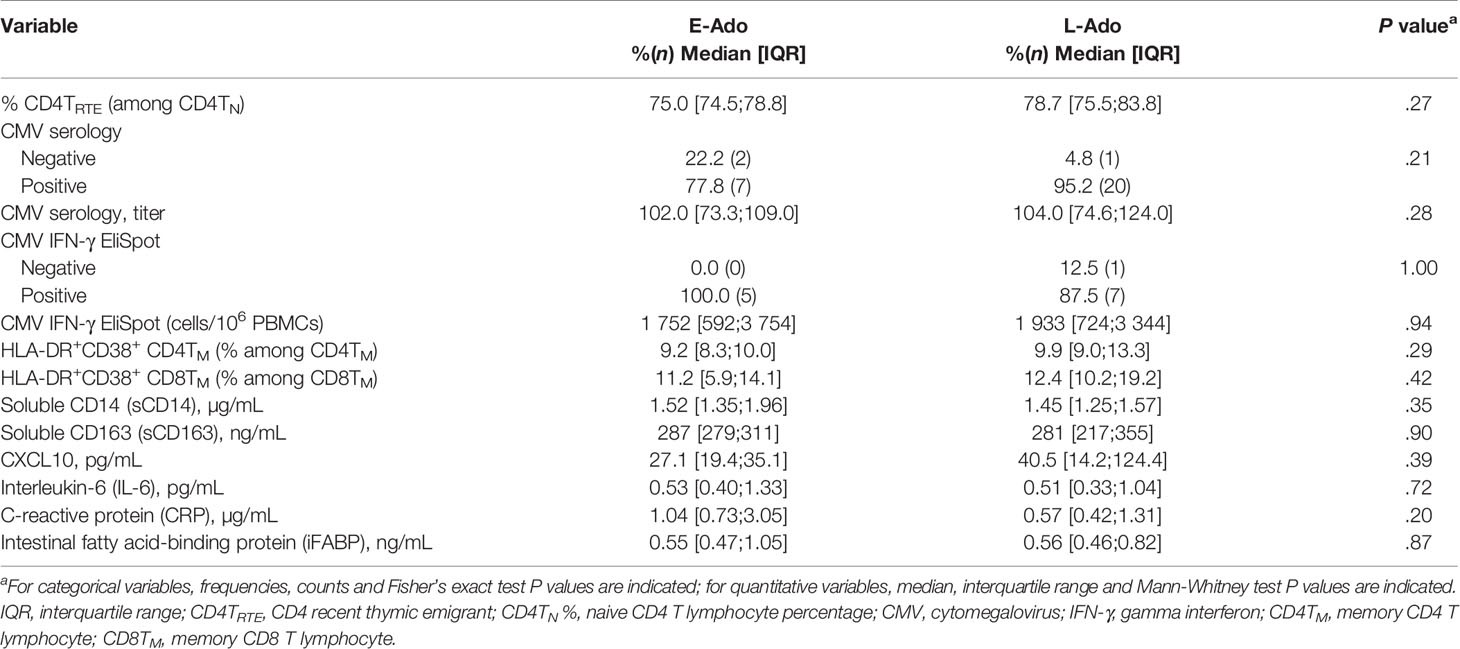
Table 3 Immune parameters in E-Ado and L-Ado.
The immune response to pathogens, inflammation and immune activation drive the recruitment of naive lymphocytes to the memory compartment and reduce thymic function (24). CMV prevalence, and the magnitude of CMV-specific serologic and T-cell responses were similar in L-Ado and E-Ado (Table 3), and blood samples from all patients tested negative for CMV DNA. These data argue against the low CD4TN levels being driven by an expansion of CMV-specific lymphocytes. E-Ado and L-Ado had similar levels of cellular and plasma markers of immune activation, inflammation, and gut permeability (Table 3). Overall, we did not observe higher levels of immune factors specific to E-Ado that could explain their low CD4TN levels.
In conclusion, we considered three explanations for the low CD4TN levels in E-Ado and analyzed the characteristics of the patients and correlations between variables. We found no indication for either a more severe disease profile or a more activated immune profile in E-Ado than in the other groups. Our data suggest that E-Ado have lower CD4TN levels than L-Ado because thymic activity is enhanced to maintain T-cell homeostasis in L-ado.
Discussion
The ANRS-EP59-CLEAC study included children and adolescents on ART with a good immunovirological status at the time of evaluation. In children, early ART had a beneficial effect for the maintenance of higher proportions of CD8TN lymphocytes and a marginal impact on CD4TN lymphocyte levels. By contrast, early ART had no beneficial effect in adolescents. CD4TN and CD8TN levels were negatively related to cellular activation or gut permeability.
The reported percentages of CD4TN were in the range of those published for age-matched healthy subjects, and only two E-Ado had values below the 10th percentile (20). CD4TN percentages were strongly associated with CD4 T-cell counts, as previously reported (21, 25–27). In our study population, CD4 T-cell counts were in the same range in the early and late treatment groups for both age strata (Table 1), reflecting a lessening of the benefits of early ART after several years of treatment (28–31). Most CD4TN are CD4TRTE, consistent with the thymus being active in pediatric patients (22, 23, 25, 32).
By contrast to these findings for CD4 T lymphocytes, early ART had a beneficial effect on CD8 T lymphocytes during childhood. E-Ch had median CD8TN values in the same range as uninfected controls, whereas the median values for L-Ch, E-Ado and L-Ado were lower than those of the reference groups (20); eight children and seven adolescents had values below the 10th percentile. E-Ch had a higher CD8TN and CD4/CD8 ratio, and lower total and CD8 T-cell counts than L-Ch (Table 1). The dichotomy between robust thymus-driven CD4TN recovery and the persistence of HIV-driven CD8TN loss is a key feature of pediatric HIV infection, reported in ART-treated young adults infected during the perinatal period, until their third decade of life (33). We show here that early ART keeps CD8TN levels high in children, with a possible impact on the future health of the child, because TN lymphocytes have a broad TCR repertoire and a high capacity to respond to new antigens (33).
TN percentages displayed a high level of interindividual variability in all groups. Among the expected factors (17, 34, 35), we found a trend towards lower CD8TN percentages in CMV-infected patients. CD4TN percentages were lower in patients born to mothers of Sub-Saharan origin than in those born to mothers of other geographic origins, although this difference was not significant. We found no association between TN levels and sex.
We document associations between higher immune activation and gut permeability biomarkers and lower TN percentages in at least one of the two age strata. In children, activated CD4TM were more strongly associated with CD4TN than CD4 T-cell counts. iFABP is a marker of intestinal permeability, and impaired mucosal immunity and decreases in thymic output influence each other (3, 36, 37). Higher CD4TN and CD8TN levels were found to be associated with lower iFABP levels, but only in adolescents. No difference in iFABP levels was observed between HIV-1-infected and uninfected infants (38–40), by contrast to what has been observed in adults (41). Thus, the data reported here and in other pediatric studies indicate that gut permeability and its impact on the immune system vary with age.
We found no association between CD4TN and virological history in either children or adolescents. Indeed, E-Ado had the lowest CD4TN, despite having the lowest normalized exposure to HIV RNA > 400 copies/ml (median 9% of their lifetime) and low HIV-DNA levels (16). In HIV-infected adolescents who had no access to early ART, CD4TN and TRTE levels were, paradoxically, higher in those with uncontrolled viremia, and HIV replication was correlated with CD4TN levels (21). Our observations are consistent with lower de novo thymic TN production in E-Ado than in L-Ado, whose CD4 T-cell compartment was less depleted by HIV replication. Reduced thymic activity may also account for the lack of association between early ART and higher CD8TN in adolescents. Such an association was expected because HIV replication is the main driver of CD8TN loss due to recruitment to the memory pool. However, this association was observed in children but not in adolescents. Thus, at adolescence, thymus-driven T-cell reconstitution may have a stronger influence on naive T lymphocytes than HIV replication.
We identified a small number of adolescents with low TN levels despite long-term viral suppression. Thymic failure may occur in pediatric patients, albeit less frequently than in adults (32). Rare cases of perinatally infected adults on suppressive ART with low CD4 T-cell counts were reported in an Italian study (33). By contrast, low CD4TN counts were mostly associated with poor adherence and active viral replication in an older population of perinatally infected patients living in France (42).
One limitation of this study is a probable survivor bias, particularly for adolescents, potentially resulting in an underestimation of the deleterious impact of late ART initiation. It was not possible to attribute differences (or the lack of difference) between E and L groups definitively to the timing of first ART, because data from an observational study necessarily reflect differences in care according to chronological time and place of birth. Nevertheless, pathophysiological description according to the timing of ART initiation is worthwhile, because it reflects the diversity of patients managed in France and other high-income countries.
In conclusion, we report that, in children over five years of age and adolescents, early ART clearly has a beneficial effect on CD8TN cells. Children in whom treatment was initiated late had high CD4TN levels, probably because of their robust thymic activity. They achieved CD4 T-cell counts similar to those of their peers receiving early treatment, whilst on suppressive therapy. Thus, CD8TN levels, and their principal clinical correlate, CD4/CD8 ratio, are valuable indicators for use in long-term immune reconstitution studies in pediatric patients.
Data Availability Statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Ethics Statement
The studies involving human participants were reviewed and approved by Comité de protection des personnes Île-de-France V. Written informed consent to participate in this study was provided by the participants’ legal guardian/next of kin.
Author Contributions
PF, JLC, CD, VA-F, JW and FB conceived the study and analyzed the data. PF supervised the clinical study. VA-F supervised the virological study. JW supervised the study methodology and statistical analyses. FB supervised the immunological study. TM, DB, and IF performed the immunological tests and analyzed immunological data. JLC performed the statistical analyses. PF, CD, AF, SB, AC, CF, IH, ML, NM, VM, M-DT and FV followed the patients and contributed to clinical data. AH participated to data collection, TW managed the database. FB wrote the manuscript. All authors contributed to the article and approved the submitted version.
Funding
ANRS (France Recherche Nord et Sud SIDA-HIV Hépatites) funded this study.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We thank the children and adolescents who participated in the study. We thank M. Hasan, S. Novault, P-H. Commere and T. Stephen from the Unité de Technologie et Service Cytometrie et Biomarqueurs (UTechS CB) at Institut Pasteur. The CMV pp65 peptide pool #11549 was obtained through the AIDS Reagent Program, Division of AIDS, NIAID, NIH. This text has been verified by a native English speaker.
The ANRS CLEAC Study Group
Hôpital Armand Trousseau, Paris: Mary-France Courcoux, Catherine Dollfus, Marie-Dominique Tabone, Geneviève Vaudre; Hôpital Bicêtre, Le Kremlin-Bicêtre: Corinne Fourcade, Josiane Warsazawski, Jérôme Lechenadec, Olivia Dialla, Laura Nailler, Lamya Ait-Si-Selmi, Isabelle Leymarie, Thierry Wack, Alexandre Hoctin, Razika Feraon-Nanache; Centre hospitalier intercommunal de Créteil: Isabelle Hau; Hôpital Delafontaine, Saint-Denis: Cécile Gakobwa; Hôpital Necker-Enfants Malades, Paris: Véronique Avettand-Fenoël, Stéphane Blanche, Marine Fillion, Pierre Frange, Nizar Mahlaoui, Adeline Mélard, Florence Veber, Marie-Christine Mourey; Maternité Port-Royal, Paris: Valérie Marcou; Hôpital Robert Debré, Paris: Albert Faye, Martine Lévine, Sandrine Richard; Pitié-Salpêtrière, Paris: Brigitte Autran, Assia Samri, Mariama Diallo; Hôpital Saint-Louis: Sophie Caillat-Zucman, Kahina Amokrane, Rayna Ivanova-Derin, Centre hospitalier intercommunal de Villeneuve-Saint-Georges: Anne Chacé; Institut Pasteur Paris: Florence Buseyne, Thomas Montange, Damien Batalie, Ingrid Fert, Asier Saez-Cirion, Valérie Monceaux, Daniel Scott-Algara; ANRS: Lucie Marchand, Delphine Lebrasseur, Axel Levier. ANRS (France Recherche Nord et Sud SIDA-HIV Hépatites) was the sponsor of the study.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fimmu.2021.662894/full#supplementary-material
References
1. Dollfus C, Le Chenadec J, Faye A, Blanche S, Briand N, Rouzioux C, et al. Long-term outcomes in adolescents perinatally infected with HIV-1 and followed up since birth in the French Perinatal Cohort (EPF/ANRS CO10). Clin Infect Dis (2010) 51:214–24. doi: 10.1086/653674
2. Chiappini E, Bianconi M, Dalzini A, Petrara MR, Galli L, Giaquinto C, et al. Accelerated aging in perinatally HIV-infected children: clinical manifestations and pathogenetic mechanisms. Aging (2018) 10:3610–25. doi: 10.18632/aging.101622
3. Zicari S, Sessa L, Cotugno N, Ruggiero A, Morrocchi E, Concato C, et al. Immune activation, inflammation, and non-AIDS co-morbidities in HIV-infected patients under long-term ART. Viruses (2019) 11:200. doi: 10.3390/v11030200
4. Cotton MF, Violari A, Otwombe K, Panchia R, Dobbels E, Rabie H, et al. Early time-limited antiretroviral therapy versus deferred therapy in South African infants infected with HIV: results from the children with HIV early antiretroviral (CHER) randomised trial. Lancet (2013) 382:1555–63. doi: 10.1016/S0140-6736(13)61409-9
5. Shiau S, Abrams EJ, Arpadi SM, Kuhn L. Early antiretroviral therapy in HIV-infected infants: can it lead to HIV remission? Lancet HIV (2018) 5:e250–e58. doi: 10.1016/s2352-3018(18)30012-2
6. Sabin CA, Smith CJ, Monforte AD, Battegay M, Gabiano C, Galli L, et al. Response to combination antiretroviral therapy: variation by age - The Collaboration of Observational HIV Epidemiological Research Europe (COHERE) study group. AIDS (2008) 22:1463–73. doi: 10.1016/S0140-6736(11)60208-0
7. Picat MQ, Lewis J, Musiime V, Prendergast A, Nathoo K, Kekitiinwa A, et al. Predicting patterns of long-term CD4 reconstitution in HIV-infected children starting antiretroviral therapy in sub-Saharan Africa: a cohort-based modelling study. PloS Med (2013) 10:e1001542. doi: 10.1371/journal.pmed.1001542
8. Azzoni L, Barbour R, Papasavvas E, Glencross DK, Stevens WS, Cotton MF, et al. Early ART results in greater immune reconstitution benefits in HIV-infected infants: Working with data missingness in a longitudinal dataset. PloS One (2015) 10:e0145320. doi: 10.1371/journal.pone.0145320
9. Guillen S, Prieto L, Jimenez de Ory S, Gonzalez-Tome MI, Rojo P, Navarro ML, et al. Prognostic factors of a lower CD4/CD8 ratio in long term viral suppression HIV infected children. PloS One (2019) 14:e0220552. doi: 10.1371/journal.pone.0220552
10. Buseyne F, Scott-Algara D, Bellal N, Burgard M, Rouzioux C, Blanche S, et al. The frequency of HIV-specific interferon-gamma-producing CD8 T cells is associated with both age and level of antigenic stimulation in HIV-1-infected children. J Infect Dis (2005) 192:1781–86. doi: 10.1086/497152
11. Scott ZA, Chadwick EG, Gibson LL, Catalina MD, McManus MM, Yogev R, et al. Infrequent detection of HIV-1-specific, but not cytomegalovirus-specific, CD8+ T cell responses in young HIV-1-infected infants. J Immunol (2001) 167:7134–40. doi: 10.4049/jimmunol.167.12.7134
12. Rinaldi S, Pallikkuth S, Cameron M, de Armas LR, Cotugno N, Dinh V, et al. Impact of early antiretroviral therapy initiation on HIV-specific CD4 and CD8 T-cell function in perinatally infected children. J Immunol (2020) 204:540–49. doi: 10.4049/jimmunol.1900856
13. Aupiais C, Faye A, Le Chenadec J, Rouzioux C, Bouallag N, Laurent C, et al. Interruption of cART in clinical practice is associated with an increase in the long-term risk of subsequent immunosuppression in HIV-1-infected children. Pediatr Infect Dis J (2014) 33:1237–45. doi: 10.1097/inf.0000000000000450
14. Palladino C, Briz V, Bellon JM, Climent FJ, de Ory SJ, Mellado MJ, et al. Determinants of highly active antiretroviral therapy duration in HIV-1-infected children and adolescents in Madrid, Spain, from 1996 to 2012. PloS One (2014) 9:e96307. doi: 10.1371/journal.pone.0096307
15. Kakkar F, Lee T, Hawkes MT, Brophy J, Lindy S, Singer J, et al. Challenges to achieving and maintaining viral suppression among children living with HIV. AIDS (2020) 34:687–97. doi: 10.1097/qad.0000000000002454
16. Avettand-Fenoel V, Le Chenadec J, Diallo M, Fillion M, Mélard A, Dollfus C, et al. Initiating antiretroviral treatment early in infancy has long-term benefits on the HIV reservoir in late childhood and adolescence. Clin Infect Dis (2021). doi: 10.1093/cid/ciaa1931. Advanced online publication.
17. Gaballa A, Clave E, Uhlin M, Toubert A, Arruda LCM. Evaluating thymic function after human hematopoietic stem cell transplantation in the personalized medicine era. Front Immunol (2020) 11:1341. doi: 10.3389/fimmu.2020.01341
18. Avettand-Fenoel V, Chaix ML, Blanche S, Burgard A, Floch C, Toure K, et al. LTR real-time PCR for HIV-1 DNA quantitation in blood cells for early diagnosis in infants born to seropositive mothers treated in HAART Area (ANRS CO 01). J Med Virol (2009) 81:217–23. doi: 10.1002/jmv.21390
19. Avettand-Fenoel V, Blanche S, Le Chenadec J, Scott-Algara D, Dollfus C, Viard JP, et al. Relationships between HIV disease history and blood HIV-1 DNA load in perinatally infected adolescents and young adults: the ANRS-EP38-IMMIP Study. J Infect Dis (2012) 205:1520–28. doi: 10.1093/infdis/jis233
20. Garcia-Prat M, Álvarez-Sierra D, Aguiló-Cucurull A, Salgado-Perandrés S, Briongos-Sebastian S, Franco-Jarava C, et al. Extended immunophenotyping reference values in a healthy pediatric population. Cytom B Clin Cytom (2019) 96:223–33. doi: 10.1002/cyto.b.21728
21. Blanche S, Scott-Algara D, Le Chenadec J, Didier C, Montange T, Avettand-Fenoel V, et al. Naive T lymphocytes and recent thymic emigrants are associated with HIV-1 disease history in French adolescents and young adults infected in the perinatal period: The ANRS-EP38-IMMIP study. Clin Infect Dis (2014) 58:573–87. doi: 10.1093/cid/cit729[doi
22. Nobile M, Correa R, Borghans JAM, D’Agostino C, Schneider P, de Boer RJ, et al. De novo T-cell generation in patients at different ages and stages of HIV-1 disease. Blood (2004) 104:470–77. doi: 10.1182/blood-2003-12-4265
23. Aguilera-Sandoval CR, Yang OO, Jojic N, Lovato P, Chen DY, Boechat MI, et al. Supranormal thymic output up to 2 decades after HIV-1 infection. AIDS (2016) 30:701–11. doi: 10.1097/QAD.0000000000001010
24. Appay V, Fastenackels S, Katlama C, Ait-Mohand H, Schneider L, Guihot A, et al. Old age and anti-cytomegalovirus immunity are associated with altered T-cell reconstitution in HIV-1-infected patients. AIDS (2011) 25:1813–22. doi: 10.1097/QAD.0b013e32834640e6
25. Lewis J, Walker AS, Castro H, De Rossi A, Gibb DM, Giaquinto C, et al. Age and CD4 count at initiation of antiretroviral therapy in HIV-infected children: effects on long-term T-cell reconstitution. J Infect Dis (2012) 205:548–56. doi: 10.1093/infdis/jir787
26. Zakhour R, Tran DQ, Degaffe G, Bell CS, Donnachie E, Zhang W, et al. Recent thymus emigrant CD4+ T cells predict HIV disease progression in patients with perinatally acquired HIV. Clin Infect Dis (2016) 62:1029–35. doi: 10.1093/cid/ciw030
27. Lewis J, Payne H, Walker AS, Otwombe K, Gibb DM, Babiker AG, et al. Thymic output and CD4 T-cell reconstitution in HIV-infected children on early and interrupted antiretroviral treatment: Evidence from the children with HIV early antiretroviral therapy trial. Front Immunol (2017) 8:1162. doi: 10.3389/fimmu.2017.01162
28. Goetghebuer T, Le Chenadec J, Haelterman E, Galli L, Dollfus C, Thorne C, et al. Short- and long-term immunological and virological outcome in HIV-infected infants according to the age at antiretroviral treatment initiation. Clin Infect Dis (2012) 54:878–81. doi: 10.1093/cid/cir950
29. Cohen S, Smit C, van Rossum AM, Fraaij PL, Wolfs TF, Geelen SP, et al. Long-term response to combination antiretroviral therapy in HIV-infected children in the Netherlands registered from 1996 to 2012. AIDS (2013) 27:2567–75. doi: 10.1097/01.aids.0000432451.75980.1b
30. Desmonde S, Dicko F, Koueta F, Eboua T, Balestre E, Amani-Bosse C, et al. Association between age at antiretroviral therapy initiation and 24-month immune response in West-African HIV-infected children. AIDS (2014) 28:1645–55. doi: 10.1097/QAD.0000000000000272
31. Novelli S, Lecuroux C, Avettand-Fenoel V, Seng R, Essat A, Morlat P, et al. Long-term therapeutic impact of the timing of antiretroviral therapy in patients diagnosed with primary human immunodeficiency virus type 1 infection. Clin Infect Dis (2018) 66:1519–27. doi: 10.1093/cid/cix1068
32. Ferrando-Martinez S, De Pablo-Bernal RS, De Luna-Romero M, De Ory SJ, Genebat M, Pacheco YM, et al. Thymic function failure is associated with human immunodeficiency virus disease progression. Clin Infect Dis (2017) 64:1191–97. doi: 10.1093/cid/cix095
33. Paghera S, Quiros-Roldan E, Sottini A, Properzi M, Castelli F, Imberti L. Lymphocyte homeostasis is maintained in perinatally HIV-infected patients after three decades of life. Immun Ageing (2019) 16:26. doi: 10.1186/s12979-019-0166-7
34. Turner JE, Campbell JP, Edwards KM, Howarth LJ, Pawelec G, Aldred S, et al. Rudimentary signs of immunosenescence in Cytomegalovirus-seropositive healthy young adults. Age (2014) 36:287–97. doi: 10.1007/s11357-013-9557-4
35. van den Heuvel D, Jansen MAE, Nasserinejad K, Dik WA, van Lochem EG, Bakker-Jonges LE, et al. Effects of nongenetic factors on immune cell dynamics in early childhood: The Generation R Study. J Allergy Clin Immunol (2017) 139:1923–34.e17. doi: 10.1016/j.jaci.2016.10.023
36. Bourgeois C, Hao Z, Rajewsky K, Potocnik AJ, Stockinger B. Ablation of thymic export causes accelerated decay of naive CD4 T cells in the periphery because of activation by environmental antigen. PNAS (2008) 105:8691–96. doi: 10.1073/pnas.0803732105
37. De Voeght A, Martens H, Renard C, Vaira D, Debruche M, Simonet J, et al. Exploring the link between innate immune activation and thymic function by measuring sCD14 and TRECs in HIV patients living in Belgium. PloS One (2017) 12:e0185761. doi: 10.1371/journal.pone.0185761
38. Koay WLA, Lindsey JC, Uprety P, Bwakura-Dangarembizi M, Weinberg A, Levin MJ, et al. Intestinal integrity biomarkers in early antiretroviral-treated perinatally HIV-1-infected infants. J Infect Dis (2018) 218:1085–89. doi: 10.1093/infdis/jiy271
39. Prendergast AJ, Chasekwa B, Rukobo S, Govha M, Mutasa K, Ntozini R, et al. Intestinal damage and inflammatory biomarkers in human immunodeficiency virus (HIV)–exposed and HIV-infected Zimbabwean infants. J Inf Dis (2017) 216:651–61. doi: 10.1093/infdis/jix367
40. Dirajlal-Fargo S, El-Kamari V, Weiner L, Shan L, Sattar A, Kulkarni M, et al. Altered intestinal permeability and fungal translocation in Ugandan children with human immunodeficiency virus. Clin Infect Dis (2020) 70:2413–22. doi: 10.1093/cid/ciz561
41. Sandler NG, Wand H, Roque A, Law M, Nason MC, Nixon DE, et al. Plasma levels of soluble CD14 independently predict mortality in HIV infection. J Infect Dis (2011) 203:780–90. doi: 10.1093/infdis/jiq118
Keywords: HIV-1, children, adolescents, early ART, T lymphocyte, naive T lymphocyte
Citation: Frange P, Montange T, Le Chenadec J, Batalie D, Fert I, Dollfus C, Faye A, Blanche S, Chacé A, Fourcade C, Hau I, Levine M, Mahlaoui N, Marcou V, Tabone M-D, Veber F, Hoctin A, Wack T, Avettand-Fenoël V, Warszawski J and Buseyne F (2021) Impact of Early Versus Late Antiretroviral Treatment Initiation on Naive T Lymphocytes in HIV-1-Infected Children and Adolescents – The-ANRS-EP59-CLEAC Study. Front. Immunol. 12:662894. doi: 10.3389/fimmu.2021.662894
Received: 01 February 2021; Accepted: 24 March 2021;
Published: 22 April 2021.
Edited by:
Francesca Chiodi, Karolinska Institutet (KI), SwedenReviewed by:
Daria Trabattoni, University of Milan, ItalyMartyn Andrew French, University of Western Australia, Australia
Copyright © 2021 Frange, Montange, Le Chenadec, Batalie, Fert, Dollfus, Faye, Blanche, Chacé, Fourcade, Hau, Levine, Mahlaoui, Marcou, Tabone, Veber, Hoctin, Wack, Avettand-Fenoël, Warszawski and Buseyne. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Florence Buseyne, ZmxvcmVuY2UuYnVzZXluZUBwYXN0ZXVyLmZy